胡杨( Populus euphratica
李端 , 1 , 司建华 , 2 , 李继彦 1 , 王佩将 1 , 原黎明 1
1.太原师范学院 地理科学学院,山西 晋中 030619
2.中国科学院西北生态环境资源研究院 内陆河流域生态水文重点实验室,甘肃 兰州 730000
Physiological responses and differences of Populus euphratica to salt stress and drought stress
Li Duan , 1 , Si Jianhua , 2 , Li Jiyan 1 , Wang Peijiang 1 , Yuan Liming 1
1.Institute of Geographical Science,Taiyuan Normal University,Jinzhong 030619,Shanxi,China
2.Key Laboratory of Eco-hydrology of Inland River Basin,Northwest Institute of Eco-Environment Resources,Chinese Academy of Sciences,Lanzhou 730000,China
通讯作者: 司建华(E-mail: jianhuas@lzb.ac.cn )
收稿日期: 2022-12-10
修回日期: 2023-01-16
基金资助:
山西省高等学校科技创新项目 . 2022L414 . 2022L411 山西省高等学校哲学社会科学研究项目 . 20210141
Received: 2022-12-10
Revised: 2023-01-16
摘要
采用盆栽两年生幼苗,研究胡杨(Populus euphratica )在盐胁迫(NaCl浓度0、100、200、300、400 mmol·L-1 )和干旱胁迫(干旱0、7、14、21、28 d)条件下的生理响应过程及其响应差异。结果表明:在抗氧化酶系统方面,随着盐胁迫和干旱胁迫程度的增加,保护酶超氧化物歧化酶、过氧化物酶和过氧化氢酶活性均呈现先增加后减小的趋势,不同保护酶的活性应对干旱胁迫和盐胁迫的反应速度和持续时间不同,胡杨通过将多种保护酶进行综合调节以形成整个抗氧化酶系统的防御功能。在渗透调节系统方面,随着盐胁迫和干旱胁迫程度的增加,可溶性糖含量持续增加,其积累对于不同程度的盐胁迫和干旱胁迫的反应速度和持续时间不同,胡杨通过持续性积累有机物质维持渗透平衡以形成渗透调节系统的长效防护功能。在细胞膜系统方面,随着盐胁迫程度的增加,丙二醛含量先减少后缓慢增加,基本维持在较低的水平,而随着干旱胁迫程度的增加,丙二醛含量持续增加,胡杨能够在盐胁迫和轻度干旱胁迫下通过细胞膜系统的适应性调节维持细胞膜结构功能的完整性以实现细胞膜系统的保护作用。在盐胁迫和干旱胁迫时,胡杨通过抗氧化酶系统、渗透调节系统和细胞膜系统进行适应性的调整,从而增强其耐盐性和抗旱性。进行胡杨盐胁迫和干旱胁迫生理响应研究对于黑河下游胡杨幼苗的培育和恢复具有重要意义。
关键词:
胡杨(Populus euphratica ) 盐胁迫 干旱胁迫 生理响应
Abstract
Using two-year-old Populus euphratica seedlings as test materials, the study of P. euphratica under different salt stress (0, 100, 200, 300, 400 mmol·L-1 of NaCl) and different drought stress (0, 7, 14, 21, 28 day) was conducted. The results showed that:In terms of the antioxidant enzyme system, with the increase of salt stress and drought stress, the activities of protective enzymes SOD, POD and CAT increased first and then decreased. The reaction speed and duration of different protective enzymes were different in response to drought stress and salt stress. The protective enzymes of P. euphratica were comprehensively regulated to form the defense function of the whole antioxidant enzyme system. As for the osmotic regulation system, with the increase of salt stress and drought stress, the content of soluble sugar continued to increase, and the response speed and duration of its accumulation to different levels of salt stress and drought stress were different. The osmotic balance was maintained by the continuous accumulation of organic matter to form the long-term protection function of the osmotic regulation system. In the cell membrane system, with the increase of salt stress, MDA content first decreased and then slowly increased, basically maintaining a low level, while with the increase of drought stress, MDA content continued to increase. Under salt stress and mild drought stress, P. euphratica can maintain the integrity of cell membrane structure and function through adaptive regulation to realize the protective effect of the cell membrane system. Under salt stress and drought stress, the antioxidant enzyme system, osmotic regulation system, and cell membrane system were adapted to enhance the salt tolerance and drought resistance of P. euphratica . The study on physiological response of P. euphratica to salt stress and drought stress is of great significance for the cultivation and recovery of the seedlings in the lower reaches of the Heihe River.
Keywords:
Populus euphratica salt stress drought stress physiological responses
本文引用格式
李端, 司建华, 李继彦, 王佩将, 原黎明. 胡杨( Populus euphratica . 中国沙漠 [J], 2023, 43(2): 205-215 doi:10.7522/j.issn.1000-694X.2023.00006
Li Duan, Si Jianhua, Li Jiyan, Wang Peijiang, Yuan Liming. Physiological responses and differences of Populus euphratica to salt stress and drought stress . Journal of Desert Research
0 引言
植物在不同生长环境中面临的逆境因子不同。水分对于任何生物都是必不可少的生态因子,直接参与植物体内各种生理活动和代谢过程,对植物生长变化具有重要影响,水分可用性的降低与植物的抗旱性紧密相关。耐盐性也是植物面对逆境时的重要抗逆特性,代表植物对盐胁迫环境的耐受能力,植物通过自身的生理代谢变化来适应进入细胞的盐分以抵抗危害[1 ] 。植物遇到盐胁迫或者干旱胁迫时,需要对盐胁迫和干旱胁迫做出一系列生理调节反应,减少因盐胁迫和水分亏缺等不利环境对其造成的各种损伤,并且做出最有利于生存过程的选择,从而形成生态适应[2 ] 。干旱和半干旱地区水资源极其匮乏,且土壤盐渍化不断加剧,植物耐盐性和抗旱性研究对于干旱和半干旱地区植物的保护、恢复和生长管理至关重要。在逆境环境中植物体内抗氧化酶系统中的抗氧化物质会被合成和积累以消除逆境等环境的不利影响,并且不会对其本身造成伤害以增加对环境的适应性;同时,植物渗透调节系统中一些渗透调节物质的积累能够维持细胞保水性以及植物吸水能力,在植物抗逆性中发挥重要作用;此外,植物在逆境中体内生成的膜质过氧化产物会与蛋白质和核酸反应,从而破坏膜结构,影响细胞膜系统的结构和功能[3 -8 ] ,进而影响植物生长和生存过程。植物在进行耐盐和抗旱的生理响应过程中往往涉及到植物的抗氧化酶系统、渗透调节系统和细胞膜系统,植物自身通过生理代谢和相应的物质积累以进行这些生理系统的调节,确保维持植物细胞的正常生理功能,来适应甚至抵抗盐分和干旱的危害。因此,对能够影响植物抗氧化酶系统、渗透调节系统和细胞膜系统的抗氧化物质、渗透调节物质、膜质过氧化产物进行分析,可较为全面地认识植物在盐胁迫和干旱胁迫下的生理调节机制,从而能够了解植物适应不同逆境的内在机制及其差异性。
黑河是中国西部最大的内陆河之一,胡杨(Populus euphratica )是中国极端干旱区荒漠河岸林的主要建群种,对于改善黑河下游生态环境、保护生物多样性等方面具有极为重要的作用,是维系黑河下游“绿色走廊”生态系统功能的主体[9 ] 。由于人类活动不断影响着黑河下游水文过程,河流从上游到下游不断变窄直至断流,土壤含盐量不断升高,河岸植物长期遭受干旱胁迫和盐胁迫[10 ] ,绿色走廊逐渐萎缩。胡杨林退化已成为影响区域生态环境的突出问题,盐胁迫和干旱胁迫是影响胡杨生存环境的重要原因,研究胡杨在盐胁迫和干旱胁迫下如何通过生理调节机制适应外部生存环境对于黑河下游胡杨幼苗的培育和恢复具有非常重要的意义。关于胡杨的耐盐机制和抗旱机制已有一些研究[11 -14 ] ,这些研究大部分侧重于生理调节机制的短期研究和非连续研究,对耐盐和耐旱过程中的生理调节机制的长期研究相对较少,也较少对二者生理调节机制的差异进行研究。因此,本试验研究了盆栽条件持续性的盐胁迫和干旱胁迫下胡杨的生理响应过程,旨在阐明胡杨抵御盐胁迫和干旱环境的适应机制及其差异,为黑河下游胡杨幼苗的培育和恢复以及黑河流域退化生态系统的恢复及重建提供科学的理论依据。
1 研究区概况
研究区额济纳绿洲,位于中国西北部黑河流域下游(41°40′—42°40′N,100°15′—101°15′E),东部为巴丹吉林沙漠,西部为马鬃山,南部为鼎新盆地,北部为蒙古国,面积约3 428 km2 。该地区气候极端干旱且少雨,日照充足,昼夜温差大,是典型的大陆性气候。多年平均气温为8.2 ℃,极端最低气温为-37.6 ℃,极端最高气温达43.1 ℃,年降水量为38 mm,年蒸发量大于3 390 mm,研究区所在区域是中国典型的超干旱地区。该地区地下水是植被生长的主要水分来源,地下水主要来源于河流中部和上游的排放水[15 ] 。额济纳旗生态系统可分为河岸林生态系统和荒漠草原生态系统两类,其中河岸林生态系统是绿洲的最重要组成部分,植被类型以胡杨和柽柳灌丛为主。
2 材料与方法
2.1 试验材料及处理
本研究在中国科学院阿拉善荒漠生态水文试验研究站进行,于4月初在直径约33 cm、高度25 cm的盆中种植了100株2年生的胡杨幼树,每盆种植1株保证根系独立,幼苗培育期间正常浇水,根据当地林场的育苗方式每7 d浇水3 L,幼苗在室外环境中正常生长3个月后在生长季进行试验。试验开始时,我们选择了约40 cm高、健康无虫蛀且生长良好的树苗作为试验对象,整个试验在0~28 d进行。盐胁迫试验分为5组,每4株为一组,共20株用于盐胁迫试验。综合之前学者对于胡杨盐胁迫的研究[7 -8 ] ,设置了盐分梯度,分别为对照组0 mmol·L-1 NaCl,其余4组NaCl浓度依次为100、200、300、400 mmol·L-1 。NaCl溶液为3 L,盐胁迫试验处理时一次性浇入各对应胁迫处理组。干旱胁迫试验同样分为5组,每4株为一组,共20株用于干旱胁迫试验。根据幼苗的浇水周期确定以下干旱方案:对照组(0 d干旱)、7 d处理组(干旱期持续7 d)、14 d处理组(干旱期持续14 d)、21 d处理组(干旱期持续21 d)和28 d处理组(干旱期持续28 d)。干旱处理是通过暂停浇水来减少水的供应。同时,在处理过程中,雨天在花盆上放置一个透明的塑料棚,以确保干旱的持续可控性。在盐胁迫期间和非干旱持续期间正常浇水,不同盐胁迫和干旱处理的开始时间分别是试验结束前28、21、14、7、0 d,以确保所有试验组的幼苗在同一天达到所需的盐胁迫和干旱胁迫持续天数,以避免因生长引起的测量差异。
2.2 测定方法
2.2.1 保护酶活性的测定
胁迫处理结束后,立刻采集新鲜叶片样本,并将其放入液氮中,转移到实验室后将液氮中的叶片转至超低温冰箱(-80 ℃ )中进行保存。采用中国苏州科明生物技术有限公司生产的试剂盒对样品中超氧化物歧化酶(SOD)、过氧化物酶(POD)和过氧化氢酶(CAT)活性进行分析测定。将新鲜叶片加液氮研磨后用分析天平精确称取0.1 g样品,然后加入1 mL的磷酸缓冲液(pH=7.8)进行冰浴匀浆。在4 ℃用12 000 r·min-1 离心15 min,提取上清液测定SOD、POD和CAT的活性。SOD活性采用氮蓝四脞比色法测定,POD活性采用愈创木酚染色法测定,CAT活性采用钼酸铵比色法测定[16 ] 。
2.2.2 可溶性糖含量的测定
用分析天平称取0.1~0.2 g样品,将样品置于研钵中加入少量蒸馏水充分研磨,将研磨好的匀浆倒入离心管并用蒸馏水定容至10 mL,离心管盖好后置于水浴锅中30 min,设置温度为95 ℃,冷却后,用离心机离心,设置转速为3 000 r·min-1 ,然后取上清液进行测定,测定采用蒽酮比色法。
2.2.3 丙二醛含量的测定
采用中国苏州科明生物技术有限公司生产的试剂盒对样品中丙二醛(MDA)含量进行分析测定。将新鲜叶片加液氮研磨后用分析天平精确称取0.1 g样品,加入1 mL的磷酸缓冲液进行冰浴匀浆。在4 ℃用12 000 r·min-1 离心15 min,提取上清液测定MDA的含量,MDA是过氧化脂质,采用硫代巴比妥酸色谱法测定。
2.3 数据分析
采用ANOVA分析盐胁迫和水分胁迫对胡杨幼苗的影响。多重比较采用Duncan法,显著性检验水平为P =0.05。方差分析由SPSS 19.0软件完成,作图由Origin 2017软件完成。
3 结果
3.1 保护酶活性的变化
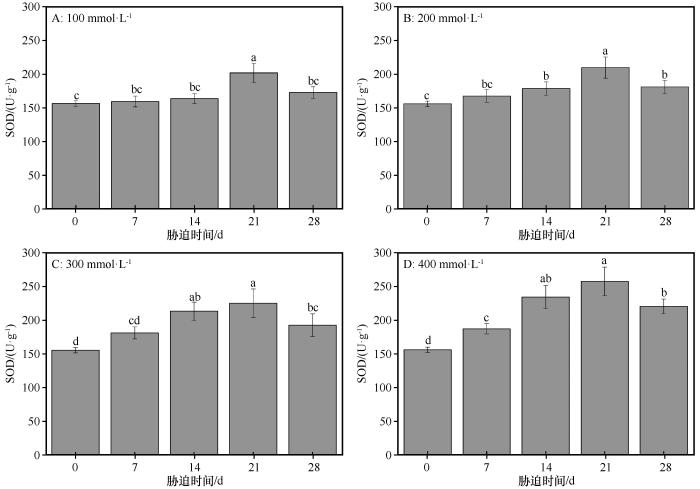
同浓度盐处理下,随着盐胁迫持续时间的延长,SOD的活性均呈现先增加后减少的趋势(图1 )。在对照组,SOD的活性为156.49 U·g-1 ,当NaCl浓度分别为100、200 mmol·L-1 时,在盐胁迫处理后的0~14 d,SOD活性增加相对平缓。当NaCl浓度分别为300、400 mmol·L-1 时,在盐胁迫处理后的0~14 d,SOD活性显著增加(P <0.05)。在NaCl浓度分别为100、200、300、400 mmol·L-1 时,SOD的活性均在盐胁迫处理后21 d时值达到最大,依次为202.09、210.39、226.20、258.32 U·g-1 ,相比对照组分别增加了29.14%、34.44%、44.55%、65.07%,然后从盐胁迫处理后21 d到盐胁迫处理后28 d,SOD的活性显著下降。在NaCl浓度分别为100、200、300、400 mmol·L-1 时,盐胁迫处理后28 d时SOD活性分别是对照组的1.10、1.16、1.24、1.41倍,除NaCl浓度为100 mmol·L-1 外,盐胁迫处理后28 d时SOD活性均显著高于对照组(P <0.05),且盐分浓度越高,差异越大。
图1
图1
盐分胁迫条件下胡杨叶片中超氧化物歧化酶(SOD)活性的变化
不同小写字母表示差异显著,P <0.05
Fig.1
The variation of SOD in the leaves of Populus euphratica under different salinity
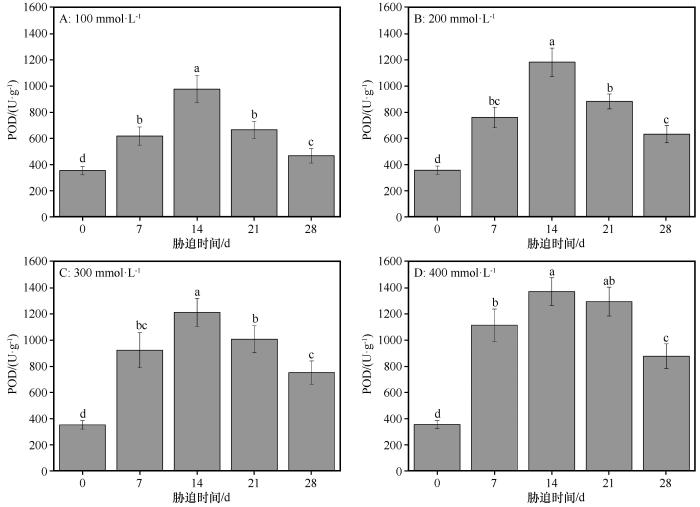
同浓度盐处理下,随着盐胁迫持续时间的延长,POD的活性均呈现先增加后减少的趋势(图2 )。在对照组,POD的活性为354.37 U·g-1 ,在NaCl浓度分别为100、200、300、400 mmol·L-1 时,POD的活性均在盐胁迫处理后14 d时值达到最大,依次为796.78、1 179.58、1 213.01、1 371.23 U·g-1 。从对照组到盐胁迫处理后14 d,相邻胁迫处理组POD活性显著增加(P <0.05),然后从盐胁迫处理后14 d到盐胁迫处理后28 d,POD的活性显著下降(P <0.05)。在NaCl浓度分别为100、200、300、400 mmol·L-1 时,盐胁迫处理后28 d时POD活性分别是对照组的1.32、1.78、2.13、2.48倍,均显著高于对照组(P <0.05)。
图2
图2
盐分胁迫条件下胡杨叶片中过氧化物酶(POD)活性的变化
不同小写字母表示差异显著,P <0.05
Fig.2
The variation of POD in the leaves of Populus euphratica under different salinity
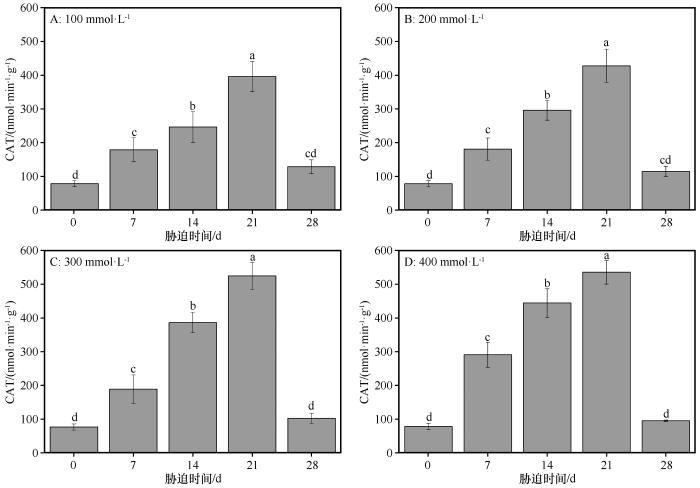
同浓度盐处理下,随着盐胁迫持续时间的延长,CAT的活性均呈现先增加后减少的趋势(图3 )。在对照组,CAT的活性为77.42 nmol·min-1 ·g-1 ,在NaCl浓度分别为100、200、300、400 mmol·L-1 时,CAT的活性均在盐胁迫处理后21 d时值达到最大,依次为396.13、427.07、524.77、535.99 nmol·min-1 ·g-1 。从对照组到盐胁迫处理后14 d,相邻胁迫处理组CAT活性显著增加(P <0.05),然后从盐胁迫处理后21 d到盐胁迫处理后28 d,CAT的活性显著下降(P <0.05)。在NaCl浓度分别为100、200、300、400 mmol·L-1 时,盐胁迫处理后28 d时CAT活性分别是对照组的1.65、1.47、1.33、1.22倍,和对照组并无显著性差异(P >0.05)。
图3
图3
盐分胁迫条件下胡杨叶片中过氧化氢酶(CAT)活性的变化
不同小写字母表示差异显著,P <0.05
Fig.3
The variation of CAT in the leaves of Populus euphratica under different salinity
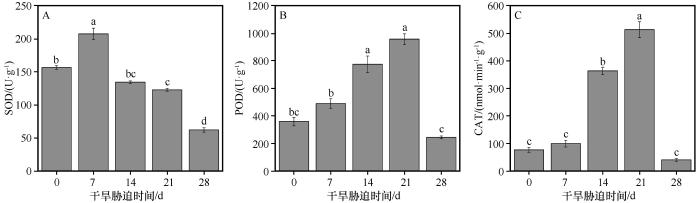
随着干旱持续时间的延长,干旱胁迫程度不断加剧,SOD、POD、CAT活性均呈现先增加后减少的趋势(图4 )。在对照组,SOD的活性为156.49 U·g-1 ,从对照组到干旱7 d,SOD的活性增加到207.48 U·g-1 ,与对照组相比增加了32.58%。从干旱7 d到干旱28 d,SOD的活性显著降低。在干旱持续28 d时,SOD的活性为62.13 U·g-1 ,与对照组相比降低了60.30%。在对照组,POD的活性为358.93 U·g-1 ,从对照组到干旱21 d,POD的活性增加到957.09 U·g-1 ,与对照组相比增加了1.67倍。从干旱21 d到干旱28 d,POD的活性迅速降低到244.49 U·g-1 ,与对照组相比降低了31.89%。在对照组,CAT的活性为76.89 nmol·min-1 ·g-1 ,从对照组到干旱21 d,CAT的活性增加到513.56 nmol·min-1 ·g-1 ,与对照组相比增加了5.68倍。从干旱21 d到干旱28 d,CAT的活性迅速降低到40.34 U·g-1 ,与对照组相比降低了47.54%。
图4
图4
干旱胁迫条件下胡杨叶片中超氧化物歧化酶(SOD),过氧化物酶(POD)和过氧化氢酶(CAT)活性的变化
不同小写字母表示差异显著,P <0.05
Fig.4
The variation of SOD, POD and CAT in the leaves of Populus euphratica under different drought
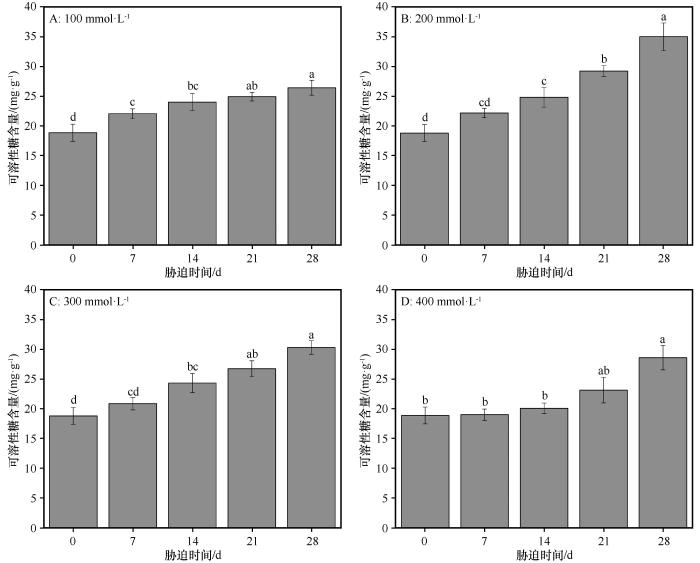
3.2 可溶性糖含量的变化
同浓度盐处理下,随着盐胁迫持续时间的延长,可溶性糖的含量均呈现逐渐增加的趋势(图5 )。在对照组,可溶性糖的含量为18.85 mg·g-1 ,在NaCl浓度分别为100、200、300、400 mmol·L-1 时,可溶性糖的含量均在盐胁迫处理后28 d时值达到最大,依次为26.41、35.01、30.36、28.54 mg·g-1 。在NaCl浓度分别为100、200、300 mmol·L-1 时,从对照组到盐胁迫处理后28 d,可溶性糖的含量显著增加(P <0.05)。在NaCl浓度为400 mmol·L-1 时,从对照组到盐胁迫处理后14 d,可溶性糖的含量基本维持稳定,从盐胁迫处理后14 d开始,一直到盐胁迫处理后28 d,可溶性糖含量开始逐渐增加。
图5
图5
盐分胁迫条件下胡杨叶片中可溶性糖含量的变化
不同小写字母表示差异显著,P <0.05
Fig.5
The variation of proline content in the leaves of Populus euphratica under different salinity
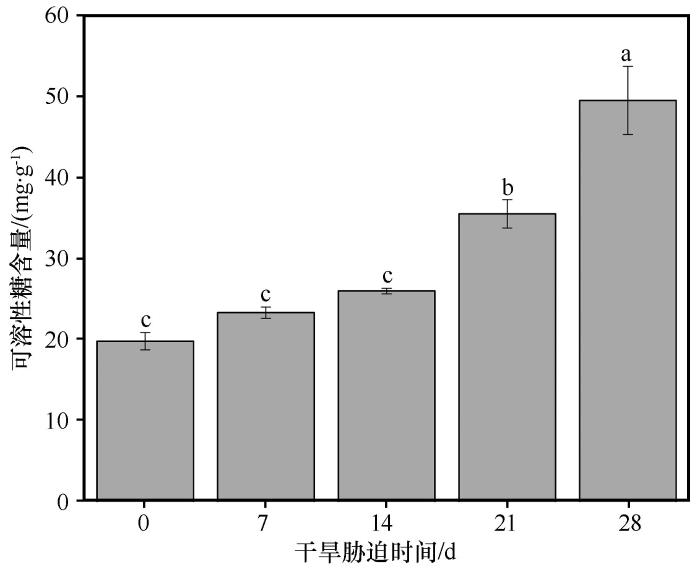
随着干旱胁迫持续时间的增加,叶片可溶性糖含量持续增加(图6 )。在对照组中,叶片可溶性糖含量为19.70 mg·g-1 。从对照组到干旱14 d时,可溶性糖的含量基本维持稳定。从干旱14 d到干旱28 d时,可溶性糖含量显著增加,到干旱28 d时,可溶性糖含量为49.30 mg·g-1 ,相比对照组增加了1.50倍。这一变化趋势与胡杨在重度盐胁迫(即NaCl浓度为400 mmol·L-1 )时可溶性糖含量的变化趋势一致。
图6
图6
干旱胁迫条件下胡杨叶片中可溶性糖含量的变化
不同小写字母表示差异显著,P <0.05
Fig.6
The variation of proline content in the leaves of Populus euphratica under different drought
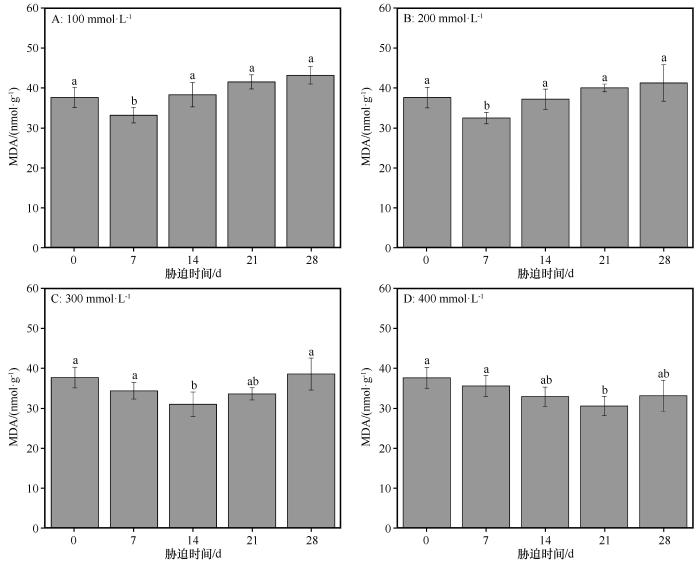
3.3 丙二醛含量的变化
植物在胁迫环境中体内自由基作用于脂质发生过氧化反应,最终氧化产物为MDA,其积累程度可以反映植物具有细胞毒性和受环境胁迫的损伤程度[17 ] 。同浓度盐处理下,随着盐胁迫持续时间的延长,MDA的含量均呈现先减少后增加的趋势(图7 )。在对照组中,叶片MDA含量为37.64 nmol·g-1 。在NaCl浓度分别为100、200 mmol·L-1 时,从对照组到盐胁迫处理后7 d时MDA含量显著降低,从盐胁迫处理后7 d到盐胁迫处理后28 d逐渐增加,相邻处理组MDA的含量无显著性差异。在NaCl浓度分别为100、200 mmol·L-1 时,从对照组到盐胁迫处理后7 d时,MDA含量比对照组分别降低了11.85%、13.71%。在NaCl浓度为300 mmol·L-1 时,从对照组到盐胁迫处理后14 d时,MDA含量比对照组降低了17.69%,从盐胁迫处理后14 d到盐胁迫处理后28 d显著增加(P <0.05)。在NaCl浓度为400 mmol·L-1 时,从对照组到盐胁迫处理后21 d时,MDA含量比对照组降低了18.78%,从盐胁迫处理后21 d到盐胁迫处理后28 d MDA含量增加。在盐胁迫28 d时各处理组MDA的含量和对照组无显著性差异(图7 )。这是由于胡杨受到盐分胁迫时,SOD、POD和CAT等保护酶活性的增加强化了其抗氧化酶系统的防御功能,并通过可溶性糖等有机物质的积累增强了渗透调节系统的功能,二者共同作用一定程度能够减少MDA的产生并促使细胞膜系统维持其生理功能,因此当胡杨受到盐胁迫时前期MDA含量降低。
图7
图7
盐分胁迫条件下胡杨叶片中丙二醛(MDA)含量的变化
不同小写字母表示差异显著,P <0.05
Fig.7
The variation of MDA content in the leaves of Populus euphratica under different salinity
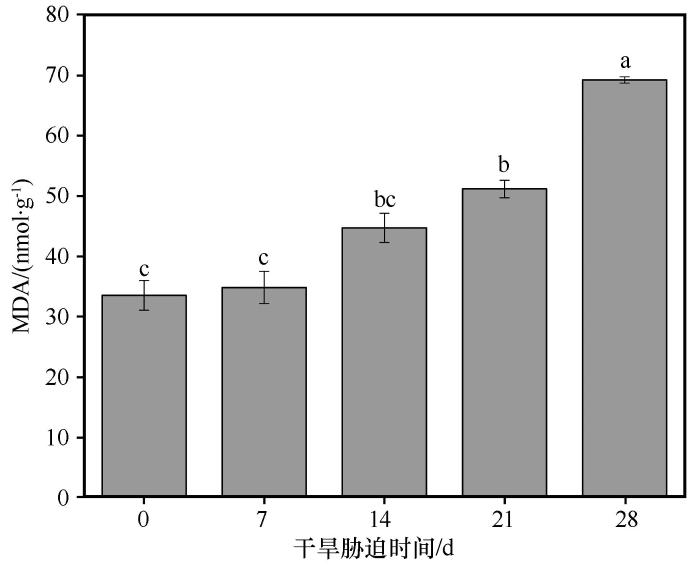
随着干旱胁迫持续时间的增加,胡杨叶片中MDA含量持续增加(图8 )。在对照组中,叶片MDA含量为33.51 nmol·g-1 。从对照组到干旱7 d时,MDA含量基本不变。从干旱7 d到干旱21 d时,MDA含量缓慢增加,MDA含量在干旱21 d值为51.13 nmol·g-1 ,比对照组增加了52.6%。从干旱21 d到干旱28 d时,MDA含量增加了35.3%,显著增加到69.19 nmol·g-1 ,使得在干旱28 d的MDA含量大约是对照组的2倍。可见干旱胁迫对胡杨叶片MDA含量有显著的影响,随着干旱胁迫程度的加剧,MDA含量持续增加,膜脂过氧化作用产生大量有害代谢产物,对细胞膜的损害程度加剧。
图8
图8
干旱胁迫条件下胡杨叶片中丙二醛(MDA)含量的变化
不同小写字母表示差异显著,P <0.05
Fig.8
The variation of MDA content in the leaves of Populus euphratica under different drought
4 讨论
4.1 胡杨抗氧化酶系统的适应性调节
植物在遭受盐胁迫和干旱胁迫时,体内会不断产生活性氧物质并进行积累,从而能导致细胞发生氧化损伤和代谢紊乱。为了抵抗盐胁迫和干旱等逆境带来的伤害,植物需要进行抗氧化酶系统的适应性调节来消除环境不利影响,增加对环境的适应性。抗氧化酶系统中最主要的保护酶是SOD、POD和CAT,它们能够在不利的生存环境中清除植物体内过量的活性氧[18 ] 。SOD能够首先阻止细胞发生氧化损伤,在众多保护酶中占据首要位置,它处于抵御活性氧伤害的“第一道防线”[19 ] 。SOD主要担任超氧自由基(O2- )的清除剂,可将O2- 歧化为H2 O2 与O2 ,H2 O2 则是被CAT和POD进一步分解和消除[20 ] ,保护酶起到清除产生的活性氧、维持植物体内动态平衡的作用,保护细胞免受伤害。以往研究表明当植物受到盐胁迫和干旱胁迫时,其体内SOD、POD和CAT等保护酶的活性会增加,从而能够清除过量的活性氧,控制细胞内脂质氧化过程和抵御活性氧的伤害,维持植物体内动态平衡[8 ,21 ] 。当超过一定的胁迫程度时,保护酶活性会产生下降,此时抗氧化防御功能和保护能力减弱,其活性进一步下降致使该种保护酶在抗氧化防御方面几乎不再起保护作用。
胡杨在应对盐胁迫的过程中,SOD的活性缓慢增强,当盐胁迫增强到一定程度时,SOD的活性逐渐下降最终接近无胁迫环境的水平。胡杨在应对干旱胁迫的过程中,SOD的活性迅速增强,其增强的活性维持时间相对较短,随着干旱胁迫程度的增加,SOD的活性迅速下降最终显著低于无胁迫环境的水平。可见,胡杨面对盐胁迫相比干旱胁迫体内SOD活性增强持续时间长,而胡杨面对干旱胁迫相比盐胁迫体内SOD活性增强更为迅速,但持续时间短,后期其提供抗氧化防御的功能相对减弱,此时其他保护酶POD和CAT承担主要的抗氧化防御功能。胡杨在应对盐胁迫和干旱胁迫的过程中,POD和CAT的活性变化趋势基本相同,前期都是随着胁迫程度的增加活性增强,再随着胁迫的进一步加剧,其活性下降。由研究我们发现,当胡杨受到盐胁迫和干旱胁迫时,胁迫前期其体内SOD、POD和CAT的活性会不断增加,SOD、POD和CAT作为保护酶能够清除过量的活性氧,维持胡杨体内动态平衡,增强胡杨对逆境的抗性[22 ] 。不同之处在于:胡杨在应对盐胁迫程度比较严重时,SOD、POD和CAT的活性接近无胁迫环境的水平。相反,胡杨在应对干旱胁迫程度比较严重时,SOD、POD和CAT的活性显著低于无胁迫环境的水平。可见,胡杨在应对盐胁迫环境下,能够维持较长时间保持SOD、POD和CAT的活性,维持自身的抗氧化防御功能和保护作用。但是,胡杨在应对干旱胁迫的环境下,重度干旱会导致其体内SOD、POD和CAT活性的明显减弱,这些保护酶在抗氧化防御方面功能弱化明显甚至几乎不再起保护作用,致使保护酶发挥其抗氧化防御功能的持续时间缩短,可见保护酶在逆境中并不能对植物细胞进行持续性的保护。综上所述,为了适应盐胁迫和干旱胁迫,胡杨将多种保护酶进行综合调节以形成整个抗氧化酶系统的防御功能,维持体内动态平衡应对不利环境和实现保护作用,但是不同的保护酶应对干旱胁迫和盐胁迫的反应速度和持续时间存在差异,这是胡杨启动抗氧化酶系统应对极端环境并维持生存的适应策略。
4.2 胡杨渗透调节系统的适应性调节
遭受逆境胁迫时,植物能够通过渗透调节系统来维持细胞内的渗透势,缓解胁迫环境对植物造成的影响和伤害。有机溶质是渗透调节系统中的主要物质,参与植物体内渗透调节过程[21 ] 。盐胁迫和干旱胁迫分别会导致土壤渗透压增高和土壤水分亏缺加剧,影响土壤水势和引发植物失水。在逆境胁迫下,植物为保证从土壤中继续吸收水分,必须使自身维持较高的渗透调节能力,降低自身水势,才能形成一个正常的水势梯度实现持续吸水[23 ] ,通过增加有机溶质的含量参与渗透调节,应对外界压力从而保护细胞,使其继续从低水势条件下有效吸水[24 ] 。可溶性糖是细胞质内重要的有机溶质,能够作为渗透保护剂,发挥稳定蛋白质的作用,成为植物遭受胁迫时一种重要的渗透调节物质[25 ] 。可溶性糖的积累能够降低叶片的渗透势,从而有助于使细胞在水分胁迫下保持膨压状态[24 ] 。
胡杨在应对盐胁迫和干旱胁迫的过程中,可溶性糖的含量变化趋势基本相同,都是随着胁迫程度的增加进行持续性积累。胡杨在应对盐胁迫和干旱胁迫的过程中,其体内可溶性糖的含量不断增加,这样体内通过可溶性糖的积累,能够提高细胞渗透压和细胞膨压,从而维持渗透调节平衡,增强胡杨对逆境的抗性。不同之处在于:胡杨在应对盐胁迫的过程中,环境盐分浓度越大,随着胁迫时间的增加,前期胡杨可溶性糖积累越慢。当盐浓度达到一定程度时,随着胁迫时间的增加,前期胡杨可溶性糖含量基本维持稳定。相反,胡杨所处盐环境的浓度越低,其可溶性糖等渗透调节物质能够进行更快的反应调节和更长时间地维持渗透调节。胡杨在应对干旱胁迫的过程中,随着胁迫时间的增加,前期胡杨可溶性糖也基本维持稳定,后期迅速积累。因此,胡杨在盐浓度较高和干旱胁迫的条件下,前期主要依靠保护酶的积累和抗氧化酶系统的抗氧化防御功能发挥保护作用,可溶性糖的含量基本不变,随着抗氧化酶系统保护作用不断减弱,可溶性糖含量持续性增加,增强渗透调节系统的保护作用。在面对长期极端环境时,抗氧化酶系统能够提供有效的前期保护,由于其保护作用缺乏长效性,后期主要依靠增强渗透调节系统的功能实现长期保护效果[26 -27 ] 。综上所述,为了适应盐胁迫和干旱胁迫,胡杨通过持续性积累可溶性糖等有机物质维持渗透平衡以形成渗透调节系统的长效防护功能,维持渗透势应对不利环境和实现保护作用,但是渗透调节物质积累应对不同程度的盐胁迫和干旱胁迫的反应速度和持续时间存在差异,这是胡杨启动渗透调节系统应对极端环境并维持生存的适应策略。
4.3 胡杨细胞膜系统的适应性调节
植物面对长期逆境胁迫时,除了渗透调节系统提供长效防护功能外,长期的保护还得依赖膜结构的功能来实现[26 -27 ] 。细胞只有在膜系统正常运转的情况下才能维持各个系统的平衡和功能的发挥,无论是抗氧化酶系统还是渗透调节系统,其发挥保护作用和渗透调节作用都需要通过细胞膜来进行物质和能量的交换,维持细胞膜系统的稳定性对植物在逆境中生存至关重要[28 ] 。MDA是膜脂过氧化的产物,使细胞膜本身受损,同时MDA自身会与蛋白质和核酸发生反应,从而破坏膜结构,因此MDA能够综合反映细胞膜功能受损情况[17 ] 。在盐胁迫下胞内大量离子、有机物质的外渗,以及有毒盐离子的进入,造成细胞内生理生化过程受到干扰,植物细胞质膜受到损伤,细胞膜系统是植物遭受盐胁迫的主要部位[29 ] 。同样,干旱胁迫会导致细胞原生质脱水,体内形成大量的自由基,膜脂过氧化水平增高和膜脂成分改变,膜系统受到破坏,干旱对细胞的影响表现在使细胞膜受到伤害[30 -31 ] 。
胡杨在应对盐胁迫的过程中,随着胁迫时间的增加,胁迫初期MDA含量缓慢降低,然后MDA含量再缓慢增加最终基本达到无胁迫环境的水平。这是因为当胡杨受到盐分胁迫时,SOD、POD和CAT等保护酶活性的增加强化了其抗氧化酶系统的防御功能,从而可以减少膜中多不饱和脂肪酸过氧化作用的发生,能够抑制MDA的产生;同时可溶性糖等有机物质的积累增强了渗透调节系统的功能,从而可以维持渗透平衡,防止胞内离子的外渗和外界有毒盐离子的进入,减少其细胞质膜受损。可见,强化抗氧化酶系统和渗透调节系统的综合作用一定程度能够减少MDA的产生并促使细胞膜系统维持其生理功能。因此,当胡杨受到盐胁迫时前期抗氧化酶系统和渗透调节系统的增强使MDA含量降低。盐胁迫增加后期MDA含量缓慢增加基本达到无胁迫环境的水平,可见,胡杨面对盐胁迫时其体内细胞膜能够相对维持长时间的稳定性。胡杨在面临盐胁迫时,MDA含量基本维持在较低的水平,说明低MDA水平可能对耐盐性有显著影响,这与之前研究结果一致[32 -35 ] 。胡杨在应对干旱胁迫的过程中,胁迫初期MDA含量基本维持稳定,然后随着胁迫程度的增加,MDA含量逐渐增加。胡杨面对轻度干旱胁迫时体内细胞膜也能够基本维持稳定,而干旱胁迫程度进一步增加时其体内细胞膜受损严重,细胞膜系统无法进行有效调节。干旱胁迫对于胡杨细胞膜系统的损伤远远大于盐胁迫的影响,这很可能是因为MDA含量不断增加会降低细胞膜的收缩性,使导管壁果胶变得松弛和膨胀,阻碍了植物水分传输过程,进一步加剧了干旱缺水环境的不利影响[36 ] 。综上所述,胡杨能够在盐胁迫和轻度干旱胁迫下通过细胞膜系统的适应性调节维持细胞膜结构功能的完整性,实现细胞膜系统的保护作用,这是胡杨启动细胞膜系统应对极端环境并维持生存的适应策略。
5 结论
随着盐胁迫和干旱程度的增加,保护酶SOD、POD和CAT活性均呈现先增加后减小的趋势,不同保护酶的活性应对干旱胁迫和盐胁迫的反应速度和持续时间不同,胡杨通过将多种保护酶进行综合调节以形成整个抗氧化酶系统的防御功能。
随着盐胁迫和干旱程度的增加,可溶性糖含量持续增加,同时可溶性糖的积累对于不同程度的盐胁迫和干旱胁迫的反应速度和持续时间不同,胡杨通过持续性积累有机物质维持渗透平衡以形成渗透调节系统的长效防护功能。
随着盐胁迫程度的增加,MDA含量先减少后缓慢增加,基本维持在较低的水平,而随着干旱胁迫程度的增加,MDA含量持续增加,胡杨能够在盐胁迫和轻度干旱胁迫下通过细胞膜系统的适应性调节维持细胞膜结构功能的完整性以实现细胞膜系统的保护作用。
总之,在盐胁迫和干旱胁迫下,胡杨通过对抗氧化酶系统、渗透调节系统和细胞膜系统进行适应性的调整,从而增强其耐盐性和抗旱性。
参考文献
View Option
[1]
潘瑞炽 ,董愚得 植物生理学 [M].北京 :高等教育出版社 ,2001 .
[本文引用: 1]
[2]
时丽冉 ,刘志华 干旱胁迫对苣荬菜抗氧化酶和渗透调节物质的影响
[J].草地学报 ,2010 ,18 (5 ):673 -677 .
[本文引用: 1]
[3]
Hoekstra F A Golovina E A Buitink J Mechanisms of plant desiccation tolerance
[J].Trends in Plant Science ,2001 ,6 (9 ):431 -438 .
[本文引用: 1]
[4]
刘振林 ,戴思兰 植物甜菜碱醛脱氢酶基因研究进展
[J].西北农林科技大学学报(自然科学版) ,2004 ,32 (3 ):104 -112 .
[5]
裘丽珍 ,黄有军 ,黄坚钦 ,等 不同耐盐性植物在盐胁迫下的生长与生理特性比较研究
[J].浙江大学学报(农业与生命科学版) ,2006 ,32 (4 ):420 -427 .
[6]
李合生 现代植物生理学 [M].北京 :高等教育出版社 ,2006 .
[7]
史军辉 ,王新英 ,刘茂秀 ,等 NaCl胁迫对胡杨幼苗叶主要渗透调节物质的影响
[J].西北林学院学报 ,2014 ,29 (6 ):6 -11 .
[本文引用: 1]
[8]
赵春彦 ,秦洁 ,贺晓慧 ,等 荒漠河岸林胡杨对盐胁迫的适应机制
[J].干旱区资源与环境 ,2022 ,36 (7 ):166 -172 .
[本文引用: 3]
[9]
Si J H Feng Q Cao S K et al Water use sources of desert riparian Populus euphratica forests
[J].Environmental Monitoring & Assessment ,2014 ,186 (9 ):5469 -5477 .
[本文引用: 1]
[10]
刘蔚 ,王涛 ,苏永红 ,等 黑河下游土壤和地下水盐分特征分析
[J].冰川冻土 ,2012 ,27 (6 ):890 -898 .
[本文引用: 1]
[11]
雷善清 ,王文娟 ,王雨辰 ,等 不同土壤水盐条件下多枝柽柳(Tamarix ramosissima )对胡杨(Populus euphratica )幼苗的影响
[J].生态学报 ,2020 ,40 (21 ):7638 -7647 .
[本文引用: 1]
[12]
曾凡江 ,张希明 , Foetzki Andrea 等 新疆策勒绿洲胡杨水分生理特性研究
[J].干旱区研究 ,2002 ,19 (2 ):26 -30 .
[13]
杨永青 ,王文棋 , Ottow Erio A 等 干旱胁迫下胡杨生理适应机制的研究
[J].北京林业大学学报 ,2006 ,28 (2 ):6 -11 .
[14]
陈亚鹏 ,陈亚宁 ,李卫红 ,等 塔里木河下游干旱胁迫下的胡杨生理特点分析
[J].西北植物学报 ,2004 ,24 (10 ):1943 -1948 .
[本文引用: 1]
[15]
Zhou H Y Chen Y N Li W et al Xylem hydraulic conductivity and embolism in riparian plants and their responses to drought stress in desert of Northwest China
[J].Ecohydrology ,2013 ,6 (6 ):984 -993 .
[本文引用: 1]
[16]
朱广廉 植物生理学实验指南 [M].北京 :北京大学出版社 ,1993 .
[本文引用: 1]
[17]
Han X He X Qiu W et al Pathogenesis-related protein PR10 from Salix matsudana Koidz exhibits resistance to salt stress in transgenic Arabidopsis thaliana
[J].Environmental and Experimental Botany ,2017 ,141 :74 -82 .
[本文引用: 2]
[18]
杨升 滨海耐盐树种筛选及评价标准研究
[D].北京 :中国林业科学研究院 ,2010 .
[本文引用: 1]
[19]
任红旭 ,陈雄 ,王亚馥 抗旱性不同的小麦幼苗在水分和盐胁迫下抗氧化酶和多胺的变化
[J].植物生态学报 ,2001 ,25 (6 ):709 -715 .
[本文引用: 1]
[20]
He X L Zhao L L Li Y P Effects of AM fungi on the growth and protective enzymes of cotton under NaCl stress
[J].Acta Ecologica Sinica ,2005 ,25 (1 ):188 -193 .
[本文引用: 1]
[21]
王有年 ,杜方 ,于同泉 ,等 水分胁迫对桃叶片碳水化合物及其相关酶活性的影响
[J].北京农学院学报 ,2001 ,16 (4 ):9 -14 .
[本文引用: 2]
[22]
Fu A Li W Chen Y The threshold of soil moisture and salinity influencing the growth of Populus euphratica and Tamarix ramosissima in the extremely arid region
[J].Environmental Earth Sciences ,2012 ,66 (8 ):2519 -2529 .
[本文引用: 1]
[23]
Zeng F Yan H Arndt S K Leaf and whole tree adaptations to mild salinity in field grown Populus euphratica
[J].Tree Physiology ,2009 ,29 (10 ):1237 -1246 .
[本文引用: 1]
[24]
Aishan T Halik Ü Kurban A et al Eco-morphological response of floodplain forests (Populus euphratica Oliv.) to water diversion in the lower Tarim River,northwest China
[J].Environmental Earth Sciences ,2015 ,73 (2 ):533 -545 .
[本文引用: 2]
[25]
田晓艳 ,刘延吉 ,张蕾 ,等 盐胁迫对景天三七保护酶系统、MDA、Pro及可溶性糖的影响
[J].草原与草坪 ,2009 (6 ):11 -14 .
[本文引用: 1]
[26]
Chris B Montagu M V Dirk I Superoxide dismutase and stress tolerance
[J].Annual Review of Plant Physiology and Plant Molecular Biology ,1992 ,43 (1 ):83 -116 .
[本文引用: 2]
[27]
Harborne J B Introduction to ecological biochemistry
[J].Biochemistry & Molecular Biology Education ,2010 ,6 (1 ):24 .
[本文引用: 2]
[28]
冯燕 ,王彦荣 ,胡小文 水分胁迫对幼苗期霸王叶片生理特性的影响
[J].草业科学 ,2011 ,28 (4 ):577 -581 .
[本文引用: 1]
[29]
赵可夫 植物抗盐生理 [M].北京 :中国科学技术出版社 ,1993 .
[本文引用: 1]
[30]
王文成 ,郭艳超 ,李克晔 ,等 盐胁迫对竹柳种苗形态及生理指标的影响
[J].华北农学报 ,2011 ,26 (1 ):143 -146 .
[本文引用: 1]
[31]
栗燕 ,黎明 ,袁晓晶 ,等 干旱胁迫下菊花叶片的生理响应及抗旱性评价
[J].石河子大学学报(自然科学版) ,2011 ,29 (1 ):30 -34 .
[本文引用: 1]
[32]
Sergio L Paola A D Cantore V et al Effect of salt stress on growth parameters,enzymatic antioxidant system,and lipid peroxidation in wild chicory (Cichorium intybus L.)
[J].Acta Physiologiae Plantarum ,2012 ,34 (6 ):2349 -2358 .
[本文引用: 1]
[33]
Valentina M Micha G Moshe T et al Salinity up-regulates the antioxidative system in root mitochondria and peroxisomes of the wild salt-tolerant tomato species Lycopersicon pennellii
[J].Journal of Experimental Botany ,2004 (399 ):1105 -1113 .
[34]
Ruiz J M Blasco B Rivero R M et al Nicotine-free and salt-tolerant tobacco plants obtained by grafting to salinity-resistant rootstocks of tomato
[J].Munksgaard International Publishers ,2005 ,124 (4 ):465 -475 .
[35]
Liang Y Chen Q Liu Q et al Exogenous silicon (Si) increases antioxidant enzyme activity and reduces lipid peroxidation in roots of salt-stressed barley (Hordeum vulgare L.)
[J].Journal of Plant Physiology ,2003 ,160 (10 ):1157 -1164 .
[本文引用: 1]
[36]
李端 ,司建华 ,张小由 ,等 胡杨(Populus euphratica )对干旱胁迫的生态适应
[J].中国沙漠 ,2020 ,40 (2 ):17 -23 .
[本文引用: 1]
1
2001
... 植物在不同生长环境中面临的逆境因子不同.水分对于任何生物都是必不可少的生态因子,直接参与植物体内各种生理活动和代谢过程,对植物生长变化具有重要影响,水分可用性的降低与植物的抗旱性紧密相关.耐盐性也是植物面对逆境时的重要抗逆特性,代表植物对盐胁迫环境的耐受能力,植物通过自身的生理代谢变化来适应进入细胞的盐分以抵抗危害[1 ] .植物遇到盐胁迫或者干旱胁迫时,需要对盐胁迫和干旱胁迫做出一系列生理调节反应,减少因盐胁迫和水分亏缺等不利环境对其造成的各种损伤,并且做出最有利于生存过程的选择,从而形成生态适应[2 ] .干旱和半干旱地区水资源极其匮乏,且土壤盐渍化不断加剧,植物耐盐性和抗旱性研究对于干旱和半干旱地区植物的保护、恢复和生长管理至关重要.在逆境环境中植物体内抗氧化酶系统中的抗氧化物质会被合成和积累以消除逆境等环境的不利影响,并且不会对其本身造成伤害以增加对环境的适应性;同时,植物渗透调节系统中一些渗透调节物质的积累能够维持细胞保水性以及植物吸水能力,在植物抗逆性中发挥重要作用;此外,植物在逆境中体内生成的膜质过氧化产物会与蛋白质和核酸反应,从而破坏膜结构,影响细胞膜系统的结构和功能[3 -8 ] ,进而影响植物生长和生存过程.植物在进行耐盐和抗旱的生理响应过程中往往涉及到植物的抗氧化酶系统、渗透调节系统和细胞膜系统,植物自身通过生理代谢和相应的物质积累以进行这些生理系统的调节,确保维持植物细胞的正常生理功能,来适应甚至抵抗盐分和干旱的危害.因此,对能够影响植物抗氧化酶系统、渗透调节系统和细胞膜系统的抗氧化物质、渗透调节物质、膜质过氧化产物进行分析,可较为全面地认识植物在盐胁迫和干旱胁迫下的生理调节机制,从而能够了解植物适应不同逆境的内在机制及其差异性. ...
干旱胁迫对苣荬菜抗氧化酶和渗透调节物质的影响
1
2010
... 植物在不同生长环境中面临的逆境因子不同.水分对于任何生物都是必不可少的生态因子,直接参与植物体内各种生理活动和代谢过程,对植物生长变化具有重要影响,水分可用性的降低与植物的抗旱性紧密相关.耐盐性也是植物面对逆境时的重要抗逆特性,代表植物对盐胁迫环境的耐受能力,植物通过自身的生理代谢变化来适应进入细胞的盐分以抵抗危害[1 ] .植物遇到盐胁迫或者干旱胁迫时,需要对盐胁迫和干旱胁迫做出一系列生理调节反应,减少因盐胁迫和水分亏缺等不利环境对其造成的各种损伤,并且做出最有利于生存过程的选择,从而形成生态适应[2 ] .干旱和半干旱地区水资源极其匮乏,且土壤盐渍化不断加剧,植物耐盐性和抗旱性研究对于干旱和半干旱地区植物的保护、恢复和生长管理至关重要.在逆境环境中植物体内抗氧化酶系统中的抗氧化物质会被合成和积累以消除逆境等环境的不利影响,并且不会对其本身造成伤害以增加对环境的适应性;同时,植物渗透调节系统中一些渗透调节物质的积累能够维持细胞保水性以及植物吸水能力,在植物抗逆性中发挥重要作用;此外,植物在逆境中体内生成的膜质过氧化产物会与蛋白质和核酸反应,从而破坏膜结构,影响细胞膜系统的结构和功能[3 -8 ] ,进而影响植物生长和生存过程.植物在进行耐盐和抗旱的生理响应过程中往往涉及到植物的抗氧化酶系统、渗透调节系统和细胞膜系统,植物自身通过生理代谢和相应的物质积累以进行这些生理系统的调节,确保维持植物细胞的正常生理功能,来适应甚至抵抗盐分和干旱的危害.因此,对能够影响植物抗氧化酶系统、渗透调节系统和细胞膜系统的抗氧化物质、渗透调节物质、膜质过氧化产物进行分析,可较为全面地认识植物在盐胁迫和干旱胁迫下的生理调节机制,从而能够了解植物适应不同逆境的内在机制及其差异性. ...
Mechanisms of plant desiccation tolerance
1
2001
... 植物在不同生长环境中面临的逆境因子不同.水分对于任何生物都是必不可少的生态因子,直接参与植物体内各种生理活动和代谢过程,对植物生长变化具有重要影响,水分可用性的降低与植物的抗旱性紧密相关.耐盐性也是植物面对逆境时的重要抗逆特性,代表植物对盐胁迫环境的耐受能力,植物通过自身的生理代谢变化来适应进入细胞的盐分以抵抗危害[1 ] .植物遇到盐胁迫或者干旱胁迫时,需要对盐胁迫和干旱胁迫做出一系列生理调节反应,减少因盐胁迫和水分亏缺等不利环境对其造成的各种损伤,并且做出最有利于生存过程的选择,从而形成生态适应[2 ] .干旱和半干旱地区水资源极其匮乏,且土壤盐渍化不断加剧,植物耐盐性和抗旱性研究对于干旱和半干旱地区植物的保护、恢复和生长管理至关重要.在逆境环境中植物体内抗氧化酶系统中的抗氧化物质会被合成和积累以消除逆境等环境的不利影响,并且不会对其本身造成伤害以增加对环境的适应性;同时,植物渗透调节系统中一些渗透调节物质的积累能够维持细胞保水性以及植物吸水能力,在植物抗逆性中发挥重要作用;此外,植物在逆境中体内生成的膜质过氧化产物会与蛋白质和核酸反应,从而破坏膜结构,影响细胞膜系统的结构和功能[3 -8 ] ,进而影响植物生长和生存过程.植物在进行耐盐和抗旱的生理响应过程中往往涉及到植物的抗氧化酶系统、渗透调节系统和细胞膜系统,植物自身通过生理代谢和相应的物质积累以进行这些生理系统的调节,确保维持植物细胞的正常生理功能,来适应甚至抵抗盐分和干旱的危害.因此,对能够影响植物抗氧化酶系统、渗透调节系统和细胞膜系统的抗氧化物质、渗透调节物质、膜质过氧化产物进行分析,可较为全面地认识植物在盐胁迫和干旱胁迫下的生理调节机制,从而能够了解植物适应不同逆境的内在机制及其差异性. ...
不同耐盐性植物在盐胁迫下的生长与生理特性比较研究
0
2006
NaCl胁迫对胡杨幼苗叶主要渗透调节物质的影响
1
2014
... 本研究在中国科学院阿拉善荒漠生态水文试验研究站进行,于4月初在直径约33 cm、高度25 cm的盆中种植了100株2年生的胡杨幼树,每盆种植1株保证根系独立,幼苗培育期间正常浇水,根据当地林场的育苗方式每7 d浇水3 L,幼苗在室外环境中正常生长3个月后在生长季进行试验.试验开始时,我们选择了约40 cm高、健康无虫蛀且生长良好的树苗作为试验对象,整个试验在0~28 d进行.盐胁迫试验分为5组,每4株为一组,共20株用于盐胁迫试验.综合之前学者对于胡杨盐胁迫的研究[7 -8 ] ,设置了盐分梯度,分别为对照组0 mmol·L-1 NaCl,其余4组NaCl浓度依次为100、200、300、400 mmol·L-1 .NaCl溶液为3 L,盐胁迫试验处理时一次性浇入各对应胁迫处理组.干旱胁迫试验同样分为5组,每4株为一组,共20株用于干旱胁迫试验.根据幼苗的浇水周期确定以下干旱方案:对照组(0 d干旱)、7 d处理组(干旱期持续7 d)、14 d处理组(干旱期持续14 d)、21 d处理组(干旱期持续21 d)和28 d处理组(干旱期持续28 d).干旱处理是通过暂停浇水来减少水的供应.同时,在处理过程中,雨天在花盆上放置一个透明的塑料棚,以确保干旱的持续可控性.在盐胁迫期间和非干旱持续期间正常浇水,不同盐胁迫和干旱处理的开始时间分别是试验结束前28、21、14、7、0 d,以确保所有试验组的幼苗在同一天达到所需的盐胁迫和干旱胁迫持续天数,以避免因生长引起的测量差异. ...
荒漠河岸林胡杨对盐胁迫的适应机制
3
2022
... 植物在不同生长环境中面临的逆境因子不同.水分对于任何生物都是必不可少的生态因子,直接参与植物体内各种生理活动和代谢过程,对植物生长变化具有重要影响,水分可用性的降低与植物的抗旱性紧密相关.耐盐性也是植物面对逆境时的重要抗逆特性,代表植物对盐胁迫环境的耐受能力,植物通过自身的生理代谢变化来适应进入细胞的盐分以抵抗危害[1 ] .植物遇到盐胁迫或者干旱胁迫时,需要对盐胁迫和干旱胁迫做出一系列生理调节反应,减少因盐胁迫和水分亏缺等不利环境对其造成的各种损伤,并且做出最有利于生存过程的选择,从而形成生态适应[2 ] .干旱和半干旱地区水资源极其匮乏,且土壤盐渍化不断加剧,植物耐盐性和抗旱性研究对于干旱和半干旱地区植物的保护、恢复和生长管理至关重要.在逆境环境中植物体内抗氧化酶系统中的抗氧化物质会被合成和积累以消除逆境等环境的不利影响,并且不会对其本身造成伤害以增加对环境的适应性;同时,植物渗透调节系统中一些渗透调节物质的积累能够维持细胞保水性以及植物吸水能力,在植物抗逆性中发挥重要作用;此外,植物在逆境中体内生成的膜质过氧化产物会与蛋白质和核酸反应,从而破坏膜结构,影响细胞膜系统的结构和功能[3 -8 ] ,进而影响植物生长和生存过程.植物在进行耐盐和抗旱的生理响应过程中往往涉及到植物的抗氧化酶系统、渗透调节系统和细胞膜系统,植物自身通过生理代谢和相应的物质积累以进行这些生理系统的调节,确保维持植物细胞的正常生理功能,来适应甚至抵抗盐分和干旱的危害.因此,对能够影响植物抗氧化酶系统、渗透调节系统和细胞膜系统的抗氧化物质、渗透调节物质、膜质过氧化产物进行分析,可较为全面地认识植物在盐胁迫和干旱胁迫下的生理调节机制,从而能够了解植物适应不同逆境的内在机制及其差异性. ...
... 本研究在中国科学院阿拉善荒漠生态水文试验研究站进行,于4月初在直径约33 cm、高度25 cm的盆中种植了100株2年生的胡杨幼树,每盆种植1株保证根系独立,幼苗培育期间正常浇水,根据当地林场的育苗方式每7 d浇水3 L,幼苗在室外环境中正常生长3个月后在生长季进行试验.试验开始时,我们选择了约40 cm高、健康无虫蛀且生长良好的树苗作为试验对象,整个试验在0~28 d进行.盐胁迫试验分为5组,每4株为一组,共20株用于盐胁迫试验.综合之前学者对于胡杨盐胁迫的研究[7 -8 ] ,设置了盐分梯度,分别为对照组0 mmol·L-1 NaCl,其余4组NaCl浓度依次为100、200、300、400 mmol·L-1 .NaCl溶液为3 L,盐胁迫试验处理时一次性浇入各对应胁迫处理组.干旱胁迫试验同样分为5组,每4株为一组,共20株用于干旱胁迫试验.根据幼苗的浇水周期确定以下干旱方案:对照组(0 d干旱)、7 d处理组(干旱期持续7 d)、14 d处理组(干旱期持续14 d)、21 d处理组(干旱期持续21 d)和28 d处理组(干旱期持续28 d).干旱处理是通过暂停浇水来减少水的供应.同时,在处理过程中,雨天在花盆上放置一个透明的塑料棚,以确保干旱的持续可控性.在盐胁迫期间和非干旱持续期间正常浇水,不同盐胁迫和干旱处理的开始时间分别是试验结束前28、21、14、7、0 d,以确保所有试验组的幼苗在同一天达到所需的盐胁迫和干旱胁迫持续天数,以避免因生长引起的测量差异. ...
... 植物在遭受盐胁迫和干旱胁迫时,体内会不断产生活性氧物质并进行积累,从而能导致细胞发生氧化损伤和代谢紊乱.为了抵抗盐胁迫和干旱等逆境带来的伤害,植物需要进行抗氧化酶系统的适应性调节来消除环境不利影响,增加对环境的适应性.抗氧化酶系统中最主要的保护酶是SOD、POD和CAT,它们能够在不利的生存环境中清除植物体内过量的活性氧[18 ] .SOD能够首先阻止细胞发生氧化损伤,在众多保护酶中占据首要位置,它处于抵御活性氧伤害的“第一道防线”[19 ] .SOD主要担任超氧自由基(O2- )的清除剂,可将O2- 歧化为H2 O2 与O2 ,H2 O2 则是被CAT和POD进一步分解和消除[20 ] ,保护酶起到清除产生的活性氧、维持植物体内动态平衡的作用,保护细胞免受伤害.以往研究表明当植物受到盐胁迫和干旱胁迫时,其体内SOD、POD和CAT等保护酶的活性会增加,从而能够清除过量的活性氧,控制细胞内脂质氧化过程和抵御活性氧的伤害,维持植物体内动态平衡[8 ,21 ] .当超过一定的胁迫程度时,保护酶活性会产生下降,此时抗氧化防御功能和保护能力减弱,其活性进一步下降致使该种保护酶在抗氧化防御方面几乎不再起保护作用. ...
Water use sources of desert riparian Populus euphratica forests
1
2014
... 黑河是中国西部最大的内陆河之一,胡杨(Populus euphratica )是中国极端干旱区荒漠河岸林的主要建群种,对于改善黑河下游生态环境、保护生物多样性等方面具有极为重要的作用,是维系黑河下游“绿色走廊”生态系统功能的主体[9 ] .由于人类活动不断影响着黑河下游水文过程,河流从上游到下游不断变窄直至断流,土壤含盐量不断升高,河岸植物长期遭受干旱胁迫和盐胁迫[10 ] ,绿色走廊逐渐萎缩.胡杨林退化已成为影响区域生态环境的突出问题,盐胁迫和干旱胁迫是影响胡杨生存环境的重要原因,研究胡杨在盐胁迫和干旱胁迫下如何通过生理调节机制适应外部生存环境对于黑河下游胡杨幼苗的培育和恢复具有非常重要的意义.关于胡杨的耐盐机制和抗旱机制已有一些研究[11 -14 ] ,这些研究大部分侧重于生理调节机制的短期研究和非连续研究,对耐盐和耐旱过程中的生理调节机制的长期研究相对较少,也较少对二者生理调节机制的差异进行研究.因此,本试验研究了盆栽条件持续性的盐胁迫和干旱胁迫下胡杨的生理响应过程,旨在阐明胡杨抵御盐胁迫和干旱环境的适应机制及其差异,为黑河下游胡杨幼苗的培育和恢复以及黑河流域退化生态系统的恢复及重建提供科学的理论依据. ...
黑河下游土壤和地下水盐分特征分析
1
2012
... 黑河是中国西部最大的内陆河之一,胡杨(Populus euphratica )是中国极端干旱区荒漠河岸林的主要建群种,对于改善黑河下游生态环境、保护生物多样性等方面具有极为重要的作用,是维系黑河下游“绿色走廊”生态系统功能的主体[9 ] .由于人类活动不断影响着黑河下游水文过程,河流从上游到下游不断变窄直至断流,土壤含盐量不断升高,河岸植物长期遭受干旱胁迫和盐胁迫[10 ] ,绿色走廊逐渐萎缩.胡杨林退化已成为影响区域生态环境的突出问题,盐胁迫和干旱胁迫是影响胡杨生存环境的重要原因,研究胡杨在盐胁迫和干旱胁迫下如何通过生理调节机制适应外部生存环境对于黑河下游胡杨幼苗的培育和恢复具有非常重要的意义.关于胡杨的耐盐机制和抗旱机制已有一些研究[11 -14 ] ,这些研究大部分侧重于生理调节机制的短期研究和非连续研究,对耐盐和耐旱过程中的生理调节机制的长期研究相对较少,也较少对二者生理调节机制的差异进行研究.因此,本试验研究了盆栽条件持续性的盐胁迫和干旱胁迫下胡杨的生理响应过程,旨在阐明胡杨抵御盐胁迫和干旱环境的适应机制及其差异,为黑河下游胡杨幼苗的培育和恢复以及黑河流域退化生态系统的恢复及重建提供科学的理论依据. ...
不同土壤水盐条件下多枝柽柳(Tamarix ramosissima )对胡杨(Populus euphratica )幼苗的影响
1
2020
... 黑河是中国西部最大的内陆河之一,胡杨(Populus euphratica )是中国极端干旱区荒漠河岸林的主要建群种,对于改善黑河下游生态环境、保护生物多样性等方面具有极为重要的作用,是维系黑河下游“绿色走廊”生态系统功能的主体[9 ] .由于人类活动不断影响着黑河下游水文过程,河流从上游到下游不断变窄直至断流,土壤含盐量不断升高,河岸植物长期遭受干旱胁迫和盐胁迫[10 ] ,绿色走廊逐渐萎缩.胡杨林退化已成为影响区域生态环境的突出问题,盐胁迫和干旱胁迫是影响胡杨生存环境的重要原因,研究胡杨在盐胁迫和干旱胁迫下如何通过生理调节机制适应外部生存环境对于黑河下游胡杨幼苗的培育和恢复具有非常重要的意义.关于胡杨的耐盐机制和抗旱机制已有一些研究[11 -14 ] ,这些研究大部分侧重于生理调节机制的短期研究和非连续研究,对耐盐和耐旱过程中的生理调节机制的长期研究相对较少,也较少对二者生理调节机制的差异进行研究.因此,本试验研究了盆栽条件持续性的盐胁迫和干旱胁迫下胡杨的生理响应过程,旨在阐明胡杨抵御盐胁迫和干旱环境的适应机制及其差异,为黑河下游胡杨幼苗的培育和恢复以及黑河流域退化生态系统的恢复及重建提供科学的理论依据. ...
塔里木河下游干旱胁迫下的胡杨生理特点分析
1
2004
... 黑河是中国西部最大的内陆河之一,胡杨(Populus euphratica )是中国极端干旱区荒漠河岸林的主要建群种,对于改善黑河下游生态环境、保护生物多样性等方面具有极为重要的作用,是维系黑河下游“绿色走廊”生态系统功能的主体[9 ] .由于人类活动不断影响着黑河下游水文过程,河流从上游到下游不断变窄直至断流,土壤含盐量不断升高,河岸植物长期遭受干旱胁迫和盐胁迫[10 ] ,绿色走廊逐渐萎缩.胡杨林退化已成为影响区域生态环境的突出问题,盐胁迫和干旱胁迫是影响胡杨生存环境的重要原因,研究胡杨在盐胁迫和干旱胁迫下如何通过生理调节机制适应外部生存环境对于黑河下游胡杨幼苗的培育和恢复具有非常重要的意义.关于胡杨的耐盐机制和抗旱机制已有一些研究[11 -14 ] ,这些研究大部分侧重于生理调节机制的短期研究和非连续研究,对耐盐和耐旱过程中的生理调节机制的长期研究相对较少,也较少对二者生理调节机制的差异进行研究.因此,本试验研究了盆栽条件持续性的盐胁迫和干旱胁迫下胡杨的生理响应过程,旨在阐明胡杨抵御盐胁迫和干旱环境的适应机制及其差异,为黑河下游胡杨幼苗的培育和恢复以及黑河流域退化生态系统的恢复及重建提供科学的理论依据. ...
Xylem hydraulic conductivity and embolism in riparian plants and their responses to drought stress in desert of Northwest China
1
2013
... 研究区额济纳绿洲,位于中国西北部黑河流域下游(41°40′—42°40′N,100°15′—101°15′E),东部为巴丹吉林沙漠,西部为马鬃山,南部为鼎新盆地,北部为蒙古国,面积约3 428 km2 .该地区气候极端干旱且少雨,日照充足,昼夜温差大,是典型的大陆性气候.多年平均气温为8.2 ℃,极端最低气温为-37.6 ℃,极端最高气温达43.1 ℃,年降水量为38 mm,年蒸发量大于3 390 mm,研究区所在区域是中国典型的超干旱地区.该地区地下水是植被生长的主要水分来源,地下水主要来源于河流中部和上游的排放水[15 ] .额济纳旗生态系统可分为河岸林生态系统和荒漠草原生态系统两类,其中河岸林生态系统是绿洲的最重要组成部分,植被类型以胡杨和柽柳灌丛为主. ...
1
1993
... 胁迫处理结束后,立刻采集新鲜叶片样本,并将其放入液氮中,转移到实验室后将液氮中的叶片转至超低温冰箱(-80 ℃ )中进行保存.采用中国苏州科明生物技术有限公司生产的试剂盒对样品中超氧化物歧化酶(SOD)、过氧化物酶(POD)和过氧化氢酶(CAT)活性进行分析测定.将新鲜叶片加液氮研磨后用分析天平精确称取0.1 g样品,然后加入1 mL的磷酸缓冲液(pH=7.8)进行冰浴匀浆.在4 ℃用12 000 r·min-1 离心15 min,提取上清液测定SOD、POD和CAT的活性.SOD活性采用氮蓝四脞比色法测定,POD活性采用愈创木酚染色法测定,CAT活性采用钼酸铵比色法测定[16 ] . ...
Pathogenesis-related protein PR10 from Salix matsudana Koidz exhibits resistance to salt stress in transgenic Arabidopsis thaliana
2
2017
... 植物在胁迫环境中体内自由基作用于脂质发生过氧化反应,最终氧化产物为MDA,其积累程度可以反映植物具有细胞毒性和受环境胁迫的损伤程度[17 ] .同浓度盐处理下,随着盐胁迫持续时间的延长,MDA的含量均呈现先减少后增加的趋势(图7 ).在对照组中,叶片MDA含量为37.64 nmol·g-1 .在NaCl浓度分别为100、200 mmol·L-1 时,从对照组到盐胁迫处理后7 d时MDA含量显著降低,从盐胁迫处理后7 d到盐胁迫处理后28 d逐渐增加,相邻处理组MDA的含量无显著性差异.在NaCl浓度分别为100、200 mmol·L-1 时,从对照组到盐胁迫处理后7 d时,MDA含量比对照组分别降低了11.85%、13.71%.在NaCl浓度为300 mmol·L-1 时,从对照组到盐胁迫处理后14 d时,MDA含量比对照组降低了17.69%,从盐胁迫处理后14 d到盐胁迫处理后28 d显著增加(P <0.05).在NaCl浓度为400 mmol·L-1 时,从对照组到盐胁迫处理后21 d时,MDA含量比对照组降低了18.78%,从盐胁迫处理后21 d到盐胁迫处理后28 d MDA含量增加.在盐胁迫28 d时各处理组MDA的含量和对照组无显著性差异(图7 ).这是由于胡杨受到盐分胁迫时,SOD、POD和CAT等保护酶活性的增加强化了其抗氧化酶系统的防御功能,并通过可溶性糖等有机物质的积累增强了渗透调节系统的功能,二者共同作用一定程度能够减少MDA的产生并促使细胞膜系统维持其生理功能,因此当胡杨受到盐胁迫时前期MDA含量降低. ...
... 植物面对长期逆境胁迫时,除了渗透调节系统提供长效防护功能外,长期的保护还得依赖膜结构的功能来实现[26 -27 ] .细胞只有在膜系统正常运转的情况下才能维持各个系统的平衡和功能的发挥,无论是抗氧化酶系统还是渗透调节系统,其发挥保护作用和渗透调节作用都需要通过细胞膜来进行物质和能量的交换,维持细胞膜系统的稳定性对植物在逆境中生存至关重要[28 ] .MDA是膜脂过氧化的产物,使细胞膜本身受损,同时MDA自身会与蛋白质和核酸发生反应,从而破坏膜结构,因此MDA能够综合反映细胞膜功能受损情况[17 ] .在盐胁迫下胞内大量离子、有机物质的外渗,以及有毒盐离子的进入,造成细胞内生理生化过程受到干扰,植物细胞质膜受到损伤,细胞膜系统是植物遭受盐胁迫的主要部位[29 ] .同样,干旱胁迫会导致细胞原生质脱水,体内形成大量的自由基,膜脂过氧化水平增高和膜脂成分改变,膜系统受到破坏,干旱对细胞的影响表现在使细胞膜受到伤害[30 -31 ] . ...
滨海耐盐树种筛选及评价标准研究
1
2010
... 植物在遭受盐胁迫和干旱胁迫时,体内会不断产生活性氧物质并进行积累,从而能导致细胞发生氧化损伤和代谢紊乱.为了抵抗盐胁迫和干旱等逆境带来的伤害,植物需要进行抗氧化酶系统的适应性调节来消除环境不利影响,增加对环境的适应性.抗氧化酶系统中最主要的保护酶是SOD、POD和CAT,它们能够在不利的生存环境中清除植物体内过量的活性氧[18 ] .SOD能够首先阻止细胞发生氧化损伤,在众多保护酶中占据首要位置,它处于抵御活性氧伤害的“第一道防线”[19 ] .SOD主要担任超氧自由基(O2- )的清除剂,可将O2- 歧化为H2 O2 与O2 ,H2 O2 则是被CAT和POD进一步分解和消除[20 ] ,保护酶起到清除产生的活性氧、维持植物体内动态平衡的作用,保护细胞免受伤害.以往研究表明当植物受到盐胁迫和干旱胁迫时,其体内SOD、POD和CAT等保护酶的活性会增加,从而能够清除过量的活性氧,控制细胞内脂质氧化过程和抵御活性氧的伤害,维持植物体内动态平衡[8 ,21 ] .当超过一定的胁迫程度时,保护酶活性会产生下降,此时抗氧化防御功能和保护能力减弱,其活性进一步下降致使该种保护酶在抗氧化防御方面几乎不再起保护作用. ...
抗旱性不同的小麦幼苗在水分和盐胁迫下抗氧化酶和多胺的变化
1
2001
... 植物在遭受盐胁迫和干旱胁迫时,体内会不断产生活性氧物质并进行积累,从而能导致细胞发生氧化损伤和代谢紊乱.为了抵抗盐胁迫和干旱等逆境带来的伤害,植物需要进行抗氧化酶系统的适应性调节来消除环境不利影响,增加对环境的适应性.抗氧化酶系统中最主要的保护酶是SOD、POD和CAT,它们能够在不利的生存环境中清除植物体内过量的活性氧[18 ] .SOD能够首先阻止细胞发生氧化损伤,在众多保护酶中占据首要位置,它处于抵御活性氧伤害的“第一道防线”[19 ] .SOD主要担任超氧自由基(O2- )的清除剂,可将O2- 歧化为H2 O2 与O2 ,H2 O2 则是被CAT和POD进一步分解和消除[20 ] ,保护酶起到清除产生的活性氧、维持植物体内动态平衡的作用,保护细胞免受伤害.以往研究表明当植物受到盐胁迫和干旱胁迫时,其体内SOD、POD和CAT等保护酶的活性会增加,从而能够清除过量的活性氧,控制细胞内脂质氧化过程和抵御活性氧的伤害,维持植物体内动态平衡[8 ,21 ] .当超过一定的胁迫程度时,保护酶活性会产生下降,此时抗氧化防御功能和保护能力减弱,其活性进一步下降致使该种保护酶在抗氧化防御方面几乎不再起保护作用. ...
Effects of AM fungi on the growth and protective enzymes of cotton under NaCl stress
1
2005
... 植物在遭受盐胁迫和干旱胁迫时,体内会不断产生活性氧物质并进行积累,从而能导致细胞发生氧化损伤和代谢紊乱.为了抵抗盐胁迫和干旱等逆境带来的伤害,植物需要进行抗氧化酶系统的适应性调节来消除环境不利影响,增加对环境的适应性.抗氧化酶系统中最主要的保护酶是SOD、POD和CAT,它们能够在不利的生存环境中清除植物体内过量的活性氧[18 ] .SOD能够首先阻止细胞发生氧化损伤,在众多保护酶中占据首要位置,它处于抵御活性氧伤害的“第一道防线”[19 ] .SOD主要担任超氧自由基(O2- )的清除剂,可将O2- 歧化为H2 O2 与O2 ,H2 O2 则是被CAT和POD进一步分解和消除[20 ] ,保护酶起到清除产生的活性氧、维持植物体内动态平衡的作用,保护细胞免受伤害.以往研究表明当植物受到盐胁迫和干旱胁迫时,其体内SOD、POD和CAT等保护酶的活性会增加,从而能够清除过量的活性氧,控制细胞内脂质氧化过程和抵御活性氧的伤害,维持植物体内动态平衡[8 ,21 ] .当超过一定的胁迫程度时,保护酶活性会产生下降,此时抗氧化防御功能和保护能力减弱,其活性进一步下降致使该种保护酶在抗氧化防御方面几乎不再起保护作用. ...
水分胁迫对桃叶片碳水化合物及其相关酶活性的影响
2
2001
... 植物在遭受盐胁迫和干旱胁迫时,体内会不断产生活性氧物质并进行积累,从而能导致细胞发生氧化损伤和代谢紊乱.为了抵抗盐胁迫和干旱等逆境带来的伤害,植物需要进行抗氧化酶系统的适应性调节来消除环境不利影响,增加对环境的适应性.抗氧化酶系统中最主要的保护酶是SOD、POD和CAT,它们能够在不利的生存环境中清除植物体内过量的活性氧[18 ] .SOD能够首先阻止细胞发生氧化损伤,在众多保护酶中占据首要位置,它处于抵御活性氧伤害的“第一道防线”[19 ] .SOD主要担任超氧自由基(O2- )的清除剂,可将O2- 歧化为H2 O2 与O2 ,H2 O2 则是被CAT和POD进一步分解和消除[20 ] ,保护酶起到清除产生的活性氧、维持植物体内动态平衡的作用,保护细胞免受伤害.以往研究表明当植物受到盐胁迫和干旱胁迫时,其体内SOD、POD和CAT等保护酶的活性会增加,从而能够清除过量的活性氧,控制细胞内脂质氧化过程和抵御活性氧的伤害,维持植物体内动态平衡[8 ,21 ] .当超过一定的胁迫程度时,保护酶活性会产生下降,此时抗氧化防御功能和保护能力减弱,其活性进一步下降致使该种保护酶在抗氧化防御方面几乎不再起保护作用. ...
... 遭受逆境胁迫时,植物能够通过渗透调节系统来维持细胞内的渗透势,缓解胁迫环境对植物造成的影响和伤害.有机溶质是渗透调节系统中的主要物质,参与植物体内渗透调节过程[21 ] .盐胁迫和干旱胁迫分别会导致土壤渗透压增高和土壤水分亏缺加剧,影响土壤水势和引发植物失水.在逆境胁迫下,植物为保证从土壤中继续吸收水分,必须使自身维持较高的渗透调节能力,降低自身水势,才能形成一个正常的水势梯度实现持续吸水[23 ] ,通过增加有机溶质的含量参与渗透调节,应对外界压力从而保护细胞,使其继续从低水势条件下有效吸水[24 ] .可溶性糖是细胞质内重要的有机溶质,能够作为渗透保护剂,发挥稳定蛋白质的作用,成为植物遭受胁迫时一种重要的渗透调节物质[25 ] .可溶性糖的积累能够降低叶片的渗透势,从而有助于使细胞在水分胁迫下保持膨压状态[24 ] . ...
The threshold of soil moisture and salinity influencing the growth of Populus euphratica and Tamarix ramosissima in the extremely arid region
1
2012
... 胡杨在应对盐胁迫的过程中,SOD的活性缓慢增强,当盐胁迫增强到一定程度时,SOD的活性逐渐下降最终接近无胁迫环境的水平.胡杨在应对干旱胁迫的过程中,SOD的活性迅速增强,其增强的活性维持时间相对较短,随着干旱胁迫程度的增加,SOD的活性迅速下降最终显著低于无胁迫环境的水平.可见,胡杨面对盐胁迫相比干旱胁迫体内SOD活性增强持续时间长,而胡杨面对干旱胁迫相比盐胁迫体内SOD活性增强更为迅速,但持续时间短,后期其提供抗氧化防御的功能相对减弱,此时其他保护酶POD和CAT承担主要的抗氧化防御功能.胡杨在应对盐胁迫和干旱胁迫的过程中,POD和CAT的活性变化趋势基本相同,前期都是随着胁迫程度的增加活性增强,再随着胁迫的进一步加剧,其活性下降.由研究我们发现,当胡杨受到盐胁迫和干旱胁迫时,胁迫前期其体内SOD、POD和CAT的活性会不断增加,SOD、POD和CAT作为保护酶能够清除过量的活性氧,维持胡杨体内动态平衡,增强胡杨对逆境的抗性[22 ] .不同之处在于:胡杨在应对盐胁迫程度比较严重时,SOD、POD和CAT的活性接近无胁迫环境的水平.相反,胡杨在应对干旱胁迫程度比较严重时,SOD、POD和CAT的活性显著低于无胁迫环境的水平.可见,胡杨在应对盐胁迫环境下,能够维持较长时间保持SOD、POD和CAT的活性,维持自身的抗氧化防御功能和保护作用.但是,胡杨在应对干旱胁迫的环境下,重度干旱会导致其体内SOD、POD和CAT活性的明显减弱,这些保护酶在抗氧化防御方面功能弱化明显甚至几乎不再起保护作用,致使保护酶发挥其抗氧化防御功能的持续时间缩短,可见保护酶在逆境中并不能对植物细胞进行持续性的保护.综上所述,为了适应盐胁迫和干旱胁迫,胡杨将多种保护酶进行综合调节以形成整个抗氧化酶系统的防御功能,维持体内动态平衡应对不利环境和实现保护作用,但是不同的保护酶应对干旱胁迫和盐胁迫的反应速度和持续时间存在差异,这是胡杨启动抗氧化酶系统应对极端环境并维持生存的适应策略. ...
Leaf and whole tree adaptations to mild salinity in field grown Populus euphratica
1
2009
... 遭受逆境胁迫时,植物能够通过渗透调节系统来维持细胞内的渗透势,缓解胁迫环境对植物造成的影响和伤害.有机溶质是渗透调节系统中的主要物质,参与植物体内渗透调节过程[21 ] .盐胁迫和干旱胁迫分别会导致土壤渗透压增高和土壤水分亏缺加剧,影响土壤水势和引发植物失水.在逆境胁迫下,植物为保证从土壤中继续吸收水分,必须使自身维持较高的渗透调节能力,降低自身水势,才能形成一个正常的水势梯度实现持续吸水[23 ] ,通过增加有机溶质的含量参与渗透调节,应对外界压力从而保护细胞,使其继续从低水势条件下有效吸水[24 ] .可溶性糖是细胞质内重要的有机溶质,能够作为渗透保护剂,发挥稳定蛋白质的作用,成为植物遭受胁迫时一种重要的渗透调节物质[25 ] .可溶性糖的积累能够降低叶片的渗透势,从而有助于使细胞在水分胁迫下保持膨压状态[24 ] . ...
Eco-morphological response of floodplain forests (Populus euphratica Oliv.) to water diversion in the lower Tarim River,northwest China
2
2015
... 遭受逆境胁迫时,植物能够通过渗透调节系统来维持细胞内的渗透势,缓解胁迫环境对植物造成的影响和伤害.有机溶质是渗透调节系统中的主要物质,参与植物体内渗透调节过程[21 ] .盐胁迫和干旱胁迫分别会导致土壤渗透压增高和土壤水分亏缺加剧,影响土壤水势和引发植物失水.在逆境胁迫下,植物为保证从土壤中继续吸收水分,必须使自身维持较高的渗透调节能力,降低自身水势,才能形成一个正常的水势梯度实现持续吸水[23 ] ,通过增加有机溶质的含量参与渗透调节,应对外界压力从而保护细胞,使其继续从低水势条件下有效吸水[24 ] .可溶性糖是细胞质内重要的有机溶质,能够作为渗透保护剂,发挥稳定蛋白质的作用,成为植物遭受胁迫时一种重要的渗透调节物质[25 ] .可溶性糖的积累能够降低叶片的渗透势,从而有助于使细胞在水分胁迫下保持膨压状态[24 ] . ...
... [24 ]. ...
盐胁迫对景天三七保护酶系统、MDA、Pro及可溶性糖的影响
1
2009
... 遭受逆境胁迫时,植物能够通过渗透调节系统来维持细胞内的渗透势,缓解胁迫环境对植物造成的影响和伤害.有机溶质是渗透调节系统中的主要物质,参与植物体内渗透调节过程[21 ] .盐胁迫和干旱胁迫分别会导致土壤渗透压增高和土壤水分亏缺加剧,影响土壤水势和引发植物失水.在逆境胁迫下,植物为保证从土壤中继续吸收水分,必须使自身维持较高的渗透调节能力,降低自身水势,才能形成一个正常的水势梯度实现持续吸水[23 ] ,通过增加有机溶质的含量参与渗透调节,应对外界压力从而保护细胞,使其继续从低水势条件下有效吸水[24 ] .可溶性糖是细胞质内重要的有机溶质,能够作为渗透保护剂,发挥稳定蛋白质的作用,成为植物遭受胁迫时一种重要的渗透调节物质[25 ] .可溶性糖的积累能够降低叶片的渗透势,从而有助于使细胞在水分胁迫下保持膨压状态[24 ] . ...
Superoxide dismutase and stress tolerance
2
1992
... 胡杨在应对盐胁迫和干旱胁迫的过程中,可溶性糖的含量变化趋势基本相同,都是随着胁迫程度的增加进行持续性积累.胡杨在应对盐胁迫和干旱胁迫的过程中,其体内可溶性糖的含量不断增加,这样体内通过可溶性糖的积累,能够提高细胞渗透压和细胞膨压,从而维持渗透调节平衡,增强胡杨对逆境的抗性.不同之处在于:胡杨在应对盐胁迫的过程中,环境盐分浓度越大,随着胁迫时间的增加,前期胡杨可溶性糖积累越慢.当盐浓度达到一定程度时,随着胁迫时间的增加,前期胡杨可溶性糖含量基本维持稳定.相反,胡杨所处盐环境的浓度越低,其可溶性糖等渗透调节物质能够进行更快的反应调节和更长时间地维持渗透调节.胡杨在应对干旱胁迫的过程中,随着胁迫时间的增加,前期胡杨可溶性糖也基本维持稳定,后期迅速积累.因此,胡杨在盐浓度较高和干旱胁迫的条件下,前期主要依靠保护酶的积累和抗氧化酶系统的抗氧化防御功能发挥保护作用,可溶性糖的含量基本不变,随着抗氧化酶系统保护作用不断减弱,可溶性糖含量持续性增加,增强渗透调节系统的保护作用.在面对长期极端环境时,抗氧化酶系统能够提供有效的前期保护,由于其保护作用缺乏长效性,后期主要依靠增强渗透调节系统的功能实现长期保护效果[26 -27 ] .综上所述,为了适应盐胁迫和干旱胁迫,胡杨通过持续性积累可溶性糖等有机物质维持渗透平衡以形成渗透调节系统的长效防护功能,维持渗透势应对不利环境和实现保护作用,但是渗透调节物质积累应对不同程度的盐胁迫和干旱胁迫的反应速度和持续时间存在差异,这是胡杨启动渗透调节系统应对极端环境并维持生存的适应策略. ...
... 植物面对长期逆境胁迫时,除了渗透调节系统提供长效防护功能外,长期的保护还得依赖膜结构的功能来实现[26 -27 ] .细胞只有在膜系统正常运转的情况下才能维持各个系统的平衡和功能的发挥,无论是抗氧化酶系统还是渗透调节系统,其发挥保护作用和渗透调节作用都需要通过细胞膜来进行物质和能量的交换,维持细胞膜系统的稳定性对植物在逆境中生存至关重要[28 ] .MDA是膜脂过氧化的产物,使细胞膜本身受损,同时MDA自身会与蛋白质和核酸发生反应,从而破坏膜结构,因此MDA能够综合反映细胞膜功能受损情况[17 ] .在盐胁迫下胞内大量离子、有机物质的外渗,以及有毒盐离子的进入,造成细胞内生理生化过程受到干扰,植物细胞质膜受到损伤,细胞膜系统是植物遭受盐胁迫的主要部位[29 ] .同样,干旱胁迫会导致细胞原生质脱水,体内形成大量的自由基,膜脂过氧化水平增高和膜脂成分改变,膜系统受到破坏,干旱对细胞的影响表现在使细胞膜受到伤害[30 -31 ] . ...
Introduction to ecological biochemistry
2
2010
... 胡杨在应对盐胁迫和干旱胁迫的过程中,可溶性糖的含量变化趋势基本相同,都是随着胁迫程度的增加进行持续性积累.胡杨在应对盐胁迫和干旱胁迫的过程中,其体内可溶性糖的含量不断增加,这样体内通过可溶性糖的积累,能够提高细胞渗透压和细胞膨压,从而维持渗透调节平衡,增强胡杨对逆境的抗性.不同之处在于:胡杨在应对盐胁迫的过程中,环境盐分浓度越大,随着胁迫时间的增加,前期胡杨可溶性糖积累越慢.当盐浓度达到一定程度时,随着胁迫时间的增加,前期胡杨可溶性糖含量基本维持稳定.相反,胡杨所处盐环境的浓度越低,其可溶性糖等渗透调节物质能够进行更快的反应调节和更长时间地维持渗透调节.胡杨在应对干旱胁迫的过程中,随着胁迫时间的增加,前期胡杨可溶性糖也基本维持稳定,后期迅速积累.因此,胡杨在盐浓度较高和干旱胁迫的条件下,前期主要依靠保护酶的积累和抗氧化酶系统的抗氧化防御功能发挥保护作用,可溶性糖的含量基本不变,随着抗氧化酶系统保护作用不断减弱,可溶性糖含量持续性增加,增强渗透调节系统的保护作用.在面对长期极端环境时,抗氧化酶系统能够提供有效的前期保护,由于其保护作用缺乏长效性,后期主要依靠增强渗透调节系统的功能实现长期保护效果[26 -27 ] .综上所述,为了适应盐胁迫和干旱胁迫,胡杨通过持续性积累可溶性糖等有机物质维持渗透平衡以形成渗透调节系统的长效防护功能,维持渗透势应对不利环境和实现保护作用,但是渗透调节物质积累应对不同程度的盐胁迫和干旱胁迫的反应速度和持续时间存在差异,这是胡杨启动渗透调节系统应对极端环境并维持生存的适应策略. ...
... 植物面对长期逆境胁迫时,除了渗透调节系统提供长效防护功能外,长期的保护还得依赖膜结构的功能来实现[26 -27 ] .细胞只有在膜系统正常运转的情况下才能维持各个系统的平衡和功能的发挥,无论是抗氧化酶系统还是渗透调节系统,其发挥保护作用和渗透调节作用都需要通过细胞膜来进行物质和能量的交换,维持细胞膜系统的稳定性对植物在逆境中生存至关重要[28 ] .MDA是膜脂过氧化的产物,使细胞膜本身受损,同时MDA自身会与蛋白质和核酸发生反应,从而破坏膜结构,因此MDA能够综合反映细胞膜功能受损情况[17 ] .在盐胁迫下胞内大量离子、有机物质的外渗,以及有毒盐离子的进入,造成细胞内生理生化过程受到干扰,植物细胞质膜受到损伤,细胞膜系统是植物遭受盐胁迫的主要部位[29 ] .同样,干旱胁迫会导致细胞原生质脱水,体内形成大量的自由基,膜脂过氧化水平增高和膜脂成分改变,膜系统受到破坏,干旱对细胞的影响表现在使细胞膜受到伤害[30 -31 ] . ...
水分胁迫对幼苗期霸王叶片生理特性的影响
1
2011
... 植物面对长期逆境胁迫时,除了渗透调节系统提供长效防护功能外,长期的保护还得依赖膜结构的功能来实现[26 -27 ] .细胞只有在膜系统正常运转的情况下才能维持各个系统的平衡和功能的发挥,无论是抗氧化酶系统还是渗透调节系统,其发挥保护作用和渗透调节作用都需要通过细胞膜来进行物质和能量的交换,维持细胞膜系统的稳定性对植物在逆境中生存至关重要[28 ] .MDA是膜脂过氧化的产物,使细胞膜本身受损,同时MDA自身会与蛋白质和核酸发生反应,从而破坏膜结构,因此MDA能够综合反映细胞膜功能受损情况[17 ] .在盐胁迫下胞内大量离子、有机物质的外渗,以及有毒盐离子的进入,造成细胞内生理生化过程受到干扰,植物细胞质膜受到损伤,细胞膜系统是植物遭受盐胁迫的主要部位[29 ] .同样,干旱胁迫会导致细胞原生质脱水,体内形成大量的自由基,膜脂过氧化水平增高和膜脂成分改变,膜系统受到破坏,干旱对细胞的影响表现在使细胞膜受到伤害[30 -31 ] . ...
1
1993
... 植物面对长期逆境胁迫时,除了渗透调节系统提供长效防护功能外,长期的保护还得依赖膜结构的功能来实现[26 -27 ] .细胞只有在膜系统正常运转的情况下才能维持各个系统的平衡和功能的发挥,无论是抗氧化酶系统还是渗透调节系统,其发挥保护作用和渗透调节作用都需要通过细胞膜来进行物质和能量的交换,维持细胞膜系统的稳定性对植物在逆境中生存至关重要[28 ] .MDA是膜脂过氧化的产物,使细胞膜本身受损,同时MDA自身会与蛋白质和核酸发生反应,从而破坏膜结构,因此MDA能够综合反映细胞膜功能受损情况[17 ] .在盐胁迫下胞内大量离子、有机物质的外渗,以及有毒盐离子的进入,造成细胞内生理生化过程受到干扰,植物细胞质膜受到损伤,细胞膜系统是植物遭受盐胁迫的主要部位[29 ] .同样,干旱胁迫会导致细胞原生质脱水,体内形成大量的自由基,膜脂过氧化水平增高和膜脂成分改变,膜系统受到破坏,干旱对细胞的影响表现在使细胞膜受到伤害[30 -31 ] . ...
盐胁迫对竹柳种苗形态及生理指标的影响
1
2011
... 植物面对长期逆境胁迫时,除了渗透调节系统提供长效防护功能外,长期的保护还得依赖膜结构的功能来实现[26 -27 ] .细胞只有在膜系统正常运转的情况下才能维持各个系统的平衡和功能的发挥,无论是抗氧化酶系统还是渗透调节系统,其发挥保护作用和渗透调节作用都需要通过细胞膜来进行物质和能量的交换,维持细胞膜系统的稳定性对植物在逆境中生存至关重要[28 ] .MDA是膜脂过氧化的产物,使细胞膜本身受损,同时MDA自身会与蛋白质和核酸发生反应,从而破坏膜结构,因此MDA能够综合反映细胞膜功能受损情况[17 ] .在盐胁迫下胞内大量离子、有机物质的外渗,以及有毒盐离子的进入,造成细胞内生理生化过程受到干扰,植物细胞质膜受到损伤,细胞膜系统是植物遭受盐胁迫的主要部位[29 ] .同样,干旱胁迫会导致细胞原生质脱水,体内形成大量的自由基,膜脂过氧化水平增高和膜脂成分改变,膜系统受到破坏,干旱对细胞的影响表现在使细胞膜受到伤害[30 -31 ] . ...
干旱胁迫下菊花叶片的生理响应及抗旱性评价
1
2011
... 植物面对长期逆境胁迫时,除了渗透调节系统提供长效防护功能外,长期的保护还得依赖膜结构的功能来实现[26 -27 ] .细胞只有在膜系统正常运转的情况下才能维持各个系统的平衡和功能的发挥,无论是抗氧化酶系统还是渗透调节系统,其发挥保护作用和渗透调节作用都需要通过细胞膜来进行物质和能量的交换,维持细胞膜系统的稳定性对植物在逆境中生存至关重要[28 ] .MDA是膜脂过氧化的产物,使细胞膜本身受损,同时MDA自身会与蛋白质和核酸发生反应,从而破坏膜结构,因此MDA能够综合反映细胞膜功能受损情况[17 ] .在盐胁迫下胞内大量离子、有机物质的外渗,以及有毒盐离子的进入,造成细胞内生理生化过程受到干扰,植物细胞质膜受到损伤,细胞膜系统是植物遭受盐胁迫的主要部位[29 ] .同样,干旱胁迫会导致细胞原生质脱水,体内形成大量的自由基,膜脂过氧化水平增高和膜脂成分改变,膜系统受到破坏,干旱对细胞的影响表现在使细胞膜受到伤害[30 -31 ] . ...
Effect of salt stress on growth parameters,enzymatic antioxidant system,and lipid peroxidation in wild chicory (Cichorium intybus L.)
1
2012
... 胡杨在应对盐胁迫的过程中,随着胁迫时间的增加,胁迫初期MDA含量缓慢降低,然后MDA含量再缓慢增加最终基本达到无胁迫环境的水平.这是因为当胡杨受到盐分胁迫时,SOD、POD和CAT等保护酶活性的增加强化了其抗氧化酶系统的防御功能,从而可以减少膜中多不饱和脂肪酸过氧化作用的发生,能够抑制MDA的产生;同时可溶性糖等有机物质的积累增强了渗透调节系统的功能,从而可以维持渗透平衡,防止胞内离子的外渗和外界有毒盐离子的进入,减少其细胞质膜受损.可见,强化抗氧化酶系统和渗透调节系统的综合作用一定程度能够减少MDA的产生并促使细胞膜系统维持其生理功能.因此,当胡杨受到盐胁迫时前期抗氧化酶系统和渗透调节系统的增强使MDA含量降低.盐胁迫增加后期MDA含量缓慢增加基本达到无胁迫环境的水平,可见,胡杨面对盐胁迫时其体内细胞膜能够相对维持长时间的稳定性.胡杨在面临盐胁迫时,MDA含量基本维持在较低的水平,说明低MDA水平可能对耐盐性有显著影响,这与之前研究结果一致[32 -35 ] .胡杨在应对干旱胁迫的过程中,胁迫初期MDA含量基本维持稳定,然后随着胁迫程度的增加,MDA含量逐渐增加.胡杨面对轻度干旱胁迫时体内细胞膜也能够基本维持稳定,而干旱胁迫程度进一步增加时其体内细胞膜受损严重,细胞膜系统无法进行有效调节.干旱胁迫对于胡杨细胞膜系统的损伤远远大于盐胁迫的影响,这很可能是因为MDA含量不断增加会降低细胞膜的收缩性,使导管壁果胶变得松弛和膨胀,阻碍了植物水分传输过程,进一步加剧了干旱缺水环境的不利影响[36 ] .综上所述,胡杨能够在盐胁迫和轻度干旱胁迫下通过细胞膜系统的适应性调节维持细胞膜结构功能的完整性,实现细胞膜系统的保护作用,这是胡杨启动细胞膜系统应对极端环境并维持生存的适应策略. ...
Salinity up-regulates the antioxidative system in root mitochondria and peroxisomes of the wild salt-tolerant tomato species Lycopersicon pennellii
0
2004
Nicotine-free and salt-tolerant tobacco plants obtained by grafting to salinity-resistant rootstocks of tomato
0
2005
Exogenous silicon (Si) increases antioxidant enzyme activity and reduces lipid peroxidation in roots of salt-stressed barley (Hordeum vulgare L.)
1
2003
... 胡杨在应对盐胁迫的过程中,随着胁迫时间的增加,胁迫初期MDA含量缓慢降低,然后MDA含量再缓慢增加最终基本达到无胁迫环境的水平.这是因为当胡杨受到盐分胁迫时,SOD、POD和CAT等保护酶活性的增加强化了其抗氧化酶系统的防御功能,从而可以减少膜中多不饱和脂肪酸过氧化作用的发生,能够抑制MDA的产生;同时可溶性糖等有机物质的积累增强了渗透调节系统的功能,从而可以维持渗透平衡,防止胞内离子的外渗和外界有毒盐离子的进入,减少其细胞质膜受损.可见,强化抗氧化酶系统和渗透调节系统的综合作用一定程度能够减少MDA的产生并促使细胞膜系统维持其生理功能.因此,当胡杨受到盐胁迫时前期抗氧化酶系统和渗透调节系统的增强使MDA含量降低.盐胁迫增加后期MDA含量缓慢增加基本达到无胁迫环境的水平,可见,胡杨面对盐胁迫时其体内细胞膜能够相对维持长时间的稳定性.胡杨在面临盐胁迫时,MDA含量基本维持在较低的水平,说明低MDA水平可能对耐盐性有显著影响,这与之前研究结果一致[32 -35 ] .胡杨在应对干旱胁迫的过程中,胁迫初期MDA含量基本维持稳定,然后随着胁迫程度的增加,MDA含量逐渐增加.胡杨面对轻度干旱胁迫时体内细胞膜也能够基本维持稳定,而干旱胁迫程度进一步增加时其体内细胞膜受损严重,细胞膜系统无法进行有效调节.干旱胁迫对于胡杨细胞膜系统的损伤远远大于盐胁迫的影响,这很可能是因为MDA含量不断增加会降低细胞膜的收缩性,使导管壁果胶变得松弛和膨胀,阻碍了植物水分传输过程,进一步加剧了干旱缺水环境的不利影响[36 ] .综上所述,胡杨能够在盐胁迫和轻度干旱胁迫下通过细胞膜系统的适应性调节维持细胞膜结构功能的完整性,实现细胞膜系统的保护作用,这是胡杨启动细胞膜系统应对极端环境并维持生存的适应策略. ...
胡杨(Populus euphratica )对干旱胁迫的生态适应
1
2020
... 胡杨在应对盐胁迫的过程中,随着胁迫时间的增加,胁迫初期MDA含量缓慢降低,然后MDA含量再缓慢增加最终基本达到无胁迫环境的水平.这是因为当胡杨受到盐分胁迫时,SOD、POD和CAT等保护酶活性的增加强化了其抗氧化酶系统的防御功能,从而可以减少膜中多不饱和脂肪酸过氧化作用的发生,能够抑制MDA的产生;同时可溶性糖等有机物质的积累增强了渗透调节系统的功能,从而可以维持渗透平衡,防止胞内离子的外渗和外界有毒盐离子的进入,减少其细胞质膜受损.可见,强化抗氧化酶系统和渗透调节系统的综合作用一定程度能够减少MDA的产生并促使细胞膜系统维持其生理功能.因此,当胡杨受到盐胁迫时前期抗氧化酶系统和渗透调节系统的增强使MDA含量降低.盐胁迫增加后期MDA含量缓慢增加基本达到无胁迫环境的水平,可见,胡杨面对盐胁迫时其体内细胞膜能够相对维持长时间的稳定性.胡杨在面临盐胁迫时,MDA含量基本维持在较低的水平,说明低MDA水平可能对耐盐性有显著影响,这与之前研究结果一致[32 -35 ] .胡杨在应对干旱胁迫的过程中,胁迫初期MDA含量基本维持稳定,然后随着胁迫程度的增加,MDA含量逐渐增加.胡杨面对轻度干旱胁迫时体内细胞膜也能够基本维持稳定,而干旱胁迫程度进一步增加时其体内细胞膜受损严重,细胞膜系统无法进行有效调节.干旱胁迫对于胡杨细胞膜系统的损伤远远大于盐胁迫的影响,这很可能是因为MDA含量不断增加会降低细胞膜的收缩性,使导管壁果胶变得松弛和膨胀,阻碍了植物水分传输过程,进一步加剧了干旱缺水环境的不利影响[36 ] .综上所述,胡杨能够在盐胁迫和轻度干旱胁迫下通过细胞膜系统的适应性调节维持细胞膜结构功能的完整性,实现细胞膜系统的保护作用,这是胡杨启动细胞膜系统应对极端环境并维持生存的适应策略. ...




 甘公网安备 62010202000688号
甘公网安备 62010202000688号
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}