晋西北丘陵风沙区不同种植年限柠条( Caragana korshinskii
张宇 , 1 , 缑倩倩 , 1 , 2 , 高敏 1 , 张妍 1 , 郭文婷 1 , 王国华 1 , 2 , 3
1.山西师范大学 地理科学学院,山西 太原 030031
2.中国科学院西北生态环境资源研究院,临泽内陆河流域研究站,甘肃 兰州 730000
3.中国科学院西北生态环境资源研究院,沙漠与沙漠化重点实验室,甘肃 兰州 730000
The response mechanism of early seed regeneration process of Caragana korshinskii bushes with different plantation ages in sandy-hilly region of northwest Shanxi Province
Zhang Yu , 1 , Gou Qianqian , 1 , 2 , Gao Min 1 , Zhang Yan 1 , Guo Wentin 1 , Wang Guohua 1 , 2 , 3
1.College of Geographic Sciences,Shanxi Normal University,Taiyuan 030031,China
2.Linze Inland River Basin Research Station /, Northwest Institute of Eco-Environment and Resources,Chinese Academy of Sciences,Lanzhou 730000,China
3.Key Laboratory of Desert and Desertification, Northwest Institute of Eco-Environment and Resources,Chinese Academy of Sciences,Lanzhou 730000,China
通讯作者: 缑倩倩(E-mail: gqqqianqian@163.com )
收稿日期: 2022-03-21
修回日期: 2022-05-20
基金资助:
国家自然科学基金项目 . 41807518 . 42171033 山西省高等学校科技创新项目 . 2019L0457 . 2019L0463 中国科学院沙漠和沙漠化重点实验室开放基金项目 . KLDD-2020-05
Received: 2022-03-21
Revised: 2022-05-20
摘要
土壤干燥化是风沙区人工林种子萌发、更新幼苗存活与生长、种子更新的关键限制因子。选取4个种植年限(7、13、19、51 a)柠条(Caragana korshinskii )种子为研究对象,通过盆栽控水模拟不同程度干旱胁迫,分析重度、中度和轻度干旱对不同年限柠条种子萌发、幼苗生长、生理及光合特性的影响。结果表明:干旱胁迫显著抑制各年限柠条种子出苗率,出苗率随干旱程度增加而降低,13 a出苗率均高于其他年限,19 a出苗率次之,51 a最低。随着干旱程度的增加,各年限幼苗株高、叶片数及总生物量均显著下降,13 a与其他年限相比减少量最小;同时,各年限幼苗根冠比均增加。不同年限柠条幼苗叶片可溶性糖含量随干旱程度增加均呈上升趋势,其中13 a含量最高,为30.19 mg·g-1 ;游离脯氨酸含量也随干旱的加剧呈增加趋势,其中19 a含量最高,为2 683.73 μg·g-1 。各年限幼苗丙二醛(MDA)含量在干旱胁迫下逐渐升高,在重度胁迫时达到最大,51 a增幅最大。不同年限柠条幼苗叶绿素a、b及叶绿素含量在干旱胁迫下总体呈先降低再升高最后降低的趋势,下降最多的为51 a幼苗叶片。在不同程度的干旱胁迫下,各年限柠条幼苗的生长均受到一定程度的抑制,但13 a和19 a能够通过自身渗透调节物质来减轻伤害,维持正常生理代谢,保障幼苗生长发育,而51 a幼苗渗透调节能力较低,MDA含量最大,使细胞膜损坏严重,导致死亡率显著增加。隶属函数法表明,不同年限柠条幼苗抗旱能力13 a>19 a>7 a>51 a。
关键词:
干旱胁迫 柠条(Caragana korshinskii )幼苗 生理特征 生长性状 天然更新
Abstract
Soil desiccation is the key limiting factor for seed germination, survival and growth of regenerated seedlings, and seed regeneration in artificial bushes in sandy areas. In this study, we selected Caragana korshinskii of different planting ages (7,13,19,51 years) as research subjects, and analyzed the effects of severe, moderate and light drought stress on seed germination, seedling growth, physiology and photosynthetic characteristics of C. korshinskii through controlling of soil water in pot culture. The results showed that the seed emergence rate of C.korshinskii significantly decreased with the increased drought stress. The seed emergence rate of C. korshinskii at 13 years was higher than other years, followed by 19 years, and the lowest kept at 51 years. The plant heigh, leaf numbers and total biomass of seedlings decreased significantly with increased of drought severity, and the plant traits were higher at 13 years compared with other ages. The root-shoot ratio of seedlings increased in each age. With increased of drought severity, the soluble sugar content of C. korshinskii seedlings in different ages increased, and the content at 13 years was the highest, which was 30.19 mg·g-1 . The content of free proline also increased, and the content of 19 years was the highest,which was 2 683.73 μg·g-1 . MDA content in seedlings of all ages increased gradually under drought stress, and reached a maximum value under severe drought stress, the content about MDA had the largest increase in 51 years. Under drought stress, chlorophyll a, b and chlorophyll content of C. korshinskii seedlings in different ages firstly decreased, then increased, and finally decreased again, and the most decreased occurred at 51 years. The plant traits of C. korshinskii seedlings were influenced by different degree of drought stress, but the seedlings at 13 years and 19 years could adjust the osmotically regulated substance to reduce damage and to maintain normal physiological metabolism and seedling growth. However, the lower osmotic regulation ability with the highest MDA content of 51 years, which caused serious damage to the cell membrane, resulting in a significant increase in mortality. According to the membership functions showed that the drought tolerance of C. korshinskii seedlings of different ages was 13 years>19 years>7 years>51 years.
Keywords:
drought stress Caragana korshinskii seedlingsphysiological characteristics growth traits natural regeneration
本文引用格式
张宇, 缑倩倩, 高敏, 张妍, 郭文婷, 王国华. 晋西北丘陵风沙区不同种植年限柠条( Caragana korshinskii . 中国沙漠 [J], 2023, 43(2): 243-254 doi:10.7522/j.issn.1000-694X.2022.00083
Zhang Yu, Gou Qianqian, Gao Min, Zhang Yan, Guo Wentin, Wang Guohua. The response mechanism of early seed regeneration process of Caragana korshinskii bushes with different plantation ages in sandy-hilly region of northwest Shanxi Province . Journal of Desert Research
0 引言
沙漠与荒漠草原过渡区是中国沙漠化与风沙活动危害最严重的区域,横跨中国北方地区且分布于极端干旱、干旱、半干旱等不同气候带[1 -2 ] 。为防止沙化土地进一步扩张,国家先后在风沙区启动了“三北”防护林、退耕还林和京津风沙源治理等以人工植被建设为主的一批重大生态建设工程[3 ] 。目前,中国人工植被面积已达6 933万hm2 ,占全国林地面积的36%[1 ] 。但在风沙治理区,大面积人工植被产生了退化、老化以及更新困难等问题[4 ] 。引起固沙人工林生长衰退的因素有很多,如固沙树种选择、种植密度、林分结构以及环境条件等[5 ] 。因此,了解并掌握人工林天然更新所需条件对其实践经营具有重要意义。
柠条(Caragana korshinskill )是中国干旱半干旱地区的主要固沙植物,具有耐旱、抗风沙、根系发达等特征,在保持水土以及改善生态环境、缓解北方沙尘暴灾害等方面起到了积极有效的作用[6 ] ,作为主要固沙树种受到极大关注。目前,国内外关于干旱对柠条锦鸡儿属植物在萌发、生长、繁殖及生理代谢等方面的影响已有大量报道。在宁夏荒漠草原地区研究发现,柠条种子萌发率和萌发速率随着干旱的加剧呈先上升后下降的趋势[7 ] ;在阿拉善地区有研究表明,中度干旱使得柠条叶生物量、地上生物量和总叶片数目显著降低,但根冠比有所增加[8 ] ;在西北干旱风沙区有研究发现,干旱的加剧可使柠条叶片中游离脯氨酸和可溶性糖含量的积累量均持续上升[9 ] ,且严重干旱对其叶片叶绿体结构造成不可恢复的破坏[10 ] 。关于不同年限柠条的研究集中在生物量分配[11 -12 ] 以及根系特征的变化[13 -14 ] 等。植物营养可利用能力随林龄的增加而降低[15 ] ,且植物叶片的光合能力均低于幼龄[16 ] 。然而,目前对长时间尺度下人工柠条林地干旱导致人工柠条林天然更新困难的研究还较少。
晋西北丘陵风沙区地处中国沙漠化扩展前沿,土壤沙化严重,是干旱半干旱地区中典型的生态过渡带与环境脆弱区,也是京津能源、水源和沙源之地,该地区长期以来成为“三北”防护林建设的重点区域[17 ] 。随着植被恢复工程的深入实施,柠条逐渐成为当地的优势种,并且形成了老龄、中龄和幼龄3个不同发育阶段,这使得人工柠条林稳定且持续性的发展受到了极大关注。在干旱半干旱地区,土壤水分在林木生长中起重要作用,同时也是治沙造林的主要限制因子[18 ] 。一般认为,种子更新发生的必要条件是有充足且具备生命力的种子以及有利于种子萌发的环境和幼苗生长的气候条件[19 ] ,而更新的关键过程在于种子萌发与幼苗存活生长[20 ] 。在天然条件下,柠条虽然可以成功结实并能够完成有性繁殖,但种子的萌发率和幼苗的存活率极低[21 ] 。在毛乌素沙地柠条种子遇少量降雨能够萌发,但降雨之后长期的干旱会极度增大幼苗死亡的风险[22 ] ;在黄土高原丘陵沟壑区,由于水分限制,也极少有实生幼苗能够成活[23 -24 ] ;同时,种子萌发的好坏也会直接影响到幼苗的成活率[25 ] ,尤其是在干旱、半干旱的荒漠地区,适合种子萌发和低龄幼苗生长、存活的环境条件极其有限[26 ] ,虽然柠条根系较为发达,耐旱能力强,但低龄柠条幼苗也同样极易遭受干旱胁迫[27 ] 。在晋西北丘陵风沙区,不同年限柠条均能顺利结实[28 ] ,但由于当地土壤干燥化发生频繁,使得人工柠条林地出现大面积退化现象,严重制约了当地植被建设的成效以及区域生态的稳定[29 ] 。因此,无论是在干旱、半干旱的荒漠区还是晋西北丘陵风沙区,柠条种子更新都与水分有关。但是,不同种植年限柠条林种子更新早期过程在干旱胁迫下是否能够顺利进行却不十分清楚。因此,本研究采用盆栽控制水分模拟干旱的方法,以晋西北丘陵风沙区不同年限柠条种子为试验材料,分析干旱胁迫对不同年限柠条种子萌发以及幼苗生理、生长状况的影响,通过主成分分析和隶属函数法对其抗旱性进行综合评价,探讨各年限柠条幼苗渗透调节物质、膜脂过氧化物、光合指标、株高、主根长以及叶片数等指标与抗旱性的关系,明确在苗期抗旱性的关键影响因素,为晋西北丘陵风沙区人工柠条林的经营管理与可持续发展提供指导,保障其发挥生态防护功能。
1 材料与方法
1.1 研究区概况
研究区位于晋西北丘陵风沙区的忻州市五寨县,气候属于温带大陆性气候,春季干旱、多大风天气,夏秋季节降水多,年降水量478.5 mm,季节分配不均而且变率较大。年平均气温4.1~5.5 ℃,最冷月(1月)平均气温-13.2 ℃,最热月(7月)平均气温20.0 ℃。年蒸发量2 000~2 300 mm,平均年日照时间2 872 h,平均无霜期125 d。研究区土壤类型以黄绵土为主,土壤肥力较低。研究区内人工植被覆盖,乔木主要有小叶杨(Populus simonii )、油松(Pinus tabulaeformis )和旱柳(Salix matsudana );灌木主要为柠条;人工林下主要有狗尾草(Setaria viridis )、胡枝子(Lespedeza bicolor )和披碱草(Elymus dahuricus )等。
1.2 试验设计
1.2.1 试验材料
试验所用种子于2020年7—9月采自晋西北丘陵风沙区不同年限(7、13、19、51 a)柠条灌木,同一年限柠条选取5株长势良好且均匀的柠条灌丛,采取成熟健康的种子,各年限种子自然风干并充分混合(表1 )。将所采集的土壤过1 mm孔径土壤筛,并于105 ℃烘箱中烘干48 h,使土壤中没有种子可以萌发。
1.2.2 试验方法
试验依据梁海斌等[30 ] 对晋西北丘陵风沙区土壤水分的研究,结合相关学者提出关于植物水分梯度划分的方法[31 ] ,设置适宜水分及轻度、中度和重度干旱胁迫,分别为土壤含水量占田间持水量的70%~75%、55%~60%、40%~45%和30%~35%。
盆栽培植试验[32 ] 于2021年4月19日进行人工控制条件下的胁迫处理,将所采集的种子置于室内黑暗干燥处风干储存备用。花盆直径和深度均为15 cm,内装黄绵土,每个年限各处理3盆为3个重复。所用花盆用烧杯等量浇水,通过自然消耗至设定土壤含水量开始进行播种,选取籽粒饱满、大小基本一致且无病虫害的种子,不同年限的种子单独播种于花盆内,每盆播种20粒,行距和播深均为2 cm。为防止系统误差,将花盆随机摆放并用标签进行标记。在种子出苗及植株生长期间按不同水分梯度进行胁迫处理,用烘干和称重相结合的方法控制土壤含水量,每天18:00补充水分,使各处理土壤水分含量控制在设定范围内,为防止盆内水分蒸发,对花盆进行套袋处理。晴天自然光照,雨天遮雨棚覆盖,以防止天然降水的影响。每天记录出苗数,当幼苗数量达到最大后2周内再无幼苗出土时,视为萌发结束。在萌发结束3 d后开始干旱胁迫处理,胁迫持续时长为30 d,试验结束统一收取植株,将样品迅速带回实验室,放入4 ℃低温冰箱保存,测定各项指标。
1.2.3 测定指标及方法
土壤 含水 量 = 原土 重 - 烘干 土重 / 烘干 土重 × 100 % (1)
出苗 率 = 出土 幼苗 数 / 供试 种子 数 × 100 % (2)
死亡 率 = 出土 幼苗 数 - 收获 幼苗 数 / 出土 幼苗 数 × 100 % (3)
丙二醛含量的测定用硫代巴比妥酸(TBA)法[33 ] ;游离脯氨酸含量的测定用茚三酮显色法[33 ] ;叶绿素含量的测定用80%丙酮法[33 ] ;可溶性糖含量的测定用蒽酮比色法[33 ] 。
在试验结束时测定幼苗的根长、株高和叶片数;茎长和根长用直接测量法,用精确到0.01 cm的直尺测量。株高测量根部以上部分;根长测量植物的主根长。
将植株的根、茎和叶于实验室晾干至恒重,测定茎生物量、叶生物量和根生物量,其中根生物量即为地下生物量。根据以下公式计算地上生物量、总生物量、根冠比。
地上 生物 量 = 茎生 物量 + 叶生 物量 (4)
总生 物量 = 地上 生物 量 + 地下 生物 量 (5)
根冠 比 = 根生 物量 / 地上 生物 量 (6)
1.3 抗旱性综合评价
利用主成分分析法将4个年限柠条苗期的15个指标(种子出苗率;幼苗生理指标:游离脯氨酸、可溶性糖和丙二醛;幼苗光合指标:叶绿素、叶绿素a和叶绿素b;幼苗生长指标:主根长、株高、叶片数、根生物量、茎生物量、叶生物量和根冠比;幼苗死亡率)降维并筛选出抗旱性综合指标,并采用隶属函数法进行抗旱性综合评价。
X u = ( X - X m i n ) / ( X m a x - X m i n ) (7)
式中:X u X X m a x X m i n
X u = 1 - ( X - X m i n ) / ( X m a x - X m i n ) (8)
运用式(7)或式(8)求出不同年限柠条种子在干旱胁迫下的隶属函数值,累加各指标的具体隶属值,并求出平均值后进行比较,平均值越大植物的抗旱性越强[34 ] 。
1.4 数据处理
采用SPSS21.0基于one-way ANOVA和Duncan法进行方差分析和显著性检验(α =0.05),用主成分分析及相关性方法分析各指标之间的关系。利用Origin2018作图软件完成图件绘制。
2 结果与分析
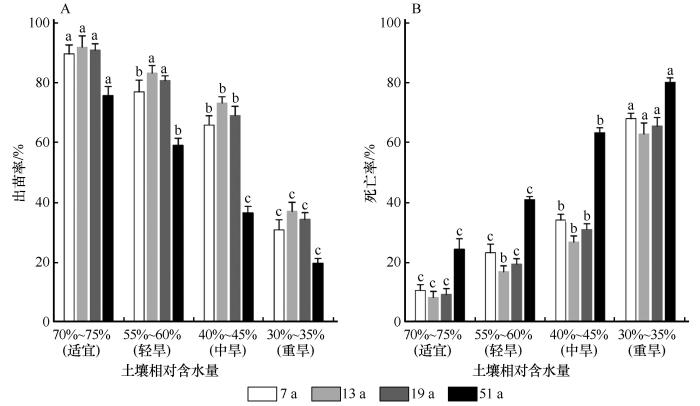
2.1 干旱胁迫对不同年限柠条种子出苗率和死亡率的影响
不同年限柠条种子出苗率在干旱胁迫下差异显著(P <0.05),均随着干旱胁迫程度的增加呈下降趋势(图1 )。在对照处理下,7 a、13 a和19 a种子出苗率均达到了90%及以上,51 a最低为75%;在重度胁迫下,51 a种子出苗率最低,仅有19.56%,而其他年限出苗率均高于51 a。不同年限柠条幼苗死亡率在干旱胁迫下差异显著(P <0.05),均随干旱程度的增加呈上升趋势。各年限幼苗死亡率在重度胁迫下达到最大,分别为69%、62.83%、65.5%和80.17%,51 a在重度胁迫下死亡率最高。
图1
图1
干旱胁迫下不同年限柠条种子出苗率和死亡率(不同字母代表同一年限不同干旱胁迫差异显著,P <0.05)
Fig.1
Seed emergence rate and mortality of Caragana korshinskii in different ages under drought stress (Different letters indicate the same age plants has significant differences in different drought stress,P <0.05)
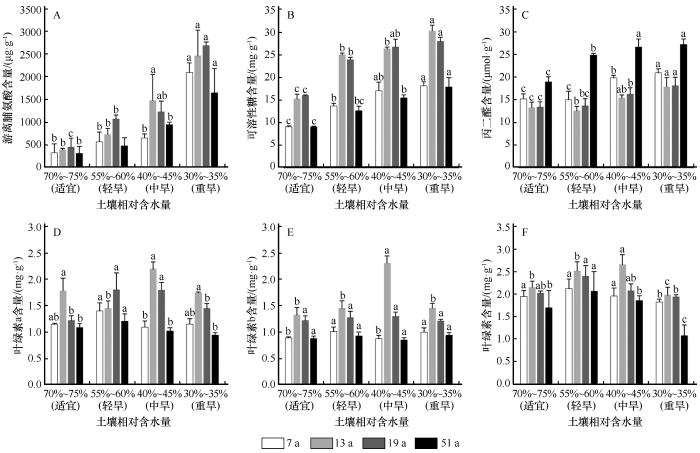
2.2 干旱胁迫对不同年限柠条幼苗生理代谢的影响
不同年限柠条幼苗叶片渗透调节物质在干旱胁迫下差异显著(P <0.05,图2 )。游离脯氨酸含量随着干旱程度的加剧呈上升趋势。轻度干旱下,19 a幼苗含量明显增加,其他年限幼苗游离脯氨酸含量增长缓慢;不同年限柠条幼苗均在重度胁迫时达到最大值,其中19 a最高,为2 682.73 μg·g-1 。峰值与对照相比,13 a游离脯氨酸含量上升幅度最多为524.01%,51 a增幅最小且积累量均低于其他年限。
图2
图2
干旱胁迫下不同年限柠条幼苗叶片的游离脯氨酸、可溶性糖、丙二醛、叶绿素a、叶绿素b和叶绿素含量的变化(不同字母代表同一年限不同干旱胁迫差异显著,P <0.05)
Fig.2
Changes of free proline, soluble sugar, malondialdehyde, chlorophyll a, chlorophyll b and chlorophyll in leaves of Caragana korshinskii seedlings in different ages under drought stress (Different letters indicate the same age plants has significant differences in different drought stress,P <0.05)
不同年限柠条幼苗可溶性糖含量在干旱胁迫下逐渐升高。在轻度胁迫时,各年限柠条幼苗可溶性糖含量均有小幅上升;在重度干旱时,13 a可溶性糖含量与对照相比增幅最大,为98.82%;19 a可溶性糖含量在中度胁迫下最高,而51 a含量在各处理下值最低。7 a在中度和重度胁迫下差异不显著。
不同年限柠条幼苗叶片丙二醛含量在干旱胁迫下差异显著(P <0.05)。当幼苗遭受干旱胁迫时,丙二醛含量表现出先减少后增加的趋势。轻度干旱下丙二醛含量最小(51 a除外),分别为14.10、12.57、13.64 μmol·g-1 ;在中度干旱时增幅较为缓慢,重度胁迫时,丙二醛含量快速积累,明显高于对照。51 a幼苗丙二醛含量均高于其他年限。
2.3 干旱胁迫对不同年限柠条幼苗光合特性的影响
不同年限柠条幼苗叶片光合生理指标在干旱胁迫下差异显著(P <0.05)。随着胁迫程度的增加,叶绿素a、b含量及叶绿素含量总体呈先上升后下降的趋势。在干旱胁迫下,7 a叶片叶绿素a、b含量呈现升—降—升的趋势;13 a叶绿素a含量在中度胁迫下达到最大,为2.18 mg·g-1 ,随后降低;19 a叶绿素a含量在轻度和中度胁迫时最高,后逐渐减少;51 a叶绿素a含量最低(图2 )。在中度干旱下,13 a叶片叶绿素b含量最高,与对照相比增加了74.31%;19 a和51 a叶绿素b含量差异不显著。在轻度和中度干旱下,13 a和19 a叶绿素含量均高于对照组,分别为2.40、2.07 mg·g-1 ;在重度胁迫下,不同年限幼苗叶片叶绿素含量降至最低,与对照相比分别下降了54.27%、41.07%、45.39%和56.69%。
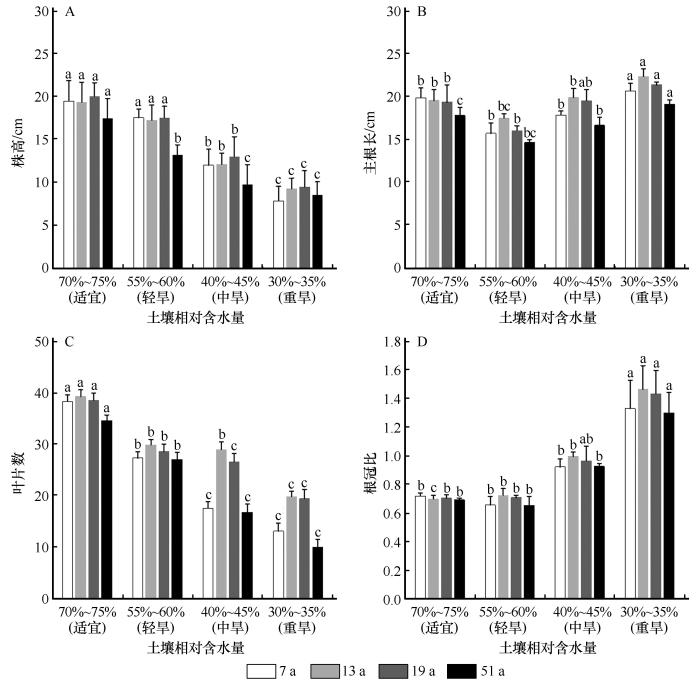
2.4 干旱胁迫对不同年限柠条幼苗生长状况的影响
2.4.1 干旱胁迫对不同年限柠条幼苗株高、主根长和叶片数的影响
干旱胁迫下,不同年限柠条幼苗株高差异显著(P <0.05),均随着干旱胁迫程度的加剧呈下降趋势。在重度干旱时,株高分别减少了59.69%、51.97%、52.75%和51.34%(图3 )。幼苗主根长在轻度和中度干旱下与对照相比差异不显著,在重度干旱下,根长较长,较对照分别增长了4.15%、14.39%、10.57%和7.308%。随着干旱程度的增加,不同年限幼苗单株叶片数量逐渐降低,在重度干旱下各年限单株叶片数目最少,与对照相比分别减少了65.68%、49.45%、49.48%和71.10%;13 a幼苗叶片数均高于其他年限。
图3
图3
干旱胁迫下不同年限柠条幼苗的株高、主根长、叶片数和根冠比的变化(不同字母代表同一年限不同干旱胁迫差异显著,P <0.05)
Fig.3
Changes of plant height, main root length, number of leaves and root-shoot ratio of Caragana korshinskii seedlings in different ages under drought stress (Different letters indicate the same age plants has significant differences in different drought stress,P <0.05)
2.4.2 干旱胁迫对不同年限柠条幼苗生物量的积累及分配的影响
干旱胁迫下,不同年限柠条幼苗地上生物量、地下生物量和总生物量积累差异显著(P <0.05),随着干旱胁迫程度的增加,各部分生物量的积累均呈下降趋势。在重度胁迫下,各年限柠条幼苗生物量的积累量降至最低,与对照相比分别减少了77.57%、74.89%、75.34%和80.00%;13 a幼苗生物量积累均高于其他年限,51 a的生物量积累最少(表2 )。
根冠比是植物地上、地下生物量分配策略的体现,由图3 可以看出,干旱胁迫下不同年限柠条幼苗的根冠比均随干旱程度的增加呈上升趋势,13 a幼苗根冠比在重度干旱下达到最大,较对照增加了104.29%;19 a增长幅度次之,较对照增加了98.61%;51 a柠条幼苗根冠比相较于其他年限最低。
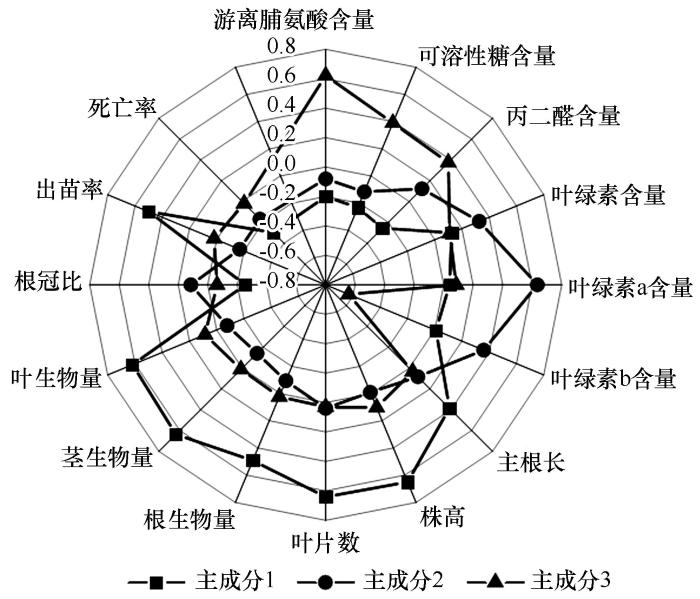
2.5 不同年限柠条幼苗抗旱性主成分分析
为避免各指标间因相关性而造成的信息重叠,利用多元方法对不同年限柠条幼苗的抗旱性进行科学评价与分析[35 ] 。通过主成分分析,以累计方差贡献率大于80%且特征值≥1作为判断条件[36 ] ,将干旱胁迫下15个鉴定指标转换成3个主成分,作为综合指标(comprehensive index,Cl )。同一指标特征向量的最大绝对值所在主成分即为所属主成分[37 ] 。第1主成分中株高系数最大,其次是叶片数、茎生物量和叶生物量,大致概括为生长状况因子,解释59.323%的贡献率;第2主成分中叶绿素a系数最大,其次是叶绿素b和叶绿素含量,可概括为光合因子,解释14.260%的贡献率;第3主成分中游离脯氨酸系数最大,其次是可溶性糖和丙二醛,可概括为生理代谢因子,解释9.433%的贡献率。因此,生长状况因子可概括为反映幼苗抗旱性能的重要指标,其次是光合因子和生理代谢因子(表3 和图4 )。
图4
图4
所测15个相关指标主成分特征向量
Fig.4
Principal component eigenvectors with the 15 measured indexes
2.6 不同年限柠条幼苗抗旱性隶属函数分析
采用单一指标评价植物的抗旱性具有片面性,隶属函数提供了在多指标测定基础上对植物进行综合评价的途径,可以克服利用少数指标对植物抗旱性进行评价的不足[33 ] 。不同年限柠条幼苗的各项隶属函数的平均值分别为0.443、0.469、0.450和0.428,抗旱能力13 a>19 a>7 a>51 a(表4 )。
3 讨论
3.1 干旱胁迫对不同年限柠条种子出苗率及死亡率的影响
在植物实生更新过程中,种子经散布、萌发,到形成完全独立的幼苗,是其有性繁殖更新过程得以实现的关键环节[38 ] 。幼苗阶段是生长最为脆弱、对环境变化最为敏感的时期,也是个体数量变化最大的时期,因此也成为植物种群生活史中亏损的主要阶段[39 ] 。本研究表明,干旱胁迫对不同年限柠条种子出苗率有明显的抑制作用,出苗率均随干旱胁迫的加剧逐渐下降。7、13 a和19 a柠条种子出苗率在土壤极度缺水的情况下能够达到36%、37.16%和34.50%,而51 a出苗率最低为19.56%;随着土壤水分含量降至最低时,51 a幼苗死亡率最高,可达80.17%。这可能是由于质量较大的种子具有较为充实的储存物质,从而能够提供更多的营养物质[40 ] ,使其具有更大的萌发能力。这与曾彦军等[41 ] 的研究结果一致,说明土壤水分的极度缺乏使得干旱半干旱地区植物的有性生殖在落种萌发时受到抑制。
3.2 干旱胁迫对不同年限柠条幼苗生理指标的影响
苗期是植物对水分胁迫最为敏感的阶段,此时遭受干旱会对后续的生长发育造成不良影响,甚至会导致死亡[42 ] 。植物受到干旱胁迫时,叶片内均会产生大量的渗透调节物质,通过产生有机质溶质来维持渗透势以保持细胞继续吸水,使叶片细胞组织具有一定的持水力或使其免于脱水,从而对胁迫起到缓冲保护作用[43 ] 。本研究表明,13 a和19 a叶片在轻度和中度胁迫下可溶性糖及游离脯氨酸含量均有增加,对环境胁迫的抵抗发挥着重要作用。相关分析表明,与各年限柠条幼苗生长呈显著相关的生理指标多集中于渗透调节物质和叶绿素,且大多呈极显著相关关系(P <0.01,表5 )。这说明在干旱的影响下,渗透调节物质对维持叶片水分平衡具有重要影响,而叶绿素是保证植物正常光合作用的重要指标,二者共同对植物的生长起着关键作用。
丙二醛是膜脂过氧化的主要产物,对细胞具有毒性,当植物受到胁迫时,细胞内氧自由基会大量积累,使膜脂脂肪酸中的不饱和键被过氧化形成丙二醛,这是造成细胞膜损伤、导致细胞死亡的重要原因[44 ] 。本研究表明,各年限幼苗丙二醛含量随干旱程度的增加均呈现逐渐增加的趋势,但在轻度和中度干旱下,13 a和19 a幼苗丙二醛含量虽有增加,但增加幅度不大;重度干旱下丙二醛含量增加较为显著,具有持续、快速累加的效应,说明极度的缺水使幼苗受到较重的伤害。因此在重度干旱下,51 a幼苗丙二醛明显升高,细胞膜损坏严重,导致死亡率增加。
植物遭受水分胁迫达到一定程度时,光合作用便完全或部分受到抑制,影响叶绿体的结构和活性[45 ] ,叶绿素a相较于叶绿素b来说对水分胁迫更为敏感,因而更加容易被分解破坏[46 ] 。本研究表明,13 a和19 a幼苗在轻度和中度干旱胁迫下叶绿素a、b以及叶绿素含量较高,此时正处于渗透调节物质的高值区,可通过维持气孔开放,以保证光合作用的正常进行,保障植物的生长;在重度胁迫处理下,叶片叶绿素含量降低,这可能是由于幼苗叶片水分含量减少的原因。这与裴保华等[47 ] 对柠条耐旱性研究结果相似。
3.3 干旱胁迫对不同年限柠条幼苗生长指标及生物量指标的影响
植物是一个功能平衡体,各功能单位的大小与整个植株是相互协调的,地上部和地下部所占的生物量是有一定比例的,比例失调会对植物的正常生长不利[48 ] 。本研究表明,在各处理初期,不同年限柠条幼苗在外部形态上表现差异不明显;在中度和重度胁迫处理的后半期,幼苗叶片变黄、卷曲发干等。本研究表明,不同年限柠条幼苗株高和叶片数均随干旱程度的加剧呈减少的趋势,根长则有所增加,尤其是13 a和19 a幼苗增幅较大,这可能是由于此时游离脯氨酸含量较高,能够促进幼苗根系生长。同时,在干旱程度加剧的情况下,各年限幼苗根冠比显著增大,表明生物量分配向根系转移,可见各功能器官对于干旱胁迫响应的敏感性不同,幼苗将更多的生物量用于根的生长发育。这与牛存洋等[49 ] 对小叶锦鸡儿的研究结果相似。由此可见,随着干旱胁迫的加剧,幼苗将相对多的光合产物用于构建营养器官(根系),以加强自身的竞争能力来维持幼苗自身的水分和其他物质的平衡。
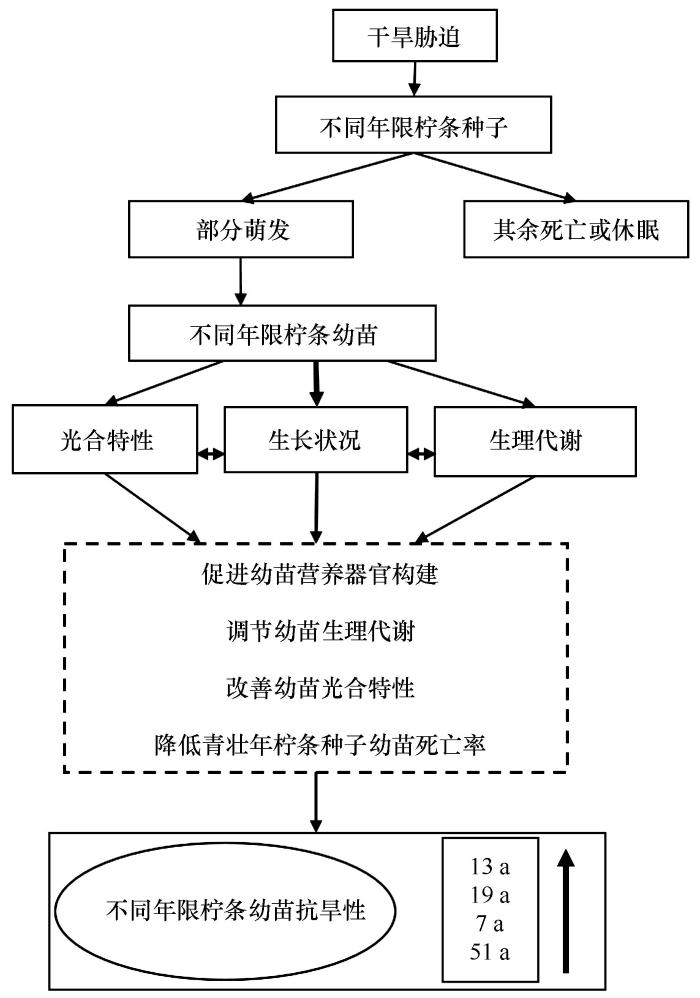
3.4 干旱胁迫对不同年限柠条种子天然更新过程可能的影响机制
本研究表明,干旱以及老龄柠条种子是造成人工柠条林天然更新困难的障碍因子,当人工柠条林达到老龄阶段时,其种子萌发及幼苗抗旱性逐渐减弱。本研究进一步推测了干旱对不同年限柠条种子天然更新过程的影响(图5 ),即干旱环境信号作用下,幼苗以营养生长为主,通常还会出现一些明显可见的表型特征,如叶片萎蔫、发干,株高、叶片数下降等。青壮年(13 a和19 a)柠条种子幼苗在轻度和中度胁迫下以地上部分为主,在重度干旱下加大对地下营养器官的构建。通过调节生理代谢、改善光合特性,降低幼苗死亡率,保障幼苗存活率,最终达到耐旱的目的。
图5
图5
干旱胁迫对不同年限柠条种子天然更新过程可能的影响机制
Fig.5
Possible mechanism of drought stress on the natural regeneration process of Caragana korshinskii seeds with different ages
4 结论
不同年限柠条种子抗旱性存在一定的差异,13 a和19 a柠条种子在重度干旱条件下仍有较高的出苗率,而51 a出苗率则最低;干旱对不同年限柠条幼苗的影响主要通过生长状况表现出来,幼苗在轻度和中度干旱下主要以地上部分进行光合作用促进生长,13 a和19 a幼苗在重度胁迫下,相较于地上部分而言,主要增加根系投入,从而有效促进根系吸收水分;在生理代谢方面,13 a和19 a幼苗能够积累大量游离脯氨酸和可溶性糖来维持叶片渗透压,以避免细胞脱水,使得叶绿素在轻度、中度干旱下轻微增加,同时将丙二醛维持在较低水平,使幼苗维持正常体内生长环境。综合生长因子、生理代谢以及光合因子的互相影响,使青壮年(13 a和19 a)柠条幼苗减少死亡率,保障幼苗生长发育。因此,在促进人工柠条林种子更新时,创造有利于青壮年柠条种子天然更新的生境,在林龄较大的衰退林分,选用青壮年柠条种子进行人工补播。
参考文献
View Option
[1]
李新荣 ,张志山 ,黄磊 ,等 我国沙区人工植被系统生态-水文过程和互馈机理研究评述
[J].科学通报 ,2013 ,58 ():397 -410 .
[本文引用: 2]
[2]
朱震达 ,刘恕 中国的荒漠化及其治理 [M].北京 :科学出版社 ,1989 :27 -42 .
[本文引用: 1]
[3]
Wang G Innes J L Lei J et al China's forestry reforms
[J].Science ,2007 ,318 (5856 ):1556 -1557 .
[本文引用: 1]
[4]
郭惠清 内蒙中部地区小老树成因及改造途径的研究
[J].干旱区资源与环境 ,1997 (4 ):73 -80 .
[本文引用: 1]
[5]
鲍婧婷 ,王进 ,苏洁琼 不同林龄柠条(Caragana korshinskii )的光合特性和水分利用特征
[J].中国沙漠 ,2016 ,36 (1 ):199 -205 .
[本文引用: 1]
[6]
史建伟 ,王孟本 ,陈建文 ,等 柠条细根的空间分布特征及其季节动态
[J].生态学报 ,2011 ,31 (3 ):726 -733 .
[本文引用: 1]
[7]
于露 ,周玉蓉 ,赵亚楠 ,等 荒漠草原土壤种子库对灌丛引入和降水梯度的响应特征
[J].草业学报 ,2020 ,29 (4 ):41 -50 .
[本文引用: 1]
[8]
Wu Y P Hu X W Wang Y R Growth, water relations, and stomatal development of Caragana korshinskii Kom. and Zygophyllum xanthoxylum (Bunge) Maxim. seedlings in response to water deficits
[J].New Zealand Journal of Agricultural Research ,2009 ,52 (2 ):185 -193 .
[本文引用: 1]
[9]
姚华 ,赵晓英 ,李晓梅 ,等 四种灌木幼苗对水分胁迫的生理响应
[J].生态科学 ,2009 ,28 (6 ):537 -542 .
[本文引用: 1]
[10]
徐当会 ,方向文 ,宾振钧 ,等 柠条适应极端干旱的生理生态机制:叶片脱落和枝条中叶绿体保持完整性
[J].中国沙漠 ,2012 ,32 (3 ):691 -697 .
[本文引用: 1]
[11]
杨明秀 ,宋乃平 ,杨新国 人工柠条林枝、叶构件生物量的分配格局与估测模型
[J].江苏农业科学 ,2013 ,41 (12 ):331 -333 .
[本文引用: 1]
[12]
马占英 平茬对黄土高原人工柠条林土壤碳输入的影响
[D].陕西杨凌 :西北农林科技大学 ,2020 .
[本文引用: 1]
[13]
牛西午 ,丁玉川 ,张强 ,等 柠条根系发育特征及有关生理特性研究
[J].西北植物学报 ,2003 (5 ):860 -865 .
[本文引用: 1]
[14]
张帆 ,陈建文 ,王孟本 幼龄柠条细根的空间分布和季节动态
[J].生态学报 ,2012 ,32 (17 ):5484 -5493 .
[本文引用: 1]
[15]
Binkley D Smith F W Son Y Nutrient supply and declines in leaf area and production in lodgepoie pine
[J].Canadian Journal of Forest Research ,1995 ,25 (4 ):621 -628 .
[本文引用: 1]
[16]
Schoettle A W Influence of tree size on shoot structure and physiology of Pinus contorta and Pinus aristata
[J].Tree Physiology ,1994 (7 ):1055 -1068 .
[本文引用: 1]
[17]
韩锦涛 ,李素清 ,赵德怀 ,等 晋西北丘陵风沙区人工植物群落优势种种间关系研究
[J].干旱区资源与环境 ,2016 ,30 (12 ):164 -169 .
[本文引用: 1]
[18]
王晗生 干旱影响下人工林的天然更新进程
[J].干旱区研究 ,2012 ,29 (5 ):743 -750 .
[本文引用: 1]
[19]
朱教君 ,李智辉 ,康宏樟 ,等 聚乙二醇模拟水分胁迫对沙地樟子松种子萌发影响研究
[J].应用生态学报 ,2005 ,16 (5 ):801 -804 .
[本文引用: 1]
[20]
Walck J L The ecology of seeds
[J].Seed Science Research ,2005 ,15 (4 ):365 -366 .
[本文引用: 1]
[21]
保长虎 ,张文辉 ,何景峰 黄土高原丘陵沟壑区30年柠条人工种群动态研究
[J].西北植物学报 ,2010 ,30 (8 ):1681 -1688 .
[本文引用: 1]
[22]
郑明清 ,郑元润 ,姜联合 毛乌素沙地4种沙生植物种子萌发及出苗对沙埋及单次供水的响应
[J].生态学报 ,2006 ,26 (8 ):2474 -2484 .
[本文引用: 1]
[23]
李茜 ,任运涛 ,牛得草 ,等 几种旱生灌木种子萌发特性及化学成分
[J].中国沙漠 ,2015 ,35 (2 ):345 -351 .
[本文引用: 1]
[24]
保长虎 黄土高原丘陵沟壑区柠条人工种群繁殖特征及天然化发育
[D].陕西杨凌 :西北农林科技大学 ,2011 .
[本文引用: 1]
[25]
杨体强 ,朱海英 ,华宏旭 ,等 电场对柠条(Caragana korshinskii )种子萌发和幼苗抗旱性的影响
[J].中国沙漠 ,2013 ,33 (6 ):1717 -1722 .
[本文引用: 1]
[26]
孙毅 宁夏白芨滩自然保护区柠条种群实生更新研究
[D].银川 :北方民族大学 ,2016 .
[本文引用: 1]
[27]
王孟本 ,荀俊杰 ,陈建文 ,等 晋西北黄土区幼龄柠条细根的净生长速率
[J].生态学报 ,2010 ,30 (5 ):1117 -1124 .
[本文引用: 1]
[28]
王国华 ,宋冰 ,席璐璐 ,等 晋西北丘陵风沙区不同林龄人工柠条生长与繁殖动态特征
[J].应用生态学报 ,2021 ,32 (6 ):2079 -2088 .
[本文引用: 1]
[29]
刘丙霞 ,任健 ,邵明安 ,等 黄土高原北部人工灌草植被土壤干燥化过程研究
[J].生态学报 ,2020 ,40 (11 ):3795 -3803 .
[本文引用: 1]
[30]
梁海斌 ,史建伟 ,牛俊杰 ,等 晋西北黄土丘陵区不同林龄柠条地土壤水分变化特征研究
[J].干旱区资源与环境 ,2014 ,28 (6 ):143 -148 .
[本文引用: 1]
[31]
Hsiao T C Plant responses to water stress
[J].Annual Review of Plant Physiology ,1973 ,24 :519 -570 .
[本文引用: 1]
[32]
陈春晓 ,谢秀华 ,王宇鹏 ,等 盐分和干旱对沙枣幼苗生理特性的影响
[J].生态学报 ,2019 ,39 (12 ):4540 -4550 .
[本文引用: 1]
[33]
张志良 ,瞿伟菁 植物生理学实验指导 [M].北京 :高等教育出版社 ,2009 :51 -106 .
[本文引用: 5]
[34]
郭郁频 ,米福贵 ,闫利军 ,等 不同早熟禾品种对干旱胁迫的生理响应及抗旱性评价
[J].草业学报 ,2014 ,23 (4 ):220 -228 .
[本文引用: 1]
[35]
赵小强 ,陆晏天 ,白明兴 ,等 不同株型玉米基因型对干旱胁迫的响应分析
[J].草业学报 ,2020 ,29 (2 ):149 -162 .
[本文引用: 1]
[36]
蔺豆豆 ,赵桂琴 ,琚泽亮 ,等 15份燕麦材料苗期抗旱性综合评价
[J].草业学报 ,2021 ,30 (11 ):108 -121 .
[本文引用: 1]
[37]
仝倩 ,施明 ,贺建勋 ,等 5种葡萄砧木耐旱性评价及鉴定指标的筛选
[J].核农学报 ,2018 ,32 (9 ):1814 -1820 .
[本文引用: 1]
[38]
Walck J L Hidayati S N Dixon K W et al Climate change and plant regeneration from seed
[J].Global Change Biology ,2011 ,17 (6 ):2145 -2161 .
[本文引用: 1]
[39]
刘从 ,田甜 ,李珊 ,等 中国木本植物幼苗生长对光照强度的响应
[J].生态学报 ,2018 ,38 (2 ):518 -527 .
[本文引用: 1]
[40]
傅家瑞 种子的活力及其生理生化基础
[J].种子 ,1984 (3 ):3 -8 .
[本文引用: 1]
[41]
曾彦军 ,王彦荣 ,萨仁 ,等 几种旱生灌木种子萌发对干旱胁迫的响应
[J].应用生态学报 ,2002 ,13 (8 ):953 -956 .
[本文引用: 1]
[42]
汤章城 植物干旱生态生理的研究
[J].生态学报 ,1983 ,3 (3 ):14 -22 .
[本文引用: 1]
[43]
Ashraf M Foolad M R Roles of glycine betaine and proline in improving plant abiotic stress resistance
[J].Environmental and Experimental Botany ,2007 ,59 (2 ):206 -216 .
[本文引用: 1]
[44]
李磊 ,贾志清 ,朱雅娟 ,等 我国干旱区植物抗旱机理研究进展
[J].中国沙漠 ,2010 ,30 (5 ):1053 -1059 .
[本文引用: 1]
[45]
云建英 ,杨甲定 ,赵哈林 干旱和高温对植物光合作用的影响机制研究进展
[J].西北植物学报 ,2006 (3 ):641 -648 .
[本文引用: 1]
[46]
张明生 ,谈锋 水分胁迫下甘薯叶绿素a/b比值的变化及其与抗旱性的关系
[J].种子 ,2001 (4 ):23 -25 .
[本文引用: 1]
[47]
裴保华 ,周宝顺 三种灌木耐旱性研究
[J].林业科学研究 ,1993 (6 ):597 -602 .
[本文引用: 1]
[48]
薛海霞 ,李清河 ,徐军 ,等 沙埋对唐古特白刺幼苗生长和生物量分配的影响
[J].草业科学 ,2016 ,33 (10 ):2062 -2070 .
[本文引用: 1]
[49]
牛存洋 ,阿拉木萨 ,宗芹 ,等 科尔沁沙地小叶锦鸡儿地上-地下生物量分配格局
[J].生态学杂志 ,2013 ,32 (8 ):1980 -1986 .
[本文引用: 1]
我国沙区人工植被系统生态-水文过程和互馈机理研究评述
2
2013
... 沙漠与荒漠草原过渡区是中国沙漠化与风沙活动危害最严重的区域,横跨中国北方地区且分布于极端干旱、干旱、半干旱等不同气候带[1 -2 ] .为防止沙化土地进一步扩张,国家先后在风沙区启动了“三北”防护林、退耕还林和京津风沙源治理等以人工植被建设为主的一批重大生态建设工程[3 ] .目前,中国人工植被面积已达6 933万hm2 ,占全国林地面积的36%[1 ] .但在风沙治理区,大面积人工植被产生了退化、老化以及更新困难等问题[4 ] .引起固沙人工林生长衰退的因素有很多,如固沙树种选择、种植密度、林分结构以及环境条件等[5 ] .因此,了解并掌握人工林天然更新所需条件对其实践经营具有重要意义. ...
... [1 ].但在风沙治理区,大面积人工植被产生了退化、老化以及更新困难等问题[4 ] .引起固沙人工林生长衰退的因素有很多,如固沙树种选择、种植密度、林分结构以及环境条件等[5 ] .因此,了解并掌握人工林天然更新所需条件对其实践经营具有重要意义. ...
1
1989
... 沙漠与荒漠草原过渡区是中国沙漠化与风沙活动危害最严重的区域,横跨中国北方地区且分布于极端干旱、干旱、半干旱等不同气候带[1 -2 ] .为防止沙化土地进一步扩张,国家先后在风沙区启动了“三北”防护林、退耕还林和京津风沙源治理等以人工植被建设为主的一批重大生态建设工程[3 ] .目前,中国人工植被面积已达6 933万hm2 ,占全国林地面积的36%[1 ] .但在风沙治理区,大面积人工植被产生了退化、老化以及更新困难等问题[4 ] .引起固沙人工林生长衰退的因素有很多,如固沙树种选择、种植密度、林分结构以及环境条件等[5 ] .因此,了解并掌握人工林天然更新所需条件对其实践经营具有重要意义. ...
China's forestry reforms
1
2007
... 沙漠与荒漠草原过渡区是中国沙漠化与风沙活动危害最严重的区域,横跨中国北方地区且分布于极端干旱、干旱、半干旱等不同气候带[1 -2 ] .为防止沙化土地进一步扩张,国家先后在风沙区启动了“三北”防护林、退耕还林和京津风沙源治理等以人工植被建设为主的一批重大生态建设工程[3 ] .目前,中国人工植被面积已达6 933万hm2 ,占全国林地面积的36%[1 ] .但在风沙治理区,大面积人工植被产生了退化、老化以及更新困难等问题[4 ] .引起固沙人工林生长衰退的因素有很多,如固沙树种选择、种植密度、林分结构以及环境条件等[5 ] .因此,了解并掌握人工林天然更新所需条件对其实践经营具有重要意义. ...
内蒙中部地区小老树成因及改造途径的研究
1
1997
... 沙漠与荒漠草原过渡区是中国沙漠化与风沙活动危害最严重的区域,横跨中国北方地区且分布于极端干旱、干旱、半干旱等不同气候带[1 -2 ] .为防止沙化土地进一步扩张,国家先后在风沙区启动了“三北”防护林、退耕还林和京津风沙源治理等以人工植被建设为主的一批重大生态建设工程[3 ] .目前,中国人工植被面积已达6 933万hm2 ,占全国林地面积的36%[1 ] .但在风沙治理区,大面积人工植被产生了退化、老化以及更新困难等问题[4 ] .引起固沙人工林生长衰退的因素有很多,如固沙树种选择、种植密度、林分结构以及环境条件等[5 ] .因此,了解并掌握人工林天然更新所需条件对其实践经营具有重要意义. ...
不同林龄柠条(Caragana korshinskii )的光合特性和水分利用特征
1
2016
... 沙漠与荒漠草原过渡区是中国沙漠化与风沙活动危害最严重的区域,横跨中国北方地区且分布于极端干旱、干旱、半干旱等不同气候带[1 -2 ] .为防止沙化土地进一步扩张,国家先后在风沙区启动了“三北”防护林、退耕还林和京津风沙源治理等以人工植被建设为主的一批重大生态建设工程[3 ] .目前,中国人工植被面积已达6 933万hm2 ,占全国林地面积的36%[1 ] .但在风沙治理区,大面积人工植被产生了退化、老化以及更新困难等问题[4 ] .引起固沙人工林生长衰退的因素有很多,如固沙树种选择、种植密度、林分结构以及环境条件等[5 ] .因此,了解并掌握人工林天然更新所需条件对其实践经营具有重要意义. ...
柠条细根的空间分布特征及其季节动态
1
2011
... 柠条(Caragana korshinskill )是中国干旱半干旱地区的主要固沙植物,具有耐旱、抗风沙、根系发达等特征,在保持水土以及改善生态环境、缓解北方沙尘暴灾害等方面起到了积极有效的作用[6 ] ,作为主要固沙树种受到极大关注.目前,国内外关于干旱对柠条锦鸡儿属植物在萌发、生长、繁殖及生理代谢等方面的影响已有大量报道.在宁夏荒漠草原地区研究发现,柠条种子萌发率和萌发速率随着干旱的加剧呈先上升后下降的趋势[7 ] ;在阿拉善地区有研究表明,中度干旱使得柠条叶生物量、地上生物量和总叶片数目显著降低,但根冠比有所增加[8 ] ;在西北干旱风沙区有研究发现,干旱的加剧可使柠条叶片中游离脯氨酸和可溶性糖含量的积累量均持续上升[9 ] ,且严重干旱对其叶片叶绿体结构造成不可恢复的破坏[10 ] .关于不同年限柠条的研究集中在生物量分配[11 -12 ] 以及根系特征的变化[13 -14 ] 等.植物营养可利用能力随林龄的增加而降低[15 ] ,且植物叶片的光合能力均低于幼龄[16 ] .然而,目前对长时间尺度下人工柠条林地干旱导致人工柠条林天然更新困难的研究还较少. ...
荒漠草原土壤种子库对灌丛引入和降水梯度的响应特征
1
2020
... 柠条(Caragana korshinskill )是中国干旱半干旱地区的主要固沙植物,具有耐旱、抗风沙、根系发达等特征,在保持水土以及改善生态环境、缓解北方沙尘暴灾害等方面起到了积极有效的作用[6 ] ,作为主要固沙树种受到极大关注.目前,国内外关于干旱对柠条锦鸡儿属植物在萌发、生长、繁殖及生理代谢等方面的影响已有大量报道.在宁夏荒漠草原地区研究发现,柠条种子萌发率和萌发速率随着干旱的加剧呈先上升后下降的趋势[7 ] ;在阿拉善地区有研究表明,中度干旱使得柠条叶生物量、地上生物量和总叶片数目显著降低,但根冠比有所增加[8 ] ;在西北干旱风沙区有研究发现,干旱的加剧可使柠条叶片中游离脯氨酸和可溶性糖含量的积累量均持续上升[9 ] ,且严重干旱对其叶片叶绿体结构造成不可恢复的破坏[10 ] .关于不同年限柠条的研究集中在生物量分配[11 -12 ] 以及根系特征的变化[13 -14 ] 等.植物营养可利用能力随林龄的增加而降低[15 ] ,且植物叶片的光合能力均低于幼龄[16 ] .然而,目前对长时间尺度下人工柠条林地干旱导致人工柠条林天然更新困难的研究还较少. ...
Growth, water relations, and stomatal development of Caragana korshinskii Kom. and Zygophyllum xanthoxylum (Bunge) Maxim. seedlings in response to water deficits
1
2009
... 柠条(Caragana korshinskill )是中国干旱半干旱地区的主要固沙植物,具有耐旱、抗风沙、根系发达等特征,在保持水土以及改善生态环境、缓解北方沙尘暴灾害等方面起到了积极有效的作用[6 ] ,作为主要固沙树种受到极大关注.目前,国内外关于干旱对柠条锦鸡儿属植物在萌发、生长、繁殖及生理代谢等方面的影响已有大量报道.在宁夏荒漠草原地区研究发现,柠条种子萌发率和萌发速率随着干旱的加剧呈先上升后下降的趋势[7 ] ;在阿拉善地区有研究表明,中度干旱使得柠条叶生物量、地上生物量和总叶片数目显著降低,但根冠比有所增加[8 ] ;在西北干旱风沙区有研究发现,干旱的加剧可使柠条叶片中游离脯氨酸和可溶性糖含量的积累量均持续上升[9 ] ,且严重干旱对其叶片叶绿体结构造成不可恢复的破坏[10 ] .关于不同年限柠条的研究集中在生物量分配[11 -12 ] 以及根系特征的变化[13 -14 ] 等.植物营养可利用能力随林龄的增加而降低[15 ] ,且植物叶片的光合能力均低于幼龄[16 ] .然而,目前对长时间尺度下人工柠条林地干旱导致人工柠条林天然更新困难的研究还较少. ...
四种灌木幼苗对水分胁迫的生理响应
1
2009
... 柠条(Caragana korshinskill )是中国干旱半干旱地区的主要固沙植物,具有耐旱、抗风沙、根系发达等特征,在保持水土以及改善生态环境、缓解北方沙尘暴灾害等方面起到了积极有效的作用[6 ] ,作为主要固沙树种受到极大关注.目前,国内外关于干旱对柠条锦鸡儿属植物在萌发、生长、繁殖及生理代谢等方面的影响已有大量报道.在宁夏荒漠草原地区研究发现,柠条种子萌发率和萌发速率随着干旱的加剧呈先上升后下降的趋势[7 ] ;在阿拉善地区有研究表明,中度干旱使得柠条叶生物量、地上生物量和总叶片数目显著降低,但根冠比有所增加[8 ] ;在西北干旱风沙区有研究发现,干旱的加剧可使柠条叶片中游离脯氨酸和可溶性糖含量的积累量均持续上升[9 ] ,且严重干旱对其叶片叶绿体结构造成不可恢复的破坏[10 ] .关于不同年限柠条的研究集中在生物量分配[11 -12 ] 以及根系特征的变化[13 -14 ] 等.植物营养可利用能力随林龄的增加而降低[15 ] ,且植物叶片的光合能力均低于幼龄[16 ] .然而,目前对长时间尺度下人工柠条林地干旱导致人工柠条林天然更新困难的研究还较少. ...
柠条适应极端干旱的生理生态机制:叶片脱落和枝条中叶绿体保持完整性
1
2012
... 柠条(Caragana korshinskill )是中国干旱半干旱地区的主要固沙植物,具有耐旱、抗风沙、根系发达等特征,在保持水土以及改善生态环境、缓解北方沙尘暴灾害等方面起到了积极有效的作用[6 ] ,作为主要固沙树种受到极大关注.目前,国内外关于干旱对柠条锦鸡儿属植物在萌发、生长、繁殖及生理代谢等方面的影响已有大量报道.在宁夏荒漠草原地区研究发现,柠条种子萌发率和萌发速率随着干旱的加剧呈先上升后下降的趋势[7 ] ;在阿拉善地区有研究表明,中度干旱使得柠条叶生物量、地上生物量和总叶片数目显著降低,但根冠比有所增加[8 ] ;在西北干旱风沙区有研究发现,干旱的加剧可使柠条叶片中游离脯氨酸和可溶性糖含量的积累量均持续上升[9 ] ,且严重干旱对其叶片叶绿体结构造成不可恢复的破坏[10 ] .关于不同年限柠条的研究集中在生物量分配[11 -12 ] 以及根系特征的变化[13 -14 ] 等.植物营养可利用能力随林龄的增加而降低[15 ] ,且植物叶片的光合能力均低于幼龄[16 ] .然而,目前对长时间尺度下人工柠条林地干旱导致人工柠条林天然更新困难的研究还较少. ...
人工柠条林枝、叶构件生物量的分配格局与估测模型
1
2013
... 柠条(Caragana korshinskill )是中国干旱半干旱地区的主要固沙植物,具有耐旱、抗风沙、根系发达等特征,在保持水土以及改善生态环境、缓解北方沙尘暴灾害等方面起到了积极有效的作用[6 ] ,作为主要固沙树种受到极大关注.目前,国内外关于干旱对柠条锦鸡儿属植物在萌发、生长、繁殖及生理代谢等方面的影响已有大量报道.在宁夏荒漠草原地区研究发现,柠条种子萌发率和萌发速率随着干旱的加剧呈先上升后下降的趋势[7 ] ;在阿拉善地区有研究表明,中度干旱使得柠条叶生物量、地上生物量和总叶片数目显著降低,但根冠比有所增加[8 ] ;在西北干旱风沙区有研究发现,干旱的加剧可使柠条叶片中游离脯氨酸和可溶性糖含量的积累量均持续上升[9 ] ,且严重干旱对其叶片叶绿体结构造成不可恢复的破坏[10 ] .关于不同年限柠条的研究集中在生物量分配[11 -12 ] 以及根系特征的变化[13 -14 ] 等.植物营养可利用能力随林龄的增加而降低[15 ] ,且植物叶片的光合能力均低于幼龄[16 ] .然而,目前对长时间尺度下人工柠条林地干旱导致人工柠条林天然更新困难的研究还较少. ...
平茬对黄土高原人工柠条林土壤碳输入的影响
1
2020
... 柠条(Caragana korshinskill )是中国干旱半干旱地区的主要固沙植物,具有耐旱、抗风沙、根系发达等特征,在保持水土以及改善生态环境、缓解北方沙尘暴灾害等方面起到了积极有效的作用[6 ] ,作为主要固沙树种受到极大关注.目前,国内外关于干旱对柠条锦鸡儿属植物在萌发、生长、繁殖及生理代谢等方面的影响已有大量报道.在宁夏荒漠草原地区研究发现,柠条种子萌发率和萌发速率随着干旱的加剧呈先上升后下降的趋势[7 ] ;在阿拉善地区有研究表明,中度干旱使得柠条叶生物量、地上生物量和总叶片数目显著降低,但根冠比有所增加[8 ] ;在西北干旱风沙区有研究发现,干旱的加剧可使柠条叶片中游离脯氨酸和可溶性糖含量的积累量均持续上升[9 ] ,且严重干旱对其叶片叶绿体结构造成不可恢复的破坏[10 ] .关于不同年限柠条的研究集中在生物量分配[11 -12 ] 以及根系特征的变化[13 -14 ] 等.植物营养可利用能力随林龄的增加而降低[15 ] ,且植物叶片的光合能力均低于幼龄[16 ] .然而,目前对长时间尺度下人工柠条林地干旱导致人工柠条林天然更新困难的研究还较少. ...
柠条根系发育特征及有关生理特性研究
1
2003
... 柠条(Caragana korshinskill )是中国干旱半干旱地区的主要固沙植物,具有耐旱、抗风沙、根系发达等特征,在保持水土以及改善生态环境、缓解北方沙尘暴灾害等方面起到了积极有效的作用[6 ] ,作为主要固沙树种受到极大关注.目前,国内外关于干旱对柠条锦鸡儿属植物在萌发、生长、繁殖及生理代谢等方面的影响已有大量报道.在宁夏荒漠草原地区研究发现,柠条种子萌发率和萌发速率随着干旱的加剧呈先上升后下降的趋势[7 ] ;在阿拉善地区有研究表明,中度干旱使得柠条叶生物量、地上生物量和总叶片数目显著降低,但根冠比有所增加[8 ] ;在西北干旱风沙区有研究发现,干旱的加剧可使柠条叶片中游离脯氨酸和可溶性糖含量的积累量均持续上升[9 ] ,且严重干旱对其叶片叶绿体结构造成不可恢复的破坏[10 ] .关于不同年限柠条的研究集中在生物量分配[11 -12 ] 以及根系特征的变化[13 -14 ] 等.植物营养可利用能力随林龄的增加而降低[15 ] ,且植物叶片的光合能力均低于幼龄[16 ] .然而,目前对长时间尺度下人工柠条林地干旱导致人工柠条林天然更新困难的研究还较少. ...
幼龄柠条细根的空间分布和季节动态
1
2012
... 柠条(Caragana korshinskill )是中国干旱半干旱地区的主要固沙植物,具有耐旱、抗风沙、根系发达等特征,在保持水土以及改善生态环境、缓解北方沙尘暴灾害等方面起到了积极有效的作用[6 ] ,作为主要固沙树种受到极大关注.目前,国内外关于干旱对柠条锦鸡儿属植物在萌发、生长、繁殖及生理代谢等方面的影响已有大量报道.在宁夏荒漠草原地区研究发现,柠条种子萌发率和萌发速率随着干旱的加剧呈先上升后下降的趋势[7 ] ;在阿拉善地区有研究表明,中度干旱使得柠条叶生物量、地上生物量和总叶片数目显著降低,但根冠比有所增加[8 ] ;在西北干旱风沙区有研究发现,干旱的加剧可使柠条叶片中游离脯氨酸和可溶性糖含量的积累量均持续上升[9 ] ,且严重干旱对其叶片叶绿体结构造成不可恢复的破坏[10 ] .关于不同年限柠条的研究集中在生物量分配[11 -12 ] 以及根系特征的变化[13 -14 ] 等.植物营养可利用能力随林龄的增加而降低[15 ] ,且植物叶片的光合能力均低于幼龄[16 ] .然而,目前对长时间尺度下人工柠条林地干旱导致人工柠条林天然更新困难的研究还较少. ...
Nutrient supply and declines in leaf area and production in lodgepoie pine
1
1995
... 柠条(Caragana korshinskill )是中国干旱半干旱地区的主要固沙植物,具有耐旱、抗风沙、根系发达等特征,在保持水土以及改善生态环境、缓解北方沙尘暴灾害等方面起到了积极有效的作用[6 ] ,作为主要固沙树种受到极大关注.目前,国内外关于干旱对柠条锦鸡儿属植物在萌发、生长、繁殖及生理代谢等方面的影响已有大量报道.在宁夏荒漠草原地区研究发现,柠条种子萌发率和萌发速率随着干旱的加剧呈先上升后下降的趋势[7 ] ;在阿拉善地区有研究表明,中度干旱使得柠条叶生物量、地上生物量和总叶片数目显著降低,但根冠比有所增加[8 ] ;在西北干旱风沙区有研究发现,干旱的加剧可使柠条叶片中游离脯氨酸和可溶性糖含量的积累量均持续上升[9 ] ,且严重干旱对其叶片叶绿体结构造成不可恢复的破坏[10 ] .关于不同年限柠条的研究集中在生物量分配[11 -12 ] 以及根系特征的变化[13 -14 ] 等.植物营养可利用能力随林龄的增加而降低[15 ] ,且植物叶片的光合能力均低于幼龄[16 ] .然而,目前对长时间尺度下人工柠条林地干旱导致人工柠条林天然更新困难的研究还较少. ...
Influence of tree size on shoot structure and physiology of Pinus contorta and Pinus aristata
1
1994
... 柠条(Caragana korshinskill )是中国干旱半干旱地区的主要固沙植物,具有耐旱、抗风沙、根系发达等特征,在保持水土以及改善生态环境、缓解北方沙尘暴灾害等方面起到了积极有效的作用[6 ] ,作为主要固沙树种受到极大关注.目前,国内外关于干旱对柠条锦鸡儿属植物在萌发、生长、繁殖及生理代谢等方面的影响已有大量报道.在宁夏荒漠草原地区研究发现,柠条种子萌发率和萌发速率随着干旱的加剧呈先上升后下降的趋势[7 ] ;在阿拉善地区有研究表明,中度干旱使得柠条叶生物量、地上生物量和总叶片数目显著降低,但根冠比有所增加[8 ] ;在西北干旱风沙区有研究发现,干旱的加剧可使柠条叶片中游离脯氨酸和可溶性糖含量的积累量均持续上升[9 ] ,且严重干旱对其叶片叶绿体结构造成不可恢复的破坏[10 ] .关于不同年限柠条的研究集中在生物量分配[11 -12 ] 以及根系特征的变化[13 -14 ] 等.植物营养可利用能力随林龄的增加而降低[15 ] ,且植物叶片的光合能力均低于幼龄[16 ] .然而,目前对长时间尺度下人工柠条林地干旱导致人工柠条林天然更新困难的研究还较少. ...
晋西北丘陵风沙区人工植物群落优势种种间关系研究
1
2016
... 晋西北丘陵风沙区地处中国沙漠化扩展前沿,土壤沙化严重,是干旱半干旱地区中典型的生态过渡带与环境脆弱区,也是京津能源、水源和沙源之地,该地区长期以来成为“三北”防护林建设的重点区域[17 ] .随着植被恢复工程的深入实施,柠条逐渐成为当地的优势种,并且形成了老龄、中龄和幼龄3个不同发育阶段,这使得人工柠条林稳定且持续性的发展受到了极大关注.在干旱半干旱地区,土壤水分在林木生长中起重要作用,同时也是治沙造林的主要限制因子[18 ] .一般认为,种子更新发生的必要条件是有充足且具备生命力的种子以及有利于种子萌发的环境和幼苗生长的气候条件[19 ] ,而更新的关键过程在于种子萌发与幼苗存活生长[20 ] .在天然条件下,柠条虽然可以成功结实并能够完成有性繁殖,但种子的萌发率和幼苗的存活率极低[21 ] .在毛乌素沙地柠条种子遇少量降雨能够萌发,但降雨之后长期的干旱会极度增大幼苗死亡的风险[22 ] ;在黄土高原丘陵沟壑区,由于水分限制,也极少有实生幼苗能够成活[23 -24 ] ;同时,种子萌发的好坏也会直接影响到幼苗的成活率[25 ] ,尤其是在干旱、半干旱的荒漠地区,适合种子萌发和低龄幼苗生长、存活的环境条件极其有限[26 ] ,虽然柠条根系较为发达,耐旱能力强,但低龄柠条幼苗也同样极易遭受干旱胁迫[27 ] .在晋西北丘陵风沙区,不同年限柠条均能顺利结实[28 ] ,但由于当地土壤干燥化发生频繁,使得人工柠条林地出现大面积退化现象,严重制约了当地植被建设的成效以及区域生态的稳定[29 ] .因此,无论是在干旱、半干旱的荒漠区还是晋西北丘陵风沙区,柠条种子更新都与水分有关.但是,不同种植年限柠条林种子更新早期过程在干旱胁迫下是否能够顺利进行却不十分清楚.因此,本研究采用盆栽控制水分模拟干旱的方法,以晋西北丘陵风沙区不同年限柠条种子为试验材料,分析干旱胁迫对不同年限柠条种子萌发以及幼苗生理、生长状况的影响,通过主成分分析和隶属函数法对其抗旱性进行综合评价,探讨各年限柠条幼苗渗透调节物质、膜脂过氧化物、光合指标、株高、主根长以及叶片数等指标与抗旱性的关系,明确在苗期抗旱性的关键影响因素,为晋西北丘陵风沙区人工柠条林的经营管理与可持续发展提供指导,保障其发挥生态防护功能. ...
干旱影响下人工林的天然更新进程
1
2012
... 晋西北丘陵风沙区地处中国沙漠化扩展前沿,土壤沙化严重,是干旱半干旱地区中典型的生态过渡带与环境脆弱区,也是京津能源、水源和沙源之地,该地区长期以来成为“三北”防护林建设的重点区域[17 ] .随着植被恢复工程的深入实施,柠条逐渐成为当地的优势种,并且形成了老龄、中龄和幼龄3个不同发育阶段,这使得人工柠条林稳定且持续性的发展受到了极大关注.在干旱半干旱地区,土壤水分在林木生长中起重要作用,同时也是治沙造林的主要限制因子[18 ] .一般认为,种子更新发生的必要条件是有充足且具备生命力的种子以及有利于种子萌发的环境和幼苗生长的气候条件[19 ] ,而更新的关键过程在于种子萌发与幼苗存活生长[20 ] .在天然条件下,柠条虽然可以成功结实并能够完成有性繁殖,但种子的萌发率和幼苗的存活率极低[21 ] .在毛乌素沙地柠条种子遇少量降雨能够萌发,但降雨之后长期的干旱会极度增大幼苗死亡的风险[22 ] ;在黄土高原丘陵沟壑区,由于水分限制,也极少有实生幼苗能够成活[23 -24 ] ;同时,种子萌发的好坏也会直接影响到幼苗的成活率[25 ] ,尤其是在干旱、半干旱的荒漠地区,适合种子萌发和低龄幼苗生长、存活的环境条件极其有限[26 ] ,虽然柠条根系较为发达,耐旱能力强,但低龄柠条幼苗也同样极易遭受干旱胁迫[27 ] .在晋西北丘陵风沙区,不同年限柠条均能顺利结实[28 ] ,但由于当地土壤干燥化发生频繁,使得人工柠条林地出现大面积退化现象,严重制约了当地植被建设的成效以及区域生态的稳定[29 ] .因此,无论是在干旱、半干旱的荒漠区还是晋西北丘陵风沙区,柠条种子更新都与水分有关.但是,不同种植年限柠条林种子更新早期过程在干旱胁迫下是否能够顺利进行却不十分清楚.因此,本研究采用盆栽控制水分模拟干旱的方法,以晋西北丘陵风沙区不同年限柠条种子为试验材料,分析干旱胁迫对不同年限柠条种子萌发以及幼苗生理、生长状况的影响,通过主成分分析和隶属函数法对其抗旱性进行综合评价,探讨各年限柠条幼苗渗透调节物质、膜脂过氧化物、光合指标、株高、主根长以及叶片数等指标与抗旱性的关系,明确在苗期抗旱性的关键影响因素,为晋西北丘陵风沙区人工柠条林的经营管理与可持续发展提供指导,保障其发挥生态防护功能. ...
聚乙二醇模拟水分胁迫对沙地樟子松种子萌发影响研究
1
2005
... 晋西北丘陵风沙区地处中国沙漠化扩展前沿,土壤沙化严重,是干旱半干旱地区中典型的生态过渡带与环境脆弱区,也是京津能源、水源和沙源之地,该地区长期以来成为“三北”防护林建设的重点区域[17 ] .随着植被恢复工程的深入实施,柠条逐渐成为当地的优势种,并且形成了老龄、中龄和幼龄3个不同发育阶段,这使得人工柠条林稳定且持续性的发展受到了极大关注.在干旱半干旱地区,土壤水分在林木生长中起重要作用,同时也是治沙造林的主要限制因子[18 ] .一般认为,种子更新发生的必要条件是有充足且具备生命力的种子以及有利于种子萌发的环境和幼苗生长的气候条件[19 ] ,而更新的关键过程在于种子萌发与幼苗存活生长[20 ] .在天然条件下,柠条虽然可以成功结实并能够完成有性繁殖,但种子的萌发率和幼苗的存活率极低[21 ] .在毛乌素沙地柠条种子遇少量降雨能够萌发,但降雨之后长期的干旱会极度增大幼苗死亡的风险[22 ] ;在黄土高原丘陵沟壑区,由于水分限制,也极少有实生幼苗能够成活[23 -24 ] ;同时,种子萌发的好坏也会直接影响到幼苗的成活率[25 ] ,尤其是在干旱、半干旱的荒漠地区,适合种子萌发和低龄幼苗生长、存活的环境条件极其有限[26 ] ,虽然柠条根系较为发达,耐旱能力强,但低龄柠条幼苗也同样极易遭受干旱胁迫[27 ] .在晋西北丘陵风沙区,不同年限柠条均能顺利结实[28 ] ,但由于当地土壤干燥化发生频繁,使得人工柠条林地出现大面积退化现象,严重制约了当地植被建设的成效以及区域生态的稳定[29 ] .因此,无论是在干旱、半干旱的荒漠区还是晋西北丘陵风沙区,柠条种子更新都与水分有关.但是,不同种植年限柠条林种子更新早期过程在干旱胁迫下是否能够顺利进行却不十分清楚.因此,本研究采用盆栽控制水分模拟干旱的方法,以晋西北丘陵风沙区不同年限柠条种子为试验材料,分析干旱胁迫对不同年限柠条种子萌发以及幼苗生理、生长状况的影响,通过主成分分析和隶属函数法对其抗旱性进行综合评价,探讨各年限柠条幼苗渗透调节物质、膜脂过氧化物、光合指标、株高、主根长以及叶片数等指标与抗旱性的关系,明确在苗期抗旱性的关键影响因素,为晋西北丘陵风沙区人工柠条林的经营管理与可持续发展提供指导,保障其发挥生态防护功能. ...
The ecology of seeds
1
2005
... 晋西北丘陵风沙区地处中国沙漠化扩展前沿,土壤沙化严重,是干旱半干旱地区中典型的生态过渡带与环境脆弱区,也是京津能源、水源和沙源之地,该地区长期以来成为“三北”防护林建设的重点区域[17 ] .随着植被恢复工程的深入实施,柠条逐渐成为当地的优势种,并且形成了老龄、中龄和幼龄3个不同发育阶段,这使得人工柠条林稳定且持续性的发展受到了极大关注.在干旱半干旱地区,土壤水分在林木生长中起重要作用,同时也是治沙造林的主要限制因子[18 ] .一般认为,种子更新发生的必要条件是有充足且具备生命力的种子以及有利于种子萌发的环境和幼苗生长的气候条件[19 ] ,而更新的关键过程在于种子萌发与幼苗存活生长[20 ] .在天然条件下,柠条虽然可以成功结实并能够完成有性繁殖,但种子的萌发率和幼苗的存活率极低[21 ] .在毛乌素沙地柠条种子遇少量降雨能够萌发,但降雨之后长期的干旱会极度增大幼苗死亡的风险[22 ] ;在黄土高原丘陵沟壑区,由于水分限制,也极少有实生幼苗能够成活[23 -24 ] ;同时,种子萌发的好坏也会直接影响到幼苗的成活率[25 ] ,尤其是在干旱、半干旱的荒漠地区,适合种子萌发和低龄幼苗生长、存活的环境条件极其有限[26 ] ,虽然柠条根系较为发达,耐旱能力强,但低龄柠条幼苗也同样极易遭受干旱胁迫[27 ] .在晋西北丘陵风沙区,不同年限柠条均能顺利结实[28 ] ,但由于当地土壤干燥化发生频繁,使得人工柠条林地出现大面积退化现象,严重制约了当地植被建设的成效以及区域生态的稳定[29 ] .因此,无论是在干旱、半干旱的荒漠区还是晋西北丘陵风沙区,柠条种子更新都与水分有关.但是,不同种植年限柠条林种子更新早期过程在干旱胁迫下是否能够顺利进行却不十分清楚.因此,本研究采用盆栽控制水分模拟干旱的方法,以晋西北丘陵风沙区不同年限柠条种子为试验材料,分析干旱胁迫对不同年限柠条种子萌发以及幼苗生理、生长状况的影响,通过主成分分析和隶属函数法对其抗旱性进行综合评价,探讨各年限柠条幼苗渗透调节物质、膜脂过氧化物、光合指标、株高、主根长以及叶片数等指标与抗旱性的关系,明确在苗期抗旱性的关键影响因素,为晋西北丘陵风沙区人工柠条林的经营管理与可持续发展提供指导,保障其发挥生态防护功能. ...
黄土高原丘陵沟壑区30年柠条人工种群动态研究
1
2010
... 晋西北丘陵风沙区地处中国沙漠化扩展前沿,土壤沙化严重,是干旱半干旱地区中典型的生态过渡带与环境脆弱区,也是京津能源、水源和沙源之地,该地区长期以来成为“三北”防护林建设的重点区域[17 ] .随着植被恢复工程的深入实施,柠条逐渐成为当地的优势种,并且形成了老龄、中龄和幼龄3个不同发育阶段,这使得人工柠条林稳定且持续性的发展受到了极大关注.在干旱半干旱地区,土壤水分在林木生长中起重要作用,同时也是治沙造林的主要限制因子[18 ] .一般认为,种子更新发生的必要条件是有充足且具备生命力的种子以及有利于种子萌发的环境和幼苗生长的气候条件[19 ] ,而更新的关键过程在于种子萌发与幼苗存活生长[20 ] .在天然条件下,柠条虽然可以成功结实并能够完成有性繁殖,但种子的萌发率和幼苗的存活率极低[21 ] .在毛乌素沙地柠条种子遇少量降雨能够萌发,但降雨之后长期的干旱会极度增大幼苗死亡的风险[22 ] ;在黄土高原丘陵沟壑区,由于水分限制,也极少有实生幼苗能够成活[23 -24 ] ;同时,种子萌发的好坏也会直接影响到幼苗的成活率[25 ] ,尤其是在干旱、半干旱的荒漠地区,适合种子萌发和低龄幼苗生长、存活的环境条件极其有限[26 ] ,虽然柠条根系较为发达,耐旱能力强,但低龄柠条幼苗也同样极易遭受干旱胁迫[27 ] .在晋西北丘陵风沙区,不同年限柠条均能顺利结实[28 ] ,但由于当地土壤干燥化发生频繁,使得人工柠条林地出现大面积退化现象,严重制约了当地植被建设的成效以及区域生态的稳定[29 ] .因此,无论是在干旱、半干旱的荒漠区还是晋西北丘陵风沙区,柠条种子更新都与水分有关.但是,不同种植年限柠条林种子更新早期过程在干旱胁迫下是否能够顺利进行却不十分清楚.因此,本研究采用盆栽控制水分模拟干旱的方法,以晋西北丘陵风沙区不同年限柠条种子为试验材料,分析干旱胁迫对不同年限柠条种子萌发以及幼苗生理、生长状况的影响,通过主成分分析和隶属函数法对其抗旱性进行综合评价,探讨各年限柠条幼苗渗透调节物质、膜脂过氧化物、光合指标、株高、主根长以及叶片数等指标与抗旱性的关系,明确在苗期抗旱性的关键影响因素,为晋西北丘陵风沙区人工柠条林的经营管理与可持续发展提供指导,保障其发挥生态防护功能. ...
毛乌素沙地4种沙生植物种子萌发及出苗对沙埋及单次供水的响应
1
2006
... 晋西北丘陵风沙区地处中国沙漠化扩展前沿,土壤沙化严重,是干旱半干旱地区中典型的生态过渡带与环境脆弱区,也是京津能源、水源和沙源之地,该地区长期以来成为“三北”防护林建设的重点区域[17 ] .随着植被恢复工程的深入实施,柠条逐渐成为当地的优势种,并且形成了老龄、中龄和幼龄3个不同发育阶段,这使得人工柠条林稳定且持续性的发展受到了极大关注.在干旱半干旱地区,土壤水分在林木生长中起重要作用,同时也是治沙造林的主要限制因子[18 ] .一般认为,种子更新发生的必要条件是有充足且具备生命力的种子以及有利于种子萌发的环境和幼苗生长的气候条件[19 ] ,而更新的关键过程在于种子萌发与幼苗存活生长[20 ] .在天然条件下,柠条虽然可以成功结实并能够完成有性繁殖,但种子的萌发率和幼苗的存活率极低[21 ] .在毛乌素沙地柠条种子遇少量降雨能够萌发,但降雨之后长期的干旱会极度增大幼苗死亡的风险[22 ] ;在黄土高原丘陵沟壑区,由于水分限制,也极少有实生幼苗能够成活[23 -24 ] ;同时,种子萌发的好坏也会直接影响到幼苗的成活率[25 ] ,尤其是在干旱、半干旱的荒漠地区,适合种子萌发和低龄幼苗生长、存活的环境条件极其有限[26 ] ,虽然柠条根系较为发达,耐旱能力强,但低龄柠条幼苗也同样极易遭受干旱胁迫[27 ] .在晋西北丘陵风沙区,不同年限柠条均能顺利结实[28 ] ,但由于当地土壤干燥化发生频繁,使得人工柠条林地出现大面积退化现象,严重制约了当地植被建设的成效以及区域生态的稳定[29 ] .因此,无论是在干旱、半干旱的荒漠区还是晋西北丘陵风沙区,柠条种子更新都与水分有关.但是,不同种植年限柠条林种子更新早期过程在干旱胁迫下是否能够顺利进行却不十分清楚.因此,本研究采用盆栽控制水分模拟干旱的方法,以晋西北丘陵风沙区不同年限柠条种子为试验材料,分析干旱胁迫对不同年限柠条种子萌发以及幼苗生理、生长状况的影响,通过主成分分析和隶属函数法对其抗旱性进行综合评价,探讨各年限柠条幼苗渗透调节物质、膜脂过氧化物、光合指标、株高、主根长以及叶片数等指标与抗旱性的关系,明确在苗期抗旱性的关键影响因素,为晋西北丘陵风沙区人工柠条林的经营管理与可持续发展提供指导,保障其发挥生态防护功能. ...
几种旱生灌木种子萌发特性及化学成分
1
2015
... 晋西北丘陵风沙区地处中国沙漠化扩展前沿,土壤沙化严重,是干旱半干旱地区中典型的生态过渡带与环境脆弱区,也是京津能源、水源和沙源之地,该地区长期以来成为“三北”防护林建设的重点区域[17 ] .随着植被恢复工程的深入实施,柠条逐渐成为当地的优势种,并且形成了老龄、中龄和幼龄3个不同发育阶段,这使得人工柠条林稳定且持续性的发展受到了极大关注.在干旱半干旱地区,土壤水分在林木生长中起重要作用,同时也是治沙造林的主要限制因子[18 ] .一般认为,种子更新发生的必要条件是有充足且具备生命力的种子以及有利于种子萌发的环境和幼苗生长的气候条件[19 ] ,而更新的关键过程在于种子萌发与幼苗存活生长[20 ] .在天然条件下,柠条虽然可以成功结实并能够完成有性繁殖,但种子的萌发率和幼苗的存活率极低[21 ] .在毛乌素沙地柠条种子遇少量降雨能够萌发,但降雨之后长期的干旱会极度增大幼苗死亡的风险[22 ] ;在黄土高原丘陵沟壑区,由于水分限制,也极少有实生幼苗能够成活[23 -24 ] ;同时,种子萌发的好坏也会直接影响到幼苗的成活率[25 ] ,尤其是在干旱、半干旱的荒漠地区,适合种子萌发和低龄幼苗生长、存活的环境条件极其有限[26 ] ,虽然柠条根系较为发达,耐旱能力强,但低龄柠条幼苗也同样极易遭受干旱胁迫[27 ] .在晋西北丘陵风沙区,不同年限柠条均能顺利结实[28 ] ,但由于当地土壤干燥化发生频繁,使得人工柠条林地出现大面积退化现象,严重制约了当地植被建设的成效以及区域生态的稳定[29 ] .因此,无论是在干旱、半干旱的荒漠区还是晋西北丘陵风沙区,柠条种子更新都与水分有关.但是,不同种植年限柠条林种子更新早期过程在干旱胁迫下是否能够顺利进行却不十分清楚.因此,本研究采用盆栽控制水分模拟干旱的方法,以晋西北丘陵风沙区不同年限柠条种子为试验材料,分析干旱胁迫对不同年限柠条种子萌发以及幼苗生理、生长状况的影响,通过主成分分析和隶属函数法对其抗旱性进行综合评价,探讨各年限柠条幼苗渗透调节物质、膜脂过氧化物、光合指标、株高、主根长以及叶片数等指标与抗旱性的关系,明确在苗期抗旱性的关键影响因素,为晋西北丘陵风沙区人工柠条林的经营管理与可持续发展提供指导,保障其发挥生态防护功能. ...
黄土高原丘陵沟壑区柠条人工种群繁殖特征及天然化发育
1
2011
... 晋西北丘陵风沙区地处中国沙漠化扩展前沿,土壤沙化严重,是干旱半干旱地区中典型的生态过渡带与环境脆弱区,也是京津能源、水源和沙源之地,该地区长期以来成为“三北”防护林建设的重点区域[17 ] .随着植被恢复工程的深入实施,柠条逐渐成为当地的优势种,并且形成了老龄、中龄和幼龄3个不同发育阶段,这使得人工柠条林稳定且持续性的发展受到了极大关注.在干旱半干旱地区,土壤水分在林木生长中起重要作用,同时也是治沙造林的主要限制因子[18 ] .一般认为,种子更新发生的必要条件是有充足且具备生命力的种子以及有利于种子萌发的环境和幼苗生长的气候条件[19 ] ,而更新的关键过程在于种子萌发与幼苗存活生长[20 ] .在天然条件下,柠条虽然可以成功结实并能够完成有性繁殖,但种子的萌发率和幼苗的存活率极低[21 ] .在毛乌素沙地柠条种子遇少量降雨能够萌发,但降雨之后长期的干旱会极度增大幼苗死亡的风险[22 ] ;在黄土高原丘陵沟壑区,由于水分限制,也极少有实生幼苗能够成活[23 -24 ] ;同时,种子萌发的好坏也会直接影响到幼苗的成活率[25 ] ,尤其是在干旱、半干旱的荒漠地区,适合种子萌发和低龄幼苗生长、存活的环境条件极其有限[26 ] ,虽然柠条根系较为发达,耐旱能力强,但低龄柠条幼苗也同样极易遭受干旱胁迫[27 ] .在晋西北丘陵风沙区,不同年限柠条均能顺利结实[28 ] ,但由于当地土壤干燥化发生频繁,使得人工柠条林地出现大面积退化现象,严重制约了当地植被建设的成效以及区域生态的稳定[29 ] .因此,无论是在干旱、半干旱的荒漠区还是晋西北丘陵风沙区,柠条种子更新都与水分有关.但是,不同种植年限柠条林种子更新早期过程在干旱胁迫下是否能够顺利进行却不十分清楚.因此,本研究采用盆栽控制水分模拟干旱的方法,以晋西北丘陵风沙区不同年限柠条种子为试验材料,分析干旱胁迫对不同年限柠条种子萌发以及幼苗生理、生长状况的影响,通过主成分分析和隶属函数法对其抗旱性进行综合评价,探讨各年限柠条幼苗渗透调节物质、膜脂过氧化物、光合指标、株高、主根长以及叶片数等指标与抗旱性的关系,明确在苗期抗旱性的关键影响因素,为晋西北丘陵风沙区人工柠条林的经营管理与可持续发展提供指导,保障其发挥生态防护功能. ...
电场对柠条(Caragana korshinskii )种子萌发和幼苗抗旱性的影响
1
2013
... 晋西北丘陵风沙区地处中国沙漠化扩展前沿,土壤沙化严重,是干旱半干旱地区中典型的生态过渡带与环境脆弱区,也是京津能源、水源和沙源之地,该地区长期以来成为“三北”防护林建设的重点区域[17 ] .随着植被恢复工程的深入实施,柠条逐渐成为当地的优势种,并且形成了老龄、中龄和幼龄3个不同发育阶段,这使得人工柠条林稳定且持续性的发展受到了极大关注.在干旱半干旱地区,土壤水分在林木生长中起重要作用,同时也是治沙造林的主要限制因子[18 ] .一般认为,种子更新发生的必要条件是有充足且具备生命力的种子以及有利于种子萌发的环境和幼苗生长的气候条件[19 ] ,而更新的关键过程在于种子萌发与幼苗存活生长[20 ] .在天然条件下,柠条虽然可以成功结实并能够完成有性繁殖,但种子的萌发率和幼苗的存活率极低[21 ] .在毛乌素沙地柠条种子遇少量降雨能够萌发,但降雨之后长期的干旱会极度增大幼苗死亡的风险[22 ] ;在黄土高原丘陵沟壑区,由于水分限制,也极少有实生幼苗能够成活[23 -24 ] ;同时,种子萌发的好坏也会直接影响到幼苗的成活率[25 ] ,尤其是在干旱、半干旱的荒漠地区,适合种子萌发和低龄幼苗生长、存活的环境条件极其有限[26 ] ,虽然柠条根系较为发达,耐旱能力强,但低龄柠条幼苗也同样极易遭受干旱胁迫[27 ] .在晋西北丘陵风沙区,不同年限柠条均能顺利结实[28 ] ,但由于当地土壤干燥化发生频繁,使得人工柠条林地出现大面积退化现象,严重制约了当地植被建设的成效以及区域生态的稳定[29 ] .因此,无论是在干旱、半干旱的荒漠区还是晋西北丘陵风沙区,柠条种子更新都与水分有关.但是,不同种植年限柠条林种子更新早期过程在干旱胁迫下是否能够顺利进行却不十分清楚.因此,本研究采用盆栽控制水分模拟干旱的方法,以晋西北丘陵风沙区不同年限柠条种子为试验材料,分析干旱胁迫对不同年限柠条种子萌发以及幼苗生理、生长状况的影响,通过主成分分析和隶属函数法对其抗旱性进行综合评价,探讨各年限柠条幼苗渗透调节物质、膜脂过氧化物、光合指标、株高、主根长以及叶片数等指标与抗旱性的关系,明确在苗期抗旱性的关键影响因素,为晋西北丘陵风沙区人工柠条林的经营管理与可持续发展提供指导,保障其发挥生态防护功能. ...
宁夏白芨滩自然保护区柠条种群实生更新研究
1
2016
... 晋西北丘陵风沙区地处中国沙漠化扩展前沿,土壤沙化严重,是干旱半干旱地区中典型的生态过渡带与环境脆弱区,也是京津能源、水源和沙源之地,该地区长期以来成为“三北”防护林建设的重点区域[17 ] .随着植被恢复工程的深入实施,柠条逐渐成为当地的优势种,并且形成了老龄、中龄和幼龄3个不同发育阶段,这使得人工柠条林稳定且持续性的发展受到了极大关注.在干旱半干旱地区,土壤水分在林木生长中起重要作用,同时也是治沙造林的主要限制因子[18 ] .一般认为,种子更新发生的必要条件是有充足且具备生命力的种子以及有利于种子萌发的环境和幼苗生长的气候条件[19 ] ,而更新的关键过程在于种子萌发与幼苗存活生长[20 ] .在天然条件下,柠条虽然可以成功结实并能够完成有性繁殖,但种子的萌发率和幼苗的存活率极低[21 ] .在毛乌素沙地柠条种子遇少量降雨能够萌发,但降雨之后长期的干旱会极度增大幼苗死亡的风险[22 ] ;在黄土高原丘陵沟壑区,由于水分限制,也极少有实生幼苗能够成活[23 -24 ] ;同时,种子萌发的好坏也会直接影响到幼苗的成活率[25 ] ,尤其是在干旱、半干旱的荒漠地区,适合种子萌发和低龄幼苗生长、存活的环境条件极其有限[26 ] ,虽然柠条根系较为发达,耐旱能力强,但低龄柠条幼苗也同样极易遭受干旱胁迫[27 ] .在晋西北丘陵风沙区,不同年限柠条均能顺利结实[28 ] ,但由于当地土壤干燥化发生频繁,使得人工柠条林地出现大面积退化现象,严重制约了当地植被建设的成效以及区域生态的稳定[29 ] .因此,无论是在干旱、半干旱的荒漠区还是晋西北丘陵风沙区,柠条种子更新都与水分有关.但是,不同种植年限柠条林种子更新早期过程在干旱胁迫下是否能够顺利进行却不十分清楚.因此,本研究采用盆栽控制水分模拟干旱的方法,以晋西北丘陵风沙区不同年限柠条种子为试验材料,分析干旱胁迫对不同年限柠条种子萌发以及幼苗生理、生长状况的影响,通过主成分分析和隶属函数法对其抗旱性进行综合评价,探讨各年限柠条幼苗渗透调节物质、膜脂过氧化物、光合指标、株高、主根长以及叶片数等指标与抗旱性的关系,明确在苗期抗旱性的关键影响因素,为晋西北丘陵风沙区人工柠条林的经营管理与可持续发展提供指导,保障其发挥生态防护功能. ...
晋西北黄土区幼龄柠条细根的净生长速率
1
2010
... 晋西北丘陵风沙区地处中国沙漠化扩展前沿,土壤沙化严重,是干旱半干旱地区中典型的生态过渡带与环境脆弱区,也是京津能源、水源和沙源之地,该地区长期以来成为“三北”防护林建设的重点区域[17 ] .随着植被恢复工程的深入实施,柠条逐渐成为当地的优势种,并且形成了老龄、中龄和幼龄3个不同发育阶段,这使得人工柠条林稳定且持续性的发展受到了极大关注.在干旱半干旱地区,土壤水分在林木生长中起重要作用,同时也是治沙造林的主要限制因子[18 ] .一般认为,种子更新发生的必要条件是有充足且具备生命力的种子以及有利于种子萌发的环境和幼苗生长的气候条件[19 ] ,而更新的关键过程在于种子萌发与幼苗存活生长[20 ] .在天然条件下,柠条虽然可以成功结实并能够完成有性繁殖,但种子的萌发率和幼苗的存活率极低[21 ] .在毛乌素沙地柠条种子遇少量降雨能够萌发,但降雨之后长期的干旱会极度增大幼苗死亡的风险[22 ] ;在黄土高原丘陵沟壑区,由于水分限制,也极少有实生幼苗能够成活[23 -24 ] ;同时,种子萌发的好坏也会直接影响到幼苗的成活率[25 ] ,尤其是在干旱、半干旱的荒漠地区,适合种子萌发和低龄幼苗生长、存活的环境条件极其有限[26 ] ,虽然柠条根系较为发达,耐旱能力强,但低龄柠条幼苗也同样极易遭受干旱胁迫[27 ] .在晋西北丘陵风沙区,不同年限柠条均能顺利结实[28 ] ,但由于当地土壤干燥化发生频繁,使得人工柠条林地出现大面积退化现象,严重制约了当地植被建设的成效以及区域生态的稳定[29 ] .因此,无论是在干旱、半干旱的荒漠区还是晋西北丘陵风沙区,柠条种子更新都与水分有关.但是,不同种植年限柠条林种子更新早期过程在干旱胁迫下是否能够顺利进行却不十分清楚.因此,本研究采用盆栽控制水分模拟干旱的方法,以晋西北丘陵风沙区不同年限柠条种子为试验材料,分析干旱胁迫对不同年限柠条种子萌发以及幼苗生理、生长状况的影响,通过主成分分析和隶属函数法对其抗旱性进行综合评价,探讨各年限柠条幼苗渗透调节物质、膜脂过氧化物、光合指标、株高、主根长以及叶片数等指标与抗旱性的关系,明确在苗期抗旱性的关键影响因素,为晋西北丘陵风沙区人工柠条林的经营管理与可持续发展提供指导,保障其发挥生态防护功能. ...
晋西北丘陵风沙区不同林龄人工柠条生长与繁殖动态特征
1
2021
... 晋西北丘陵风沙区地处中国沙漠化扩展前沿,土壤沙化严重,是干旱半干旱地区中典型的生态过渡带与环境脆弱区,也是京津能源、水源和沙源之地,该地区长期以来成为“三北”防护林建设的重点区域[17 ] .随着植被恢复工程的深入实施,柠条逐渐成为当地的优势种,并且形成了老龄、中龄和幼龄3个不同发育阶段,这使得人工柠条林稳定且持续性的发展受到了极大关注.在干旱半干旱地区,土壤水分在林木生长中起重要作用,同时也是治沙造林的主要限制因子[18 ] .一般认为,种子更新发生的必要条件是有充足且具备生命力的种子以及有利于种子萌发的环境和幼苗生长的气候条件[19 ] ,而更新的关键过程在于种子萌发与幼苗存活生长[20 ] .在天然条件下,柠条虽然可以成功结实并能够完成有性繁殖,但种子的萌发率和幼苗的存活率极低[21 ] .在毛乌素沙地柠条种子遇少量降雨能够萌发,但降雨之后长期的干旱会极度增大幼苗死亡的风险[22 ] ;在黄土高原丘陵沟壑区,由于水分限制,也极少有实生幼苗能够成活[23 -24 ] ;同时,种子萌发的好坏也会直接影响到幼苗的成活率[25 ] ,尤其是在干旱、半干旱的荒漠地区,适合种子萌发和低龄幼苗生长、存活的环境条件极其有限[26 ] ,虽然柠条根系较为发达,耐旱能力强,但低龄柠条幼苗也同样极易遭受干旱胁迫[27 ] .在晋西北丘陵风沙区,不同年限柠条均能顺利结实[28 ] ,但由于当地土壤干燥化发生频繁,使得人工柠条林地出现大面积退化现象,严重制约了当地植被建设的成效以及区域生态的稳定[29 ] .因此,无论是在干旱、半干旱的荒漠区还是晋西北丘陵风沙区,柠条种子更新都与水分有关.但是,不同种植年限柠条林种子更新早期过程在干旱胁迫下是否能够顺利进行却不十分清楚.因此,本研究采用盆栽控制水分模拟干旱的方法,以晋西北丘陵风沙区不同年限柠条种子为试验材料,分析干旱胁迫对不同年限柠条种子萌发以及幼苗生理、生长状况的影响,通过主成分分析和隶属函数法对其抗旱性进行综合评价,探讨各年限柠条幼苗渗透调节物质、膜脂过氧化物、光合指标、株高、主根长以及叶片数等指标与抗旱性的关系,明确在苗期抗旱性的关键影响因素,为晋西北丘陵风沙区人工柠条林的经营管理与可持续发展提供指导,保障其发挥生态防护功能. ...
黄土高原北部人工灌草植被土壤干燥化过程研究
1
2020
... 晋西北丘陵风沙区地处中国沙漠化扩展前沿,土壤沙化严重,是干旱半干旱地区中典型的生态过渡带与环境脆弱区,也是京津能源、水源和沙源之地,该地区长期以来成为“三北”防护林建设的重点区域[17 ] .随着植被恢复工程的深入实施,柠条逐渐成为当地的优势种,并且形成了老龄、中龄和幼龄3个不同发育阶段,这使得人工柠条林稳定且持续性的发展受到了极大关注.在干旱半干旱地区,土壤水分在林木生长中起重要作用,同时也是治沙造林的主要限制因子[18 ] .一般认为,种子更新发生的必要条件是有充足且具备生命力的种子以及有利于种子萌发的环境和幼苗生长的气候条件[19 ] ,而更新的关键过程在于种子萌发与幼苗存活生长[20 ] .在天然条件下,柠条虽然可以成功结实并能够完成有性繁殖,但种子的萌发率和幼苗的存活率极低[21 ] .在毛乌素沙地柠条种子遇少量降雨能够萌发,但降雨之后长期的干旱会极度增大幼苗死亡的风险[22 ] ;在黄土高原丘陵沟壑区,由于水分限制,也极少有实生幼苗能够成活[23 -24 ] ;同时,种子萌发的好坏也会直接影响到幼苗的成活率[25 ] ,尤其是在干旱、半干旱的荒漠地区,适合种子萌发和低龄幼苗生长、存活的环境条件极其有限[26 ] ,虽然柠条根系较为发达,耐旱能力强,但低龄柠条幼苗也同样极易遭受干旱胁迫[27 ] .在晋西北丘陵风沙区,不同年限柠条均能顺利结实[28 ] ,但由于当地土壤干燥化发生频繁,使得人工柠条林地出现大面积退化现象,严重制约了当地植被建设的成效以及区域生态的稳定[29 ] .因此,无论是在干旱、半干旱的荒漠区还是晋西北丘陵风沙区,柠条种子更新都与水分有关.但是,不同种植年限柠条林种子更新早期过程在干旱胁迫下是否能够顺利进行却不十分清楚.因此,本研究采用盆栽控制水分模拟干旱的方法,以晋西北丘陵风沙区不同年限柠条种子为试验材料,分析干旱胁迫对不同年限柠条种子萌发以及幼苗生理、生长状况的影响,通过主成分分析和隶属函数法对其抗旱性进行综合评价,探讨各年限柠条幼苗渗透调节物质、膜脂过氧化物、光合指标、株高、主根长以及叶片数等指标与抗旱性的关系,明确在苗期抗旱性的关键影响因素,为晋西北丘陵风沙区人工柠条林的经营管理与可持续发展提供指导,保障其发挥生态防护功能. ...
晋西北黄土丘陵区不同林龄柠条地土壤水分变化特征研究
1
2014
... 试验依据梁海斌等[30 ] 对晋西北丘陵风沙区土壤水分的研究,结合相关学者提出关于植物水分梯度划分的方法[31 ] ,设置适宜水分及轻度、中度和重度干旱胁迫,分别为土壤含水量占田间持水量的70%~75%、55%~60%、40%~45%和30%~35%. ...
Plant responses to water stress
1
1973
... 试验依据梁海斌等[30 ] 对晋西北丘陵风沙区土壤水分的研究,结合相关学者提出关于植物水分梯度划分的方法[31 ] ,设置适宜水分及轻度、中度和重度干旱胁迫,分别为土壤含水量占田间持水量的70%~75%、55%~60%、40%~45%和30%~35%. ...
盐分和干旱对沙枣幼苗生理特性的影响
1
2019
... 盆栽培植试验[32 ] 于2021年4月19日进行人工控制条件下的胁迫处理,将所采集的种子置于室内黑暗干燥处风干储存备用.花盆直径和深度均为15 cm,内装黄绵土,每个年限各处理3盆为3个重复.所用花盆用烧杯等量浇水,通过自然消耗至设定土壤含水量开始进行播种,选取籽粒饱满、大小基本一致且无病虫害的种子,不同年限的种子单独播种于花盆内,每盆播种20粒,行距和播深均为2 cm.为防止系统误差,将花盆随机摆放并用标签进行标记.在种子出苗及植株生长期间按不同水分梯度进行胁迫处理,用烘干和称重相结合的方法控制土壤含水量,每天18:00补充水分,使各处理土壤水分含量控制在设定范围内,为防止盆内水分蒸发,对花盆进行套袋处理.晴天自然光照,雨天遮雨棚覆盖,以防止天然降水的影响.每天记录出苗数,当幼苗数量达到最大后2周内再无幼苗出土时,视为萌发结束.在萌发结束3 d后开始干旱胁迫处理,胁迫持续时长为30 d,试验结束统一收取植株,将样品迅速带回实验室,放入4 ℃低温冰箱保存,测定各项指标. ...
5
2009
... 丙二醛含量的测定用硫代巴比妥酸(TBA)法[33 ] ;游离脯氨酸含量的测定用茚三酮显色法[33 ] ;叶绿素含量的测定用80%丙酮法[33 ] ;可溶性糖含量的测定用蒽酮比色法[33 ] . ...
... [33 ];叶绿素含量的测定用80%丙酮法[33 ] ;可溶性糖含量的测定用蒽酮比色法[33 ] . ...
... [33 ];可溶性糖含量的测定用蒽酮比色法[33 ] . ...
... [33 ]. ...
... 采用单一指标评价植物的抗旱性具有片面性,隶属函数提供了在多指标测定基础上对植物进行综合评价的途径,可以克服利用少数指标对植物抗旱性进行评价的不足[33 ] .不同年限柠条幼苗的各项隶属函数的平均值分别为0.443、0.469、0.450和0.428,抗旱能力13 a>19 a>7 a>51 a(表4 ). ...
不同早熟禾品种对干旱胁迫的生理响应及抗旱性评价
1
2014
... 运用式(7) 或式(8) 求出不同年限柠条种子在干旱胁迫下的隶属函数值,累加各指标的具体隶属值,并求出平均值后进行比较,平均值越大植物的抗旱性越强[34 ] . ...
不同株型玉米基因型对干旱胁迫的响应分析
1
2020
... 为避免各指标间因相关性而造成的信息重叠,利用多元方法对不同年限柠条幼苗的抗旱性进行科学评价与分析[35 ] .通过主成分分析,以累计方差贡献率大于80%且特征值≥1作为判断条件[36 ] ,将干旱胁迫下15个鉴定指标转换成3个主成分,作为综合指标(comprehensive index,Cl ).同一指标特征向量的最大绝对值所在主成分即为所属主成分[37 ] .第1主成分中株高系数最大,其次是叶片数、茎生物量和叶生物量,大致概括为生长状况因子,解释59.323%的贡献率;第2主成分中叶绿素a系数最大,其次是叶绿素b和叶绿素含量,可概括为光合因子,解释14.260%的贡献率;第3主成分中游离脯氨酸系数最大,其次是可溶性糖和丙二醛,可概括为生理代谢因子,解释9.433%的贡献率.因此,生长状况因子可概括为反映幼苗抗旱性能的重要指标,其次是光合因子和生理代谢因子(表3 和图4 ). ...
15份燕麦材料苗期抗旱性综合评价
1
2021
... 为避免各指标间因相关性而造成的信息重叠,利用多元方法对不同年限柠条幼苗的抗旱性进行科学评价与分析[35 ] .通过主成分分析,以累计方差贡献率大于80%且特征值≥1作为判断条件[36 ] ,将干旱胁迫下15个鉴定指标转换成3个主成分,作为综合指标(comprehensive index,Cl ).同一指标特征向量的最大绝对值所在主成分即为所属主成分[37 ] .第1主成分中株高系数最大,其次是叶片数、茎生物量和叶生物量,大致概括为生长状况因子,解释59.323%的贡献率;第2主成分中叶绿素a系数最大,其次是叶绿素b和叶绿素含量,可概括为光合因子,解释14.260%的贡献率;第3主成分中游离脯氨酸系数最大,其次是可溶性糖和丙二醛,可概括为生理代谢因子,解释9.433%的贡献率.因此,生长状况因子可概括为反映幼苗抗旱性能的重要指标,其次是光合因子和生理代谢因子(表3 和图4 ). ...
5种葡萄砧木耐旱性评价及鉴定指标的筛选
1
2018
... 为避免各指标间因相关性而造成的信息重叠,利用多元方法对不同年限柠条幼苗的抗旱性进行科学评价与分析[35 ] .通过主成分分析,以累计方差贡献率大于80%且特征值≥1作为判断条件[36 ] ,将干旱胁迫下15个鉴定指标转换成3个主成分,作为综合指标(comprehensive index,Cl ).同一指标特征向量的最大绝对值所在主成分即为所属主成分[37 ] .第1主成分中株高系数最大,其次是叶片数、茎生物量和叶生物量,大致概括为生长状况因子,解释59.323%的贡献率;第2主成分中叶绿素a系数最大,其次是叶绿素b和叶绿素含量,可概括为光合因子,解释14.260%的贡献率;第3主成分中游离脯氨酸系数最大,其次是可溶性糖和丙二醛,可概括为生理代谢因子,解释9.433%的贡献率.因此,生长状况因子可概括为反映幼苗抗旱性能的重要指标,其次是光合因子和生理代谢因子(表3 和图4 ). ...
Climate change and plant regeneration from seed
1
2011
... 在植物实生更新过程中,种子经散布、萌发,到形成完全独立的幼苗,是其有性繁殖更新过程得以实现的关键环节[38 ] .幼苗阶段是生长最为脆弱、对环境变化最为敏感的时期,也是个体数量变化最大的时期,因此也成为植物种群生活史中亏损的主要阶段[39 ] .本研究表明,干旱胁迫对不同年限柠条种子出苗率有明显的抑制作用,出苗率均随干旱胁迫的加剧逐渐下降.7、13 a和19 a柠条种子出苗率在土壤极度缺水的情况下能够达到36%、37.16%和34.50%,而51 a出苗率最低为19.56%;随着土壤水分含量降至最低时,51 a幼苗死亡率最高,可达80.17%.这可能是由于质量较大的种子具有较为充实的储存物质,从而能够提供更多的营养物质[40 ] ,使其具有更大的萌发能力.这与曾彦军等[41 ] 的研究结果一致,说明土壤水分的极度缺乏使得干旱半干旱地区植物的有性生殖在落种萌发时受到抑制. ...
中国木本植物幼苗生长对光照强度的响应
1
2018
... 在植物实生更新过程中,种子经散布、萌发,到形成完全独立的幼苗,是其有性繁殖更新过程得以实现的关键环节[38 ] .幼苗阶段是生长最为脆弱、对环境变化最为敏感的时期,也是个体数量变化最大的时期,因此也成为植物种群生活史中亏损的主要阶段[39 ] .本研究表明,干旱胁迫对不同年限柠条种子出苗率有明显的抑制作用,出苗率均随干旱胁迫的加剧逐渐下降.7、13 a和19 a柠条种子出苗率在土壤极度缺水的情况下能够达到36%、37.16%和34.50%,而51 a出苗率最低为19.56%;随着土壤水分含量降至最低时,51 a幼苗死亡率最高,可达80.17%.这可能是由于质量较大的种子具有较为充实的储存物质,从而能够提供更多的营养物质[40 ] ,使其具有更大的萌发能力.这与曾彦军等[41 ] 的研究结果一致,说明土壤水分的极度缺乏使得干旱半干旱地区植物的有性生殖在落种萌发时受到抑制. ...
种子的活力及其生理生化基础
1
1984
... 在植物实生更新过程中,种子经散布、萌发,到形成完全独立的幼苗,是其有性繁殖更新过程得以实现的关键环节[38 ] .幼苗阶段是生长最为脆弱、对环境变化最为敏感的时期,也是个体数量变化最大的时期,因此也成为植物种群生活史中亏损的主要阶段[39 ] .本研究表明,干旱胁迫对不同年限柠条种子出苗率有明显的抑制作用,出苗率均随干旱胁迫的加剧逐渐下降.7、13 a和19 a柠条种子出苗率在土壤极度缺水的情况下能够达到36%、37.16%和34.50%,而51 a出苗率最低为19.56%;随着土壤水分含量降至最低时,51 a幼苗死亡率最高,可达80.17%.这可能是由于质量较大的种子具有较为充实的储存物质,从而能够提供更多的营养物质[40 ] ,使其具有更大的萌发能力.这与曾彦军等[41 ] 的研究结果一致,说明土壤水分的极度缺乏使得干旱半干旱地区植物的有性生殖在落种萌发时受到抑制. ...
几种旱生灌木种子萌发对干旱胁迫的响应
1
2002
... 在植物实生更新过程中,种子经散布、萌发,到形成完全独立的幼苗,是其有性繁殖更新过程得以实现的关键环节[38 ] .幼苗阶段是生长最为脆弱、对环境变化最为敏感的时期,也是个体数量变化最大的时期,因此也成为植物种群生活史中亏损的主要阶段[39 ] .本研究表明,干旱胁迫对不同年限柠条种子出苗率有明显的抑制作用,出苗率均随干旱胁迫的加剧逐渐下降.7、13 a和19 a柠条种子出苗率在土壤极度缺水的情况下能够达到36%、37.16%和34.50%,而51 a出苗率最低为19.56%;随着土壤水分含量降至最低时,51 a幼苗死亡率最高,可达80.17%.这可能是由于质量较大的种子具有较为充实的储存物质,从而能够提供更多的营养物质[40 ] ,使其具有更大的萌发能力.这与曾彦军等[41 ] 的研究结果一致,说明土壤水分的极度缺乏使得干旱半干旱地区植物的有性生殖在落种萌发时受到抑制. ...
植物干旱生态生理的研究
1
1983
... 苗期是植物对水分胁迫最为敏感的阶段,此时遭受干旱会对后续的生长发育造成不良影响,甚至会导致死亡[42 ] .植物受到干旱胁迫时,叶片内均会产生大量的渗透调节物质,通过产生有机质溶质来维持渗透势以保持细胞继续吸水,使叶片细胞组织具有一定的持水力或使其免于脱水,从而对胁迫起到缓冲保护作用[43 ] .本研究表明,13 a和19 a叶片在轻度和中度胁迫下可溶性糖及游离脯氨酸含量均有增加,对环境胁迫的抵抗发挥着重要作用.相关分析表明,与各年限柠条幼苗生长呈显著相关的生理指标多集中于渗透调节物质和叶绿素,且大多呈极显著相关关系(P <0.01,表5 ).这说明在干旱的影响下,渗透调节物质对维持叶片水分平衡具有重要影响,而叶绿素是保证植物正常光合作用的重要指标,二者共同对植物的生长起着关键作用. ...
Roles of glycine betaine and proline in improving plant abiotic stress resistance
1
2007
... 苗期是植物对水分胁迫最为敏感的阶段,此时遭受干旱会对后续的生长发育造成不良影响,甚至会导致死亡[42 ] .植物受到干旱胁迫时,叶片内均会产生大量的渗透调节物质,通过产生有机质溶质来维持渗透势以保持细胞继续吸水,使叶片细胞组织具有一定的持水力或使其免于脱水,从而对胁迫起到缓冲保护作用[43 ] .本研究表明,13 a和19 a叶片在轻度和中度胁迫下可溶性糖及游离脯氨酸含量均有增加,对环境胁迫的抵抗发挥着重要作用.相关分析表明,与各年限柠条幼苗生长呈显著相关的生理指标多集中于渗透调节物质和叶绿素,且大多呈极显著相关关系(P <0.01,表5 ).这说明在干旱的影响下,渗透调节物质对维持叶片水分平衡具有重要影响,而叶绿素是保证植物正常光合作用的重要指标,二者共同对植物的生长起着关键作用. ...
我国干旱区植物抗旱机理研究进展
1
2010
... 丙二醛是膜脂过氧化的主要产物,对细胞具有毒性,当植物受到胁迫时,细胞内氧自由基会大量积累,使膜脂脂肪酸中的不饱和键被过氧化形成丙二醛,这是造成细胞膜损伤、导致细胞死亡的重要原因[44 ] .本研究表明,各年限幼苗丙二醛含量随干旱程度的增加均呈现逐渐增加的趋势,但在轻度和中度干旱下,13 a和19 a幼苗丙二醛含量虽有增加,但增加幅度不大;重度干旱下丙二醛含量增加较为显著,具有持续、快速累加的效应,说明极度的缺水使幼苗受到较重的伤害.因此在重度干旱下,51 a幼苗丙二醛明显升高,细胞膜损坏严重,导致死亡率增加. ...
干旱和高温对植物光合作用的影响机制研究进展
1
2006
... 植物遭受水分胁迫达到一定程度时,光合作用便完全或部分受到抑制,影响叶绿体的结构和活性[45 ] ,叶绿素a相较于叶绿素b来说对水分胁迫更为敏感,因而更加容易被分解破坏[46 ] .本研究表明,13 a和19 a幼苗在轻度和中度干旱胁迫下叶绿素a、b以及叶绿素含量较高,此时正处于渗透调节物质的高值区,可通过维持气孔开放,以保证光合作用的正常进行,保障植物的生长;在重度胁迫处理下,叶片叶绿素含量降低,这可能是由于幼苗叶片水分含量减少的原因.这与裴保华等[47 ] 对柠条耐旱性研究结果相似. ...
水分胁迫下甘薯叶绿素a/b比值的变化及其与抗旱性的关系
1
2001
... 植物遭受水分胁迫达到一定程度时,光合作用便完全或部分受到抑制,影响叶绿体的结构和活性[45 ] ,叶绿素a相较于叶绿素b来说对水分胁迫更为敏感,因而更加容易被分解破坏[46 ] .本研究表明,13 a和19 a幼苗在轻度和中度干旱胁迫下叶绿素a、b以及叶绿素含量较高,此时正处于渗透调节物质的高值区,可通过维持气孔开放,以保证光合作用的正常进行,保障植物的生长;在重度胁迫处理下,叶片叶绿素含量降低,这可能是由于幼苗叶片水分含量减少的原因.这与裴保华等[47 ] 对柠条耐旱性研究结果相似. ...
三种灌木耐旱性研究
1
1993
... 植物遭受水分胁迫达到一定程度时,光合作用便完全或部分受到抑制,影响叶绿体的结构和活性[45 ] ,叶绿素a相较于叶绿素b来说对水分胁迫更为敏感,因而更加容易被分解破坏[46 ] .本研究表明,13 a和19 a幼苗在轻度和中度干旱胁迫下叶绿素a、b以及叶绿素含量较高,此时正处于渗透调节物质的高值区,可通过维持气孔开放,以保证光合作用的正常进行,保障植物的生长;在重度胁迫处理下,叶片叶绿素含量降低,这可能是由于幼苗叶片水分含量减少的原因.这与裴保华等[47 ] 对柠条耐旱性研究结果相似. ...
沙埋对唐古特白刺幼苗生长和生物量分配的影响
1
2016
... 植物是一个功能平衡体,各功能单位的大小与整个植株是相互协调的,地上部和地下部所占的生物量是有一定比例的,比例失调会对植物的正常生长不利[48 ] .本研究表明,在各处理初期,不同年限柠条幼苗在外部形态上表现差异不明显;在中度和重度胁迫处理的后半期,幼苗叶片变黄、卷曲发干等.本研究表明,不同年限柠条幼苗株高和叶片数均随干旱程度的加剧呈减少的趋势,根长则有所增加,尤其是13 a和19 a幼苗增幅较大,这可能是由于此时游离脯氨酸含量较高,能够促进幼苗根系生长.同时,在干旱程度加剧的情况下,各年限幼苗根冠比显著增大,表明生物量分配向根系转移,可见各功能器官对于干旱胁迫响应的敏感性不同,幼苗将更多的生物量用于根的生长发育.这与牛存洋等[49 ] 对小叶锦鸡儿的研究结果相似.由此可见,随着干旱胁迫的加剧,幼苗将相对多的光合产物用于构建营养器官(根系),以加强自身的竞争能力来维持幼苗自身的水分和其他物质的平衡. ...
科尔沁沙地小叶锦鸡儿地上-地下生物量分配格局
1
2013
... 植物是一个功能平衡体,各功能单位的大小与整个植株是相互协调的,地上部和地下部所占的生物量是有一定比例的,比例失调会对植物的正常生长不利[48 ] .本研究表明,在各处理初期,不同年限柠条幼苗在外部形态上表现差异不明显;在中度和重度胁迫处理的后半期,幼苗叶片变黄、卷曲发干等.本研究表明,不同年限柠条幼苗株高和叶片数均随干旱程度的加剧呈减少的趋势,根长则有所增加,尤其是13 a和19 a幼苗增幅较大,这可能是由于此时游离脯氨酸含量较高,能够促进幼苗根系生长.同时,在干旱程度加剧的情况下,各年限幼苗根冠比显著增大,表明生物量分配向根系转移,可见各功能器官对于干旱胁迫响应的敏感性不同,幼苗将更多的生物量用于根的生长发育.这与牛存洋等[49 ] 对小叶锦鸡儿的研究结果相似.由此可见,随着干旱胁迫的加剧,幼苗将相对多的光合产物用于构建营养器官(根系),以加强自身的竞争能力来维持幼苗自身的水分和其他物质的平衡. ...




 甘公网安备 62010202000688号
甘公网安备 62010202000688号
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}