巴丹吉林沙漠植被群落特征与土壤水分关系研究
1
2021
... 生物多样性对环境因子的响应已经成为生态学研究的热点[1-2].β多样性表示不同群落间物种组成差异,是联系α多样性与γ多样性的桥梁和纽带[3-4],能较为直观地反映环境与物种多样性的相互作用[5].β多样性格局的形成可以分为两种途径.一是物种的周转,表示不同群落间物种的替换;二是物种的嵌套,表示一个低丰富度的群落是另一高丰富度群落非随机物种丢失的结果,前者是后者的子集,形成嵌套[6].为了量化两种途径,基于Sørensen相异性指数,β多样性可被分解为周转和嵌套两部分[7-8].该分解方法有利于我们更好地了解β多样性的形成机制. ...
额济纳荒漠绿洲植物群落的数量分类及其与地下水环境的关系分析
1
2011
... 生物多样性对环境因子的响应已经成为生态学研究的热点[1-2].β多样性表示不同群落间物种组成差异,是联系α多样性与γ多样性的桥梁和纽带[3-4],能较为直观地反映环境与物种多样性的相互作用[5].β多样性格局的形成可以分为两种途径.一是物种的周转,表示不同群落间物种的替换;二是物种的嵌套,表示一个低丰富度的群落是另一高丰富度群落非随机物种丢失的结果,前者是后者的子集,形成嵌套[6].为了量化两种途径,基于Sørensen相异性指数,β多样性可被分解为周转和嵌套两部分[7-8].该分解方法有利于我们更好地了解β多样性的形成机制. ...
Evolution and measurement of species diversity
2
1972
... 生物多样性对环境因子的响应已经成为生态学研究的热点[1-2].β多样性表示不同群落间物种组成差异,是联系α多样性与γ多样性的桥梁和纽带[3-4],能较为直观地反映环境与物种多样性的相互作用[5].β多样性格局的形成可以分为两种途径.一是物种的周转,表示不同群落间物种的替换;二是物种的嵌套,表示一个低丰富度的群落是另一高丰富度群落非随机物种丢失的结果,前者是后者的子集,形成嵌套[6].为了量化两种途径,基于Sørensen相异性指数,β多样性可被分解为周转和嵌套两部分[7-8].该分解方法有利于我们更好地了解β多样性的形成机制. ...
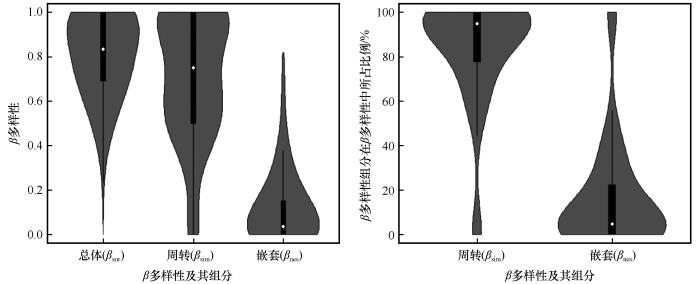
... β多样性不仅代表着一个地区物种多样性的分布情况,而且还体现着物种与环境之间的关系[3,10,16,29].通过对乌兰布和沙漠植物群落的研究,利用Sørensen相异性指数来表征植物群落β多样性.结果发现该区域植物群落相异性指数较大(均值为0.82,图2),表明该区域植物群落的物种组成之间存在较大差异[30].为了量化植物群落β多样性的过程,将总体β多样性(βsor)分解为周转(βsim)和嵌套(βnes)两部分,发现周转占有很大比例,嵌套比例较低.这与库姆塔格沙漠南缘的植物群落β多样性研究结果一致[30].因此乌兰布和沙漠的植物群落β多样性可能主要受植物群落物种在空间上或群落间的周转影响. ...
Beta-diversity in tropical forest trees
1
2009
... 生物多样性对环境因子的响应已经成为生态学研究的热点[1-2].β多样性表示不同群落间物种组成差异,是联系α多样性与γ多样性的桥梁和纽带[3-4],能较为直观地反映环境与物种多样性的相互作用[5].β多样性格局的形成可以分为两种途径.一是物种的周转,表示不同群落间物种的替换;二是物种的嵌套,表示一个低丰富度的群落是另一高丰富度群落非随机物种丢失的结果,前者是后者的子集,形成嵌套[6].为了量化两种途径,基于Sørensen相异性指数,β多样性可被分解为周转和嵌套两部分[7-8].该分解方法有利于我们更好地了解β多样性的形成机制. ...
千岛湖岛屿维管植物β多样性及其影响因素
2
2014
... 生物多样性对环境因子的响应已经成为生态学研究的热点[1-2].β多样性表示不同群落间物种组成差异,是联系α多样性与γ多样性的桥梁和纽带[3-4],能较为直观地反映环境与物种多样性的相互作用[5].β多样性格局的形成可以分为两种途径.一是物种的周转,表示不同群落间物种的替换;二是物种的嵌套,表示一个低丰富度的群落是另一高丰富度群落非随机物种丢失的结果,前者是后者的子集,形成嵌套[6].为了量化两种途径,基于Sørensen相异性指数,β多样性可被分解为周转和嵌套两部分[7-8].该分解方法有利于我们更好地了解β多样性的形成机制. ...
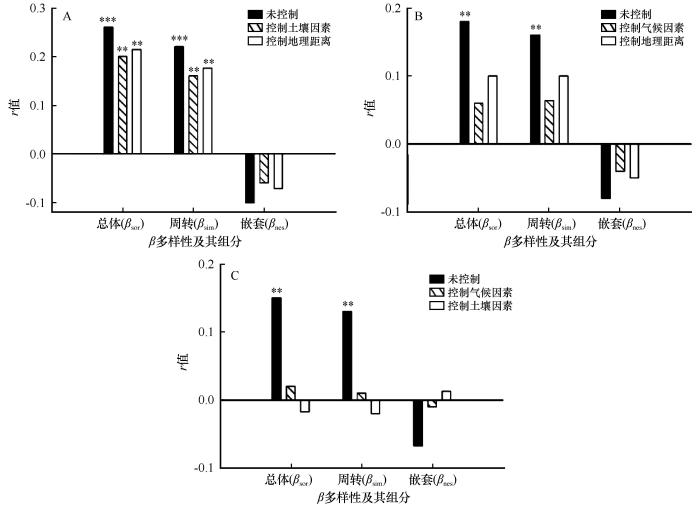
... 一般认为β多样性主要由扩散过程和生态位过程两种机制共同作用形成[9-10].生态位理论认为物种组成差异由生境变化导致,扩散理论认为物种分布格局完全由其扩散能力确定[5].扩散限制和生境过滤并不是互斥关系,有时同时起着作用[37].本文研究表明环境差异和地理距离均与乌兰布和沙漠植物群落β多样性及其组分存在显著的相关性(图3).在荒漠生态系统中,干旱起着强烈的环境过滤作用[38].随着干旱程度的增加,有利于比叶面积小和根系能在短时间获取最大养分和水分的植物生存[39-40].植物(如梭梭、沙拐枣等)会通过减少地上生物量的分配和叶片退化减少水分的散失,并把更多光合作用产物分配给地下部分[41].物种相似性随着地理距离的增大而减小,说明扩散限制在植物群落β多样性中起一定作用.但二者对于研究区植物群落β多样性及其周转组分的影响存在差异,与气候因素、土壤因素的相关系数分别为0.26、0.18,与地理距离的相关系数是0.16(图3).环境筛选作用远大于扩散限制,在荒漠区的库姆塔格沙漠南缘和古尔班通古特沙漠研究中也得到过类似结果[30,34].这说明在环境贫瘠、物种结构简单的乌兰布和沙漠,环境过滤作用对植物群落β多样性的作用大.值得注意的是,在研究区域中物种周转主导植物群落β多样性的变化,且对环境变量的响应与植物群落β多样性一致.因此,随着环境距离(包括气候和土壤等环境条件的差异)和地理距离的增加,空间异质性也明显增强,植物种子的扩散受到限制[42],在长期自然选择和群落演替中,不同生境下形成与之相适应的物种[43].本研究表明,以环境过滤为主,加之扩散限制共同影响着乌兰布和沙漠的植物群落β多样性的格局,即生态位理论和中性理论共同驱动乌兰布和沙漠植物群落的构建. ...
The geographical structure of British bird distributions:diversity,spatial turnover and scale
1
2001
... 生物多样性对环境因子的响应已经成为生态学研究的热点[1-2].β多样性表示不同群落间物种组成差异,是联系α多样性与γ多样性的桥梁和纽带[3-4],能较为直观地反映环境与物种多样性的相互作用[5].β多样性格局的形成可以分为两种途径.一是物种的周转,表示不同群落间物种的替换;二是物种的嵌套,表示一个低丰富度的群落是另一高丰富度群落非随机物种丢失的结果,前者是后者的子集,形成嵌套[6].为了量化两种途径,基于Sørensen相异性指数,β多样性可被分解为周转和嵌套两部分[7-8].该分解方法有利于我们更好地了解β多样性的形成机制. ...
Partitioning the turnover and nestedness components of beta diversity
1
2010
... 生物多样性对环境因子的响应已经成为生态学研究的热点[1-2].β多样性表示不同群落间物种组成差异,是联系α多样性与γ多样性的桥梁和纽带[3-4],能较为直观地反映环境与物种多样性的相互作用[5].β多样性格局的形成可以分为两种途径.一是物种的周转,表示不同群落间物种的替换;二是物种的嵌套,表示一个低丰富度的群落是另一高丰富度群落非随机物种丢失的结果,前者是后者的子集,形成嵌套[6].为了量化两种途径,基于Sørensen相异性指数,β多样性可被分解为周转和嵌套两部分[7-8].该分解方法有利于我们更好地了解β多样性的形成机制. ...
Disentangling nestedness from models of ecological complexity
1
2012
... 生物多样性对环境因子的响应已经成为生态学研究的热点[1-2].β多样性表示不同群落间物种组成差异,是联系α多样性与γ多样性的桥梁和纽带[3-4],能较为直观地反映环境与物种多样性的相互作用[5].β多样性格局的形成可以分为两种途径.一是物种的周转,表示不同群落间物种的替换;二是物种的嵌套,表示一个低丰富度的群落是另一高丰富度群落非随机物种丢失的结果,前者是后者的子集,形成嵌套[6].为了量化两种途径,基于Sørensen相异性指数,β多样性可被分解为周转和嵌套两部分[7-8].该分解方法有利于我们更好地了解β多样性的形成机制. ...
The distance decay of similarity in ecological communities
2
2007
... 一般认为物种β多样性主要由生态位过程和扩散过程共同作用形成[9-10].按照生态位理论,历史过程决定了区域的物种库,环境因素决定着群落中物种的共存[11].不同的物种有着各自适应的生境,生境差异越大,物种组成差异也越大[12].气候作为重要的环境因子,对不同物种空间更替的作用一直是研究的热点[13-14].除了气候,土壤异质性在小尺度上也是影响植被特征的重要因子[15].中性理论认为,物种随机扩散过程是植物群落构建的重要机制[16].在研究群落周转中,广泛采用地理距离来表征物种扩散限制[17].在扩散限制的影响下,群落物种组成的相似性随着地理距离增加而减小[12].环境过滤和扩散限制对植物群落β多样性的形成至关重要. ...
... 一般认为β多样性主要由扩散过程和生态位过程两种机制共同作用形成[9-10].生态位理论认为物种组成差异由生境变化导致,扩散理论认为物种分布格局完全由其扩散能力确定[5].扩散限制和生境过滤并不是互斥关系,有时同时起着作用[37].本文研究表明环境差异和地理距离均与乌兰布和沙漠植物群落β多样性及其组分存在显著的相关性(图3).在荒漠生态系统中,干旱起着强烈的环境过滤作用[38].随着干旱程度的增加,有利于比叶面积小和根系能在短时间获取最大养分和水分的植物生存[39-40].植物(如梭梭、沙拐枣等)会通过减少地上生物量的分配和叶片退化减少水分的散失,并把更多光合作用产物分配给地下部分[41].物种相似性随着地理距离的增大而减小,说明扩散限制在植物群落β多样性中起一定作用.但二者对于研究区植物群落β多样性及其周转组分的影响存在差异,与气候因素、土壤因素的相关系数分别为0.26、0.18,与地理距离的相关系数是0.16(图3).环境筛选作用远大于扩散限制,在荒漠区的库姆塔格沙漠南缘和古尔班通古特沙漠研究中也得到过类似结果[30,34].这说明在环境贫瘠、物种结构简单的乌兰布和沙漠,环境过滤作用对植物群落β多样性的作用大.值得注意的是,在研究区域中物种周转主导植物群落β多样性的变化,且对环境变量的响应与植物群落β多样性一致.因此,随着环境距离(包括气候和土壤等环境条件的差异)和地理距离的增加,空间异质性也明显增强,植物种子的扩散受到限制[42],在长期自然选择和群落演替中,不同生境下形成与之相适应的物种[43].本研究表明,以环境过滤为主,加之扩散限制共同影响着乌兰布和沙漠的植物群落β多样性的格局,即生态位理论和中性理论共同驱动乌兰布和沙漠植物群落的构建. ...
Partitioning beta diversity in a subtropical broadleaved forest of China
6
2009
... 一般认为物种β多样性主要由生态位过程和扩散过程共同作用形成[9-10].按照生态位理论,历史过程决定了区域的物种库,环境因素决定着群落中物种的共存[11].不同的物种有着各自适应的生境,生境差异越大,物种组成差异也越大[12].气候作为重要的环境因子,对不同物种空间更替的作用一直是研究的热点[13-14].除了气候,土壤异质性在小尺度上也是影响植被特征的重要因子[15].中性理论认为,物种随机扩散过程是植物群落构建的重要机制[16].在研究群落周转中,广泛采用地理距离来表征物种扩散限制[17].在扩散限制的影响下,群落物种组成的相似性随着地理距离增加而减小[12].环境过滤和扩散限制对植物群落β多样性的形成至关重要. ...
... β多样性不仅代表着一个地区物种多样性的分布情况,而且还体现着物种与环境之间的关系[3,10,16,29].通过对乌兰布和沙漠植物群落的研究,利用Sørensen相异性指数来表征植物群落β多样性.结果发现该区域植物群落相异性指数较大(均值为0.82,图2),表明该区域植物群落的物种组成之间存在较大差异[30].为了量化植物群落β多样性的过程,将总体β多样性(βsor)分解为周转(βsim)和嵌套(βnes)两部分,发现周转占有很大比例,嵌套比例较低.这与库姆塔格沙漠南缘的植物群落β多样性研究结果一致[30].因此乌兰布和沙漠的植物群落β多样性可能主要受植物群落物种在空间上或群落间的周转影响. ...
... 目前学者们普遍认为扩散限制和生境过滤对植物群落β多样性的形成均起作用[16,10,35].但有研究表明,地理距离对植物群落β多样性的形成的作用不显著[36].本研究结果表明,地理距离的改变对乌兰布和沙漠植物群落β多样性存在显著作用,随着地理距离的增大,植物群落的相异性增大.进一步分析表明,地理距离与植物群落β多样性的相关系数始终低于环境因子的影响.这说明在所研究的范围内,生境条件复杂多变,如梭梭、沙冬青和白刺等群落,其生境中沙粒含量较高;盐爪爪(Kalidium foliatum)、马蔺(Iris lactea)和红砂群落等,其生境中土壤水分、盐分和黏粒含量较高[21].地理距离的改变对植物群落结构有一定的影响,但不是主导因素.在库姆塔格沙漠南缘、古尔班通古特沙漠和新疆艾比湖荒漠植被的研究也证实环境条件是影响荒漠植物群落β多样性的主导因素[10,33-34]. ...
... [10,33-34]. ...
... 一般认为β多样性主要由扩散过程和生态位过程两种机制共同作用形成[9-10].生态位理论认为物种组成差异由生境变化导致,扩散理论认为物种分布格局完全由其扩散能力确定[5].扩散限制和生境过滤并不是互斥关系,有时同时起着作用[37].本文研究表明环境差异和地理距离均与乌兰布和沙漠植物群落β多样性及其组分存在显著的相关性(图3).在荒漠生态系统中,干旱起着强烈的环境过滤作用[38].随着干旱程度的增加,有利于比叶面积小和根系能在短时间获取最大养分和水分的植物生存[39-40].植物(如梭梭、沙拐枣等)会通过减少地上生物量的分配和叶片退化减少水分的散失,并把更多光合作用产物分配给地下部分[41].物种相似性随着地理距离的增大而减小,说明扩散限制在植物群落β多样性中起一定作用.但二者对于研究区植物群落β多样性及其周转组分的影响存在差异,与气候因素、土壤因素的相关系数分别为0.26、0.18,与地理距离的相关系数是0.16(图3).环境筛选作用远大于扩散限制,在荒漠区的库姆塔格沙漠南缘和古尔班通古特沙漠研究中也得到过类似结果[30,34].这说明在环境贫瘠、物种结构简单的乌兰布和沙漠,环境过滤作用对植物群落β多样性的作用大.值得注意的是,在研究区域中物种周转主导植物群落β多样性的变化,且对环境变量的响应与植物群落β多样性一致.因此,随着环境距离(包括气候和土壤等环境条件的差异)和地理距离的增加,空间异质性也明显增强,植物种子的扩散受到限制[42],在长期自然选择和群落演替中,不同生境下形成与之相适应的物种[43].本研究表明,以环境过滤为主,加之扩散限制共同影响着乌兰布和沙漠的植物群落β多样性的格局,即生态位理论和中性理论共同驱动乌兰布和沙漠植物群落的构建. ...
... 本研究虽发现环境过滤和扩散限制是影响植物群落β多样性及周转(βsim)的重要过程,但相关性均较低.这表明存在未测量的因子起着作用[10].如植物群落的自身特性[36,44]、人为干扰[34]、研究尺度[45-47]都会在不同程度上影响植物群落β多样性. ...
Relative contribution of niche and neutral processes on tree species turnover across scales in seasonal forests of NW Argentina
1
2016
... 一般认为物种β多样性主要由生态位过程和扩散过程共同作用形成[9-10].按照生态位理论,历史过程决定了区域的物种库,环境因素决定着群落中物种的共存[11].不同的物种有着各自适应的生境,生境差异越大,物种组成差异也越大[12].气候作为重要的环境因子,对不同物种空间更替的作用一直是研究的热点[13-14].除了气候,土壤异质性在小尺度上也是影响植被特征的重要因子[15].中性理论认为,物种随机扩散过程是植物群落构建的重要机制[16].在研究群落周转中,广泛采用地理距离来表征物种扩散限制[17].在扩散限制的影响下,群落物种组成的相似性随着地理距离增加而减小[12].环境过滤和扩散限制对植物群落β多样性的形成至关重要. ...
群落构建的中性理论和生态位理论
2
2009
... 一般认为物种β多样性主要由生态位过程和扩散过程共同作用形成[9-10].按照生态位理论,历史过程决定了区域的物种库,环境因素决定着群落中物种的共存[11].不同的物种有着各自适应的生境,生境差异越大,物种组成差异也越大[12].气候作为重要的环境因子,对不同物种空间更替的作用一直是研究的热点[13-14].除了气候,土壤异质性在小尺度上也是影响植被特征的重要因子[15].中性理论认为,物种随机扩散过程是植物群落构建的重要机制[16].在研究群落周转中,广泛采用地理距离来表征物种扩散限制[17].在扩散限制的影响下,群落物种组成的相似性随着地理距离增加而减小[12].环境过滤和扩散限制对植物群落β多样性的形成至关重要. ...
... [12].环境过滤和扩散限制对植物群落β多样性的形成至关重要. ...
Alpha,Beta and Gamma diversity differ in response to precipitation in the Inner Mongolia grassland
1
2014
... 一般认为物种β多样性主要由生态位过程和扩散过程共同作用形成[9-10].按照生态位理论,历史过程决定了区域的物种库,环境因素决定着群落中物种的共存[11].不同的物种有着各自适应的生境,生境差异越大,物种组成差异也越大[12].气候作为重要的环境因子,对不同物种空间更替的作用一直是研究的热点[13-14].除了气候,土壤异质性在小尺度上也是影响植被特征的重要因子[15].中性理论认为,物种随机扩散过程是植物群落构建的重要机制[16].在研究群落周转中,广泛采用地理距离来表征物种扩散限制[17].在扩散限制的影响下,群落物种组成的相似性随着地理距离增加而减小[12].环境过滤和扩散限制对植物群落β多样性的形成至关重要. ...
Geographical divergence of species richness and local homogenization of plant assemblages due to climate change in grasslands
1
2022
... 一般认为物种β多样性主要由生态位过程和扩散过程共同作用形成[9-10].按照生态位理论,历史过程决定了区域的物种库,环境因素决定着群落中物种的共存[11].不同的物种有着各自适应的生境,生境差异越大,物种组成差异也越大[12].气候作为重要的环境因子,对不同物种空间更替的作用一直是研究的热点[13-14].除了气候,土壤异质性在小尺度上也是影响植被特征的重要因子[15].中性理论认为,物种随机扩散过程是植物群落构建的重要机制[16].在研究群落周转中,广泛采用地理距离来表征物种扩散限制[17].在扩散限制的影响下,群落物种组成的相似性随着地理距离增加而减小[12].环境过滤和扩散限制对植物群落β多样性的形成至关重要. ...
Feedbacks between nutrient cycling and vegetation predict plant species coexistence and invasion
2
2002
... 一般认为物种β多样性主要由生态位过程和扩散过程共同作用形成[9-10].按照生态位理论,历史过程决定了区域的物种库,环境因素决定着群落中物种的共存[11].不同的物种有着各自适应的生境,生境差异越大,物种组成差异也越大[12].气候作为重要的环境因子,对不同物种空间更替的作用一直是研究的热点[13-14].除了气候,土壤异质性在小尺度上也是影响植被特征的重要因子[15].中性理论认为,物种随机扩散过程是植物群落构建的重要机制[16].在研究群落周转中,广泛采用地理距离来表征物种扩散限制[17].在扩散限制的影响下,群落物种组成的相似性随着地理距离增加而减小[12].环境过滤和扩散限制对植物群落β多样性的形成至关重要. ...
... 一些研究表明,土壤的变化在小尺度上是植被特征的重要决定因子[15].本研究结果表明,土壤因素对植物群落β多样性及周转组分有显著影响,但对嵌套组分影响不显著.相对气候而言,土壤因素相关性较低,这与在古尔班通古特沙漠的研究结果相同[34].这说明在大尺度上,沙漠土壤异质性对植被组成变化影响较小. ...
The unified neutral theory of biodiversity and biogeography:reply
3
2004
... 一般认为物种β多样性主要由生态位过程和扩散过程共同作用形成[9-10].按照生态位理论,历史过程决定了区域的物种库,环境因素决定着群落中物种的共存[11].不同的物种有着各自适应的生境,生境差异越大,物种组成差异也越大[12].气候作为重要的环境因子,对不同物种空间更替的作用一直是研究的热点[13-14].除了气候,土壤异质性在小尺度上也是影响植被特征的重要因子[15].中性理论认为,物种随机扩散过程是植物群落构建的重要机制[16].在研究群落周转中,广泛采用地理距离来表征物种扩散限制[17].在扩散限制的影响下,群落物种组成的相似性随着地理距离增加而减小[12].环境过滤和扩散限制对植物群落β多样性的形成至关重要. ...
... β多样性不仅代表着一个地区物种多样性的分布情况,而且还体现着物种与环境之间的关系[3,10,16,29].通过对乌兰布和沙漠植物群落的研究,利用Sørensen相异性指数来表征植物群落β多样性.结果发现该区域植物群落相异性指数较大(均值为0.82,图2),表明该区域植物群落的物种组成之间存在较大差异[30].为了量化植物群落β多样性的过程,将总体β多样性(βsor)分解为周转(βsim)和嵌套(βnes)两部分,发现周转占有很大比例,嵌套比例较低.这与库姆塔格沙漠南缘的植物群落β多样性研究结果一致[30].因此乌兰布和沙漠的植物群落β多样性可能主要受植物群落物种在空间上或群落间的周转影响. ...
... 目前学者们普遍认为扩散限制和生境过滤对植物群落β多样性的形成均起作用[16,10,35].但有研究表明,地理距离对植物群落β多样性的形成的作用不显著[36].本研究结果表明,地理距离的改变对乌兰布和沙漠植物群落β多样性存在显著作用,随着地理距离的增大,植物群落的相异性增大.进一步分析表明,地理距离与植物群落β多样性的相关系数始终低于环境因子的影响.这说明在所研究的范围内,生境条件复杂多变,如梭梭、沙冬青和白刺等群落,其生境中沙粒含量较高;盐爪爪(Kalidium foliatum)、马蔺(Iris lactea)和红砂群落等,其生境中土壤水分、盐分和黏粒含量较高[21].地理距离的改变对植物群落结构有一定的影响,但不是主导因素.在库姆塔格沙漠南缘、古尔班通古特沙漠和新疆艾比湖荒漠植被的研究也证实环境条件是影响荒漠植物群落β多样性的主导因素[10,33-34]. ...
Dispersal and neutral sampling mediate contingent effects of disturbance on plant beta‐diversity:a meta‐analysis
1
2017
... 一般认为物种β多样性主要由生态位过程和扩散过程共同作用形成[9-10].按照生态位理论,历史过程决定了区域的物种库,环境因素决定着群落中物种的共存[11].不同的物种有着各自适应的生境,生境差异越大,物种组成差异也越大[12].气候作为重要的环境因子,对不同物种空间更替的作用一直是研究的热点[13-14].除了气候,土壤异质性在小尺度上也是影响植被特征的重要因子[15].中性理论认为,物种随机扩散过程是植物群落构建的重要机制[16].在研究群落周转中,广泛采用地理距离来表征物种扩散限制[17].在扩散限制的影响下,群落物种组成的相似性随着地理距离增加而减小[12].环境过滤和扩散限制对植物群落β多样性的形成至关重要. ...
1
2002
... 干旱区、半干旱区占全球陆地面积的25%,是陆地生态系统的重要组成部分.干旱区、半干旱区生态系统脆弱,是研究生物多样性的重点区域[18].荒漠植被的生存和分布受到气候、土壤和水分的影响[19-20].乌兰布和沙漠是中国第八大沙漠,目前对该区域物种组成、群落类型和多样性的调查已经比较充分[21-23].这些研究为乌兰布和沙漠植被资源保护奠定了基础.但是,在乌兰布和沙漠中植物群落β多样性形成机制未见报道.本研究通过分析乌兰布和沙漠植物群落β多样性的格局及影响因素,定量评价样地间气候、土壤和地理距离与植物群落β多样性的关系,以期深入了解该区域植物群落对特殊环境的适应及群落维持机制,为沙漠地区的植物群落β多样性形成和维持机制的研究提供事实证据. ...
Different ecological processes determined the alpha and beta components of taxonomic,functional,and phylogenetic diversity for plant communities in dryland regions of Northwest China
1
2019
... 干旱区、半干旱区占全球陆地面积的25%,是陆地生态系统的重要组成部分.干旱区、半干旱区生态系统脆弱,是研究生物多样性的重点区域[18].荒漠植被的生存和分布受到气候、土壤和水分的影响[19-20].乌兰布和沙漠是中国第八大沙漠,目前对该区域物种组成、群落类型和多样性的调查已经比较充分[21-23].这些研究为乌兰布和沙漠植被资源保护奠定了基础.但是,在乌兰布和沙漠中植物群落β多样性形成机制未见报道.本研究通过分析乌兰布和沙漠植物群落β多样性的格局及影响因素,定量评价样地间气候、土壤和地理距离与植物群落β多样性的关系,以期深入了解该区域植物群落对特殊环境的适应及群落维持机制,为沙漠地区的植物群落β多样性形成和维持机制的研究提供事实证据. ...
Species richness patterns and water-energy dynamics in the drylands of Northwest China
1
2013
... 干旱区、半干旱区占全球陆地面积的25%,是陆地生态系统的重要组成部分.干旱区、半干旱区生态系统脆弱,是研究生物多样性的重点区域[18].荒漠植被的生存和分布受到气候、土壤和水分的影响[19-20].乌兰布和沙漠是中国第八大沙漠,目前对该区域物种组成、群落类型和多样性的调查已经比较充分[21-23].这些研究为乌兰布和沙漠植被资源保护奠定了基础.但是,在乌兰布和沙漠中植物群落β多样性形成机制未见报道.本研究通过分析乌兰布和沙漠植物群落β多样性的格局及影响因素,定量评价样地间气候、土壤和地理距离与植物群落β多样性的关系,以期深入了解该区域植物群落对特殊环境的适应及群落维持机制,为沙漠地区的植物群落β多样性形成和维持机制的研究提供事实证据. ...
乌兰布和沙漠典型灌木群落结构及数量特征
2
2012
... 干旱区、半干旱区占全球陆地面积的25%,是陆地生态系统的重要组成部分.干旱区、半干旱区生态系统脆弱,是研究生物多样性的重点区域[18].荒漠植被的生存和分布受到气候、土壤和水分的影响[19-20].乌兰布和沙漠是中国第八大沙漠,目前对该区域物种组成、群落类型和多样性的调查已经比较充分[21-23].这些研究为乌兰布和沙漠植被资源保护奠定了基础.但是,在乌兰布和沙漠中植物群落β多样性形成机制未见报道.本研究通过分析乌兰布和沙漠植物群落β多样性的格局及影响因素,定量评价样地间气候、土壤和地理距离与植物群落β多样性的关系,以期深入了解该区域植物群落对特殊环境的适应及群落维持机制,为沙漠地区的植物群落β多样性形成和维持机制的研究提供事实证据. ...
... 目前学者们普遍认为扩散限制和生境过滤对植物群落β多样性的形成均起作用[16,10,35].但有研究表明,地理距离对植物群落β多样性的形成的作用不显著[36].本研究结果表明,地理距离的改变对乌兰布和沙漠植物群落β多样性存在显著作用,随着地理距离的增大,植物群落的相异性增大.进一步分析表明,地理距离与植物群落β多样性的相关系数始终低于环境因子的影响.这说明在所研究的范围内,生境条件复杂多变,如梭梭、沙冬青和白刺等群落,其生境中沙粒含量较高;盐爪爪(Kalidium foliatum)、马蔺(Iris lactea)和红砂群落等,其生境中土壤水分、盐分和黏粒含量较高[21].地理距离的改变对植物群落结构有一定的影响,但不是主导因素.在库姆塔格沙漠南缘、古尔班通古特沙漠和新疆艾比湖荒漠植被的研究也证实环境条件是影响荒漠植物群落β多样性的主导因素[10,33-34]. ...
乌兰布和沙漠典型灌木群落多样性及其生态位
1
2020
... 干旱区、半干旱区占全球陆地面积的25%,是陆地生态系统的重要组成部分.干旱区、半干旱区生态系统脆弱,是研究生物多样性的重点区域[18].荒漠植被的生存和分布受到气候、土壤和水分的影响[19-20].乌兰布和沙漠是中国第八大沙漠,目前对该区域物种组成、群落类型和多样性的调查已经比较充分[21-23].这些研究为乌兰布和沙漠植被资源保护奠定了基础.但是,在乌兰布和沙漠中植物群落β多样性形成机制未见报道.本研究通过分析乌兰布和沙漠植物群落β多样性的格局及影响因素,定量评价样地间气候、土壤和地理距离与植物群落β多样性的关系,以期深入了解该区域植物群落对特殊环境的适应及群落维持机制,为沙漠地区的植物群落β多样性形成和维持机制的研究提供事实证据. ...
基于3S技术的乌兰布和沙漠范围和面积分析
1
2015
... 乌兰布和沙漠(海拔1 030—1 474 m)位于内蒙古自治区西部的阿拉善盟和巴彦淖尔市境内,南连贺兰山、吉兰泰盐池,西至阿木乌苏拉乌山、哈乌拉山、浩温多尔山,东临黄河磴口-临河段,北到狼山、河套平原,面积约10 000 km2.沙漠南部多为戈壁、盐碱地和沙山,北部多为半流动和半固定沙地[24-25].地貌以沙漠、山地和戈壁为主.该地区属温带大陆性气候,年降水量110—160 mm,年潜在蒸发量2 400—3 200 mm,降水多在6—9月;年平均气温7.5—8.5 ℃,昼夜温差较大.主要土壤为风沙土[26].优势植物为梭梭(Haloxylon ammodendron)、沙冬青(Ammopiptanthus mongolicus)、白刺(Nitraria tangutorum)、霸王(Sarcozygium xanthoxylon)、红砂(Reaumuria soongarica)和芦苇(Phragmites australis)等. ...
乌兰布和沙漠的形成与环境变化
1
2007
... 乌兰布和沙漠(海拔1 030—1 474 m)位于内蒙古自治区西部的阿拉善盟和巴彦淖尔市境内,南连贺兰山、吉兰泰盐池,西至阿木乌苏拉乌山、哈乌拉山、浩温多尔山,东临黄河磴口-临河段,北到狼山、河套平原,面积约10 000 km2.沙漠南部多为戈壁、盐碱地和沙山,北部多为半流动和半固定沙地[24-25].地貌以沙漠、山地和戈壁为主.该地区属温带大陆性气候,年降水量110—160 mm,年潜在蒸发量2 400—3 200 mm,降水多在6—9月;年平均气温7.5—8.5 ℃,昼夜温差较大.主要土壤为风沙土[26].优势植物为梭梭(Haloxylon ammodendron)、沙冬青(Ammopiptanthus mongolicus)、白刺(Nitraria tangutorum)、霸王(Sarcozygium xanthoxylon)、红砂(Reaumuria soongarica)和芦苇(Phragmites australis)等. ...
1
2018
... 乌兰布和沙漠(海拔1 030—1 474 m)位于内蒙古自治区西部的阿拉善盟和巴彦淖尔市境内,南连贺兰山、吉兰泰盐池,西至阿木乌苏拉乌山、哈乌拉山、浩温多尔山,东临黄河磴口-临河段,北到狼山、河套平原,面积约10 000 km2.沙漠南部多为戈壁、盐碱地和沙山,北部多为半流动和半固定沙地[24-25].地貌以沙漠、山地和戈壁为主.该地区属温带大陆性气候,年降水量110—160 mm,年潜在蒸发量2 400—3 200 mm,降水多在6—9月;年平均气温7.5—8.5 ℃,昼夜温差较大.主要土壤为风沙土[26].优势植物为梭梭(Haloxylon ammodendron)、沙冬青(Ammopiptanthus mongolicus)、白刺(Nitraria tangutorum)、霸王(Sarcozygium xanthoxylon)、红砂(Reaumuria soongarica)和芦苇(Phragmites australis)等. ...
1
2000
... 在调查点的每个小样方中,沿对角线用土钻钻取3钻土,取样深度为10 cm,将每个小样方的土壤样品混合均匀,保证每个调查点有3个土壤样品用于理化性质测定.将混合均匀的土壤样品带回实验室风干,过2 mm土壤筛,剔除土壤中的根系和杂质等.经H2SO4-HClO4法消煮后,用钼锑抗比色法测定全磷(STP)含量;经H2SO4-H2O2法消煮后,运用凯氏定氮法测定全氮(STN);经HF-HClO4法消煮后,运用火焰光度计法测定全钾(STK);有机质(SOM) 采用重铬酸钾氧还原滴定法测定;速效磷(SAP) 测定用碳酸氢钠提取-钼锑抗比色法;速效钾(SAK)测定用醋酸铵浸提-火焰光度法[27]. ...
Betapart:an R package for the study of beta diversity
1
2012
... 本文基于物种有无计算Sørensen相异性指数,使用R语言中betapart包[28]将总体β多样性βsor分解为物种周转(βsim)和嵌套(βnes)两部分. ...
Analyzing beta diversity: partitioning the spatial variation of community composition data
1
2005
... β多样性不仅代表着一个地区物种多样性的分布情况,而且还体现着物种与环境之间的关系[3,10,16,29].通过对乌兰布和沙漠植物群落的研究,利用Sørensen相异性指数来表征植物群落β多样性.结果发现该区域植物群落相异性指数较大(均值为0.82,图2),表明该区域植物群落的物种组成之间存在较大差异[30].为了量化植物群落β多样性的过程,将总体β多样性(βsor)分解为周转(βsim)和嵌套(βnes)两部分,发现周转占有很大比例,嵌套比例较低.这与库姆塔格沙漠南缘的植物群落β多样性研究结果一致[30].因此乌兰布和沙漠的植物群落β多样性可能主要受植物群落物种在空间上或群落间的周转影响. ...
环境过滤和扩散限制对库姆塔格沙漠南缘植物群落β-多样性的影响
3
2021
... β多样性不仅代表着一个地区物种多样性的分布情况,而且还体现着物种与环境之间的关系[3,10,16,29].通过对乌兰布和沙漠植物群落的研究,利用Sørensen相异性指数来表征植物群落β多样性.结果发现该区域植物群落相异性指数较大(均值为0.82,图2),表明该区域植物群落的物种组成之间存在较大差异[30].为了量化植物群落β多样性的过程,将总体β多样性(βsor)分解为周转(βsim)和嵌套(βnes)两部分,发现周转占有很大比例,嵌套比例较低.这与库姆塔格沙漠南缘的植物群落β多样性研究结果一致[30].因此乌兰布和沙漠的植物群落β多样性可能主要受植物群落物种在空间上或群落间的周转影响. ...
... [30].因此乌兰布和沙漠的植物群落β多样性可能主要受植物群落物种在空间上或群落间的周转影响. ...
... 一般认为β多样性主要由扩散过程和生态位过程两种机制共同作用形成[9-10].生态位理论认为物种组成差异由生境变化导致,扩散理论认为物种分布格局完全由其扩散能力确定[5].扩散限制和生境过滤并不是互斥关系,有时同时起着作用[37].本文研究表明环境差异和地理距离均与乌兰布和沙漠植物群落β多样性及其组分存在显著的相关性(图3).在荒漠生态系统中,干旱起着强烈的环境过滤作用[38].随着干旱程度的增加,有利于比叶面积小和根系能在短时间获取最大养分和水分的植物生存[39-40].植物(如梭梭、沙拐枣等)会通过减少地上生物量的分配和叶片退化减少水分的散失,并把更多光合作用产物分配给地下部分[41].物种相似性随着地理距离的增大而减小,说明扩散限制在植物群落β多样性中起一定作用.但二者对于研究区植物群落β多样性及其周转组分的影响存在差异,与气候因素、土壤因素的相关系数分别为0.26、0.18,与地理距离的相关系数是0.16(图3).环境筛选作用远大于扩散限制,在荒漠区的库姆塔格沙漠南缘和古尔班通古特沙漠研究中也得到过类似结果[30,34].这说明在环境贫瘠、物种结构简单的乌兰布和沙漠,环境过滤作用对植物群落β多样性的作用大.值得注意的是,在研究区域中物种周转主导植物群落β多样性的变化,且对环境变量的响应与植物群落β多样性一致.因此,随着环境距离(包括气候和土壤等环境条件的差异)和地理距离的增加,空间异质性也明显增强,植物种子的扩散受到限制[42],在长期自然选择和群落演替中,不同生境下形成与之相适应的物种[43].本研究表明,以环境过滤为主,加之扩散限制共同影响着乌兰布和沙漠的植物群落β多样性的格局,即生态位理论和中性理论共同驱动乌兰布和沙漠植物群落的构建. ...
Relationships between plant spatial patterns,water infiltration capacity,and plant community composition in semi-arid Mediterranean ecosystems along atress gradients
1
2013
... 通过主成分分析,最终筛选出影响乌兰布和沙漠植物群落β多样性的主要气候因子,主要包括年降水量、最湿润月降水量、最湿润季降水量、最干旱季降水量、最干旱月降水量,说明这些气候因子是造成植物群落差异的重要变量.有研究发现,降水是影响干旱区和半干旱区生态系统的重要因子[31-32].在乌兰布和沙漠,降水也是影响本地区植物群落β多样性和周转组分的主要气候因子.我们调查发现,降水差异影响植物的分布,沙冬青群落生长在降水较好的条件下,白沙蒿(Artemisia sphaerocephala)群落的生境条件极为恶劣.此外,水资源的利用方式也会影响植物群落的物种组成,如荒漠区深根系的植物利用的是地下水,浅根系植物利用的是土壤表层水和地下水,水分利用策略也使物种组成有较大差异[33]. ...
Soil water availability shapes species richness in mid-latitude shrub steppe plant communities
1
2020
... 通过主成分分析,最终筛选出影响乌兰布和沙漠植物群落β多样性的主要气候因子,主要包括年降水量、最湿润月降水量、最湿润季降水量、最干旱季降水量、最干旱月降水量,说明这些气候因子是造成植物群落差异的重要变量.有研究发现,降水是影响干旱区和半干旱区生态系统的重要因子[31-32].在乌兰布和沙漠,降水也是影响本地区植物群落β多样性和周转组分的主要气候因子.我们调查发现,降水差异影响植物的分布,沙冬青群落生长在降水较好的条件下,白沙蒿(Artemisia sphaerocephala)群落的生境条件极为恶劣.此外,水资源的利用方式也会影响植物群落的物种组成,如荒漠区深根系的植物利用的是地下水,浅根系植物利用的是土壤表层水和地下水,水分利用策略也使物种组成有较大差异[33]. ...
Characteristics and driving mechanisms of species beta diversity in desert plant communities
2
2021
... 通过主成分分析,最终筛选出影响乌兰布和沙漠植物群落β多样性的主要气候因子,主要包括年降水量、最湿润月降水量、最湿润季降水量、最干旱季降水量、最干旱月降水量,说明这些气候因子是造成植物群落差异的重要变量.有研究发现,降水是影响干旱区和半干旱区生态系统的重要因子[31-32].在乌兰布和沙漠,降水也是影响本地区植物群落β多样性和周转组分的主要气候因子.我们调查发现,降水差异影响植物的分布,沙冬青群落生长在降水较好的条件下,白沙蒿(Artemisia sphaerocephala)群落的生境条件极为恶劣.此外,水资源的利用方式也会影响植物群落的物种组成,如荒漠区深根系的植物利用的是地下水,浅根系植物利用的是土壤表层水和地下水,水分利用策略也使物种组成有较大差异[33]. ...
... 目前学者们普遍认为扩散限制和生境过滤对植物群落β多样性的形成均起作用[16,10,35].但有研究表明,地理距离对植物群落β多样性的形成的作用不显著[36].本研究结果表明,地理距离的改变对乌兰布和沙漠植物群落β多样性存在显著作用,随着地理距离的增大,植物群落的相异性增大.进一步分析表明,地理距离与植物群落β多样性的相关系数始终低于环境因子的影响.这说明在所研究的范围内,生境条件复杂多变,如梭梭、沙冬青和白刺等群落,其生境中沙粒含量较高;盐爪爪(Kalidium foliatum)、马蔺(Iris lactea)和红砂群落等,其生境中土壤水分、盐分和黏粒含量较高[21].地理距离的改变对植物群落结构有一定的影响,但不是主导因素.在库姆塔格沙漠南缘、古尔班通古特沙漠和新疆艾比湖荒漠植被的研究也证实环境条件是影响荒漠植物群落β多样性的主导因素[10,33-34]. ...
古尔班通古特沙漠南部植物群落β多样性及其解释
4
2010
... 一些研究表明,土壤的变化在小尺度上是植被特征的重要决定因子[15].本研究结果表明,土壤因素对植物群落β多样性及周转组分有显著影响,但对嵌套组分影响不显著.相对气候而言,土壤因素相关性较低,这与在古尔班通古特沙漠的研究结果相同[34].这说明在大尺度上,沙漠土壤异质性对植被组成变化影响较小. ...
... 目前学者们普遍认为扩散限制和生境过滤对植物群落β多样性的形成均起作用[16,10,35].但有研究表明,地理距离对植物群落β多样性的形成的作用不显著[36].本研究结果表明,地理距离的改变对乌兰布和沙漠植物群落β多样性存在显著作用,随着地理距离的增大,植物群落的相异性增大.进一步分析表明,地理距离与植物群落β多样性的相关系数始终低于环境因子的影响.这说明在所研究的范围内,生境条件复杂多变,如梭梭、沙冬青和白刺等群落,其生境中沙粒含量较高;盐爪爪(Kalidium foliatum)、马蔺(Iris lactea)和红砂群落等,其生境中土壤水分、盐分和黏粒含量较高[21].地理距离的改变对植物群落结构有一定的影响,但不是主导因素.在库姆塔格沙漠南缘、古尔班通古特沙漠和新疆艾比湖荒漠植被的研究也证实环境条件是影响荒漠植物群落β多样性的主导因素[10,33-34]. ...
... 一般认为β多样性主要由扩散过程和生态位过程两种机制共同作用形成[9-10].生态位理论认为物种组成差异由生境变化导致,扩散理论认为物种分布格局完全由其扩散能力确定[5].扩散限制和生境过滤并不是互斥关系,有时同时起着作用[37].本文研究表明环境差异和地理距离均与乌兰布和沙漠植物群落β多样性及其组分存在显著的相关性(图3).在荒漠生态系统中,干旱起着强烈的环境过滤作用[38].随着干旱程度的增加,有利于比叶面积小和根系能在短时间获取最大养分和水分的植物生存[39-40].植物(如梭梭、沙拐枣等)会通过减少地上生物量的分配和叶片退化减少水分的散失,并把更多光合作用产物分配给地下部分[41].物种相似性随着地理距离的增大而减小,说明扩散限制在植物群落β多样性中起一定作用.但二者对于研究区植物群落β多样性及其周转组分的影响存在差异,与气候因素、土壤因素的相关系数分别为0.26、0.18,与地理距离的相关系数是0.16(图3).环境筛选作用远大于扩散限制,在荒漠区的库姆塔格沙漠南缘和古尔班通古特沙漠研究中也得到过类似结果[30,34].这说明在环境贫瘠、物种结构简单的乌兰布和沙漠,环境过滤作用对植物群落β多样性的作用大.值得注意的是,在研究区域中物种周转主导植物群落β多样性的变化,且对环境变量的响应与植物群落β多样性一致.因此,随着环境距离(包括气候和土壤等环境条件的差异)和地理距离的增加,空间异质性也明显增强,植物种子的扩散受到限制[42],在长期自然选择和群落演替中,不同生境下形成与之相适应的物种[43].本研究表明,以环境过滤为主,加之扩散限制共同影响着乌兰布和沙漠的植物群落β多样性的格局,即生态位理论和中性理论共同驱动乌兰布和沙漠植物群落的构建. ...
... 本研究虽发现环境过滤和扩散限制是影响植物群落β多样性及周转(βsim)的重要过程,但相关性均较低.这表明存在未测量的因子起着作用[10].如植物群落的自身特性[36,44]、人为干扰[34]、研究尺度[45-47]都会在不同程度上影响植物群落β多样性. ...
Habitat associations of trees and shrubs in a 50-ha Neotropical forest plots
1
2001
... 目前学者们普遍认为扩散限制和生境过滤对植物群落β多样性的形成均起作用[16,10,35].但有研究表明,地理距离对植物群落β多样性的形成的作用不显著[36].本研究结果表明,地理距离的改变对乌兰布和沙漠植物群落β多样性存在显著作用,随着地理距离的增大,植物群落的相异性增大.进一步分析表明,地理距离与植物群落β多样性的相关系数始终低于环境因子的影响.这说明在所研究的范围内,生境条件复杂多变,如梭梭、沙冬青和白刺等群落,其生境中沙粒含量较高;盐爪爪(Kalidium foliatum)、马蔺(Iris lactea)和红砂群落等,其生境中土壤水分、盐分和黏粒含量较高[21].地理距离的改变对植物群落结构有一定的影响,但不是主导因素.在库姆塔格沙漠南缘、古尔班通古特沙漠和新疆艾比湖荒漠植被的研究也证实环境条件是影响荒漠植物群落β多样性的主导因素[10,33-34]. ...
古田山10种主要森林群落类型的α和β多样性格局及影响因素
2
2019
... 目前学者们普遍认为扩散限制和生境过滤对植物群落β多样性的形成均起作用[16,10,35].但有研究表明,地理距离对植物群落β多样性的形成的作用不显著[36].本研究结果表明,地理距离的改变对乌兰布和沙漠植物群落β多样性存在显著作用,随着地理距离的增大,植物群落的相异性增大.进一步分析表明,地理距离与植物群落β多样性的相关系数始终低于环境因子的影响.这说明在所研究的范围内,生境条件复杂多变,如梭梭、沙冬青和白刺等群落,其生境中沙粒含量较高;盐爪爪(Kalidium foliatum)、马蔺(Iris lactea)和红砂群落等,其生境中土壤水分、盐分和黏粒含量较高[21].地理距离的改变对植物群落结构有一定的影响,但不是主导因素.在库姆塔格沙漠南缘、古尔班通古特沙漠和新疆艾比湖荒漠植被的研究也证实环境条件是影响荒漠植物群落β多样性的主导因素[10,33-34]. ...
... 本研究虽发现环境过滤和扩散限制是影响植物群落β多样性及周转(βsim)的重要过程,但相关性均较低.这表明存在未测量的因子起着作用[10].如植物群落的自身特性[36,44]、人为干扰[34]、研究尺度[45-47]都会在不同程度上影响植物群落β多样性. ...
Dispersal,environment and floristic variation of western Amazonian forests
1
2003
... 一般认为β多样性主要由扩散过程和生态位过程两种机制共同作用形成[9-10].生态位理论认为物种组成差异由生境变化导致,扩散理论认为物种分布格局完全由其扩散能力确定[5].扩散限制和生境过滤并不是互斥关系,有时同时起着作用[37].本文研究表明环境差异和地理距离均与乌兰布和沙漠植物群落β多样性及其组分存在显著的相关性(图3).在荒漠生态系统中,干旱起着强烈的环境过滤作用[38].随着干旱程度的增加,有利于比叶面积小和根系能在短时间获取最大养分和水分的植物生存[39-40].植物(如梭梭、沙拐枣等)会通过减少地上生物量的分配和叶片退化减少水分的散失,并把更多光合作用产物分配给地下部分[41].物种相似性随着地理距离的增大而减小,说明扩散限制在植物群落β多样性中起一定作用.但二者对于研究区植物群落β多样性及其周转组分的影响存在差异,与气候因素、土壤因素的相关系数分别为0.26、0.18,与地理距离的相关系数是0.16(图3).环境筛选作用远大于扩散限制,在荒漠区的库姆塔格沙漠南缘和古尔班通古特沙漠研究中也得到过类似结果[30,34].这说明在环境贫瘠、物种结构简单的乌兰布和沙漠,环境过滤作用对植物群落β多样性的作用大.值得注意的是,在研究区域中物种周转主导植物群落β多样性的变化,且对环境变量的响应与植物群落β多样性一致.因此,随着环境距离(包括气候和土壤等环境条件的差异)和地理距离的增加,空间异质性也明显增强,植物种子的扩散受到限制[42],在长期自然选择和群落演替中,不同生境下形成与之相适应的物种[43].本研究表明,以环境过滤为主,加之扩散限制共同影响着乌兰布和沙漠的植物群落β多样性的格局,即生态位理论和中性理论共同驱动乌兰布和沙漠植物群落的构建. ...
Global ecosystem thresholds driven by aridity
1
2020
... 一般认为β多样性主要由扩散过程和生态位过程两种机制共同作用形成[9-10].生态位理论认为物种组成差异由生境变化导致,扩散理论认为物种分布格局完全由其扩散能力确定[5].扩散限制和生境过滤并不是互斥关系,有时同时起着作用[37].本文研究表明环境差异和地理距离均与乌兰布和沙漠植物群落β多样性及其组分存在显著的相关性(图3).在荒漠生态系统中,干旱起着强烈的环境过滤作用[38].随着干旱程度的增加,有利于比叶面积小和根系能在短时间获取最大养分和水分的植物生存[39-40].植物(如梭梭、沙拐枣等)会通过减少地上生物量的分配和叶片退化减少水分的散失,并把更多光合作用产物分配给地下部分[41].物种相似性随着地理距离的增大而减小,说明扩散限制在植物群落β多样性中起一定作用.但二者对于研究区植物群落β多样性及其周转组分的影响存在差异,与气候因素、土壤因素的相关系数分别为0.26、0.18,与地理距离的相关系数是0.16(图3).环境筛选作用远大于扩散限制,在荒漠区的库姆塔格沙漠南缘和古尔班通古特沙漠研究中也得到过类似结果[30,34].这说明在环境贫瘠、物种结构简单的乌兰布和沙漠,环境过滤作用对植物群落β多样性的作用大.值得注意的是,在研究区域中物种周转主导植物群落β多样性的变化,且对环境变量的响应与植物群落β多样性一致.因此,随着环境距离(包括气候和土壤等环境条件的差异)和地理距离的增加,空间异质性也明显增强,植物种子的扩散受到限制[42],在长期自然选择和群落演替中,不同生境下形成与之相适应的物种[43].本研究表明,以环境过滤为主,加之扩散限制共同影响着乌兰布和沙漠的植物群落β多样性的格局,即生态位理论和中性理论共同驱动乌兰布和沙漠植物群落的构建. ...
Causes and consequences of variation in leaf mass per area (LMA):a meta-analysis
1
2010
... 一般认为β多样性主要由扩散过程和生态位过程两种机制共同作用形成[9-10].生态位理论认为物种组成差异由生境变化导致,扩散理论认为物种分布格局完全由其扩散能力确定[5].扩散限制和生境过滤并不是互斥关系,有时同时起着作用[37].本文研究表明环境差异和地理距离均与乌兰布和沙漠植物群落β多样性及其组分存在显著的相关性(图3).在荒漠生态系统中,干旱起着强烈的环境过滤作用[38].随着干旱程度的增加,有利于比叶面积小和根系能在短时间获取最大养分和水分的植物生存[39-40].植物(如梭梭、沙拐枣等)会通过减少地上生物量的分配和叶片退化减少水分的散失,并把更多光合作用产物分配给地下部分[41].物种相似性随着地理距离的增大而减小,说明扩散限制在植物群落β多样性中起一定作用.但二者对于研究区植物群落β多样性及其周转组分的影响存在差异,与气候因素、土壤因素的相关系数分别为0.26、0.18,与地理距离的相关系数是0.16(图3).环境筛选作用远大于扩散限制,在荒漠区的库姆塔格沙漠南缘和古尔班通古特沙漠研究中也得到过类似结果[30,34].这说明在环境贫瘠、物种结构简单的乌兰布和沙漠,环境过滤作用对植物群落β多样性的作用大.值得注意的是,在研究区域中物种周转主导植物群落β多样性的变化,且对环境变量的响应与植物群落β多样性一致.因此,随着环境距离(包括气候和土壤等环境条件的差异)和地理距离的增加,空间异质性也明显增强,植物种子的扩散受到限制[42],在长期自然选择和群落演替中,不同生境下形成与之相适应的物种[43].本研究表明,以环境过滤为主,加之扩散限制共同影响着乌兰布和沙漠的植物群落β多样性的格局,即生态位理论和中性理论共同驱动乌兰布和沙漠植物群落的构建. ...
Rooting depths,lateral root spreads and below-ground/above-ground allometries of plants in water-limited ecosystems
1
2002
... 一般认为β多样性主要由扩散过程和生态位过程两种机制共同作用形成[9-10].生态位理论认为物种组成差异由生境变化导致,扩散理论认为物种分布格局完全由其扩散能力确定[5].扩散限制和生境过滤并不是互斥关系,有时同时起着作用[37].本文研究表明环境差异和地理距离均与乌兰布和沙漠植物群落β多样性及其组分存在显著的相关性(图3).在荒漠生态系统中,干旱起着强烈的环境过滤作用[38].随着干旱程度的增加,有利于比叶面积小和根系能在短时间获取最大养分和水分的植物生存[39-40].植物(如梭梭、沙拐枣等)会通过减少地上生物量的分配和叶片退化减少水分的散失,并把更多光合作用产物分配给地下部分[41].物种相似性随着地理距离的增大而减小,说明扩散限制在植物群落β多样性中起一定作用.但二者对于研究区植物群落β多样性及其周转组分的影响存在差异,与气候因素、土壤因素的相关系数分别为0.26、0.18,与地理距离的相关系数是0.16(图3).环境筛选作用远大于扩散限制,在荒漠区的库姆塔格沙漠南缘和古尔班通古特沙漠研究中也得到过类似结果[30,34].这说明在环境贫瘠、物种结构简单的乌兰布和沙漠,环境过滤作用对植物群落β多样性的作用大.值得注意的是,在研究区域中物种周转主导植物群落β多样性的变化,且对环境变量的响应与植物群落β多样性一致.因此,随着环境距离(包括气候和土壤等环境条件的差异)和地理距离的增加,空间异质性也明显增强,植物种子的扩散受到限制[42],在长期自然选择和群落演替中,不同生境下形成与之相适应的物种[43].本研究表明,以环境过滤为主,加之扩散限制共同影响着乌兰布和沙漠的植物群落β多样性的格局,即生态位理论和中性理论共同驱动乌兰布和沙漠植物群落的构建. ...
柴达木盆地荒漠植物功能性状及其对环境因子的响应
1
2022
... 一般认为β多样性主要由扩散过程和生态位过程两种机制共同作用形成[9-10].生态位理论认为物种组成差异由生境变化导致,扩散理论认为物种分布格局完全由其扩散能力确定[5].扩散限制和生境过滤并不是互斥关系,有时同时起着作用[37].本文研究表明环境差异和地理距离均与乌兰布和沙漠植物群落β多样性及其组分存在显著的相关性(图3).在荒漠生态系统中,干旱起着强烈的环境过滤作用[38].随着干旱程度的增加,有利于比叶面积小和根系能在短时间获取最大养分和水分的植物生存[39-40].植物(如梭梭、沙拐枣等)会通过减少地上生物量的分配和叶片退化减少水分的散失,并把更多光合作用产物分配给地下部分[41].物种相似性随着地理距离的增大而减小,说明扩散限制在植物群落β多样性中起一定作用.但二者对于研究区植物群落β多样性及其周转组分的影响存在差异,与气候因素、土壤因素的相关系数分别为0.26、0.18,与地理距离的相关系数是0.16(图3).环境筛选作用远大于扩散限制,在荒漠区的库姆塔格沙漠南缘和古尔班通古特沙漠研究中也得到过类似结果[30,34].这说明在环境贫瘠、物种结构简单的乌兰布和沙漠,环境过滤作用对植物群落β多样性的作用大.值得注意的是,在研究区域中物种周转主导植物群落β多样性的变化,且对环境变量的响应与植物群落β多样性一致.因此,随着环境距离(包括气候和土壤等环境条件的差异)和地理距离的增加,空间异质性也明显增强,植物种子的扩散受到限制[42],在长期自然选择和群落演替中,不同生境下形成与之相适应的物种[43].本研究表明,以环境过滤为主,加之扩散限制共同影响着乌兰布和沙漠的植物群落β多样性的格局,即生态位理论和中性理论共同驱动乌兰布和沙漠植物群落的构建. ...
汾河中上游湿地植被β多样性
1
2016
... 一般认为β多样性主要由扩散过程和生态位过程两种机制共同作用形成[9-10].生态位理论认为物种组成差异由生境变化导致,扩散理论认为物种分布格局完全由其扩散能力确定[5].扩散限制和生境过滤并不是互斥关系,有时同时起着作用[37].本文研究表明环境差异和地理距离均与乌兰布和沙漠植物群落β多样性及其组分存在显著的相关性(图3).在荒漠生态系统中,干旱起着强烈的环境过滤作用[38].随着干旱程度的增加,有利于比叶面积小和根系能在短时间获取最大养分和水分的植物生存[39-40].植物(如梭梭、沙拐枣等)会通过减少地上生物量的分配和叶片退化减少水分的散失,并把更多光合作用产物分配给地下部分[41].物种相似性随着地理距离的增大而减小,说明扩散限制在植物群落β多样性中起一定作用.但二者对于研究区植物群落β多样性及其周转组分的影响存在差异,与气候因素、土壤因素的相关系数分别为0.26、0.18,与地理距离的相关系数是0.16(图3).环境筛选作用远大于扩散限制,在荒漠区的库姆塔格沙漠南缘和古尔班通古特沙漠研究中也得到过类似结果[30,34].这说明在环境贫瘠、物种结构简单的乌兰布和沙漠,环境过滤作用对植物群落β多样性的作用大.值得注意的是,在研究区域中物种周转主导植物群落β多样性的变化,且对环境变量的响应与植物群落β多样性一致.因此,随着环境距离(包括气候和土壤等环境条件的差异)和地理距离的增加,空间异质性也明显增强,植物种子的扩散受到限制[42],在长期自然选择和群落演替中,不同生境下形成与之相适应的物种[43].本研究表明,以环境过滤为主,加之扩散限制共同影响着乌兰布和沙漠的植物群落β多样性的格局,即生态位理论和中性理论共同驱动乌兰布和沙漠植物群落的构建. ...
地理距离及环境差异对云南元江干热河谷植物群落beta多样性的影响
1
2016
... 一般认为β多样性主要由扩散过程和生态位过程两种机制共同作用形成[9-10].生态位理论认为物种组成差异由生境变化导致,扩散理论认为物种分布格局完全由其扩散能力确定[5].扩散限制和生境过滤并不是互斥关系,有时同时起着作用[37].本文研究表明环境差异和地理距离均与乌兰布和沙漠植物群落β多样性及其组分存在显著的相关性(图3).在荒漠生态系统中,干旱起着强烈的环境过滤作用[38].随着干旱程度的增加,有利于比叶面积小和根系能在短时间获取最大养分和水分的植物生存[39-40].植物(如梭梭、沙拐枣等)会通过减少地上生物量的分配和叶片退化减少水分的散失,并把更多光合作用产物分配给地下部分[41].物种相似性随着地理距离的增大而减小,说明扩散限制在植物群落β多样性中起一定作用.但二者对于研究区植物群落β多样性及其周转组分的影响存在差异,与气候因素、土壤因素的相关系数分别为0.26、0.18,与地理距离的相关系数是0.16(图3).环境筛选作用远大于扩散限制,在荒漠区的库姆塔格沙漠南缘和古尔班通古特沙漠研究中也得到过类似结果[30,34].这说明在环境贫瘠、物种结构简单的乌兰布和沙漠,环境过滤作用对植物群落β多样性的作用大.值得注意的是,在研究区域中物种周转主导植物群落β多样性的变化,且对环境变量的响应与植物群落β多样性一致.因此,随着环境距离(包括气候和土壤等环境条件的差异)和地理距离的增加,空间异质性也明显增强,植物种子的扩散受到限制[42],在长期自然选择和群落演替中,不同生境下形成与之相适应的物种[43].本研究表明,以环境过滤为主,加之扩散限制共同影响着乌兰布和沙漠的植物群落β多样性的格局,即生态位理论和中性理论共同驱动乌兰布和沙漠植物群落的构建. ...
百山祖自然保护区植物群落beta多样性
1
2013
... 本研究虽发现环境过滤和扩散限制是影响植物群落β多样性及周转(βsim)的重要过程,但相关性均较低.这表明存在未测量的因子起着作用[10].如植物群落的自身特性[36,44]、人为干扰[34]、研究尺度[45-47]都会在不同程度上影响植物群落β多样性. ...
Phylogenetic structure of floridian plant communities depends on taxonomic and spatial scale
1
2006
... 本研究虽发现环境过滤和扩散限制是影响植物群落β多样性及周转(βsim)的重要过程,但相关性均较低.这表明存在未测量的因子起着作用[10].如植物群落的自身特性[36,44]、人为干扰[34]、研究尺度[45-47]都会在不同程度上影响植物群落β多样性. ...
Spatial scale and cross-taxon congruence of terrestrial vertebrate and vascular plant species richness in China
2010
Phylogenetic beta diversity in tropical forests:implications for the roles of geographical and environmental distance
1
2013
... 本研究虽发现环境过滤和扩散限制是影响植物群落β多样性及周转(βsim)的重要过程,但相关性均较低.这表明存在未测量的因子起着作用[10].如植物群落的自身特性[36,44]、人为干扰[34]、研究尺度[45-47]都会在不同程度上影响植物群落β多样性. ...





 甘公网安备 62010202000688号
甘公网安备 62010202000688号
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}