宁夏荒漠草原蒙古冰草种群小尺度空间格局研究
1
2019
... 荒漠草原是草原的物种多样性和潜在生产力最低的极限状态[1-2],生态环境脆弱,易受人类活动和气候变化的影响.蒙古冰草(Agropyron mongolicum var. mongolicum),又称沙芦草,禾本科冰草属多年生草本植物,广泛分布于中国西北、华北地区干草原和荒漠草原的沙质生境[3],是宁夏荒漠草原主要优势物种[4],在饲喂家畜、保护生态和遗传育种方面具有重要价值[5-7].在荒漠草原管理方面,蒙古冰草与其他种混播后可控制流沙移动,具有改善脆弱生态环境的作用,是改良荒漠草原和建设人工草地的重要草种[8-9]. ...
中国荒漠草原的研究态势与热点分析:基于文献计量研究
1
2021
... 荒漠草原是草原的物种多样性和潜在生产力最低的极限状态[1-2],生态环境脆弱,易受人类活动和气候变化的影响.蒙古冰草(Agropyron mongolicum var. mongolicum),又称沙芦草,禾本科冰草属多年生草本植物,广泛分布于中国西北、华北地区干草原和荒漠草原的沙质生境[3],是宁夏荒漠草原主要优势物种[4],在饲喂家畜、保护生态和遗传育种方面具有重要价值[5-7].在荒漠草原管理方面,蒙古冰草与其他种混播后可控制流沙移动,具有改善脆弱生态环境的作用,是改良荒漠草原和建设人工草地的重要草种[8-9]. ...
围封对荒漠草原沙芦草种群构件生物量分配特性的影响
3
2017
... 荒漠草原是草原的物种多样性和潜在生产力最低的极限状态[1-2],生态环境脆弱,易受人类活动和气候变化的影响.蒙古冰草(Agropyron mongolicum var. mongolicum),又称沙芦草,禾本科冰草属多年生草本植物,广泛分布于中国西北、华北地区干草原和荒漠草原的沙质生境[3],是宁夏荒漠草原主要优势物种[4],在饲喂家畜、保护生态和遗传育种方面具有重要价值[5-7].在荒漠草原管理方面,蒙古冰草与其他种混播后可控制流沙移动,具有改善脆弱生态环境的作用,是改良荒漠草原和建设人工草地的重要草种[8-9]. ...
... 植物在环境变化和胁迫因子下具有各自生长特征和物质分配规律[29].有研究表明干旱荒漠区草本植物的地上地下生物量分配遵从异速分配原则[30],一些荒漠植物生物量分配对土壤因子的响应也具有物种特异性[31].宁夏荒漠草原土壤类型空间异质性强,蒙古冰草广泛分布于多种土壤类型上[32],如风沙土、灰钙土等[33].目前,国内外关于蒙古冰草生物量分配对放牧方式、混播方式和CO2浓度变化等的响应已有报道[3,34-35],但很少有研究考虑土壤类型的影响.因此,生长在同一气候条件下、不同土壤类型的蒙古冰草是否符合最优分配理论和异速生长生物量分配理论还需要进一步的试验验证. ...
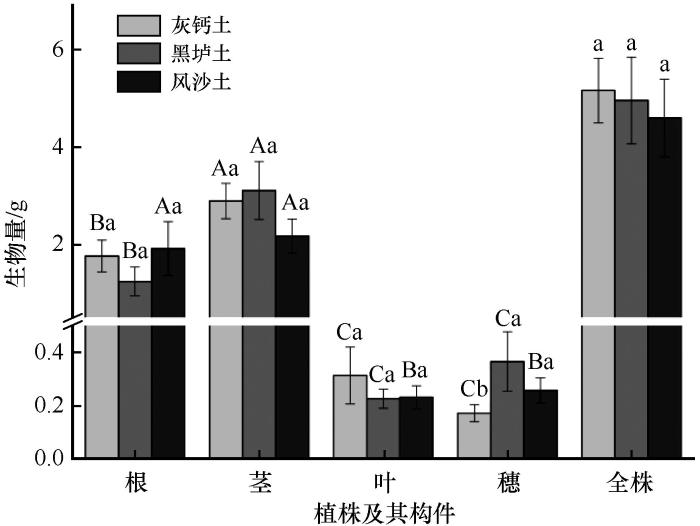
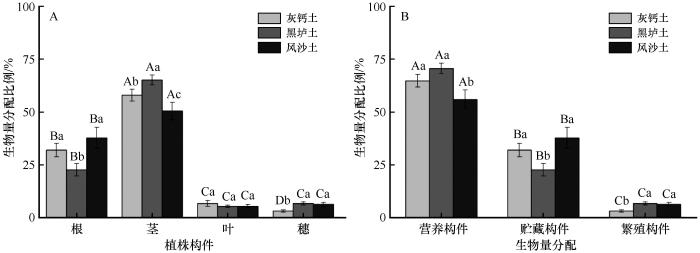
... 植物如何响应环境中资源可用性的变化是植物生态学的核心问题[46].植物构件生物量的分配反映了植物的发育规律和生存策略,影响植物个体的生长和植被结构,以及草地土壤碳输入和生态系统碳循环[11,47-49].有研究表明,土壤质地、碳氮比等因素对全球植物根系生物量分配比例起重要作用[50].在3种土壤类型条件下,蒙古冰草的构件生物量分配都表现出贮藏分配最多、营养分配次之、繁殖分配最少的规律,这与赵盼盼等[3]的研究结果一致.荒漠草原植物生长的主要限制性因子是水分和土壤养分[50],本控制试验中3种土壤类型具有相同的降水条件,尤其是在试验结束前经历了长达80 d的干旱,可以很好地探讨干旱条件下,不同土壤类型蒙古冰草是否符合最优分配理论.蒙古冰草根、茎、穗、营养分配、贮藏分配和繁殖分配比率在不同土壤类型间存在显著差异,各构件生物量比在不同土壤类型间也存在差异,且叶/根、叶/穗、茎/穗、叶/茎、根/茎、根/穗、繁殖构件生物量/营养构件生物量、贮藏构件生物量/营养构件生物量、繁殖构件生物量/贮藏构件生物量的排序在3种土壤类型中发生转变(表1).这表明蒙古冰草叶、茎、根、穗等构件以及繁殖、营养和贮藏构件的功能分配在不同土壤间存在权衡,蒙古冰草通过调整生物量分配策略以适应不同土壤类型,如在养分较好的灰钙土中以营养分配和贮藏分配为主,在黑垆土中则是减少贮藏分配而增加营养分配和繁殖分配,而在养分较差的风沙土中则是减少营养分配而增加贮藏分配和繁殖分配.另外,我们发现根、茎和穗生物量分配比例在不同类型土壤中一般存在显著差异,但叶生物量分配在不同类型土壤中无显著差异(图2).这表明蒙古冰草构件生物量分配主要受土壤水分和养分资源等限制. ...
小麦近缘植物沙芦草围封保护效果研究
1
2016
... 荒漠草原是草原的物种多样性和潜在生产力最低的极限状态[1-2],生态环境脆弱,易受人类活动和气候变化的影响.蒙古冰草(Agropyron mongolicum var. mongolicum),又称沙芦草,禾本科冰草属多年生草本植物,广泛分布于中国西北、华北地区干草原和荒漠草原的沙质生境[3],是宁夏荒漠草原主要优势物种[4],在饲喂家畜、保护生态和遗传育种方面具有重要价值[5-7].在荒漠草原管理方面,蒙古冰草与其他种混播后可控制流沙移动,具有改善脆弱生态环境的作用,是改良荒漠草原和建设人工草地的重要草种[8-9]. ...
Genetic diversity of prolamines in Agropyron mongolicum Keng indigenous to northern China
1
2007
... 荒漠草原是草原的物种多样性和潜在生产力最低的极限状态[1-2],生态环境脆弱,易受人类活动和气候变化的影响.蒙古冰草(Agropyron mongolicum var. mongolicum),又称沙芦草,禾本科冰草属多年生草本植物,广泛分布于中国西北、华北地区干草原和荒漠草原的沙质生境[3],是宁夏荒漠草原主要优势物种[4],在饲喂家畜、保护生态和遗传育种方面具有重要价值[5-7].在荒漠草原管理方面,蒙古冰草与其他种混播后可控制流沙移动,具有改善脆弱生态环境的作用,是改良荒漠草原和建设人工草地的重要草种[8-9]. ...
Reference genes for quantitative real-time PCR analysis and quantitative expression of P5CS in Agropyron mongolicum under drought stress
0
2016
宁夏荒漠草原沙芦草种群生长适应性研究
1
2019
... 荒漠草原是草原的物种多样性和潜在生产力最低的极限状态[1-2],生态环境脆弱,易受人类活动和气候变化的影响.蒙古冰草(Agropyron mongolicum var. mongolicum),又称沙芦草,禾本科冰草属多年生草本植物,广泛分布于中国西北、华北地区干草原和荒漠草原的沙质生境[3],是宁夏荒漠草原主要优势物种[4],在饲喂家畜、保护生态和遗传育种方面具有重要价值[5-7].在荒漠草原管理方面,蒙古冰草与其他种混播后可控制流沙移动,具有改善脆弱生态环境的作用,是改良荒漠草原和建设人工草地的重要草种[8-9]. ...
宁夏荒漠草原沙芦草种群结构及繁殖对策的研究
1
2013
... 荒漠草原是草原的物种多样性和潜在生产力最低的极限状态[1-2],生态环境脆弱,易受人类活动和气候变化的影响.蒙古冰草(Agropyron mongolicum var. mongolicum),又称沙芦草,禾本科冰草属多年生草本植物,广泛分布于中国西北、华北地区干草原和荒漠草原的沙质生境[3],是宁夏荒漠草原主要优势物种[4],在饲喂家畜、保护生态和遗传育种方面具有重要价值[5-7].在荒漠草原管理方面,蒙古冰草与其他种混播后可控制流沙移动,具有改善脆弱生态环境的作用,是改良荒漠草原和建设人工草地的重要草种[8-9]. ...
Functional analysis of three miRNAs in Agropyron mongolicum Keng under drought stress
1
2019
... 荒漠草原是草原的物种多样性和潜在生产力最低的极限状态[1-2],生态环境脆弱,易受人类活动和气候变化的影响.蒙古冰草(Agropyron mongolicum var. mongolicum),又称沙芦草,禾本科冰草属多年生草本植物,广泛分布于中国西北、华北地区干草原和荒漠草原的沙质生境[3],是宁夏荒漠草原主要优势物种[4],在饲喂家畜、保护生态和遗传育种方面具有重要价值[5-7].在荒漠草原管理方面,蒙古冰草与其他种混播后可控制流沙移动,具有改善脆弱生态环境的作用,是改良荒漠草原和建设人工草地的重要草种[8-9]. ...
岷江干旱河谷主要灌丛类型地上生物量研究
1
2003
... 自国际生物学计划(IBP)实行以来,植被生物量一直是生态学的重要研究方向[10].植株构件生物量能够表征植物个体及种群的生长发育节律,体现植物个体及种群适应环境的生存策略,其大小和变化反映了生态系统的物质流、能量流和重要功能[11-13].生物量分配则反映了植物对环境的适应能力和调节自身资源配置以应对特定环境的生长和生殖策略[14-15],是植物生命史和生态学理论的基础[16],对生态动力学和全球变化起重要作用[17].最优分配理论(Optimal Partitioning Theory)认为植物将生物量分配给获取最有限资源的器官,是叶、茎和根等构件功能分配的权衡[18-19],受气候、植物种类、土壤理化性质、土壤养分、土壤水分、光照和CO2浓度等因素的影响[17,19].如植物受到水分和养分的胁迫时会增加根系生物量的投入,而受到光胁迫时会增加冠层生物量的投入.最优分配理论虽然已得到多数研究的证实,但其普遍性仍受到质疑[20-21].如在考虑了植物大小时,种内的生物量分配存在变异,异速生长生物量分配理论(Allometric Biomass Partitioning Theory)有助于解释这一变异.异速生长关系能够揭示植物器官生长的内在规律,是生物量分配和器官功能关系的重要研究方式[22-25].一般表现为植物的某一器官生物量或总生物量与其他性状呈幂函数关系[22,26-28].如异速生长生物量分配理论认为,对于异速生长的理想植物,叶质量(Ml)与根质量(Mr)的关系为(Ml=1/βMr3/4),茎质量(Ms)和根质量(Mr)呈等比例关系[19].异速生长生物量分配理论是最优分配理论的有效补充,二者结合应用可以更好地揭示植物如何分配生物量以响应植物个体大小和环境的变化. ...
Seasonal not annual rainfall determines grassland biomass response to carbon dioxide
2
2014
... 自国际生物学计划(IBP)实行以来,植被生物量一直是生态学的重要研究方向[10].植株构件生物量能够表征植物个体及种群的生长发育节律,体现植物个体及种群适应环境的生存策略,其大小和变化反映了生态系统的物质流、能量流和重要功能[11-13].生物量分配则反映了植物对环境的适应能力和调节自身资源配置以应对特定环境的生长和生殖策略[14-15],是植物生命史和生态学理论的基础[16],对生态动力学和全球变化起重要作用[17].最优分配理论(Optimal Partitioning Theory)认为植物将生物量分配给获取最有限资源的器官,是叶、茎和根等构件功能分配的权衡[18-19],受气候、植物种类、土壤理化性质、土壤养分、土壤水分、光照和CO2浓度等因素的影响[17,19].如植物受到水分和养分的胁迫时会增加根系生物量的投入,而受到光胁迫时会增加冠层生物量的投入.最优分配理论虽然已得到多数研究的证实,但其普遍性仍受到质疑[20-21].如在考虑了植物大小时,种内的生物量分配存在变异,异速生长生物量分配理论(Allometric Biomass Partitioning Theory)有助于解释这一变异.异速生长关系能够揭示植物器官生长的内在规律,是生物量分配和器官功能关系的重要研究方式[22-25].一般表现为植物的某一器官生物量或总生物量与其他性状呈幂函数关系[22,26-28].如异速生长生物量分配理论认为,对于异速生长的理想植物,叶质量(Ml)与根质量(Mr)的关系为(Ml=1/βMr3/4),茎质量(Ms)和根质量(Mr)呈等比例关系[19].异速生长生物量分配理论是最优分配理论的有效补充,二者结合应用可以更好地揭示植物如何分配生物量以响应植物个体大小和环境的变化. ...
... 植物如何响应环境中资源可用性的变化是植物生态学的核心问题[46].植物构件生物量的分配反映了植物的发育规律和生存策略,影响植物个体的生长和植被结构,以及草地土壤碳输入和生态系统碳循环[11,47-49].有研究表明,土壤质地、碳氮比等因素对全球植物根系生物量分配比例起重要作用[50].在3种土壤类型条件下,蒙古冰草的构件生物量分配都表现出贮藏分配最多、营养分配次之、繁殖分配最少的规律,这与赵盼盼等[3]的研究结果一致.荒漠草原植物生长的主要限制性因子是水分和土壤养分[50],本控制试验中3种土壤类型具有相同的降水条件,尤其是在试验结束前经历了长达80 d的干旱,可以很好地探讨干旱条件下,不同土壤类型蒙古冰草是否符合最优分配理论.蒙古冰草根、茎、穗、营养分配、贮藏分配和繁殖分配比率在不同土壤类型间存在显著差异,各构件生物量比在不同土壤类型间也存在差异,且叶/根、叶/穗、茎/穗、叶/茎、根/茎、根/穗、繁殖构件生物量/营养构件生物量、贮藏构件生物量/营养构件生物量、繁殖构件生物量/贮藏构件生物量的排序在3种土壤类型中发生转变(表1).这表明蒙古冰草叶、茎、根、穗等构件以及繁殖、营养和贮藏构件的功能分配在不同土壤间存在权衡,蒙古冰草通过调整生物量分配策略以适应不同土壤类型,如在养分较好的灰钙土中以营养分配和贮藏分配为主,在黑垆土中则是减少贮藏分配而增加营养分配和繁殖分配,而在养分较差的风沙土中则是减少营养分配而增加贮藏分配和繁殖分配.另外,我们发现根、茎和穗生物量分配比例在不同类型土壤中一般存在显著差异,但叶生物量分配在不同类型土壤中无显著差异(图2).这表明蒙古冰草构件生物量分配主要受土壤水分和养分资源等限制. ...
祁连山东段不同植物群落的特征及生物量分配
0
2020
不同海拔梯度牧草生物型种群构件的生物量分配策略
2
2021
... 自国际生物学计划(IBP)实行以来,植被生物量一直是生态学的重要研究方向[10].植株构件生物量能够表征植物个体及种群的生长发育节律,体现植物个体及种群适应环境的生存策略,其大小和变化反映了生态系统的物质流、能量流和重要功能[11-13].生物量分配则反映了植物对环境的适应能力和调节自身资源配置以应对特定环境的生长和生殖策略[14-15],是植物生命史和生态学理论的基础[16],对生态动力学和全球变化起重要作用[17].最优分配理论(Optimal Partitioning Theory)认为植物将生物量分配给获取最有限资源的器官,是叶、茎和根等构件功能分配的权衡[18-19],受气候、植物种类、土壤理化性质、土壤养分、土壤水分、光照和CO2浓度等因素的影响[17,19].如植物受到水分和养分的胁迫时会增加根系生物量的投入,而受到光胁迫时会增加冠层生物量的投入.最优分配理论虽然已得到多数研究的证实,但其普遍性仍受到质疑[20-21].如在考虑了植物大小时,种内的生物量分配存在变异,异速生长生物量分配理论(Allometric Biomass Partitioning Theory)有助于解释这一变异.异速生长关系能够揭示植物器官生长的内在规律,是生物量分配和器官功能关系的重要研究方式[22-25].一般表现为植物的某一器官生物量或总生物量与其他性状呈幂函数关系[22,26-28].如异速生长生物量分配理论认为,对于异速生长的理想植物,叶质量(Ml)与根质量(Mr)的关系为(Ml=1/βMr3/4),茎质量(Ms)和根质量(Mr)呈等比例关系[19].异速生长生物量分配理论是最优分配理论的有效补充,二者结合应用可以更好地揭示植物如何分配生物量以响应植物个体大小和环境的变化. ...
... 为获取标准样株,于2021年8月蒙古冰草种子成熟后,分别测量每个样方中蒙古冰草的株高,每个土壤类型各记录20组数据,然后在每个样方中随机选取6~7株挖取整株,带回实验室,将地上部分按照茎、叶和穗分开;地下部分去除死根,冲洗干净.将茎、叶、穗、根分别置于信封中.于105 ℃杀青30 min后,置于65 ℃下烘干至恒重,采用分析天平称其干重,记录每株植物各构件的重量,并按照张鲜花等[13]的方法计算: ...
荒漠草地4种灌木生物量分配特征
1
2013
... 自国际生物学计划(IBP)实行以来,植被生物量一直是生态学的重要研究方向[10].植株构件生物量能够表征植物个体及种群的生长发育节律,体现植物个体及种群适应环境的生存策略,其大小和变化反映了生态系统的物质流、能量流和重要功能[11-13].生物量分配则反映了植物对环境的适应能力和调节自身资源配置以应对特定环境的生长和生殖策略[14-15],是植物生命史和生态学理论的基础[16],对生态动力学和全球变化起重要作用[17].最优分配理论(Optimal Partitioning Theory)认为植物将生物量分配给获取最有限资源的器官,是叶、茎和根等构件功能分配的权衡[18-19],受气候、植物种类、土壤理化性质、土壤养分、土壤水分、光照和CO2浓度等因素的影响[17,19].如植物受到水分和养分的胁迫时会增加根系生物量的投入,而受到光胁迫时会增加冠层生物量的投入.最优分配理论虽然已得到多数研究的证实,但其普遍性仍受到质疑[20-21].如在考虑了植物大小时,种内的生物量分配存在变异,异速生长生物量分配理论(Allometric Biomass Partitioning Theory)有助于解释这一变异.异速生长关系能够揭示植物器官生长的内在规律,是生物量分配和器官功能关系的重要研究方式[22-25].一般表现为植物的某一器官生物量或总生物量与其他性状呈幂函数关系[22,26-28].如异速生长生物量分配理论认为,对于异速生长的理想植物,叶质量(Ml)与根质量(Mr)的关系为(Ml=1/βMr3/4),茎质量(Ms)和根质量(Mr)呈等比例关系[19].异速生长生物量分配理论是最优分配理论的有效补充,二者结合应用可以更好地揭示植物如何分配生物量以响应植物个体大小和环境的变化. ...
不同株高野生无芒雀麦表型特征及生物量分配研究
1
2020
... 自国际生物学计划(IBP)实行以来,植被生物量一直是生态学的重要研究方向[10].植株构件生物量能够表征植物个体及种群的生长发育节律,体现植物个体及种群适应环境的生存策略,其大小和变化反映了生态系统的物质流、能量流和重要功能[11-13].生物量分配则反映了植物对环境的适应能力和调节自身资源配置以应对特定环境的生长和生殖策略[14-15],是植物生命史和生态学理论的基础[16],对生态动力学和全球变化起重要作用[17].最优分配理论(Optimal Partitioning Theory)认为植物将生物量分配给获取最有限资源的器官,是叶、茎和根等构件功能分配的权衡[18-19],受气候、植物种类、土壤理化性质、土壤养分、土壤水分、光照和CO2浓度等因素的影响[17,19].如植物受到水分和养分的胁迫时会增加根系生物量的投入,而受到光胁迫时会增加冠层生物量的投入.最优分配理论虽然已得到多数研究的证实,但其普遍性仍受到质疑[20-21].如在考虑了植物大小时,种内的生物量分配存在变异,异速生长生物量分配理论(Allometric Biomass Partitioning Theory)有助于解释这一变异.异速生长关系能够揭示植物器官生长的内在规律,是生物量分配和器官功能关系的重要研究方式[22-25].一般表现为植物的某一器官生物量或总生物量与其他性状呈幂函数关系[22,26-28].如异速生长生物量分配理论认为,对于异速生长的理想植物,叶质量(Ml)与根质量(Mr)的关系为(Ml=1/βMr3/4),茎质量(Ms)和根质量(Mr)呈等比例关系[19].异速生长生物量分配理论是最优分配理论的有效补充,二者结合应用可以更好地揭示植物如何分配生物量以响应植物个体大小和环境的变化. ...
Dynamic optimization of plant growth
1
2000
... 自国际生物学计划(IBP)实行以来,植被生物量一直是生态学的重要研究方向[10].植株构件生物量能够表征植物个体及种群的生长发育节律,体现植物个体及种群适应环境的生存策略,其大小和变化反映了生态系统的物质流、能量流和重要功能[11-13].生物量分配则反映了植物对环境的适应能力和调节自身资源配置以应对特定环境的生长和生殖策略[14-15],是植物生命史和生态学理论的基础[16],对生态动力学和全球变化起重要作用[17].最优分配理论(Optimal Partitioning Theory)认为植物将生物量分配给获取最有限资源的器官,是叶、茎和根等构件功能分配的权衡[18-19],受气候、植物种类、土壤理化性质、土壤养分、土壤水分、光照和CO2浓度等因素的影响[17,19].如植物受到水分和养分的胁迫时会增加根系生物量的投入,而受到光胁迫时会增加冠层生物量的投入.最优分配理论虽然已得到多数研究的证实,但其普遍性仍受到质疑[20-21].如在考虑了植物大小时,种内的生物量分配存在变异,异速生长生物量分配理论(Allometric Biomass Partitioning Theory)有助于解释这一变异.异速生长关系能够揭示植物器官生长的内在规律,是生物量分配和器官功能关系的重要研究方式[22-25].一般表现为植物的某一器官生物量或总生物量与其他性状呈幂函数关系[22,26-28].如异速生长生物量分配理论认为,对于异速生长的理想植物,叶质量(Ml)与根质量(Mr)的关系为(Ml=1/βMr3/4),茎质量(Ms)和根质量(Mr)呈等比例关系[19].异速生长生物量分配理论是最优分配理论的有效补充,二者结合应用可以更好地揭示植物如何分配生物量以响应植物个体大小和环境的变化. ...
Biomass allocation in old-field annual species grown in elevated CO2 environments: no evidence for optimal partitioning
2
2000
... 自国际生物学计划(IBP)实行以来,植被生物量一直是生态学的重要研究方向[10].植株构件生物量能够表征植物个体及种群的生长发育节律,体现植物个体及种群适应环境的生存策略,其大小和变化反映了生态系统的物质流、能量流和重要功能[11-13].生物量分配则反映了植物对环境的适应能力和调节自身资源配置以应对特定环境的生长和生殖策略[14-15],是植物生命史和生态学理论的基础[16],对生态动力学和全球变化起重要作用[17].最优分配理论(Optimal Partitioning Theory)认为植物将生物量分配给获取最有限资源的器官,是叶、茎和根等构件功能分配的权衡[18-19],受气候、植物种类、土壤理化性质、土壤养分、土壤水分、光照和CO2浓度等因素的影响[17,19].如植物受到水分和养分的胁迫时会增加根系生物量的投入,而受到光胁迫时会增加冠层生物量的投入.最优分配理论虽然已得到多数研究的证实,但其普遍性仍受到质疑[20-21].如在考虑了植物大小时,种内的生物量分配存在变异,异速生长生物量分配理论(Allometric Biomass Partitioning Theory)有助于解释这一变异.异速生长关系能够揭示植物器官生长的内在规律,是生物量分配和器官功能关系的重要研究方式[22-25].一般表现为植物的某一器官生物量或总生物量与其他性状呈幂函数关系[22,26-28].如异速生长生物量分配理论认为,对于异速生长的理想植物,叶质量(Ml)与根质量(Mr)的关系为(Ml=1/βMr3/4),茎质量(Ms)和根质量(Mr)呈等比例关系[19].异速生长生物量分配理论是最优分配理论的有效补充,二者结合应用可以更好地揭示植物如何分配生物量以响应植物个体大小和环境的变化. ...
... [17,19].如植物受到水分和养分的胁迫时会增加根系生物量的投入,而受到光胁迫时会增加冠层生物量的投入.最优分配理论虽然已得到多数研究的证实,但其普遍性仍受到质疑[20-21].如在考虑了植物大小时,种内的生物量分配存在变异,异速生长生物量分配理论(Allometric Biomass Partitioning Theory)有助于解释这一变异.异速生长关系能够揭示植物器官生长的内在规律,是生物量分配和器官功能关系的重要研究方式[22-25].一般表现为植物的某一器官生物量或总生物量与其他性状呈幂函数关系[22,26-28].如异速生长生物量分配理论认为,对于异速生长的理想植物,叶质量(Ml)与根质量(Mr)的关系为(Ml=1/βMr3/4),茎质量(Ms)和根质量(Mr)呈等比例关系[19].异速生长生物量分配理论是最优分配理论的有效补充,二者结合应用可以更好地揭示植物如何分配生物量以响应植物个体大小和环境的变化. ...
Resource limitation in plants:an economic analogy
1
1985
... 自国际生物学计划(IBP)实行以来,植被生物量一直是生态学的重要研究方向[10].植株构件生物量能够表征植物个体及种群的生长发育节律,体现植物个体及种群适应环境的生存策略,其大小和变化反映了生态系统的物质流、能量流和重要功能[11-13].生物量分配则反映了植物对环境的适应能力和调节自身资源配置以应对特定环境的生长和生殖策略[14-15],是植物生命史和生态学理论的基础[16],对生态动力学和全球变化起重要作用[17].最优分配理论(Optimal Partitioning Theory)认为植物将生物量分配给获取最有限资源的器官,是叶、茎和根等构件功能分配的权衡[18-19],受气候、植物种类、土壤理化性质、土壤养分、土壤水分、光照和CO2浓度等因素的影响[17,19].如植物受到水分和养分的胁迫时会增加根系生物量的投入,而受到光胁迫时会增加冠层生物量的投入.最优分配理论虽然已得到多数研究的证实,但其普遍性仍受到质疑[20-21].如在考虑了植物大小时,种内的生物量分配存在变异,异速生长生物量分配理论(Allometric Biomass Partitioning Theory)有助于解释这一变异.异速生长关系能够揭示植物器官生长的内在规律,是生物量分配和器官功能关系的重要研究方式[22-25].一般表现为植物的某一器官生物量或总生物量与其他性状呈幂函数关系[22,26-28].如异速生长生物量分配理论认为,对于异速生长的理想植物,叶质量(Ml)与根质量(Mr)的关系为(Ml=1/βMr3/4),茎质量(Ms)和根质量(Mr)呈等比例关系[19].异速生长生物量分配理论是最优分配理论的有效补充,二者结合应用可以更好地揭示植物如何分配生物量以响应植物个体大小和环境的变化. ...
Consistency between an allometric approach and optimal partitioning theory in global patterns of plant biomass allocation
3
2007
... 自国际生物学计划(IBP)实行以来,植被生物量一直是生态学的重要研究方向[10].植株构件生物量能够表征植物个体及种群的生长发育节律,体现植物个体及种群适应环境的生存策略,其大小和变化反映了生态系统的物质流、能量流和重要功能[11-13].生物量分配则反映了植物对环境的适应能力和调节自身资源配置以应对特定环境的生长和生殖策略[14-15],是植物生命史和生态学理论的基础[16],对生态动力学和全球变化起重要作用[17].最优分配理论(Optimal Partitioning Theory)认为植物将生物量分配给获取最有限资源的器官,是叶、茎和根等构件功能分配的权衡[18-19],受气候、植物种类、土壤理化性质、土壤养分、土壤水分、光照和CO2浓度等因素的影响[17,19].如植物受到水分和养分的胁迫时会增加根系生物量的投入,而受到光胁迫时会增加冠层生物量的投入.最优分配理论虽然已得到多数研究的证实,但其普遍性仍受到质疑[20-21].如在考虑了植物大小时,种内的生物量分配存在变异,异速生长生物量分配理论(Allometric Biomass Partitioning Theory)有助于解释这一变异.异速生长关系能够揭示植物器官生长的内在规律,是生物量分配和器官功能关系的重要研究方式[22-25].一般表现为植物的某一器官生物量或总生物量与其他性状呈幂函数关系[22,26-28].如异速生长生物量分配理论认为,对于异速生长的理想植物,叶质量(Ml)与根质量(Mr)的关系为(Ml=1/βMr3/4),茎质量(Ms)和根质量(Mr)呈等比例关系[19].异速生长生物量分配理论是最优分配理论的有效补充,二者结合应用可以更好地揭示植物如何分配生物量以响应植物个体大小和环境的变化. ...
... ,19].如植物受到水分和养分的胁迫时会增加根系生物量的投入,而受到光胁迫时会增加冠层生物量的投入.最优分配理论虽然已得到多数研究的证实,但其普遍性仍受到质疑[20-21].如在考虑了植物大小时,种内的生物量分配存在变异,异速生长生物量分配理论(Allometric Biomass Partitioning Theory)有助于解释这一变异.异速生长关系能够揭示植物器官生长的内在规律,是生物量分配和器官功能关系的重要研究方式[22-25].一般表现为植物的某一器官生物量或总生物量与其他性状呈幂函数关系[22,26-28].如异速生长生物量分配理论认为,对于异速生长的理想植物,叶质量(Ml)与根质量(Mr)的关系为(Ml=1/βMr3/4),茎质量(Ms)和根质量(Mr)呈等比例关系[19].异速生长生物量分配理论是最优分配理论的有效补充,二者结合应用可以更好地揭示植物如何分配生物量以响应植物个体大小和环境的变化. ...
... [19].异速生长生物量分配理论是最优分配理论的有效补充,二者结合应用可以更好地揭示植物如何分配生物量以响应植物个体大小和环境的变化. ...
The effect of nutrient availability on biomass allocation patterns in 27 species of herbaceous plants
1
2000
... 自国际生物学计划(IBP)实行以来,植被生物量一直是生态学的重要研究方向[10].植株构件生物量能够表征植物个体及种群的生长发育节律,体现植物个体及种群适应环境的生存策略,其大小和变化反映了生态系统的物质流、能量流和重要功能[11-13].生物量分配则反映了植物对环境的适应能力和调节自身资源配置以应对特定环境的生长和生殖策略[14-15],是植物生命史和生态学理论的基础[16],对生态动力学和全球变化起重要作用[17].最优分配理论(Optimal Partitioning Theory)认为植物将生物量分配给获取最有限资源的器官,是叶、茎和根等构件功能分配的权衡[18-19],受气候、植物种类、土壤理化性质、土壤养分、土壤水分、光照和CO2浓度等因素的影响[17,19].如植物受到水分和养分的胁迫时会增加根系生物量的投入,而受到光胁迫时会增加冠层生物量的投入.最优分配理论虽然已得到多数研究的证实,但其普遍性仍受到质疑[20-21].如在考虑了植物大小时,种内的生物量分配存在变异,异速生长生物量分配理论(Allometric Biomass Partitioning Theory)有助于解释这一变异.异速生长关系能够揭示植物器官生长的内在规律,是生物量分配和器官功能关系的重要研究方式[22-25].一般表现为植物的某一器官生物量或总生物量与其他性状呈幂函数关系[22,26-28].如异速生长生物量分配理论认为,对于异速生长的理想植物,叶质量(Ml)与根质量(Mr)的关系为(Ml=1/βMr3/4),茎质量(Ms)和根质量(Mr)呈等比例关系[19].异速生长生物量分配理论是最优分配理论的有效补充,二者结合应用可以更好地揭示植物如何分配生物量以响应植物个体大小和环境的变化. ...
Root-shoot relations:optimality in acclimation and adaptation or the ′Emperor′s New Clothes
1
2002
... 自国际生物学计划(IBP)实行以来,植被生物量一直是生态学的重要研究方向[10].植株构件生物量能够表征植物个体及种群的生长发育节律,体现植物个体及种群适应环境的生存策略,其大小和变化反映了生态系统的物质流、能量流和重要功能[11-13].生物量分配则反映了植物对环境的适应能力和调节自身资源配置以应对特定环境的生长和生殖策略[14-15],是植物生命史和生态学理论的基础[16],对生态动力学和全球变化起重要作用[17].最优分配理论(Optimal Partitioning Theory)认为植物将生物量分配给获取最有限资源的器官,是叶、茎和根等构件功能分配的权衡[18-19],受气候、植物种类、土壤理化性质、土壤养分、土壤水分、光照和CO2浓度等因素的影响[17,19].如植物受到水分和养分的胁迫时会增加根系生物量的投入,而受到光胁迫时会增加冠层生物量的投入.最优分配理论虽然已得到多数研究的证实,但其普遍性仍受到质疑[20-21].如在考虑了植物大小时,种内的生物量分配存在变异,异速生长生物量分配理论(Allometric Biomass Partitioning Theory)有助于解释这一变异.异速生长关系能够揭示植物器官生长的内在规律,是生物量分配和器官功能关系的重要研究方式[22-25].一般表现为植物的某一器官生物量或总生物量与其他性状呈幂函数关系[22,26-28].如异速生长生物量分配理论认为,对于异速生长的理想植物,叶质量(Ml)与根质量(Mr)的关系为(Ml=1/βMr3/4),茎质量(Ms)和根质量(Mr)呈等比例关系[19].异速生长生物量分配理论是最优分配理论的有效补充,二者结合应用可以更好地揭示植物如何分配生物量以响应植物个体大小和环境的变化. ...
Plant allometry:is there a grand unifying theory?
2
2004
... 自国际生物学计划(IBP)实行以来,植被生物量一直是生态学的重要研究方向[10].植株构件生物量能够表征植物个体及种群的生长发育节律,体现植物个体及种群适应环境的生存策略,其大小和变化反映了生态系统的物质流、能量流和重要功能[11-13].生物量分配则反映了植物对环境的适应能力和调节自身资源配置以应对特定环境的生长和生殖策略[14-15],是植物生命史和生态学理论的基础[16],对生态动力学和全球变化起重要作用[17].最优分配理论(Optimal Partitioning Theory)认为植物将生物量分配给获取最有限资源的器官,是叶、茎和根等构件功能分配的权衡[18-19],受气候、植物种类、土壤理化性质、土壤养分、土壤水分、光照和CO2浓度等因素的影响[17,19].如植物受到水分和养分的胁迫时会增加根系生物量的投入,而受到光胁迫时会增加冠层生物量的投入.最优分配理论虽然已得到多数研究的证实,但其普遍性仍受到质疑[20-21].如在考虑了植物大小时,种内的生物量分配存在变异,异速生长生物量分配理论(Allometric Biomass Partitioning Theory)有助于解释这一变异.异速生长关系能够揭示植物器官生长的内在规律,是生物量分配和器官功能关系的重要研究方式[22-25].一般表现为植物的某一器官生物量或总生物量与其他性状呈幂函数关系[22,26-28].如异速生长生物量分配理论认为,对于异速生长的理想植物,叶质量(Ml)与根质量(Mr)的关系为(Ml=1/βMr3/4),茎质量(Ms)和根质量(Mr)呈等比例关系[19].异速生长生物量分配理论是最优分配理论的有效补充,二者结合应用可以更好地揭示植物如何分配生物量以响应植物个体大小和环境的变化. ...
... [22,26-28].如异速生长生物量分配理论认为,对于异速生长的理想植物,叶质量(Ml)与根质量(Mr)的关系为(Ml=1/βMr3/4),茎质量(Ms)和根质量(Mr)呈等比例关系[19].异速生长生物量分配理论是最优分配理论的有效补充,二者结合应用可以更好地揭示植物如何分配生物量以响应植物个体大小和环境的变化. ...
植物生物量分配模式与生长速率的相关规律研究
0
2007
植物代谢速率与个体生物量关系研究进展
1
2011
... 自国际生物学计划(IBP)实行以来,植被生物量一直是生态学的重要研究方向[10].植株构件生物量能够表征植物个体及种群的生长发育节律,体现植物个体及种群适应环境的生存策略,其大小和变化反映了生态系统的物质流、能量流和重要功能[11-13].生物量分配则反映了植物对环境的适应能力和调节自身资源配置以应对特定环境的生长和生殖策略[14-15],是植物生命史和生态学理论的基础[16],对生态动力学和全球变化起重要作用[17].最优分配理论(Optimal Partitioning Theory)认为植物将生物量分配给获取最有限资源的器官,是叶、茎和根等构件功能分配的权衡[18-19],受气候、植物种类、土壤理化性质、土壤养分、土壤水分、光照和CO2浓度等因素的影响[17,19].如植物受到水分和养分的胁迫时会增加根系生物量的投入,而受到光胁迫时会增加冠层生物量的投入.最优分配理论虽然已得到多数研究的证实,但其普遍性仍受到质疑[20-21].如在考虑了植物大小时,种内的生物量分配存在变异,异速生长生物量分配理论(Allometric Biomass Partitioning Theory)有助于解释这一变异.异速生长关系能够揭示植物器官生长的内在规律,是生物量分配和器官功能关系的重要研究方式[22-25].一般表现为植物的某一器官生物量或总生物量与其他性状呈幂函数关系[22,26-28].如异速生长生物量分配理论认为,对于异速生长的理想植物,叶质量(Ml)与根质量(Mr)的关系为(Ml=1/βMr3/4),茎质量(Ms)和根质量(Mr)呈等比例关系[19].异速生长生物量分配理论是最优分配理论的有效补充,二者结合应用可以更好地揭示植物如何分配生物量以响应植物个体大小和环境的变化. ...
A general model for the origin of allometric scaling laws in biology
1
1997
... 自国际生物学计划(IBP)实行以来,植被生物量一直是生态学的重要研究方向[10].植株构件生物量能够表征植物个体及种群的生长发育节律,体现植物个体及种群适应环境的生存策略,其大小和变化反映了生态系统的物质流、能量流和重要功能[11-13].生物量分配则反映了植物对环境的适应能力和调节自身资源配置以应对特定环境的生长和生殖策略[14-15],是植物生命史和生态学理论的基础[16],对生态动力学和全球变化起重要作用[17].最优分配理论(Optimal Partitioning Theory)认为植物将生物量分配给获取最有限资源的器官,是叶、茎和根等构件功能分配的权衡[18-19],受气候、植物种类、土壤理化性质、土壤养分、土壤水分、光照和CO2浓度等因素的影响[17,19].如植物受到水分和养分的胁迫时会增加根系生物量的投入,而受到光胁迫时会增加冠层生物量的投入.最优分配理论虽然已得到多数研究的证实,但其普遍性仍受到质疑[20-21].如在考虑了植物大小时,种内的生物量分配存在变异,异速生长生物量分配理论(Allometric Biomass Partitioning Theory)有助于解释这一变异.异速生长关系能够揭示植物器官生长的内在规律,是生物量分配和器官功能关系的重要研究方式[22-25].一般表现为植物的某一器官生物量或总生物量与其他性状呈幂函数关系[22,26-28].如异速生长生物量分配理论认为,对于异速生长的理想植物,叶质量(Ml)与根质量(Mr)的关系为(Ml=1/βMr3/4),茎质量(Ms)和根质量(Mr)呈等比例关系[19].异速生长生物量分配理论是最优分配理论的有效补充,二者结合应用可以更好地揭示植物如何分配生物量以响应植物个体大小和环境的变化. ...
准噶尔荒漠6种类短命植物生物量分配与异速生长关系
0
2014
四种一年生荒漠植物构件形态与生物量间的异速生长关系
1
2015
... 自国际生物学计划(IBP)实行以来,植被生物量一直是生态学的重要研究方向[10].植株构件生物量能够表征植物个体及种群的生长发育节律,体现植物个体及种群适应环境的生存策略,其大小和变化反映了生态系统的物质流、能量流和重要功能[11-13].生物量分配则反映了植物对环境的适应能力和调节自身资源配置以应对特定环境的生长和生殖策略[14-15],是植物生命史和生态学理论的基础[16],对生态动力学和全球变化起重要作用[17].最优分配理论(Optimal Partitioning Theory)认为植物将生物量分配给获取最有限资源的器官,是叶、茎和根等构件功能分配的权衡[18-19],受气候、植物种类、土壤理化性质、土壤养分、土壤水分、光照和CO2浓度等因素的影响[17,19].如植物受到水分和养分的胁迫时会增加根系生物量的投入,而受到光胁迫时会增加冠层生物量的投入.最优分配理论虽然已得到多数研究的证实,但其普遍性仍受到质疑[20-21].如在考虑了植物大小时,种内的生物量分配存在变异,异速生长生物量分配理论(Allometric Biomass Partitioning Theory)有助于解释这一变异.异速生长关系能够揭示植物器官生长的内在规律,是生物量分配和器官功能关系的重要研究方式[22-25].一般表现为植物的某一器官生物量或总生物量与其他性状呈幂函数关系[22,26-28].如异速生长生物量分配理论认为,对于异速生长的理想植物,叶质量(Ml)与根质量(Mr)的关系为(Ml=1/βMr3/4),茎质量(Ms)和根质量(Mr)呈等比例关系[19].异速生长生物量分配理论是最优分配理论的有效补充,二者结合应用可以更好地揭示植物如何分配生物量以响应植物个体大小和环境的变化. ...
不同雪被厚度下典型高山草地早春植物叶片性状、株高及生物量分配的研究
1
2016
... 植物在环境变化和胁迫因子下具有各自生长特征和物质分配规律[29].有研究表明干旱荒漠区草本植物的地上地下生物量分配遵从异速分配原则[30],一些荒漠植物生物量分配对土壤因子的响应也具有物种特异性[31].宁夏荒漠草原土壤类型空间异质性强,蒙古冰草广泛分布于多种土壤类型上[32],如风沙土、灰钙土等[33].目前,国内外关于蒙古冰草生物量分配对放牧方式、混播方式和CO2浓度变化等的响应已有报道[3,34-35],但很少有研究考虑土壤类型的影响.因此,生长在同一气候条件下、不同土壤类型的蒙古冰草是否符合最优分配理论和异速生长生物量分配理论还需要进一步的试验验证. ...
生物与非生物因子对不同类型植物生物量分配模式的影响
1
2019
... 植物在环境变化和胁迫因子下具有各自生长特征和物质分配规律[29].有研究表明干旱荒漠区草本植物的地上地下生物量分配遵从异速分配原则[30],一些荒漠植物生物量分配对土壤因子的响应也具有物种特异性[31].宁夏荒漠草原土壤类型空间异质性强,蒙古冰草广泛分布于多种土壤类型上[32],如风沙土、灰钙土等[33].目前,国内外关于蒙古冰草生物量分配对放牧方式、混播方式和CO2浓度变化等的响应已有报道[3,34-35],但很少有研究考虑土壤类型的影响.因此,生长在同一气候条件下、不同土壤类型的蒙古冰草是否符合最优分配理论和异速生长生物量分配理论还需要进一步的试验验证. ...
四种荒漠植物生物量分配对土壤因子的响应及异速生长分析
1
2019
... 植物在环境变化和胁迫因子下具有各自生长特征和物质分配规律[29].有研究表明干旱荒漠区草本植物的地上地下生物量分配遵从异速分配原则[30],一些荒漠植物生物量分配对土壤因子的响应也具有物种特异性[31].宁夏荒漠草原土壤类型空间异质性强,蒙古冰草广泛分布于多种土壤类型上[32],如风沙土、灰钙土等[33].目前,国内外关于蒙古冰草生物量分配对放牧方式、混播方式和CO2浓度变化等的响应已有报道[3,34-35],但很少有研究考虑土壤类型的影响.因此,生长在同一气候条件下、不同土壤类型的蒙古冰草是否符合最优分配理论和异速生长生物量分配理论还需要进一步的试验验证. ...
冰草属牧草的种类与分布
1
1989
... 植物在环境变化和胁迫因子下具有各自生长特征和物质分配规律[29].有研究表明干旱荒漠区草本植物的地上地下生物量分配遵从异速分配原则[30],一些荒漠植物生物量分配对土壤因子的响应也具有物种特异性[31].宁夏荒漠草原土壤类型空间异质性强,蒙古冰草广泛分布于多种土壤类型上[32],如风沙土、灰钙土等[33].目前,国内外关于蒙古冰草生物量分配对放牧方式、混播方式和CO2浓度变化等的响应已有报道[3,34-35],但很少有研究考虑土壤类型的影响.因此,生长在同一气候条件下、不同土壤类型的蒙古冰草是否符合最优分配理论和异速生长生物量分配理论还需要进一步的试验验证. ...
宁夏荒漠草原不同群落蒙古冰草种群空间格局及种间关联性
1
2020
... 植物在环境变化和胁迫因子下具有各自生长特征和物质分配规律[29].有研究表明干旱荒漠区草本植物的地上地下生物量分配遵从异速分配原则[30],一些荒漠植物生物量分配对土壤因子的响应也具有物种特异性[31].宁夏荒漠草原土壤类型空间异质性强,蒙古冰草广泛分布于多种土壤类型上[32],如风沙土、灰钙土等[33].目前,国内外关于蒙古冰草生物量分配对放牧方式、混播方式和CO2浓度变化等的响应已有报道[3,34-35],但很少有研究考虑土壤类型的影响.因此,生长在同一气候条件下、不同土壤类型的蒙古冰草是否符合最优分配理论和异速生长生物量分配理论还需要进一步的试验验证. ...
沙芦草+胡枝子混播草地的生物量及土壤速效养分
1
2015
... 植物在环境变化和胁迫因子下具有各自生长特征和物质分配规律[29].有研究表明干旱荒漠区草本植物的地上地下生物量分配遵从异速分配原则[30],一些荒漠植物生物量分配对土壤因子的响应也具有物种特异性[31].宁夏荒漠草原土壤类型空间异质性强,蒙古冰草广泛分布于多种土壤类型上[32],如风沙土、灰钙土等[33].目前,国内外关于蒙古冰草生物量分配对放牧方式、混播方式和CO2浓度变化等的响应已有报道[3,34-35],但很少有研究考虑土壤类型的影响.因此,生长在同一气候条件下、不同土壤类型的蒙古冰草是否符合最优分配理论和异速生长生物量分配理论还需要进一步的试验验证. ...
CO2浓度升高对土壤环境及2种牧草生长的影响
1
2016
... 植物在环境变化和胁迫因子下具有各自生长特征和物质分配规律[29].有研究表明干旱荒漠区草本植物的地上地下生物量分配遵从异速分配原则[30],一些荒漠植物生物量分配对土壤因子的响应也具有物种特异性[31].宁夏荒漠草原土壤类型空间异质性强,蒙古冰草广泛分布于多种土壤类型上[32],如风沙土、灰钙土等[33].目前,国内外关于蒙古冰草生物量分配对放牧方式、混播方式和CO2浓度变化等的响应已有报道[3,34-35],但很少有研究考虑土壤类型的影响.因此,生长在同一气候条件下、不同土壤类型的蒙古冰草是否符合最优分配理论和异速生长生物量分配理论还需要进一步的试验验证. ...
宁夏盐池县荒漠草原区不同群落优势植物叶片-土壤生态化学计量特征
1
2021
... 研究区位于宁夏回族自治区盐池县(37°04′—38°10′N,106°30′—107°47′E),北接毛乌素沙地、南靠黄土高原,海拔1 279~1 954 m,地形南高北低、中部高而东西低.地处典型的半干旱与干旱气候过渡区,属典型的大陆性季风气候,具有气候干燥、降水稀少、冬冷夏热、蒸发强烈、日照充分、风沙大等特点.年平均气温为8.1 ℃,最低月平均气温-13 ℃,最高月平均气温22.7 ℃,极端最高温气温34.9 ℃,极端最低气温-24.2 ℃.年降水量250~350 mm,从东南向西北递减,年际变化幅度高于30%,降水多分布在7—9月,占全年降水量的65%以上.蒸发量为降水量的5~7倍.年无霜期165 d左右,≥10 ℃的有效积温为2 945 ℃左右,年日照时数为2 867.3 h,全年太阳辐射总量为140 kJ·cm-2.土壤以灰钙土、风沙土、黑垆土和黄绵土为主,土壤肥力较低、易受侵蚀.南部黄土丘陵区以黑垆土为主,占到整个南部地区土地面积的4/5以上;北部地区以风沙土和灰钙土为主,面积分别占北部地区的46.5%和45%[36]. ...
荒漠草原2种典型群落类型下土壤含水量与土壤粒径分布的关系
1
2019
... 植被在区系上属于亚欧草原区、亚洲中部亚区,群落表现为旱生植物与典型草原建群种多年生禾草的镶嵌式分布格局[37],处于中国典型草原与荒漠草原的过渡地带.由于土壤养分贫瘠,降水少且季节分配不均,以及风沙危害等资源限制,植被类型单一,主要有灌丛、草原、沙地植被和荒漠植被等,植物种类以旱生和中生类型为主.灌木以北沙柳(Salix psammophilia)和小叶锦鸡儿(Caragana microphylla)为主,草原植被主要有蒙古冰草、短花针茅(Stipa breviflora)和大针茅(Stipa grandis)等[38]. ...
北方农牧交错带封育草地土壤含水量变化研究:以宁夏盐池县为例
1
2010
... 植被在区系上属于亚欧草原区、亚洲中部亚区,群落表现为旱生植物与典型草原建群种多年生禾草的镶嵌式分布格局[37],处于中国典型草原与荒漠草原的过渡地带.由于土壤养分贫瘠,降水少且季节分配不均,以及风沙危害等资源限制,植被类型单一,主要有灌丛、草原、沙地植被和荒漠植被等,植物种类以旱生和中生类型为主.灌木以北沙柳(Salix psammophilia)和小叶锦鸡儿(Caragana microphylla)为主,草原植被主要有蒙古冰草、短花针茅(Stipa breviflora)和大针茅(Stipa grandis)等[38]. ...
1
1958
... 在每个样方中按照五点取样法采集0~20 cm土壤样品,同一样方中的土样混合成一个样品,总共9个混合样.样品带回实验室自然风干,去除残留的枯枝落叶后过2 mm筛,用于测定土壤理化性质.有机碳采用总有机碳分析仪(Vario TOC, Elementar, 丹麦)测定,硝态氮和铵态氮含量分别用酚二黄酸比色法和靛酚蓝比色法测定[39],速效磷采用Olsen法,速效钾采用1 mol·L-1 pH=7.0乙酸铵浸提-火焰光度法,pH和电导率分别用5∶1和2.5∶1水土比浸提测定[40]. ...
Diversity and distribution of soil fungal communities associated with biological soil crusts in the southeastern Tengger Desert (China) as revealed by 454 pyrosequencing
1
2016
... 在每个样方中按照五点取样法采集0~20 cm土壤样品,同一样方中的土样混合成一个样品,总共9个混合样.样品带回实验室自然风干,去除残留的枯枝落叶后过2 mm筛,用于测定土壤理化性质.有机碳采用总有机碳分析仪(Vario TOC, Elementar, 丹麦)测定,硝态氮和铵态氮含量分别用酚二黄酸比色法和靛酚蓝比色法测定[39],速效磷采用Olsen法,速效钾采用1 mol·L-1 pH=7.0乙酸铵浸提-火焰光度法,pH和电导率分别用5∶1和2.5∶1水土比浸提测定[40]. ...
腾格里沙漠东南缘4种灌木的生物量预测模型
2
2013
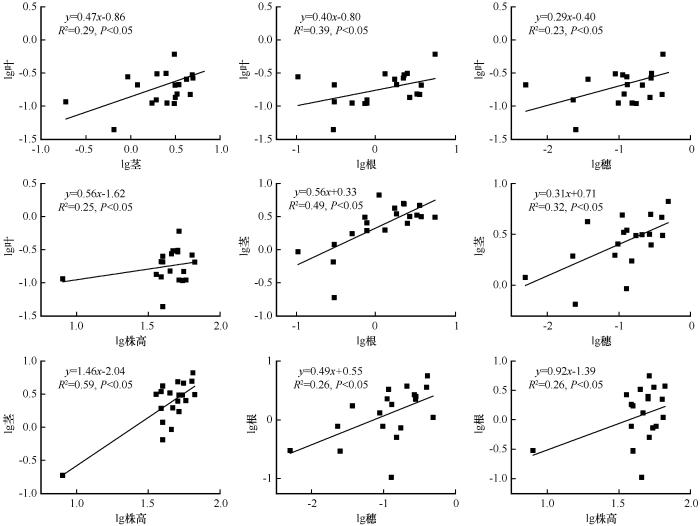
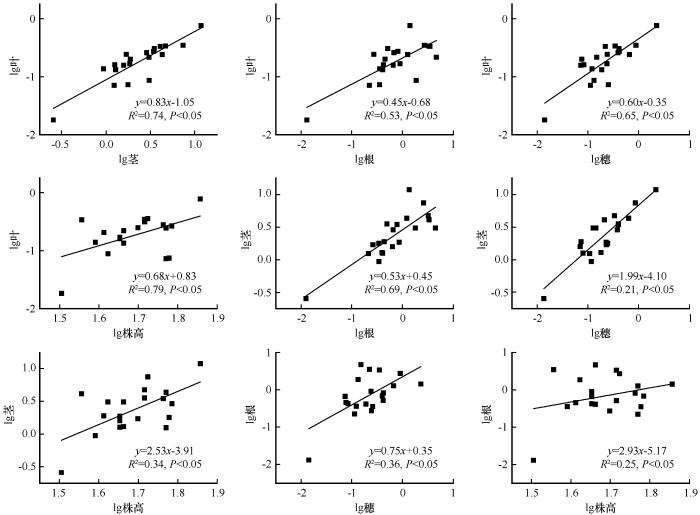
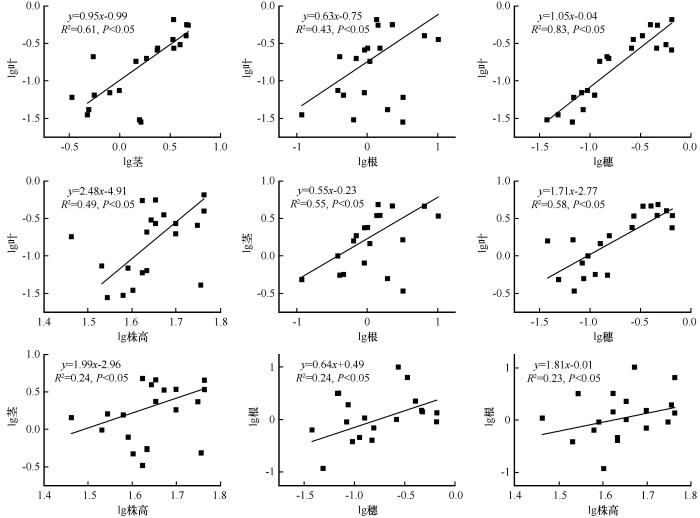
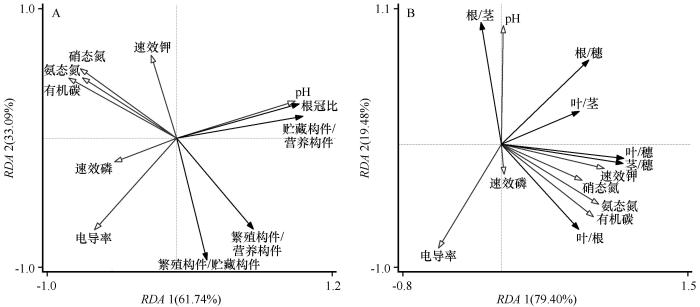
... 采用单因素方差分析法分析土壤类型对土壤理化性质、蒙古冰草各构件生物量、构件分配比例、各构件生物量比率和根冠比的影响,利用最小显著性差异法(Least-Significant Difference,LSD)进行多重比较.基于冗余分析(Redundancy Analysis, RDA)研究土壤理化性质与蒙古冰草各构件生物量分配比例之间的关系.各构件生物量之间以及与总生物量和株高之间的关系采用杨昊天等[41]的异速生长模型进行拟合,即y=axb,对方程两边取对数后转化为线性函数lgy=lga+blgx,其中b为回归直线斜率,lga为截距,通过标准主轴法(Standardized Major Axis, SMA)计算斜率和截距.若b与1.0差异显著,则表明变量间为异速生长关系,若差异不显著,则为等速关系[41].采用SPSS 26.0(SPSS Inc., Illinois, 美国)进行数据统计分析.用Origin 9.0(Origin Lab Corp., 美国)作图.用Canoco 5.0进行冗余分析. ...
... [41].采用SPSS 26.0(SPSS Inc., Illinois, 美国)进行数据统计分析.用Origin 9.0(Origin Lab Corp., 美国)作图.用Canoco 5.0进行冗余分析. ...
1
1994
... 植物生物量分配是陆地生态系统生物地球化学循环中的重要变量,定量理解生物量分配模式是生态管理的重要理论基础[42],对提升植物保护和利用以及生态系统功能具有重要作用.植物通过改变生物量的分配模式以响应环境变化[43],各构件生物量的分配比率和各构件间的比值体现了植物适应环境的策略.模拟试验表明土壤异质性对植物生物量和生物量分配有显著影响[44],但是在自然环境下还缺乏荒漠草原土壤类型对蒙古冰草构件生物量分配模式和异速生长关系的验证.本研究重点探讨了宁夏荒漠草原不同土壤类型对蒙古冰草各构件生物量分配的影响. ...
Variation in growth rate and ecophysiology among 34 grassland and savanna species under contrasting N supply:a test of functional group differences
1
2003
... 植物生物量分配是陆地生态系统生物地球化学循环中的重要变量,定量理解生物量分配模式是生态管理的重要理论基础[42],对提升植物保护和利用以及生态系统功能具有重要作用.植物通过改变生物量的分配模式以响应环境变化[43],各构件生物量的分配比率和各构件间的比值体现了植物适应环境的策略.模拟试验表明土壤异质性对植物生物量和生物量分配有显著影响[44],但是在自然环境下还缺乏荒漠草原土壤类型对蒙古冰草构件生物量分配模式和异速生长关系的验证.本研究重点探讨了宁夏荒漠草原不同土壤类型对蒙古冰草各构件生物量分配的影响. ...
Effects of three-dimensional soil heterogeneity and species composition on plant biomass and biomass allocation of grass-mixtures
1
2021
... 植物生物量分配是陆地生态系统生物地球化学循环中的重要变量,定量理解生物量分配模式是生态管理的重要理论基础[42],对提升植物保护和利用以及生态系统功能具有重要作用.植物通过改变生物量的分配模式以响应环境变化[43],各构件生物量的分配比率和各构件间的比值体现了植物适应环境的策略.模拟试验表明土壤异质性对植物生物量和生物量分配有显著影响[44],但是在自然环境下还缺乏荒漠草原土壤类型对蒙古冰草构件生物量分配模式和异速生长关系的验证.本研究重点探讨了宁夏荒漠草原不同土壤类型对蒙古冰草各构件生物量分配的影响. ...
土壤类型对建瓯雷竹单株生物量的影响
1
2017
... 生物量是植物生长情况最直观的外在表现,通过植物生物量可以对植物的生长情况进行初步判断[45].各土壤类型间蒙古冰草全株总生物量表现为灰钙土>黑垆土>风沙土,与土壤有机碳、硝态氮、铵态氮和速效钾等土壤养分的趋势相同,但差异不显著.由于试验的气候背景一致,即降水、气温、光照等因子相同,表明宁夏荒漠草原3种土壤类型对蒙古冰草总生物量大小有影响但差异不显著. ...
Biomass allocation in plants:ontogeny or optimality?a test along three resource gradients
1
1999
... 植物如何响应环境中资源可用性的变化是植物生态学的核心问题[46].植物构件生物量的分配反映了植物的发育规律和生存策略,影响植物个体的生长和植被结构,以及草地土壤碳输入和生态系统碳循环[11,47-49].有研究表明,土壤质地、碳氮比等因素对全球植物根系生物量分配比例起重要作用[50].在3种土壤类型条件下,蒙古冰草的构件生物量分配都表现出贮藏分配最多、营养分配次之、繁殖分配最少的规律,这与赵盼盼等[3]的研究结果一致.荒漠草原植物生长的主要限制性因子是水分和土壤养分[50],本控制试验中3种土壤类型具有相同的降水条件,尤其是在试验结束前经历了长达80 d的干旱,可以很好地探讨干旱条件下,不同土壤类型蒙古冰草是否符合最优分配理论.蒙古冰草根、茎、穗、营养分配、贮藏分配和繁殖分配比率在不同土壤类型间存在显著差异,各构件生物量比在不同土壤类型间也存在差异,且叶/根、叶/穗、茎/穗、叶/茎、根/茎、根/穗、繁殖构件生物量/营养构件生物量、贮藏构件生物量/营养构件生物量、繁殖构件生物量/贮藏构件生物量的排序在3种土壤类型中发生转变(表1).这表明蒙古冰草叶、茎、根、穗等构件以及繁殖、营养和贮藏构件的功能分配在不同土壤间存在权衡,蒙古冰草通过调整生物量分配策略以适应不同土壤类型,如在养分较好的灰钙土中以营养分配和贮藏分配为主,在黑垆土中则是减少贮藏分配而增加营养分配和繁殖分配,而在养分较差的风沙土中则是减少营养分配而增加贮藏分配和繁殖分配.另外,我们发现根、茎和穗生物量分配比例在不同类型土壤中一般存在显著差异,但叶生物量分配在不同类型土壤中无显著差异(图2).这表明蒙古冰草构件生物量分配主要受土壤水分和养分资源等限制. ...
Carbon allocation in forest ecosystems
1
2007
... 植物如何响应环境中资源可用性的变化是植物生态学的核心问题[46].植物构件生物量的分配反映了植物的发育规律和生存策略,影响植物个体的生长和植被结构,以及草地土壤碳输入和生态系统碳循环[11,47-49].有研究表明,土壤质地、碳氮比等因素对全球植物根系生物量分配比例起重要作用[50].在3种土壤类型条件下,蒙古冰草的构件生物量分配都表现出贮藏分配最多、营养分配次之、繁殖分配最少的规律,这与赵盼盼等[3]的研究结果一致.荒漠草原植物生长的主要限制性因子是水分和土壤养分[50],本控制试验中3种土壤类型具有相同的降水条件,尤其是在试验结束前经历了长达80 d的干旱,可以很好地探讨干旱条件下,不同土壤类型蒙古冰草是否符合最优分配理论.蒙古冰草根、茎、穗、营养分配、贮藏分配和繁殖分配比率在不同土壤类型间存在显著差异,各构件生物量比在不同土壤类型间也存在差异,且叶/根、叶/穗、茎/穗、叶/茎、根/茎、根/穗、繁殖构件生物量/营养构件生物量、贮藏构件生物量/营养构件生物量、繁殖构件生物量/贮藏构件生物量的排序在3种土壤类型中发生转变(表1).这表明蒙古冰草叶、茎、根、穗等构件以及繁殖、营养和贮藏构件的功能分配在不同土壤间存在权衡,蒙古冰草通过调整生物量分配策略以适应不同土壤类型,如在养分较好的灰钙土中以营养分配和贮藏分配为主,在黑垆土中则是减少贮藏分配而增加营养分配和繁殖分配,而在养分较差的风沙土中则是减少营养分配而增加贮藏分配和繁殖分配.另外,我们发现根、茎和穗生物量分配比例在不同类型土壤中一般存在显著差异,但叶生物量分配在不同类型土壤中无显著差异(图2).这表明蒙古冰草构件生物量分配主要受土壤水分和养分资源等限制. ...
5个象草品种的构件生物量特征及分配动态
1
2011
... 植物如何响应环境中资源可用性的变化是植物生态学的核心问题[46].植物构件生物量的分配反映了植物的发育规律和生存策略,影响植物个体的生长和植被结构,以及草地土壤碳输入和生态系统碳循环[11,47-49].有研究表明,土壤质地、碳氮比等因素对全球植物根系生物量分配比例起重要作用[50].在3种土壤类型条件下,蒙古冰草的构件生物量分配都表现出贮藏分配最多、营养分配次之、繁殖分配最少的规律,这与赵盼盼等[3]的研究结果一致.荒漠草原植物生长的主要限制性因子是水分和土壤养分[50],本控制试验中3种土壤类型具有相同的降水条件,尤其是在试验结束前经历了长达80 d的干旱,可以很好地探讨干旱条件下,不同土壤类型蒙古冰草是否符合最优分配理论.蒙古冰草根、茎、穗、营养分配、贮藏分配和繁殖分配比率在不同土壤类型间存在显著差异,各构件生物量比在不同土壤类型间也存在差异,且叶/根、叶/穗、茎/穗、叶/茎、根/茎、根/穗、繁殖构件生物量/营养构件生物量、贮藏构件生物量/营养构件生物量、繁殖构件生物量/贮藏构件生物量的排序在3种土壤类型中发生转变(表1).这表明蒙古冰草叶、茎、根、穗等构件以及繁殖、营养和贮藏构件的功能分配在不同土壤间存在权衡,蒙古冰草通过调整生物量分配策略以适应不同土壤类型,如在养分较好的灰钙土中以营养分配和贮藏分配为主,在黑垆土中则是减少贮藏分配而增加营养分配和繁殖分配,而在养分较差的风沙土中则是减少营养分配而增加贮藏分配和繁殖分配.另外,我们发现根、茎和穗生物量分配比例在不同类型土壤中一般存在显著差异,但叶生物量分配在不同类型土壤中无显著差异(图2).这表明蒙古冰草构件生物量分配主要受土壤水分和养分资源等限制. ...
The global distribution and environmental drivers of aboveground versus belowground plant biomass
2
2021
... 植物如何响应环境中资源可用性的变化是植物生态学的核心问题[46].植物构件生物量的分配反映了植物的发育规律和生存策略,影响植物个体的生长和植被结构,以及草地土壤碳输入和生态系统碳循环[11,47-49].有研究表明,土壤质地、碳氮比等因素对全球植物根系生物量分配比例起重要作用[50].在3种土壤类型条件下,蒙古冰草的构件生物量分配都表现出贮藏分配最多、营养分配次之、繁殖分配最少的规律,这与赵盼盼等[3]的研究结果一致.荒漠草原植物生长的主要限制性因子是水分和土壤养分[50],本控制试验中3种土壤类型具有相同的降水条件,尤其是在试验结束前经历了长达80 d的干旱,可以很好地探讨干旱条件下,不同土壤类型蒙古冰草是否符合最优分配理论.蒙古冰草根、茎、穗、营养分配、贮藏分配和繁殖分配比率在不同土壤类型间存在显著差异,各构件生物量比在不同土壤类型间也存在差异,且叶/根、叶/穗、茎/穗、叶/茎、根/茎、根/穗、繁殖构件生物量/营养构件生物量、贮藏构件生物量/营养构件生物量、繁殖构件生物量/贮藏构件生物量的排序在3种土壤类型中发生转变(表1).这表明蒙古冰草叶、茎、根、穗等构件以及繁殖、营养和贮藏构件的功能分配在不同土壤间存在权衡,蒙古冰草通过调整生物量分配策略以适应不同土壤类型,如在养分较好的灰钙土中以营养分配和贮藏分配为主,在黑垆土中则是减少贮藏分配而增加营养分配和繁殖分配,而在养分较差的风沙土中则是减少营养分配而增加贮藏分配和繁殖分配.另外,我们发现根、茎和穗生物量分配比例在不同类型土壤中一般存在显著差异,但叶生物量分配在不同类型土壤中无显著差异(图2).这表明蒙古冰草构件生物量分配主要受土壤水分和养分资源等限制. ...
... [50],本控制试验中3种土壤类型具有相同的降水条件,尤其是在试验结束前经历了长达80 d的干旱,可以很好地探讨干旱条件下,不同土壤类型蒙古冰草是否符合最优分配理论.蒙古冰草根、茎、穗、营养分配、贮藏分配和繁殖分配比率在不同土壤类型间存在显著差异,各构件生物量比在不同土壤类型间也存在差异,且叶/根、叶/穗、茎/穗、叶/茎、根/茎、根/穗、繁殖构件生物量/营养构件生物量、贮藏构件生物量/营养构件生物量、繁殖构件生物量/贮藏构件生物量的排序在3种土壤类型中发生转变(表1).这表明蒙古冰草叶、茎、根、穗等构件以及繁殖、营养和贮藏构件的功能分配在不同土壤间存在权衡,蒙古冰草通过调整生物量分配策略以适应不同土壤类型,如在养分较好的灰钙土中以营养分配和贮藏分配为主,在黑垆土中则是减少贮藏分配而增加营养分配和繁殖分配,而在养分较差的风沙土中则是减少营养分配而增加贮藏分配和繁殖分配.另外,我们发现根、茎和穗生物量分配比例在不同类型土壤中一般存在显著差异,但叶生物量分配在不同类型土壤中无显著差异(图2).这表明蒙古冰草构件生物量分配主要受土壤水分和养分资源等限制. ...
不同生育期入侵植物大狼把草的生物量分配格局及异速生长分析
1
2017
... 地下生物量分配比例反映了植物的生长策略,通常来说,多年生植物地下部分生物量分配比例更大[51].左有璐等[52]发现生活型影响植物构件生物量分配,一年生植物地上生物量分配比例相对更高,多年生植物地下生物量分配比例相对更高.本研究的根冠比数据表明蒙古冰草在不同类型土壤中的地下和地上生物量分配策略不同,其分配策略与土壤性质密切相关(表1,图6).在营养贫瘠的风沙土中优先将生物量分配给根系以保证能获取足够养分生长和繁殖,而在黑垆土中时则会适当减少根系生物量的分配比例而增加地上生物量.本研究证实,在干旱条件下不同土壤类型中蒙古冰草构件生物量比和根冠比支持最优分配理论. ...
川西北高寒草甸优势植物生物量分配对策
1
2018
... 地下生物量分配比例反映了植物的生长策略,通常来说,多年生植物地下部分生物量分配比例更大[51].左有璐等[52]发现生活型影响植物构件生物量分配,一年生植物地上生物量分配比例相对更高,多年生植物地下生物量分配比例相对更高.本研究的根冠比数据表明蒙古冰草在不同类型土壤中的地下和地上生物量分配策略不同,其分配策略与土壤性质密切相关(表1,图6).在营养贫瘠的风沙土中优先将生物量分配给根系以保证能获取足够养分生长和繁殖,而在黑垆土中时则会适当减少根系生物量的分配比例而增加地上生物量.本研究证实,在干旱条件下不同土壤类型中蒙古冰草构件生物量比和根冠比支持最优分配理论. ...
Direct and indirect impacts of climate change on microbial and biocrust communities alter the resistance of the N cycle in a semiarid grassland
1
2015
... 土壤类型对生物量分配的影响可能主要取决于土壤理化属性,冗余分析表明土壤理化属性可以解释部分构件生物量比.不同类型土壤中蒙古冰草构件生物量分配之所以会出现差异,主要是因为植物为了增强其生态适应性,会减少非限制性资源构件的分配,而增大获取限制性资源的构件分配.土壤有机碳含量在一定范围内,才能使植物生长最优化,有机碳含量过高会造成富营养化而不利于植物生长,但有机碳含量太低又会使得养分循环失衡而影响植物生长.氮素含量是影响干旱区植物生物量的主要营养限制因子[53].土壤速效磷缺乏已成为植物生长的主要限制因子[54],速效磷含量通常与植物生物量呈正比[55-56].宁夏荒漠草原3种土壤类型中有机碳、硝态氮、铵态氮、速效钾的变化趋势一致,且均与蒙古冰草生物量变化趋势一致,但与构件生物量比的关系较为复杂,既有趋势一致的(如叶/根、叶/穗、茎/穗),也有趋势相反的(如构件/营养构件),还有无规律的(如贮藏构件/营养构件、根冠比),还需开展更多的研究解析构件生物量比与土壤养分之间的关系.陈冠光[57]发现干旱区植物生物量随土壤pH的升高呈先增加后减少趋势,土壤pH为8.6左右时生物量最大.本研究结果显示,土壤pH和电导率是影响土壤根、茎、叶和穗生物量分配比例的主要因子(图6A),当土壤pH在最适值范围时(风沙土pH为8.81),蒙古冰草根生物量比例增加,而茎生物量比例减少,土壤pH对蒙古冰草构件生物量分配策略的影响可能是通过调整根和茎的生物量比例来权衡的. ...
农业生态系统养分循环再利用作物产量增益的地理分异
1
1998
... 土壤类型对生物量分配的影响可能主要取决于土壤理化属性,冗余分析表明土壤理化属性可以解释部分构件生物量比.不同类型土壤中蒙古冰草构件生物量分配之所以会出现差异,主要是因为植物为了增强其生态适应性,会减少非限制性资源构件的分配,而增大获取限制性资源的构件分配.土壤有机碳含量在一定范围内,才能使植物生长最优化,有机碳含量过高会造成富营养化而不利于植物生长,但有机碳含量太低又会使得养分循环失衡而影响植物生长.氮素含量是影响干旱区植物生物量的主要营养限制因子[53].土壤速效磷缺乏已成为植物生长的主要限制因子[54],速效磷含量通常与植物生物量呈正比[55-56].宁夏荒漠草原3种土壤类型中有机碳、硝态氮、铵态氮、速效钾的变化趋势一致,且均与蒙古冰草生物量变化趋势一致,但与构件生物量比的关系较为复杂,既有趋势一致的(如叶/根、叶/穗、茎/穗),也有趋势相反的(如构件/营养构件),还有无规律的(如贮藏构件/营养构件、根冠比),还需开展更多的研究解析构件生物量比与土壤养分之间的关系.陈冠光[57]发现干旱区植物生物量随土壤pH的升高呈先增加后减少趋势,土壤pH为8.6左右时生物量最大.本研究结果显示,土壤pH和电导率是影响土壤根、茎、叶和穗生物量分配比例的主要因子(图6A),当土壤pH在最适值范围时(风沙土pH为8.81),蒙古冰草根生物量比例增加,而茎生物量比例减少,土壤pH对蒙古冰草构件生物量分配策略的影响可能是通过调整根和茎的生物量比例来权衡的. ...
土壤磷水平对柱花草株高、根长及生物量的影响
1
2014
... 土壤类型对生物量分配的影响可能主要取决于土壤理化属性,冗余分析表明土壤理化属性可以解释部分构件生物量比.不同类型土壤中蒙古冰草构件生物量分配之所以会出现差异,主要是因为植物为了增强其生态适应性,会减少非限制性资源构件的分配,而增大获取限制性资源的构件分配.土壤有机碳含量在一定范围内,才能使植物生长最优化,有机碳含量过高会造成富营养化而不利于植物生长,但有机碳含量太低又会使得养分循环失衡而影响植物生长.氮素含量是影响干旱区植物生物量的主要营养限制因子[53].土壤速效磷缺乏已成为植物生长的主要限制因子[54],速效磷含量通常与植物生物量呈正比[55-56].宁夏荒漠草原3种土壤类型中有机碳、硝态氮、铵态氮、速效钾的变化趋势一致,且均与蒙古冰草生物量变化趋势一致,但与构件生物量比的关系较为复杂,既有趋势一致的(如叶/根、叶/穗、茎/穗),也有趋势相反的(如构件/营养构件),还有无规律的(如贮藏构件/营养构件、根冠比),还需开展更多的研究解析构件生物量比与土壤养分之间的关系.陈冠光[57]发现干旱区植物生物量随土壤pH的升高呈先增加后减少趋势,土壤pH为8.6左右时生物量最大.本研究结果显示,土壤pH和电导率是影响土壤根、茎、叶和穗生物量分配比例的主要因子(图6A),当土壤pH在最适值范围时(风沙土pH为8.81),蒙古冰草根生物量比例增加,而茎生物量比例减少,土壤pH对蒙古冰草构件生物量分配策略的影响可能是通过调整根和茎的生物量比例来权衡的. ...
不同磷背景土壤磷素用量对烤烟农艺性状及生物量的影响
1
2018
... 土壤类型对生物量分配的影响可能主要取决于土壤理化属性,冗余分析表明土壤理化属性可以解释部分构件生物量比.不同类型土壤中蒙古冰草构件生物量分配之所以会出现差异,主要是因为植物为了增强其生态适应性,会减少非限制性资源构件的分配,而增大获取限制性资源的构件分配.土壤有机碳含量在一定范围内,才能使植物生长最优化,有机碳含量过高会造成富营养化而不利于植物生长,但有机碳含量太低又会使得养分循环失衡而影响植物生长.氮素含量是影响干旱区植物生物量的主要营养限制因子[53].土壤速效磷缺乏已成为植物生长的主要限制因子[54],速效磷含量通常与植物生物量呈正比[55-56].宁夏荒漠草原3种土壤类型中有机碳、硝态氮、铵态氮、速效钾的变化趋势一致,且均与蒙古冰草生物量变化趋势一致,但与构件生物量比的关系较为复杂,既有趋势一致的(如叶/根、叶/穗、茎/穗),也有趋势相反的(如构件/营养构件),还有无规律的(如贮藏构件/营养构件、根冠比),还需开展更多的研究解析构件生物量比与土壤养分之间的关系.陈冠光[57]发现干旱区植物生物量随土壤pH的升高呈先增加后减少趋势,土壤pH为8.6左右时生物量最大.本研究结果显示,土壤pH和电导率是影响土壤根、茎、叶和穗生物量分配比例的主要因子(图6A),当土壤pH在最适值范围时(风沙土pH为8.81),蒙古冰草根生物量比例增加,而茎生物量比例减少,土壤pH对蒙古冰草构件生物量分配策略的影响可能是通过调整根和茎的生物量比例来权衡的. ...
疏勒河下游地下水埋深与土壤理化性质变化对植物群落的影响
1
2020
... 土壤类型对生物量分配的影响可能主要取决于土壤理化属性,冗余分析表明土壤理化属性可以解释部分构件生物量比.不同类型土壤中蒙古冰草构件生物量分配之所以会出现差异,主要是因为植物为了增强其生态适应性,会减少非限制性资源构件的分配,而增大获取限制性资源的构件分配.土壤有机碳含量在一定范围内,才能使植物生长最优化,有机碳含量过高会造成富营养化而不利于植物生长,但有机碳含量太低又会使得养分循环失衡而影响植物生长.氮素含量是影响干旱区植物生物量的主要营养限制因子[53].土壤速效磷缺乏已成为植物生长的主要限制因子[54],速效磷含量通常与植物生物量呈正比[55-56].宁夏荒漠草原3种土壤类型中有机碳、硝态氮、铵态氮、速效钾的变化趋势一致,且均与蒙古冰草生物量变化趋势一致,但与构件生物量比的关系较为复杂,既有趋势一致的(如叶/根、叶/穗、茎/穗),也有趋势相反的(如构件/营养构件),还有无规律的(如贮藏构件/营养构件、根冠比),还需开展更多的研究解析构件生物量比与土壤养分之间的关系.陈冠光[57]发现干旱区植物生物量随土壤pH的升高呈先增加后减少趋势,土壤pH为8.6左右时生物量最大.本研究结果显示,土壤pH和电导率是影响土壤根、茎、叶和穗生物量分配比例的主要因子(图6A),当土壤pH在最适值范围时(风沙土pH为8.81),蒙古冰草根生物量比例增加,而茎生物量比例减少,土壤pH对蒙古冰草构件生物量分配策略的影响可能是通过调整根和茎的生物量比例来权衡的. ...




 甘公网安备 62010202000688号
甘公网安备 62010202000688号
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}