荒漠蓝藻规模化培养试验
王楠 , 1 , 2 , 许文文 1 , 2 , 赵燕翘 1 , 2 , 赵洋 , 1
1.中国科学院西北生态环境资源研究院 沙坡头沙漠研究试验站,甘肃 兰州 730000
2.中国科学院大学,北京 100049
Large-scale culture experiment of desert cyanobacteria
Wang Nan , 1 , 2 , Xu Wenwen 1 , 2 , Zhao Yanqiao 1 , 2 , Zhao Yang , 1
1.Shapotou Desert Research and Experiment Station,Northwest Institute of Eco-Environment and Resources,Chinese Academy of Sciences,Lanzhou 730000,China
2.University of Chinese Academy of Sciences,Beijing 100049,China
通讯作者: 赵洋(E-mail: zhaoyang66@126.com )
收稿日期: 2021-10-26
修回日期: 2021-12-13
基金资助:
国家自然科学基金项目 . 32171630 中国科学院“西部之光”人才计划项目
Received: 2021-10-26
Revised: 2021-12-13
摘要
以人工生物土壤结皮治理沙化土地技术是近年来新兴的绿色环保固沙技术,是目前沙区生态恢复研究的热点。然而,用于人工生物土壤结皮培养的荒漠蓝藻等生物体的产量是限制该技术大规模应用的瓶颈。为阐明荒漠蓝藻规模化培养的生长特征及其限制因子,以分离于腾格里沙漠蓝藻结皮中的优势荒漠蓝藻——鱼腥藻(Anabaena cylindrica )、具鞘微鞘藻(Microcoleus vaginatus )、念珠藻(Nostoc carneum )、席藻(Phormidium sp. NIES-2121)、伪枝藻(Scytonema sp. NIES-4073)、爪哇伪枝藻(Scytonema javanicum )和单歧藻(Tolypothrix tenuis )为研究对象,测定了不同扰动时长下荒漠蓝藻的生长特征和产量,分析了荒漠蓝藻的生长与扰动时长、温度、光照强度、光合有效辐射、pH和电导率对荒漠蓝藻的影响。结果显示:荒漠蓝藻接种后干重先迅速增加后逐渐趋于稳定,而6 h和12 h扰动下干重在培养后期高于3 h扰动;荒漠蓝藻在3、6、12 h扰动下的总质量分别为每培养池(15 m3 )0.858、0.940、0.841 kg;在3 h和12 h扰动下荒漠蓝藻干重与日均温度呈显著二项式相关,与日均光照强度和日均光合有效辐射呈显著负相关;荒漠蓝藻总质量与有效积温显著正相关。6 h扰动时长下荒漠蓝藻的总质量最高,且6 h扰动下荒漠蓝藻长势好于3 h,较12 h扰动耗能小。在荒漠蓝藻规模化培养时应将扰动时长设置为6 h,且确保生长期内的有效积温应不低于145 ℃。
关键词:
人工生物土壤结皮 荒漠蓝藻 规模化培养 扰动时长 有效积温
Abstract
In recent years, the artificial biological soil crusts applied in land desertification has been developed to be a technology of new and green in the environment protection particular in fixing moving sand and the study of ecological restoration of desert areas. However, the yield of desert cyanobacteria and other organisms used for the cultivation of artificial biological soil crusts is a bottleneck that limits the large-scale application of this technology. Thereby, to explore the growth characteristics of mass-cultivation of desert cyanobacteria and their effect factors, seven predominated species isolating from Tengger Desert cyanobacteria crusts, including Microcolus vaginatus , Anabaena sp., Nostoc sp., Phormidium sp., Scytonema sp., Tolypothrix sp. and Scytonema javanicum , were selected as subjects. Results showed that the dry weight of desert cyanobacteria firstly quickly increased after inoculation and then slowly tend to stabilize. And, the dry weight of desert cyanobacteria under the disturbance of 6- and 12-hours was higher than that of the disturbance of 3 hours in the later cultivation. What's more, the total masses of desert cyanobacteria under the disturbance of 3-, 6- and 12-hours were 0.858-, 0.940- and 0.841-kg·pond-1 , respectively. Under the disturbance of 3- and 12-hours, the dry weight of desert cyanobacteria had a significant binomial relationship with varying temperature and a negative relationship with changes in the light intensity and the photosynthetically active radiation; also, the total mass of desert cyanobacteria was significantly positively correlated with the effective accumulated temperature. In conclusion, disturbance of 6 hours not only produced the highest total mass of desert cyanobacteria but developed the higher growth ratio of desert cyanobacteria very differently than under the disturbance of 3 hours; in the meantime, the consuming energy of desert cyanobacteria under the disturbance of 6 hours was less than that under the disturbance of 12 hours. Therefore, this study hints the disturbance duration during the growth period of desert cyanobacteria should be set to 6 hours and the effective accumulated temperature should be ≥145 ℃.
Keywords:
artificial biological soil crust desert cyanobacteria scale cultivation disturbance duration effective accumulated temperature
本文引用格式
王楠, 许文文, 赵燕翘, 赵洋. 荒漠蓝藻规模化培养试验 . 中国沙漠 [J], 2022, 42(4): 181-189 doi:10.7522/j.issn.1000-694X.2021.00153
Wang Nan, Xu Wenwen, Zhao Yanqiao, Zhao Yang. Large-scale culture experiment of desert cyanobacteria . Journal of Desert Research
0 引言
生物土壤结皮(Biological Soil Crusts,BSCs)是由蓝藻、绿藻、地衣、藓类和异养微生物及相关的其他生物体与土壤表层颗粒等非生物体胶结形成的复杂复合体,约占全球陆地面积的12.2%[1 -2 ] 。BSCs具有稳定土壤,增加C、N固定,调节土壤水文过程及提高生物多样性等功能,被誉为“荒漠生态系统工程师”[3 -4 ] 。近年来,BSCs在固沙方面的作用得到了广泛认可,其形成和发育可以降低荒漠生态系统非生物因子的胁迫水平,从而维持系统的健康[5 ] 。但BSCs自然形成需要几十年,人工培育方法可以缩短BSCs的形成时间,并迅速对沙化土壤进行固定,从而改善土壤生境,成为沙化土地治理的新模式[6 -7 ] 。荒漠蓝藻作为荒漠、半荒漠生态系统的先行者,是BSCs形成初期的核心组分,能够在恶劣的生境条件(干旱、营养贫瘠、强紫外线辐射等)下生存繁衍,而且比地衣和藓类,容易培养,从而成为人工培育BSCs的首选理想物种[3 ] 。然而,足量的荒漠蓝藻是使用该技术进行大规模沙化土地固定的基础,显然,荒漠蓝藻的规模化培养是该技术推广和应用的重要前提和保障。但以往的研究中关于荒漠蓝藻培养的容器较小,无法满足大规模应用的需求,因此亟待系统地开展荒漠蓝藻规模化培养研究。
荒漠蓝藻的生长和产量除受到温度、光照、营养盐等环境条件的影响外,也会受到不同时间尺度上扰动过程的控制[8 -10 ] 。水体扰动能够通过增加悬浮物浓度从而影响水体光照条件,进而影响藻类的光合效率 [11 ] 。不同扰动方式下藻类的敏感度和耐受能力不同,其营养盐吸收速率也会表现出明显差异[12 -13 ] 。此外,较强的扰动条件会破坏藻细胞的结构、造成细胞的机械损伤、改变其生物量和沉降量等[14 ] 。目前,水体扰动对蓝藻生长影响的研究主要在水生/水华蓝藻方面,对不同扰动时长下荒漠蓝藻生长状况有待进一步研究[15 ] 。
蓝藻完成某一生育期或完整生命史过程需要一定量的日平均温度的累积,特别要求有特定的有效积温[16 ] 。不同地区蓝藻对温度的适应范围不同,这会导致蓝藻生长期内所需有效积温差异明显[17 ] 。在太湖水体,上半年(1—6月)的逐日水温积温对同年春末夏初蓝藻爆发的强度有决定作用,且当温度累积到一定程度时,蓝藻才会大面积产生[18 ] 。而北方寒冷内陆性碱性水体在有效积温较低时有助于蓝藻爆发[17 ] 。此外,日照也是蓝藻增殖的关键条件,累积光照强度和累积光合有效辐射同样可视为影响蓝藻生长的关键因素[17 ,19 ] 。目前,关于影响荒漠蓝藻生长的环境因子研究主要围绕在N、P等营养盐浓度,最适生长温度和光照强度等方面,且集中于实验室条件,而荒漠蓝藻生长对温室生态环境的响应机制研究较少[20 -22 ] 。有效积温、累积光照强度和累积光合有效辐射显著影响水生蓝藻的生物量积累,但是否对荒漠蓝藻也会产生影响,仍鲜见报道[16 ] 。
鱼腥藻(Anabaena cylindrica )、具鞘微鞘藻(Microcoleus vaginatus )、念珠藻(Nostoc carneum )、席藻(Phormidium sp. NIES-2121)、伪枝藻(Scytonema sp. NIES-4073)、爪哇伪枝藻(Scytonema javanicum )和单歧藻(Tolypothrix tenuis )是腾格里沙漠蓝藻结皮中的优势物种[3 ,5 ] 。鉴于混合荒漠蓝藻在人工BSCs培育中的广泛应用[3 ] ,本研究将上述7种蓝藻组成的混合藻种作为研究对象,研究了规模化培养时荒漠蓝藻的生长特征,探讨了扰动时长、温度、光照强度和光合有效辐射对蓝藻生长的影响,并引入农业气象学中有效积温、累积光照强度和累积光合有效辐射探究它们与荒漠蓝藻产量间的关系,将为荒漠蓝藻的规模化培养提供理论依据。
1 材料与方法
1.1 荒漠蓝藻工厂化生产基地概况
本研究在中国科学院沙坡头沙漠试验研究站(37°27′N、105°00′E,海拔1 339 m)荒漠蓝藻工厂化生产基地进行,该基地包括日光温室(长×宽×高=40 m×9 m×3 m)一间,温室内有6个蓝藻培养池(长×宽×深=15 m×1 m×1 m,图1 )。
图1
图1
荒漠蓝藻工厂化生产基地(A,日光温室;B,蓝藻培养池)
Fig.1
Desert cyanobacteria factory production base (A, Solar greenhouse; B, Cyanobacteria cultivation pond)
1.2 试验设计和方法
具鞘微鞘藻、席藻、鱼腥藻、念珠藻、伪枝藻、单歧藻和爪哇伪枝藻均由腾格里沙漠东南缘发育良好的蓝藻结皮中分离、纯化所得,使用显微镜进行物种鉴定[23 -24 ] 。荒漠蓝藻规模化培养步骤如下:首先,将7种蓝藻先用装有无菌BG·11培养液的三角瓶在摇床上进行一级培养,培养10—15 d;随后,将7种蓝藻全部转入装有BG·11培养液塑料培养箱(长×宽×深=0.5 m×0.8 m×0.6 m)中进行10—15 d的二级扩繁。上述培养均在实验室条件下进行,培养温度25±2 ℃、光照强度4 000 lux,光暗比12 h∶12 h。最后,将培养的混合藻种以8 g·t-1 (干重)转入蓝藻培养池内进行规模化培养,培养池中加入7.5 t BG·11培养液[25 ] (水深0.5 m)。
规模化培养开展于2020年9—10月,分别设置3 h(07:00—08:00, 12:00—13:00,16:00—17:00)、6 h(07:00—09:00, 11:00—13:00,15:00—17:00)和12 h(07:00—19:00)3种扰动时长。使用潜水泵(QDX15-10-0.75L2,中国浙江省振圣泵业股份有限公司)对培养池进行扰动,并利用微电脑时空开关控制扰动时长。由于共有6个蓝藻培养池,3种扰动时长设置3次重复无法同时开展试验,因此,共进行3个周期的试验。3种扰动时长在每个周期试验中分别设置2个重复,3个周期共6个重复,每个培养周期的时间均为12 d。此外,为了消除微环境异质性带来的差异, 3个周期试验中6个蓝藻培养池对应设置的扰动时长均不同。
1.3 样品采集与指标测定
采用HOBO Pendant 数据记录仪(MX2201-MX2202,Onset,美国)自动监测温度和光照强度,采用光合有效辐射传感器(SQ-110,Apogee,美国)自动监测光和有效辐射,2种仪器均放置于培养池的几何中心,垂直于水面0.25 m处,每1 h记录1次数据。蓝藻接种完成后,从第0天开始,每隔1 d从培养池前中后位置均匀舀取1 L蓝藻液,将样品及时带回实验室。静置1 h后,用水质测试针(86505,中国台湾衡欣)测上清液的pH和电导率。然后将所有蓝藻液置于80 ℃烘箱中烘干至恒重,取出后用千分之一电子天平称质量。培养结束后,关闭扰动装置,静置12 h后收取蓝藻。蓝藻收取完成后,先测定每个培养池的蓝藻总鲜重,然后均匀地从中吸取鲜藻20 mL于西林瓶中,在80 ℃烘箱中烘干至恒重,取出后称取干重,并计算出每个培养池蓝藻总干重。
1.4 数据处理及统计分析
由于没有关于荒漠蓝藻生长下限温度的相关报道,因此以农作物生长的起始温度为参考,本研究中荒漠蓝藻生物学下限温度取值10 ℃[16 ] 。日均温度、日均光照强度和日均光合有效辐射采用每日07:00—19:00数据平均值,累积光照强度和累积光合有效辐射为逐日日均光照强度和日均光合有效辐射的累积值,有效积温的计算表达式如下所示:
K = ∑ P M ( T - T 1 ) (1)
式中:P 为开始时间;M 为结束时间;T 为日平均温度;T 1 为下限温度;K 为有效积温。
采用SPSS 21.0(IBM Corp., Armonk, 美国)统计软件进行单因素方差分析和Duncan's多重比较分析在3种扰动时长下荒漠蓝藻在整个培养周期的干重、pH、电导率、温度、光照强度、光合有效辐射以及培养末期总质量的差异显著性。根据R 2 最大原则选取最优拟合方程,分别使用线性、二项式、指数、幂函数等方程建立荒漠蓝藻生长特征与扰动时长、温度、光照强度、光合有效辐射、pH和电导率的回归模型。
2 结果与分析
2.1 荒漠蓝藻培养过程中温度、光照强度和光合有效辐射的变化特征
3种扰动时长下温度、光照强度和光合有效辐射随时间逐渐下降(表1 )。3、6、12 h扰动下温度平均值在第3天上升达到最大值,平均值分别为21.83、21.73、21.77 ℃,随后逐渐降低。与6 h扰动相比,3 h和12 h下的光照强度和光合有效辐射分别随时间变化存在显著(P <0.05)和极显著差异(P <0.001)。
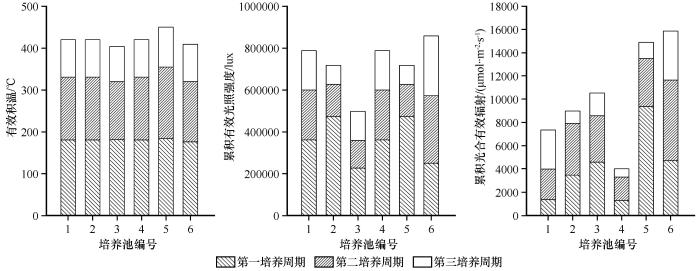
6个蓝藻培养池在3个培养周期中对应的有效积温、累积光照强度和累积光合有效辐射分别为80—185 ℃、90 000—480 000 lux和715—9 400 μmol·m-2 ·s-1
图2
图2
3个培养周期中不同蓝藻培养池对应的有效积温、累积光照强度和累积光合有效辐射
Fig.2
Effective accumulated temperature, accumulated light intensity and accumulated photosynthetically active radiation corresponding to 6 cyanobacteria culture pools in 3 cycles
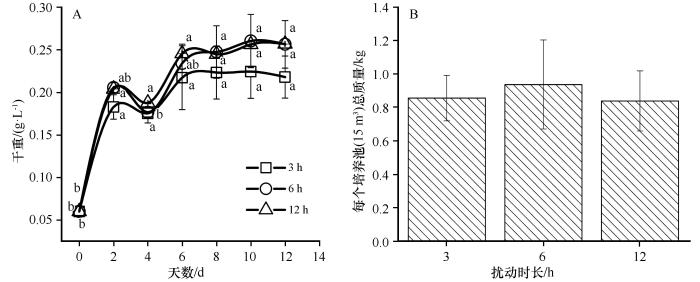
2.2 扰动时长对荒漠蓝藻日生长特征和总质量的影响
荒漠蓝藻在3种扰动时长下干重在0—2 d迅速增加,在2—4 d短暂下降后又在4—6 d小幅度增加,之后趋于稳定(图3 A)。荒漠蓝藻的总质量随扰动时长的增加先增加后减小,在3、6 h和12 h扰动下每个培养池(15 m3 )总质量分别为0.858、0.940、0.841 kg(图3 B)。
图3
图3
3种扰动时长下荒漠蓝藻生长特征变化(平均值±标准误)
不同小写字母表示3种处理随时间变化的差异显著(P <0.05)
Fig.3
Changes in the growth characteristics of desert cyanobacteria in three disturbance duration(Mean ± SE )
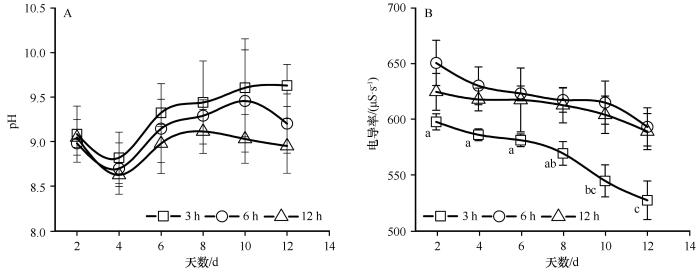
2.3 荒漠蓝藻培养过程中pH 和电导率变化
荒漠蓝藻在3种扰动时长下pH呈先下降后上升的趋势(图4 A),在3 h扰动下,第4天pH由9.08下降至8.82后,开始逐渐升高至9.63。在整个培养周期,电导率均呈持续下降的趋势,在3 h扰动下电导率随时间变化存在极显著差异(P <0.001;图4 B)。
图4
图4
3种扰动时长下pH和电导率随时间变化(平均值±标准误)
不同小写字母表示3种处理随时间变化的差异显著(P < 0.05)
Fig.4
Changes in pH and conductivity in three disturbance duration (Mean ± SE )
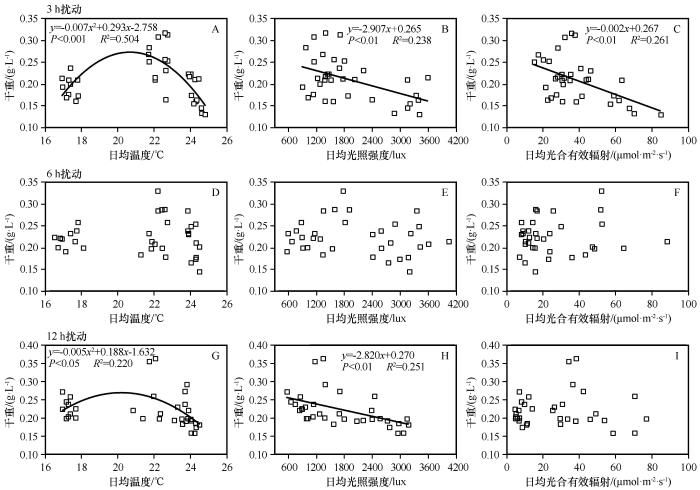
2.4 荒漠蓝藻生长特征与环境因子回归分析
3 h扰动下荒漠蓝藻干重与日均温度、日均光照强度和日均光合有效辐射分别呈极显著二项式相关(y =-0.007x 2 +0.293x -2.758, P <0.001, R 2 =0.504)、显著线性相关(y =-2.907x +0.265, P <0.01, R 2 =0.238)和显著线性相关关系(y =-0.002x +0.267, P <0.01, R 2 =0.261;图5 A,图5 B,图5 C)。12 h扰动下干重与日均温度、日均光照强度和日均光合有效辐射分别呈显著二项式相关(y =-0.005x 2 +0.188x -1.632, P <0.05, R 2 =0.220)、显著线性相关关系(y =-2.820x +0.270, P <0.01, R 2 =0.251)和不相关(图5 G,图5 H,图5 I)。
图5
图5
3种扰动时长下荒漠蓝藻干重与日均温度、日均光照强度和日均光合有效辐射的关系
图A、B和C表示3 h扰动下的相应关系,图D、E和F表示6 h扰动下的相应关系,图G、H和F表示12 h扰动下的相应关系
Fig.5
Relationship between the dry weight of desert cyanobacteria and the average daily temperature, average daily light intensity and average daily photosynthetically active radiation in three disturbance duration
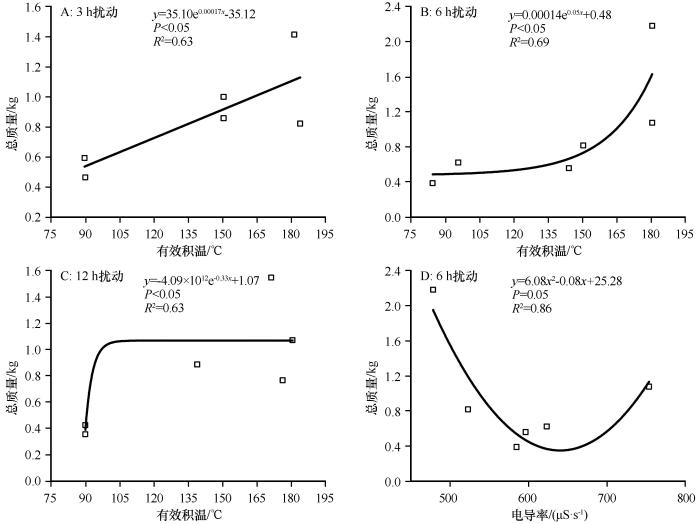
在3、6 h和12 h扰动下荒漠蓝藻总质量与有效积温分别呈显著指数相关(P <0.05,图6 )。6 h扰动的总质量与电导率呈显著二项式相关(y =6.08x 2 -0.08x +25.28,P =0.05,R 2 =0.86)。
图6
图6
3种扰动时长下荒漠蓝藻总质量(15 m3 培养池)与有效积温和电导率的关系
Fig.6
Relationship between the total mass of desert cyanobacteria and the effective accumulated temperature and conductivity in three disturbance duration
3 讨论
扰动能够影响水体中悬浮物浓度,改变水体的光照条件及促进N、P等营养盐的传递和转化速率,最终决定藻类初级生产力[26 ] 。本研究中,荒漠蓝藻在3种扰动时长下干重呈现出先迅速增加,之后趋于稳定的趋势。在3、6、12 h扰动下荒漠蓝藻的总质量分别为每培养池(15 m3 )0.858、0.940、0.841 kg。在3 h扰动下,荒漠蓝藻在培养后期表现为生长滞后且总产量较少,这可能由于3 h扰动下形成的长时间静水状态会使荒漠蓝藻更易大量沉积在底部或粘结在池壁上,降低了其光合作用速率和酶活性[15 ] 。蓝藻进行光暗反应需要一个较为稳定的环境,在12 h扰动下荒漠蓝藻的总质量最少,原因可能是12 h扰动下藻类和水体中其他浮游生物和颗粒物质长时间悬浮并相互摩擦,难以聚集生长,形成的高浊度水还会显著降低入射光强和光合有效辐射,从而破坏藻类生长原有的稳定环境[27 ] 。此外,12 h扰动下的长时间水流冲刷作用还会对藻类产生机械剪切,直接损坏藻类细胞结构的完整性[28 -29 ] 。生长后期停滞、低光合速率、环境的破坏以及机械损伤可能是本研究中3 h和12 h扰动下总质量低的重要原因。而6 h扰动下荒漠蓝藻的总质量最高,这可能因为对水体扰动的适当减弱既可以为蓝藻的光合作用提供充足的CO2 ,又不会破坏藻细胞,还能使整个培养过程中水体保持稳定的温度、光照强度和光合有效辐射,增大了水体环境的稳定性,有利于接入藻种的生长,从而获得的总质量最高。本文的研究结果与李林等[30 ] 研究中表明间歇性扰动条件下微囊藻(Microcystis aeruginosa )密度较连续性水流条件下高的结果类似;Karp-Boss等[31 ] 认为,连续扰动会对鱼腥藻(Anabaena aquae )细胞产生机械破坏,而间歇性扰动会促进其生长。
温度和光照强度同样会影响藻类的生长,高于或低于最佳温度和光照强度都会对蓝藻生长产生抑制作用,包括阻碍蛋白质的合成、降低光合作用生长速率、加快酶的失活以及阻碍细胞膜的渗透[32 -34 ] 。本研究结果显示,在3、12 h扰动下荒漠蓝藻的干重会受到日均温度、日均光照强度和日均光合有效辐射不同程度的制约,在这2种扰动时长下需保持适中温度(22—23 ℃)、低光照强度(500—1 500 lux)和低光合有效辐射(15—40 μmol·m-2 ·s-1 )才适宜其生长。谢作明等[20 ] 研究发现在温室中的开放式循环培养池培养荒漠蓝藻时应采用遮阳网控制光照强度在 600—700 μE·m-2 ·s-1 ,并将温度保持在35 ℃以下。此外,雷亚萍[34 ] 研究发现辐射强度越高对爪哇伪枝藻的光合活性、叶绿素a、膜结构和蛋白质结构的损伤也越严重,当光强从20 μE·m-2 ·s-1 转到 100 μE·m-2 ·s-1 时,藻体的最大相对电子传递速率和饱和光照强度明显降低。
蓝藻的生长发育是一个热量的累积过程[16 ] 。本文对荒漠蓝藻总质量与有效积温进行研究发现,荒漠蓝藻总质量与有效积温呈显著正相关,较低的积温对蓝藻生长发育有抑制作用。在9—10月,温室条件下荒漠蓝藻完成其生长发育所需有效积温为80—185 ℃,蓝藻总质量的增长速率随积温的增加逐渐增加,快速增长转折点对应的有效积温为145 ℃,在有效积温为180 ℃时,其生物量达到最大值。朱广伟等[18 ] 在分析2005—2017年5—7月太湖北部水体蓝藻叶绿素a浓度的影响因素时,发现积温在调节太湖叶绿素a浓度年际变化过程中发挥正向主导作用。谢小萍等[35 ] 试验发现在太湖蓝藻复苏期,水中的叶绿素a浓度随着有效积温的变化而变化,有效积温越高,水体中蓝藻生物量越高。李星[36 ] 研究发现,积温过低不利于大量蓝藻的产生,黄岐附近水体在积温达到2 300 ℃时有利于加快蓝藻的生长。关于蓝藻生长与积温之间的关系也有不同的结果,北方内陆寒冷地区碱性水体中大量蓝藻的形成相比于热带和亚热带地区需要的有效积温较低,蓝藻爆发与有效积温负相关[17 ] 。范裕祥等[37 ] 通过对巢湖蓝藻暴发期间气象观测资料的分析,发现蓝藻发生面积与前5 d积温呈高度显著的线性负相关,持续的高温对蓝藻积聚有明显的抑制作用,5 d积温 165 ℃·d 可作为蓝藻积聚的上限。
本研究结果显示,在3种扰动时长下水体的pH均升高至9左右,这可能由于蓝藻在生长过程中消耗H2 CO3 所导致[33 ] 。在本试验期间,随着时间的推移,水体的电导率逐渐降低,可能由于藻种加入水体中,经过一段时间,开始吸收利用水体中的营养盐来进行自身的生长繁殖,从而使水体中N、P等营养盐降低[33 ] 。其中,3 h扰动下水体营养盐消耗剧烈,而6 h扰动下营养盐消耗较为平缓,可以维持稳定的营养盐水平来保证藻细胞的充分生长和分裂。在6 h扰动下,荒漠蓝藻生长末期水体的电导率为470—630 μS·s-1 ,其中在470—500 μS·s-1 下对应的总质量较高,表明荒漠蓝藻适宜生长在低电导率水环境中。这与前人研究中盐胁迫会抑制荒漠蓝藻的生长,其生物量会随盐浓度的升高而下降的结论相似[21 -22 ] 。
综上所述,在3种扰动时长下荒漠蓝藻的生长呈现出骤然增长,然后维持在成熟期。其中,3 h扰动下荒漠蓝藻在培养后期生长出现延滞现象。荒漠蓝藻的生物量在6 h扰动下的总质量最高,为每培养池(15 m3 )0.940 kg。日均温度、日均光照强度、日均光合有效辐射和有效积温显著影响荒漠蓝藻的生长和产量。根据荒漠蓝藻总产量、生长过程和耗能等多方面综合考虑,荒漠蓝藻规模化培养的最佳扰动时长为6 h。积温对荒漠蓝藻大量生长发挥关键作用,建议在荒漠蓝藻生长过程中,有效积温应不低于145 ℃。
参考文献
View Option
[1]
West N E Structure and function of microphytic soil crusts in wildland ecosystems of arid to semi-arid regions
[J].Advances in Ecological Research ,1990 ,20 :179 -223 .
[本文引用: 1]
[2]
Rodriguez-Caballero E Belnap J Budel B et al Dryland photoautotrophic soil surface communities endangered by global change
[J].Nature Geoscience ,2008 ,11 :185 -189 .
[本文引用: 1]
[3]
李新荣 ,回嵘 ,赵洋 中国荒漠生物土壤结皮生态生理学研究 [M].北京 :高等教育出版社 ,2016 :20 -25 .
[本文引用: 4]
[4]
Xiao B Hu K Moss-dominated biocrusts decrease soil moisture and result in the degradation of artificially planted shrubs under semiarid climate
[J].Geoderma ,2017 ,291 :47 -54 .
[本文引用: 1]
[5]
Zhao Y Wang N Zhang Z S et al Accelerating the development of artificial biocrusts using covers for restoration of degraded land in dryland ecosystem
[J].Land Degradation and Development ,2020 ,32 :1 -11 .
[本文引用: 2]
[6]
赵洋 ,潘颜霞 ,苏洁琼 ,等 中国干旱区沙化土地绿色环保治理技术综述
[J].中国沙漠 ,2021 ,41 (1 ):195 -202 .
[本文引用: 1]
[7]
Xiao B Hu K L Veste M et al Natural recovery rates of moss biocrusts after severe disturbance in a semiarid climate of the Chinese Loess Plateau
[J].Geoderma ,2019 ,337 :402 -412 .
[本文引用: 1]
[8]
刘永定 ,陈兰周 ,胡春香 荒漠蓝藻环境生物学与生物土壤结皮固沙 [M].北京 :科学出版 ,2013 :231 -242 .
[本文引用: 1]
[9]
Giraldo-Silva A Nelson C Barger N N et al Nursing biocrusts:isolation,cultivation,and fitness test of indigenous cyanobacteria
[J].Restoration Ecology ,2019 ,27 :793 -803 .
[10]
D'Acqui L P Use of indigenous cyanobacteria for sustainable improvement of biogeochemical and physical fertility of marginal soils in semiarid tropics
[M]// Arora N K,Mehnaz S,Balestrini R,eds.Bioformulations:For Sustainable Agriculture .New Delhi,India :Springer ,2016 :213 -232 .
[本文引用: 1]
[11]
潘雯雯 ,杨桂军 ,芮政 ,等 野外模拟扰动方式对太湖浮游植物群落结构的影响
[J].环境科学研究 ,2020 ,33 (6 ):1421 -1430 .
[本文引用: 1]
[12]
Sullivan J M Swift E Effects of small-scale turbulence on net growth rate and size of ten species of marine dinoflagellates
[J].Journal of Phycology ,2010 ,39 :83 -94 .
[本文引用: 1]
[13]
张文慧 水动力对太湖营养盐循环及藻类生长的影响
[D].北京 :中国环境科学研究院 ,2017 .
[本文引用: 1]
[14]
Xiao Y Li Z Li C et al Effect of small-scale turbulence on the physiology and morphology of two bloom-forming cyanobacteria
[J].PloS One ,2016 ,11 :1 -14 .
[本文引用: 1]
[15]
王文超 ,徐绍峰 ,徐梦瑶 ,等 水体扰动对铜绿微囊藻生长和酶活性的影响
[J].军事医学 ,2019 ,43 (6 ):448 -453 .
[本文引用: 2]
[16]
齐尚红 ,王冰洁 ,武作书 农业生产与温度的关系
[J].河南科技学院学报 ,2007 ,35 (4 ):20 -23 .
[本文引用: 4]
[17]
潘翰 ,刘琰冉 ,马成学 北方寒冷内陆碱性水体蓝藻暴发特点及影响因子
[J].东北林业大学学报 ,2017 ,45 (7 ):79 -83 .
[本文引用: 4]
[18]
朱广伟 ,钟春妮 ,秦伯强 ,等 2005-2017年北部太湖水体叶绿素a和营养盐变化及影响因素
[J].湖泊科学 ,2018 ,30 (2 ):279 -295 .
[本文引用: 2]
[19]
孙祥 水文气象对天目湖沙河水库藻类群落结构动态变化的影响
[D].安徽芜湖 :安徽师范大学 ,2018 .
[本文引用: 1]
[20]
谢作明 ,刘永定 ,陈兰洲 ,等 不同培养条件对具鞘微鞘藻生物量和多糖产量的影响
[J].水生生物学报 ,2008 ,32 (2 ):272 -275 .
[本文引用: 2]
[21]
陈兰周 ,刘永定 ,李敦海 盐胁迫对爪哇伪枝藻(Scytonema javanicum )生理生化特性的影响
[J].中国沙漠 ,2003 ,23 (3 ):79 -82 .
[本文引用: 1]
[22]
Chen L Z Li D H Song L R et al Effects of salt stress on carbohydrate metabolism in desert soil alga Microcoleus vaginatus Gom
[J].Journal of Integrative Plant Biology ,2006 ,8 :914 -919 .
[本文引用: 2]
[23]
Taton A Stana G Brambilla E et al Cyanobacterial diversity in natural and artificial microbial mats of Lake Fryxell (McMurdo Dry Valleys,Antarctica):a morphological and molecular approach
[J].Applied and Environmental Microbiology ,2003 ,69 :5157 -5169 .
[本文引用: 1]
[24]
Park C H Li X R Jia R L et al Effects of superabsorbent polymer on cyanobacterial biological soil crust formation in laboratory
[J].Arid Soil Research and Rehabilitation ,2014 ,29 :55 -71 .
[本文引用: 1]
[25]
Rippka R Deruelles J Waterbury J B et al Generic assignments,strain histories and properties of pure cultures of cyanobacteria
[J].Journal of General Microbiology ,1979 ,111 :1 -61 .
[本文引用: 1]
[26]
Li Zhe Xiao Y Yang J et al Response of cellular stoichiometry and phosphorus storage of the cyanobacteria Aphanizomenon flos-aquae to small-scale turbulence
[J].Chinese Journal of Oceanology and Limnology ,2017 ,35 :1409 -1416 .
[本文引用: 1]
[27]
任杰 ,朱广伟 ,金颖薇 换水率和营养水平对太湖流域横山水库硅藻水华的影响
[J].湖泊科学 ,2017 ,29 (3 ):604 -616 .
[本文引用: 1]
[28]
Kinsman R Ibelings B W Walsby A E Gas vesicle collapse by turgor pressure and its role in buoyancy regulation by Anabaena flosaquae
[J].Journal of General Microbiology ,1991 ,137 :1171 -1178 .
[本文引用: 1]
[29]
江林燕 ,江成 ,周伟 ,等 水体扰动对铜绿微囊藻生长影响的规律及原因
[J].环境化学 ,2012 ,31 (2 ):216 -220 .
[本文引用: 1]
[30]
李林 ,朱伟 连续水流和间歇水流对微囊藻生长的影响
[J].环境科学与技术 ,2012 ,35 (10 ):34 -37 .
[本文引用: 1]
[31]
Karp-Boss L Boss E Jumars P A Nutrient fluxes to planktonic osmotrophs in the presence of fluid motion
[J].Oceanography and Marine Biology ,1996 ,34 :71 -107 .
[本文引用: 1]
[32]
颜润润 ,逄勇 ,陈晓峰 ,等 不同风等级扰动对贫富营养下铜绿微囊藻生长的影响
[J].环境科学 ,2008 ,29 (10 ):63 -67 .
[本文引用: 1]
[33]
赵秀侠 ,方婷 ,杨坤 ,等 安徽沱湖夏季浮游植物群落结构特征与环境因子关系
[J].植物科学学报 ,2018 ,36 (5 ):687 -695 .
[本文引用: 2]
[34]
雷亚萍 共生关系对爪哇伪枝藻抗荒漠胁迫能力的影响
[D].武汉 :武汉理工大学 ,2017 .
[本文引用: 2]
[35]
谢小萍 ,李亚春 ,杭鑫 ,等 气温对太湖蓝藻复苏和休眠进程的影响
[J].湖泊科学 ,2016 ,28 (4 ):818 -824 .
[本文引用: 1]
[36]
李星 海表温度对连江黄岐赤潮影响的研究
[J].海洋预报 ,2021 ,38 (3 ):98 -103 .
[本文引用: 1]
[37]
范裕祥 ,金社军 ,周培 ,等 巢湖蓝藻水华分布特征和气象条件分析
[J].安徽农业科学 ,2015 ,43 (4 ):191 -193 .
[本文引用: 1]
Structure and function of microphytic soil crusts in wildland ecosystems of arid to semi-arid regions
1
1990
... 生物土壤结皮(Biological Soil Crusts,BSCs)是由蓝藻、绿藻、地衣、藓类和异养微生物及相关的其他生物体与土壤表层颗粒等非生物体胶结形成的复杂复合体,约占全球陆地面积的12.2%[1 -2 ] .BSCs具有稳定土壤,增加C、N固定,调节土壤水文过程及提高生物多样性等功能,被誉为“荒漠生态系统工程师”[3 -4 ] .近年来,BSCs在固沙方面的作用得到了广泛认可,其形成和发育可以降低荒漠生态系统非生物因子的胁迫水平,从而维持系统的健康[5 ] .但BSCs自然形成需要几十年,人工培育方法可以缩短BSCs的形成时间,并迅速对沙化土壤进行固定,从而改善土壤生境,成为沙化土地治理的新模式[6 -7 ] .荒漠蓝藻作为荒漠、半荒漠生态系统的先行者,是BSCs形成初期的核心组分,能够在恶劣的生境条件(干旱、营养贫瘠、强紫外线辐射等)下生存繁衍,而且比地衣和藓类,容易培养,从而成为人工培育BSCs的首选理想物种[3 ] .然而,足量的荒漠蓝藻是使用该技术进行大规模沙化土地固定的基础,显然,荒漠蓝藻的规模化培养是该技术推广和应用的重要前提和保障.但以往的研究中关于荒漠蓝藻培养的容器较小,无法满足大规模应用的需求,因此亟待系统地开展荒漠蓝藻规模化培养研究. ...
Dryland photoautotrophic soil surface communities endangered by global change
1
2008
... 生物土壤结皮(Biological Soil Crusts,BSCs)是由蓝藻、绿藻、地衣、藓类和异养微生物及相关的其他生物体与土壤表层颗粒等非生物体胶结形成的复杂复合体,约占全球陆地面积的12.2%[1 -2 ] .BSCs具有稳定土壤,增加C、N固定,调节土壤水文过程及提高生物多样性等功能,被誉为“荒漠生态系统工程师”[3 -4 ] .近年来,BSCs在固沙方面的作用得到了广泛认可,其形成和发育可以降低荒漠生态系统非生物因子的胁迫水平,从而维持系统的健康[5 ] .但BSCs自然形成需要几十年,人工培育方法可以缩短BSCs的形成时间,并迅速对沙化土壤进行固定,从而改善土壤生境,成为沙化土地治理的新模式[6 -7 ] .荒漠蓝藻作为荒漠、半荒漠生态系统的先行者,是BSCs形成初期的核心组分,能够在恶劣的生境条件(干旱、营养贫瘠、强紫外线辐射等)下生存繁衍,而且比地衣和藓类,容易培养,从而成为人工培育BSCs的首选理想物种[3 ] .然而,足量的荒漠蓝藻是使用该技术进行大规模沙化土地固定的基础,显然,荒漠蓝藻的规模化培养是该技术推广和应用的重要前提和保障.但以往的研究中关于荒漠蓝藻培养的容器较小,无法满足大规模应用的需求,因此亟待系统地开展荒漠蓝藻规模化培养研究. ...
4
2016
... 生物土壤结皮(Biological Soil Crusts,BSCs)是由蓝藻、绿藻、地衣、藓类和异养微生物及相关的其他生物体与土壤表层颗粒等非生物体胶结形成的复杂复合体,约占全球陆地面积的12.2%[1 -2 ] .BSCs具有稳定土壤,增加C、N固定,调节土壤水文过程及提高生物多样性等功能,被誉为“荒漠生态系统工程师”[3 -4 ] .近年来,BSCs在固沙方面的作用得到了广泛认可,其形成和发育可以降低荒漠生态系统非生物因子的胁迫水平,从而维持系统的健康[5 ] .但BSCs自然形成需要几十年,人工培育方法可以缩短BSCs的形成时间,并迅速对沙化土壤进行固定,从而改善土壤生境,成为沙化土地治理的新模式[6 -7 ] .荒漠蓝藻作为荒漠、半荒漠生态系统的先行者,是BSCs形成初期的核心组分,能够在恶劣的生境条件(干旱、营养贫瘠、强紫外线辐射等)下生存繁衍,而且比地衣和藓类,容易培养,从而成为人工培育BSCs的首选理想物种[3 ] .然而,足量的荒漠蓝藻是使用该技术进行大规模沙化土地固定的基础,显然,荒漠蓝藻的规模化培养是该技术推广和应用的重要前提和保障.但以往的研究中关于荒漠蓝藻培养的容器较小,无法满足大规模应用的需求,因此亟待系统地开展荒漠蓝藻规模化培养研究. ...
... [3 ].然而,足量的荒漠蓝藻是使用该技术进行大规模沙化土地固定的基础,显然,荒漠蓝藻的规模化培养是该技术推广和应用的重要前提和保障.但以往的研究中关于荒漠蓝藻培养的容器较小,无法满足大规模应用的需求,因此亟待系统地开展荒漠蓝藻规模化培养研究. ...
... 鱼腥藻(Anabaena cylindrica )、具鞘微鞘藻(Microcoleus vaginatus )、念珠藻(Nostoc carneum )、席藻(Phormidium sp. NIES-2121)、伪枝藻(Scytonema sp. NIES-4073)、爪哇伪枝藻(Scytonema javanicum )和单歧藻(Tolypothrix tenuis )是腾格里沙漠蓝藻结皮中的优势物种[3 ,5 ] .鉴于混合荒漠蓝藻在人工BSCs培育中的广泛应用[3 ] ,本研究将上述7种蓝藻组成的混合藻种作为研究对象,研究了规模化培养时荒漠蓝藻的生长特征,探讨了扰动时长、温度、光照强度和光合有效辐射对蓝藻生长的影响,并引入农业气象学中有效积温、累积光照强度和累积光合有效辐射探究它们与荒漠蓝藻产量间的关系,将为荒漠蓝藻的规模化培养提供理论依据. ...
... [3 ],本研究将上述7种蓝藻组成的混合藻种作为研究对象,研究了规模化培养时荒漠蓝藻的生长特征,探讨了扰动时长、温度、光照强度和光合有效辐射对蓝藻生长的影响,并引入农业气象学中有效积温、累积光照强度和累积光合有效辐射探究它们与荒漠蓝藻产量间的关系,将为荒漠蓝藻的规模化培养提供理论依据. ...
Moss-dominated biocrusts decrease soil moisture and result in the degradation of artificially planted shrubs under semiarid climate
1
2017
... 生物土壤结皮(Biological Soil Crusts,BSCs)是由蓝藻、绿藻、地衣、藓类和异养微生物及相关的其他生物体与土壤表层颗粒等非生物体胶结形成的复杂复合体,约占全球陆地面积的12.2%[1 -2 ] .BSCs具有稳定土壤,增加C、N固定,调节土壤水文过程及提高生物多样性等功能,被誉为“荒漠生态系统工程师”[3 -4 ] .近年来,BSCs在固沙方面的作用得到了广泛认可,其形成和发育可以降低荒漠生态系统非生物因子的胁迫水平,从而维持系统的健康[5 ] .但BSCs自然形成需要几十年,人工培育方法可以缩短BSCs的形成时间,并迅速对沙化土壤进行固定,从而改善土壤生境,成为沙化土地治理的新模式[6 -7 ] .荒漠蓝藻作为荒漠、半荒漠生态系统的先行者,是BSCs形成初期的核心组分,能够在恶劣的生境条件(干旱、营养贫瘠、强紫外线辐射等)下生存繁衍,而且比地衣和藓类,容易培养,从而成为人工培育BSCs的首选理想物种[3 ] .然而,足量的荒漠蓝藻是使用该技术进行大规模沙化土地固定的基础,显然,荒漠蓝藻的规模化培养是该技术推广和应用的重要前提和保障.但以往的研究中关于荒漠蓝藻培养的容器较小,无法满足大规模应用的需求,因此亟待系统地开展荒漠蓝藻规模化培养研究. ...
Accelerating the development of artificial biocrusts using covers for restoration of degraded land in dryland ecosystem
2
2020
... 生物土壤结皮(Biological Soil Crusts,BSCs)是由蓝藻、绿藻、地衣、藓类和异养微生物及相关的其他生物体与土壤表层颗粒等非生物体胶结形成的复杂复合体,约占全球陆地面积的12.2%[1 -2 ] .BSCs具有稳定土壤,增加C、N固定,调节土壤水文过程及提高生物多样性等功能,被誉为“荒漠生态系统工程师”[3 -4 ] .近年来,BSCs在固沙方面的作用得到了广泛认可,其形成和发育可以降低荒漠生态系统非生物因子的胁迫水平,从而维持系统的健康[5 ] .但BSCs自然形成需要几十年,人工培育方法可以缩短BSCs的形成时间,并迅速对沙化土壤进行固定,从而改善土壤生境,成为沙化土地治理的新模式[6 -7 ] .荒漠蓝藻作为荒漠、半荒漠生态系统的先行者,是BSCs形成初期的核心组分,能够在恶劣的生境条件(干旱、营养贫瘠、强紫外线辐射等)下生存繁衍,而且比地衣和藓类,容易培养,从而成为人工培育BSCs的首选理想物种[3 ] .然而,足量的荒漠蓝藻是使用该技术进行大规模沙化土地固定的基础,显然,荒漠蓝藻的规模化培养是该技术推广和应用的重要前提和保障.但以往的研究中关于荒漠蓝藻培养的容器较小,无法满足大规模应用的需求,因此亟待系统地开展荒漠蓝藻规模化培养研究. ...
... 鱼腥藻(Anabaena cylindrica )、具鞘微鞘藻(Microcoleus vaginatus )、念珠藻(Nostoc carneum )、席藻(Phormidium sp. NIES-2121)、伪枝藻(Scytonema sp. NIES-4073)、爪哇伪枝藻(Scytonema javanicum )和单歧藻(Tolypothrix tenuis )是腾格里沙漠蓝藻结皮中的优势物种[3 ,5 ] .鉴于混合荒漠蓝藻在人工BSCs培育中的广泛应用[3 ] ,本研究将上述7种蓝藻组成的混合藻种作为研究对象,研究了规模化培养时荒漠蓝藻的生长特征,探讨了扰动时长、温度、光照强度和光合有效辐射对蓝藻生长的影响,并引入农业气象学中有效积温、累积光照强度和累积光合有效辐射探究它们与荒漠蓝藻产量间的关系,将为荒漠蓝藻的规模化培养提供理论依据. ...
中国干旱区沙化土地绿色环保治理技术综述
1
2021
... 生物土壤结皮(Biological Soil Crusts,BSCs)是由蓝藻、绿藻、地衣、藓类和异养微生物及相关的其他生物体与土壤表层颗粒等非生物体胶结形成的复杂复合体,约占全球陆地面积的12.2%[1 -2 ] .BSCs具有稳定土壤,增加C、N固定,调节土壤水文过程及提高生物多样性等功能,被誉为“荒漠生态系统工程师”[3 -4 ] .近年来,BSCs在固沙方面的作用得到了广泛认可,其形成和发育可以降低荒漠生态系统非生物因子的胁迫水平,从而维持系统的健康[5 ] .但BSCs自然形成需要几十年,人工培育方法可以缩短BSCs的形成时间,并迅速对沙化土壤进行固定,从而改善土壤生境,成为沙化土地治理的新模式[6 -7 ] .荒漠蓝藻作为荒漠、半荒漠生态系统的先行者,是BSCs形成初期的核心组分,能够在恶劣的生境条件(干旱、营养贫瘠、强紫外线辐射等)下生存繁衍,而且比地衣和藓类,容易培养,从而成为人工培育BSCs的首选理想物种[3 ] .然而,足量的荒漠蓝藻是使用该技术进行大规模沙化土地固定的基础,显然,荒漠蓝藻的规模化培养是该技术推广和应用的重要前提和保障.但以往的研究中关于荒漠蓝藻培养的容器较小,无法满足大规模应用的需求,因此亟待系统地开展荒漠蓝藻规模化培养研究. ...
Natural recovery rates of moss biocrusts after severe disturbance in a semiarid climate of the Chinese Loess Plateau
1
2019
... 生物土壤结皮(Biological Soil Crusts,BSCs)是由蓝藻、绿藻、地衣、藓类和异养微生物及相关的其他生物体与土壤表层颗粒等非生物体胶结形成的复杂复合体,约占全球陆地面积的12.2%[1 -2 ] .BSCs具有稳定土壤,增加C、N固定,调节土壤水文过程及提高生物多样性等功能,被誉为“荒漠生态系统工程师”[3 -4 ] .近年来,BSCs在固沙方面的作用得到了广泛认可,其形成和发育可以降低荒漠生态系统非生物因子的胁迫水平,从而维持系统的健康[5 ] .但BSCs自然形成需要几十年,人工培育方法可以缩短BSCs的形成时间,并迅速对沙化土壤进行固定,从而改善土壤生境,成为沙化土地治理的新模式[6 -7 ] .荒漠蓝藻作为荒漠、半荒漠生态系统的先行者,是BSCs形成初期的核心组分,能够在恶劣的生境条件(干旱、营养贫瘠、强紫外线辐射等)下生存繁衍,而且比地衣和藓类,容易培养,从而成为人工培育BSCs的首选理想物种[3 ] .然而,足量的荒漠蓝藻是使用该技术进行大规模沙化土地固定的基础,显然,荒漠蓝藻的规模化培养是该技术推广和应用的重要前提和保障.但以往的研究中关于荒漠蓝藻培养的容器较小,无法满足大规模应用的需求,因此亟待系统地开展荒漠蓝藻规模化培养研究. ...
1
2013
... 荒漠蓝藻的生长和产量除受到温度、光照、营养盐等环境条件的影响外,也会受到不同时间尺度上扰动过程的控制[8 -10 ] .水体扰动能够通过增加悬浮物浓度从而影响水体光照条件,进而影响藻类的光合效率 [11 ] .不同扰动方式下藻类的敏感度和耐受能力不同,其营养盐吸收速率也会表现出明显差异[12 -13 ] .此外,较强的扰动条件会破坏藻细胞的结构、造成细胞的机械损伤、改变其生物量和沉降量等[14 ] .目前,水体扰动对蓝藻生长影响的研究主要在水生/水华蓝藻方面,对不同扰动时长下荒漠蓝藻生长状况有待进一步研究[15 ] . ...
Nursing biocrusts:isolation,cultivation,and fitness test of indigenous cyanobacteria
0
2019
Use of indigenous cyanobacteria for sustainable improvement of biogeochemical and physical fertility of marginal soils in semiarid tropics
1
2016
... 荒漠蓝藻的生长和产量除受到温度、光照、营养盐等环境条件的影响外,也会受到不同时间尺度上扰动过程的控制[8 -10 ] .水体扰动能够通过增加悬浮物浓度从而影响水体光照条件,进而影响藻类的光合效率 [11 ] .不同扰动方式下藻类的敏感度和耐受能力不同,其营养盐吸收速率也会表现出明显差异[12 -13 ] .此外,较强的扰动条件会破坏藻细胞的结构、造成细胞的机械损伤、改变其生物量和沉降量等[14 ] .目前,水体扰动对蓝藻生长影响的研究主要在水生/水华蓝藻方面,对不同扰动时长下荒漠蓝藻生长状况有待进一步研究[15 ] . ...
野外模拟扰动方式对太湖浮游植物群落结构的影响
1
2020
... 荒漠蓝藻的生长和产量除受到温度、光照、营养盐等环境条件的影响外,也会受到不同时间尺度上扰动过程的控制[8 -10 ] .水体扰动能够通过增加悬浮物浓度从而影响水体光照条件,进而影响藻类的光合效率 [11 ] .不同扰动方式下藻类的敏感度和耐受能力不同,其营养盐吸收速率也会表现出明显差异[12 -13 ] .此外,较强的扰动条件会破坏藻细胞的结构、造成细胞的机械损伤、改变其生物量和沉降量等[14 ] .目前,水体扰动对蓝藻生长影响的研究主要在水生/水华蓝藻方面,对不同扰动时长下荒漠蓝藻生长状况有待进一步研究[15 ] . ...
Effects of small-scale turbulence on net growth rate and size of ten species of marine dinoflagellates
1
2010
... 荒漠蓝藻的生长和产量除受到温度、光照、营养盐等环境条件的影响外,也会受到不同时间尺度上扰动过程的控制[8 -10 ] .水体扰动能够通过增加悬浮物浓度从而影响水体光照条件,进而影响藻类的光合效率 [11 ] .不同扰动方式下藻类的敏感度和耐受能力不同,其营养盐吸收速率也会表现出明显差异[12 -13 ] .此外,较强的扰动条件会破坏藻细胞的结构、造成细胞的机械损伤、改变其生物量和沉降量等[14 ] .目前,水体扰动对蓝藻生长影响的研究主要在水生/水华蓝藻方面,对不同扰动时长下荒漠蓝藻生长状况有待进一步研究[15 ] . ...
水动力对太湖营养盐循环及藻类生长的影响
1
2017
... 荒漠蓝藻的生长和产量除受到温度、光照、营养盐等环境条件的影响外,也会受到不同时间尺度上扰动过程的控制[8 -10 ] .水体扰动能够通过增加悬浮物浓度从而影响水体光照条件,进而影响藻类的光合效率 [11 ] .不同扰动方式下藻类的敏感度和耐受能力不同,其营养盐吸收速率也会表现出明显差异[12 -13 ] .此外,较强的扰动条件会破坏藻细胞的结构、造成细胞的机械损伤、改变其生物量和沉降量等[14 ] .目前,水体扰动对蓝藻生长影响的研究主要在水生/水华蓝藻方面,对不同扰动时长下荒漠蓝藻生长状况有待进一步研究[15 ] . ...
Effect of small-scale turbulence on the physiology and morphology of two bloom-forming cyanobacteria
1
2016
... 荒漠蓝藻的生长和产量除受到温度、光照、营养盐等环境条件的影响外,也会受到不同时间尺度上扰动过程的控制[8 -10 ] .水体扰动能够通过增加悬浮物浓度从而影响水体光照条件,进而影响藻类的光合效率 [11 ] .不同扰动方式下藻类的敏感度和耐受能力不同,其营养盐吸收速率也会表现出明显差异[12 -13 ] .此外,较强的扰动条件会破坏藻细胞的结构、造成细胞的机械损伤、改变其生物量和沉降量等[14 ] .目前,水体扰动对蓝藻生长影响的研究主要在水生/水华蓝藻方面,对不同扰动时长下荒漠蓝藻生长状况有待进一步研究[15 ] . ...
水体扰动对铜绿微囊藻生长和酶活性的影响
2
2019
... 荒漠蓝藻的生长和产量除受到温度、光照、营养盐等环境条件的影响外,也会受到不同时间尺度上扰动过程的控制[8 -10 ] .水体扰动能够通过增加悬浮物浓度从而影响水体光照条件,进而影响藻类的光合效率 [11 ] .不同扰动方式下藻类的敏感度和耐受能力不同,其营养盐吸收速率也会表现出明显差异[12 -13 ] .此外,较强的扰动条件会破坏藻细胞的结构、造成细胞的机械损伤、改变其生物量和沉降量等[14 ] .目前,水体扰动对蓝藻生长影响的研究主要在水生/水华蓝藻方面,对不同扰动时长下荒漠蓝藻生长状况有待进一步研究[15 ] . ...
... 扰动能够影响水体中悬浮物浓度,改变水体的光照条件及促进N、P等营养盐的传递和转化速率,最终决定藻类初级生产力[26 ] .本研究中,荒漠蓝藻在3种扰动时长下干重呈现出先迅速增加,之后趋于稳定的趋势.在3、6、12 h扰动下荒漠蓝藻的总质量分别为每培养池(15 m3 )0.858、0.940、0.841 kg.在3 h扰动下,荒漠蓝藻在培养后期表现为生长滞后且总产量较少,这可能由于3 h扰动下形成的长时间静水状态会使荒漠蓝藻更易大量沉积在底部或粘结在池壁上,降低了其光合作用速率和酶活性[15 ] .蓝藻进行光暗反应需要一个较为稳定的环境,在12 h扰动下荒漠蓝藻的总质量最少,原因可能是12 h扰动下藻类和水体中其他浮游生物和颗粒物质长时间悬浮并相互摩擦,难以聚集生长,形成的高浊度水还会显著降低入射光强和光合有效辐射,从而破坏藻类生长原有的稳定环境[27 ] .此外,12 h扰动下的长时间水流冲刷作用还会对藻类产生机械剪切,直接损坏藻类细胞结构的完整性[28 -29 ] .生长后期停滞、低光合速率、环境的破坏以及机械损伤可能是本研究中3 h和12 h扰动下总质量低的重要原因.而6 h扰动下荒漠蓝藻的总质量最高,这可能因为对水体扰动的适当减弱既可以为蓝藻的光合作用提供充足的CO2 ,又不会破坏藻细胞,还能使整个培养过程中水体保持稳定的温度、光照强度和光合有效辐射,增大了水体环境的稳定性,有利于接入藻种的生长,从而获得的总质量最高.本文的研究结果与李林等[30 ] 研究中表明间歇性扰动条件下微囊藻(Microcystis aeruginosa )密度较连续性水流条件下高的结果类似;Karp-Boss等[31 ] 认为,连续扰动会对鱼腥藻(Anabaena aquae )细胞产生机械破坏,而间歇性扰动会促进其生长. ...
农业生产与温度的关系
4
2007
... 蓝藻完成某一生育期或完整生命史过程需要一定量的日平均温度的累积,特别要求有特定的有效积温[16 ] .不同地区蓝藻对温度的适应范围不同,这会导致蓝藻生长期内所需有效积温差异明显[17 ] .在太湖水体,上半年(1—6月)的逐日水温积温对同年春末夏初蓝藻爆发的强度有决定作用,且当温度累积到一定程度时,蓝藻才会大面积产生[18 ] .而北方寒冷内陆性碱性水体在有效积温较低时有助于蓝藻爆发[17 ] .此外,日照也是蓝藻增殖的关键条件,累积光照强度和累积光合有效辐射同样可视为影响蓝藻生长的关键因素[17 ,19 ] .目前,关于影响荒漠蓝藻生长的环境因子研究主要围绕在N、P等营养盐浓度,最适生长温度和光照强度等方面,且集中于实验室条件,而荒漠蓝藻生长对温室生态环境的响应机制研究较少[20 -22 ] .有效积温、累积光照强度和累积光合有效辐射显著影响水生蓝藻的生物量积累,但是否对荒漠蓝藻也会产生影响,仍鲜见报道[16 ] . ...
... [16 ]. ...
... 由于没有关于荒漠蓝藻生长下限温度的相关报道,因此以农作物生长的起始温度为参考,本研究中荒漠蓝藻生物学下限温度取值10 ℃[16 ] .日均温度、日均光照强度和日均光合有效辐射采用每日07:00—19:00数据平均值,累积光照强度和累积光合有效辐射为逐日日均光照强度和日均光合有效辐射的累积值,有效积温的计算表达式如下所示: ...
... 蓝藻的生长发育是一个热量的累积过程[16 ] .本文对荒漠蓝藻总质量与有效积温进行研究发现,荒漠蓝藻总质量与有效积温呈显著正相关,较低的积温对蓝藻生长发育有抑制作用.在9—10月,温室条件下荒漠蓝藻完成其生长发育所需有效积温为80—185 ℃,蓝藻总质量的增长速率随积温的增加逐渐增加,快速增长转折点对应的有效积温为145 ℃,在有效积温为180 ℃时,其生物量达到最大值.朱广伟等[18 ] 在分析2005—2017年5—7月太湖北部水体蓝藻叶绿素a浓度的影响因素时,发现积温在调节太湖叶绿素a浓度年际变化过程中发挥正向主导作用.谢小萍等[35 ] 试验发现在太湖蓝藻复苏期,水中的叶绿素a浓度随着有效积温的变化而变化,有效积温越高,水体中蓝藻生物量越高.李星[36 ] 研究发现,积温过低不利于大量蓝藻的产生,黄岐附近水体在积温达到2 300 ℃时有利于加快蓝藻的生长.关于蓝藻生长与积温之间的关系也有不同的结果,北方内陆寒冷地区碱性水体中大量蓝藻的形成相比于热带和亚热带地区需要的有效积温较低,蓝藻爆发与有效积温负相关[17 ] .范裕祥等[37 ] 通过对巢湖蓝藻暴发期间气象观测资料的分析,发现蓝藻发生面积与前5 d积温呈高度显著的线性负相关,持续的高温对蓝藻积聚有明显的抑制作用,5 d积温 165 ℃·d 可作为蓝藻积聚的上限. ...
北方寒冷内陆碱性水体蓝藻暴发特点及影响因子
4
2017
... 蓝藻完成某一生育期或完整生命史过程需要一定量的日平均温度的累积,特别要求有特定的有效积温[16 ] .不同地区蓝藻对温度的适应范围不同,这会导致蓝藻生长期内所需有效积温差异明显[17 ] .在太湖水体,上半年(1—6月)的逐日水温积温对同年春末夏初蓝藻爆发的强度有决定作用,且当温度累积到一定程度时,蓝藻才会大面积产生[18 ] .而北方寒冷内陆性碱性水体在有效积温较低时有助于蓝藻爆发[17 ] .此外,日照也是蓝藻增殖的关键条件,累积光照强度和累积光合有效辐射同样可视为影响蓝藻生长的关键因素[17 ,19 ] .目前,关于影响荒漠蓝藻生长的环境因子研究主要围绕在N、P等营养盐浓度,最适生长温度和光照强度等方面,且集中于实验室条件,而荒漠蓝藻生长对温室生态环境的响应机制研究较少[20 -22 ] .有效积温、累积光照强度和累积光合有效辐射显著影响水生蓝藻的生物量积累,但是否对荒漠蓝藻也会产生影响,仍鲜见报道[16 ] . ...
... [17 ].此外,日照也是蓝藻增殖的关键条件,累积光照强度和累积光合有效辐射同样可视为影响蓝藻生长的关键因素[17 ,19 ] .目前,关于影响荒漠蓝藻生长的环境因子研究主要围绕在N、P等营养盐浓度,最适生长温度和光照强度等方面,且集中于实验室条件,而荒漠蓝藻生长对温室生态环境的响应机制研究较少[20 -22 ] .有效积温、累积光照强度和累积光合有效辐射显著影响水生蓝藻的生物量积累,但是否对荒漠蓝藻也会产生影响,仍鲜见报道[16 ] . ...
... [17 ,19 ].目前,关于影响荒漠蓝藻生长的环境因子研究主要围绕在N、P等营养盐浓度,最适生长温度和光照强度等方面,且集中于实验室条件,而荒漠蓝藻生长对温室生态环境的响应机制研究较少[20 -22 ] .有效积温、累积光照强度和累积光合有效辐射显著影响水生蓝藻的生物量积累,但是否对荒漠蓝藻也会产生影响,仍鲜见报道[16 ] . ...
... 蓝藻的生长发育是一个热量的累积过程[16 ] .本文对荒漠蓝藻总质量与有效积温进行研究发现,荒漠蓝藻总质量与有效积温呈显著正相关,较低的积温对蓝藻生长发育有抑制作用.在9—10月,温室条件下荒漠蓝藻完成其生长发育所需有效积温为80—185 ℃,蓝藻总质量的增长速率随积温的增加逐渐增加,快速增长转折点对应的有效积温为145 ℃,在有效积温为180 ℃时,其生物量达到最大值.朱广伟等[18 ] 在分析2005—2017年5—7月太湖北部水体蓝藻叶绿素a浓度的影响因素时,发现积温在调节太湖叶绿素a浓度年际变化过程中发挥正向主导作用.谢小萍等[35 ] 试验发现在太湖蓝藻复苏期,水中的叶绿素a浓度随着有效积温的变化而变化,有效积温越高,水体中蓝藻生物量越高.李星[36 ] 研究发现,积温过低不利于大量蓝藻的产生,黄岐附近水体在积温达到2 300 ℃时有利于加快蓝藻的生长.关于蓝藻生长与积温之间的关系也有不同的结果,北方内陆寒冷地区碱性水体中大量蓝藻的形成相比于热带和亚热带地区需要的有效积温较低,蓝藻爆发与有效积温负相关[17 ] .范裕祥等[37 ] 通过对巢湖蓝藻暴发期间气象观测资料的分析,发现蓝藻发生面积与前5 d积温呈高度显著的线性负相关,持续的高温对蓝藻积聚有明显的抑制作用,5 d积温 165 ℃·d 可作为蓝藻积聚的上限. ...
2005-2017年北部太湖水体叶绿素a和营养盐变化及影响因素
2
2018
... 蓝藻完成某一生育期或完整生命史过程需要一定量的日平均温度的累积,特别要求有特定的有效积温[16 ] .不同地区蓝藻对温度的适应范围不同,这会导致蓝藻生长期内所需有效积温差异明显[17 ] .在太湖水体,上半年(1—6月)的逐日水温积温对同年春末夏初蓝藻爆发的强度有决定作用,且当温度累积到一定程度时,蓝藻才会大面积产生[18 ] .而北方寒冷内陆性碱性水体在有效积温较低时有助于蓝藻爆发[17 ] .此外,日照也是蓝藻增殖的关键条件,累积光照强度和累积光合有效辐射同样可视为影响蓝藻生长的关键因素[17 ,19 ] .目前,关于影响荒漠蓝藻生长的环境因子研究主要围绕在N、P等营养盐浓度,最适生长温度和光照强度等方面,且集中于实验室条件,而荒漠蓝藻生长对温室生态环境的响应机制研究较少[20 -22 ] .有效积温、累积光照强度和累积光合有效辐射显著影响水生蓝藻的生物量积累,但是否对荒漠蓝藻也会产生影响,仍鲜见报道[16 ] . ...
... 蓝藻的生长发育是一个热量的累积过程[16 ] .本文对荒漠蓝藻总质量与有效积温进行研究发现,荒漠蓝藻总质量与有效积温呈显著正相关,较低的积温对蓝藻生长发育有抑制作用.在9—10月,温室条件下荒漠蓝藻完成其生长发育所需有效积温为80—185 ℃,蓝藻总质量的增长速率随积温的增加逐渐增加,快速增长转折点对应的有效积温为145 ℃,在有效积温为180 ℃时,其生物量达到最大值.朱广伟等[18 ] 在分析2005—2017年5—7月太湖北部水体蓝藻叶绿素a浓度的影响因素时,发现积温在调节太湖叶绿素a浓度年际变化过程中发挥正向主导作用.谢小萍等[35 ] 试验发现在太湖蓝藻复苏期,水中的叶绿素a浓度随着有效积温的变化而变化,有效积温越高,水体中蓝藻生物量越高.李星[36 ] 研究发现,积温过低不利于大量蓝藻的产生,黄岐附近水体在积温达到2 300 ℃时有利于加快蓝藻的生长.关于蓝藻生长与积温之间的关系也有不同的结果,北方内陆寒冷地区碱性水体中大量蓝藻的形成相比于热带和亚热带地区需要的有效积温较低,蓝藻爆发与有效积温负相关[17 ] .范裕祥等[37 ] 通过对巢湖蓝藻暴发期间气象观测资料的分析,发现蓝藻发生面积与前5 d积温呈高度显著的线性负相关,持续的高温对蓝藻积聚有明显的抑制作用,5 d积温 165 ℃·d 可作为蓝藻积聚的上限. ...
水文气象对天目湖沙河水库藻类群落结构动态变化的影响
1
2018
... 蓝藻完成某一生育期或完整生命史过程需要一定量的日平均温度的累积,特别要求有特定的有效积温[16 ] .不同地区蓝藻对温度的适应范围不同,这会导致蓝藻生长期内所需有效积温差异明显[17 ] .在太湖水体,上半年(1—6月)的逐日水温积温对同年春末夏初蓝藻爆发的强度有决定作用,且当温度累积到一定程度时,蓝藻才会大面积产生[18 ] .而北方寒冷内陆性碱性水体在有效积温较低时有助于蓝藻爆发[17 ] .此外,日照也是蓝藻增殖的关键条件,累积光照强度和累积光合有效辐射同样可视为影响蓝藻生长的关键因素[17 ,19 ] .目前,关于影响荒漠蓝藻生长的环境因子研究主要围绕在N、P等营养盐浓度,最适生长温度和光照强度等方面,且集中于实验室条件,而荒漠蓝藻生长对温室生态环境的响应机制研究较少[20 -22 ] .有效积温、累积光照强度和累积光合有效辐射显著影响水生蓝藻的生物量积累,但是否对荒漠蓝藻也会产生影响,仍鲜见报道[16 ] . ...
不同培养条件对具鞘微鞘藻生物量和多糖产量的影响
2
2008
... 蓝藻完成某一生育期或完整生命史过程需要一定量的日平均温度的累积,特别要求有特定的有效积温[16 ] .不同地区蓝藻对温度的适应范围不同,这会导致蓝藻生长期内所需有效积温差异明显[17 ] .在太湖水体,上半年(1—6月)的逐日水温积温对同年春末夏初蓝藻爆发的强度有决定作用,且当温度累积到一定程度时,蓝藻才会大面积产生[18 ] .而北方寒冷内陆性碱性水体在有效积温较低时有助于蓝藻爆发[17 ] .此外,日照也是蓝藻增殖的关键条件,累积光照强度和累积光合有效辐射同样可视为影响蓝藻生长的关键因素[17 ,19 ] .目前,关于影响荒漠蓝藻生长的环境因子研究主要围绕在N、P等营养盐浓度,最适生长温度和光照强度等方面,且集中于实验室条件,而荒漠蓝藻生长对温室生态环境的响应机制研究较少[20 -22 ] .有效积温、累积光照强度和累积光合有效辐射显著影响水生蓝藻的生物量积累,但是否对荒漠蓝藻也会产生影响,仍鲜见报道[16 ] . ...
... 温度和光照强度同样会影响藻类的生长,高于或低于最佳温度和光照强度都会对蓝藻生长产生抑制作用,包括阻碍蛋白质的合成、降低光合作用生长速率、加快酶的失活以及阻碍细胞膜的渗透[32 -34 ] .本研究结果显示,在3、12 h扰动下荒漠蓝藻的干重会受到日均温度、日均光照强度和日均光合有效辐射不同程度的制约,在这2种扰动时长下需保持适中温度(22—23 ℃)、低光照强度(500—1 500 lux)和低光合有效辐射(15—40 μmol·m-2 ·s-1 )才适宜其生长.谢作明等[20 ] 研究发现在温室中的开放式循环培养池培养荒漠蓝藻时应采用遮阳网控制光照强度在 600—700 μE·m-2 ·s-1 ,并将温度保持在35 ℃以下.此外,雷亚萍[34 ] 研究发现辐射强度越高对爪哇伪枝藻的光合活性、叶绿素a、膜结构和蛋白质结构的损伤也越严重,当光强从20 μE·m-2 ·s-1 转到 100 μE·m-2 ·s-1 时,藻体的最大相对电子传递速率和饱和光照强度明显降低. ...
盐胁迫对爪哇伪枝藻(Scytonema javanicum )生理生化特性的影响
1
2003
... 本研究结果显示,在3种扰动时长下水体的pH均升高至9左右,这可能由于蓝藻在生长过程中消耗H2 CO3 所导致[33 ] .在本试验期间,随着时间的推移,水体的电导率逐渐降低,可能由于藻种加入水体中,经过一段时间,开始吸收利用水体中的营养盐来进行自身的生长繁殖,从而使水体中N、P等营养盐降低[33 ] .其中,3 h扰动下水体营养盐消耗剧烈,而6 h扰动下营养盐消耗较为平缓,可以维持稳定的营养盐水平来保证藻细胞的充分生长和分裂.在6 h扰动下,荒漠蓝藻生长末期水体的电导率为470—630 μS·s-1 ,其中在470—500 μS·s-1 下对应的总质量较高,表明荒漠蓝藻适宜生长在低电导率水环境中.这与前人研究中盐胁迫会抑制荒漠蓝藻的生长,其生物量会随盐浓度的升高而下降的结论相似[21 -22 ] . ...
Effects of salt stress on carbohydrate metabolism in desert soil alga Microcoleus vaginatus Gom
2
2006
... 蓝藻完成某一生育期或完整生命史过程需要一定量的日平均温度的累积,特别要求有特定的有效积温[16 ] .不同地区蓝藻对温度的适应范围不同,这会导致蓝藻生长期内所需有效积温差异明显[17 ] .在太湖水体,上半年(1—6月)的逐日水温积温对同年春末夏初蓝藻爆发的强度有决定作用,且当温度累积到一定程度时,蓝藻才会大面积产生[18 ] .而北方寒冷内陆性碱性水体在有效积温较低时有助于蓝藻爆发[17 ] .此外,日照也是蓝藻增殖的关键条件,累积光照强度和累积光合有效辐射同样可视为影响蓝藻生长的关键因素[17 ,19 ] .目前,关于影响荒漠蓝藻生长的环境因子研究主要围绕在N、P等营养盐浓度,最适生长温度和光照强度等方面,且集中于实验室条件,而荒漠蓝藻生长对温室生态环境的响应机制研究较少[20 -22 ] .有效积温、累积光照强度和累积光合有效辐射显著影响水生蓝藻的生物量积累,但是否对荒漠蓝藻也会产生影响,仍鲜见报道[16 ] . ...
... 本研究结果显示,在3种扰动时长下水体的pH均升高至9左右,这可能由于蓝藻在生长过程中消耗H2 CO3 所导致[33 ] .在本试验期间,随着时间的推移,水体的电导率逐渐降低,可能由于藻种加入水体中,经过一段时间,开始吸收利用水体中的营养盐来进行自身的生长繁殖,从而使水体中N、P等营养盐降低[33 ] .其中,3 h扰动下水体营养盐消耗剧烈,而6 h扰动下营养盐消耗较为平缓,可以维持稳定的营养盐水平来保证藻细胞的充分生长和分裂.在6 h扰动下,荒漠蓝藻生长末期水体的电导率为470—630 μS·s-1 ,其中在470—500 μS·s-1 下对应的总质量较高,表明荒漠蓝藻适宜生长在低电导率水环境中.这与前人研究中盐胁迫会抑制荒漠蓝藻的生长,其生物量会随盐浓度的升高而下降的结论相似[21 -22 ] . ...
Cyanobacterial diversity in natural and artificial microbial mats of Lake Fryxell (McMurdo Dry Valleys,Antarctica):a morphological and molecular approach
1
2003
... 具鞘微鞘藻、席藻、鱼腥藻、念珠藻、伪枝藻、单歧藻和爪哇伪枝藻均由腾格里沙漠东南缘发育良好的蓝藻结皮中分离、纯化所得,使用显微镜进行物种鉴定[23 -24 ] .荒漠蓝藻规模化培养步骤如下:首先,将7种蓝藻先用装有无菌BG·11培养液的三角瓶在摇床上进行一级培养,培养10—15 d;随后,将7种蓝藻全部转入装有BG·11培养液塑料培养箱(长×宽×深=0.5 m×0.8 m×0.6 m)中进行10—15 d的二级扩繁.上述培养均在实验室条件下进行,培养温度25±2 ℃、光照强度4 000 lux,光暗比12 h∶12 h.最后,将培养的混合藻种以8 g·t-1 (干重)转入蓝藻培养池内进行规模化培养,培养池中加入7.5 t BG·11培养液[25 ] (水深0.5 m). ...
Effects of superabsorbent polymer on cyanobacterial biological soil crust formation in laboratory
1
2014
... 具鞘微鞘藻、席藻、鱼腥藻、念珠藻、伪枝藻、单歧藻和爪哇伪枝藻均由腾格里沙漠东南缘发育良好的蓝藻结皮中分离、纯化所得,使用显微镜进行物种鉴定[23 -24 ] .荒漠蓝藻规模化培养步骤如下:首先,将7种蓝藻先用装有无菌BG·11培养液的三角瓶在摇床上进行一级培养,培养10—15 d;随后,将7种蓝藻全部转入装有BG·11培养液塑料培养箱(长×宽×深=0.5 m×0.8 m×0.6 m)中进行10—15 d的二级扩繁.上述培养均在实验室条件下进行,培养温度25±2 ℃、光照强度4 000 lux,光暗比12 h∶12 h.最后,将培养的混合藻种以8 g·t-1 (干重)转入蓝藻培养池内进行规模化培养,培养池中加入7.5 t BG·11培养液[25 ] (水深0.5 m). ...
Generic assignments,strain histories and properties of pure cultures of cyanobacteria
1
1979
... 具鞘微鞘藻、席藻、鱼腥藻、念珠藻、伪枝藻、单歧藻和爪哇伪枝藻均由腾格里沙漠东南缘发育良好的蓝藻结皮中分离、纯化所得,使用显微镜进行物种鉴定[23 -24 ] .荒漠蓝藻规模化培养步骤如下:首先,将7种蓝藻先用装有无菌BG·11培养液的三角瓶在摇床上进行一级培养,培养10—15 d;随后,将7种蓝藻全部转入装有BG·11培养液塑料培养箱(长×宽×深=0.5 m×0.8 m×0.6 m)中进行10—15 d的二级扩繁.上述培养均在实验室条件下进行,培养温度25±2 ℃、光照强度4 000 lux,光暗比12 h∶12 h.最后,将培养的混合藻种以8 g·t-1 (干重)转入蓝藻培养池内进行规模化培养,培养池中加入7.5 t BG·11培养液[25 ] (水深0.5 m). ...
Response of cellular stoichiometry and phosphorus storage of the cyanobacteria Aphanizomenon flos-aquae to small-scale turbulence
1
2017
... 扰动能够影响水体中悬浮物浓度,改变水体的光照条件及促进N、P等营养盐的传递和转化速率,最终决定藻类初级生产力[26 ] .本研究中,荒漠蓝藻在3种扰动时长下干重呈现出先迅速增加,之后趋于稳定的趋势.在3、6、12 h扰动下荒漠蓝藻的总质量分别为每培养池(15 m3 )0.858、0.940、0.841 kg.在3 h扰动下,荒漠蓝藻在培养后期表现为生长滞后且总产量较少,这可能由于3 h扰动下形成的长时间静水状态会使荒漠蓝藻更易大量沉积在底部或粘结在池壁上,降低了其光合作用速率和酶活性[15 ] .蓝藻进行光暗反应需要一个较为稳定的环境,在12 h扰动下荒漠蓝藻的总质量最少,原因可能是12 h扰动下藻类和水体中其他浮游生物和颗粒物质长时间悬浮并相互摩擦,难以聚集生长,形成的高浊度水还会显著降低入射光强和光合有效辐射,从而破坏藻类生长原有的稳定环境[27 ] .此外,12 h扰动下的长时间水流冲刷作用还会对藻类产生机械剪切,直接损坏藻类细胞结构的完整性[28 -29 ] .生长后期停滞、低光合速率、环境的破坏以及机械损伤可能是本研究中3 h和12 h扰动下总质量低的重要原因.而6 h扰动下荒漠蓝藻的总质量最高,这可能因为对水体扰动的适当减弱既可以为蓝藻的光合作用提供充足的CO2 ,又不会破坏藻细胞,还能使整个培养过程中水体保持稳定的温度、光照强度和光合有效辐射,增大了水体环境的稳定性,有利于接入藻种的生长,从而获得的总质量最高.本文的研究结果与李林等[30 ] 研究中表明间歇性扰动条件下微囊藻(Microcystis aeruginosa )密度较连续性水流条件下高的结果类似;Karp-Boss等[31 ] 认为,连续扰动会对鱼腥藻(Anabaena aquae )细胞产生机械破坏,而间歇性扰动会促进其生长. ...
换水率和营养水平对太湖流域横山水库硅藻水华的影响
1
2017
... 扰动能够影响水体中悬浮物浓度,改变水体的光照条件及促进N、P等营养盐的传递和转化速率,最终决定藻类初级生产力[26 ] .本研究中,荒漠蓝藻在3种扰动时长下干重呈现出先迅速增加,之后趋于稳定的趋势.在3、6、12 h扰动下荒漠蓝藻的总质量分别为每培养池(15 m3 )0.858、0.940、0.841 kg.在3 h扰动下,荒漠蓝藻在培养后期表现为生长滞后且总产量较少,这可能由于3 h扰动下形成的长时间静水状态会使荒漠蓝藻更易大量沉积在底部或粘结在池壁上,降低了其光合作用速率和酶活性[15 ] .蓝藻进行光暗反应需要一个较为稳定的环境,在12 h扰动下荒漠蓝藻的总质量最少,原因可能是12 h扰动下藻类和水体中其他浮游生物和颗粒物质长时间悬浮并相互摩擦,难以聚集生长,形成的高浊度水还会显著降低入射光强和光合有效辐射,从而破坏藻类生长原有的稳定环境[27 ] .此外,12 h扰动下的长时间水流冲刷作用还会对藻类产生机械剪切,直接损坏藻类细胞结构的完整性[28 -29 ] .生长后期停滞、低光合速率、环境的破坏以及机械损伤可能是本研究中3 h和12 h扰动下总质量低的重要原因.而6 h扰动下荒漠蓝藻的总质量最高,这可能因为对水体扰动的适当减弱既可以为蓝藻的光合作用提供充足的CO2 ,又不会破坏藻细胞,还能使整个培养过程中水体保持稳定的温度、光照强度和光合有效辐射,增大了水体环境的稳定性,有利于接入藻种的生长,从而获得的总质量最高.本文的研究结果与李林等[30 ] 研究中表明间歇性扰动条件下微囊藻(Microcystis aeruginosa )密度较连续性水流条件下高的结果类似;Karp-Boss等[31 ] 认为,连续扰动会对鱼腥藻(Anabaena aquae )细胞产生机械破坏,而间歇性扰动会促进其生长. ...
Gas vesicle collapse by turgor pressure and its role in buoyancy regulation by Anabaena flosaquae
1
1991
... 扰动能够影响水体中悬浮物浓度,改变水体的光照条件及促进N、P等营养盐的传递和转化速率,最终决定藻类初级生产力[26 ] .本研究中,荒漠蓝藻在3种扰动时长下干重呈现出先迅速增加,之后趋于稳定的趋势.在3、6、12 h扰动下荒漠蓝藻的总质量分别为每培养池(15 m3 )0.858、0.940、0.841 kg.在3 h扰动下,荒漠蓝藻在培养后期表现为生长滞后且总产量较少,这可能由于3 h扰动下形成的长时间静水状态会使荒漠蓝藻更易大量沉积在底部或粘结在池壁上,降低了其光合作用速率和酶活性[15 ] .蓝藻进行光暗反应需要一个较为稳定的环境,在12 h扰动下荒漠蓝藻的总质量最少,原因可能是12 h扰动下藻类和水体中其他浮游生物和颗粒物质长时间悬浮并相互摩擦,难以聚集生长,形成的高浊度水还会显著降低入射光强和光合有效辐射,从而破坏藻类生长原有的稳定环境[27 ] .此外,12 h扰动下的长时间水流冲刷作用还会对藻类产生机械剪切,直接损坏藻类细胞结构的完整性[28 -29 ] .生长后期停滞、低光合速率、环境的破坏以及机械损伤可能是本研究中3 h和12 h扰动下总质量低的重要原因.而6 h扰动下荒漠蓝藻的总质量最高,这可能因为对水体扰动的适当减弱既可以为蓝藻的光合作用提供充足的CO2 ,又不会破坏藻细胞,还能使整个培养过程中水体保持稳定的温度、光照强度和光合有效辐射,增大了水体环境的稳定性,有利于接入藻种的生长,从而获得的总质量最高.本文的研究结果与李林等[30 ] 研究中表明间歇性扰动条件下微囊藻(Microcystis aeruginosa )密度较连续性水流条件下高的结果类似;Karp-Boss等[31 ] 认为,连续扰动会对鱼腥藻(Anabaena aquae )细胞产生机械破坏,而间歇性扰动会促进其生长. ...
水体扰动对铜绿微囊藻生长影响的规律及原因
1
2012
... 扰动能够影响水体中悬浮物浓度,改变水体的光照条件及促进N、P等营养盐的传递和转化速率,最终决定藻类初级生产力[26 ] .本研究中,荒漠蓝藻在3种扰动时长下干重呈现出先迅速增加,之后趋于稳定的趋势.在3、6、12 h扰动下荒漠蓝藻的总质量分别为每培养池(15 m3 )0.858、0.940、0.841 kg.在3 h扰动下,荒漠蓝藻在培养后期表现为生长滞后且总产量较少,这可能由于3 h扰动下形成的长时间静水状态会使荒漠蓝藻更易大量沉积在底部或粘结在池壁上,降低了其光合作用速率和酶活性[15 ] .蓝藻进行光暗反应需要一个较为稳定的环境,在12 h扰动下荒漠蓝藻的总质量最少,原因可能是12 h扰动下藻类和水体中其他浮游生物和颗粒物质长时间悬浮并相互摩擦,难以聚集生长,形成的高浊度水还会显著降低入射光强和光合有效辐射,从而破坏藻类生长原有的稳定环境[27 ] .此外,12 h扰动下的长时间水流冲刷作用还会对藻类产生机械剪切,直接损坏藻类细胞结构的完整性[28 -29 ] .生长后期停滞、低光合速率、环境的破坏以及机械损伤可能是本研究中3 h和12 h扰动下总质量低的重要原因.而6 h扰动下荒漠蓝藻的总质量最高,这可能因为对水体扰动的适当减弱既可以为蓝藻的光合作用提供充足的CO2 ,又不会破坏藻细胞,还能使整个培养过程中水体保持稳定的温度、光照强度和光合有效辐射,增大了水体环境的稳定性,有利于接入藻种的生长,从而获得的总质量最高.本文的研究结果与李林等[30 ] 研究中表明间歇性扰动条件下微囊藻(Microcystis aeruginosa )密度较连续性水流条件下高的结果类似;Karp-Boss等[31 ] 认为,连续扰动会对鱼腥藻(Anabaena aquae )细胞产生机械破坏,而间歇性扰动会促进其生长. ...
连续水流和间歇水流对微囊藻生长的影响
1
2012
... 扰动能够影响水体中悬浮物浓度,改变水体的光照条件及促进N、P等营养盐的传递和转化速率,最终决定藻类初级生产力[26 ] .本研究中,荒漠蓝藻在3种扰动时长下干重呈现出先迅速增加,之后趋于稳定的趋势.在3、6、12 h扰动下荒漠蓝藻的总质量分别为每培养池(15 m3 )0.858、0.940、0.841 kg.在3 h扰动下,荒漠蓝藻在培养后期表现为生长滞后且总产量较少,这可能由于3 h扰动下形成的长时间静水状态会使荒漠蓝藻更易大量沉积在底部或粘结在池壁上,降低了其光合作用速率和酶活性[15 ] .蓝藻进行光暗反应需要一个较为稳定的环境,在12 h扰动下荒漠蓝藻的总质量最少,原因可能是12 h扰动下藻类和水体中其他浮游生物和颗粒物质长时间悬浮并相互摩擦,难以聚集生长,形成的高浊度水还会显著降低入射光强和光合有效辐射,从而破坏藻类生长原有的稳定环境[27 ] .此外,12 h扰动下的长时间水流冲刷作用还会对藻类产生机械剪切,直接损坏藻类细胞结构的完整性[28 -29 ] .生长后期停滞、低光合速率、环境的破坏以及机械损伤可能是本研究中3 h和12 h扰动下总质量低的重要原因.而6 h扰动下荒漠蓝藻的总质量最高,这可能因为对水体扰动的适当减弱既可以为蓝藻的光合作用提供充足的CO2 ,又不会破坏藻细胞,还能使整个培养过程中水体保持稳定的温度、光照强度和光合有效辐射,增大了水体环境的稳定性,有利于接入藻种的生长,从而获得的总质量最高.本文的研究结果与李林等[30 ] 研究中表明间歇性扰动条件下微囊藻(Microcystis aeruginosa )密度较连续性水流条件下高的结果类似;Karp-Boss等[31 ] 认为,连续扰动会对鱼腥藻(Anabaena aquae )细胞产生机械破坏,而间歇性扰动会促进其生长. ...
Nutrient fluxes to planktonic osmotrophs in the presence of fluid motion
1
1996
... 扰动能够影响水体中悬浮物浓度,改变水体的光照条件及促进N、P等营养盐的传递和转化速率,最终决定藻类初级生产力[26 ] .本研究中,荒漠蓝藻在3种扰动时长下干重呈现出先迅速增加,之后趋于稳定的趋势.在3、6、12 h扰动下荒漠蓝藻的总质量分别为每培养池(15 m3 )0.858、0.940、0.841 kg.在3 h扰动下,荒漠蓝藻在培养后期表现为生长滞后且总产量较少,这可能由于3 h扰动下形成的长时间静水状态会使荒漠蓝藻更易大量沉积在底部或粘结在池壁上,降低了其光合作用速率和酶活性[15 ] .蓝藻进行光暗反应需要一个较为稳定的环境,在12 h扰动下荒漠蓝藻的总质量最少,原因可能是12 h扰动下藻类和水体中其他浮游生物和颗粒物质长时间悬浮并相互摩擦,难以聚集生长,形成的高浊度水还会显著降低入射光强和光合有效辐射,从而破坏藻类生长原有的稳定环境[27 ] .此外,12 h扰动下的长时间水流冲刷作用还会对藻类产生机械剪切,直接损坏藻类细胞结构的完整性[28 -29 ] .生长后期停滞、低光合速率、环境的破坏以及机械损伤可能是本研究中3 h和12 h扰动下总质量低的重要原因.而6 h扰动下荒漠蓝藻的总质量最高,这可能因为对水体扰动的适当减弱既可以为蓝藻的光合作用提供充足的CO2 ,又不会破坏藻细胞,还能使整个培养过程中水体保持稳定的温度、光照强度和光合有效辐射,增大了水体环境的稳定性,有利于接入藻种的生长,从而获得的总质量最高.本文的研究结果与李林等[30 ] 研究中表明间歇性扰动条件下微囊藻(Microcystis aeruginosa )密度较连续性水流条件下高的结果类似;Karp-Boss等[31 ] 认为,连续扰动会对鱼腥藻(Anabaena aquae )细胞产生机械破坏,而间歇性扰动会促进其生长. ...
不同风等级扰动对贫富营养下铜绿微囊藻生长的影响
1
2008
... 温度和光照强度同样会影响藻类的生长,高于或低于最佳温度和光照强度都会对蓝藻生长产生抑制作用,包括阻碍蛋白质的合成、降低光合作用生长速率、加快酶的失活以及阻碍细胞膜的渗透[32 -34 ] .本研究结果显示,在3、12 h扰动下荒漠蓝藻的干重会受到日均温度、日均光照强度和日均光合有效辐射不同程度的制约,在这2种扰动时长下需保持适中温度(22—23 ℃)、低光照强度(500—1 500 lux)和低光合有效辐射(15—40 μmol·m-2 ·s-1 )才适宜其生长.谢作明等[20 ] 研究发现在温室中的开放式循环培养池培养荒漠蓝藻时应采用遮阳网控制光照强度在 600—700 μE·m-2 ·s-1 ,并将温度保持在35 ℃以下.此外,雷亚萍[34 ] 研究发现辐射强度越高对爪哇伪枝藻的光合活性、叶绿素a、膜结构和蛋白质结构的损伤也越严重,当光强从20 μE·m-2 ·s-1 转到 100 μE·m-2 ·s-1 时,藻体的最大相对电子传递速率和饱和光照强度明显降低. ...
安徽沱湖夏季浮游植物群落结构特征与环境因子关系
2
2018
... 本研究结果显示,在3种扰动时长下水体的pH均升高至9左右,这可能由于蓝藻在生长过程中消耗H2 CO3 所导致[33 ] .在本试验期间,随着时间的推移,水体的电导率逐渐降低,可能由于藻种加入水体中,经过一段时间,开始吸收利用水体中的营养盐来进行自身的生长繁殖,从而使水体中N、P等营养盐降低[33 ] .其中,3 h扰动下水体营养盐消耗剧烈,而6 h扰动下营养盐消耗较为平缓,可以维持稳定的营养盐水平来保证藻细胞的充分生长和分裂.在6 h扰动下,荒漠蓝藻生长末期水体的电导率为470—630 μS·s-1 ,其中在470—500 μS·s-1 下对应的总质量较高,表明荒漠蓝藻适宜生长在低电导率水环境中.这与前人研究中盐胁迫会抑制荒漠蓝藻的生长,其生物量会随盐浓度的升高而下降的结论相似[21 -22 ] . ...
... [33 ].其中,3 h扰动下水体营养盐消耗剧烈,而6 h扰动下营养盐消耗较为平缓,可以维持稳定的营养盐水平来保证藻细胞的充分生长和分裂.在6 h扰动下,荒漠蓝藻生长末期水体的电导率为470—630 μS·s-1 ,其中在470—500 μS·s-1 下对应的总质量较高,表明荒漠蓝藻适宜生长在低电导率水环境中.这与前人研究中盐胁迫会抑制荒漠蓝藻的生长,其生物量会随盐浓度的升高而下降的结论相似[21 -22 ] . ...
共生关系对爪哇伪枝藻抗荒漠胁迫能力的影响
2
2017
... 温度和光照强度同样会影响藻类的生长,高于或低于最佳温度和光照强度都会对蓝藻生长产生抑制作用,包括阻碍蛋白质的合成、降低光合作用生长速率、加快酶的失活以及阻碍细胞膜的渗透[32 -34 ] .本研究结果显示,在3、12 h扰动下荒漠蓝藻的干重会受到日均温度、日均光照强度和日均光合有效辐射不同程度的制约,在这2种扰动时长下需保持适中温度(22—23 ℃)、低光照强度(500—1 500 lux)和低光合有效辐射(15—40 μmol·m-2 ·s-1 )才适宜其生长.谢作明等[20 ] 研究发现在温室中的开放式循环培养池培养荒漠蓝藻时应采用遮阳网控制光照强度在 600—700 μE·m-2 ·s-1 ,并将温度保持在35 ℃以下.此外,雷亚萍[34 ] 研究发现辐射强度越高对爪哇伪枝藻的光合活性、叶绿素a、膜结构和蛋白质结构的损伤也越严重,当光强从20 μE·m-2 ·s-1 转到 100 μE·m-2 ·s-1 时,藻体的最大相对电子传递速率和饱和光照强度明显降低. ...
... [34 ]研究发现辐射强度越高对爪哇伪枝藻的光合活性、叶绿素a、膜结构和蛋白质结构的损伤也越严重,当光强从20 μE·m-2 ·s-1 转到 100 μE·m-2 ·s-1 时,藻体的最大相对电子传递速率和饱和光照强度明显降低. ...
气温对太湖蓝藻复苏和休眠进程的影响
1
2016
... 蓝藻的生长发育是一个热量的累积过程[16 ] .本文对荒漠蓝藻总质量与有效积温进行研究发现,荒漠蓝藻总质量与有效积温呈显著正相关,较低的积温对蓝藻生长发育有抑制作用.在9—10月,温室条件下荒漠蓝藻完成其生长发育所需有效积温为80—185 ℃,蓝藻总质量的增长速率随积温的增加逐渐增加,快速增长转折点对应的有效积温为145 ℃,在有效积温为180 ℃时,其生物量达到最大值.朱广伟等[18 ] 在分析2005—2017年5—7月太湖北部水体蓝藻叶绿素a浓度的影响因素时,发现积温在调节太湖叶绿素a浓度年际变化过程中发挥正向主导作用.谢小萍等[35 ] 试验发现在太湖蓝藻复苏期,水中的叶绿素a浓度随着有效积温的变化而变化,有效积温越高,水体中蓝藻生物量越高.李星[36 ] 研究发现,积温过低不利于大量蓝藻的产生,黄岐附近水体在积温达到2 300 ℃时有利于加快蓝藻的生长.关于蓝藻生长与积温之间的关系也有不同的结果,北方内陆寒冷地区碱性水体中大量蓝藻的形成相比于热带和亚热带地区需要的有效积温较低,蓝藻爆发与有效积温负相关[17 ] .范裕祥等[37 ] 通过对巢湖蓝藻暴发期间气象观测资料的分析,发现蓝藻发生面积与前5 d积温呈高度显著的线性负相关,持续的高温对蓝藻积聚有明显的抑制作用,5 d积温 165 ℃·d 可作为蓝藻积聚的上限. ...
海表温度对连江黄岐赤潮影响的研究
1
2021
... 蓝藻的生长发育是一个热量的累积过程[16 ] .本文对荒漠蓝藻总质量与有效积温进行研究发现,荒漠蓝藻总质量与有效积温呈显著正相关,较低的积温对蓝藻生长发育有抑制作用.在9—10月,温室条件下荒漠蓝藻完成其生长发育所需有效积温为80—185 ℃,蓝藻总质量的增长速率随积温的增加逐渐增加,快速增长转折点对应的有效积温为145 ℃,在有效积温为180 ℃时,其生物量达到最大值.朱广伟等[18 ] 在分析2005—2017年5—7月太湖北部水体蓝藻叶绿素a浓度的影响因素时,发现积温在调节太湖叶绿素a浓度年际变化过程中发挥正向主导作用.谢小萍等[35 ] 试验发现在太湖蓝藻复苏期,水中的叶绿素a浓度随着有效积温的变化而变化,有效积温越高,水体中蓝藻生物量越高.李星[36 ] 研究发现,积温过低不利于大量蓝藻的产生,黄岐附近水体在积温达到2 300 ℃时有利于加快蓝藻的生长.关于蓝藻生长与积温之间的关系也有不同的结果,北方内陆寒冷地区碱性水体中大量蓝藻的形成相比于热带和亚热带地区需要的有效积温较低,蓝藻爆发与有效积温负相关[17 ] .范裕祥等[37 ] 通过对巢湖蓝藻暴发期间气象观测资料的分析,发现蓝藻发生面积与前5 d积温呈高度显著的线性负相关,持续的高温对蓝藻积聚有明显的抑制作用,5 d积温 165 ℃·d 可作为蓝藻积聚的上限. ...
巢湖蓝藻水华分布特征和气象条件分析
1
2015
... 蓝藻的生长发育是一个热量的累积过程[16 ] .本文对荒漠蓝藻总质量与有效积温进行研究发现,荒漠蓝藻总质量与有效积温呈显著正相关,较低的积温对蓝藻生长发育有抑制作用.在9—10月,温室条件下荒漠蓝藻完成其生长发育所需有效积温为80—185 ℃,蓝藻总质量的增长速率随积温的增加逐渐增加,快速增长转折点对应的有效积温为145 ℃,在有效积温为180 ℃时,其生物量达到最大值.朱广伟等[18 ] 在分析2005—2017年5—7月太湖北部水体蓝藻叶绿素a浓度的影响因素时,发现积温在调节太湖叶绿素a浓度年际变化过程中发挥正向主导作用.谢小萍等[35 ] 试验发现在太湖蓝藻复苏期,水中的叶绿素a浓度随着有效积温的变化而变化,有效积温越高,水体中蓝藻生物量越高.李星[36 ] 研究发现,积温过低不利于大量蓝藻的产生,黄岐附近水体在积温达到2 300 ℃时有利于加快蓝藻的生长.关于蓝藻生长与积温之间的关系也有不同的结果,北方内陆寒冷地区碱性水体中大量蓝藻的形成相比于热带和亚热带地区需要的有效积温较低,蓝藻爆发与有效积温负相关[17 ] .范裕祥等[37 ] 通过对巢湖蓝藻暴发期间气象观测资料的分析,发现蓝藻发生面积与前5 d积温呈高度显著的线性负相关,持续的高温对蓝藻积聚有明显的抑制作用,5 d积温 165 ℃·d 可作为蓝藻积聚的上限. ...





 甘公网安备 62010202000688号
甘公网安备 62010202000688号
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}