Biocrusts enhance non-rainfall water deposition and alter its distribution in dryland soils
2
2021
... 生物土壤结皮(简称“生物结皮”)是干旱区生态系统的特殊地被层,由细菌、蓝藻、真核藻类、真菌、地衣和苔藓与土壤颗粒胶结而成[1].生物结皮在养分积累、水分保持、减少风蚀水蚀、生物地球化学循环、促进植被演替等过程中发挥着重要的生态功能[2-3].根据生物结皮的优势物种,可将生物结皮分为藻结皮、地衣结皮和苔藓结皮,这种不同类型反映了生物结皮的演替趋势[4].研究普遍认为,生物结皮的发育影响了土壤理化环境和生物学特性,尤其是显著提高了土壤养分含量和微生物活性[5].生物结皮不仅在荒漠生态系统广泛分布,在中国黄土高原植被间隙中亦普遍存在,尤其是近年来随着退耕还林还草工程的实施,大面积退耕撂荒地进入人为扰动减少、植被逐渐恢复的过程,土壤有机碳含量和有效养分增加,促使生物结皮广泛发育,并在生态恢复中扮演着不可忽视的角色[6]. ...
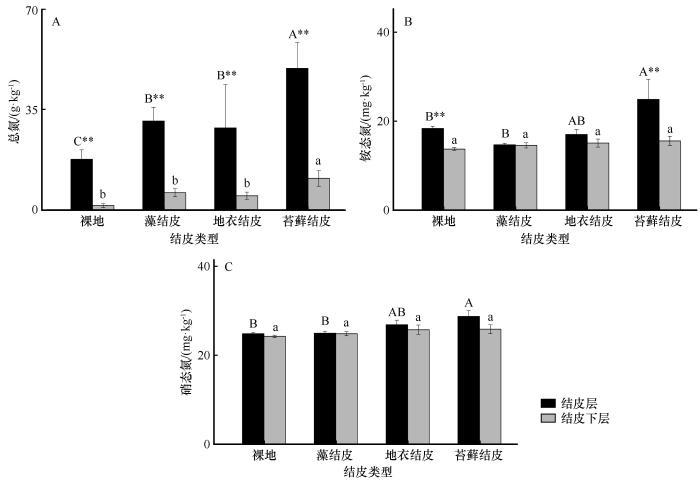
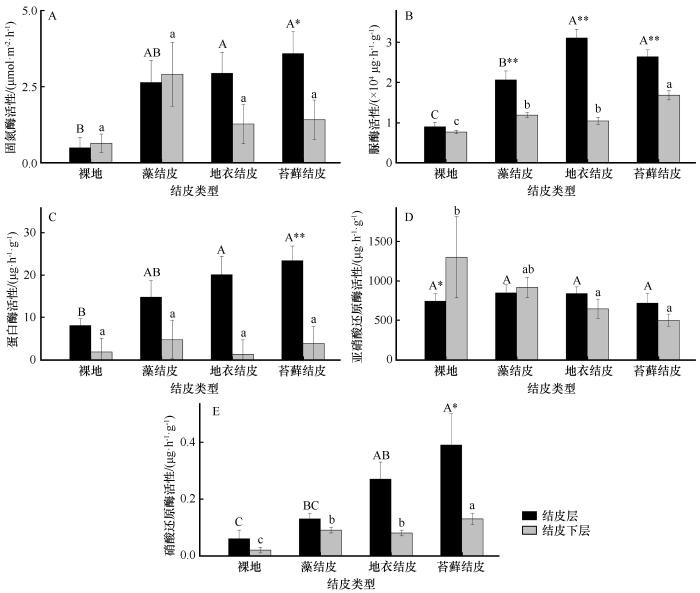
... 土壤氮素的生物化学循环几乎全部依靠微生物介导的氧化还原反应完成[28-29].荒漠中藻结皮、地衣结皮和苔藓结皮的固氮速率差异显著,氮输入强烈依赖于生物结皮中的固氮微生物[30-31].本研究中,TN、NH-N、NO-N含量以及固氮酶活性随生物结皮演替其变化总体呈增加趋势,这在很大程度上取决于生物结皮固氮微生物的物种组成和丰度.在古尔班通古特沙漠中的相关研究表明,藻结皮和地衣结皮中固氮微生物以念珠藻属(Nostoc)和伪枝藻属(Syctonema)为主,且固氮微生物多样性与nifH基因丰度均在藻结皮阶段最高,其次为苔藓结皮[32];也有研究发现固氮蓝细菌是结皮演替早期主要的固氮功能群,随着生物结皮演替,地衣真菌(Leotiomycetes和Lecanoromycetes)及异养固氮菌(放线菌Actinomycetales、根瘤菌Rhizobiales等)成为地衣及苔藓结皮的主要固氮类群[33];而在浑善达克沙地,研究发现固氮微生物多样性指数随生物结皮的演替而上升,其中变形菌门的斯克尔曼氏菌(Skermanella)相对丰度显著增加,在苔藓结皮阶段成为仅次于未分类念珠藻科和伪枝藻属的第三优势固氮类群[34];这种固氮优势类群的差异可能与其多样性和群落组成相关.但在黄土高原水蚀风蚀交错区,生物结皮演替后期尤其是苔藓结皮的固氮酶活性最高,这可能是因为固氮微生物的组成在结皮演替后期发生了重要变化,还需进一步研究证实.在有机氮矿化过程中,有机氮常通过氨化作用被降解为NH[19],因此,NH-N含量在生物结皮后期演替阶段显著升高可能也与有机氮矿化逐渐升高有关.本研究发现黄土高原水蚀风蚀交错区生物结皮演替过程中脲酶和蛋白酶活性显著升高,在苔藓结皮或地衣结皮达到峰值.而脲酶和蛋白酶是蛋白质降解和尿素水解的重要酶类,在有机氮矿化过程中发挥着重要作用[35],说明有机氮的矿化作用随生物结皮演替也呈升高趋势.这种变化趋势可能有两方面原因,一是生物结皮的演替在后期积累了丰富的有机氮,为蛋白酶和脲酶提供了丰富的底物[1,25];二是生物结皮的演替使微生物种类和数量增加,尤其是与编码蛋白酶和脲酶(ureC)相关的功能微生物可能会增加,但有待于实验进一步验证.执行反硝化反应第一步的硝酸还原酶(编码基因narG)是控制反硝化过程的关键酶[36-37],在本研究中,硝酸还原酶在生物结皮演替过程中呈显著升高趋势,预示着硝酸还原酶相关的功能微生物丰度可能在结皮演替过程中有明显的升高. ...
Biological soil crusts enhance the recovery of nutrient levels of surface dune soil in arid desert regions
1
2019
... 生物土壤结皮(简称“生物结皮”)是干旱区生态系统的特殊地被层,由细菌、蓝藻、真核藻类、真菌、地衣和苔藓与土壤颗粒胶结而成[1].生物结皮在养分积累、水分保持、减少风蚀水蚀、生物地球化学循环、促进植被演替等过程中发挥着重要的生态功能[2-3].根据生物结皮的优势物种,可将生物结皮分为藻结皮、地衣结皮和苔藓结皮,这种不同类型反映了生物结皮的演替趋势[4].研究普遍认为,生物结皮的发育影响了土壤理化环境和生物学特性,尤其是显著提高了土壤养分含量和微生物活性[5].生物结皮不仅在荒漠生态系统广泛分布,在中国黄土高原植被间隙中亦普遍存在,尤其是近年来随着退耕还林还草工程的实施,大面积退耕撂荒地进入人为扰动减少、植被逐渐恢复的过程,土壤有机碳含量和有效养分增加,促使生物结皮广泛发育,并在生态恢复中扮演着不可忽视的角色[6]. ...
生物土壤结皮研究:进展、前沿与展望
1
2009
... 生物土壤结皮(简称“生物结皮”)是干旱区生态系统的特殊地被层,由细菌、蓝藻、真核藻类、真菌、地衣和苔藓与土壤颗粒胶结而成[1].生物结皮在养分积累、水分保持、减少风蚀水蚀、生物地球化学循环、促进植被演替等过程中发挥着重要的生态功能[2-3].根据生物结皮的优势物种,可将生物结皮分为藻结皮、地衣结皮和苔藓结皮,这种不同类型反映了生物结皮的演替趋势[4].研究普遍认为,生物结皮的发育影响了土壤理化环境和生物学特性,尤其是显著提高了土壤养分含量和微生物活性[5].生物结皮不仅在荒漠生态系统广泛分布,在中国黄土高原植被间隙中亦普遍存在,尤其是近年来随着退耕还林还草工程的实施,大面积退耕撂荒地进入人为扰动减少、植被逐渐恢复的过程,土壤有机碳含量和有效养分增加,促使生物结皮广泛发育,并在生态恢复中扮演着不可忽视的角色[6]. ...
Successional stages of biological soil crusts and their microstructure variability in Shapotou region (China)
2
2012
... 生物土壤结皮(简称“生物结皮”)是干旱区生态系统的特殊地被层,由细菌、蓝藻、真核藻类、真菌、地衣和苔藓与土壤颗粒胶结而成[1].生物结皮在养分积累、水分保持、减少风蚀水蚀、生物地球化学循环、促进植被演替等过程中发挥着重要的生态功能[2-3].根据生物结皮的优势物种,可将生物结皮分为藻结皮、地衣结皮和苔藓结皮,这种不同类型反映了生物结皮的演替趋势[4].研究普遍认为,生物结皮的发育影响了土壤理化环境和生物学特性,尤其是显著提高了土壤养分含量和微生物活性[5].生物结皮不仅在荒漠生态系统广泛分布,在中国黄土高原植被间隙中亦普遍存在,尤其是近年来随着退耕还林还草工程的实施,大面积退耕撂荒地进入人为扰动减少、植被逐渐恢复的过程,土壤有机碳含量和有效养分增加,促使生物结皮广泛发育,并在生态恢复中扮演着不可忽视的角色[6]. ...
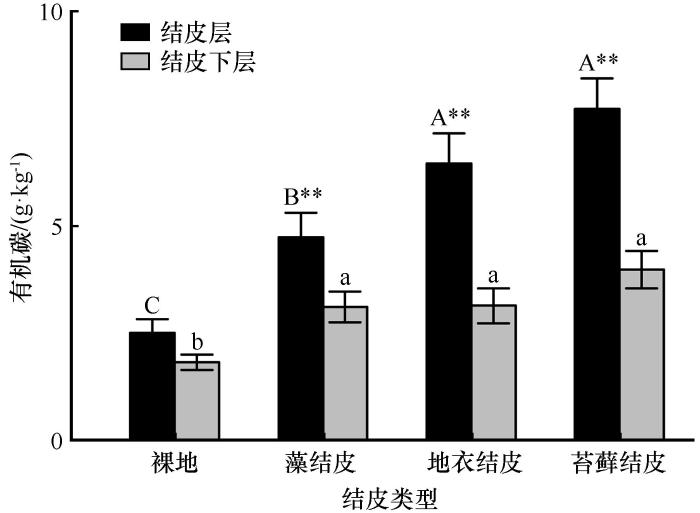
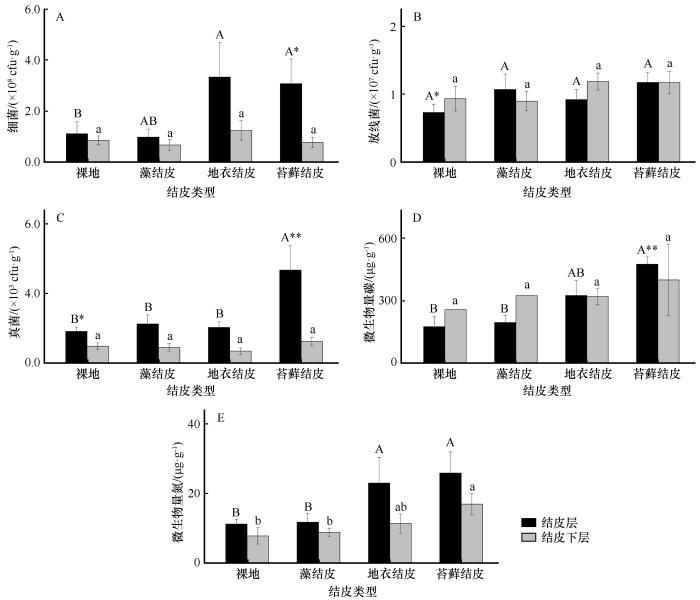
... 生物结皮在演替过程中,物种组成发生明显的变化,裸地阶段光合微生物数量极少,其固定的SOC含量最低,当发育到藻结皮时,结皮中有丰富的丝状蓝藻以及真核藻类,它们通过光合作用,向土壤中输入SOC;当生物结皮进一步发育到地衣结皮或苔藓结皮时,其光合固碳能力更强,即可向土壤中输入更多的SOC[25].诸多研究表明,随生物结皮的演替,其光合固碳能力显著增强,固定的SOC亦显著提高[4,26].本研究中MBC、MBN随生物结皮演替呈显著升高趋势,细菌与真菌数量在结皮演替后期也逐渐增加,是因为生物结皮演替过程中逐渐增加的SOC为异养细菌和真菌提供了能源物质,促进了微生物的生长和繁殖.在不同土壤层次中,结皮层SOC含量显著高于下层土壤已成研究共识[9,27],本研究中,氮素含量、氮转化相关酶活性和微生物数量在结皮层和结皮下层土壤中多数具有明显的差异,主要是结皮层和结皮下层积累的SOC巨大差异所致. ...
Responses of microbial activities and soil physical-chemical properties to the successional process of biological soil crusts in the Gurbantunggut Desert,Xinjiang
2
2015
... 生物土壤结皮(简称“生物结皮”)是干旱区生态系统的特殊地被层,由细菌、蓝藻、真核藻类、真菌、地衣和苔藓与土壤颗粒胶结而成[1].生物结皮在养分积累、水分保持、减少风蚀水蚀、生物地球化学循环、促进植被演替等过程中发挥着重要的生态功能[2-3].根据生物结皮的优势物种,可将生物结皮分为藻结皮、地衣结皮和苔藓结皮,这种不同类型反映了生物结皮的演替趋势[4].研究普遍认为,生物结皮的发育影响了土壤理化环境和生物学特性,尤其是显著提高了土壤养分含量和微生物活性[5].生物结皮不仅在荒漠生态系统广泛分布,在中国黄土高原植被间隙中亦普遍存在,尤其是近年来随着退耕还林还草工程的实施,大面积退耕撂荒地进入人为扰动减少、植被逐渐恢复的过程,土壤有机碳含量和有效养分增加,促使生物结皮广泛发育,并在生态恢复中扮演着不可忽视的角色[6]. ...
... 已有研究表明,生物结皮不仅能显著提高表层土壤有机质和土壤养分的含量,也可显著提高碳、氮、磷等元素相关的土壤酶活性[5,15-16].在黄土高原水蚀风蚀交错区,生物结皮层的土壤碱性磷酸酶、脲酶和过氧化氢酶活性明显高于结皮下层[17];黄绵土和风沙土藓结皮中的蔗糖酶、碱性磷酸酶、脲酶和蛋白酶活性显著高于无结皮土壤[18].土壤中氮素转化过程复杂,土壤酶在其生物化学转化过程中发挥着重要的催化功能,如矿化过程、固氮过程、硝化与反硝化过程,土壤酶能够反映土壤中生化过程作用的强度和方向,可表征土壤肥力水平和养分代谢能力等[19].土壤微生物是营养元素生物化学转化的重要驱动者,也是土壤的活性养分库,土壤微生物因其极强的适应性和生存力在生物结皮演替的各个阶段都起着促进作用[20].然而,黄土高原水蚀风蚀交错区有关生物结皮对氮转化过程的相关酶活性和微生物数量的影响规律尚不清楚.鉴于此,本文拟对黄土高原水蚀风蚀交错区六道沟小流域的不同生物结皮类型中氮素含量、氮转化相关酶活性、微生物数量开展研究,以期揭示黄土高原水蚀风蚀交错区氮素转化对生物结皮演替的响应规律,为认识生物结皮的生态功能提供科学依据. ...
Biological soil crusts decrease erodibility by modifying inherent soil properties on the Loess Plateau,China
1
2017
... 生物土壤结皮(简称“生物结皮”)是干旱区生态系统的特殊地被层,由细菌、蓝藻、真核藻类、真菌、地衣和苔藓与土壤颗粒胶结而成[1].生物结皮在养分积累、水分保持、减少风蚀水蚀、生物地球化学循环、促进植被演替等过程中发挥着重要的生态功能[2-3].根据生物结皮的优势物种,可将生物结皮分为藻结皮、地衣结皮和苔藓结皮,这种不同类型反映了生物结皮的演替趋势[4].研究普遍认为,生物结皮的发育影响了土壤理化环境和生物学特性,尤其是显著提高了土壤养分含量和微生物活性[5].生物结皮不仅在荒漠生态系统广泛分布,在中国黄土高原植被间隙中亦普遍存在,尤其是近年来随着退耕还林还草工程的实施,大面积退耕撂荒地进入人为扰动减少、植被逐渐恢复的过程,土壤有机碳含量和有效养分增加,促使生物结皮广泛发育,并在生态恢复中扮演着不可忽视的角色[6]. ...
Runoff and soil loss from revegetated grasslands in the hilsy loess plateau region,China:influence of biocrust patches and plant canopies
1
2013
... 黄土高原水蚀风蚀交错区位于黄土高原中北部,属于黄土高原向荒漠过渡、森林草原向典型干旱草原过渡的地带,也是黄河流域的生态屏障[7-8].该区域降雨主要在夏季,通常以暴雨形式出现,特殊的地貌、极不均匀的降雨分布以及单次暴雨的强度使该区受水蚀、风蚀的双重侵蚀[9].黄土高原严重的水土流失伴随着土壤有效养分的大量损失,尤其是氮素,极易随降雨形成的地表径流和侵蚀泥沙而流失[10-11].氮素是调节陆地生态系统结构和功能的关键元素,也是植物净生产力和土壤生态系统生产力的主要限制性元素[12].研究表明,随着降雨强度的增加,氮素损失总体呈上升趋势,由水土流失引起的淤泥总量决定其氮素损失量,可吸附氮和溶解氮损失分别占总氮损失的7%~82%(平均58.6%)和18%~93%(平均41.4%)[13].在黄土高原,氮素匮乏会限制植物的生长繁殖,进而影响生态系统功能[14]. ...
Evolution of ecosystem services in the Chinese Loess Plateau under climatic and land use changes
1
2013
... 黄土高原水蚀风蚀交错区位于黄土高原中北部,属于黄土高原向荒漠过渡、森林草原向典型干旱草原过渡的地带,也是黄河流域的生态屏障[7-8].该区域降雨主要在夏季,通常以暴雨形式出现,特殊的地貌、极不均匀的降雨分布以及单次暴雨的强度使该区受水蚀、风蚀的双重侵蚀[9].黄土高原严重的水土流失伴随着土壤有效养分的大量损失,尤其是氮素,极易随降雨形成的地表径流和侵蚀泥沙而流失[10-11].氮素是调节陆地生态系统结构和功能的关键元素,也是植物净生产力和土壤生态系统生产力的主要限制性元素[12].研究表明,随着降雨强度的增加,氮素损失总体呈上升趋势,由水土流失引起的淤泥总量决定其氮素损失量,可吸附氮和溶解氮损失分别占总氮损失的7%~82%(平均58.6%)和18%~93%(平均41.4%)[13].在黄土高原,氮素匮乏会限制植物的生长繁殖,进而影响生态系统功能[14]. ...
2
2022
... 黄土高原水蚀风蚀交错区位于黄土高原中北部,属于黄土高原向荒漠过渡、森林草原向典型干旱草原过渡的地带,也是黄河流域的生态屏障[7-8].该区域降雨主要在夏季,通常以暴雨形式出现,特殊的地貌、极不均匀的降雨分布以及单次暴雨的强度使该区受水蚀、风蚀的双重侵蚀[9].黄土高原严重的水土流失伴随着土壤有效养分的大量损失,尤其是氮素,极易随降雨形成的地表径流和侵蚀泥沙而流失[10-11].氮素是调节陆地生态系统结构和功能的关键元素,也是植物净生产力和土壤生态系统生产力的主要限制性元素[12].研究表明,随着降雨强度的增加,氮素损失总体呈上升趋势,由水土流失引起的淤泥总量决定其氮素损失量,可吸附氮和溶解氮损失分别占总氮损失的7%~82%(平均58.6%)和18%~93%(平均41.4%)[13].在黄土高原,氮素匮乏会限制植物的生长繁殖,进而影响生态系统功能[14]. ...
... 生物结皮在演替过程中,物种组成发生明显的变化,裸地阶段光合微生物数量极少,其固定的SOC含量最低,当发育到藻结皮时,结皮中有丰富的丝状蓝藻以及真核藻类,它们通过光合作用,向土壤中输入SOC;当生物结皮进一步发育到地衣结皮或苔藓结皮时,其光合固碳能力更强,即可向土壤中输入更多的SOC[25].诸多研究表明,随生物结皮的演替,其光合固碳能力显著增强,固定的SOC亦显著提高[4,26].本研究中MBC、MBN随生物结皮演替呈显著升高趋势,细菌与真菌数量在结皮演替后期也逐渐增加,是因为生物结皮演替过程中逐渐增加的SOC为异养细菌和真菌提供了能源物质,促进了微生物的生长和繁殖.在不同土壤层次中,结皮层SOC含量显著高于下层土壤已成研究共识[9,27],本研究中,氮素含量、氮转化相关酶活性和微生物数量在结皮层和结皮下层土壤中多数具有明显的差异,主要是结皮层和结皮下层积累的SOC巨大差异所致. ...
Role of soil erodibility in affecting available nitrogen and phosphorus losses under simulated rainfall
1
2014
... 黄土高原水蚀风蚀交错区位于黄土高原中北部,属于黄土高原向荒漠过渡、森林草原向典型干旱草原过渡的地带,也是黄河流域的生态屏障[7-8].该区域降雨主要在夏季,通常以暴雨形式出现,特殊的地貌、极不均匀的降雨分布以及单次暴雨的强度使该区受水蚀、风蚀的双重侵蚀[9].黄土高原严重的水土流失伴随着土壤有效养分的大量损失,尤其是氮素,极易随降雨形成的地表径流和侵蚀泥沙而流失[10-11].氮素是调节陆地生态系统结构和功能的关键元素,也是植物净生产力和土壤生态系统生产力的主要限制性元素[12].研究表明,随着降雨强度的增加,氮素损失总体呈上升趋势,由水土流失引起的淤泥总量决定其氮素损失量,可吸附氮和溶解氮损失分别占总氮损失的7%~82%(平均58.6%)和18%~93%(平均41.4%)[13].在黄土高原,氮素匮乏会限制植物的生长繁殖,进而影响生态系统功能[14]. ...
Assessing impacts of rainfall intensity and slope on dissolved and adsorbed nitrogen loss under bare loessial soil by simulated rainfalls
1
2018
... 黄土高原水蚀风蚀交错区位于黄土高原中北部,属于黄土高原向荒漠过渡、森林草原向典型干旱草原过渡的地带,也是黄河流域的生态屏障[7-8].该区域降雨主要在夏季,通常以暴雨形式出现,特殊的地貌、极不均匀的降雨分布以及单次暴雨的强度使该区受水蚀、风蚀的双重侵蚀[9].黄土高原严重的水土流失伴随着土壤有效养分的大量损失,尤其是氮素,极易随降雨形成的地表径流和侵蚀泥沙而流失[10-11].氮素是调节陆地生态系统结构和功能的关键元素,也是植物净生产力和土壤生态系统生产力的主要限制性元素[12].研究表明,随着降雨强度的增加,氮素损失总体呈上升趋势,由水土流失引起的淤泥总量决定其氮素损失量,可吸附氮和溶解氮损失分别占总氮损失的7%~82%(平均58.6%)和18%~93%(平均41.4%)[13].在黄土高原,氮素匮乏会限制植物的生长繁殖,进而影响生态系统功能[14]. ...
中国土壤氮素研究
1
2008
... 黄土高原水蚀风蚀交错区位于黄土高原中北部,属于黄土高原向荒漠过渡、森林草原向典型干旱草原过渡的地带,也是黄河流域的生态屏障[7-8].该区域降雨主要在夏季,通常以暴雨形式出现,特殊的地貌、极不均匀的降雨分布以及单次暴雨的强度使该区受水蚀、风蚀的双重侵蚀[9].黄土高原严重的水土流失伴随着土壤有效养分的大量损失,尤其是氮素,极易随降雨形成的地表径流和侵蚀泥沙而流失[10-11].氮素是调节陆地生态系统结构和功能的关键元素,也是植物净生产力和土壤生态系统生产力的主要限制性元素[12].研究表明,随着降雨强度的增加,氮素损失总体呈上升趋势,由水土流失引起的淤泥总量决定其氮素损失量,可吸附氮和溶解氮损失分别占总氮损失的7%~82%(平均58.6%)和18%~93%(平均41.4%)[13].在黄土高原,氮素匮乏会限制植物的生长繁殖,进而影响生态系统功能[14]. ...
Characteristics and numeric simulation of soil evaporation in biological soil crusts
1
2010
... 黄土高原水蚀风蚀交错区位于黄土高原中北部,属于黄土高原向荒漠过渡、森林草原向典型干旱草原过渡的地带,也是黄河流域的生态屏障[7-8].该区域降雨主要在夏季,通常以暴雨形式出现,特殊的地貌、极不均匀的降雨分布以及单次暴雨的强度使该区受水蚀、风蚀的双重侵蚀[9].黄土高原严重的水土流失伴随着土壤有效养分的大量损失,尤其是氮素,极易随降雨形成的地表径流和侵蚀泥沙而流失[10-11].氮素是调节陆地生态系统结构和功能的关键元素,也是植物净生产力和土壤生态系统生产力的主要限制性元素[12].研究表明,随着降雨强度的增加,氮素损失总体呈上升趋势,由水土流失引起的淤泥总量决定其氮素损失量,可吸附氮和溶解氮损失分别占总氮损失的7%~82%(平均58.6%)和18%~93%(平均41.4%)[13].在黄土高原,氮素匮乏会限制植物的生长繁殖,进而影响生态系统功能[14]. ...
晋西北黄土丘陵区不同植被恢复下的土壤碳氮累积特征
1
2021
... 黄土高原水蚀风蚀交错区位于黄土高原中北部,属于黄土高原向荒漠过渡、森林草原向典型干旱草原过渡的地带,也是黄河流域的生态屏障[7-8].该区域降雨主要在夏季,通常以暴雨形式出现,特殊的地貌、极不均匀的降雨分布以及单次暴雨的强度使该区受水蚀、风蚀的双重侵蚀[9].黄土高原严重的水土流失伴随着土壤有效养分的大量损失,尤其是氮素,极易随降雨形成的地表径流和侵蚀泥沙而流失[10-11].氮素是调节陆地生态系统结构和功能的关键元素,也是植物净生产力和土壤生态系统生产力的主要限制性元素[12].研究表明,随着降雨强度的增加,氮素损失总体呈上升趋势,由水土流失引起的淤泥总量决定其氮素损失量,可吸附氮和溶解氮损失分别占总氮损失的7%~82%(平均58.6%)和18%~93%(平均41.4%)[13].在黄土高原,氮素匮乏会限制植物的生长繁殖,进而影响生态系统功能[14]. ...
不同土壤和植被生境下生物结皮对土壤性质的影响
1
2019
... 已有研究表明,生物结皮不仅能显著提高表层土壤有机质和土壤养分的含量,也可显著提高碳、氮、磷等元素相关的土壤酶活性[5,15-16].在黄土高原水蚀风蚀交错区,生物结皮层的土壤碱性磷酸酶、脲酶和过氧化氢酶活性明显高于结皮下层[17];黄绵土和风沙土藓结皮中的蔗糖酶、碱性磷酸酶、脲酶和蛋白酶活性显著高于无结皮土壤[18].土壤中氮素转化过程复杂,土壤酶在其生物化学转化过程中发挥着重要的催化功能,如矿化过程、固氮过程、硝化与反硝化过程,土壤酶能够反映土壤中生化过程作用的强度和方向,可表征土壤肥力水平和养分代谢能力等[19].土壤微生物是营养元素生物化学转化的重要驱动者,也是土壤的活性养分库,土壤微生物因其极强的适应性和生存力在生物结皮演替的各个阶段都起着促进作用[20].然而,黄土高原水蚀风蚀交错区有关生物结皮对氮转化过程的相关酶活性和微生物数量的影响规律尚不清楚.鉴于此,本文拟对黄土高原水蚀风蚀交错区六道沟小流域的不同生物结皮类型中氮素含量、氮转化相关酶活性、微生物数量开展研究,以期揭示黄土高原水蚀风蚀交错区氮素转化对生物结皮演替的响应规律,为认识生物结皮的生态功能提供科学依据. ...
Impact of human trampling on biological soil crusts determined by soil microbial biomass,enzyme activities and nematode communities in a desert ecosystem
1
2018
... 已有研究表明,生物结皮不仅能显著提高表层土壤有机质和土壤养分的含量,也可显著提高碳、氮、磷等元素相关的土壤酶活性[5,15-16].在黄土高原水蚀风蚀交错区,生物结皮层的土壤碱性磷酸酶、脲酶和过氧化氢酶活性明显高于结皮下层[17];黄绵土和风沙土藓结皮中的蔗糖酶、碱性磷酸酶、脲酶和蛋白酶活性显著高于无结皮土壤[18].土壤中氮素转化过程复杂,土壤酶在其生物化学转化过程中发挥着重要的催化功能,如矿化过程、固氮过程、硝化与反硝化过程,土壤酶能够反映土壤中生化过程作用的强度和方向,可表征土壤肥力水平和养分代谢能力等[19].土壤微生物是营养元素生物化学转化的重要驱动者,也是土壤的活性养分库,土壤微生物因其极强的适应性和生存力在生物结皮演替的各个阶段都起着促进作用[20].然而,黄土高原水蚀风蚀交错区有关生物结皮对氮转化过程的相关酶活性和微生物数量的影响规律尚不清楚.鉴于此,本文拟对黄土高原水蚀风蚀交错区六道沟小流域的不同生物结皮类型中氮素含量、氮转化相关酶活性、微生物数量开展研究,以期揭示黄土高原水蚀风蚀交错区氮素转化对生物结皮演替的响应规律,为认识生物结皮的生态功能提供科学依据. ...
侵蚀条件下生物结皮对坡面土壤碳氮的影响
1
2011
... 已有研究表明,生物结皮不仅能显著提高表层土壤有机质和土壤养分的含量,也可显著提高碳、氮、磷等元素相关的土壤酶活性[5,15-16].在黄土高原水蚀风蚀交错区,生物结皮层的土壤碱性磷酸酶、脲酶和过氧化氢酶活性明显高于结皮下层[17];黄绵土和风沙土藓结皮中的蔗糖酶、碱性磷酸酶、脲酶和蛋白酶活性显著高于无结皮土壤[18].土壤中氮素转化过程复杂,土壤酶在其生物化学转化过程中发挥着重要的催化功能,如矿化过程、固氮过程、硝化与反硝化过程,土壤酶能够反映土壤中生化过程作用的强度和方向,可表征土壤肥力水平和养分代谢能力等[19].土壤微生物是营养元素生物化学转化的重要驱动者,也是土壤的活性养分库,土壤微生物因其极强的适应性和生存力在生物结皮演替的各个阶段都起着促进作用[20].然而,黄土高原水蚀风蚀交错区有关生物结皮对氮转化过程的相关酶活性和微生物数量的影响规律尚不清楚.鉴于此,本文拟对黄土高原水蚀风蚀交错区六道沟小流域的不同生物结皮类型中氮素含量、氮转化相关酶活性、微生物数量开展研究,以期揭示黄土高原水蚀风蚀交错区氮素转化对生物结皮演替的响应规律,为认识生物结皮的生态功能提供科学依据. ...
Vertical distribution of soil microbial biomass and its association with shrubs from the Negev Desert
1
2012
... 已有研究表明,生物结皮不仅能显著提高表层土壤有机质和土壤养分的含量,也可显著提高碳、氮、磷等元素相关的土壤酶活性[5,15-16].在黄土高原水蚀风蚀交错区,生物结皮层的土壤碱性磷酸酶、脲酶和过氧化氢酶活性明显高于结皮下层[17];黄绵土和风沙土藓结皮中的蔗糖酶、碱性磷酸酶、脲酶和蛋白酶活性显著高于无结皮土壤[18].土壤中氮素转化过程复杂,土壤酶在其生物化学转化过程中发挥着重要的催化功能,如矿化过程、固氮过程、硝化与反硝化过程,土壤酶能够反映土壤中生化过程作用的强度和方向,可表征土壤肥力水平和养分代谢能力等[19].土壤微生物是营养元素生物化学转化的重要驱动者,也是土壤的活性养分库,土壤微生物因其极强的适应性和生存力在生物结皮演替的各个阶段都起着促进作用[20].然而,黄土高原水蚀风蚀交错区有关生物结皮对氮转化过程的相关酶活性和微生物数量的影响规律尚不清楚.鉴于此,本文拟对黄土高原水蚀风蚀交错区六道沟小流域的不同生物结皮类型中氮素含量、氮转化相关酶活性、微生物数量开展研究,以期揭示黄土高原水蚀风蚀交错区氮素转化对生物结皮演替的响应规律,为认识生物结皮的生态功能提供科学依据. ...
3
2014
... 已有研究表明,生物结皮不仅能显著提高表层土壤有机质和土壤养分的含量,也可显著提高碳、氮、磷等元素相关的土壤酶活性[5,15-16].在黄土高原水蚀风蚀交错区,生物结皮层的土壤碱性磷酸酶、脲酶和过氧化氢酶活性明显高于结皮下层[17];黄绵土和风沙土藓结皮中的蔗糖酶、碱性磷酸酶、脲酶和蛋白酶活性显著高于无结皮土壤[18].土壤中氮素转化过程复杂,土壤酶在其生物化学转化过程中发挥着重要的催化功能,如矿化过程、固氮过程、硝化与反硝化过程,土壤酶能够反映土壤中生化过程作用的强度和方向,可表征土壤肥力水平和养分代谢能力等[19].土壤微生物是营养元素生物化学转化的重要驱动者,也是土壤的活性养分库,土壤微生物因其极强的适应性和生存力在生物结皮演替的各个阶段都起着促进作用[20].然而,黄土高原水蚀风蚀交错区有关生物结皮对氮转化过程的相关酶活性和微生物数量的影响规律尚不清楚.鉴于此,本文拟对黄土高原水蚀风蚀交错区六道沟小流域的不同生物结皮类型中氮素含量、氮转化相关酶活性、微生物数量开展研究,以期揭示黄土高原水蚀风蚀交错区氮素转化对生物结皮演替的响应规律,为认识生物结皮的生态功能提供科学依据. ...
... 土壤氮素的生物化学循环几乎全部依靠微生物介导的氧化还原反应完成[28-29].荒漠中藻结皮、地衣结皮和苔藓结皮的固氮速率差异显著,氮输入强烈依赖于生物结皮中的固氮微生物[30-31].本研究中,TN、NH-N、NO-N含量以及固氮酶活性随生物结皮演替其变化总体呈增加趋势,这在很大程度上取决于生物结皮固氮微生物的物种组成和丰度.在古尔班通古特沙漠中的相关研究表明,藻结皮和地衣结皮中固氮微生物以念珠藻属(Nostoc)和伪枝藻属(Syctonema)为主,且固氮微生物多样性与nifH基因丰度均在藻结皮阶段最高,其次为苔藓结皮[32];也有研究发现固氮蓝细菌是结皮演替早期主要的固氮功能群,随着生物结皮演替,地衣真菌(Leotiomycetes和Lecanoromycetes)及异养固氮菌(放线菌Actinomycetales、根瘤菌Rhizobiales等)成为地衣及苔藓结皮的主要固氮类群[33];而在浑善达克沙地,研究发现固氮微生物多样性指数随生物结皮的演替而上升,其中变形菌门的斯克尔曼氏菌(Skermanella)相对丰度显著增加,在苔藓结皮阶段成为仅次于未分类念珠藻科和伪枝藻属的第三优势固氮类群[34];这种固氮优势类群的差异可能与其多样性和群落组成相关.但在黄土高原水蚀风蚀交错区,生物结皮演替后期尤其是苔藓结皮的固氮酶活性最高,这可能是因为固氮微生物的组成在结皮演替后期发生了重要变化,还需进一步研究证实.在有机氮矿化过程中,有机氮常通过氨化作用被降解为NH[19],因此,NH-N含量在生物结皮后期演替阶段显著升高可能也与有机氮矿化逐渐升高有关.本研究发现黄土高原水蚀风蚀交错区生物结皮演替过程中脲酶和蛋白酶活性显著升高,在苔藓结皮或地衣结皮达到峰值.而脲酶和蛋白酶是蛋白质降解和尿素水解的重要酶类,在有机氮矿化过程中发挥着重要作用[35],说明有机氮的矿化作用随生物结皮演替也呈升高趋势.这种变化趋势可能有两方面原因,一是生物结皮的演替在后期积累了丰富的有机氮,为蛋白酶和脲酶提供了丰富的底物[1,25];二是生物结皮的演替使微生物种类和数量增加,尤其是与编码蛋白酶和脲酶(ureC)相关的功能微生物可能会增加,但有待于实验进一步验证.执行反硝化反应第一步的硝酸还原酶(编码基因narG)是控制反硝化过程的关键酶[36-37],在本研究中,硝酸还原酶在生物结皮演替过程中呈显著升高趋势,预示着硝酸还原酶相关的功能微生物丰度可能在结皮演替过程中有明显的升高. ...
... 生物结皮具有丰富的微生物物种、生态功能和生态多样性,是研究“生物多样性-功能”的一个模式系统[25,36].本文集中研究了黄土高原水蚀风蚀交错区生物结皮不同演替阶段氮素含量、氮素转化过程(固氮过程、矿化过程和反硝化过程)中相关酶活性以及微生物数量,但因方法的限制,未对硝化过程的相关酶活性进行探讨,而硝化作用是连接固氮过程、有机氮矿化和反硝化过程的纽带,通过与以上过程偶联,共同决定着土壤生态系统中氮的收支与归趋[19].氮循环功能微生物在土壤氮元素生物化学循环过程中发挥着驱动作用,近十几年来,分子生物学技术和宏基因组学的快速发展,在农田、森林、草地生态系统开展了大量研究工作,在很大程度上揭示了氮循环过程的作用机制[33,38-39].本文还未对影响这些过程的功能微生物多样性开展相关研究,文中分析判断依据是基于生物结皮微生物多样性的已有文献.因此,在研究区系统研究生物结皮氮素各个转化过程的耦合关系,并利用现代分子生物学技术将微生物功能群多样性与其生态功能相结合,将为揭示氮素转化过程的微生物学机制提供科学依据,也是生物结皮研究领域的重要拓展. ...
荒漠地表生物土壤结皮形成与演替特征概述
1
2010
... 已有研究表明,生物结皮不仅能显著提高表层土壤有机质和土壤养分的含量,也可显著提高碳、氮、磷等元素相关的土壤酶活性[5,15-16].在黄土高原水蚀风蚀交错区,生物结皮层的土壤碱性磷酸酶、脲酶和过氧化氢酶活性明显高于结皮下层[17];黄绵土和风沙土藓结皮中的蔗糖酶、碱性磷酸酶、脲酶和蛋白酶活性显著高于无结皮土壤[18].土壤中氮素转化过程复杂,土壤酶在其生物化学转化过程中发挥着重要的催化功能,如矿化过程、固氮过程、硝化与反硝化过程,土壤酶能够反映土壤中生化过程作用的强度和方向,可表征土壤肥力水平和养分代谢能力等[19].土壤微生物是营养元素生物化学转化的重要驱动者,也是土壤的活性养分库,土壤微生物因其极强的适应性和生存力在生物结皮演替的各个阶段都起着促进作用[20].然而,黄土高原水蚀风蚀交错区有关生物结皮对氮转化过程的相关酶活性和微生物数量的影响规律尚不清楚.鉴于此,本文拟对黄土高原水蚀风蚀交错区六道沟小流域的不同生物结皮类型中氮素含量、氮转化相关酶活性、微生物数量开展研究,以期揭示黄土高原水蚀风蚀交错区氮素转化对生物结皮演替的响应规律,为认识生物结皮的生态功能提供科学依据. ...
黄土高原水蚀风蚀交错区藓结皮覆盖土壤的蒸发特征
1
2020
... 研究区位于陕西省神木县六道沟小流域(38°46′—38°51′N,110°21′—110°23′E),属于典型的水蚀风蚀交错带.该区主体地貌由风沙地貌和黄土丘陵共同构成[21],属中温带半干旱气候,年均降水量408.5 mm,降水量80%以上发生在7—9月,多以暴雨形式出现,气候变化剧烈,水土流失严重,是典型的水蚀风蚀生态脆弱区,又因不合理的人为活动,植被退化,土地沙化严重[22]. ...
陕北水蚀风蚀交错区小流域苔藓结皮的空间特征及其影响因子
1
2014
... 研究区位于陕西省神木县六道沟小流域(38°46′—38°51′N,110°21′—110°23′E),属于典型的水蚀风蚀交错带.该区主体地貌由风沙地貌和黄土丘陵共同构成[21],属中温带半干旱气候,年均降水量408.5 mm,降水量80%以上发生在7—9月,多以暴雨形式出现,气候变化剧烈,水土流失严重,是典型的水蚀风蚀生态脆弱区,又因不合理的人为活动,植被退化,土地沙化严重[22]. ...
2
2008
... 土壤的有机碳(SOC)、总氮(TN)和pH值均采用风干土样,通过常规方法测定[23].土壤NH-N和NO-N、土壤含水量、微生物量和土壤酶活性的测定采用新鲜土样;土壤中的NH-N和NO-N通过2 mol·L-1的KCl溶液提取,并采用连续流动分析仪测定[23];微生物数量(真菌、细菌、放线菌)的测定采用稀释平板计数法;微生物量碳和氮(MBC、MBN)的测定采用氯仿熏蒸法[24];固氮酶活性采用乙炔还原法,脲酶活性采用靛酚比色法测定,蛋白酶采用茚三酮比色法测定,亚硝酸还原酶、硝酸还原酶采用酚二磺酸比色法测定[24]. ...
... [23];微生物数量(真菌、细菌、放线菌)的测定采用稀释平板计数法;微生物量碳和氮(MBC、MBN)的测定采用氯仿熏蒸法[24];固氮酶活性采用乙炔还原法,脲酶活性采用靛酚比色法测定,蛋白酶采用茚三酮比色法测定,亚硝酸还原酶、硝酸还原酶采用酚二磺酸比色法测定[24]. ...
2
2010
... 土壤的有机碳(SOC)、总氮(TN)和pH值均采用风干土样,通过常规方法测定[23].土壤NH-N和NO-N、土壤含水量、微生物量和土壤酶活性的测定采用新鲜土样;土壤中的NH-N和NO-N通过2 mol·L-1的KCl溶液提取,并采用连续流动分析仪测定[23];微生物数量(真菌、细菌、放线菌)的测定采用稀释平板计数法;微生物量碳和氮(MBC、MBN)的测定采用氯仿熏蒸法[24];固氮酶活性采用乙炔还原法,脲酶活性采用靛酚比色法测定,蛋白酶采用茚三酮比色法测定,亚硝酸还原酶、硝酸还原酶采用酚二磺酸比色法测定[24]. ...
... [24]. ...
3
2016
... 生物结皮在演替过程中,物种组成发生明显的变化,裸地阶段光合微生物数量极少,其固定的SOC含量最低,当发育到藻结皮时,结皮中有丰富的丝状蓝藻以及真核藻类,它们通过光合作用,向土壤中输入SOC;当生物结皮进一步发育到地衣结皮或苔藓结皮时,其光合固碳能力更强,即可向土壤中输入更多的SOC[25].诸多研究表明,随生物结皮的演替,其光合固碳能力显著增强,固定的SOC亦显著提高[4,26].本研究中MBC、MBN随生物结皮演替呈显著升高趋势,细菌与真菌数量在结皮演替后期也逐渐增加,是因为生物结皮演替过程中逐渐增加的SOC为异养细菌和真菌提供了能源物质,促进了微生物的生长和繁殖.在不同土壤层次中,结皮层SOC含量显著高于下层土壤已成研究共识[9,27],本研究中,氮素含量、氮转化相关酶活性和微生物数量在结皮层和结皮下层土壤中多数具有明显的差异,主要是结皮层和结皮下层积累的SOC巨大差异所致. ...
... 土壤氮素的生物化学循环几乎全部依靠微生物介导的氧化还原反应完成[28-29].荒漠中藻结皮、地衣结皮和苔藓结皮的固氮速率差异显著,氮输入强烈依赖于生物结皮中的固氮微生物[30-31].本研究中,TN、NH-N、NO-N含量以及固氮酶活性随生物结皮演替其变化总体呈增加趋势,这在很大程度上取决于生物结皮固氮微生物的物种组成和丰度.在古尔班通古特沙漠中的相关研究表明,藻结皮和地衣结皮中固氮微生物以念珠藻属(Nostoc)和伪枝藻属(Syctonema)为主,且固氮微生物多样性与nifH基因丰度均在藻结皮阶段最高,其次为苔藓结皮[32];也有研究发现固氮蓝细菌是结皮演替早期主要的固氮功能群,随着生物结皮演替,地衣真菌(Leotiomycetes和Lecanoromycetes)及异养固氮菌(放线菌Actinomycetales、根瘤菌Rhizobiales等)成为地衣及苔藓结皮的主要固氮类群[33];而在浑善达克沙地,研究发现固氮微生物多样性指数随生物结皮的演替而上升,其中变形菌门的斯克尔曼氏菌(Skermanella)相对丰度显著增加,在苔藓结皮阶段成为仅次于未分类念珠藻科和伪枝藻属的第三优势固氮类群[34];这种固氮优势类群的差异可能与其多样性和群落组成相关.但在黄土高原水蚀风蚀交错区,生物结皮演替后期尤其是苔藓结皮的固氮酶活性最高,这可能是因为固氮微生物的组成在结皮演替后期发生了重要变化,还需进一步研究证实.在有机氮矿化过程中,有机氮常通过氨化作用被降解为NH[19],因此,NH-N含量在生物结皮后期演替阶段显著升高可能也与有机氮矿化逐渐升高有关.本研究发现黄土高原水蚀风蚀交错区生物结皮演替过程中脲酶和蛋白酶活性显著升高,在苔藓结皮或地衣结皮达到峰值.而脲酶和蛋白酶是蛋白质降解和尿素水解的重要酶类,在有机氮矿化过程中发挥着重要作用[35],说明有机氮的矿化作用随生物结皮演替也呈升高趋势.这种变化趋势可能有两方面原因,一是生物结皮的演替在后期积累了丰富的有机氮,为蛋白酶和脲酶提供了丰富的底物[1,25];二是生物结皮的演替使微生物种类和数量增加,尤其是与编码蛋白酶和脲酶(ureC)相关的功能微生物可能会增加,但有待于实验进一步验证.执行反硝化反应第一步的硝酸还原酶(编码基因narG)是控制反硝化过程的关键酶[36-37],在本研究中,硝酸还原酶在生物结皮演替过程中呈显著升高趋势,预示着硝酸还原酶相关的功能微生物丰度可能在结皮演替过程中有明显的升高. ...
... 生物结皮具有丰富的微生物物种、生态功能和生态多样性,是研究“生物多样性-功能”的一个模式系统[25,36].本文集中研究了黄土高原水蚀风蚀交错区生物结皮不同演替阶段氮素含量、氮素转化过程(固氮过程、矿化过程和反硝化过程)中相关酶活性以及微生物数量,但因方法的限制,未对硝化过程的相关酶活性进行探讨,而硝化作用是连接固氮过程、有机氮矿化和反硝化过程的纽带,通过与以上过程偶联,共同决定着土壤生态系统中氮的收支与归趋[19].氮循环功能微生物在土壤氮元素生物化学循环过程中发挥着驱动作用,近十几年来,分子生物学技术和宏基因组学的快速发展,在农田、森林、草地生态系统开展了大量研究工作,在很大程度上揭示了氮循环过程的作用机制[33,38-39].本文还未对影响这些过程的功能微生物多样性开展相关研究,文中分析判断依据是基于生物结皮微生物多样性的已有文献.因此,在研究区系统研究生物结皮氮素各个转化过程的耦合关系,并利用现代分子生物学技术将微生物功能群多样性与其生态功能相结合,将为揭示氮素转化过程的微生物学机制提供科学依据,也是生物结皮研究领域的重要拓展. ...
The seasonal and successional variations of carbon release from biological soil crust-covered soil
1
2016
... 生物结皮在演替过程中,物种组成发生明显的变化,裸地阶段光合微生物数量极少,其固定的SOC含量最低,当发育到藻结皮时,结皮中有丰富的丝状蓝藻以及真核藻类,它们通过光合作用,向土壤中输入SOC;当生物结皮进一步发育到地衣结皮或苔藓结皮时,其光合固碳能力更强,即可向土壤中输入更多的SOC[25].诸多研究表明,随生物结皮的演替,其光合固碳能力显著增强,固定的SOC亦显著提高[4,26].本研究中MBC、MBN随生物结皮演替呈显著升高趋势,细菌与真菌数量在结皮演替后期也逐渐增加,是因为生物结皮演替过程中逐渐增加的SOC为异养细菌和真菌提供了能源物质,促进了微生物的生长和繁殖.在不同土壤层次中,结皮层SOC含量显著高于下层土壤已成研究共识[9,27],本研究中,氮素含量、氮转化相关酶活性和微生物数量在结皮层和结皮下层土壤中多数具有明显的差异,主要是结皮层和结皮下层积累的SOC巨大差异所致. ...
中国干旱沙区的生态重建与恢复:沙坡头站60年重要研究进展综述
1
2016
... 生物结皮在演替过程中,物种组成发生明显的变化,裸地阶段光合微生物数量极少,其固定的SOC含量最低,当发育到藻结皮时,结皮中有丰富的丝状蓝藻以及真核藻类,它们通过光合作用,向土壤中输入SOC;当生物结皮进一步发育到地衣结皮或苔藓结皮时,其光合固碳能力更强,即可向土壤中输入更多的SOC[25].诸多研究表明,随生物结皮的演替,其光合固碳能力显著增强,固定的SOC亦显著提高[4,26].本研究中MBC、MBN随生物结皮演替呈显著升高趋势,细菌与真菌数量在结皮演替后期也逐渐增加,是因为生物结皮演替过程中逐渐增加的SOC为异养细菌和真菌提供了能源物质,促进了微生物的生长和繁殖.在不同土壤层次中,结皮层SOC含量显著高于下层土壤已成研究共识[9,27],本研究中,氮素含量、氮转化相关酶活性和微生物数量在结皮层和结皮下层土壤中多数具有明显的差异,主要是结皮层和结皮下层积累的SOC巨大差异所致. ...
Species pool and local ecological assembly processes shape the β-diversity of diazotrophs in grassland soils
1
2021
... 土壤氮素的生物化学循环几乎全部依靠微生物介导的氧化还原反应完成[28-29].荒漠中藻结皮、地衣结皮和苔藓结皮的固氮速率差异显著,氮输入强烈依赖于生物结皮中的固氮微生物[30-31].本研究中,TN、NH-N、NO-N含量以及固氮酶活性随生物结皮演替其变化总体呈增加趋势,这在很大程度上取决于生物结皮固氮微生物的物种组成和丰度.在古尔班通古特沙漠中的相关研究表明,藻结皮和地衣结皮中固氮微生物以念珠藻属(Nostoc)和伪枝藻属(Syctonema)为主,且固氮微生物多样性与nifH基因丰度均在藻结皮阶段最高,其次为苔藓结皮[32];也有研究发现固氮蓝细菌是结皮演替早期主要的固氮功能群,随着生物结皮演替,地衣真菌(Leotiomycetes和Lecanoromycetes)及异养固氮菌(放线菌Actinomycetales、根瘤菌Rhizobiales等)成为地衣及苔藓结皮的主要固氮类群[33];而在浑善达克沙地,研究发现固氮微生物多样性指数随生物结皮的演替而上升,其中变形菌门的斯克尔曼氏菌(Skermanella)相对丰度显著增加,在苔藓结皮阶段成为仅次于未分类念珠藻科和伪枝藻属的第三优势固氮类群[34];这种固氮优势类群的差异可能与其多样性和群落组成相关.但在黄土高原水蚀风蚀交错区,生物结皮演替后期尤其是苔藓结皮的固氮酶活性最高,这可能是因为固氮微生物的组成在结皮演替后期发生了重要变化,还需进一步研究证实.在有机氮矿化过程中,有机氮常通过氨化作用被降解为NH[19],因此,NH-N含量在生物结皮后期演替阶段显著升高可能也与有机氮矿化逐渐升高有关.本研究发现黄土高原水蚀风蚀交错区生物结皮演替过程中脲酶和蛋白酶活性显著升高,在苔藓结皮或地衣结皮达到峰值.而脲酶和蛋白酶是蛋白质降解和尿素水解的重要酶类,在有机氮矿化过程中发挥着重要作用[35],说明有机氮的矿化作用随生物结皮演替也呈升高趋势.这种变化趋势可能有两方面原因,一是生物结皮的演替在后期积累了丰富的有机氮,为蛋白酶和脲酶提供了丰富的底物[1,25];二是生物结皮的演替使微生物种类和数量增加,尤其是与编码蛋白酶和脲酶(ureC)相关的功能微生物可能会增加,但有待于实验进一步验证.执行反硝化反应第一步的硝酸还原酶(编码基因narG)是控制反硝化过程的关键酶[36-37],在本研究中,硝酸还原酶在生物结皮演替过程中呈显著升高趋势,预示着硝酸还原酶相关的功能微生物丰度可能在结皮演替过程中有明显的升高. ...
宏基因组技术在氮循环功能微生物分子检测研究中的应用
1
2018
... 土壤氮素的生物化学循环几乎全部依靠微生物介导的氧化还原反应完成[28-29].荒漠中藻结皮、地衣结皮和苔藓结皮的固氮速率差异显著,氮输入强烈依赖于生物结皮中的固氮微生物[30-31].本研究中,TN、NH-N、NO-N含量以及固氮酶活性随生物结皮演替其变化总体呈增加趋势,这在很大程度上取决于生物结皮固氮微生物的物种组成和丰度.在古尔班通古特沙漠中的相关研究表明,藻结皮和地衣结皮中固氮微生物以念珠藻属(Nostoc)和伪枝藻属(Syctonema)为主,且固氮微生物多样性与nifH基因丰度均在藻结皮阶段最高,其次为苔藓结皮[32];也有研究发现固氮蓝细菌是结皮演替早期主要的固氮功能群,随着生物结皮演替,地衣真菌(Leotiomycetes和Lecanoromycetes)及异养固氮菌(放线菌Actinomycetales、根瘤菌Rhizobiales等)成为地衣及苔藓结皮的主要固氮类群[33];而在浑善达克沙地,研究发现固氮微生物多样性指数随生物结皮的演替而上升,其中变形菌门的斯克尔曼氏菌(Skermanella)相对丰度显著增加,在苔藓结皮阶段成为仅次于未分类念珠藻科和伪枝藻属的第三优势固氮类群[34];这种固氮优势类群的差异可能与其多样性和群落组成相关.但在黄土高原水蚀风蚀交错区,生物结皮演替后期尤其是苔藓结皮的固氮酶活性最高,这可能是因为固氮微生物的组成在结皮演替后期发生了重要变化,还需进一步研究证实.在有机氮矿化过程中,有机氮常通过氨化作用被降解为NH[19],因此,NH-N含量在生物结皮后期演替阶段显著升高可能也与有机氮矿化逐渐升高有关.本研究发现黄土高原水蚀风蚀交错区生物结皮演替过程中脲酶和蛋白酶活性显著升高,在苔藓结皮或地衣结皮达到峰值.而脲酶和蛋白酶是蛋白质降解和尿素水解的重要酶类,在有机氮矿化过程中发挥着重要作用[35],说明有机氮的矿化作用随生物结皮演替也呈升高趋势.这种变化趋势可能有两方面原因,一是生物结皮的演替在后期积累了丰富的有机氮,为蛋白酶和脲酶提供了丰富的底物[1,25];二是生物结皮的演替使微生物种类和数量增加,尤其是与编码蛋白酶和脲酶(ureC)相关的功能微生物可能会增加,但有待于实验进一步验证.执行反硝化反应第一步的硝酸还原酶(编码基因narG)是控制反硝化过程的关键酶[36-37],在本研究中,硝酸还原酶在生物结皮演替过程中呈显著升高趋势,预示着硝酸还原酶相关的功能微生物丰度可能在结皮演替过程中有明显的升高. ...
腾格里沙漠东南缘生物土壤结皮的固氮潜力
1
2012
... 土壤氮素的生物化学循环几乎全部依靠微生物介导的氧化还原反应完成[28-29].荒漠中藻结皮、地衣结皮和苔藓结皮的固氮速率差异显著,氮输入强烈依赖于生物结皮中的固氮微生物[30-31].本研究中,TN、NH-N、NO-N含量以及固氮酶活性随生物结皮演替其变化总体呈增加趋势,这在很大程度上取决于生物结皮固氮微生物的物种组成和丰度.在古尔班通古特沙漠中的相关研究表明,藻结皮和地衣结皮中固氮微生物以念珠藻属(Nostoc)和伪枝藻属(Syctonema)为主,且固氮微生物多样性与nifH基因丰度均在藻结皮阶段最高,其次为苔藓结皮[32];也有研究发现固氮蓝细菌是结皮演替早期主要的固氮功能群,随着生物结皮演替,地衣真菌(Leotiomycetes和Lecanoromycetes)及异养固氮菌(放线菌Actinomycetales、根瘤菌Rhizobiales等)成为地衣及苔藓结皮的主要固氮类群[33];而在浑善达克沙地,研究发现固氮微生物多样性指数随生物结皮的演替而上升,其中变形菌门的斯克尔曼氏菌(Skermanella)相对丰度显著增加,在苔藓结皮阶段成为仅次于未分类念珠藻科和伪枝藻属的第三优势固氮类群[34];这种固氮优势类群的差异可能与其多样性和群落组成相关.但在黄土高原水蚀风蚀交错区,生物结皮演替后期尤其是苔藓结皮的固氮酶活性最高,这可能是因为固氮微生物的组成在结皮演替后期发生了重要变化,还需进一步研究证实.在有机氮矿化过程中,有机氮常通过氨化作用被降解为NH[19],因此,NH-N含量在生物结皮后期演替阶段显著升高可能也与有机氮矿化逐渐升高有关.本研究发现黄土高原水蚀风蚀交错区生物结皮演替过程中脲酶和蛋白酶活性显著升高,在苔藓结皮或地衣结皮达到峰值.而脲酶和蛋白酶是蛋白质降解和尿素水解的重要酶类,在有机氮矿化过程中发挥着重要作用[35],说明有机氮的矿化作用随生物结皮演替也呈升高趋势.这种变化趋势可能有两方面原因,一是生物结皮的演替在后期积累了丰富的有机氮,为蛋白酶和脲酶提供了丰富的底物[1,25];二是生物结皮的演替使微生物种类和数量增加,尤其是与编码蛋白酶和脲酶(ureC)相关的功能微生物可能会增加,但有待于实验进一步验证.执行反硝化反应第一步的硝酸还原酶(编码基因narG)是控制反硝化过程的关键酶[36-37],在本研究中,硝酸还原酶在生物结皮演替过程中呈显著升高趋势,预示着硝酸还原酶相关的功能微生物丰度可能在结皮演替过程中有明显的升高. ...
Diversity of nitrogen-fixing microorganisms in biological soil crusts of copper mine wastelands
1
2014
... 土壤氮素的生物化学循环几乎全部依靠微生物介导的氧化还原反应完成[28-29].荒漠中藻结皮、地衣结皮和苔藓结皮的固氮速率差异显著,氮输入强烈依赖于生物结皮中的固氮微生物[30-31].本研究中,TN、NH-N、NO-N含量以及固氮酶活性随生物结皮演替其变化总体呈增加趋势,这在很大程度上取决于生物结皮固氮微生物的物种组成和丰度.在古尔班通古特沙漠中的相关研究表明,藻结皮和地衣结皮中固氮微生物以念珠藻属(Nostoc)和伪枝藻属(Syctonema)为主,且固氮微生物多样性与nifH基因丰度均在藻结皮阶段最高,其次为苔藓结皮[32];也有研究发现固氮蓝细菌是结皮演替早期主要的固氮功能群,随着生物结皮演替,地衣真菌(Leotiomycetes和Lecanoromycetes)及异养固氮菌(放线菌Actinomycetales、根瘤菌Rhizobiales等)成为地衣及苔藓结皮的主要固氮类群[33];而在浑善达克沙地,研究发现固氮微生物多样性指数随生物结皮的演替而上升,其中变形菌门的斯克尔曼氏菌(Skermanella)相对丰度显著增加,在苔藓结皮阶段成为仅次于未分类念珠藻科和伪枝藻属的第三优势固氮类群[34];这种固氮优势类群的差异可能与其多样性和群落组成相关.但在黄土高原水蚀风蚀交错区,生物结皮演替后期尤其是苔藓结皮的固氮酶活性最高,这可能是因为固氮微生物的组成在结皮演替后期发生了重要变化,还需进一步研究证实.在有机氮矿化过程中,有机氮常通过氨化作用被降解为NH[19],因此,NH-N含量在生物结皮后期演替阶段显著升高可能也与有机氮矿化逐渐升高有关.本研究发现黄土高原水蚀风蚀交错区生物结皮演替过程中脲酶和蛋白酶活性显著升高,在苔藓结皮或地衣结皮达到峰值.而脲酶和蛋白酶是蛋白质降解和尿素水解的重要酶类,在有机氮矿化过程中发挥着重要作用[35],说明有机氮的矿化作用随生物结皮演替也呈升高趋势.这种变化趋势可能有两方面原因,一是生物结皮的演替在后期积累了丰富的有机氮,为蛋白酶和脲酶提供了丰富的底物[1,25];二是生物结皮的演替使微生物种类和数量增加,尤其是与编码蛋白酶和脲酶(ureC)相关的功能微生物可能会增加,但有待于实验进一步验证.执行反硝化反应第一步的硝酸还原酶(编码基因narG)是控制反硝化过程的关键酶[36-37],在本研究中,硝酸还原酶在生物结皮演替过程中呈显著升高趋势,预示着硝酸还原酶相关的功能微生物丰度可能在结皮演替过程中有明显的升高. ...
Soil total organic carbon/total nitrogen ratio as a key driver deterministically shapes diazotrophic community assemblages during the succession of biological soil crusts
1
2021
... 土壤氮素的生物化学循环几乎全部依靠微生物介导的氧化还原反应完成[28-29].荒漠中藻结皮、地衣结皮和苔藓结皮的固氮速率差异显著,氮输入强烈依赖于生物结皮中的固氮微生物[30-31].本研究中,TN、NH-N、NO-N含量以及固氮酶活性随生物结皮演替其变化总体呈增加趋势,这在很大程度上取决于生物结皮固氮微生物的物种组成和丰度.在古尔班通古特沙漠中的相关研究表明,藻结皮和地衣结皮中固氮微生物以念珠藻属(Nostoc)和伪枝藻属(Syctonema)为主,且固氮微生物多样性与nifH基因丰度均在藻结皮阶段最高,其次为苔藓结皮[32];也有研究发现固氮蓝细菌是结皮演替早期主要的固氮功能群,随着生物结皮演替,地衣真菌(Leotiomycetes和Lecanoromycetes)及异养固氮菌(放线菌Actinomycetales、根瘤菌Rhizobiales等)成为地衣及苔藓结皮的主要固氮类群[33];而在浑善达克沙地,研究发现固氮微生物多样性指数随生物结皮的演替而上升,其中变形菌门的斯克尔曼氏菌(Skermanella)相对丰度显著增加,在苔藓结皮阶段成为仅次于未分类念珠藻科和伪枝藻属的第三优势固氮类群[34];这种固氮优势类群的差异可能与其多样性和群落组成相关.但在黄土高原水蚀风蚀交错区,生物结皮演替后期尤其是苔藓结皮的固氮酶活性最高,这可能是因为固氮微生物的组成在结皮演替后期发生了重要变化,还需进一步研究证实.在有机氮矿化过程中,有机氮常通过氨化作用被降解为NH[19],因此,NH-N含量在生物结皮后期演替阶段显著升高可能也与有机氮矿化逐渐升高有关.本研究发现黄土高原水蚀风蚀交错区生物结皮演替过程中脲酶和蛋白酶活性显著升高,在苔藓结皮或地衣结皮达到峰值.而脲酶和蛋白酶是蛋白质降解和尿素水解的重要酶类,在有机氮矿化过程中发挥着重要作用[35],说明有机氮的矿化作用随生物结皮演替也呈升高趋势.这种变化趋势可能有两方面原因,一是生物结皮的演替在后期积累了丰富的有机氮,为蛋白酶和脲酶提供了丰富的底物[1,25];二是生物结皮的演替使微生物种类和数量增加,尤其是与编码蛋白酶和脲酶(ureC)相关的功能微生物可能会增加,但有待于实验进一步验证.执行反硝化反应第一步的硝酸还原酶(编码基因narG)是控制反硝化过程的关键酶[36-37],在本研究中,硝酸还原酶在生物结皮演替过程中呈显著升高趋势,预示着硝酸还原酶相关的功能微生物丰度可能在结皮演替过程中有明显的升高. ...
The simulated N deposition accelerates net N mineralization and nitrification in a tropical forest soil
2
2019
... 土壤氮素的生物化学循环几乎全部依靠微生物介导的氧化还原反应完成[28-29].荒漠中藻结皮、地衣结皮和苔藓结皮的固氮速率差异显著,氮输入强烈依赖于生物结皮中的固氮微生物[30-31].本研究中,TN、NH-N、NO-N含量以及固氮酶活性随生物结皮演替其变化总体呈增加趋势,这在很大程度上取决于生物结皮固氮微生物的物种组成和丰度.在古尔班通古特沙漠中的相关研究表明,藻结皮和地衣结皮中固氮微生物以念珠藻属(Nostoc)和伪枝藻属(Syctonema)为主,且固氮微生物多样性与nifH基因丰度均在藻结皮阶段最高,其次为苔藓结皮[32];也有研究发现固氮蓝细菌是结皮演替早期主要的固氮功能群,随着生物结皮演替,地衣真菌(Leotiomycetes和Lecanoromycetes)及异养固氮菌(放线菌Actinomycetales、根瘤菌Rhizobiales等)成为地衣及苔藓结皮的主要固氮类群[33];而在浑善达克沙地,研究发现固氮微生物多样性指数随生物结皮的演替而上升,其中变形菌门的斯克尔曼氏菌(Skermanella)相对丰度显著增加,在苔藓结皮阶段成为仅次于未分类念珠藻科和伪枝藻属的第三优势固氮类群[34];这种固氮优势类群的差异可能与其多样性和群落组成相关.但在黄土高原水蚀风蚀交错区,生物结皮演替后期尤其是苔藓结皮的固氮酶活性最高,这可能是因为固氮微生物的组成在结皮演替后期发生了重要变化,还需进一步研究证实.在有机氮矿化过程中,有机氮常通过氨化作用被降解为NH[19],因此,NH-N含量在生物结皮后期演替阶段显著升高可能也与有机氮矿化逐渐升高有关.本研究发现黄土高原水蚀风蚀交错区生物结皮演替过程中脲酶和蛋白酶活性显著升高,在苔藓结皮或地衣结皮达到峰值.而脲酶和蛋白酶是蛋白质降解和尿素水解的重要酶类,在有机氮矿化过程中发挥着重要作用[35],说明有机氮的矿化作用随生物结皮演替也呈升高趋势.这种变化趋势可能有两方面原因,一是生物结皮的演替在后期积累了丰富的有机氮,为蛋白酶和脲酶提供了丰富的底物[1,25];二是生物结皮的演替使微生物种类和数量增加,尤其是与编码蛋白酶和脲酶(ureC)相关的功能微生物可能会增加,但有待于实验进一步验证.执行反硝化反应第一步的硝酸还原酶(编码基因narG)是控制反硝化过程的关键酶[36-37],在本研究中,硝酸还原酶在生物结皮演替过程中呈显著升高趋势,预示着硝酸还原酶相关的功能微生物丰度可能在结皮演替过程中有明显的升高. ...
... 生物结皮具有丰富的微生物物种、生态功能和生态多样性,是研究“生物多样性-功能”的一个模式系统[25,36].本文集中研究了黄土高原水蚀风蚀交错区生物结皮不同演替阶段氮素含量、氮素转化过程(固氮过程、矿化过程和反硝化过程)中相关酶活性以及微生物数量,但因方法的限制,未对硝化过程的相关酶活性进行探讨,而硝化作用是连接固氮过程、有机氮矿化和反硝化过程的纽带,通过与以上过程偶联,共同决定着土壤生态系统中氮的收支与归趋[19].氮循环功能微生物在土壤氮元素生物化学循环过程中发挥着驱动作用,近十几年来,分子生物学技术和宏基因组学的快速发展,在农田、森林、草地生态系统开展了大量研究工作,在很大程度上揭示了氮循环过程的作用机制[33,38-39].本文还未对影响这些过程的功能微生物多样性开展相关研究,文中分析判断依据是基于生物结皮微生物多样性的已有文献.因此,在研究区系统研究生物结皮氮素各个转化过程的耦合关系,并利用现代分子生物学技术将微生物功能群多样性与其生态功能相结合,将为揭示氮素转化过程的微生物学机制提供科学依据,也是生物结皮研究领域的重要拓展. ...
浑善达克沙地生物土壤结皮及其下层土壤中固氮细菌群落结构和多样性
1
2018
... 土壤氮素的生物化学循环几乎全部依靠微生物介导的氧化还原反应完成[28-29].荒漠中藻结皮、地衣结皮和苔藓结皮的固氮速率差异显著,氮输入强烈依赖于生物结皮中的固氮微生物[30-31].本研究中,TN、NH-N、NO-N含量以及固氮酶活性随生物结皮演替其变化总体呈增加趋势,这在很大程度上取决于生物结皮固氮微生物的物种组成和丰度.在古尔班通古特沙漠中的相关研究表明,藻结皮和地衣结皮中固氮微生物以念珠藻属(Nostoc)和伪枝藻属(Syctonema)为主,且固氮微生物多样性与nifH基因丰度均在藻结皮阶段最高,其次为苔藓结皮[32];也有研究发现固氮蓝细菌是结皮演替早期主要的固氮功能群,随着生物结皮演替,地衣真菌(Leotiomycetes和Lecanoromycetes)及异养固氮菌(放线菌Actinomycetales、根瘤菌Rhizobiales等)成为地衣及苔藓结皮的主要固氮类群[33];而在浑善达克沙地,研究发现固氮微生物多样性指数随生物结皮的演替而上升,其中变形菌门的斯克尔曼氏菌(Skermanella)相对丰度显著增加,在苔藓结皮阶段成为仅次于未分类念珠藻科和伪枝藻属的第三优势固氮类群[34];这种固氮优势类群的差异可能与其多样性和群落组成相关.但在黄土高原水蚀风蚀交错区,生物结皮演替后期尤其是苔藓结皮的固氮酶活性最高,这可能是因为固氮微生物的组成在结皮演替后期发生了重要变化,还需进一步研究证实.在有机氮矿化过程中,有机氮常通过氨化作用被降解为NH[19],因此,NH-N含量在生物结皮后期演替阶段显著升高可能也与有机氮矿化逐渐升高有关.本研究发现黄土高原水蚀风蚀交错区生物结皮演替过程中脲酶和蛋白酶活性显著升高,在苔藓结皮或地衣结皮达到峰值.而脲酶和蛋白酶是蛋白质降解和尿素水解的重要酶类,在有机氮矿化过程中发挥着重要作用[35],说明有机氮的矿化作用随生物结皮演替也呈升高趋势.这种变化趋势可能有两方面原因,一是生物结皮的演替在后期积累了丰富的有机氮,为蛋白酶和脲酶提供了丰富的底物[1,25];二是生物结皮的演替使微生物种类和数量增加,尤其是与编码蛋白酶和脲酶(ureC)相关的功能微生物可能会增加,但有待于实验进一步验证.执行反硝化反应第一步的硝酸还原酶(编码基因narG)是控制反硝化过程的关键酶[36-37],在本研究中,硝酸还原酶在生物结皮演替过程中呈显著升高趋势,预示着硝酸还原酶相关的功能微生物丰度可能在结皮演替过程中有明显的升高. ...
1
2006
... 土壤氮素的生物化学循环几乎全部依靠微生物介导的氧化还原反应完成[28-29].荒漠中藻结皮、地衣结皮和苔藓结皮的固氮速率差异显著,氮输入强烈依赖于生物结皮中的固氮微生物[30-31].本研究中,TN、NH-N、NO-N含量以及固氮酶活性随生物结皮演替其变化总体呈增加趋势,这在很大程度上取决于生物结皮固氮微生物的物种组成和丰度.在古尔班通古特沙漠中的相关研究表明,藻结皮和地衣结皮中固氮微生物以念珠藻属(Nostoc)和伪枝藻属(Syctonema)为主,且固氮微生物多样性与nifH基因丰度均在藻结皮阶段最高,其次为苔藓结皮[32];也有研究发现固氮蓝细菌是结皮演替早期主要的固氮功能群,随着生物结皮演替,地衣真菌(Leotiomycetes和Lecanoromycetes)及异养固氮菌(放线菌Actinomycetales、根瘤菌Rhizobiales等)成为地衣及苔藓结皮的主要固氮类群[33];而在浑善达克沙地,研究发现固氮微生物多样性指数随生物结皮的演替而上升,其中变形菌门的斯克尔曼氏菌(Skermanella)相对丰度显著增加,在苔藓结皮阶段成为仅次于未分类念珠藻科和伪枝藻属的第三优势固氮类群[34];这种固氮优势类群的差异可能与其多样性和群落组成相关.但在黄土高原水蚀风蚀交错区,生物结皮演替后期尤其是苔藓结皮的固氮酶活性最高,这可能是因为固氮微生物的组成在结皮演替后期发生了重要变化,还需进一步研究证实.在有机氮矿化过程中,有机氮常通过氨化作用被降解为NH[19],因此,NH-N含量在生物结皮后期演替阶段显著升高可能也与有机氮矿化逐渐升高有关.本研究发现黄土高原水蚀风蚀交错区生物结皮演替过程中脲酶和蛋白酶活性显著升高,在苔藓结皮或地衣结皮达到峰值.而脲酶和蛋白酶是蛋白质降解和尿素水解的重要酶类,在有机氮矿化过程中发挥着重要作用[35],说明有机氮的矿化作用随生物结皮演替也呈升高趋势.这种变化趋势可能有两方面原因,一是生物结皮的演替在后期积累了丰富的有机氮,为蛋白酶和脲酶提供了丰富的底物[1,25];二是生物结皮的演替使微生物种类和数量增加,尤其是与编码蛋白酶和脲酶(ureC)相关的功能微生物可能会增加,但有待于实验进一步验证.执行反硝化反应第一步的硝酸还原酶(编码基因narG)是控制反硝化过程的关键酶[36-37],在本研究中,硝酸还原酶在生物结皮演替过程中呈显著升高趋势,预示着硝酸还原酶相关的功能微生物丰度可能在结皮演替过程中有明显的升高. ...
Functional microarray analysis of nitrogen and carbon cycling genes across an Antarctic latitudinal transect
2
2007
... 土壤氮素的生物化学循环几乎全部依靠微生物介导的氧化还原反应完成[28-29].荒漠中藻结皮、地衣结皮和苔藓结皮的固氮速率差异显著,氮输入强烈依赖于生物结皮中的固氮微生物[30-31].本研究中,TN、NH-N、NO-N含量以及固氮酶活性随生物结皮演替其变化总体呈增加趋势,这在很大程度上取决于生物结皮固氮微生物的物种组成和丰度.在古尔班通古特沙漠中的相关研究表明,藻结皮和地衣结皮中固氮微生物以念珠藻属(Nostoc)和伪枝藻属(Syctonema)为主,且固氮微生物多样性与nifH基因丰度均在藻结皮阶段最高,其次为苔藓结皮[32];也有研究发现固氮蓝细菌是结皮演替早期主要的固氮功能群,随着生物结皮演替,地衣真菌(Leotiomycetes和Lecanoromycetes)及异养固氮菌(放线菌Actinomycetales、根瘤菌Rhizobiales等)成为地衣及苔藓结皮的主要固氮类群[33];而在浑善达克沙地,研究发现固氮微生物多样性指数随生物结皮的演替而上升,其中变形菌门的斯克尔曼氏菌(Skermanella)相对丰度显著增加,在苔藓结皮阶段成为仅次于未分类念珠藻科和伪枝藻属的第三优势固氮类群[34];这种固氮优势类群的差异可能与其多样性和群落组成相关.但在黄土高原水蚀风蚀交错区,生物结皮演替后期尤其是苔藓结皮的固氮酶活性最高,这可能是因为固氮微生物的组成在结皮演替后期发生了重要变化,还需进一步研究证实.在有机氮矿化过程中,有机氮常通过氨化作用被降解为NH[19],因此,NH-N含量在生物结皮后期演替阶段显著升高可能也与有机氮矿化逐渐升高有关.本研究发现黄土高原水蚀风蚀交错区生物结皮演替过程中脲酶和蛋白酶活性显著升高,在苔藓结皮或地衣结皮达到峰值.而脲酶和蛋白酶是蛋白质降解和尿素水解的重要酶类,在有机氮矿化过程中发挥着重要作用[35],说明有机氮的矿化作用随生物结皮演替也呈升高趋势.这种变化趋势可能有两方面原因,一是生物结皮的演替在后期积累了丰富的有机氮,为蛋白酶和脲酶提供了丰富的底物[1,25];二是生物结皮的演替使微生物种类和数量增加,尤其是与编码蛋白酶和脲酶(ureC)相关的功能微生物可能会增加,但有待于实验进一步验证.执行反硝化反应第一步的硝酸还原酶(编码基因narG)是控制反硝化过程的关键酶[36-37],在本研究中,硝酸还原酶在生物结皮演替过程中呈显著升高趋势,预示着硝酸还原酶相关的功能微生物丰度可能在结皮演替过程中有明显的升高. ...
... 生物结皮具有丰富的微生物物种、生态功能和生态多样性,是研究“生物多样性-功能”的一个模式系统[25,36].本文集中研究了黄土高原水蚀风蚀交错区生物结皮不同演替阶段氮素含量、氮素转化过程(固氮过程、矿化过程和反硝化过程)中相关酶活性以及微生物数量,但因方法的限制,未对硝化过程的相关酶活性进行探讨,而硝化作用是连接固氮过程、有机氮矿化和反硝化过程的纽带,通过与以上过程偶联,共同决定着土壤生态系统中氮的收支与归趋[19].氮循环功能微生物在土壤氮元素生物化学循环过程中发挥着驱动作用,近十几年来,分子生物学技术和宏基因组学的快速发展,在农田、森林、草地生态系统开展了大量研究工作,在很大程度上揭示了氮循环过程的作用机制[33,38-39].本文还未对影响这些过程的功能微生物多样性开展相关研究,文中分析判断依据是基于生物结皮微生物多样性的已有文献.因此,在研究区系统研究生物结皮氮素各个转化过程的耦合关系,并利用现代分子生物学技术将微生物功能群多样性与其生态功能相结合,将为揭示氮素转化过程的微生物学机制提供科学依据,也是生物结皮研究领域的重要拓展. ...
Changes in functional gene structure and metabolic potential of the microbial community in biological soil crusts along a revegetation chronosequence in the Tengger Desert
1
2018
... 土壤氮素的生物化学循环几乎全部依靠微生物介导的氧化还原反应完成[28-29].荒漠中藻结皮、地衣结皮和苔藓结皮的固氮速率差异显著,氮输入强烈依赖于生物结皮中的固氮微生物[30-31].本研究中,TN、NH-N、NO-N含量以及固氮酶活性随生物结皮演替其变化总体呈增加趋势,这在很大程度上取决于生物结皮固氮微生物的物种组成和丰度.在古尔班通古特沙漠中的相关研究表明,藻结皮和地衣结皮中固氮微生物以念珠藻属(Nostoc)和伪枝藻属(Syctonema)为主,且固氮微生物多样性与nifH基因丰度均在藻结皮阶段最高,其次为苔藓结皮[32];也有研究发现固氮蓝细菌是结皮演替早期主要的固氮功能群,随着生物结皮演替,地衣真菌(Leotiomycetes和Lecanoromycetes)及异养固氮菌(放线菌Actinomycetales、根瘤菌Rhizobiales等)成为地衣及苔藓结皮的主要固氮类群[33];而在浑善达克沙地,研究发现固氮微生物多样性指数随生物结皮的演替而上升,其中变形菌门的斯克尔曼氏菌(Skermanella)相对丰度显著增加,在苔藓结皮阶段成为仅次于未分类念珠藻科和伪枝藻属的第三优势固氮类群[34];这种固氮优势类群的差异可能与其多样性和群落组成相关.但在黄土高原水蚀风蚀交错区,生物结皮演替后期尤其是苔藓结皮的固氮酶活性最高,这可能是因为固氮微生物的组成在结皮演替后期发生了重要变化,还需进一步研究证实.在有机氮矿化过程中,有机氮常通过氨化作用被降解为NH[19],因此,NH-N含量在生物结皮后期演替阶段显著升高可能也与有机氮矿化逐渐升高有关.本研究发现黄土高原水蚀风蚀交错区生物结皮演替过程中脲酶和蛋白酶活性显著升高,在苔藓结皮或地衣结皮达到峰值.而脲酶和蛋白酶是蛋白质降解和尿素水解的重要酶类,在有机氮矿化过程中发挥着重要作用[35],说明有机氮的矿化作用随生物结皮演替也呈升高趋势.这种变化趋势可能有两方面原因,一是生物结皮的演替在后期积累了丰富的有机氮,为蛋白酶和脲酶提供了丰富的底物[1,25];二是生物结皮的演替使微生物种类和数量增加,尤其是与编码蛋白酶和脲酶(ureC)相关的功能微生物可能会增加,但有待于实验进一步验证.执行反硝化反应第一步的硝酸还原酶(编码基因narG)是控制反硝化过程的关键酶[36-37],在本研究中,硝酸还原酶在生物结皮演替过程中呈显著升高趋势,预示着硝酸还原酶相关的功能微生物丰度可能在结皮演替过程中有明显的升高. ...
Impact of long-term fertilization on the composition of denitrifier communities based on nitrite reductase analyses in a paddy soil
1
2010
... 生物结皮具有丰富的微生物物种、生态功能和生态多样性,是研究“生物多样性-功能”的一个模式系统[25,36].本文集中研究了黄土高原水蚀风蚀交错区生物结皮不同演替阶段氮素含量、氮素转化过程(固氮过程、矿化过程和反硝化过程)中相关酶活性以及微生物数量,但因方法的限制,未对硝化过程的相关酶活性进行探讨,而硝化作用是连接固氮过程、有机氮矿化和反硝化过程的纽带,通过与以上过程偶联,共同决定着土壤生态系统中氮的收支与归趋[19].氮循环功能微生物在土壤氮元素生物化学循环过程中发挥着驱动作用,近十几年来,分子生物学技术和宏基因组学的快速发展,在农田、森林、草地生态系统开展了大量研究工作,在很大程度上揭示了氮循环过程的作用机制[33,38-39].本文还未对影响这些过程的功能微生物多样性开展相关研究,文中分析判断依据是基于生物结皮微生物多样性的已有文献.因此,在研究区系统研究生物结皮氮素各个转化过程的耦合关系,并利用现代分子生物学技术将微生物功能群多样性与其生态功能相结合,将为揭示氮素转化过程的微生物学机制提供科学依据,也是生物结皮研究领域的重要拓展. ...
The microbial gene diversity along an elevation gradient of the Tibetan grassland
1
2014
... 生物结皮具有丰富的微生物物种、生态功能和生态多样性,是研究“生物多样性-功能”的一个模式系统[25,36].本文集中研究了黄土高原水蚀风蚀交错区生物结皮不同演替阶段氮素含量、氮素转化过程(固氮过程、矿化过程和反硝化过程)中相关酶活性以及微生物数量,但因方法的限制,未对硝化过程的相关酶活性进行探讨,而硝化作用是连接固氮过程、有机氮矿化和反硝化过程的纽带,通过与以上过程偶联,共同决定着土壤生态系统中氮的收支与归趋[19].氮循环功能微生物在土壤氮元素生物化学循环过程中发挥着驱动作用,近十几年来,分子生物学技术和宏基因组学的快速发展,在农田、森林、草地生态系统开展了大量研究工作,在很大程度上揭示了氮循环过程的作用机制[33,38-39].本文还未对影响这些过程的功能微生物多样性开展相关研究,文中分析判断依据是基于生物结皮微生物多样性的已有文献.因此,在研究区系统研究生物结皮氮素各个转化过程的耦合关系,并利用现代分子生物学技术将微生物功能群多样性与其生态功能相结合,将为揭示氮素转化过程的微生物学机制提供科学依据,也是生物结皮研究领域的重要拓展. ...




 甘公网安备 62010202000688号
甘公网安备 62010202000688号
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}