0 引言
气候变化是全球各国共同面临的巨大挑战[1 ] 。18世纪50年代以来,全球地表平均温度持续升高,在人类活动作用下,现代气候变暖速率是过去2 000年来前所未有的[2 ] 。气候变化改变了全球陆地生态系统,引发区域物种损失、疾病增加和大规模动植物死亡等不良现象,并且其实际影响远比预测更为深远[3 ] 。为准确研究全球变化的影响及后果,国际地圈与生物圈计划(International Geosphere-Biosphere Programme,IGBP)早已将全球变化与陆地生态系统关系作为研究重点[4 ] 。
干旱区作为陆地生态系统的重要组成,占据着全球约41%的陆地面积[5 ] ,具有降水稀缺、土壤贫瘠等特点[6 ] ,既是生态脆弱区,又是区域生态对气候变化的响应敏感区[7 ] 。中国是世界干旱区分布最多的国家之一[8 ] ,受全球变暖影响,中国的干旱化程度与范围均有加剧[9 -10 ] ,暖干化导致的植被覆盖度降低、水土流失等问题愈发突出[11 ] 。近年来,位于干旱半干旱区的黄河流域,气候暖干化和人口增长,水资源紧缺,干旱灾害的范围、频次和持续时间均有加剧,直接影响区域生态环境和社会经济发展[12 ] 。故针对这一问题,开展中国北方干旱半干旱区生态系统与气候变化的响应关系研究尤为重要。
植被是陆地生态系统的主体,可以指示生态与气候间的交互关系,也能反映干旱影响程度[9 ,13 ] ,因此可以将其作为气候变化与干旱区陆地生态系统研究的主要对象[14 ] 。现状植被类型与地带性分布是长期演替的结果,是对环境变化最准确、直接的反应[15 ] ,但现状植被常在人类活动干扰下,转化为栽培植被或被破坏发生逆行演替[16 ] ,因而使植被与气候间的真实响应关系被掩盖。“潜在自然植被”的概念排除人类活动与自然灾害等强烈扰动,仅考虑植被与自然因素(如气候、土壤、地形等)间响应关系,立地所形成的到达稳定阶段的顶极植被类型[17 ] 。潜在自然植被类型基于现状植被,但不一定完全一致,它是与立地环境达到平衡的演替终态[14 ] ,故采用潜在自然植被作为植被定义更为恰当,以真实地揭示自然状态下植被气候的相互作用关系,反映区域现状植被天然发展趋势。
Holdridge Life Zone(HLZ)模型是目前研究气候与植被生态系统定量关系的主流模型。该模型使用年平均生物温度、年降水量和潜在蒸散比率3个生物气候指标刻画各植被地带类型[18 -19 ] ,以其简明、适用而广泛应用于国内外气候植被模拟等研究领域。Zou等[20 ] 对黄土高原潜在植被的空间分布进行了模拟分析,范泽孟等[21 -22 ] 研究了中国生态过渡带类型及其分布的未来变化趋势与特征,揭示了气候变化驱动下欧亚大陆植被的时空变化模式及生态多样性,Fan等[23 ] 基于HASM方法内插气候数据分析了青藏高原不同高度带的潜在植被分布,Bourque[24 ] 结合DEM和气象数据识别了圣卢西亚岛的植被区划。
以上研究利用HLZ模型直接识别研究区植被类型,该方法判别潜在植被多缺乏与现状植被间关系的理论说明。此外,模型无法反映植被分布间高程差异的问题[25 ] 也未能解决,可能对地形多为高原山地、起伏明显的中国北方干旱区植被研究造成影响。鉴于此,如何划分与现状植被对应良好的潜在植被类型,构建适用于中国北方干旱半干旱区气候植被定量研究的模型,是厘清自然条件下干旱区生态系统主体与气候变化关系的核心。因此,论文旨在建立中国北方干旱半干旱区潜在植被分类体系,对HLZ模型进行修正、拓展,运用1981—2020年中国气象模拟数据,对潜在植被的空间分布格局进行模拟,揭示气候变化背景下干旱区潜在植被时空变化特征,以期为北方地区植被恢复、国家自然保护区建设等方面提供理论支撑。
1 数据及研究方法
1.1 研究区概况
研究区为北方干旱半干旱区所在省市区[8 ,26 ] ,31°36′—53°20′N、73°27′—126°20′E,包括新疆、内蒙古、甘肃、青海、宁夏、陕西、山西、河北和北京9省市区,总面积约451.5万km2 (图1 )。地势西高东低,横跨中国三级阶梯,地形以盆地、山地和高原为主,包括柴达木盆地、塔里木盆地、准噶尔盆地、天山山脉、祁连山、青藏高原、内蒙古高原和黄土高原等。区内温带东西部由于距海远近差异,年平均降水量跨度较大,东部可达500 mm,而西部多低于50 mm且气温日较差更大,反映了温带季风气候与温带大陆性气候差异。青藏高原区域年平均降水量约为260 mm,越靠近柴达木盆地越干旱,年平均气温多在0 ℃以下,气温日较差较大。自然景观丰富,森林、灌丛、草原、沼泽、荒漠等在区内广泛分布[27 ] 。
图1
图1
研究区高程及气象站点分布
Fig.1
Distribution of elevation and meteorological stations in study area
1.2 数据来源与处理
1.2.1 气候数据收集与处理
气象数据来源于国家青藏高原科学数据中心(http://data.tpdc.ac.cn/ )发布的中国1 km分辨率逐月平均气温、降水数据集,所选时间跨度为1981年1月至2020年12月。由于该数据集是根据CRU发布的全球0.5°气候数据集以及WorldClim发布的全球高分辨率气候数据集,通过Delta空间降尺度方案在中国地区降尺度生成的,故选用同期国家科学气象数据中心(http://data.cma.cn/ )发布的中国地面气候标准值月值数据集研究区范围内157个气象站点(图1 ),通过计算二者年平均降水量、年平均生物温度以检验近40年气象模拟数据精度。选用标准为站点包含近40年间所有月份的气象观测数据,不存在缺值或间断。最终检验结果可信(图2 )。
图2
图2
气象数据精度检验
Fig.2
Accuracy inspection of meteorological data
1.2.2 植被数据采集与分类
植被数据来源于国家青藏高原科学数据中心(http://data.tpdc.ac.cn/ )发布的1∶100万中国植被图,该数据可以准确详细地反映现状植被的空间分布。将研究区范围的植被图按植被型字段合并后,仅保留植被大类为自然植被的记录,再根据植被数据说明[16 ] 删除非气候顶级性质的植被型,以作为进一步划分潜在自然植被类型的基础,最终保留29种有效植被型。
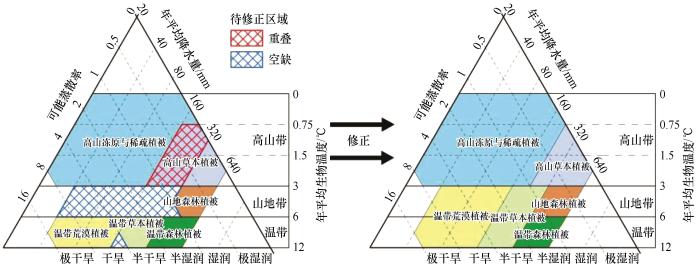
原HLZ模型无法反映植被分布间高程差异的问题主要体现在其根据年平均生物温度不同参数层级划分出不同的纬度带与垂直带,而两者在数值上具有等值性[25 ,28 ] 。等值性产生的问题会在生命地带即植被的命名上体现。命名方式为所属纬度带、垂直带以及植被类型的组合,其中纬度带需要根据当地年平均生物温度与海拔按标准气温直减率转换为海平面处的年平均生物温度进行判别[29 ] 。以研究区青藏高原植被命名为例,该地平均海拔为4 082 m,年平均生物温度为2.74 ℃,垂直带为高山带,根据标准气温直减率,海平面处的年平均生物温度高于24 ℃,纬度带为热带,故命名为热带高山植被。而实际中青藏高原北部不属于热带,该错误命名主要是由温度转换、纬度带与垂直带分级完全一致引起的,故原模型的植被命名法也不适用于潜在植被的命名,在命名上需建立合适的标准。
考虑不将年平均生物温度划分为两侧相同层级的纬度带与垂直带,而根据原划分方法合并为单侧温度带。由于研究区地形多为山地、高原,高程变化显著,故将原垂直带作为主要划分依据。鉴于研究区主体的温度带属于温带,故将原垂直带的基带(如山前地带)换为温带,以上仍记为山地带与高山带,这样可以反映温带中随着海拔升高年平均生物温度降低的特点。
拓展后的模型以干旱半干旱区潜在自然植被类型作为生命地带。为建立其与现状植被的紧密联系,根据潜在自然植被定义、现状植被说明[16 ] 及其修订案[30 ] ,直接从现状植被中合并潜在植被。潜在植被的分类与命名主要遵循“植物群落学-生态学”分类原则,即以植被的立地生态条件与外貌作为依据,表现形式为温度带与植被型组的组合,限定各植被型组的温度带也可防止不合理植被命名的发生;其次,通过实验与重要性比较,若植被型组不同的植被需要划入某类中,则根据面积占比大于75%的植被类型命名。研究区最终共划分出高山冻原与稀疏植被、高山草本植被、山地森林植被、温带荒漠植被、温带草本植被和温带森林植被6类潜在自然植被(表1 )。
其中,将垫状矮半灌木高寒荒漠纳入高山冻原与稀疏植被的依据为:第一,建群种与高山垫状植被相似,以垫型小半灌木为优势种;第二,与其他组成植被的分布位置接近,所处生态条件相似,其虽属于荒漠植被,但在年平均生物温度上与其他荒漠植被差异明显。将亚高山落叶阔叶灌丛与亚高山革质常绿阔叶灌丛纳入高山草本植被的依据为:第一,数据说明指出,两类灌丛植被与高寒草原草甸构成了“亚高山灌丛草甸带”;第二,它们在年平均生物温度和年平均降水量上无法分离,呈包含关系。将温带丛生矮禾草、矮半灌木荒漠草原纳入温带荒漠植被的依据为:第一,修订案指出,极端干旱环境中的该类植被不属于草本植被;第二,该类植被的年平均生物温度、可能蒸散率与温带草本植被差距过大,更接近温带荒漠植被。将温带落叶灌丛纳入温带森林植被的依据为:第一,数据说明指出,目前大多数灌丛为森林破坏后形成的次生植被,短时间内难以恢复成森林,成为相对稳定的植被类型。根据潜在自然植被定义,该类植被受人类活动影响前属于森林植被型组。第二,与其他组成植被的分布位置接近,所处生态条件相似。将亚热带森林植被亚型纳入温带森林植被的依据为:该类植被为非干旱区植被,本身面积极小且与其他温带森林植被混合分布,无法根据其年平均生物温度划分更高层的温度带。
1.3 研究方法
1.3.1 北方干旱半干旱区生命地带模型
HLZ模型以年降水量(P )、年平均生物温度(ABT )和可能蒸散率(PER )为气候指标,指标参数遵循二倍变化关系,通过取交集在一个各自成边的正三角形中划分若干代表不同植被类型的区域,即“生命地带”[18 ] ,可以反映生成该植被类型的水热阈值,故可通过建立现状植被与气候间的响应模式来反演区域潜在植被分布。
指标中,年降水量是一年降水量的总和,反映水分因素。ABT 是植物有效生长范围内的平均温度,为0~30 ℃,反映热量因素。可能蒸散率是年平均生物温度在一定比例下与年平均降水量的比率,反映干燥程度。三者的空间求算方程[21 -22 ] 如下:
P ( x , y , t ) = ∑ i = 1 12 P ( i , x , y , t ) (1)
A B T ( x , y , t ) = 1 / 12 × ∑ i = 1 12 T ( i , x , y , t ) (2)
P E R ( x , y , t ) = 58.93 × A B T ( x , y , t ) / P ( x , y , t ) (3)
式中:P x , y , t A B T x , y , t P E R x , y , t t x , y P i , x , y , t i T i , x , y , t i
假设3个指标的数值服从正态分布,可根据各潜在自然植被的指标平均值(μ σ μ ± σ μ ± 2 σ μ ± 3 σ 表2 )。根据表2 与指标参数二倍变化关系计算模拟值,要求模拟值在能划分出各类潜在植被对应的温度带基础上,尽量逼近一倍标准差阈值(表3 )。
根据表3 ,初步构建中国北方干旱半干旱区生命地带模型。由新模型可知,3 ℃以下形成了适宜高山植被的高山带,3~6 ℃形成了适宜山地植被的山地带,6~12 ℃形成了适宜温带植被的温带,3种温度带分界鲜明。而高山草本植被和高山冻原与稀疏植被在ABT 为0.75~3 ℃及年平均降水量160~320 mm出现重叠;山地带有大量植被空缺,使得温带植被与高山植被分布不连续;山地森林植被与温带草本植被受可能蒸散率限制,与相邻植被不连续。综上,该结果需要进行调整,具体方案如下:第一,高山草本植被的年平均降水量一般多于高山冻原与稀疏植被,为求均衡,取160 mm与320 mm的均值,即240 mm,将高山冻原与稀疏植被的年平均降水量上限下调至此处,将高山草本植被的下限上调至此处;第二,针对山地带的植被空缺,首先提取各潜在植被的高程平均值与标准差,将其他植被与山地森林植被比较,发现温带荒漠植被、温带草本植被与山地森林植被非常接近,说明这两类植被多数也分布于山地,而高山植被与山地森林植被的差距很大,故考虑将温带荒漠植被、温带草本植被的年平均生物温度下限下调至3 ℃,形成跨越两个温度带的生命地带;第三,由于可能蒸散率基于年平均降水量和年平均生物温度,若利用二者的模拟值计算可能蒸散率,则能发现它比表3 中的结果更宽泛,故可对其上下限进行小幅度调整,即将山地森林植被与温带草本植被的可能蒸散率上限分别上调至1.5与6(图3 )。最终实现参数修正,构建中国北方干旱半干旱区生命地带模型(表4 )。
图3
图3
中国北方干旱半干旱区生命地带模型
Fig.3
Life Zone Model of arid and semi-arid region of northern China
1.3.2 植被面积转移矩阵
转移矩阵可以反映同一事物在不同状态下其内部结构间的转换关系与变化方向[31 -32 ] 。通过建立模拟潜在植被与实际潜在植被(由现状植被模拟所得)间面积转移矩阵,作为模拟精度检验的基础,然后构建不同年代际间模拟潜在植被类型的面积转移矩阵,用以进一步分析不同时间段下各个植被类型的具体转换关系。该转移矩阵模型为:
A m n = A 11 A 12 ⋯ A 1 n A 21 A 22 ⋯ A 2 n ⋮ ⋮ ⋮ ⋮ A m 1 A m 2 ⋯ A m n (4)
式中:A m n m n A i j m i n j
1.3.3 Kappa 系数
Kappa系数被用来检验潜在自然植被模拟精度,通过在SPSS中导入模拟与实际潜在植被的面积转移矩阵计算Kappa系数。Kappa系数值一般为[0,1],根据值的大小可以划分5种级别的一致性,即0~0.2为极低一致性、0.2~0.4为一般一致性、0.4~0.6为中度一致性、0.6~0.8为高度一致性、0.8~1为极高一致性[33 ] 。其理论计算公式为:
K = N ∑ i n P i i - ∑ i n ( P i + P + i ) N 2 - ∑ i n ( P i + P + i ) (5)
式中:K n P i i i i Pi + 、P + i i i N
1.3.4 潜在植被空间分布平均中心模型
改进的平均中心模型[21 ,23 ] 被用于识别中国北方干旱半干旱区各类潜在自然植被在1981—1990、1991—2000、2001—2010年及2011—2020年4个时间段平均中心偏移特征,其理论计算公式为:
x j t = ∑ i = 1 I j ( t ) S i j ( t ) X i j ( t ) S j ( t ) (6)
y j t = ∑ i = 1 I j ( t ) S i j ( t ) Y i j ( t ) S j ( t ) (7)
式中:t I j ( t ) t j S i j ( t ) t j i S j ( t ) t j ( X i j ( t ) , Y i j ( t ) ) t j i ( X j ( t ) , Y j ( t ) ) t j
2 结果及分析
2.1 北方干旱半干旱区气候指标特征
2.1.1 时间变化特征
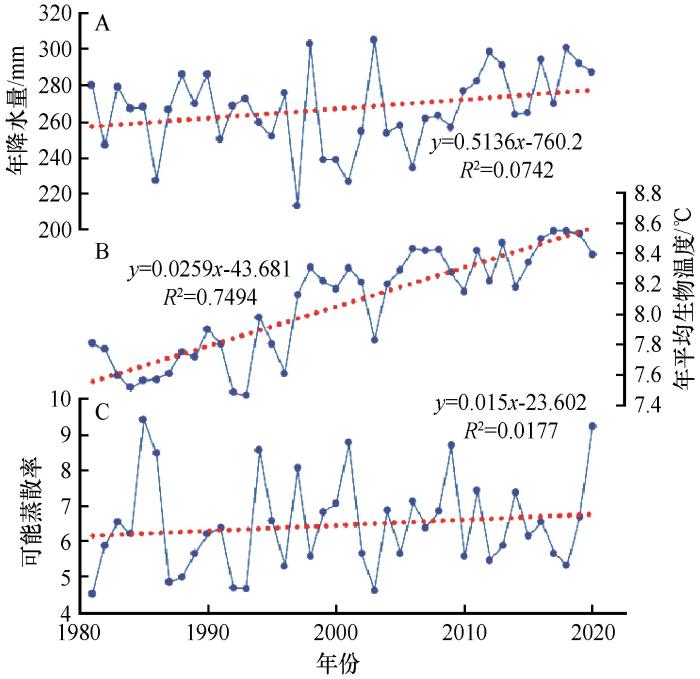
1981—2020年中国北方干旱半干旱区年降水量呈微弱上升趋势,上升速率为5.1 mm/10a,最大值为305.4 mm,最小值为213.3 mm,分别出现在2003年和1997年。降水的年际变化整体上很小,20世纪80年代与21世纪初降水微弱增加,90年代有小幅下降趋势(图4 )。
图4
图4
1981—2020年中国北方干旱半干旱区年降水量、年平均生物温度和可能蒸散率变化趋势
Fig.4
Trends of mean annual precipitation, mean annual bio-temperature and potential evapotranspiration rate in arid and semi-arid region of northern China from 1981 to 2010
年平均生物温度40年间呈微弱上升趋势,增长速率为0.3 ℃/10a,最大值为8.6 ℃,最小值为7.5 ℃,分别出现在2017年和1993年。年平均生物温度的年际变化整体上很小,20世纪80年代、90年代及21世纪初均呈上升趋势,其中90年代增长速率较快,高达0.8 ℃/10a。
可能蒸散率40年间呈微弱上升趋势,增长速率为0.2/10a,最大值为9.4,最小值为4.5,分别出现在1985年和1981年。可能蒸散率的年际变化整体上较小,20世纪80年代趋于水平,90年代与21世纪初均呈上升趋势,其中90年代增长速率较快,高达1.4/10a。
2.1.2 空间分布特征
1981—2020年中国北方干旱半干旱区年平均降水量、年平均生物温度和可能蒸散率在空间中变化幅度均较大。其中年平均降水量呈东南多、西北少的分布格局,最大值(1 226 mm)出现在陕西省安康市,低值出现在新疆南部、甘肃西北与内蒙古西部,具有明显的东南向西北方向阶梯式递减趋势,而在新疆西北的伊犁河谷一带又明显高于其他西北地区(图5 )。年平均生物温度主要与高程成反比,海拔越高则温度越低,昆仑山脉、祁连山、天山山脉、阿尔泰山脉等年平均生物温度多低于5 ℃,高值除研究区东南部外均出现在盆地区域。可能蒸散率西部明显高于东部,高值出现在吐鲁番盆地、哈密盆地、塔里木盆地和柴达木盆地,最高可达130,其他区域都低于6。
图5
图5
1981—2020年中国北方干旱半干旱区气候指标平均空间分布
Fig.5
Average spatial distribution of climate indicators in arid and semi-arid region of northern China from 1981 to 2010
2.2 北方干旱半干旱区潜在植被特征
2.2.1 空间分布特征
根据表4 模拟1981—2020年中国北方干旱半干旱区潜在植被的平均分布,通过建立现状植被与模拟植被的面积转移矩阵(表5 ),计算Kappa系数,所得值为0.505,表明模拟结果具有中度一致性,较令人满意,以此模拟1981—1990、1991—2000、2001—2010年和2011—2020年4个年代潜在自然植被分布(图6 )。模拟结果显示,不同年代际的各植被类型尽管在不同区域有所变动,但总体上所处地带不变,呈现一定规律性。其中,东部地区的潜在植被自东南向西北依次表现为温带森林植被、温带草本植被、温带荒漠植被,温带森林植被与温带草本植被呈西南向东北的条带状分布,温带荒漠植被呈东西向的条带状分布并延伸至天山山脉周边,大兴安岭西侧植被则以山地森林植被为主。青藏高原的潜在植被自东向西依次表现为山地森林植被、高山草本植被、高山冻原与稀疏植被,在柴达木盆地周围有少量温带荒漠植被与温带草本植被分布。准噶尔盆地的潜在植被主要为温带草本植被。各类潜在植被的分布基本与其背后的高程内涵相吻合,即高山植被分布于高原、山脉,山地植被分布于山地。
图6
图6
中国北方干旱半干旱区潜在自然植被各年代空间分布
Fig.6
Interdecadal spatial distribution of potential vegetation of arid and semi-arid region of northern China
2.2.2 分布面积与格局变化特征
对中国北方干旱半干旱区1980s、1990s、2000s及2010s潜在自然植被的模拟结果(图6 )进行面积统计(表6 ),结果表明,温带荒漠植被、温带草本植被、温带森林植被是研究区的主要植被类型,它们在各年代的面积基本均高于总面积的五分之一。其中,温带荒漠植被在1980s的占比为23.2%,随后在1990s、2000s和2010s出现持续下降,温带草本植被则呈相反变化趋势,前3个年代际面积占比持续增加,至2010s出现小幅度下降。温带森林植被除2000s面积均有增加,并在2010s达到最大面积占比28.5%。高山草本植被的面积也较多,占比均超过总面积的10%,于1990s出现微弱减少后在2000s有小幅增加,又于2010s出现微弱减少。高山冻原与稀疏植被、山地森林植被的面积较小,占比均低于总面积的10%,主体均呈降低趋势。
通过建立1980s与1990s、1990s与2000s、2000s与2010s潜在自然植被面积转移矩阵(表7 ~9 ),进一步研究不同年代际间植被类型转换关系。1990s新增的高山冻原与稀疏植被基本由高山草本植被转换而来,反映局地降水的减少。高山草本植被与山地森林植被变动很小,两个年代际间二者存在少量相互转换关系,体现了局地温度的升降。新增的温带荒漠植被主要由温带草本植被转换而来,而温带草本植被的新增面积也主要由温带荒漠植被构成,其占总新增面积的57.1%,说明局地降水出现增减。温带森林植被与温带草本植被在两个年代际间也存在多数转变,温带草本植被占其总新增面积的40.1%,其又占温带草本植被总新增面积的32.3%。此外,山地森林植被是温带森林植被新增部分的主要组成,占总新增面积的59.9%,揭示了局地温度的升高。
由表8 可知,2000s的高山冻原与稀疏植被变动极小,新增面积不到1%。新增的高山草本植被主要由高山冻原与稀疏植被转换而来,反映局地降水的增加。山地森林植被变动较小,但相比前两个年代际间的新增面积有所增加,增幅均超过两倍。温带荒漠植被的新增面积仍是由温带草本植被构成,而它在新温带草本植被中的面积明显下降,仅占总新增面积的约22.1%。与之对应的是温带森林植被在新温带草本植被中的面积大幅上升,占总新增面积的约71.6%。山地森林植被与温带草本植被仍是新增温带森林植被的主要构成,不同在于温带草本植被的占比小幅增加,几乎与山地森林植被相等。
由表9 可知,2010s高山冻原与稀疏植被的新增面积主要仍由高山草本植被构成,但其中温带荒漠植被所占面积明显增加,体现局部气温的减少。高山草本植被的面积变动很小,类似于1980s与1990s间的变化。对于新增的山地森林植被,高山草本植被与温带草本植被仍是其主要构成,且转换程度十分接近1990s与2000s。新增温带荒漠植被的面积很少,高山冻原与稀疏植被和温带草本植被向其转换程度明显下降。相反,温带荒漠植被在新增温带草本植被中的占比大幅上升,约占93.9%,而温带森林植被的占比明显下降,不足1%。温带草本植被是新增温带森林植被的主要构成,但相比1990s与2000s,其转换程度明显增大,约占总新增面积的93.6%。
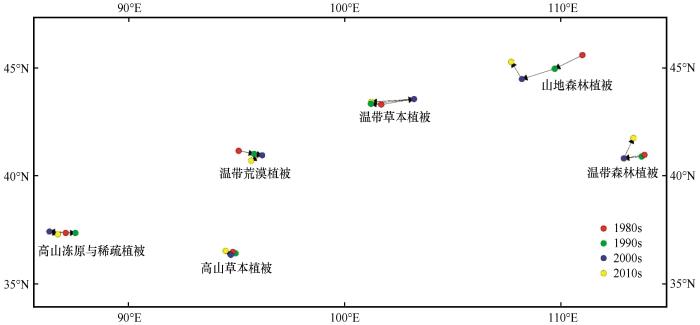
对中国北方干旱半干旱区3个年代际的各类潜在植被计算平均中心及其时空偏移(表10 ),并进行时空变化趋势制图(图7 ),发现1981—2020年平均中心总体偏移距离大于50 km的潜在植被共有3类,分别为山地森林植被、温带荒漠植被和温带森林植被。其中,山地森林植被偏移幅度尤为显著,40年偏移距离为235.9 km。除温带荒漠植被外,其他植被的平均中心总体均往西偏移。6类植被中,平均中心位于南部的偏移幅度总体小于北部,且越靠近南部,偏移幅度越小。1990s—2000s潜在植被偏移幅度整体大于其他时段,与同期干燥程度较快上升相对应。
图7
图7
各年代际潜在植被平均中心偏移趋势
Fig.7
Interdecadal shift trends of potential vegetation's mean center
3 结论与讨论
3.1 结论
基于1981—2020年中国逐月气象数据和现状植被数据,采用Holdridge生命地带模型指标探究中国北方干旱半干旱区气候特征,并根据修正后的模型定量分析了该气候条件下区域内潜在自然植被的时空变化特征。
研究期内年平均生物温度与可能蒸散率总体呈上升趋势,年降水量虽呈上升趋势,但研究区总体仍存在暖干化现象。东南地区至西北地区的年降水量变化明显,低海拔区域具有更高的年平均生物温度,盆地拥有较高的可能蒸散率。
研究区温度带分为高山带、山地带和温带,以高山冻原与稀疏植被、高山草本植被、山地森林植被、温带荒漠植被、温带草本植被和温带森林植被6类潜在自然植被为生命地带。其中,所属不同温度带的植被基本能体现高程差异。经检验,能较好地刻画和表征研究区气候与植被关系。
各潜在自然植被在1980s、1990s、2000s和2010s均具有相同的地带性分布规律。其中,温带荒漠植被、温带草本植被和温带森林植被是中国北方干旱半干旱区分布面积最多的植被类型,其在各年代际的总面积均占植被总面积的约70%,而高山冻原与稀疏植被的分布面积最少,平均低于植被总面积的9%。两个相邻年代际间,温带森林植被的面积平均增加最快,而温带荒漠植被减少最快。植被相邻年代际间的面积转换关系较为稳定,差别在于转换程度,温带草本植被的新增面积构成最为复杂。40年间除温带荒漠植被,其他植被的平均中心总体均往西移。
3.2 讨论
基于代表现状植被的1∶100万中国植被图,划分出6种北方干旱半干旱区潜在自然植被类型,再根据各类植被的气候指标阈值,构建适应研究区的生命地带模型的方法,相较于前人基于原模型和气象数据确认潜在植被,能更精确地反映无人类活动影响下研究区的植被类型,进而有助于真实地揭示气候与当地植被的响应关系。本研究于中国黄土高原西部(34°30′—36°30′N,104°—107°E、)模拟潜在植被,其中山地森林植被的空间平均分布与Zou等[20 ] 研究结果基本一致,均分布于该区域较高海拔处,而在他们的研究中,“草原”与本文温带森林植被的分布一致,这是由双方潜在自然植被分类的差异所造成,实际上这两类植被在气候指标阈值上是较为接近的,均体现了与周围植被类型的差异;于研究区西部模拟的潜在植被空间分布大致与修丽娜[34 ] 的研究结果一致,反映出植被的垂直地带性分布;于内蒙古西部模拟的温带草原植被自东向西向温带荒漠植被变化基本与李飞等[7 ] 研究结果基本一致,反映出植被的经向地带性变化。
在中国北方干旱半干旱区潜在自然植被分类体系中并未出现独立的灌丛植被类型,这是由于划分的植被是非过渡性质的,而现状灌丛植被多因极端干旱或寒冷并不典型,与其他植被形成过渡带,致使其被周围生态幅更宽植被在气候指标阈值上所掩盖,这点在相关研究[32 ,34 -36 ] 中也有体现。
中国北方干旱半干旱区生命地带模型指标模拟值的划分要求遵循二倍变化关系,且尽量逼近参考值。当参考值较小时,如最小参考值为0.15,是否也要逼近该参考值则需要进行考量,因为尝试逼近可能使层级增加2层以上,而过多的层级会使生命地带不连续或重叠的区域增加,对进一步修正造成困难,增加人为误差。因此使用该方法划分时,也需要忽略逼近较小的模拟值,以减少模拟值层数,如年平均生物温度0.5 ℃以下的多级划分通常没有意义。此外,在潜在自然植被的空间分布中,根据东南部水热条件,理应存在植被,而模拟结果却不存在。这是由于当地多为栽培植被,在处理时并未考虑当地植被转化为栽培植被前的潜在植被类型,而是直接删除,并未读取当地的气候指标阈值,致使东南部更高的水热条件下无植被分布。虽然东南部潜在植被已是半湿润区范畴,并不影响研究结果,但后续研究应充分考虑栽培植被区域的潜在植被类型,引入生态过渡带等概念划分出更全面的植被分类体系,实现更准确的区域植被模拟。
参考文献
View Option
[1]
魏一鸣 ,袁潇晨 ,吴刚 ,等 气候变化风险评估研究现状与热点:基于Web of Science的文献计量分析
[J].中国科学基金 ,2014 ,28 (5 ):347 -356 .
[本文引用: 1]
[2]
IPCC Climate Change 2021:The Physical Science Basis.Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change
[R].Cambridge,UK :Cambridge University Press ,2021 .
[本文引用: 1]
[3]
IPCC Climate Change 2022:Impacts, Adaptation, and Vulnerability.Contribution of Working Group II to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change
[R].Cambridge, UK : Cambridge University Press ,2022 .
[本文引用: 1]
[4]
倪健 植被-气候分类指标及其应用
[J].生态学杂志 ,1998 (5 ):34 -45 .
[本文引用: 1]
[6]
陈亚宁 ,李玉朋 ,李稚 ,等 全球气候变化对干旱区影响分析
[J].地球科学进展 ,2022 (2 ):111 -119 .
[本文引用: 1]
[7]
李飞 ,赵军 ,赵传燕 ,等 中国北方干旱半干旱区潜在植被演替
[J].生态学报 ,2011 ,31 (3 ):689 -697 .
[本文引用: 2]
[8]
陈曦 ,胡汝骥 ,姜逢清 ,等 中国干旱区自然地理 [M].北京 :科学出版社 ,2015 .
[本文引用: 2]
[9]
杨思遥 ,孟丹 ,李小娟 ,等 华北地区2001-2014年植被变化对SPEI气象干旱指数多尺度的响应
[J].生态学报 ,2018 ,38 (3 ):1028 -1039 .
[本文引用: 2]
[10]
安洁 ,付博 ,李玮 ,等 东亚地区典型极端气候指标未来预估及高温下人口暴露度研究
[J].北京大学学报(自然科学版) ,2020 ,56 (5 ):884 -892 .
[本文引用: 1]
[11]
杨舒畅 ,杨恒山 1982-2013年内蒙古地区干旱变化及植被响应
[J].自然灾害学报 ,2019 ,28 (1 ):175 -183 .
[本文引用: 1]
[12]
韩兰英 ,张强 ,马鹏里 ,等 气候变暖背景下黄河流域干旱灾害风险空间特征
[J].中国沙漠 ,2021 ,41 (4 ):225 -234 .
[本文引用: 1]
[13]
张华 ,徐存刚 ,王浩 2001-2018年西北地区植被变化对气象干旱的响应
[J].地理科学 ,2020 ,40 (6 ):1029 -1038 .
[本文引用: 1]
[14]
刘华民 ,吴绍洪 ,郑度 ,等 潜在自然植被研究与展望
[J].地理科学进展 ,2004 (1 ):62 -70 .
[本文引用: 2]
[15]
赵传燕 ,冯兆东 ,南忠仁 ,等 黄土高原祖厉河流域潜在植被分布模拟研究
[J].地理学报 ,2007 ,62 (1 ):52 -61 .
[本文引用: 1]
[16]
张新时 中国植被及其地理格局中华人民共和国植被图集(1∶100万)说明书(上卷) [M].北京 :地质出版社 ,2007 .
[本文引用: 3]
[17]
Tüxen R Die heutige potentielle natürliche vegetation als gegenstand der vegetationskartierung: mit 10 Tabellen
[J]. Berichte zur deutschen Landeskunde ,1956 ,13 .
[本文引用: 1]
[18]
Holdridge L R Determination of world plant formations from simple climatic data
[J].Science ,1947 ,105 (2727 ):367 .
[本文引用: 2]
[19]
Fan Z Fan B Shifts of the mean centers of potential vegetation ecosystems under future climate change in Eurasia
[J].Forests ,2019 ,10 (10 ):873 .
[本文引用: 1]
[20]
Zou S B Cheng G D Xiao H L et al Holocene natural rhythms of vegetation and present potential ecology in the Western Chinese Loess Plateau
[J].Quaternary International ,2009 ,194 :55 -67 .
[本文引用: 2]
[21]
范泽孟 中国生态过渡带分布的空间识别及情景模拟
[J].地理学报 ,2021 ,76 (3 ):626 -644 .
[本文引用: 3]
[22]
范泽孟 ,范斌 ,岳天祥 欧亚大陆植被生态系统潜在分布情景及其对气候变化的响应
[J].中国科学:地球科学 ,2019 ,49 (11 ):1817 -1830 .
[本文引用: 2]
[23]
Fan Z Bai X Scenarios of potential vegetation distribution in the different gradient zones of Qinghai-Tibet Plateau under future climate change
[J].Science of The Total Environment ,2021 ,796 (2 ):148918 .
[本文引用: 2]
[24]
Bourque P A Ecological life zones of Saint Lucia
.[J].Global Ecology & Biogeography ,2001 ,10 (5 ):549 -566 .
[本文引用: 1]
[25]
程姗岭 ,于海鹏 ,任钰 ,等 中国干旱半干旱区气候异常影响机理研究进展
[J].中国沙漠 ,2023 ,43 (3 ):21 -35 .
[本文引用: 2]
[26]
尹云鹤 ,马丹阳 ,邓浩宇 ,等 中国北方干湿过渡区生态系统生产力的气候变化风险评估
[J].地理学报 ,2021 ,76 (7 ):1605 -1617 .
[本文引用: 1]
[28]
张新时 研究全球变化的植被-气候分类系统
[J].第四纪研究 ,1993 (2 ):157 -169 .
[本文引用: 1]
[29]
MacMahon J A Wieboldt T F Applying biogeographic principles to resource management: a case study evaluating holdridge's life zone model
[J].Great Basin Naturalist Memoirs ,1978 (2 ):245 -257 .
[本文引用: 1]
[30]
郭柯 ,方精云 ,王国宏 ,等 中国植被分类系统修订方案
[J].植物生态学报 ,2020 ,44 (2 ):111 -127 .
[本文引用: 1]
[31]
高萌 基于Holdridge生命地带模型的我国南北过渡带潜在植被分布模拟
[D].兰州 :兰州大学 ,2019 .
[本文引用: 1]
[32]
李玉 ,牛路 ,赵泉华 抚顺矿区1989-2019年土地利用/覆盖变化分析
[J].测绘科学 ,2021 ,46 (8 ):96 -104 .
[本文引用: 2]
[33]
郝君明 中国Holdridge生命地带与潜在植被空间格局研究
[D].兰州 :西北师范大学 ,2009 .
[本文引用: 1]
[34]
修丽娜 基于CSCS模型的中国潜在自然植被时空分布特征研究
[D].兰州 :兰州大学 ,2014 .
[本文引用: 2]
[35]
车彦军 ,赵军 ,张明军 ,等 不同气候变化情景下2070-2099年中国潜在植被及其敏感性
[J].生态学报 ,2016 ,36 (10 ):2885 -2895 .
[36]
杜怀玉 ,赵军 ,师银芳 ,等 气候变化下中国潜在植被演替及其敏感性
[J].生态学杂志 ,2018 ,37 (5 ):1459 -1466 .
[本文引用: 1]
气候变化风险评估研究现状与热点:基于Web of Science的文献计量分析
1
2014
... 气候变化是全球各国共同面临的巨大挑战[1 ] .18世纪50年代以来,全球地表平均温度持续升高,在人类活动作用下,现代气候变暖速率是过去2 000年来前所未有的[2 ] .气候变化改变了全球陆地生态系统,引发区域物种损失、疾病增加和大规模动植物死亡等不良现象,并且其实际影响远比预测更为深远[3 ] .为准确研究全球变化的影响及后果,国际地圈与生物圈计划(International Geosphere-Biosphere Programme,IGBP)早已将全球变化与陆地生态系统关系作为研究重点[4 ] . ...
Climate Change 2021:The Physical Science Basis.Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change
1
2021
... 气候变化是全球各国共同面临的巨大挑战[1 ] .18世纪50年代以来,全球地表平均温度持续升高,在人类活动作用下,现代气候变暖速率是过去2 000年来前所未有的[2 ] .气候变化改变了全球陆地生态系统,引发区域物种损失、疾病增加和大规模动植物死亡等不良现象,并且其实际影响远比预测更为深远[3 ] .为准确研究全球变化的影响及后果,国际地圈与生物圈计划(International Geosphere-Biosphere Programme,IGBP)早已将全球变化与陆地生态系统关系作为研究重点[4 ] . ...
Climate Change 2022:Impacts, Adaptation, and Vulnerability.Contribution of Working Group II to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change
1
2022
... 气候变化是全球各国共同面临的巨大挑战[1 ] .18世纪50年代以来,全球地表平均温度持续升高,在人类活动作用下,现代气候变暖速率是过去2 000年来前所未有的[2 ] .气候变化改变了全球陆地生态系统,引发区域物种损失、疾病增加和大规模动植物死亡等不良现象,并且其实际影响远比预测更为深远[3 ] .为准确研究全球变化的影响及后果,国际地圈与生物圈计划(International Geosphere-Biosphere Programme,IGBP)早已将全球变化与陆地生态系统关系作为研究重点[4 ] . ...
植被-气候分类指标及其应用
1
1998
... 气候变化是全球各国共同面临的巨大挑战[1 ] .18世纪50年代以来,全球地表平均温度持续升高,在人类活动作用下,现代气候变暖速率是过去2 000年来前所未有的[2 ] .气候变化改变了全球陆地生态系统,引发区域物种损失、疾病增加和大规模动植物死亡等不良现象,并且其实际影响远比预测更为深远[3 ] .为准确研究全球变化的影响及后果,国际地圈与生物圈计划(International Geosphere-Biosphere Programme,IGBP)早已将全球变化与陆地生态系统关系作为研究重点[4 ] . ...
1
2021
... 干旱区作为陆地生态系统的重要组成,占据着全球约41%的陆地面积[5 ] ,具有降水稀缺、土壤贫瘠等特点[6 ] ,既是生态脆弱区,又是区域生态对气候变化的响应敏感区[7 ] .中国是世界干旱区分布最多的国家之一[8 ] ,受全球变暖影响,中国的干旱化程度与范围均有加剧[9 -10 ] ,暖干化导致的植被覆盖度降低、水土流失等问题愈发突出[11 ] .近年来,位于干旱半干旱区的黄河流域,气候暖干化和人口增长,水资源紧缺,干旱灾害的范围、频次和持续时间均有加剧,直接影响区域生态环境和社会经济发展[12 ] .故针对这一问题,开展中国北方干旱半干旱区生态系统与气候变化的响应关系研究尤为重要. ...
全球气候变化对干旱区影响分析
1
2022
... 干旱区作为陆地生态系统的重要组成,占据着全球约41%的陆地面积[5 ] ,具有降水稀缺、土壤贫瘠等特点[6 ] ,既是生态脆弱区,又是区域生态对气候变化的响应敏感区[7 ] .中国是世界干旱区分布最多的国家之一[8 ] ,受全球变暖影响,中国的干旱化程度与范围均有加剧[9 -10 ] ,暖干化导致的植被覆盖度降低、水土流失等问题愈发突出[11 ] .近年来,位于干旱半干旱区的黄河流域,气候暖干化和人口增长,水资源紧缺,干旱灾害的范围、频次和持续时间均有加剧,直接影响区域生态环境和社会经济发展[12 ] .故针对这一问题,开展中国北方干旱半干旱区生态系统与气候变化的响应关系研究尤为重要. ...
中国北方干旱半干旱区潜在植被演替
2
2011
... 干旱区作为陆地生态系统的重要组成,占据着全球约41%的陆地面积[5 ] ,具有降水稀缺、土壤贫瘠等特点[6 ] ,既是生态脆弱区,又是区域生态对气候变化的响应敏感区[7 ] .中国是世界干旱区分布最多的国家之一[8 ] ,受全球变暖影响,中国的干旱化程度与范围均有加剧[9 -10 ] ,暖干化导致的植被覆盖度降低、水土流失等问题愈发突出[11 ] .近年来,位于干旱半干旱区的黄河流域,气候暖干化和人口增长,水资源紧缺,干旱灾害的范围、频次和持续时间均有加剧,直接影响区域生态环境和社会经济发展[12 ] .故针对这一问题,开展中国北方干旱半干旱区生态系统与气候变化的响应关系研究尤为重要. ...
... 基于代表现状植被的1∶100万中国植被图,划分出6种北方干旱半干旱区潜在自然植被类型,再根据各类植被的气候指标阈值,构建适应研究区的生命地带模型的方法,相较于前人基于原模型和气象数据确认潜在植被,能更精确地反映无人类活动影响下研究区的植被类型,进而有助于真实地揭示气候与当地植被的响应关系.本研究于中国黄土高原西部(34°30′—36°30′N,104°—107°E、)模拟潜在植被,其中山地森林植被的空间平均分布与Zou等[20 ] 研究结果基本一致,均分布于该区域较高海拔处,而在他们的研究中,“草原”与本文温带森林植被的分布一致,这是由双方潜在自然植被分类的差异所造成,实际上这两类植被在气候指标阈值上是较为接近的,均体现了与周围植被类型的差异;于研究区西部模拟的潜在植被空间分布大致与修丽娜[34 ] 的研究结果一致,反映出植被的垂直地带性分布;于内蒙古西部模拟的温带草原植被自东向西向温带荒漠植被变化基本与李飞等[7 ] 研究结果基本一致,反映出植被的经向地带性变化. ...
2
2015
... 干旱区作为陆地生态系统的重要组成,占据着全球约41%的陆地面积[5 ] ,具有降水稀缺、土壤贫瘠等特点[6 ] ,既是生态脆弱区,又是区域生态对气候变化的响应敏感区[7 ] .中国是世界干旱区分布最多的国家之一[8 ] ,受全球变暖影响,中国的干旱化程度与范围均有加剧[9 -10 ] ,暖干化导致的植被覆盖度降低、水土流失等问题愈发突出[11 ] .近年来,位于干旱半干旱区的黄河流域,气候暖干化和人口增长,水资源紧缺,干旱灾害的范围、频次和持续时间均有加剧,直接影响区域生态环境和社会经济发展[12 ] .故针对这一问题,开展中国北方干旱半干旱区生态系统与气候变化的响应关系研究尤为重要. ...
... 研究区为北方干旱半干旱区所在省市区[8 ,26 ] ,31°36′—53°20′N、73°27′—126°20′E,包括新疆、内蒙古、甘肃、青海、宁夏、陕西、山西、河北和北京9省市区,总面积约451.5万km2 (图1 ).地势西高东低,横跨中国三级阶梯,地形以盆地、山地和高原为主,包括柴达木盆地、塔里木盆地、准噶尔盆地、天山山脉、祁连山、青藏高原、内蒙古高原和黄土高原等.区内温带东西部由于距海远近差异,年平均降水量跨度较大,东部可达500 mm,而西部多低于50 mm且气温日较差更大,反映了温带季风气候与温带大陆性气候差异.青藏高原区域年平均降水量约为260 mm,越靠近柴达木盆地越干旱,年平均气温多在0 ℃以下,气温日较差较大.自然景观丰富,森林、灌丛、草原、沼泽、荒漠等在区内广泛分布[27 ] . ...
华北地区2001-2014年植被变化对SPEI气象干旱指数多尺度的响应
2
2018
... 干旱区作为陆地生态系统的重要组成,占据着全球约41%的陆地面积[5 ] ,具有降水稀缺、土壤贫瘠等特点[6 ] ,既是生态脆弱区,又是区域生态对气候变化的响应敏感区[7 ] .中国是世界干旱区分布最多的国家之一[8 ] ,受全球变暖影响,中国的干旱化程度与范围均有加剧[9 -10 ] ,暖干化导致的植被覆盖度降低、水土流失等问题愈发突出[11 ] .近年来,位于干旱半干旱区的黄河流域,气候暖干化和人口增长,水资源紧缺,干旱灾害的范围、频次和持续时间均有加剧,直接影响区域生态环境和社会经济发展[12 ] .故针对这一问题,开展中国北方干旱半干旱区生态系统与气候变化的响应关系研究尤为重要. ...
... 植被是陆地生态系统的主体,可以指示生态与气候间的交互关系,也能反映干旱影响程度[9 ,13 ] ,因此可以将其作为气候变化与干旱区陆地生态系统研究的主要对象[14 ] .现状植被类型与地带性分布是长期演替的结果,是对环境变化最准确、直接的反应[15 ] ,但现状植被常在人类活动干扰下,转化为栽培植被或被破坏发生逆行演替[16 ] ,因而使植被与气候间的真实响应关系被掩盖.“潜在自然植被”的概念排除人类活动与自然灾害等强烈扰动,仅考虑植被与自然因素(如气候、土壤、地形等)间响应关系,立地所形成的到达稳定阶段的顶极植被类型[17 ] .潜在自然植被类型基于现状植被,但不一定完全一致,它是与立地环境达到平衡的演替终态[14 ] ,故采用潜在自然植被作为植被定义更为恰当,以真实地揭示自然状态下植被气候的相互作用关系,反映区域现状植被天然发展趋势. ...
东亚地区典型极端气候指标未来预估及高温下人口暴露度研究
1
2020
... 干旱区作为陆地生态系统的重要组成,占据着全球约41%的陆地面积[5 ] ,具有降水稀缺、土壤贫瘠等特点[6 ] ,既是生态脆弱区,又是区域生态对气候变化的响应敏感区[7 ] .中国是世界干旱区分布最多的国家之一[8 ] ,受全球变暖影响,中国的干旱化程度与范围均有加剧[9 -10 ] ,暖干化导致的植被覆盖度降低、水土流失等问题愈发突出[11 ] .近年来,位于干旱半干旱区的黄河流域,气候暖干化和人口增长,水资源紧缺,干旱灾害的范围、频次和持续时间均有加剧,直接影响区域生态环境和社会经济发展[12 ] .故针对这一问题,开展中国北方干旱半干旱区生态系统与气候变化的响应关系研究尤为重要. ...
1982-2013年内蒙古地区干旱变化及植被响应
1
2019
... 干旱区作为陆地生态系统的重要组成,占据着全球约41%的陆地面积[5 ] ,具有降水稀缺、土壤贫瘠等特点[6 ] ,既是生态脆弱区,又是区域生态对气候变化的响应敏感区[7 ] .中国是世界干旱区分布最多的国家之一[8 ] ,受全球变暖影响,中国的干旱化程度与范围均有加剧[9 -10 ] ,暖干化导致的植被覆盖度降低、水土流失等问题愈发突出[11 ] .近年来,位于干旱半干旱区的黄河流域,气候暖干化和人口增长,水资源紧缺,干旱灾害的范围、频次和持续时间均有加剧,直接影响区域生态环境和社会经济发展[12 ] .故针对这一问题,开展中国北方干旱半干旱区生态系统与气候变化的响应关系研究尤为重要. ...
气候变暖背景下黄河流域干旱灾害风险空间特征
1
2021
... 干旱区作为陆地生态系统的重要组成,占据着全球约41%的陆地面积[5 ] ,具有降水稀缺、土壤贫瘠等特点[6 ] ,既是生态脆弱区,又是区域生态对气候变化的响应敏感区[7 ] .中国是世界干旱区分布最多的国家之一[8 ] ,受全球变暖影响,中国的干旱化程度与范围均有加剧[9 -10 ] ,暖干化导致的植被覆盖度降低、水土流失等问题愈发突出[11 ] .近年来,位于干旱半干旱区的黄河流域,气候暖干化和人口增长,水资源紧缺,干旱灾害的范围、频次和持续时间均有加剧,直接影响区域生态环境和社会经济发展[12 ] .故针对这一问题,开展中国北方干旱半干旱区生态系统与气候变化的响应关系研究尤为重要. ...
2001-2018年西北地区植被变化对气象干旱的响应
1
2020
... 植被是陆地生态系统的主体,可以指示生态与气候间的交互关系,也能反映干旱影响程度[9 ,13 ] ,因此可以将其作为气候变化与干旱区陆地生态系统研究的主要对象[14 ] .现状植被类型与地带性分布是长期演替的结果,是对环境变化最准确、直接的反应[15 ] ,但现状植被常在人类活动干扰下,转化为栽培植被或被破坏发生逆行演替[16 ] ,因而使植被与气候间的真实响应关系被掩盖.“潜在自然植被”的概念排除人类活动与自然灾害等强烈扰动,仅考虑植被与自然因素(如气候、土壤、地形等)间响应关系,立地所形成的到达稳定阶段的顶极植被类型[17 ] .潜在自然植被类型基于现状植被,但不一定完全一致,它是与立地环境达到平衡的演替终态[14 ] ,故采用潜在自然植被作为植被定义更为恰当,以真实地揭示自然状态下植被气候的相互作用关系,反映区域现状植被天然发展趋势. ...
潜在自然植被研究与展望
2
2004
... 植被是陆地生态系统的主体,可以指示生态与气候间的交互关系,也能反映干旱影响程度[9 ,13 ] ,因此可以将其作为气候变化与干旱区陆地生态系统研究的主要对象[14 ] .现状植被类型与地带性分布是长期演替的结果,是对环境变化最准确、直接的反应[15 ] ,但现状植被常在人类活动干扰下,转化为栽培植被或被破坏发生逆行演替[16 ] ,因而使植被与气候间的真实响应关系被掩盖.“潜在自然植被”的概念排除人类活动与自然灾害等强烈扰动,仅考虑植被与自然因素(如气候、土壤、地形等)间响应关系,立地所形成的到达稳定阶段的顶极植被类型[17 ] .潜在自然植被类型基于现状植被,但不一定完全一致,它是与立地环境达到平衡的演替终态[14 ] ,故采用潜在自然植被作为植被定义更为恰当,以真实地揭示自然状态下植被气候的相互作用关系,反映区域现状植被天然发展趋势. ...
... [14 ],故采用潜在自然植被作为植被定义更为恰当,以真实地揭示自然状态下植被气候的相互作用关系,反映区域现状植被天然发展趋势. ...
黄土高原祖厉河流域潜在植被分布模拟研究
1
2007
... 植被是陆地生态系统的主体,可以指示生态与气候间的交互关系,也能反映干旱影响程度[9 ,13 ] ,因此可以将其作为气候变化与干旱区陆地生态系统研究的主要对象[14 ] .现状植被类型与地带性分布是长期演替的结果,是对环境变化最准确、直接的反应[15 ] ,但现状植被常在人类活动干扰下,转化为栽培植被或被破坏发生逆行演替[16 ] ,因而使植被与气候间的真实响应关系被掩盖.“潜在自然植被”的概念排除人类活动与自然灾害等强烈扰动,仅考虑植被与自然因素(如气候、土壤、地形等)间响应关系,立地所形成的到达稳定阶段的顶极植被类型[17 ] .潜在自然植被类型基于现状植被,但不一定完全一致,它是与立地环境达到平衡的演替终态[14 ] ,故采用潜在自然植被作为植被定义更为恰当,以真实地揭示自然状态下植被气候的相互作用关系,反映区域现状植被天然发展趋势. ...
3
2007
... 植被是陆地生态系统的主体,可以指示生态与气候间的交互关系,也能反映干旱影响程度[9 ,13 ] ,因此可以将其作为气候变化与干旱区陆地生态系统研究的主要对象[14 ] .现状植被类型与地带性分布是长期演替的结果,是对环境变化最准确、直接的反应[15 ] ,但现状植被常在人类活动干扰下,转化为栽培植被或被破坏发生逆行演替[16 ] ,因而使植被与气候间的真实响应关系被掩盖.“潜在自然植被”的概念排除人类活动与自然灾害等强烈扰动,仅考虑植被与自然因素(如气候、土壤、地形等)间响应关系,立地所形成的到达稳定阶段的顶极植被类型[17 ] .潜在自然植被类型基于现状植被,但不一定完全一致,它是与立地环境达到平衡的演替终态[14 ] ,故采用潜在自然植被作为植被定义更为恰当,以真实地揭示自然状态下植被气候的相互作用关系,反映区域现状植被天然发展趋势. ...
... 植被数据来源于国家青藏高原科学数据中心(http://data.tpdc.ac.cn/ )发布的1∶100万中国植被图,该数据可以准确详细地反映现状植被的空间分布.将研究区范围的植被图按植被型字段合并后,仅保留植被大类为自然植被的记录,再根据植被数据说明[16 ] 删除非气候顶级性质的植被型,以作为进一步划分潜在自然植被类型的基础,最终保留29种有效植被型. ...
... 拓展后的模型以干旱半干旱区潜在自然植被类型作为生命地带.为建立其与现状植被的紧密联系,根据潜在自然植被定义、现状植被说明[16 ] 及其修订案[30 ] ,直接从现状植被中合并潜在植被.潜在植被的分类与命名主要遵循“植物群落学-生态学”分类原则,即以植被的立地生态条件与外貌作为依据,表现形式为温度带与植被型组的组合,限定各植被型组的温度带也可防止不合理植被命名的发生;其次,通过实验与重要性比较,若植被型组不同的植被需要划入某类中,则根据面积占比大于75%的植被类型命名.研究区最终共划分出高山冻原与稀疏植被、高山草本植被、山地森林植被、温带荒漠植被、温带草本植被和温带森林植被6类潜在自然植被(表1 ). ...
Die heutige potentielle natürliche vegetation als gegenstand der vegetationskartierung: mit 10 Tabellen
1
1956
... 植被是陆地生态系统的主体,可以指示生态与气候间的交互关系,也能反映干旱影响程度[9 ,13 ] ,因此可以将其作为气候变化与干旱区陆地生态系统研究的主要对象[14 ] .现状植被类型与地带性分布是长期演替的结果,是对环境变化最准确、直接的反应[15 ] ,但现状植被常在人类活动干扰下,转化为栽培植被或被破坏发生逆行演替[16 ] ,因而使植被与气候间的真实响应关系被掩盖.“潜在自然植被”的概念排除人类活动与自然灾害等强烈扰动,仅考虑植被与自然因素(如气候、土壤、地形等)间响应关系,立地所形成的到达稳定阶段的顶极植被类型[17 ] .潜在自然植被类型基于现状植被,但不一定完全一致,它是与立地环境达到平衡的演替终态[14 ] ,故采用潜在自然植被作为植被定义更为恰当,以真实地揭示自然状态下植被气候的相互作用关系,反映区域现状植被天然发展趋势. ...
Determination of world plant formations from simple climatic data
2
1947
... Holdridge Life Zone(HLZ)模型是目前研究气候与植被生态系统定量关系的主流模型.该模型使用年平均生物温度、年降水量和潜在蒸散比率3个生物气候指标刻画各植被地带类型[18 -19 ] ,以其简明、适用而广泛应用于国内外气候植被模拟等研究领域.Zou等[20 ] 对黄土高原潜在植被的空间分布进行了模拟分析,范泽孟等[21 -22 ] 研究了中国生态过渡带类型及其分布的未来变化趋势与特征,揭示了气候变化驱动下欧亚大陆植被的时空变化模式及生态多样性,Fan等[23 ] 基于HASM方法内插气候数据分析了青藏高原不同高度带的潜在植被分布,Bourque[24 ] 结合DEM和气象数据识别了圣卢西亚岛的植被区划. ...
... HLZ模型以年降水量(P )、年平均生物温度(ABT )和可能蒸散率(PER )为气候指标,指标参数遵循二倍变化关系,通过取交集在一个各自成边的正三角形中划分若干代表不同植被类型的区域,即“生命地带”[18 ] ,可以反映生成该植被类型的水热阈值,故可通过建立现状植被与气候间的响应模式来反演区域潜在植被分布. ...
Shifts of the mean centers of potential vegetation ecosystems under future climate change in Eurasia
1
2019
... Holdridge Life Zone(HLZ)模型是目前研究气候与植被生态系统定量关系的主流模型.该模型使用年平均生物温度、年降水量和潜在蒸散比率3个生物气候指标刻画各植被地带类型[18 -19 ] ,以其简明、适用而广泛应用于国内外气候植被模拟等研究领域.Zou等[20 ] 对黄土高原潜在植被的空间分布进行了模拟分析,范泽孟等[21 -22 ] 研究了中国生态过渡带类型及其分布的未来变化趋势与特征,揭示了气候变化驱动下欧亚大陆植被的时空变化模式及生态多样性,Fan等[23 ] 基于HASM方法内插气候数据分析了青藏高原不同高度带的潜在植被分布,Bourque[24 ] 结合DEM和气象数据识别了圣卢西亚岛的植被区划. ...
Holocene natural rhythms of vegetation and present potential ecology in the Western Chinese Loess Plateau
2
2009
... Holdridge Life Zone(HLZ)模型是目前研究气候与植被生态系统定量关系的主流模型.该模型使用年平均生物温度、年降水量和潜在蒸散比率3个生物气候指标刻画各植被地带类型[18 -19 ] ,以其简明、适用而广泛应用于国内外气候植被模拟等研究领域.Zou等[20 ] 对黄土高原潜在植被的空间分布进行了模拟分析,范泽孟等[21 -22 ] 研究了中国生态过渡带类型及其分布的未来变化趋势与特征,揭示了气候变化驱动下欧亚大陆植被的时空变化模式及生态多样性,Fan等[23 ] 基于HASM方法内插气候数据分析了青藏高原不同高度带的潜在植被分布,Bourque[24 ] 结合DEM和气象数据识别了圣卢西亚岛的植被区划. ...
... 基于代表现状植被的1∶100万中国植被图,划分出6种北方干旱半干旱区潜在自然植被类型,再根据各类植被的气候指标阈值,构建适应研究区的生命地带模型的方法,相较于前人基于原模型和气象数据确认潜在植被,能更精确地反映无人类活动影响下研究区的植被类型,进而有助于真实地揭示气候与当地植被的响应关系.本研究于中国黄土高原西部(34°30′—36°30′N,104°—107°E、)模拟潜在植被,其中山地森林植被的空间平均分布与Zou等[20 ] 研究结果基本一致,均分布于该区域较高海拔处,而在他们的研究中,“草原”与本文温带森林植被的分布一致,这是由双方潜在自然植被分类的差异所造成,实际上这两类植被在气候指标阈值上是较为接近的,均体现了与周围植被类型的差异;于研究区西部模拟的潜在植被空间分布大致与修丽娜[34 ] 的研究结果一致,反映出植被的垂直地带性分布;于内蒙古西部模拟的温带草原植被自东向西向温带荒漠植被变化基本与李飞等[7 ] 研究结果基本一致,反映出植被的经向地带性变化. ...
中国生态过渡带分布的空间识别及情景模拟
3
2021
... Holdridge Life Zone(HLZ)模型是目前研究气候与植被生态系统定量关系的主流模型.该模型使用年平均生物温度、年降水量和潜在蒸散比率3个生物气候指标刻画各植被地带类型[18 -19 ] ,以其简明、适用而广泛应用于国内外气候植被模拟等研究领域.Zou等[20 ] 对黄土高原潜在植被的空间分布进行了模拟分析,范泽孟等[21 -22 ] 研究了中国生态过渡带类型及其分布的未来变化趋势与特征,揭示了气候变化驱动下欧亚大陆植被的时空变化模式及生态多样性,Fan等[23 ] 基于HASM方法内插气候数据分析了青藏高原不同高度带的潜在植被分布,Bourque[24 ] 结合DEM和气象数据识别了圣卢西亚岛的植被区划. ...
... 指标中,年降水量是一年降水量的总和,反映水分因素.ABT 是植物有效生长范围内的平均温度,为0~30 ℃,反映热量因素.可能蒸散率是年平均生物温度在一定比例下与年平均降水量的比率,反映干燥程度.三者的空间求算方程[21 -22 ] 如下: ...
... 改进的平均中心模型[21 ,23 ] 被用于识别中国北方干旱半干旱区各类潜在自然植被在1981—1990、1991—2000、2001—2010年及2011—2020年4个时间段平均中心偏移特征,其理论计算公式为: ...
欧亚大陆植被生态系统潜在分布情景及其对气候变化的响应
2
2019
... Holdridge Life Zone(HLZ)模型是目前研究气候与植被生态系统定量关系的主流模型.该模型使用年平均生物温度、年降水量和潜在蒸散比率3个生物气候指标刻画各植被地带类型[18 -19 ] ,以其简明、适用而广泛应用于国内外气候植被模拟等研究领域.Zou等[20 ] 对黄土高原潜在植被的空间分布进行了模拟分析,范泽孟等[21 -22 ] 研究了中国生态过渡带类型及其分布的未来变化趋势与特征,揭示了气候变化驱动下欧亚大陆植被的时空变化模式及生态多样性,Fan等[23 ] 基于HASM方法内插气候数据分析了青藏高原不同高度带的潜在植被分布,Bourque[24 ] 结合DEM和气象数据识别了圣卢西亚岛的植被区划. ...
... 指标中,年降水量是一年降水量的总和,反映水分因素.ABT 是植物有效生长范围内的平均温度,为0~30 ℃,反映热量因素.可能蒸散率是年平均生物温度在一定比例下与年平均降水量的比率,反映干燥程度.三者的空间求算方程[21 -22 ] 如下: ...
Scenarios of potential vegetation distribution in the different gradient zones of Qinghai-Tibet Plateau under future climate change
2
2021
... Holdridge Life Zone(HLZ)模型是目前研究气候与植被生态系统定量关系的主流模型.该模型使用年平均生物温度、年降水量和潜在蒸散比率3个生物气候指标刻画各植被地带类型[18 -19 ] ,以其简明、适用而广泛应用于国内外气候植被模拟等研究领域.Zou等[20 ] 对黄土高原潜在植被的空间分布进行了模拟分析,范泽孟等[21 -22 ] 研究了中国生态过渡带类型及其分布的未来变化趋势与特征,揭示了气候变化驱动下欧亚大陆植被的时空变化模式及生态多样性,Fan等[23 ] 基于HASM方法内插气候数据分析了青藏高原不同高度带的潜在植被分布,Bourque[24 ] 结合DEM和气象数据识别了圣卢西亚岛的植被区划. ...
... 改进的平均中心模型[21 ,23 ] 被用于识别中国北方干旱半干旱区各类潜在自然植被在1981—1990、1991—2000、2001—2010年及2011—2020年4个时间段平均中心偏移特征,其理论计算公式为: ...
Ecological life zones of Saint Lucia
1
2001
... Holdridge Life Zone(HLZ)模型是目前研究气候与植被生态系统定量关系的主流模型.该模型使用年平均生物温度、年降水量和潜在蒸散比率3个生物气候指标刻画各植被地带类型[18 -19 ] ,以其简明、适用而广泛应用于国内外气候植被模拟等研究领域.Zou等[20 ] 对黄土高原潜在植被的空间分布进行了模拟分析,范泽孟等[21 -22 ] 研究了中国生态过渡带类型及其分布的未来变化趋势与特征,揭示了气候变化驱动下欧亚大陆植被的时空变化模式及生态多样性,Fan等[23 ] 基于HASM方法内插气候数据分析了青藏高原不同高度带的潜在植被分布,Bourque[24 ] 结合DEM和气象数据识别了圣卢西亚岛的植被区划. ...
中国干旱半干旱区气候异常影响机理研究进展
2
2023
... 以上研究利用HLZ模型直接识别研究区植被类型,该方法判别潜在植被多缺乏与现状植被间关系的理论说明.此外,模型无法反映植被分布间高程差异的问题[25 ] 也未能解决,可能对地形多为高原山地、起伏明显的中国北方干旱区植被研究造成影响.鉴于此,如何划分与现状植被对应良好的潜在植被类型,构建适用于中国北方干旱半干旱区气候植被定量研究的模型,是厘清自然条件下干旱区生态系统主体与气候变化关系的核心.因此,论文旨在建立中国北方干旱半干旱区潜在植被分类体系,对HLZ模型进行修正、拓展,运用1981—2020年中国气象模拟数据,对潜在植被的空间分布格局进行模拟,揭示气候变化背景下干旱区潜在植被时空变化特征,以期为北方地区植被恢复、国家自然保护区建设等方面提供理论支撑. ...
... 原HLZ模型无法反映植被分布间高程差异的问题主要体现在其根据年平均生物温度不同参数层级划分出不同的纬度带与垂直带,而两者在数值上具有等值性[25 ,28 ] .等值性产生的问题会在生命地带即植被的命名上体现.命名方式为所属纬度带、垂直带以及植被类型的组合,其中纬度带需要根据当地年平均生物温度与海拔按标准气温直减率转换为海平面处的年平均生物温度进行判别[29 ] .以研究区青藏高原植被命名为例,该地平均海拔为4 082 m,年平均生物温度为2.74 ℃,垂直带为高山带,根据标准气温直减率,海平面处的年平均生物温度高于24 ℃,纬度带为热带,故命名为热带高山植被.而实际中青藏高原北部不属于热带,该错误命名主要是由温度转换、纬度带与垂直带分级完全一致引起的,故原模型的植被命名法也不适用于潜在植被的命名,在命名上需建立合适的标准. ...
中国北方干湿过渡区生态系统生产力的气候变化风险评估
1
2021
... 研究区为北方干旱半干旱区所在省市区[8 ,26 ] ,31°36′—53°20′N、73°27′—126°20′E,包括新疆、内蒙古、甘肃、青海、宁夏、陕西、山西、河北和北京9省市区,总面积约451.5万km2 (图1 ).地势西高东低,横跨中国三级阶梯,地形以盆地、山地和高原为主,包括柴达木盆地、塔里木盆地、准噶尔盆地、天山山脉、祁连山、青藏高原、内蒙古高原和黄土高原等.区内温带东西部由于距海远近差异,年平均降水量跨度较大,东部可达500 mm,而西部多低于50 mm且气温日较差更大,反映了温带季风气候与温带大陆性气候差异.青藏高原区域年平均降水量约为260 mm,越靠近柴达木盆地越干旱,年平均气温多在0 ℃以下,气温日较差较大.自然景观丰富,森林、灌丛、草原、沼泽、荒漠等在区内广泛分布[27 ] . ...
1
1960
... 研究区为北方干旱半干旱区所在省市区[8 ,26 ] ,31°36′—53°20′N、73°27′—126°20′E,包括新疆、内蒙古、甘肃、青海、宁夏、陕西、山西、河北和北京9省市区,总面积约451.5万km2 (图1 ).地势西高东低,横跨中国三级阶梯,地形以盆地、山地和高原为主,包括柴达木盆地、塔里木盆地、准噶尔盆地、天山山脉、祁连山、青藏高原、内蒙古高原和黄土高原等.区内温带东西部由于距海远近差异,年平均降水量跨度较大,东部可达500 mm,而西部多低于50 mm且气温日较差更大,反映了温带季风气候与温带大陆性气候差异.青藏高原区域年平均降水量约为260 mm,越靠近柴达木盆地越干旱,年平均气温多在0 ℃以下,气温日较差较大.自然景观丰富,森林、灌丛、草原、沼泽、荒漠等在区内广泛分布[27 ] . ...
研究全球变化的植被-气候分类系统
1
1993
... 原HLZ模型无法反映植被分布间高程差异的问题主要体现在其根据年平均生物温度不同参数层级划分出不同的纬度带与垂直带,而两者在数值上具有等值性[25 ,28 ] .等值性产生的问题会在生命地带即植被的命名上体现.命名方式为所属纬度带、垂直带以及植被类型的组合,其中纬度带需要根据当地年平均生物温度与海拔按标准气温直减率转换为海平面处的年平均生物温度进行判别[29 ] .以研究区青藏高原植被命名为例,该地平均海拔为4 082 m,年平均生物温度为2.74 ℃,垂直带为高山带,根据标准气温直减率,海平面处的年平均生物温度高于24 ℃,纬度带为热带,故命名为热带高山植被.而实际中青藏高原北部不属于热带,该错误命名主要是由温度转换、纬度带与垂直带分级完全一致引起的,故原模型的植被命名法也不适用于潜在植被的命名,在命名上需建立合适的标准. ...
Applying biogeographic principles to resource management: a case study evaluating holdridge's life zone model
1
1978
... 原HLZ模型无法反映植被分布间高程差异的问题主要体现在其根据年平均生物温度不同参数层级划分出不同的纬度带与垂直带,而两者在数值上具有等值性[25 ,28 ] .等值性产生的问题会在生命地带即植被的命名上体现.命名方式为所属纬度带、垂直带以及植被类型的组合,其中纬度带需要根据当地年平均生物温度与海拔按标准气温直减率转换为海平面处的年平均生物温度进行判别[29 ] .以研究区青藏高原植被命名为例,该地平均海拔为4 082 m,年平均生物温度为2.74 ℃,垂直带为高山带,根据标准气温直减率,海平面处的年平均生物温度高于24 ℃,纬度带为热带,故命名为热带高山植被.而实际中青藏高原北部不属于热带,该错误命名主要是由温度转换、纬度带与垂直带分级完全一致引起的,故原模型的植被命名法也不适用于潜在植被的命名,在命名上需建立合适的标准. ...
中国植被分类系统修订方案
1
2020
... 拓展后的模型以干旱半干旱区潜在自然植被类型作为生命地带.为建立其与现状植被的紧密联系,根据潜在自然植被定义、现状植被说明[16 ] 及其修订案[30 ] ,直接从现状植被中合并潜在植被.潜在植被的分类与命名主要遵循“植物群落学-生态学”分类原则,即以植被的立地生态条件与外貌作为依据,表现形式为温度带与植被型组的组合,限定各植被型组的温度带也可防止不合理植被命名的发生;其次,通过实验与重要性比较,若植被型组不同的植被需要划入某类中,则根据面积占比大于75%的植被类型命名.研究区最终共划分出高山冻原与稀疏植被、高山草本植被、山地森林植被、温带荒漠植被、温带草本植被和温带森林植被6类潜在自然植被(表1 ). ...
基于Holdridge生命地带模型的我国南北过渡带潜在植被分布模拟
1
2019
... 转移矩阵可以反映同一事物在不同状态下其内部结构间的转换关系与变化方向[31 -32 ] .通过建立模拟潜在植被与实际潜在植被(由现状植被模拟所得)间面积转移矩阵,作为模拟精度检验的基础,然后构建不同年代际间模拟潜在植被类型的面积转移矩阵,用以进一步分析不同时间段下各个植被类型的具体转换关系.该转移矩阵模型为: ...
抚顺矿区1989-2019年土地利用/覆盖变化分析
2
2021
... 转移矩阵可以反映同一事物在不同状态下其内部结构间的转换关系与变化方向[31 -32 ] .通过建立模拟潜在植被与实际潜在植被(由现状植被模拟所得)间面积转移矩阵,作为模拟精度检验的基础,然后构建不同年代际间模拟潜在植被类型的面积转移矩阵,用以进一步分析不同时间段下各个植被类型的具体转换关系.该转移矩阵模型为: ...
... 在中国北方干旱半干旱区潜在自然植被分类体系中并未出现独立的灌丛植被类型,这是由于划分的植被是非过渡性质的,而现状灌丛植被多因极端干旱或寒冷并不典型,与其他植被形成过渡带,致使其被周围生态幅更宽植被在气候指标阈值上所掩盖,这点在相关研究[32 ,34 -36 ] 中也有体现. ...
中国Holdridge生命地带与潜在植被空间格局研究
1
2009
... Kappa系数被用来检验潜在自然植被模拟精度,通过在SPSS中导入模拟与实际潜在植被的面积转移矩阵计算Kappa系数.Kappa系数值一般为[0,1],根据值的大小可以划分5种级别的一致性,即0~0.2为极低一致性、0.2~0.4为一般一致性、0.4~0.6为中度一致性、0.6~0.8为高度一致性、0.8~1为极高一致性[33 ] .其理论计算公式为: ...
基于CSCS模型的中国潜在自然植被时空分布特征研究
2
2014
... 基于代表现状植被的1∶100万中国植被图,划分出6种北方干旱半干旱区潜在自然植被类型,再根据各类植被的气候指标阈值,构建适应研究区的生命地带模型的方法,相较于前人基于原模型和气象数据确认潜在植被,能更精确地反映无人类活动影响下研究区的植被类型,进而有助于真实地揭示气候与当地植被的响应关系.本研究于中国黄土高原西部(34°30′—36°30′N,104°—107°E、)模拟潜在植被,其中山地森林植被的空间平均分布与Zou等[20 ] 研究结果基本一致,均分布于该区域较高海拔处,而在他们的研究中,“草原”与本文温带森林植被的分布一致,这是由双方潜在自然植被分类的差异所造成,实际上这两类植被在气候指标阈值上是较为接近的,均体现了与周围植被类型的差异;于研究区西部模拟的潜在植被空间分布大致与修丽娜[34 ] 的研究结果一致,反映出植被的垂直地带性分布;于内蒙古西部模拟的温带草原植被自东向西向温带荒漠植被变化基本与李飞等[7 ] 研究结果基本一致,反映出植被的经向地带性变化. ...
... 在中国北方干旱半干旱区潜在自然植被分类体系中并未出现独立的灌丛植被类型,这是由于划分的植被是非过渡性质的,而现状灌丛植被多因极端干旱或寒冷并不典型,与其他植被形成过渡带,致使其被周围生态幅更宽植被在气候指标阈值上所掩盖,这点在相关研究[32 ,34 -36 ] 中也有体现. ...
不同气候变化情景下2070-2099年中国潜在植被及其敏感性
0
2016
气候变化下中国潜在植被演替及其敏感性
1
2018
... 在中国北方干旱半干旱区潜在自然植被分类体系中并未出现独立的灌丛植被类型,这是由于划分的植被是非过渡性质的,而现状灌丛植被多因极端干旱或寒冷并不典型,与其他植被形成过渡带,致使其被周围生态幅更宽植被在气候指标阈值上所掩盖,这点在相关研究[32 ,34 -36 ] 中也有体现. ...








 甘公网安备 62010202000688号
甘公网安备 62010202000688号
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}