0 引言
生态位指群落中各种群所利用的资源总和及与其他相关种群间的功能关系[1 ] ,表示生态系统中每种生物生存必需的生境最小阈值[2 ] 。生态位理论对于研究物种间关系、种群间共存机制和群落动态演替等方面有着重要的作用[3 -6 ] 。生态位宽度和生态位重叠度,通过种间关系反映种群在生态系统中的时空位置,可供预测群落演替发展方向[7 ] 。生态位宽度反映物种对各种资源的利用程度,与物种对资源利用效率、适应环境能力呈正比[8 ] 。生态位重叠指不同物种对同一资源共同利用程度,反映物种间生态相似性与竞争强度[9 ] 。种间联结指不同物种在空间分布上的相互关联性,反映群落中各物种在不同生境中相互影响、相互作用所形成的有机关系[10 ] ,利用二元数据定性或定量分析植物种间的相关性[11 ] 。生态位与种间联结可以解释物种对资源的利用能力,反映种间协同与竞争关系,可被用于分析群落稳定性和演替趋势[12 ] ,在生态修复中具有一定的实践参考价值。生态位、n 维生态位理论[13 -17 ] 被广泛应用于物种多样性、群落结构演替与种群进化等研究[18 -21 ] 。植物生态位和种间联结性是种群生态学和群落生态学的重要内容,生态位特征和种间关系对群落的结构、功能和分类的作用,为当前热点研究内容。
目前诸多学者对多种脆弱生态地区进行研究,如砒砂岩区[22 ] 、喀斯特地貌区[1 ] 、干热河谷[23 ] 、黄土高原[24 ] 、林火后[9 ] 、岛屿[25 ] 等,认为脆弱生态系统下植被保护对生态建设有着重要意义。针对不同植被类型的研究(如苔藓植物[12 ] 、浮游植物[5 ] 、木本植物[26 ] 、草本植物[27 ] 等)表明物种资源利用的相似性导致种间竞争。研究者分别探究了乔木层[19 ] 、灌木层[25 ] 、草本层[27 ] ,认识群落的组成、结构和功能及群落与环境的相互关系,预测群落演替方向。西北区域物种多样性较低、生境较脆弱,天山是一道气候和生态环境的天然屏障[28 ] 。天山北麓绿洲荒漠过渡带受植被的特殊生境条件和人为干扰双重影响,是具有双重特征且敏感退化趋势的生态脆弱带,是绿洲与荒漠进退消涨地带[29 ] 。近年来天山北麓绿洲荒漠过渡带植被研究,侧重于生态位特征即物种组成、优势物种的多样性与生态位特征等方向[30 -31 ] ,关于优势植物生态位特征和种间联结的研究尚未有报道。
研究天山北麓绿洲荒漠过渡带优势植物的生态位特征与种间联结,探讨群落的稳定性,对今后生态系统的恢复、构建都具有重要意义。本文以天山北麓绿洲荒漠过渡带为研究区,采用Levins生态位宽度(B L )、Shannon生态位宽度(B S )、Schoener重叠指数(Cik )、Pianka重叠指数(Oik )、方差比率(VR )及χ 2 检验、Pearson相关检验、Spearman秩相关检验等方法进行分析,以期揭示该区优势物种生态位宽度和生态位重叠特征,进而推测其演替趋势,为生态保护提供理论基础或参考建议。
1 数据来源及研究方法
1.1 研究区概况
研究区位于新疆维吾尔自治区天山北麓,地理坐标为44°05′45″—44°37′45″N、87°48′07″—88°19′23″E,海拔383.29—683.56 m。研究区中包含了荒漠地带(古尔班通古特沙漠)、荒漠草原地带以及森林草原地带,存在一定过渡性。该地气候为温带荒漠气候,平均气温4~6 ℃,年降水量115 mm左右,年蒸发量1 942 mm。年降水量地区差异大,南部年降水量130—150 mm,沙漠区少于130 mm。无霜期达140~150 d。≥10 ℃的积温3 000~3 500 ℃,全年日照时数3 100~3 200 h,干燥度达16。土壤为碱化漠钙土,局部出现草甸盐土或盐化草甸土。植被组成简单、稀疏,乔灌草三层结构明显。其中主要物种包括榆树(Ulmus pumila )、角果藜(Ceratocarpus arenarius )、早熟禾(Poa annua )等。
1.2 数据来源
经野外实地调查,于2022年5—6月在天山北麓绿洲荒漠过渡带地区,选取具有代表性的地段共设置39个10 m×10 m样方(表1 ),其中受人为干扰较轻的过渡荒漠群落分布区9个, 古尔班通古特沙漠边缘地段30个。对样方内胸径≥ 2.5 cm的树种采用每木调查法。在每个样方设置4个5 m × 5 m的小样方,调查并记录灌木层内所有灌木个体(包括胸径≤2.5 cm的乔木幼树)。调查数据包括样地内灌草总盖度,灌木、草本层的平均高度和盖度,样方内灌木层各物种个体坐标值、高度、基径、冠幅,草本层不同物种的个体数量、高度、盖度和基径,并调查灌草层各物种的频度。同时,各样方采用GPS定位,记录其经度、纬度、海拔、生境类型等生态因子。
1.3 研究方法
1.3.1 总体联结性检验[11 ]
δ T 2 = ∑ i = 1 S P i ( 1 - P i ) (1)
S T 2 = 1 N ∑ i = 1 N ( T j - t ) 2 (2)
V R = S T 2 / δ T 2 (3)
W = V R × N (4)
式中:δ T 2 S T 2 S 为总物种数;N 为总样地数;Pi 为物种i 出现的频度;ni 为物种i 出现的样地数;Tj 为样地j 中出现的物种数;t 为样地中物种的平均数。VR 为灌木层主要物种间的总体联结指数,在独立性零假设条件下,VR 的期望值为1,其中种间独立则VR =1,种间正关联则VR >1,种间负关联则VR <1。W >χ 0.05 2 N )或W <χ 0.95 2 N ),为关联显著(P <0.05), χ 0.95 2 N )<W <χ 0.05 2 N )内,为关联不显著(P >0.05)。
1.3.2 种间联结性分析
用卡方检验、相关系数分析反映种间联结性[11 ] 。数据为非连续性取样,采用Yates的连续校正公式进行卡方检验:
χ 2 = N a d - b c - 0.5 N 2 ( a + b ) ( b + d ) ( c + d ) ( a + c ) (5)
式中:N 为总样地数;a 为2个物种同时出现的样地数;b 、c 分别为2个物种单独出现的样地数;d 为2个物种都不出现的样地数。当ad =bc 时,两个种相互独立;当ad >bc 时,两个种之间呈正联结;当ad <bc 时,两个种之间呈负联结。若χ 2 <3.841(P >0.05),为不显著联结,种对间基本独立;若3.841≤χ 2 ≤6.635(0.01≤P ≤0.05),为显著联结;若χ 2 >6.635(P <0.01),则是极显著联结。
χ 2 检验仅定性判断了种间联结性是否显著,补充采用Pearson相关检验和Spearman秩相关检验,分析物种间相伴出现的概率,确定物种间联结程度和差异。
r p ( i , k ) = ∑ j = 1 N ( x i j - x ¯ i ) 2 ( x k j - x ¯ k ) 2 ∑ j = 1 N ( x i j - x ¯ i ) 2 ∑ j = 1 N ( x k j - x ¯ k ) 2 (6)
r s ( i , k ) = 1 - 6 ∑ j = 1 N ( x i j - x ¯ i ) ( x k j - x ¯ k ) 2 N 3 - N (7)
式中:rp (i ,k )和rs (i ,k )分别是物种i 和物种k 在样地j 中的Pearson相关系数和Spearman秩相关系数;N 为样地总数;xij 和xkj 分别是物种i 和物种k 在样地j 中的重要值;xj 和x ¯ k j 个样地中物种i 和物种k 重要值的平均值。rp (i ,k )和rs (i ,k )的值域为[-1,1],反映正负相关性。
最后,对天山北麓主要物种间的Pearson相关系数、Spearman秩相关系数与生态位相似性和生态位重叠值运用SPSS17.0进行线性回归分析,计算重要值和生态位与种间联结的相关性特征。
2 结果与分析
2.1 重要值与生态位宽度
样地统计植物共有117种,选取重要值(大于0.5%)的前28位物种视为种群优势种,占比80.32%(表2 ),并进行生态位特征和种间联结分析。其中榆树的相对显著度在28个优势种中最高,早熟禾次之。群落中榆树重要值最高,角果藜、白梭梭(Haloxylon persicum )、早熟禾次之,而簇生泉卷耳(Cerastium fontanum )重要值最小,说明榆树和角果藜在植物群落中起主导作用,为群落中的主要优势种。主要物种的B L 和B S 的变化范围分别为1.00~12.11和0.00~2.69。群落中骆驼刺(Alhagi camelorum )的B L 和B S 分别为12.11和2.77,生态位宽度最大。角果藜、梭梭(Haloxylon ammodendron )、白梭梭等物种的生态位宽度较大。青甘杨(Populus przewalskii )、旱柳(Salix matsudana )、白车轴草(Trifolium repens )、簇生泉卷耳这4个物种的生态位宽度最小。重要值较大(小)的物种,生态位宽度一般较大(小),但两者排序并非完全一致。如灰绿藜(Oxybasis glauca )的重要值比蒌蒿(Artemisia selengensis )大,但生态位宽度较小。
2.2 生态位重叠程度
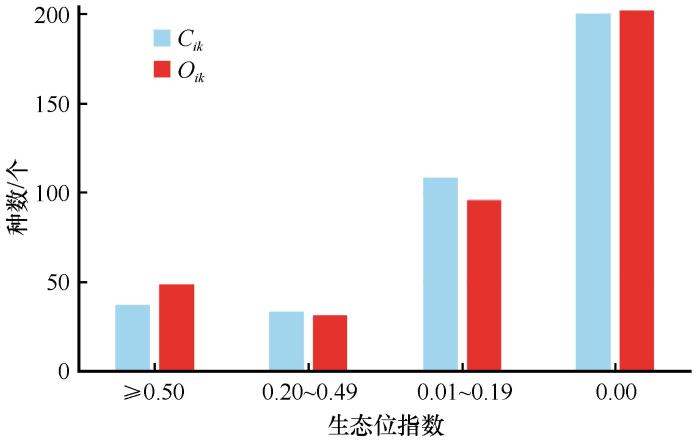
生态位重叠指数能量化种间资源的竞争程度,是判断物种间生态相似性与竞争强弱的重要指标。Schoener重叠指数(Cik ),为物种i 和k 的生态位相似性系数;Pianka重叠指数(Oik ),为物种i 和k 的生态位重叠指数,值域均为[0,1]。天山北麓绿洲荒漠过渡带的28个主要物种,共有378个种对,其Cik 平均值为0.121,Cik ≥0.50有37个种对数,簇生泉卷耳和白车轴草Cik 最大,青甘杨和白车轴草,青甘杨和簇生泉卷耳的重叠值高达1,表明生态位完全重叠无法共存,竞争关系明显。Cik 处于(0.2,0.49)的有33个种对数, Cik 介于(0.01,0.19)的有108个种对数,而角果藜和早熟禾,早熟禾和拉拉藤,早熟禾和骆驼刺等200个种对Cik 均为0(图1 )。
图1
图1
天山北麓绿洲荒漠过渡带优势植物Schoener重叠指数(Cik )和Pianka重叠指数(Oik )分布
Fig.1
Schoener overlap index (Cik ) and Pianka overlap index (Oik ), the main plant species in the northern foot of Tianshan Mountains
Oik 平均值为0.15,其中Oik ≥0.50有49个种对数,Oik 最大的是白车轴草和早熟禾,簇生泉卷耳和早熟禾等种间的生态位重叠值高达1,表明了生态位完全重叠二者无法共存。而角果藜和早熟禾,早熟禾和骆驼刺等202个种对Oik 均为0。有31个种对数Oik 为(0.20,0.49),有96个种对数Oik 介于(0.01,0.19),有202个种对数Oik 为0(图1 )。总体上来看天山北麓绿洲荒漠过渡带优势植物的少数种间存在较强竞争,其余多数竞争较弱,只有部分资源会被共同利用,而其他部分被各个物种各自占据。
2.3 总体联结性
由计算所得,总体联结性的方差比率VR = 1.18>1,表明天山北麓绿洲荒漠过渡带优势物种间总体呈正联结关系(表3 )。检验统计量W =45.87,而χ 0.95 ( 56 ) 2 W <χ 0.05 ( 56 ) 2 W 值落入χ 2
2.4 主要种对间的联结性
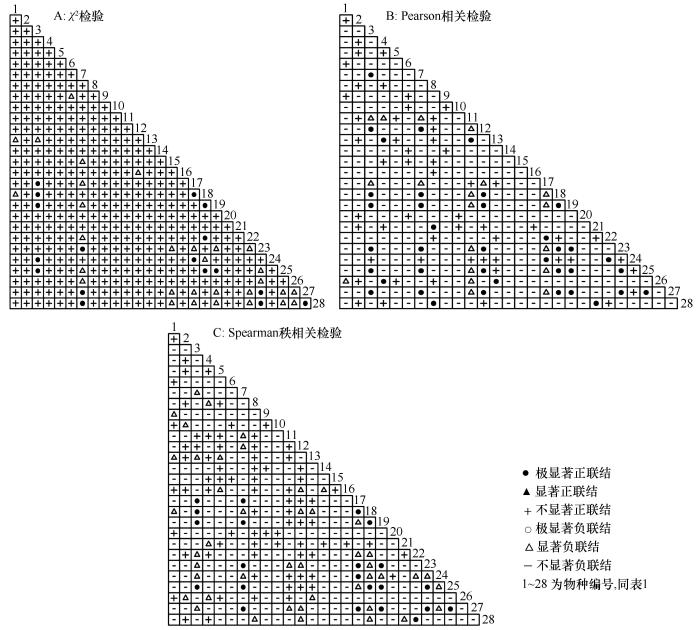
天山北麓绿洲荒漠过渡带主要物种经χ 2 检验全部显示正联结,其中呈极显著正联结的有17对,呈显著正联结的有30对,呈不显著正联结的有331对(87.57%,表4 、图2 A)。Pearson相关检验显著率为10.53%,显示正负相关种对比值0.43(表4 、图2 B)。正相关种对数有113对(其中呈极显著正相关的有38对,呈显著正相关的有19对,呈不显著正相关的有56对);呈不显著负相关的有265对(70.11%)。不显著相关种对有321对,绝大部分种对呈不显著相关,与χ 2
图2
图2
主要物种的χ 2 检验、Pearson相关检验、Spearman秩相关检验半矩阵图
Fig.2
Semi-matrix diagrams of chisquare test, Pearson correlation test, Spearman rank correlation test among main species
Spearman秩相关检验的结果显示正相关的种对数有142对,其中呈不显著正相关占比最高,显著正相关次之,有42对(表4 、图2 C)。负相关种对数有236对,呈不显著负相关233对。正负相关种对比值为0.60,检验显著率为11.90%,表明大部分种间呈不显著相关,如表4 所列。3种检验方法具有一致性,说明种对间联结性较差,空间分布呈一定的独立性。
2.5 种间联结系数与生态位重叠指数的回归分析
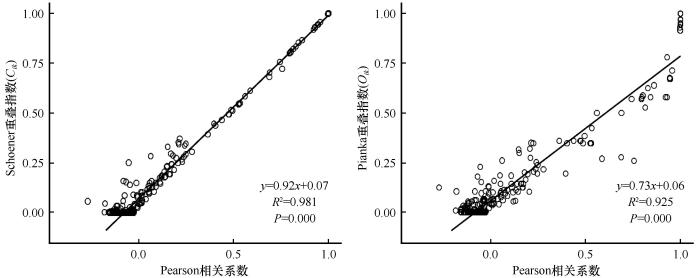
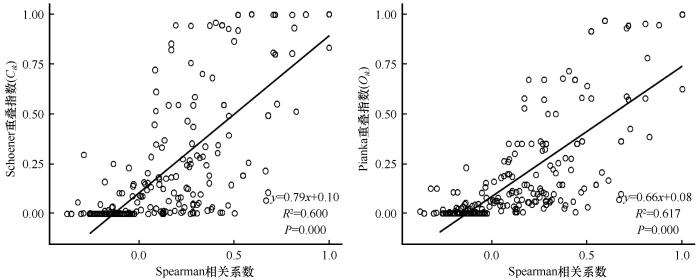
对天山北麓绿洲荒漠过渡带主要物种的Pearson相关系数、Spearman秩相关系数与生态位重叠指数进行回归分析,三者之间均呈极显著正相关(P <0.01,斜率>0),表明该过渡带主要物种的生态位重叠程度与Pearson相关系数、Spearman秩相关系数呈正比(图3 ~4 ),即样地主要物种的种间正联结性越强,种对间相伴生长几率越高,其生态位重叠指数越大。
图3
图3
主要物种Pearson相关系数和生态位重叠指数的回归分析
Fig.3
Regression analysis between Pearson correlation coefficient and niche overlap among main species
图4
图4
主要物种Spearman秩相关系数和生态位重叠指数的回归分析
Fig.4
Regression analysis between Spearman rank correlation coefficient and niche overlap among main species
3 讨论
3.1 绿洲荒漠过渡带优势树种的生态位特征
天山北麓绿洲荒漠过渡地带中不同生态环境孕育出不同类型植被,其群落Levins和Shannon生态位宽度测定结果具有一致性,表明未达到稳定阶段,处于群落结构变化较的时期[32 ] 。其中白梭梭、梭梭等物种生态位宽度较宽,具有一定发展潜力,具有耐高温、耐旱、耐盐等特性,生态适应性强,进而对养分资源的利用能力大,因此植被出现概率较高,空间分布范围广,成为绿洲荒漠过渡带生态系统中的优势种,可作为防沙治沙的优良品种进行开发与利用。另有部分物种重要值较大而生态位宽度却较小,如灰绿藜与蒌蒿、猪毛菜与早熟禾等。物种的分布频度与数量也会影响生态位宽度[1 ,10 ] ,其中白车轴草、簇生泉卷耳等喜湿物种在湿度较大的样地中优势明显,大量出现在某一样地中,提高了其在整体重要值的排名。但研究区内湿热气候存在较少,在调查时出现特定生境的样方内,对资源环境存在着一定的依赖性,生态位宽度很低,有较大可能会发展为特化种。而干旱气候地貌占大部分区域,降水量小,蒸发量大,因此耐旱植被适应性强,不同植被出现频率、数量不同。研究发现,骆驼刺的生态位宽度和重要值大小的排序结果并非完全一致。骆驼刺的生态位宽度最大,表明其在研究区分布范围较均匀,其性喜光、耐旱、耐贫瘠土壤,喜欢沙漠地带,因此在恶劣的干旱环境下的群落内部具有较强的竞争适应能力。天山北麓绿洲荒漠过渡带的环境异质性使得两者的相关性并不显著,说明群落资源还有待充分利用,群落正在朝着结构复杂化,功能不断完整的方向发展。
通常认为重叠指数(Cik 和Oik )≥0.500,则种间具有较大的生态和资源利用相似性,竞争比较严重[33 -35 ] 。种间生态位宽度与生态位重叠值一般呈正比。但本研究发现研究区出现物种的生态位宽度与生态位重叠呈反比的现象,例如角果藜和白车轴草等物种的生态位宽度较大,而生态位重叠程度较小。这可能是由天山北麓绿洲荒漠过渡带生境的脆弱与恶劣引起的,该区土壤分区差异性较大;其次不同空间区域内出现不同强度人为干扰破坏,造成部分地块土层浅薄、土壤含水量低和土壤养分差,不适应植物生长[29 ] ,物种间资源利用竞争激烈,某些物种为生存发育抢夺资源,导致对资源利用能力较差的物种多聚集在局部适宜的生境斑块中,其他空间分布较少,最终出现物种生态位宽度较小,但生态位重叠却较大[10 ] 。这与陈龙涛等[36 ] 研究发现的现象一致。对于生态位重叠指数为0的物种,说明物种生态习性和对环境资源要求不同,不存在竞争关系。天山北麓绿洲荒漠过渡带约一半物种生态位重叠指数为0,这可能与土壤理化性质不同而形成的生境异质性有关。研究区多数优势物种间的生态位重叠指数较低,生态位分化比较明显,说明优势种占据的生境资源不在同一空间,对资源的需求差异较大,造成物种分布不连续,受外界人为破坏严重时对植物群落的破坏较为严重,使得不同优势种难以形成同一稳定的生存环境。
3.2 种间联结特征
种间联系能很好地反映植物之间的相互作用,也能反映群落的动态变化。在次生演替的早期,物种之间正、负相关系数都很小,说明群落处在一个不稳定的时期,种间关系也是离散的。随演替进程的进行,种间连接趋向于正向连接,正-负连接比例增大,群落稳定性提高,进而向顶极演化。通过对天山北麓绿洲荒漠过渡带种间关系的测量,发现该地区主要植物的整体联结度呈极低的正相关性,与χ ²试验的正、负相关性相符合, Spearman秩相关和 Pearson相关系数都小于1,表明该区域处于不稳定状态,群落结构和物种构成尚不完备,处于演替早期。综合χ ²检验、Pearson相关检验和Spearman秩相关检验,群落内主要物种多呈不显著联结,种对间联结程度较低,多呈独立空间分布[37 -38 ] 。3种检验方法的结果均表明呈显著正联结的种对数大于显著负联结种对数,种对的显著率较低,说明群落演替初期物种间竞争较弱,种间联结较为分散,这在一定程度上可以缓和物种间对资源和空间的竞争关系,有利于演替初期物种的共存[9 ] 。此外Pearson相关检验与Spearman秩相关检验相对的显著率明显高于χ ²检验。这是因为Pearson相关对数据的正态分布有较高的要求,而野外采集的数据难以保证呈正态分布。而 Spearman 秩相关分析与物种如何分布不相关,所以为补充相关系数检验的不足, 需引入Spearman 秩相关分析做补充。χ ²是基于二元数据的检验结果,虽能够较好地反映物种间关联的显著程度,但在反映物种间联结大小方面存在不足,还需结合Spearman秩相关及Pearson相关进行分析[32 -33 ] 。
植物的种间连接关系与植物的生态位交叠关系紧密,它们之间的正向关系反映了植物的生态位重叠,负联结表明植物间呈生态位分离[35 ] 。通过对天山北麓绿洲荒漠的研究,发现天山北麓绿洲荒漠过渡带主要物种间Pearson相关系数、Spearman秩相关系数与生态位重叠值回归分析均呈极显著正相关,即生态位重叠程度随种间正联结增加而增大。这与诸多研究结果一致[39 -41 ] ,若对研究区进行植被修复,选择显著正相关种对更有利于提高群落稳定性。
4 结论
从天山北麓绿洲荒漠过渡带主要物种的生态位与种间联结特征来看,群落优势种种间整体呈不显著的正联结,有正向演替的趋势群落优势物种的生态位宽度差异较大,种间重叠与联结程度均较低,种间关系分散,竞争较缓和。在对天山北麓绿洲荒漠过渡带进行植被保护与恢复时,应参照当前群落演替的初级阶段特征,将生物学特点、生态学习性和生境需求相似,种间正联结较强的物种进行搭配种植,先锋种选择生态位宽度高的物种,伴生种选取生态位重叠程度低的物种。结合有限的环境资源,适当进行人工植被修复,促使群落形成有利的协调关系,维持群落稳定性和丰富群落多样性,促进群落可持续发展。
参考文献
View Option
[1]
杜忠毓 ,邢文黎 ,薛亮 ,等 喀斯特石漠化锑矿区植物群落主要物种生态位特征及其种间联结
[J].生态学报 ,2023 ,43 (7 ):1 -16 .
[本文引用: 3]
[2]
薛卫星 ,李春辉 ,艾训儒 ,等 鹅掌楸天然林优势树种生态位与种间联结性
[J].森林与环境学报 ,2023 ,43 (1 ):26 -34 .
[本文引用: 1]
[3]
Winemiller K O Fitzgerald D B Bower L M et al Functional traits,convergent evolution,and periodic tables of niches
[J].Ecology Letters ,2015 ,18 (8 ):737 -751 .
[本文引用: 1]
[4]
Rota E Caruso T Bargagli R Community structure,diversity and spatial organization of enchytraeids in Mediterranean urban holm oak stands
[J].European Journal of Soil Biology ,2014 ,62 :83 -91 .
[5]
马一明 ,李秋华 ,潘少朴 ,等 贵州高原花溪水库浮游植物优势种生态位及种间联结性动态分析
[J].湖泊科学 ,2021 ,33 (3 ):785 -796 .
[本文引用: 1]
[6]
王自力 ,陈勇 ,陈晓鸣 ,等 白蜡虫及其3种优势寄生蜂的时空生态位
[J].生态学报 ,2011 ,31 (4 ):914 -922 .
[本文引用: 1]
[7]
Vannette R L Fukami T Historical contingency in species interactions:towards niche‐based predictions
[J].Ecology Letters ,2014 ,17 (1 ):115 -124 .
[本文引用: 1]
[8]
张婷 ,张建利 ,张晨 ,等 FAST周边喀斯特土壤养分资源轴的植物优势种生态位
[J].生态学杂志 ,2023 ,42 (8 ):1820 -1828 .
[本文引用: 1]
[9]
刘鲁光 ,朱兆棋 ,陈曦 ,等 喜德县中坝村火烧迹地植被恢复初期优势种生态位与种间联结
[J].西部林业科学 ,2022 ,51 (1 ):110 -117 .
[本文引用: 3]
[10]
刘润红 ,陈乐 ,涂洪润 ,等 桂林岩溶石山青冈群落灌木层主要物种生态位与种间联结
[J].生态学报 ,2020 ,40 (6 ):2057 -2071 .
[本文引用: 3]
[11]
徐满厚 ,刘敏 ,翟大彤 ,等 植物种间联结研究内容与方法评述
[J].生态学报 ,2016 ,36 (24 ):8224 -8233 .
[本文引用: 3]
[12]
刘艳 ,郑越月 ,敖艳艳 不同生长基质的苔藓植物优势种生态位与种间联结
[J].生态学报 ,2019 ,39 (1 ):286 -293 .
[本文引用: 2]
[13]
Colwell R K Futuyma D J On the measurement of niche breadth and overlap
[J].Ecology ,1971 ,52 (4 ):567 -576 .
[本文引用: 1]
[14]
Mueller L D Altenberg L Statistical inference on measures of niche overlap
[J].Ecology ,1985 ,66 (4 ):1204 -1210 .
[15]
Yu S X Orloci L On the fundamental,realized niche and niche center
[J].Acta Scientiarum Naturalium Universitatis Sunyatseni ,1993 ,32 (4 ):70 -80 .
[16]
Leibold M A The niche concept revisited:mechanistic models and community context
[J].Ecology ,1995 ,76 (5 ):1371 -1382 .
[17]
Vandermeer J The niche construction paradigm in ecological time
[J].Ecological Modelling ,2008 ,214(2 /4 ):385 -390
[本文引用: 1]
[18]
张甜 ,贾荣亮 ,高艳红 ,等 沙坡头人工固沙植被演替过程中主要结皮生物生态位和种间关联变化特征
[J].中国沙漠 ,2021 ,41 (4 ):100 -108 .
[本文引用: 1]
[19]
肖艳梅 ,解婧媛 ,姚义鹏 ,等 桂林岩溶石山常绿落叶阔叶混交林乔木层优势物种生态位研究
[J].生态学报 ,2021 ,41 (20 ):8159 -8170 .
[本文引用: 1]
[20]
郝建锋 ,李艳 ,齐锦秋 ,等 人为干扰对碧峰峡栲树次生林群落物种多样性及其优势种群生态位的影响
[J].生态学报 ,2016 ,36 (23 ):7678 -7688 .
[21]
梁玉莲 ,国庆喜 基于地形汇流值的东北天然次生林主要树种的生态位
[J].生态学报 ,2010 ,30 (11 ):2874 -2882 .
[本文引用: 1]
[22]
孙成 ,秦富仓 ,包雪源 ,等 砒砂岩区人工油松林下草本植物的生态位特征
[J].水土保持通报 ,2021 ,41 (3 ):81 -86 .
[本文引用: 1]
[23]
何聪 ,吴世祥 ,杨丹 ,等 元谋干热河谷干湿季侵蚀沟草本群落优势种种间关系研究
[J].西部林业科学 ,2020 ,49 (4 ):124 -129 .
[本文引用: 1]
[24]
郭连金 ,张文辉 ,刘国彬 黄土丘陵区沙棘人工林发育过程中物种多样性及种间关联变化
[J].应用生态学报 ,2007 ,18 (1 ):9 -15 .
[本文引用: 1]
[25]
肖集泓 ,熊宽洪 ,陈越琳 ,等 福建琅岐岛台湾相思群落灌木层主要物种生态位与种间联结
[J].热带亚热带植物学报 ,2023 ,31 (6 ):805 -815 .
[本文引用: 2]
[26]
魏亚情 ,宋希强 ,赵莹 ,等 吊罗山石碌含笑群落木本植物种间联结性研究
[J].热带作物学报 ,2022 ,43 (12 ):2606 -2613 .
[本文引用: 1]
[27]
胡文杰 ,潘磊 ,唐万鹏 ,等 虫害侵扰初期华山松群落草本层生态位特征和种间关系
[J].东北林业大学学报 ,2023 ,51 (3 ):36 -40 .
[本文引用: 2]
[28]
郭文章 ,井长青 ,邓小进 ,等 天山北坡典型草地土壤呼吸特征及其对环境因子的响应
[J].中国农业科技导报 ,2022 ,24 (10 ):189 -199 .
[本文引用: 1]
[29]
班卫强 ,严成 ,尹林克 ,等 古尔班通古特沙漠南缘不同立地条件植物多样性和优势种群生态位特征研究
[J].中国沙漠 ,2012 ,32 (6 ):1632 -1638 .
[本文引用: 2]
[30]
张凯 ,王润元 ,张强 ,等 绿洲荒漠过渡带夏季晴天地表辐射和能量平衡及小气候特征
[J].中国沙漠 ,2007 ,27 (6 ):1055 -1061 .
[本文引用: 1]
[31]
张林静 ,岳明 ,张远东 ,等 新疆阜康绿洲荒漠过渡带植物群落物种多样性特征
[J].地理科学 ,2003 ,23 (3 ):329 -334 .
[本文引用: 1]
[32]
金俊彦 ,覃文更 ,谭卫宁 ,等 濒危植物单性木兰群落主要种群种间联结性研究
[J].西部林业科学 ,2013 ,42 (3 ):86 -94 .
[本文引用: 2]
[33]
Yang Q Liu S Hu C et al Ecological species groups and interspecific association of vegetation in natural recovery process at ejiadian landslide after 2008 Wenchuan earthquake
[J].Journal of Mountain Science ,2016 ,13 :1609 -1620 .
[本文引用: 2]
[34]
Walker B Conserving biological diversity through ecosystem resilience
[J].Conservation Biology ,1995 ,9 (4 ):747 -752 .
[35]
沈雪梨 ,吴远密 ,朱艺璇 ,等 濒危植物百花山葡萄群落物种生态位特征与种间联结研究
[J].植物科学学报 ,2020 ,38 (2 ):195 -204 .
[本文引用: 2]
[36]
陈龙涛 ,石晓东 ,高润梅 山西陵川南方红豆杉群落种间联结与生态位特征研究
[J].植物科学学报 ,2016 ,34 (4 ):521 -529 .
[本文引用: 1]
[37]
张零念 ,朱贵青 ,杨宽 ,等 滇中云南杨梅灌丛主要木本植物生态位与种间联结
[J].植物生态学报 ,2022 ,46 (11 ):1400 -1410 .
[本文引用: 1]
[38]
李帅锋 ,刘万德 ,苏建荣 ,等 滇西北云南红豆杉群落物种生态位与种间联结
[J].植物科学学报 ,2012 ,30 (6 ):568 -576 .
[本文引用: 1]
[39]
郭忠玲 ,马元丹 ,郑金萍 ,等 长白山落叶阔叶混交林的物种多样性、种群空间分布格局及种间关联性研究
[J].应用生态学报 ,2004 (11 ):2013 -2018 .
[本文引用: 1]
[40]
李丘霖 ,宗秀虹 ,邓洪平 ,等 赤水桫椤群落乔木层优势物种生态位与种间联结性研究
[J].西北植物学报 ,2017 ,37 (7 ):1422 -1428 .
[41]
白晓航 ,施佳颖 黑龙江丰林国家级自然保护区红松+紫椴+硕桦群系优势树种生态位特征与种间联结分析
[J].园林 ,2023 ,40 (10 ):14 -21 .
[本文引用: 1]
喀斯特石漠化锑矿区植物群落主要物种生态位特征及其种间联结
3
2023
... 生态位指群落中各种群所利用的资源总和及与其他相关种群间的功能关系[1 ] ,表示生态系统中每种生物生存必需的生境最小阈值[2 ] .生态位理论对于研究物种间关系、种群间共存机制和群落动态演替等方面有着重要的作用[3 -6 ] .生态位宽度和生态位重叠度,通过种间关系反映种群在生态系统中的时空位置,可供预测群落演替发展方向[7 ] .生态位宽度反映物种对各种资源的利用程度,与物种对资源利用效率、适应环境能力呈正比[8 ] .生态位重叠指不同物种对同一资源共同利用程度,反映物种间生态相似性与竞争强度[9 ] .种间联结指不同物种在空间分布上的相互关联性,反映群落中各物种在不同生境中相互影响、相互作用所形成的有机关系[10 ] ,利用二元数据定性或定量分析植物种间的相关性[11 ] .生态位与种间联结可以解释物种对资源的利用能力,反映种间协同与竞争关系,可被用于分析群落稳定性和演替趋势[12 ] ,在生态修复中具有一定的实践参考价值.生态位、n 维生态位理论[13 -17 ] 被广泛应用于物种多样性、群落结构演替与种群进化等研究[18 -21 ] .植物生态位和种间联结性是种群生态学和群落生态学的重要内容,生态位特征和种间关系对群落的结构、功能和分类的作用,为当前热点研究内容. ...
... 目前诸多学者对多种脆弱生态地区进行研究,如砒砂岩区[22 ] 、喀斯特地貌区[1 ] 、干热河谷[23 ] 、黄土高原[24 ] 、林火后[9 ] 、岛屿[25 ] 等,认为脆弱生态系统下植被保护对生态建设有着重要意义.针对不同植被类型的研究(如苔藓植物[12 ] 、浮游植物[5 ] 、木本植物[26 ] 、草本植物[27 ] 等)表明物种资源利用的相似性导致种间竞争.研究者分别探究了乔木层[19 ] 、灌木层[25 ] 、草本层[27 ] ,认识群落的组成、结构和功能及群落与环境的相互关系,预测群落演替方向.西北区域物种多样性较低、生境较脆弱,天山是一道气候和生态环境的天然屏障[28 ] .天山北麓绿洲荒漠过渡带受植被的特殊生境条件和人为干扰双重影响,是具有双重特征且敏感退化趋势的生态脆弱带,是绿洲与荒漠进退消涨地带[29 ] .近年来天山北麓绿洲荒漠过渡带植被研究,侧重于生态位特征即物种组成、优势物种的多样性与生态位特征等方向[30 -31 ] ,关于优势植物生态位特征和种间联结的研究尚未有报道. ...
... 天山北麓绿洲荒漠过渡地带中不同生态环境孕育出不同类型植被,其群落Levins和Shannon生态位宽度测定结果具有一致性,表明未达到稳定阶段,处于群落结构变化较的时期[32 ] .其中白梭梭、梭梭等物种生态位宽度较宽,具有一定发展潜力,具有耐高温、耐旱、耐盐等特性,生态适应性强,进而对养分资源的利用能力大,因此植被出现概率较高,空间分布范围广,成为绿洲荒漠过渡带生态系统中的优势种,可作为防沙治沙的优良品种进行开发与利用.另有部分物种重要值较大而生态位宽度却较小,如灰绿藜与蒌蒿、猪毛菜与早熟禾等.物种的分布频度与数量也会影响生态位宽度[1 ,10 ] ,其中白车轴草、簇生泉卷耳等喜湿物种在湿度较大的样地中优势明显,大量出现在某一样地中,提高了其在整体重要值的排名.但研究区内湿热气候存在较少,在调查时出现特定生境的样方内,对资源环境存在着一定的依赖性,生态位宽度很低,有较大可能会发展为特化种.而干旱气候地貌占大部分区域,降水量小,蒸发量大,因此耐旱植被适应性强,不同植被出现频率、数量不同.研究发现,骆驼刺的生态位宽度和重要值大小的排序结果并非完全一致.骆驼刺的生态位宽度最大,表明其在研究区分布范围较均匀,其性喜光、耐旱、耐贫瘠土壤,喜欢沙漠地带,因此在恶劣的干旱环境下的群落内部具有较强的竞争适应能力.天山北麓绿洲荒漠过渡带的环境异质性使得两者的相关性并不显著,说明群落资源还有待充分利用,群落正在朝着结构复杂化,功能不断完整的方向发展. ...
鹅掌楸天然林优势树种生态位与种间联结性
1
2023
... 生态位指群落中各种群所利用的资源总和及与其他相关种群间的功能关系[1 ] ,表示生态系统中每种生物生存必需的生境最小阈值[2 ] .生态位理论对于研究物种间关系、种群间共存机制和群落动态演替等方面有着重要的作用[3 -6 ] .生态位宽度和生态位重叠度,通过种间关系反映种群在生态系统中的时空位置,可供预测群落演替发展方向[7 ] .生态位宽度反映物种对各种资源的利用程度,与物种对资源利用效率、适应环境能力呈正比[8 ] .生态位重叠指不同物种对同一资源共同利用程度,反映物种间生态相似性与竞争强度[9 ] .种间联结指不同物种在空间分布上的相互关联性,反映群落中各物种在不同生境中相互影响、相互作用所形成的有机关系[10 ] ,利用二元数据定性或定量分析植物种间的相关性[11 ] .生态位与种间联结可以解释物种对资源的利用能力,反映种间协同与竞争关系,可被用于分析群落稳定性和演替趋势[12 ] ,在生态修复中具有一定的实践参考价值.生态位、n 维生态位理论[13 -17 ] 被广泛应用于物种多样性、群落结构演替与种群进化等研究[18 -21 ] .植物生态位和种间联结性是种群生态学和群落生态学的重要内容,生态位特征和种间关系对群落的结构、功能和分类的作用,为当前热点研究内容. ...
Functional traits,convergent evolution,and periodic tables of niches
1
2015
... 生态位指群落中各种群所利用的资源总和及与其他相关种群间的功能关系[1 ] ,表示生态系统中每种生物生存必需的生境最小阈值[2 ] .生态位理论对于研究物种间关系、种群间共存机制和群落动态演替等方面有着重要的作用[3 -6 ] .生态位宽度和生态位重叠度,通过种间关系反映种群在生态系统中的时空位置,可供预测群落演替发展方向[7 ] .生态位宽度反映物种对各种资源的利用程度,与物种对资源利用效率、适应环境能力呈正比[8 ] .生态位重叠指不同物种对同一资源共同利用程度,反映物种间生态相似性与竞争强度[9 ] .种间联结指不同物种在空间分布上的相互关联性,反映群落中各物种在不同生境中相互影响、相互作用所形成的有机关系[10 ] ,利用二元数据定性或定量分析植物种间的相关性[11 ] .生态位与种间联结可以解释物种对资源的利用能力,反映种间协同与竞争关系,可被用于分析群落稳定性和演替趋势[12 ] ,在生态修复中具有一定的实践参考价值.生态位、n 维生态位理论[13 -17 ] 被广泛应用于物种多样性、群落结构演替与种群进化等研究[18 -21 ] .植物生态位和种间联结性是种群生态学和群落生态学的重要内容,生态位特征和种间关系对群落的结构、功能和分类的作用,为当前热点研究内容. ...
Community structure,diversity and spatial organization of enchytraeids in Mediterranean urban holm oak stands
0
2014
贵州高原花溪水库浮游植物优势种生态位及种间联结性动态分析
1
2021
... 目前诸多学者对多种脆弱生态地区进行研究,如砒砂岩区[22 ] 、喀斯特地貌区[1 ] 、干热河谷[23 ] 、黄土高原[24 ] 、林火后[9 ] 、岛屿[25 ] 等,认为脆弱生态系统下植被保护对生态建设有着重要意义.针对不同植被类型的研究(如苔藓植物[12 ] 、浮游植物[5 ] 、木本植物[26 ] 、草本植物[27 ] 等)表明物种资源利用的相似性导致种间竞争.研究者分别探究了乔木层[19 ] 、灌木层[25 ] 、草本层[27 ] ,认识群落的组成、结构和功能及群落与环境的相互关系,预测群落演替方向.西北区域物种多样性较低、生境较脆弱,天山是一道气候和生态环境的天然屏障[28 ] .天山北麓绿洲荒漠过渡带受植被的特殊生境条件和人为干扰双重影响,是具有双重特征且敏感退化趋势的生态脆弱带,是绿洲与荒漠进退消涨地带[29 ] .近年来天山北麓绿洲荒漠过渡带植被研究,侧重于生态位特征即物种组成、优势物种的多样性与生态位特征等方向[30 -31 ] ,关于优势植物生态位特征和种间联结的研究尚未有报道. ...
白蜡虫及其3种优势寄生蜂的时空生态位
1
2011
... 生态位指群落中各种群所利用的资源总和及与其他相关种群间的功能关系[1 ] ,表示生态系统中每种生物生存必需的生境最小阈值[2 ] .生态位理论对于研究物种间关系、种群间共存机制和群落动态演替等方面有着重要的作用[3 -6 ] .生态位宽度和生态位重叠度,通过种间关系反映种群在生态系统中的时空位置,可供预测群落演替发展方向[7 ] .生态位宽度反映物种对各种资源的利用程度,与物种对资源利用效率、适应环境能力呈正比[8 ] .生态位重叠指不同物种对同一资源共同利用程度,反映物种间生态相似性与竞争强度[9 ] .种间联结指不同物种在空间分布上的相互关联性,反映群落中各物种在不同生境中相互影响、相互作用所形成的有机关系[10 ] ,利用二元数据定性或定量分析植物种间的相关性[11 ] .生态位与种间联结可以解释物种对资源的利用能力,反映种间协同与竞争关系,可被用于分析群落稳定性和演替趋势[12 ] ,在生态修复中具有一定的实践参考价值.生态位、n 维生态位理论[13 -17 ] 被广泛应用于物种多样性、群落结构演替与种群进化等研究[18 -21 ] .植物生态位和种间联结性是种群生态学和群落生态学的重要内容,生态位特征和种间关系对群落的结构、功能和分类的作用,为当前热点研究内容. ...
Historical contingency in species interactions:towards niche‐based predictions
1
2014
... 生态位指群落中各种群所利用的资源总和及与其他相关种群间的功能关系[1 ] ,表示生态系统中每种生物生存必需的生境最小阈值[2 ] .生态位理论对于研究物种间关系、种群间共存机制和群落动态演替等方面有着重要的作用[3 -6 ] .生态位宽度和生态位重叠度,通过种间关系反映种群在生态系统中的时空位置,可供预测群落演替发展方向[7 ] .生态位宽度反映物种对各种资源的利用程度,与物种对资源利用效率、适应环境能力呈正比[8 ] .生态位重叠指不同物种对同一资源共同利用程度,反映物种间生态相似性与竞争强度[9 ] .种间联结指不同物种在空间分布上的相互关联性,反映群落中各物种在不同生境中相互影响、相互作用所形成的有机关系[10 ] ,利用二元数据定性或定量分析植物种间的相关性[11 ] .生态位与种间联结可以解释物种对资源的利用能力,反映种间协同与竞争关系,可被用于分析群落稳定性和演替趋势[12 ] ,在生态修复中具有一定的实践参考价值.生态位、n 维生态位理论[13 -17 ] 被广泛应用于物种多样性、群落结构演替与种群进化等研究[18 -21 ] .植物生态位和种间联结性是种群生态学和群落生态学的重要内容,生态位特征和种间关系对群落的结构、功能和分类的作用,为当前热点研究内容. ...
FAST周边喀斯特土壤养分资源轴的植物优势种生态位
1
2023
... 生态位指群落中各种群所利用的资源总和及与其他相关种群间的功能关系[1 ] ,表示生态系统中每种生物生存必需的生境最小阈值[2 ] .生态位理论对于研究物种间关系、种群间共存机制和群落动态演替等方面有着重要的作用[3 -6 ] .生态位宽度和生态位重叠度,通过种间关系反映种群在生态系统中的时空位置,可供预测群落演替发展方向[7 ] .生态位宽度反映物种对各种资源的利用程度,与物种对资源利用效率、适应环境能力呈正比[8 ] .生态位重叠指不同物种对同一资源共同利用程度,反映物种间生态相似性与竞争强度[9 ] .种间联结指不同物种在空间分布上的相互关联性,反映群落中各物种在不同生境中相互影响、相互作用所形成的有机关系[10 ] ,利用二元数据定性或定量分析植物种间的相关性[11 ] .生态位与种间联结可以解释物种对资源的利用能力,反映种间协同与竞争关系,可被用于分析群落稳定性和演替趋势[12 ] ,在生态修复中具有一定的实践参考价值.生态位、n 维生态位理论[13 -17 ] 被广泛应用于物种多样性、群落结构演替与种群进化等研究[18 -21 ] .植物生态位和种间联结性是种群生态学和群落生态学的重要内容,生态位特征和种间关系对群落的结构、功能和分类的作用,为当前热点研究内容. ...
喜德县中坝村火烧迹地植被恢复初期优势种生态位与种间联结
3
2022
... 生态位指群落中各种群所利用的资源总和及与其他相关种群间的功能关系[1 ] ,表示生态系统中每种生物生存必需的生境最小阈值[2 ] .生态位理论对于研究物种间关系、种群间共存机制和群落动态演替等方面有着重要的作用[3 -6 ] .生态位宽度和生态位重叠度,通过种间关系反映种群在生态系统中的时空位置,可供预测群落演替发展方向[7 ] .生态位宽度反映物种对各种资源的利用程度,与物种对资源利用效率、适应环境能力呈正比[8 ] .生态位重叠指不同物种对同一资源共同利用程度,反映物种间生态相似性与竞争强度[9 ] .种间联结指不同物种在空间分布上的相互关联性,反映群落中各物种在不同生境中相互影响、相互作用所形成的有机关系[10 ] ,利用二元数据定性或定量分析植物种间的相关性[11 ] .生态位与种间联结可以解释物种对资源的利用能力,反映种间协同与竞争关系,可被用于分析群落稳定性和演替趋势[12 ] ,在生态修复中具有一定的实践参考价值.生态位、n 维生态位理论[13 -17 ] 被广泛应用于物种多样性、群落结构演替与种群进化等研究[18 -21 ] .植物生态位和种间联结性是种群生态学和群落生态学的重要内容,生态位特征和种间关系对群落的结构、功能和分类的作用,为当前热点研究内容. ...
... 目前诸多学者对多种脆弱生态地区进行研究,如砒砂岩区[22 ] 、喀斯特地貌区[1 ] 、干热河谷[23 ] 、黄土高原[24 ] 、林火后[9 ] 、岛屿[25 ] 等,认为脆弱生态系统下植被保护对生态建设有着重要意义.针对不同植被类型的研究(如苔藓植物[12 ] 、浮游植物[5 ] 、木本植物[26 ] 、草本植物[27 ] 等)表明物种资源利用的相似性导致种间竞争.研究者分别探究了乔木层[19 ] 、灌木层[25 ] 、草本层[27 ] ,认识群落的组成、结构和功能及群落与环境的相互关系,预测群落演替方向.西北区域物种多样性较低、生境较脆弱,天山是一道气候和生态环境的天然屏障[28 ] .天山北麓绿洲荒漠过渡带受植被的特殊生境条件和人为干扰双重影响,是具有双重特征且敏感退化趋势的生态脆弱带,是绿洲与荒漠进退消涨地带[29 ] .近年来天山北麓绿洲荒漠过渡带植被研究,侧重于生态位特征即物种组成、优势物种的多样性与生态位特征等方向[30 -31 ] ,关于优势植物生态位特征和种间联结的研究尚未有报道. ...
... 种间联系能很好地反映植物之间的相互作用,也能反映群落的动态变化.在次生演替的早期,物种之间正、负相关系数都很小,说明群落处在一个不稳定的时期,种间关系也是离散的.随演替进程的进行,种间连接趋向于正向连接,正-负连接比例增大,群落稳定性提高,进而向顶极演化.通过对天山北麓绿洲荒漠过渡带种间关系的测量,发现该地区主要植物的整体联结度呈极低的正相关性,与χ ²试验的正、负相关性相符合, Spearman秩相关和 Pearson相关系数都小于1,表明该区域处于不稳定状态,群落结构和物种构成尚不完备,处于演替早期.综合χ ²检验、Pearson相关检验和Spearman秩相关检验,群落内主要物种多呈不显著联结,种对间联结程度较低,多呈独立空间分布[37 -38 ] .3种检验方法的结果均表明呈显著正联结的种对数大于显著负联结种对数,种对的显著率较低,说明群落演替初期物种间竞争较弱,种间联结较为分散,这在一定程度上可以缓和物种间对资源和空间的竞争关系,有利于演替初期物种的共存[9 ] .此外Pearson相关检验与Spearman秩相关检验相对的显著率明显高于χ ²检验.这是因为Pearson相关对数据的正态分布有较高的要求,而野外采集的数据难以保证呈正态分布.而 Spearman 秩相关分析与物种如何分布不相关,所以为补充相关系数检验的不足, 需引入Spearman 秩相关分析做补充.χ ²是基于二元数据的检验结果,虽能够较好地反映物种间关联的显著程度,但在反映物种间联结大小方面存在不足,还需结合Spearman秩相关及Pearson相关进行分析[32 -33 ] . ...
桂林岩溶石山青冈群落灌木层主要物种生态位与种间联结
3
2020
... 生态位指群落中各种群所利用的资源总和及与其他相关种群间的功能关系[1 ] ,表示生态系统中每种生物生存必需的生境最小阈值[2 ] .生态位理论对于研究物种间关系、种群间共存机制和群落动态演替等方面有着重要的作用[3 -6 ] .生态位宽度和生态位重叠度,通过种间关系反映种群在生态系统中的时空位置,可供预测群落演替发展方向[7 ] .生态位宽度反映物种对各种资源的利用程度,与物种对资源利用效率、适应环境能力呈正比[8 ] .生态位重叠指不同物种对同一资源共同利用程度,反映物种间生态相似性与竞争强度[9 ] .种间联结指不同物种在空间分布上的相互关联性,反映群落中各物种在不同生境中相互影响、相互作用所形成的有机关系[10 ] ,利用二元数据定性或定量分析植物种间的相关性[11 ] .生态位与种间联结可以解释物种对资源的利用能力,反映种间协同与竞争关系,可被用于分析群落稳定性和演替趋势[12 ] ,在生态修复中具有一定的实践参考价值.生态位、n 维生态位理论[13 -17 ] 被广泛应用于物种多样性、群落结构演替与种群进化等研究[18 -21 ] .植物生态位和种间联结性是种群生态学和群落生态学的重要内容,生态位特征和种间关系对群落的结构、功能和分类的作用,为当前热点研究内容. ...
... 天山北麓绿洲荒漠过渡地带中不同生态环境孕育出不同类型植被,其群落Levins和Shannon生态位宽度测定结果具有一致性,表明未达到稳定阶段,处于群落结构变化较的时期[32 ] .其中白梭梭、梭梭等物种生态位宽度较宽,具有一定发展潜力,具有耐高温、耐旱、耐盐等特性,生态适应性强,进而对养分资源的利用能力大,因此植被出现概率较高,空间分布范围广,成为绿洲荒漠过渡带生态系统中的优势种,可作为防沙治沙的优良品种进行开发与利用.另有部分物种重要值较大而生态位宽度却较小,如灰绿藜与蒌蒿、猪毛菜与早熟禾等.物种的分布频度与数量也会影响生态位宽度[1 ,10 ] ,其中白车轴草、簇生泉卷耳等喜湿物种在湿度较大的样地中优势明显,大量出现在某一样地中,提高了其在整体重要值的排名.但研究区内湿热气候存在较少,在调查时出现特定生境的样方内,对资源环境存在着一定的依赖性,生态位宽度很低,有较大可能会发展为特化种.而干旱气候地貌占大部分区域,降水量小,蒸发量大,因此耐旱植被适应性强,不同植被出现频率、数量不同.研究发现,骆驼刺的生态位宽度和重要值大小的排序结果并非完全一致.骆驼刺的生态位宽度最大,表明其在研究区分布范围较均匀,其性喜光、耐旱、耐贫瘠土壤,喜欢沙漠地带,因此在恶劣的干旱环境下的群落内部具有较强的竞争适应能力.天山北麓绿洲荒漠过渡带的环境异质性使得两者的相关性并不显著,说明群落资源还有待充分利用,群落正在朝着结构复杂化,功能不断完整的方向发展. ...
... 通常认为重叠指数(Cik 和Oik )≥0.500,则种间具有较大的生态和资源利用相似性,竞争比较严重[33 -35 ] .种间生态位宽度与生态位重叠值一般呈正比.但本研究发现研究区出现物种的生态位宽度与生态位重叠呈反比的现象,例如角果藜和白车轴草等物种的生态位宽度较大,而生态位重叠程度较小.这可能是由天山北麓绿洲荒漠过渡带生境的脆弱与恶劣引起的,该区土壤分区差异性较大;其次不同空间区域内出现不同强度人为干扰破坏,造成部分地块土层浅薄、土壤含水量低和土壤养分差,不适应植物生长[29 ] ,物种间资源利用竞争激烈,某些物种为生存发育抢夺资源,导致对资源利用能力较差的物种多聚集在局部适宜的生境斑块中,其他空间分布较少,最终出现物种生态位宽度较小,但生态位重叠却较大[10 ] .这与陈龙涛等[36 ] 研究发现的现象一致.对于生态位重叠指数为0的物种,说明物种生态习性和对环境资源要求不同,不存在竞争关系.天山北麓绿洲荒漠过渡带约一半物种生态位重叠指数为0,这可能与土壤理化性质不同而形成的生境异质性有关.研究区多数优势物种间的生态位重叠指数较低,生态位分化比较明显,说明优势种占据的生境资源不在同一空间,对资源的需求差异较大,造成物种分布不连续,受外界人为破坏严重时对植物群落的破坏较为严重,使得不同优势种难以形成同一稳定的生存环境. ...
植物种间联结研究内容与方法评述
3
2016
... 生态位指群落中各种群所利用的资源总和及与其他相关种群间的功能关系[1 ] ,表示生态系统中每种生物生存必需的生境最小阈值[2 ] .生态位理论对于研究物种间关系、种群间共存机制和群落动态演替等方面有着重要的作用[3 -6 ] .生态位宽度和生态位重叠度,通过种间关系反映种群在生态系统中的时空位置,可供预测群落演替发展方向[7 ] .生态位宽度反映物种对各种资源的利用程度,与物种对资源利用效率、适应环境能力呈正比[8 ] .生态位重叠指不同物种对同一资源共同利用程度,反映物种间生态相似性与竞争强度[9 ] .种间联结指不同物种在空间分布上的相互关联性,反映群落中各物种在不同生境中相互影响、相互作用所形成的有机关系[10 ] ,利用二元数据定性或定量分析植物种间的相关性[11 ] .生态位与种间联结可以解释物种对资源的利用能力,反映种间协同与竞争关系,可被用于分析群落稳定性和演替趋势[12 ] ,在生态修复中具有一定的实践参考价值.生态位、n 维生态位理论[13 -17 ] 被广泛应用于物种多样性、群落结构演替与种群进化等研究[18 -21 ] .植物生态位和种间联结性是种群生态学和群落生态学的重要内容,生态位特征和种间关系对群落的结构、功能和分类的作用,为当前热点研究内容. ...
... Information table of sample plots the northern foot of Tianshan Mountains
Table 1 编号 经度 纬度 海拔/m 生境类型 编号 经度 纬度 海拔/m 生境类型 1 87.78°E 44.05°N 592.96 沙漠 21 87.15°E 44.06°N 369.96 沙漠 2 87.80°E 44.05°N 649.23 沙漠 22 87.14°E 44.06°N 398.12 沙漠 3 87.83°E 44.04°N 618.36 绿洲 23 87.80°E 44.40°N 396.78 沙漠 4 87.84°E 44.12°N 563.99 人工林 24 88.05°E 44.16°N 396.76 绿洲 5 87.84°E 44.13°N 468.74 绿洲 25 88.05°E 44.15°N 592.96 沙漠 6 87.84°E 44.13°N 458.05 人工林 26 87.14°E 44.06°N 401.26 绿洲 7 87.87°E 44.37°N 341.17 绿洲 27 87.87°E 44.38°N 435.12 沙漠 8 87.87°E 44.37°N 383.29 沙漠 28 87.87°E 44.37°N 389.83 沙漠 9 87.87°E 44.37°N 470.36 沙漠 29 87.88°E 44.46°N 538.23 绿洲 10 87.88°E 44.38°N 443.30 人工林 30 87.87°E 44.38°N 383.32 沙漠 11 87.98°E 44.63°N 445.56 沙漠 31 87.87°E 44.38°N 391.62 沙漠 12 87.98°E 44.62°N 431.15 沙漠 32 87.87°E 44.37°N 430.26 沙漠 13 87.88°E 44.37°N 592.96 沙漠 33 87.88°E 44.38°N 430.59 沙漠 14 87.87°E 44.15°N 390.12 绿洲 34 87.68°E 44.35°N 432.51 沙漠 15 87.88°E 44.38°N 383.29 沙漠 35 87.88°E 44.38°N 425.76 沙漠 16 87.87°E 44.63°N 384.42 绿洲 36 87.88°E 44.38°N 424.86 绿洲 17 87.88°E 44.38°N 379.53 沙漠 37 88.19°E 44.16°N 530.69 沙漠 18 87.80°E 44.05°N 592.96 绿洲 38 88.19°E 44.15°N 683.56 绿洲 19 87.14°E 44.06°N 396.76 沙漠 39 88.19°E 44.16°N 544.18 沙漠 20 87.80°E 44.65°N 396.70 绿洲
<strong>1.3</strong> 研究方法 <strong>1.3.1</strong> 总体联结性检验<sup>[<xref ref-type="bibr" rid="R11">11</xref>]</sup> 方差比率(VR )和W 值: ...
... 用卡方检验、相关系数分析反映种间联结性[11 ] .数据为非连续性取样,采用Yates的连续校正公式进行卡方检验: ...
不同生长基质的苔藓植物优势种生态位与种间联结
2
2019
... 生态位指群落中各种群所利用的资源总和及与其他相关种群间的功能关系[1 ] ,表示生态系统中每种生物生存必需的生境最小阈值[2 ] .生态位理论对于研究物种间关系、种群间共存机制和群落动态演替等方面有着重要的作用[3 -6 ] .生态位宽度和生态位重叠度,通过种间关系反映种群在生态系统中的时空位置,可供预测群落演替发展方向[7 ] .生态位宽度反映物种对各种资源的利用程度,与物种对资源利用效率、适应环境能力呈正比[8 ] .生态位重叠指不同物种对同一资源共同利用程度,反映物种间生态相似性与竞争强度[9 ] .种间联结指不同物种在空间分布上的相互关联性,反映群落中各物种在不同生境中相互影响、相互作用所形成的有机关系[10 ] ,利用二元数据定性或定量分析植物种间的相关性[11 ] .生态位与种间联结可以解释物种对资源的利用能力,反映种间协同与竞争关系,可被用于分析群落稳定性和演替趋势[12 ] ,在生态修复中具有一定的实践参考价值.生态位、n 维生态位理论[13 -17 ] 被广泛应用于物种多样性、群落结构演替与种群进化等研究[18 -21 ] .植物生态位和种间联结性是种群生态学和群落生态学的重要内容,生态位特征和种间关系对群落的结构、功能和分类的作用,为当前热点研究内容. ...
... 目前诸多学者对多种脆弱生态地区进行研究,如砒砂岩区[22 ] 、喀斯特地貌区[1 ] 、干热河谷[23 ] 、黄土高原[24 ] 、林火后[9 ] 、岛屿[25 ] 等,认为脆弱生态系统下植被保护对生态建设有着重要意义.针对不同植被类型的研究(如苔藓植物[12 ] 、浮游植物[5 ] 、木本植物[26 ] 、草本植物[27 ] 等)表明物种资源利用的相似性导致种间竞争.研究者分别探究了乔木层[19 ] 、灌木层[25 ] 、草本层[27 ] ,认识群落的组成、结构和功能及群落与环境的相互关系,预测群落演替方向.西北区域物种多样性较低、生境较脆弱,天山是一道气候和生态环境的天然屏障[28 ] .天山北麓绿洲荒漠过渡带受植被的特殊生境条件和人为干扰双重影响,是具有双重特征且敏感退化趋势的生态脆弱带,是绿洲与荒漠进退消涨地带[29 ] .近年来天山北麓绿洲荒漠过渡带植被研究,侧重于生态位特征即物种组成、优势物种的多样性与生态位特征等方向[30 -31 ] ,关于优势植物生态位特征和种间联结的研究尚未有报道. ...
On the measurement of niche breadth and overlap
1
1971
... 生态位指群落中各种群所利用的资源总和及与其他相关种群间的功能关系[1 ] ,表示生态系统中每种生物生存必需的生境最小阈值[2 ] .生态位理论对于研究物种间关系、种群间共存机制和群落动态演替等方面有着重要的作用[3 -6 ] .生态位宽度和生态位重叠度,通过种间关系反映种群在生态系统中的时空位置,可供预测群落演替发展方向[7 ] .生态位宽度反映物种对各种资源的利用程度,与物种对资源利用效率、适应环境能力呈正比[8 ] .生态位重叠指不同物种对同一资源共同利用程度,反映物种间生态相似性与竞争强度[9 ] .种间联结指不同物种在空间分布上的相互关联性,反映群落中各物种在不同生境中相互影响、相互作用所形成的有机关系[10 ] ,利用二元数据定性或定量分析植物种间的相关性[11 ] .生态位与种间联结可以解释物种对资源的利用能力,反映种间协同与竞争关系,可被用于分析群落稳定性和演替趋势[12 ] ,在生态修复中具有一定的实践参考价值.生态位、n 维生态位理论[13 -17 ] 被广泛应用于物种多样性、群落结构演替与种群进化等研究[18 -21 ] .植物生态位和种间联结性是种群生态学和群落生态学的重要内容,生态位特征和种间关系对群落的结构、功能和分类的作用,为当前热点研究内容. ...
Statistical inference on measures of niche overlap
0
1985
On the fundamental,realized niche and niche center
0
1993
The niche concept revisited:mechanistic models and community context
0
1995
The niche construction paradigm in ecological time
1
2008
... 生态位指群落中各种群所利用的资源总和及与其他相关种群间的功能关系[1 ] ,表示生态系统中每种生物生存必需的生境最小阈值[2 ] .生态位理论对于研究物种间关系、种群间共存机制和群落动态演替等方面有着重要的作用[3 -6 ] .生态位宽度和生态位重叠度,通过种间关系反映种群在生态系统中的时空位置,可供预测群落演替发展方向[7 ] .生态位宽度反映物种对各种资源的利用程度,与物种对资源利用效率、适应环境能力呈正比[8 ] .生态位重叠指不同物种对同一资源共同利用程度,反映物种间生态相似性与竞争强度[9 ] .种间联结指不同物种在空间分布上的相互关联性,反映群落中各物种在不同生境中相互影响、相互作用所形成的有机关系[10 ] ,利用二元数据定性或定量分析植物种间的相关性[11 ] .生态位与种间联结可以解释物种对资源的利用能力,反映种间协同与竞争关系,可被用于分析群落稳定性和演替趋势[12 ] ,在生态修复中具有一定的实践参考价值.生态位、n 维生态位理论[13 -17 ] 被广泛应用于物种多样性、群落结构演替与种群进化等研究[18 -21 ] .植物生态位和种间联结性是种群生态学和群落生态学的重要内容,生态位特征和种间关系对群落的结构、功能和分类的作用,为当前热点研究内容. ...
沙坡头人工固沙植被演替过程中主要结皮生物生态位和种间关联变化特征
1
2021
... 生态位指群落中各种群所利用的资源总和及与其他相关种群间的功能关系[1 ] ,表示生态系统中每种生物生存必需的生境最小阈值[2 ] .生态位理论对于研究物种间关系、种群间共存机制和群落动态演替等方面有着重要的作用[3 -6 ] .生态位宽度和生态位重叠度,通过种间关系反映种群在生态系统中的时空位置,可供预测群落演替发展方向[7 ] .生态位宽度反映物种对各种资源的利用程度,与物种对资源利用效率、适应环境能力呈正比[8 ] .生态位重叠指不同物种对同一资源共同利用程度,反映物种间生态相似性与竞争强度[9 ] .种间联结指不同物种在空间分布上的相互关联性,反映群落中各物种在不同生境中相互影响、相互作用所形成的有机关系[10 ] ,利用二元数据定性或定量分析植物种间的相关性[11 ] .生态位与种间联结可以解释物种对资源的利用能力,反映种间协同与竞争关系,可被用于分析群落稳定性和演替趋势[12 ] ,在生态修复中具有一定的实践参考价值.生态位、n 维生态位理论[13 -17 ] 被广泛应用于物种多样性、群落结构演替与种群进化等研究[18 -21 ] .植物生态位和种间联结性是种群生态学和群落生态学的重要内容,生态位特征和种间关系对群落的结构、功能和分类的作用,为当前热点研究内容. ...
桂林岩溶石山常绿落叶阔叶混交林乔木层优势物种生态位研究
1
2021
... 目前诸多学者对多种脆弱生态地区进行研究,如砒砂岩区[22 ] 、喀斯特地貌区[1 ] 、干热河谷[23 ] 、黄土高原[24 ] 、林火后[9 ] 、岛屿[25 ] 等,认为脆弱生态系统下植被保护对生态建设有着重要意义.针对不同植被类型的研究(如苔藓植物[12 ] 、浮游植物[5 ] 、木本植物[26 ] 、草本植物[27 ] 等)表明物种资源利用的相似性导致种间竞争.研究者分别探究了乔木层[19 ] 、灌木层[25 ] 、草本层[27 ] ,认识群落的组成、结构和功能及群落与环境的相互关系,预测群落演替方向.西北区域物种多样性较低、生境较脆弱,天山是一道气候和生态环境的天然屏障[28 ] .天山北麓绿洲荒漠过渡带受植被的特殊生境条件和人为干扰双重影响,是具有双重特征且敏感退化趋势的生态脆弱带,是绿洲与荒漠进退消涨地带[29 ] .近年来天山北麓绿洲荒漠过渡带植被研究,侧重于生态位特征即物种组成、优势物种的多样性与生态位特征等方向[30 -31 ] ,关于优势植物生态位特征和种间联结的研究尚未有报道. ...
人为干扰对碧峰峡栲树次生林群落物种多样性及其优势种群生态位的影响
0
2016
基于地形汇流值的东北天然次生林主要树种的生态位
1
2010
... 生态位指群落中各种群所利用的资源总和及与其他相关种群间的功能关系[1 ] ,表示生态系统中每种生物生存必需的生境最小阈值[2 ] .生态位理论对于研究物种间关系、种群间共存机制和群落动态演替等方面有着重要的作用[3 -6 ] .生态位宽度和生态位重叠度,通过种间关系反映种群在生态系统中的时空位置,可供预测群落演替发展方向[7 ] .生态位宽度反映物种对各种资源的利用程度,与物种对资源利用效率、适应环境能力呈正比[8 ] .生态位重叠指不同物种对同一资源共同利用程度,反映物种间生态相似性与竞争强度[9 ] .种间联结指不同物种在空间分布上的相互关联性,反映群落中各物种在不同生境中相互影响、相互作用所形成的有机关系[10 ] ,利用二元数据定性或定量分析植物种间的相关性[11 ] .生态位与种间联结可以解释物种对资源的利用能力,反映种间协同与竞争关系,可被用于分析群落稳定性和演替趋势[12 ] ,在生态修复中具有一定的实践参考价值.生态位、n 维生态位理论[13 -17 ] 被广泛应用于物种多样性、群落结构演替与种群进化等研究[18 -21 ] .植物生态位和种间联结性是种群生态学和群落生态学的重要内容,生态位特征和种间关系对群落的结构、功能和分类的作用,为当前热点研究内容. ...
砒砂岩区人工油松林下草本植物的生态位特征
1
2021
... 目前诸多学者对多种脆弱生态地区进行研究,如砒砂岩区[22 ] 、喀斯特地貌区[1 ] 、干热河谷[23 ] 、黄土高原[24 ] 、林火后[9 ] 、岛屿[25 ] 等,认为脆弱生态系统下植被保护对生态建设有着重要意义.针对不同植被类型的研究(如苔藓植物[12 ] 、浮游植物[5 ] 、木本植物[26 ] 、草本植物[27 ] 等)表明物种资源利用的相似性导致种间竞争.研究者分别探究了乔木层[19 ] 、灌木层[25 ] 、草本层[27 ] ,认识群落的组成、结构和功能及群落与环境的相互关系,预测群落演替方向.西北区域物种多样性较低、生境较脆弱,天山是一道气候和生态环境的天然屏障[28 ] .天山北麓绿洲荒漠过渡带受植被的特殊生境条件和人为干扰双重影响,是具有双重特征且敏感退化趋势的生态脆弱带,是绿洲与荒漠进退消涨地带[29 ] .近年来天山北麓绿洲荒漠过渡带植被研究,侧重于生态位特征即物种组成、优势物种的多样性与生态位特征等方向[30 -31 ] ,关于优势植物生态位特征和种间联结的研究尚未有报道. ...
元谋干热河谷干湿季侵蚀沟草本群落优势种种间关系研究
1
2020
... 目前诸多学者对多种脆弱生态地区进行研究,如砒砂岩区[22 ] 、喀斯特地貌区[1 ] 、干热河谷[23 ] 、黄土高原[24 ] 、林火后[9 ] 、岛屿[25 ] 等,认为脆弱生态系统下植被保护对生态建设有着重要意义.针对不同植被类型的研究(如苔藓植物[12 ] 、浮游植物[5 ] 、木本植物[26 ] 、草本植物[27 ] 等)表明物种资源利用的相似性导致种间竞争.研究者分别探究了乔木层[19 ] 、灌木层[25 ] 、草本层[27 ] ,认识群落的组成、结构和功能及群落与环境的相互关系,预测群落演替方向.西北区域物种多样性较低、生境较脆弱,天山是一道气候和生态环境的天然屏障[28 ] .天山北麓绿洲荒漠过渡带受植被的特殊生境条件和人为干扰双重影响,是具有双重特征且敏感退化趋势的生态脆弱带,是绿洲与荒漠进退消涨地带[29 ] .近年来天山北麓绿洲荒漠过渡带植被研究,侧重于生态位特征即物种组成、优势物种的多样性与生态位特征等方向[30 -31 ] ,关于优势植物生态位特征和种间联结的研究尚未有报道. ...
黄土丘陵区沙棘人工林发育过程中物种多样性及种间关联变化
1
2007
... 目前诸多学者对多种脆弱生态地区进行研究,如砒砂岩区[22 ] 、喀斯特地貌区[1 ] 、干热河谷[23 ] 、黄土高原[24 ] 、林火后[9 ] 、岛屿[25 ] 等,认为脆弱生态系统下植被保护对生态建设有着重要意义.针对不同植被类型的研究(如苔藓植物[12 ] 、浮游植物[5 ] 、木本植物[26 ] 、草本植物[27 ] 等)表明物种资源利用的相似性导致种间竞争.研究者分别探究了乔木层[19 ] 、灌木层[25 ] 、草本层[27 ] ,认识群落的组成、结构和功能及群落与环境的相互关系,预测群落演替方向.西北区域物种多样性较低、生境较脆弱,天山是一道气候和生态环境的天然屏障[28 ] .天山北麓绿洲荒漠过渡带受植被的特殊生境条件和人为干扰双重影响,是具有双重特征且敏感退化趋势的生态脆弱带,是绿洲与荒漠进退消涨地带[29 ] .近年来天山北麓绿洲荒漠过渡带植被研究,侧重于生态位特征即物种组成、优势物种的多样性与生态位特征等方向[30 -31 ] ,关于优势植物生态位特征和种间联结的研究尚未有报道. ...
福建琅岐岛台湾相思群落灌木层主要物种生态位与种间联结
2
2023
... 目前诸多学者对多种脆弱生态地区进行研究,如砒砂岩区[22 ] 、喀斯特地貌区[1 ] 、干热河谷[23 ] 、黄土高原[24 ] 、林火后[9 ] 、岛屿[25 ] 等,认为脆弱生态系统下植被保护对生态建设有着重要意义.针对不同植被类型的研究(如苔藓植物[12 ] 、浮游植物[5 ] 、木本植物[26 ] 、草本植物[27 ] 等)表明物种资源利用的相似性导致种间竞争.研究者分别探究了乔木层[19 ] 、灌木层[25 ] 、草本层[27 ] ,认识群落的组成、结构和功能及群落与环境的相互关系,预测群落演替方向.西北区域物种多样性较低、生境较脆弱,天山是一道气候和生态环境的天然屏障[28 ] .天山北麓绿洲荒漠过渡带受植被的特殊生境条件和人为干扰双重影响,是具有双重特征且敏感退化趋势的生态脆弱带,是绿洲与荒漠进退消涨地带[29 ] .近年来天山北麓绿洲荒漠过渡带植被研究,侧重于生态位特征即物种组成、优势物种的多样性与生态位特征等方向[30 -31 ] ,关于优势植物生态位特征和种间联结的研究尚未有报道. ...
... [25 ]、草本层[27 ] ,认识群落的组成、结构和功能及群落与环境的相互关系,预测群落演替方向.西北区域物种多样性较低、生境较脆弱,天山是一道气候和生态环境的天然屏障[28 ] .天山北麓绿洲荒漠过渡带受植被的特殊生境条件和人为干扰双重影响,是具有双重特征且敏感退化趋势的生态脆弱带,是绿洲与荒漠进退消涨地带[29 ] .近年来天山北麓绿洲荒漠过渡带植被研究,侧重于生态位特征即物种组成、优势物种的多样性与生态位特征等方向[30 -31 ] ,关于优势植物生态位特征和种间联结的研究尚未有报道. ...
吊罗山石碌含笑群落木本植物种间联结性研究
1
2022
... 目前诸多学者对多种脆弱生态地区进行研究,如砒砂岩区[22 ] 、喀斯特地貌区[1 ] 、干热河谷[23 ] 、黄土高原[24 ] 、林火后[9 ] 、岛屿[25 ] 等,认为脆弱生态系统下植被保护对生态建设有着重要意义.针对不同植被类型的研究(如苔藓植物[12 ] 、浮游植物[5 ] 、木本植物[26 ] 、草本植物[27 ] 等)表明物种资源利用的相似性导致种间竞争.研究者分别探究了乔木层[19 ] 、灌木层[25 ] 、草本层[27 ] ,认识群落的组成、结构和功能及群落与环境的相互关系,预测群落演替方向.西北区域物种多样性较低、生境较脆弱,天山是一道气候和生态环境的天然屏障[28 ] .天山北麓绿洲荒漠过渡带受植被的特殊生境条件和人为干扰双重影响,是具有双重特征且敏感退化趋势的生态脆弱带,是绿洲与荒漠进退消涨地带[29 ] .近年来天山北麓绿洲荒漠过渡带植被研究,侧重于生态位特征即物种组成、优势物种的多样性与生态位特征等方向[30 -31 ] ,关于优势植物生态位特征和种间联结的研究尚未有报道. ...
虫害侵扰初期华山松群落草本层生态位特征和种间关系
2
2023
... 目前诸多学者对多种脆弱生态地区进行研究,如砒砂岩区[22 ] 、喀斯特地貌区[1 ] 、干热河谷[23 ] 、黄土高原[24 ] 、林火后[9 ] 、岛屿[25 ] 等,认为脆弱生态系统下植被保护对生态建设有着重要意义.针对不同植被类型的研究(如苔藓植物[12 ] 、浮游植物[5 ] 、木本植物[26 ] 、草本植物[27 ] 等)表明物种资源利用的相似性导致种间竞争.研究者分别探究了乔木层[19 ] 、灌木层[25 ] 、草本层[27 ] ,认识群落的组成、结构和功能及群落与环境的相互关系,预测群落演替方向.西北区域物种多样性较低、生境较脆弱,天山是一道气候和生态环境的天然屏障[28 ] .天山北麓绿洲荒漠过渡带受植被的特殊生境条件和人为干扰双重影响,是具有双重特征且敏感退化趋势的生态脆弱带,是绿洲与荒漠进退消涨地带[29 ] .近年来天山北麓绿洲荒漠过渡带植被研究,侧重于生态位特征即物种组成、优势物种的多样性与生态位特征等方向[30 -31 ] ,关于优势植物生态位特征和种间联结的研究尚未有报道. ...
... [27 ],认识群落的组成、结构和功能及群落与环境的相互关系,预测群落演替方向.西北区域物种多样性较低、生境较脆弱,天山是一道气候和生态环境的天然屏障[28 ] .天山北麓绿洲荒漠过渡带受植被的特殊生境条件和人为干扰双重影响,是具有双重特征且敏感退化趋势的生态脆弱带,是绿洲与荒漠进退消涨地带[29 ] .近年来天山北麓绿洲荒漠过渡带植被研究,侧重于生态位特征即物种组成、优势物种的多样性与生态位特征等方向[30 -31 ] ,关于优势植物生态位特征和种间联结的研究尚未有报道. ...
天山北坡典型草地土壤呼吸特征及其对环境因子的响应
1
2022
... 目前诸多学者对多种脆弱生态地区进行研究,如砒砂岩区[22 ] 、喀斯特地貌区[1 ] 、干热河谷[23 ] 、黄土高原[24 ] 、林火后[9 ] 、岛屿[25 ] 等,认为脆弱生态系统下植被保护对生态建设有着重要意义.针对不同植被类型的研究(如苔藓植物[12 ] 、浮游植物[5 ] 、木本植物[26 ] 、草本植物[27 ] 等)表明物种资源利用的相似性导致种间竞争.研究者分别探究了乔木层[19 ] 、灌木层[25 ] 、草本层[27 ] ,认识群落的组成、结构和功能及群落与环境的相互关系,预测群落演替方向.西北区域物种多样性较低、生境较脆弱,天山是一道气候和生态环境的天然屏障[28 ] .天山北麓绿洲荒漠过渡带受植被的特殊生境条件和人为干扰双重影响,是具有双重特征且敏感退化趋势的生态脆弱带,是绿洲与荒漠进退消涨地带[29 ] .近年来天山北麓绿洲荒漠过渡带植被研究,侧重于生态位特征即物种组成、优势物种的多样性与生态位特征等方向[30 -31 ] ,关于优势植物生态位特征和种间联结的研究尚未有报道. ...
古尔班通古特沙漠南缘不同立地条件植物多样性和优势种群生态位特征研究
2
2012
... 目前诸多学者对多种脆弱生态地区进行研究,如砒砂岩区[22 ] 、喀斯特地貌区[1 ] 、干热河谷[23 ] 、黄土高原[24 ] 、林火后[9 ] 、岛屿[25 ] 等,认为脆弱生态系统下植被保护对生态建设有着重要意义.针对不同植被类型的研究(如苔藓植物[12 ] 、浮游植物[5 ] 、木本植物[26 ] 、草本植物[27 ] 等)表明物种资源利用的相似性导致种间竞争.研究者分别探究了乔木层[19 ] 、灌木层[25 ] 、草本层[27 ] ,认识群落的组成、结构和功能及群落与环境的相互关系,预测群落演替方向.西北区域物种多样性较低、生境较脆弱,天山是一道气候和生态环境的天然屏障[28 ] .天山北麓绿洲荒漠过渡带受植被的特殊生境条件和人为干扰双重影响,是具有双重特征且敏感退化趋势的生态脆弱带,是绿洲与荒漠进退消涨地带[29 ] .近年来天山北麓绿洲荒漠过渡带植被研究,侧重于生态位特征即物种组成、优势物种的多样性与生态位特征等方向[30 -31 ] ,关于优势植物生态位特征和种间联结的研究尚未有报道. ...
... 通常认为重叠指数(Cik 和Oik )≥0.500,则种间具有较大的生态和资源利用相似性,竞争比较严重[33 -35 ] .种间生态位宽度与生态位重叠值一般呈正比.但本研究发现研究区出现物种的生态位宽度与生态位重叠呈反比的现象,例如角果藜和白车轴草等物种的生态位宽度较大,而生态位重叠程度较小.这可能是由天山北麓绿洲荒漠过渡带生境的脆弱与恶劣引起的,该区土壤分区差异性较大;其次不同空间区域内出现不同强度人为干扰破坏,造成部分地块土层浅薄、土壤含水量低和土壤养分差,不适应植物生长[29 ] ,物种间资源利用竞争激烈,某些物种为生存发育抢夺资源,导致对资源利用能力较差的物种多聚集在局部适宜的生境斑块中,其他空间分布较少,最终出现物种生态位宽度较小,但生态位重叠却较大[10 ] .这与陈龙涛等[36 ] 研究发现的现象一致.对于生态位重叠指数为0的物种,说明物种生态习性和对环境资源要求不同,不存在竞争关系.天山北麓绿洲荒漠过渡带约一半物种生态位重叠指数为0,这可能与土壤理化性质不同而形成的生境异质性有关.研究区多数优势物种间的生态位重叠指数较低,生态位分化比较明显,说明优势种占据的生境资源不在同一空间,对资源的需求差异较大,造成物种分布不连续,受外界人为破坏严重时对植物群落的破坏较为严重,使得不同优势种难以形成同一稳定的生存环境. ...
绿洲荒漠过渡带夏季晴天地表辐射和能量平衡及小气候特征
1
2007
... 目前诸多学者对多种脆弱生态地区进行研究,如砒砂岩区[22 ] 、喀斯特地貌区[1 ] 、干热河谷[23 ] 、黄土高原[24 ] 、林火后[9 ] 、岛屿[25 ] 等,认为脆弱生态系统下植被保护对生态建设有着重要意义.针对不同植被类型的研究(如苔藓植物[12 ] 、浮游植物[5 ] 、木本植物[26 ] 、草本植物[27 ] 等)表明物种资源利用的相似性导致种间竞争.研究者分别探究了乔木层[19 ] 、灌木层[25 ] 、草本层[27 ] ,认识群落的组成、结构和功能及群落与环境的相互关系,预测群落演替方向.西北区域物种多样性较低、生境较脆弱,天山是一道气候和生态环境的天然屏障[28 ] .天山北麓绿洲荒漠过渡带受植被的特殊生境条件和人为干扰双重影响,是具有双重特征且敏感退化趋势的生态脆弱带,是绿洲与荒漠进退消涨地带[29 ] .近年来天山北麓绿洲荒漠过渡带植被研究,侧重于生态位特征即物种组成、优势物种的多样性与生态位特征等方向[30 -31 ] ,关于优势植物生态位特征和种间联结的研究尚未有报道. ...
新疆阜康绿洲荒漠过渡带植物群落物种多样性特征
1
2003
... 目前诸多学者对多种脆弱生态地区进行研究,如砒砂岩区[22 ] 、喀斯特地貌区[1 ] 、干热河谷[23 ] 、黄土高原[24 ] 、林火后[9 ] 、岛屿[25 ] 等,认为脆弱生态系统下植被保护对生态建设有着重要意义.针对不同植被类型的研究(如苔藓植物[12 ] 、浮游植物[5 ] 、木本植物[26 ] 、草本植物[27 ] 等)表明物种资源利用的相似性导致种间竞争.研究者分别探究了乔木层[19 ] 、灌木层[25 ] 、草本层[27 ] ,认识群落的组成、结构和功能及群落与环境的相互关系,预测群落演替方向.西北区域物种多样性较低、生境较脆弱,天山是一道气候和生态环境的天然屏障[28 ] .天山北麓绿洲荒漠过渡带受植被的特殊生境条件和人为干扰双重影响,是具有双重特征且敏感退化趋势的生态脆弱带,是绿洲与荒漠进退消涨地带[29 ] .近年来天山北麓绿洲荒漠过渡带植被研究,侧重于生态位特征即物种组成、优势物种的多样性与生态位特征等方向[30 -31 ] ,关于优势植物生态位特征和种间联结的研究尚未有报道. ...
濒危植物单性木兰群落主要种群种间联结性研究
2
2013
... 天山北麓绿洲荒漠过渡地带中不同生态环境孕育出不同类型植被,其群落Levins和Shannon生态位宽度测定结果具有一致性,表明未达到稳定阶段,处于群落结构变化较的时期[32 ] .其中白梭梭、梭梭等物种生态位宽度较宽,具有一定发展潜力,具有耐高温、耐旱、耐盐等特性,生态适应性强,进而对养分资源的利用能力大,因此植被出现概率较高,空间分布范围广,成为绿洲荒漠过渡带生态系统中的优势种,可作为防沙治沙的优良品种进行开发与利用.另有部分物种重要值较大而生态位宽度却较小,如灰绿藜与蒌蒿、猪毛菜与早熟禾等.物种的分布频度与数量也会影响生态位宽度[1 ,10 ] ,其中白车轴草、簇生泉卷耳等喜湿物种在湿度较大的样地中优势明显,大量出现在某一样地中,提高了其在整体重要值的排名.但研究区内湿热气候存在较少,在调查时出现特定生境的样方内,对资源环境存在着一定的依赖性,生态位宽度很低,有较大可能会发展为特化种.而干旱气候地貌占大部分区域,降水量小,蒸发量大,因此耐旱植被适应性强,不同植被出现频率、数量不同.研究发现,骆驼刺的生态位宽度和重要值大小的排序结果并非完全一致.骆驼刺的生态位宽度最大,表明其在研究区分布范围较均匀,其性喜光、耐旱、耐贫瘠土壤,喜欢沙漠地带,因此在恶劣的干旱环境下的群落内部具有较强的竞争适应能力.天山北麓绿洲荒漠过渡带的环境异质性使得两者的相关性并不显著,说明群落资源还有待充分利用,群落正在朝着结构复杂化,功能不断完整的方向发展. ...
... 种间联系能很好地反映植物之间的相互作用,也能反映群落的动态变化.在次生演替的早期,物种之间正、负相关系数都很小,说明群落处在一个不稳定的时期,种间关系也是离散的.随演替进程的进行,种间连接趋向于正向连接,正-负连接比例增大,群落稳定性提高,进而向顶极演化.通过对天山北麓绿洲荒漠过渡带种间关系的测量,发现该地区主要植物的整体联结度呈极低的正相关性,与χ ²试验的正、负相关性相符合, Spearman秩相关和 Pearson相关系数都小于1,表明该区域处于不稳定状态,群落结构和物种构成尚不完备,处于演替早期.综合χ ²检验、Pearson相关检验和Spearman秩相关检验,群落内主要物种多呈不显著联结,种对间联结程度较低,多呈独立空间分布[37 -38 ] .3种检验方法的结果均表明呈显著正联结的种对数大于显著负联结种对数,种对的显著率较低,说明群落演替初期物种间竞争较弱,种间联结较为分散,这在一定程度上可以缓和物种间对资源和空间的竞争关系,有利于演替初期物种的共存[9 ] .此外Pearson相关检验与Spearman秩相关检验相对的显著率明显高于χ ²检验.这是因为Pearson相关对数据的正态分布有较高的要求,而野外采集的数据难以保证呈正态分布.而 Spearman 秩相关分析与物种如何分布不相关,所以为补充相关系数检验的不足, 需引入Spearman 秩相关分析做补充.χ ²是基于二元数据的检验结果,虽能够较好地反映物种间关联的显著程度,但在反映物种间联结大小方面存在不足,还需结合Spearman秩相关及Pearson相关进行分析[32 -33 ] . ...
Ecological species groups and interspecific association of vegetation in natural recovery process at ejiadian landslide after 2008 Wenchuan earthquake
2
2016
... 通常认为重叠指数(Cik 和Oik )≥0.500,则种间具有较大的生态和资源利用相似性,竞争比较严重[33 -35 ] .种间生态位宽度与生态位重叠值一般呈正比.但本研究发现研究区出现物种的生态位宽度与生态位重叠呈反比的现象,例如角果藜和白车轴草等物种的生态位宽度较大,而生态位重叠程度较小.这可能是由天山北麓绿洲荒漠过渡带生境的脆弱与恶劣引起的,该区土壤分区差异性较大;其次不同空间区域内出现不同强度人为干扰破坏,造成部分地块土层浅薄、土壤含水量低和土壤养分差,不适应植物生长[29 ] ,物种间资源利用竞争激烈,某些物种为生存发育抢夺资源,导致对资源利用能力较差的物种多聚集在局部适宜的生境斑块中,其他空间分布较少,最终出现物种生态位宽度较小,但生态位重叠却较大[10 ] .这与陈龙涛等[36 ] 研究发现的现象一致.对于生态位重叠指数为0的物种,说明物种生态习性和对环境资源要求不同,不存在竞争关系.天山北麓绿洲荒漠过渡带约一半物种生态位重叠指数为0,这可能与土壤理化性质不同而形成的生境异质性有关.研究区多数优势物种间的生态位重叠指数较低,生态位分化比较明显,说明优势种占据的生境资源不在同一空间,对资源的需求差异较大,造成物种分布不连续,受外界人为破坏严重时对植物群落的破坏较为严重,使得不同优势种难以形成同一稳定的生存环境. ...
... 种间联系能很好地反映植物之间的相互作用,也能反映群落的动态变化.在次生演替的早期,物种之间正、负相关系数都很小,说明群落处在一个不稳定的时期,种间关系也是离散的.随演替进程的进行,种间连接趋向于正向连接,正-负连接比例增大,群落稳定性提高,进而向顶极演化.通过对天山北麓绿洲荒漠过渡带种间关系的测量,发现该地区主要植物的整体联结度呈极低的正相关性,与χ ²试验的正、负相关性相符合, Spearman秩相关和 Pearson相关系数都小于1,表明该区域处于不稳定状态,群落结构和物种构成尚不完备,处于演替早期.综合χ ²检验、Pearson相关检验和Spearman秩相关检验,群落内主要物种多呈不显著联结,种对间联结程度较低,多呈独立空间分布[37 -38 ] .3种检验方法的结果均表明呈显著正联结的种对数大于显著负联结种对数,种对的显著率较低,说明群落演替初期物种间竞争较弱,种间联结较为分散,这在一定程度上可以缓和物种间对资源和空间的竞争关系,有利于演替初期物种的共存[9 ] .此外Pearson相关检验与Spearman秩相关检验相对的显著率明显高于χ ²检验.这是因为Pearson相关对数据的正态分布有较高的要求,而野外采集的数据难以保证呈正态分布.而 Spearman 秩相关分析与物种如何分布不相关,所以为补充相关系数检验的不足, 需引入Spearman 秩相关分析做补充.χ ²是基于二元数据的检验结果,虽能够较好地反映物种间关联的显著程度,但在反映物种间联结大小方面存在不足,还需结合Spearman秩相关及Pearson相关进行分析[32 -33 ] . ...
Conserving biological diversity through ecosystem resilience
0
1995
濒危植物百花山葡萄群落物种生态位特征与种间联结研究
2
2020
... 通常认为重叠指数(Cik 和Oik )≥0.500,则种间具有较大的生态和资源利用相似性,竞争比较严重[33 -35 ] .种间生态位宽度与生态位重叠值一般呈正比.但本研究发现研究区出现物种的生态位宽度与生态位重叠呈反比的现象,例如角果藜和白车轴草等物种的生态位宽度较大,而生态位重叠程度较小.这可能是由天山北麓绿洲荒漠过渡带生境的脆弱与恶劣引起的,该区土壤分区差异性较大;其次不同空间区域内出现不同强度人为干扰破坏,造成部分地块土层浅薄、土壤含水量低和土壤养分差,不适应植物生长[29 ] ,物种间资源利用竞争激烈,某些物种为生存发育抢夺资源,导致对资源利用能力较差的物种多聚集在局部适宜的生境斑块中,其他空间分布较少,最终出现物种生态位宽度较小,但生态位重叠却较大[10 ] .这与陈龙涛等[36 ] 研究发现的现象一致.对于生态位重叠指数为0的物种,说明物种生态习性和对环境资源要求不同,不存在竞争关系.天山北麓绿洲荒漠过渡带约一半物种生态位重叠指数为0,这可能与土壤理化性质不同而形成的生境异质性有关.研究区多数优势物种间的生态位重叠指数较低,生态位分化比较明显,说明优势种占据的生境资源不在同一空间,对资源的需求差异较大,造成物种分布不连续,受外界人为破坏严重时对植物群落的破坏较为严重,使得不同优势种难以形成同一稳定的生存环境. ...
... 植物的种间连接关系与植物的生态位交叠关系紧密,它们之间的正向关系反映了植物的生态位重叠,负联结表明植物间呈生态位分离[35 ] .通过对天山北麓绿洲荒漠的研究,发现天山北麓绿洲荒漠过渡带主要物种间Pearson相关系数、Spearman秩相关系数与生态位重叠值回归分析均呈极显著正相关,即生态位重叠程度随种间正联结增加而增大.这与诸多研究结果一致[39 -41 ] ,若对研究区进行植被修复,选择显著正相关种对更有利于提高群落稳定性. ...
山西陵川南方红豆杉群落种间联结与生态位特征研究
1
2016
... 通常认为重叠指数(Cik 和Oik )≥0.500,则种间具有较大的生态和资源利用相似性,竞争比较严重[33 -35 ] .种间生态位宽度与生态位重叠值一般呈正比.但本研究发现研究区出现物种的生态位宽度与生态位重叠呈反比的现象,例如角果藜和白车轴草等物种的生态位宽度较大,而生态位重叠程度较小.这可能是由天山北麓绿洲荒漠过渡带生境的脆弱与恶劣引起的,该区土壤分区差异性较大;其次不同空间区域内出现不同强度人为干扰破坏,造成部分地块土层浅薄、土壤含水量低和土壤养分差,不适应植物生长[29 ] ,物种间资源利用竞争激烈,某些物种为生存发育抢夺资源,导致对资源利用能力较差的物种多聚集在局部适宜的生境斑块中,其他空间分布较少,最终出现物种生态位宽度较小,但生态位重叠却较大[10 ] .这与陈龙涛等[36 ] 研究发现的现象一致.对于生态位重叠指数为0的物种,说明物种生态习性和对环境资源要求不同,不存在竞争关系.天山北麓绿洲荒漠过渡带约一半物种生态位重叠指数为0,这可能与土壤理化性质不同而形成的生境异质性有关.研究区多数优势物种间的生态位重叠指数较低,生态位分化比较明显,说明优势种占据的生境资源不在同一空间,对资源的需求差异较大,造成物种分布不连续,受外界人为破坏严重时对植物群落的破坏较为严重,使得不同优势种难以形成同一稳定的生存环境. ...
滇中云南杨梅灌丛主要木本植物生态位与种间联结
1
2022
... 种间联系能很好地反映植物之间的相互作用,也能反映群落的动态变化.在次生演替的早期,物种之间正、负相关系数都很小,说明群落处在一个不稳定的时期,种间关系也是离散的.随演替进程的进行,种间连接趋向于正向连接,正-负连接比例增大,群落稳定性提高,进而向顶极演化.通过对天山北麓绿洲荒漠过渡带种间关系的测量,发现该地区主要植物的整体联结度呈极低的正相关性,与χ ²试验的正、负相关性相符合, Spearman秩相关和 Pearson相关系数都小于1,表明该区域处于不稳定状态,群落结构和物种构成尚不完备,处于演替早期.综合χ ²检验、Pearson相关检验和Spearman秩相关检验,群落内主要物种多呈不显著联结,种对间联结程度较低,多呈独立空间分布[37 -38 ] .3种检验方法的结果均表明呈显著正联结的种对数大于显著负联结种对数,种对的显著率较低,说明群落演替初期物种间竞争较弱,种间联结较为分散,这在一定程度上可以缓和物种间对资源和空间的竞争关系,有利于演替初期物种的共存[9 ] .此外Pearson相关检验与Spearman秩相关检验相对的显著率明显高于χ ²检验.这是因为Pearson相关对数据的正态分布有较高的要求,而野外采集的数据难以保证呈正态分布.而 Spearman 秩相关分析与物种如何分布不相关,所以为补充相关系数检验的不足, 需引入Spearman 秩相关分析做补充.χ ²是基于二元数据的检验结果,虽能够较好地反映物种间关联的显著程度,但在反映物种间联结大小方面存在不足,还需结合Spearman秩相关及Pearson相关进行分析[32 -33 ] . ...
滇西北云南红豆杉群落物种生态位与种间联结
1
2012
... 种间联系能很好地反映植物之间的相互作用,也能反映群落的动态变化.在次生演替的早期,物种之间正、负相关系数都很小,说明群落处在一个不稳定的时期,种间关系也是离散的.随演替进程的进行,种间连接趋向于正向连接,正-负连接比例增大,群落稳定性提高,进而向顶极演化.通过对天山北麓绿洲荒漠过渡带种间关系的测量,发现该地区主要植物的整体联结度呈极低的正相关性,与χ ²试验的正、负相关性相符合, Spearman秩相关和 Pearson相关系数都小于1,表明该区域处于不稳定状态,群落结构和物种构成尚不完备,处于演替早期.综合χ ²检验、Pearson相关检验和Spearman秩相关检验,群落内主要物种多呈不显著联结,种对间联结程度较低,多呈独立空间分布[37 -38 ] .3种检验方法的结果均表明呈显著正联结的种对数大于显著负联结种对数,种对的显著率较低,说明群落演替初期物种间竞争较弱,种间联结较为分散,这在一定程度上可以缓和物种间对资源和空间的竞争关系,有利于演替初期物种的共存[9 ] .此外Pearson相关检验与Spearman秩相关检验相对的显著率明显高于χ ²检验.这是因为Pearson相关对数据的正态分布有较高的要求,而野外采集的数据难以保证呈正态分布.而 Spearman 秩相关分析与物种如何分布不相关,所以为补充相关系数检验的不足, 需引入Spearman 秩相关分析做补充.χ ²是基于二元数据的检验结果,虽能够较好地反映物种间关联的显著程度,但在反映物种间联结大小方面存在不足,还需结合Spearman秩相关及Pearson相关进行分析[32 -33 ] . ...
长白山落叶阔叶混交林的物种多样性、种群空间分布格局及种间关联性研究
1
2004
... 植物的种间连接关系与植物的生态位交叠关系紧密,它们之间的正向关系反映了植物的生态位重叠,负联结表明植物间呈生态位分离[35 ] .通过对天山北麓绿洲荒漠的研究,发现天山北麓绿洲荒漠过渡带主要物种间Pearson相关系数、Spearman秩相关系数与生态位重叠值回归分析均呈极显著正相关,即生态位重叠程度随种间正联结增加而增大.这与诸多研究结果一致[39 -41 ] ,若对研究区进行植被修复,选择显著正相关种对更有利于提高群落稳定性. ...
赤水桫椤群落乔木层优势物种生态位与种间联结性研究
0
2017
黑龙江丰林国家级自然保护区红松+紫椴+硕桦群系优势树种生态位特征与种间联结分析
1
2023
... 植物的种间连接关系与植物的生态位交叠关系紧密,它们之间的正向关系反映了植物的生态位重叠,负联结表明植物间呈生态位分离[35 ] .通过对天山北麓绿洲荒漠的研究,发现天山北麓绿洲荒漠过渡带主要物种间Pearson相关系数、Spearman秩相关系数与生态位重叠值回归分析均呈极显著正相关,即生态位重叠程度随种间正联结增加而增大.这与诸多研究结果一致[39 -41 ] ,若对研究区进行植被修复,选择显著正相关种对更有利于提高群落稳定性. ...




 甘公网安备 62010202000688号
甘公网安备 62010202000688号
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}