


0 引言
荒漠先锋植物沙芥是十字花科沙芥属一年生或二年生草本植物,主要在甘肃、宁夏、内蒙古等地的沙漠或草原沙地分布,是中国特有物种[1]。根据短角果的形态及植株特征,沙芥属植物至少分为5个种。近年来,借助于形态学观察和分子遗传学方法,沙芥属植物总体分为沙芥(Pugionium cornutum)和斧翅沙芥(P. dolabratum)2个种,距果沙芥(P. calcaratum)、鸡冠沙芥(P. cristatum)和翅果沙芥(P. pterocarpum)属于斧翅沙芥[2]。沙芥属植物具有较高的食用和药用价值。沙芥富含粗纤维、蛋白质、维生素C及多种矿物质微量元素,肉质多浆、风味独特,长期以来作为蔬菜被沙区人民种植和食用[3];沙芥味辛性温,可用于治疗消化不良、食物中毒等症状,其根味辛性凉,具有止咳、清肺热等疗效[1]。因此沙芥属植物的资源开发和机制研究具有重要的生态和经济效益。
沙芥和斧翅沙芥在距今9—22.8万年开始分离,分别进化出不同的习性来适应水分缺失的环境[4]。在地理分布方面,沙芥主要分布在草原沙地、半固定沙漠或流动沙漠,而斧翅沙芥主要分布于固定或半固定沙漠[2]。在形态方面,斧翅沙芥的叶片更窄而沙芥的茎更长,较窄的叶片可减少斧翅沙芥因蒸腾作用造成的水分损失,较长的茎可防止沙芥被流沙掩埋,反映出进化的适应性[2,4-5]。在胁迫耐受方面,沙芥种子萌发对NaCl的极限耐受浓度为300 mmol·L-1,显著高于斧翅沙芥的150 mmol·L-1,表明沙芥对高盐胁迫具有更高的耐受能力[6-7];相同干旱条件下,斧翅沙芥的抗坏血酸与谷胱甘肽含量及抗氧化物酶活性均高于沙芥,表明斧翅沙芥清除活性氧的能力更强,干旱胁迫耐受能力强于沙芥[8-9]。
然而,斧翅沙芥耐受干旱胁迫的遗传机制尚不清楚,本研究以斧翅沙芥叶片为材料,采用Illumina HiSeq高通量测序技术,获得干旱胁迫前后的基因表达谱信息并进行比较分析、GO功能富集和KEGG通路富集,以期挖掘斧翅沙芥干旱胁迫条件下差异表达基因特征,为进一步系统研究其干旱胁迫耐受机制提供数据支持。
1 材料与方法
1.1 材料
1.2 方法
1.2.1 干旱胁迫条件
盆栽的斧翅沙芥幼苗随机分成对照组和干旱处理组2组,每组3个生物学平行(n=20)。参考沙芥的干旱胁迫处理方法[5],采用重度干旱条件为本研究干旱条件,即土壤含水量为田间持水量的75%—85%为对照组,土壤含水量为田间持水量的30%—40%为干旱处理组。对照组和干旱处理组分别收集第3叶片或第4叶片,液氮速冻,-80 ℃下保存。
1.2.2 RNA提取与测序
cDNA文库构建和测序服务委托北京诺禾致源科技股份有限公司进行。斧翅沙芥叶片总RNA提取后,加入带有Oligo(dT)的磁珠富集mRNA,将富集的mRNA加入fragmentation buffer打断,并以短片段化的mRNA为模板,六碱基随机引物为引物,通过逆转录PCR合成双链cDNA。双链cDNA纯化后,分别进行末端修复、加poly(A)尾巴、连接测序接头后,通过PCR扩增和再次纯化获得cDNA文库。随后进行Illumina HiSeq测序。
1.2.3 序列组装和注释
测序获得的原始测序序列(raw reads)按照质量值Qphred≤20的碱基数不超过总碱基数50%的标准过滤后得到clean reads,使用Trinity进行从头组装并使用Corset进行层次聚类,获得unigene。为获得基因功能信息,将Unigene与Nr(
1.2.4 差异表达基因分析
采用RSEM软件将每个样本的clean reads与Trinity拼接得到的转录组作为参考序列做比对,每个基因的相对表达量用RPKM(Reads per kilo bases per million reads)方法计算。采用DESeq方法筛选差异表达基因(阈值padj<0.05),FDR(false discovery rate)≤0.05并且log2 X[X为基因差异位数(Fold Change)]的绝对值≥1的Unigene被认为是具有统计学显著性意义的差异表达基因。注释过的基因分别按照GO三个大类(生物过程、细胞组分和分子功能)和参与的KEGG代谢通路进行GO功能显著性富集分析和KEGG通路显著性富集分析。
2 结果与分析
2.1 转录组数据质量评估
通过对Illumina测序数据进行质量评估,斧翅沙芥对照组(CK)和干旱处理组(DT)各3个生物学重复得到的平均clean reads数分别为51 979 319和49 043 718;各样本Q20均大于98%,Q30均大于95%(表1),表明测序结果质量可靠。通过Trinity拼接和Corset层次聚类,6个样本共获得154 875个unigenes,其中67 006(43.26%)个unigenes长度为200—1 000 nt,52 101(33.64%)个unigenes长度为1 000—2 000 nt,35 768(23.09%)个unigenes长度大于2 000 nt。Unigenes的平均长度和N50分别为1 437 nt和1 956 nt。
表1 测序数据概况
Table 1
| 样品 | 过滤数据 读段数量 | 质量值≥20碱基 百分比 Q20/% | 质量值≥30碱基 百分比 Q30/% |
|---|---|---|---|
| CK1 | 47 083 896 | 98.43 | 95.71 |
| CK2 | 51 089 578 | 98.14 | 95.04 |
| CK3 | 57 764 482 | 98.14 | 95.06 |
| DT1 | 47 438 914 | 98.26 | 95.38 |
| DT2 | 46 524 568 | 98.36 | 95.58 |
| DT3 | 53 167 672 | 98.21 | 95.22 |
2.2 基因功能注释
将获得的unigenes与7个数据库比对,136 667(88.24%)个unigenes在Nt数据库得到注释,136 008(87.81%)个unigenes在Nr数据库得到注释,在Pfam、KOG、Swiss-prot、GO和KEGG数据库分别注释了95 298(61.53%)、43 162(27.86%)、111 119(71.74%)、99 810(64.44%)和57 349(38.24%)个unigenes。
2.3 斧翅沙芥干旱胁迫差异表达基因分析
为鉴定斧翅沙芥干旱胁迫应答基因,我们将对照组和干旱处理组的斧翅沙芥叶片表达基因进行差异表达基因(differentially expressed gene,DEG)分析。对照组和干旱处理组之间共获得13 855个DEGs,其中7 182个DEGs上调表达,6 673个DEGs下调表达(图1)。上调表达基因数高于下调表达基因数。
图1
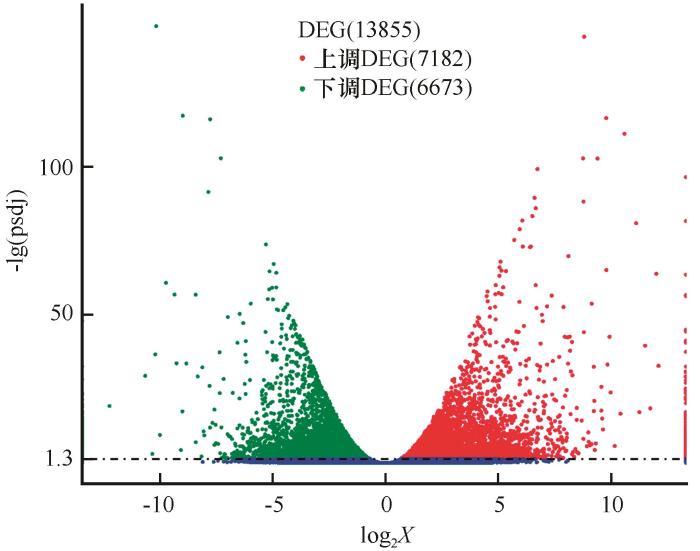
图1
斧翅沙芥干旱胁迫差异表达基因(DEG)的表达分析
Fig.1
Expression analysis of differentially expressed genes in Pugionium dolabratum under drought treatment
2.4 斧翅沙芥干旱胁迫差异表达基因的GO富集
为分析斧翅沙芥干旱胁迫后差异表达基因的功能,我们对获得的DEGs进行GO功能富集分析(图2)。这些DEGs富集到生物过程、细胞组分和分子功能3个大类的59个小类中:富集到生物过程中的DEGs主要集中在单一生物代谢过程(3 474)、氧化还原过程(1 346)、单一生物合成过程(1 281)、氧乙酸代谢过程(1 144)、有机酸代谢过程(1 144)、羧酸代谢过程(1 123)、碳水化合物代谢过程(917)和脂质代谢过程(851);富集到细胞组分中的DEGs主要集中在质体(251)、叶绿体(246)、类囊体(204)、类囊体部分(185)、光合膜(177)和光系统(150);富集到分子功能中的DEGs主要集中在催化活性(5 981)、氧化还原酶活性(1 362)。
图2
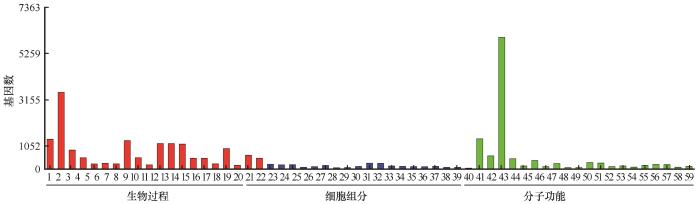
图2
斧翅沙芥干旱胁迫差异表达基因的GO富集分析
1:氧化还原过程;2:单一生物代谢过程;3:脂质代谢过程;4:单羧酸代谢过程;5:光合作用;6:单羧酸生物合成过程;7:脂肪酸代谢过程;8:单一生物合成过程;9:脂质生物合成过程;10:脂肪酸生物合成过程;11:氧乙酸代谢过程;12:有机酸代谢过程;13:羧酸代谢过程;14:有机酸生物合成过程;15:羧酸生物合成过程;16:细胞醛代谢过程;17:碳水化合物代谢过程;18:甘油醛-3-磷酸代谢过程;19:细胞脂质代谢过程;20:化学反应;21:类囊体;22:光合膜;23:类囊体部分;24:光系统II放氧复合物;25:类囊体膜;26:光系统;27:光系统I;28:质外体;29:光系统II;30:质体;31:叶绿体;32:外包裹结构;33:叶绿体部分;34:细胞壁;35:氧化还原酶复合物;36:质体部分;37:叶绿体包膜;38:质体包膜;39:系统I反应中心;40:氧化还原酶活性;41:辅因子结合;42:催化活性;43:辅酶结合;44:磷酸吡哆醛结合;45:裂解酶活性;46:单加氧酶活性;47:氧化还原酶活性,作用于成对供体,分子氧掺入或减少;48:转氨酶活性;49:转移酶活性,转移含氮基团;50:氧化还原酶活性,作用于供体的CH-OH基团;51:氧化还原酶活性,作用于供体的CH-OH基团,NAD或NADP作为受体;52:氧化还原酶活性,作用于供体的醛基;53:抗氧化活性;54:氧化还原酶活性,作用于供体的醛基,NAD或NADP作为受体;55:碳碳裂解酶活性;56:四吡咯结合;57:亚铁原卟啉结合;58:氧化还原酶活性,作用于单供体并结合分子氧;59:羧基裂解酶活性 1: oxidation-reduction process; 2: single-organism metabolic process; 3: lipid metabolic process; 4: monocarboxylic acid metabolic process; 5: photosynthesis; 6: monocarboxylic acid biosynthetic process; 7: fatty acid metabolic process; 8: single-organism biosynthetic process; 9: lipid biosynthetic process; 10: fatty acid biosynthetic process; 11: oxoacid metabolic process; 12: organic acid metabolic process; 13: carboxylic acid metabolic process; 14: organic acid biosynthetic process; 15: carboxylic acid biosynthetic process; 16: cellular aldehyde metabolic process; 17: carbohydrate metabolic process; 18: glyceraldehyde-3-phosphate metabolic process; 19: cellular lipid metabolic process; 20: response to chemical; 21: thylakoid; 22: photosynthetic membrane; 23: thylakoid part; 24: photosystem II oxygen evolving complex; 25: thylakoid membrane; 26: photosystem; 27: photosystem I; 28: apoplast; 29: photosystem II; 30: plastid; 31: chloroplast; 32: external encapsulating structure; 33: chloroplast part; 34: cell wall; 35: oxidoreductase complex; 36: plastid part; 37: chloroplast envelope; 38: plastid envelope; 39: photosystem I reaction center; 40: oxidoreductase activity; 41: cofactor binding; 42: catalytic activity; 43: coenzyme binding; 44: pyridoxal phosphate binding; 45: lyase activity; 46: monooxygenase activity; 47: oxidoreductase activity, acting on paired donors, with incorporation or reduction of molecular oxygen; 48: transaminase activity; 49: transferase activity, transferring nitrogenous groups; 50: oxidoreductase activity, acting on CH-OH group of donors; 51: oxidoreductase activity, acting on the CH-OH group of donors, NAD or NADP as acceptor; 52: oxidoreductase activity, acting on the aldehyde group of donors; 53: antioxidant activity; 54: oxidoreductase activity, acting on the aldehyde group of donors, NAD or NADP as acceptor; 55: carbon-carbon lyase activity; 56: tetrapyrrole binding; 57: heme binding; 58: oxidoreductase activity, acting on single donors with incorporation of molecular oxygen; 59: carboxy-lyase activity
Fig.2
Gene Ontology Enrichment Analysis of differentially expressed genes in Pugionium dolabratum under drought treatment
2.5 斧翅沙芥干旱胁迫差异表达基因的KEGG通路富集
对获得的DEGs进行KEGG通路富集分析的结果表明,采用P值小于0.05的通路定义为显著富集的KEGG通路,共有3 237个DEGs富集到38个KEGG通路中。其中,258个DEGs富集到植物激素信号转导,246个富集到淀粉和蔗糖代谢,218个富集到乙醛酸和二羧酸代谢,159个富集到光合生物的碳固定,156个富集到甘氨酸、丝氨酸和苏氨酸代谢(图3)。
图3
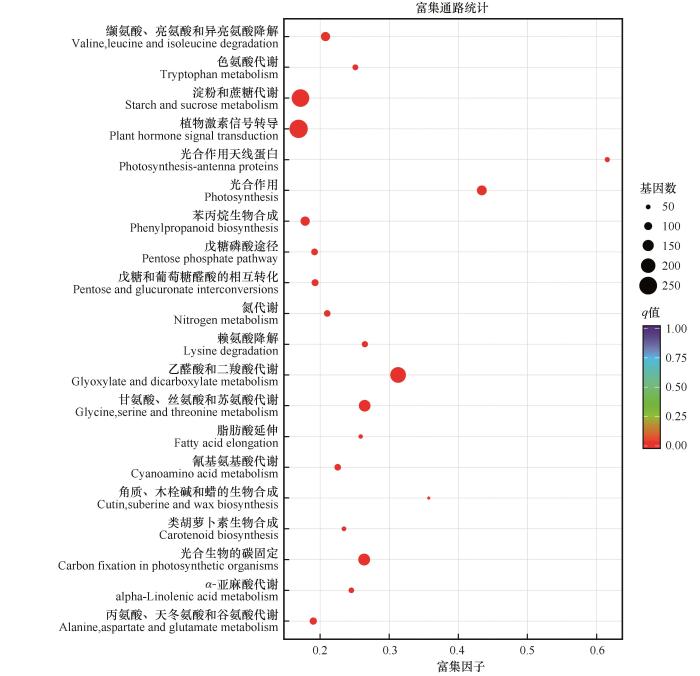
图3
斧翅沙芥干旱胁迫差异表达基因的KEGG富集分析
Fig.3
KEGG enrichment analysis of differentially expressed genes in Pugionium dolabratum under drought treatment
2.6 斧翅沙芥干旱胁迫差异表达转录因子分析
转录因子广泛参与了植物干旱等非生物胁迫应答的调控。基于Swiss-Prot注释,与对照组相比,斧翅沙芥干旱胁迫后共有47个转录因子家族的459个DEGs表达量发生显著性变化,其中277个DEGs上调表达, 182个DEGs下调表达。含有差异表达DEGs最多的10个转录因子家族分别为HSF、ERF、WRKY、MYB、bHLH、NF-Y、DIVARICATA、Trihelix、TCP、AP2/ERF(表2)。干旱处理后,DEGs上调表达数目最多的转录因子家族是热胁迫转录因子家族,该家族中有40个成员的表达水平显著上调,另有7个成员表达水平下调(表2和图4),表明热胁迫转录因子可能参与了斧翅沙芥干旱胁迫应答基因表达的调控。
表2 干旱胁迫下差异表达转录因子数目
Table 2
| 转录因子家族 | 上调 | 下调 |
|---|---|---|
| Heat stress transcription factor | 40 | 7 |
| Ethylene-responsive transcription factor | 38 | 17 |
| WRKY transcription factor | 36 | 21 |
| MYB family transcription factor | 33 | 15 |
| bHLH transcription factor | 30 | 27 |
| Nuclear transcription factor Y | 17 | 6 |
| DIVARICATA transcription factor | 13 | 8 |
| Trihelix transcription factor | 10 | 6 |
| TCP transcription factor | 6 | 10 |
| AP2-like ethylene-responsive transcription factor | 4 | 12 |
图4

图4
干旱胁迫差异表达的热胁迫转录因子家族成员表达模式分析
Fig.4
Expression pattern of differentially expressed heat stress transcription factors under drought treatment
3 讨论与结论
沙芥属植物兼具较高的经济开发和生态保护功能,既可作为食用蔬菜、中草药和饲用牧草,又可作为生态屏障用于防风固沙、水土保持[11]。近年来,其适应性更广、抗逆性更强、营养更丰富的新品种不断被培育出来[12]。已有研究表明,沙芥通过调控参与光合作用、氮代谢、植物激素信号转导途径,尤其是抗坏血酸代谢途径相关基因的表达,提高了沙芥抗氧化能力进而增强其对干旱胁迫的耐受能力[5]。同时,沙芥通过在植株中积累氯离子和上调促氯离子积累、抗氧化、光合作用、碳固定相关基因的表达,增强其渗透调节能力、活性氧清除能力和碳同化效率,从而提高沙芥对盐胁迫的耐受能力[13-14]。生理试验发现,与沙芥相比,近缘种斧翅沙芥具有更强的活性氧清除和干旱胁迫耐受能力,然而目前对斧翅沙芥抗旱机制的研究较少。
本研究利用Illumina测序技术高通量筛选斧翅沙芥干旱胁迫应答基因,共获得13 855个DEGs,GO功能富集分析表明这些DEGs显著富集到物质代谢、氧化还原和催化活性等功能,KEGG通路富集分析表明这些DEGs显著富集到氨基酸代谢、淀粉和糖代谢以及植物激素信号转导等途径,暗示这些功能和通路在斧翅沙芥干旱胁迫耐受中发挥调控作用,尤其是与沙芥干旱胁迫DEGs相比,斧翅沙芥参与淀粉和糖代谢等的DEGs显著增多,这可能通过渗透调节和能量代谢两方面功能促使斧翅沙芥具有更强的干旱胁迫耐受能力。
本研究利用转录组分析技术对干旱处理条件下的斧翅沙芥幼苗叶片进行高通量测序分析,筛选得到大量显著富集到物质代谢、氧化还原和催化活性等功能和参与氨基酸代谢、淀粉和糖代谢以及植物激素信号转导途径的DEGs,并分析了可能参与斧翅沙芥干旱胁迫调控的转录因子家族。这些结果初步揭示了斧翅沙芥对抗干旱胁迫的调控特征,为进一步理解斧翅沙芥干旱环境适应的分子机制提供线索。建议进一步深入研究热胁迫转录因子在斧翅沙芥干旱胁迫后上调表达的、对干旱的调控机制。
参考文献
Interspecific delimitation and phylogenetic origin of Pugionium (Brassicaceae)
[J].
Compositional and gastrointestinal prokinetic studies of Pugionium (L.)
[J].
Pleistocene climate change and the origin of two desert plant species,Pugionium cornutum and Pugionium dolabratum (Brassicaceae),in northwest China
[J].
De novo assembly and analysis of the Pugionium cornutum (L.) Gaertn.transcriptome and identification of genes involved in the drought response
[J].
斧翅沙芥(Pugionium dolabratum)脂质转运蛋白基因PdLTP3的克隆及其对干旱胁迫的响应
[J].
Chloride is beneficial for growth of the xerophyte Pugionium cornutum by enhancing osmotic adjustment capacity under salt and drought stresses
[J].
Transcriptomic profiling identifies candidate genes involved in the salt tolerance of the xerophyte Pugionium cornutum
[J].
Transcriptome analysis of Lolium temulentum exposed to a combination of drought and heat stress
[J].

 甘公网安备 62010202000688号
甘公网安备 62010202000688号
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}