Climate Change 2021:The Physical Science Basis-Summary for Policymakers.Contribution of Working Group I to the Sixth Assess-ment Report of the Intergovernmental Panel on Climate Change
1
2021
... 21世纪以来,全球陆地地表温度比20世纪下半叶增加了1.59 ℃[1].模型预测干旱区的增温幅度将会是湿润区的两倍,干旱程度也将持续增加[2],未来全球干旱半干旱区将很可能经历暖干化的气候变化过程[3].然而,过去57年(1961—2018年)中国西北地区的降水量整体上呈现为略有增加的趋势,且在空间和时间尺度上表现出较大的不确定性[4].这种以增温和降水变化为主要特征的全球气候变化将影响干旱生态系统的结构和功能,其中,尤为引人关注的科学问题是其如何影响碳循环关键过程而决定生态系统的碳收支,从而对气候变化产生反馈[5]. ...
Accelerated dryland expansion under climate change
1
2016
... 21世纪以来,全球陆地地表温度比20世纪下半叶增加了1.59 ℃[1].模型预测干旱区的增温幅度将会是湿润区的两倍,干旱程度也将持续增加[2],未来全球干旱半干旱区将很可能经历暖干化的气候变化过程[3].然而,过去57年(1961—2018年)中国西北地区的降水量整体上呈现为略有增加的趋势,且在空间和时间尺度上表现出较大的不确定性[4].这种以增温和降水变化为主要特征的全球气候变化将影响干旱生态系统的结构和功能,其中,尤为引人关注的科学问题是其如何影响碳循环关键过程而决定生态系统的碳收支,从而对气候变化产生反馈[5]. ...
Global warming and changes in drought
1
2014
... 21世纪以来,全球陆地地表温度比20世纪下半叶增加了1.59 ℃[1].模型预测干旱区的增温幅度将会是湿润区的两倍,干旱程度也将持续增加[2],未来全球干旱半干旱区将很可能经历暖干化的气候变化过程[3].然而,过去57年(1961—2018年)中国西北地区的降水量整体上呈现为略有增加的趋势,且在空间和时间尺度上表现出较大的不确定性[4].这种以增温和降水变化为主要特征的全球气候变化将影响干旱生态系统的结构和功能,其中,尤为引人关注的科学问题是其如何影响碳循环关键过程而决定生态系统的碳收支,从而对气候变化产生反馈[5]. ...
中国西北地区夏季降水及其东部降尺度预测模型
1
2023
... 21世纪以来,全球陆地地表温度比20世纪下半叶增加了1.59 ℃[1].模型预测干旱区的增温幅度将会是湿润区的两倍,干旱程度也将持续增加[2],未来全球干旱半干旱区将很可能经历暖干化的气候变化过程[3].然而,过去57年(1961—2018年)中国西北地区的降水量整体上呈现为略有增加的趋势,且在空间和时间尺度上表现出较大的不确定性[4].这种以增温和降水变化为主要特征的全球气候变化将影响干旱生态系统的结构和功能,其中,尤为引人关注的科学问题是其如何影响碳循环关键过程而决定生态系统的碳收支,从而对气候变化产生反馈[5]. ...
Quantifying global soil carbon losses in response to warming
1
2016
... 21世纪以来,全球陆地地表温度比20世纪下半叶增加了1.59 ℃[1].模型预测干旱区的增温幅度将会是湿润区的两倍,干旱程度也将持续增加[2],未来全球干旱半干旱区将很可能经历暖干化的气候变化过程[3].然而,过去57年(1961—2018年)中国西北地区的降水量整体上呈现为略有增加的趋势,且在空间和时间尺度上表现出较大的不确定性[4].这种以增温和降水变化为主要特征的全球气候变化将影响干旱生态系统的结构和功能,其中,尤为引人关注的科学问题是其如何影响碳循环关键过程而决定生态系统的碳收支,从而对气候变化产生反馈[5]. ...
生物土壤结皮研究:进展、前沿与展望
1
2009
... 生物结皮是由隐花植物、微小生物和土壤颗粒胶结而成的重要地表覆被物,在干旱半干旱区的盖度可高达70%[6],在维持该地区土壤肥力和稳定[7]、碳和氮的固定[8-9]、维管束植被定殖[10]、微小动物和微生物的栖息[11-12]、生态水文[13]等方面均发挥着不可替代的生态功能.就荒漠生态系统碳循环过程而言,生物结皮是重要的碳汇,通过其光合作用固定大气中的二氧化碳,全球年固碳量可达0.6 Pg[14];同时,生物结皮又以呼吸作用的形式释放碳而表现为重要的碳源.据估计,生物结皮呼吸碳排放占半干旱区42%~66%的总呼吸碳排放[15].因此,生物结皮对干旱半干旱区的碳收支与平衡具有重要意义. ...
Biological soil crusts decrease erodibility by modifying inherent soil properties on the Loess Plateau,China
1
2017
... 生物结皮是由隐花植物、微小生物和土壤颗粒胶结而成的重要地表覆被物,在干旱半干旱区的盖度可高达70%[6],在维持该地区土壤肥力和稳定[7]、碳和氮的固定[8-9]、维管束植被定殖[10]、微小动物和微生物的栖息[11-12]、生态水文[13]等方面均发挥着不可替代的生态功能.就荒漠生态系统碳循环过程而言,生物结皮是重要的碳汇,通过其光合作用固定大气中的二氧化碳,全球年固碳量可达0.6 Pg[14];同时,生物结皮又以呼吸作用的形式释放碳而表现为重要的碳源.据估计,生物结皮呼吸碳排放占半干旱区42%~66%的总呼吸碳排放[15].因此,生物结皮对干旱半干旱区的碳收支与平衡具有重要意义. ...
阿里荒漠区土壤有机碳分布特征及其与土壤物理性质的关系
1
2023
... 生物结皮是由隐花植物、微小生物和土壤颗粒胶结而成的重要地表覆被物,在干旱半干旱区的盖度可高达70%[6],在维持该地区土壤肥力和稳定[7]、碳和氮的固定[8-9]、维管束植被定殖[10]、微小动物和微生物的栖息[11-12]、生态水文[13]等方面均发挥着不可替代的生态功能.就荒漠生态系统碳循环过程而言,生物结皮是重要的碳汇,通过其光合作用固定大气中的二氧化碳,全球年固碳量可达0.6 Pg[14];同时,生物结皮又以呼吸作用的形式释放碳而表现为重要的碳源.据估计,生物结皮呼吸碳排放占半干旱区42%~66%的总呼吸碳排放[15].因此,生物结皮对干旱半干旱区的碳收支与平衡具有重要意义. ...
Biological soil crusts accelerate the nitrogen cycle through large NO and HONO emissions in drylands
1
2015
... 生物结皮是由隐花植物、微小生物和土壤颗粒胶结而成的重要地表覆被物,在干旱半干旱区的盖度可高达70%[6],在维持该地区土壤肥力和稳定[7]、碳和氮的固定[8-9]、维管束植被定殖[10]、微小动物和微生物的栖息[11-12]、生态水文[13]等方面均发挥着不可替代的生态功能.就荒漠生态系统碳循环过程而言,生物结皮是重要的碳汇,通过其光合作用固定大气中的二氧化碳,全球年固碳量可达0.6 Pg[14];同时,生物结皮又以呼吸作用的形式释放碳而表现为重要的碳源.据估计,生物结皮呼吸碳排放占半干旱区42%~66%的总呼吸碳排放[15].因此,生物结皮对干旱半干旱区的碳收支与平衡具有重要意义. ...
Biological soil crusts determine the germination and growth of two exotic plants
1
2017
... 生物结皮是由隐花植物、微小生物和土壤颗粒胶结而成的重要地表覆被物,在干旱半干旱区的盖度可高达70%[6],在维持该地区土壤肥力和稳定[7]、碳和氮的固定[8-9]、维管束植被定殖[10]、微小动物和微生物的栖息[11-12]、生态水文[13]等方面均发挥着不可替代的生态功能.就荒漠生态系统碳循环过程而言,生物结皮是重要的碳汇,通过其光合作用固定大气中的二氧化碳,全球年固碳量可达0.6 Pg[14];同时,生物结皮又以呼吸作用的形式释放碳而表现为重要的碳源.据估计,生物结皮呼吸碳排放占半干旱区42%~66%的总呼吸碳排放[15].因此,生物结皮对干旱半干旱区的碳收支与平衡具有重要意义. ...
Ants mediate soil water in arid desert ecosystems: mitigating rainfall interception induced by biological soil crusts?
1
2014
... 生物结皮是由隐花植物、微小生物和土壤颗粒胶结而成的重要地表覆被物,在干旱半干旱区的盖度可高达70%[6],在维持该地区土壤肥力和稳定[7]、碳和氮的固定[8-9]、维管束植被定殖[10]、微小动物和微生物的栖息[11-12]、生态水文[13]等方面均发挥着不可替代的生态功能.就荒漠生态系统碳循环过程而言,生物结皮是重要的碳汇,通过其光合作用固定大气中的二氧化碳,全球年固碳量可达0.6 Pg[14];同时,生物结皮又以呼吸作用的形式释放碳而表现为重要的碳源.据估计,生物结皮呼吸碳排放占半干旱区42%~66%的总呼吸碳排放[15].因此,生物结皮对干旱半干旱区的碳收支与平衡具有重要意义. ...
Shifts in soil microbial community functional gene structure across a 61-year desert revegetation chronosequence
3
2019
... 生物结皮是由隐花植物、微小生物和土壤颗粒胶结而成的重要地表覆被物,在干旱半干旱区的盖度可高达70%[6],在维持该地区土壤肥力和稳定[7]、碳和氮的固定[8-9]、维管束植被定殖[10]、微小动物和微生物的栖息[11-12]、生态水文[13]等方面均发挥着不可替代的生态功能.就荒漠生态系统碳循环过程而言,生物结皮是重要的碳汇,通过其光合作用固定大气中的二氧化碳,全球年固碳量可达0.6 Pg[14];同时,生物结皮又以呼吸作用的形式释放碳而表现为重要的碳源.据估计,生物结皮呼吸碳排放占半干旱区42%~66%的总呼吸碳排放[15].因此,生物结皮对干旱半干旱区的碳收支与平衡具有重要意义. ...
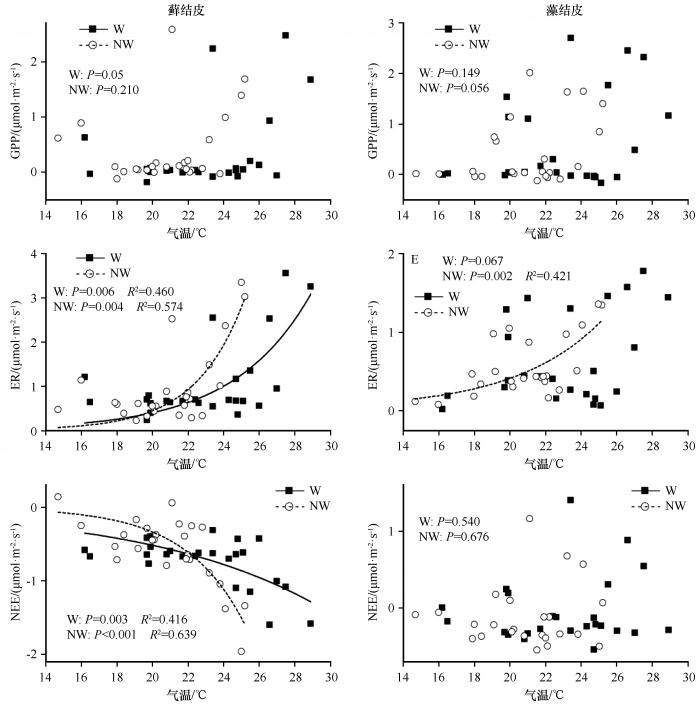
... 生物结皮是干旱半干旱陆地生态系统碳循环的重要组成部分[27].荒漠地区的生物结皮通常由早期的蓝藻结皮逐渐向地衣结皮和藓结皮演替[28],并伴随着土壤养分水平、隐花植物和微生物的多样性及生物量的增加[12,29].我们的研究结果表明,增温与不增温条件下藓结皮与藻结皮的GPP无显著差异,这与毛乌素沙地藓结皮光合速率是藻结皮2倍的研究报道并不一致[30].这可能是试验期内的高温导致结皮层水分迅速蒸发,从而缩短了其有效光合时间.藓结皮有更高的隐花植物、土壤微生物生物量和更为丰富的碳源[31],自养[14]和异养呼吸[29]均高于藻结皮,特别是在中、高降水强度下,藓结皮比藻结皮有更高的呼吸碳排放(图3),其更高的碳分解相关功能基因也为此提供了佐证[12]. ...
... [12]. ...
Reference for different sensitivities of greenhouse gases effluxes to warming climate among types of desert biological soil crust
1
2022
... 生物结皮是由隐花植物、微小生物和土壤颗粒胶结而成的重要地表覆被物,在干旱半干旱区的盖度可高达70%[6],在维持该地区土壤肥力和稳定[7]、碳和氮的固定[8-9]、维管束植被定殖[10]、微小动物和微生物的栖息[11-12]、生态水文[13]等方面均发挥着不可替代的生态功能.就荒漠生态系统碳循环过程而言,生物结皮是重要的碳汇,通过其光合作用固定大气中的二氧化碳,全球年固碳量可达0.6 Pg[14];同时,生物结皮又以呼吸作用的形式释放碳而表现为重要的碳源.据估计,生物结皮呼吸碳排放占半干旱区42%~66%的总呼吸碳排放[15].因此,生物结皮对干旱半干旱区的碳收支与平衡具有重要意义. ...
生物土壤结皮固沙理论与实践
3
2021
... 生物结皮是由隐花植物、微小生物和土壤颗粒胶结而成的重要地表覆被物,在干旱半干旱区的盖度可高达70%[6],在维持该地区土壤肥力和稳定[7]、碳和氮的固定[8-9]、维管束植被定殖[10]、微小动物和微生物的栖息[11-12]、生态水文[13]等方面均发挥着不可替代的生态功能.就荒漠生态系统碳循环过程而言,生物结皮是重要的碳汇,通过其光合作用固定大气中的二氧化碳,全球年固碳量可达0.6 Pg[14];同时,生物结皮又以呼吸作用的形式释放碳而表现为重要的碳源.据估计,生物结皮呼吸碳排放占半干旱区42%~66%的总呼吸碳排放[15].因此,生物结皮对干旱半干旱区的碳收支与平衡具有重要意义. ...
... 2014年6月,用直径为12 cm的PVC管采集藓结皮和藻结皮覆盖的12个10 cm深的原状土(2个处理×2种结皮类型×3个重复),带回沙坡头站实验室后放置30天,以保证土壤初始水分含量基本一致.试验以开顶式生长室(OTC)模拟增温,OTC边长为1.3 m、高2 m的等边八边体[14],设置增温(W)和不增温(NW)两个温度处理.根据研究区近10年的降水事件,设置2、5、8、13、20 mm共5个降水梯度,分别代表小降水事件(2、5 mm)、中降水事件(8 mm)以及大降水事件(13和20 mm).分别在OTC内部和外部搭建透明遮雨棚(长×宽×高=1 m×1 m×1.2 m),将所有样品置于遮雨棚下以避免自然降水差异的干扰.采用人工定量均匀喷洒蒸馏水模拟降水量变化,在每次模拟降水前用称重法(精度0.01)称取原状土质量,待土壤含水量接近初始值后进行模拟降水.OTC内、外分别安装了两套HOBO U30微型气象站(Onset Computer Corporation,Cape Cod,USA),每30 min测定气温和10 cm土壤湿度. ...
... 生物结皮是干旱半干旱陆地生态系统碳循环的重要组成部分[27].荒漠地区的生物结皮通常由早期的蓝藻结皮逐渐向地衣结皮和藓结皮演替[28],并伴随着土壤养分水平、隐花植物和微生物的多样性及生物量的增加[12,29].我们的研究结果表明,增温与不增温条件下藓结皮与藻结皮的GPP无显著差异,这与毛乌素沙地藓结皮光合速率是藻结皮2倍的研究报道并不一致[30].这可能是试验期内的高温导致结皮层水分迅速蒸发,从而缩短了其有效光合时间.藓结皮有更高的隐花植物、土壤微生物生物量和更为丰富的碳源[31],自养[14]和异养呼吸[29]均高于藻结皮,特别是在中、高降水强度下,藓结皮比藻结皮有更高的呼吸碳排放(图3),其更高的碳分解相关功能基因也为此提供了佐证[12]. ...
Biological soil crust microsites are the main contributor to soil respiration in a semiarid ecosystem
1
2011
... 生物结皮是由隐花植物、微小生物和土壤颗粒胶结而成的重要地表覆被物,在干旱半干旱区的盖度可高达70%[6],在维持该地区土壤肥力和稳定[7]、碳和氮的固定[8-9]、维管束植被定殖[10]、微小动物和微生物的栖息[11-12]、生态水文[13]等方面均发挥着不可替代的生态功能.就荒漠生态系统碳循环过程而言,生物结皮是重要的碳汇,通过其光合作用固定大气中的二氧化碳,全球年固碳量可达0.6 Pg[14];同时,生物结皮又以呼吸作用的形式释放碳而表现为重要的碳源.据估计,生物结皮呼吸碳排放占半干旱区42%~66%的总呼吸碳排放[15].因此,生物结皮对干旱半干旱区的碳收支与平衡具有重要意义. ...
Carbon fixation by biological soil crusts following revegetation of sand dunes in arid desert regions of China:a four-year field study
3
2012
... 荒漠生物结皮的固碳量与碳排放量和结皮类型、降水事件和温度密切相关[16-18].通常情况下,处于演替后期的藓结皮年固碳量明显高于早期的藻结皮[16].目前,许多学者研究了模拟增温、不同降水量、降水频率等单一环境因子对荒漠生物结皮总初级生产力、生态系统呼吸或净碳交换量的影响[19-21].大多研究表明降水增加会同时促进生物结皮的碳固定和排放[16,19-21],而高频率小降水事件更能刺激土壤呼吸而促进碳排放[21].一项最新的研究表明,增温和降雨频率对干旱半干旱区土壤呼吸没有显著交互作用[21],但增温也往往会因加剧干旱效应而抑制土壤呼吸,从而导致碳排放减少[22].荒漠约占全球陆地面积的41%[23],因此,增温与不同降水量及其互作效应如何影响不同类型荒漠生物结皮的碳交换关键过程关系到该区域乃至全球的碳平衡,但有关这方面的试验研究还比较欠缺,导致难以准确地预测未来全球变化背景下荒漠生态系统的碳收支与平衡. ...
... [16].目前,许多学者研究了模拟增温、不同降水量、降水频率等单一环境因子对荒漠生物结皮总初级生产力、生态系统呼吸或净碳交换量的影响[19-21].大多研究表明降水增加会同时促进生物结皮的碳固定和排放[16,19-21],而高频率小降水事件更能刺激土壤呼吸而促进碳排放[21].一项最新的研究表明,增温和降雨频率对干旱半干旱区土壤呼吸没有显著交互作用[21],但增温也往往会因加剧干旱效应而抑制土壤呼吸,从而导致碳排放减少[22].荒漠约占全球陆地面积的41%[23],因此,增温与不同降水量及其互作效应如何影响不同类型荒漠生物结皮的碳交换关键过程关系到该区域乃至全球的碳平衡,但有关这方面的试验研究还比较欠缺,导致难以准确地预测未来全球变化背景下荒漠生态系统的碳收支与平衡. ...
... [16,19-21],而高频率小降水事件更能刺激土壤呼吸而促进碳排放[21].一项最新的研究表明,增温和降雨频率对干旱半干旱区土壤呼吸没有显著交互作用[21],但增温也往往会因加剧干旱效应而抑制土壤呼吸,从而导致碳排放减少[22].荒漠约占全球陆地面积的41%[23],因此,增温与不同降水量及其互作效应如何影响不同类型荒漠生物结皮的碳交换关键过程关系到该区域乃至全球的碳平衡,但有关这方面的试验研究还比较欠缺,导致难以准确地预测未来全球变化背景下荒漠生态系统的碳收支与平衡. ...
Soil CO2 exchange controlled by the interaction of biocrust successional stage and environmental variables in two semiarid ecosystems
0
2018
Soil respiration sensitivities to water and temperature in a revegetated desert
1
2015
... 荒漠生物结皮的固碳量与碳排放量和结皮类型、降水事件和温度密切相关[16-18].通常情况下,处于演替后期的藓结皮年固碳量明显高于早期的藻结皮[16].目前,许多学者研究了模拟增温、不同降水量、降水频率等单一环境因子对荒漠生物结皮总初级生产力、生态系统呼吸或净碳交换量的影响[19-21].大多研究表明降水增加会同时促进生物结皮的碳固定和排放[16,19-21],而高频率小降水事件更能刺激土壤呼吸而促进碳排放[21].一项最新的研究表明,增温和降雨频率对干旱半干旱区土壤呼吸没有显著交互作用[21],但增温也往往会因加剧干旱效应而抑制土壤呼吸,从而导致碳排放减少[22].荒漠约占全球陆地面积的41%[23],因此,增温与不同降水量及其互作效应如何影响不同类型荒漠生物结皮的碳交换关键过程关系到该区域乃至全球的碳平衡,但有关这方面的试验研究还比较欠缺,导致难以准确地预测未来全球变化背景下荒漠生态系统的碳收支与平衡. ...
Carbon flux in deserts depends on soil cover type: a case study in the Gurbantunggute desert,North China
2
2013
... 荒漠生物结皮的固碳量与碳排放量和结皮类型、降水事件和温度密切相关[16-18].通常情况下,处于演替后期的藓结皮年固碳量明显高于早期的藻结皮[16].目前,许多学者研究了模拟增温、不同降水量、降水频率等单一环境因子对荒漠生物结皮总初级生产力、生态系统呼吸或净碳交换量的影响[19-21].大多研究表明降水增加会同时促进生物结皮的碳固定和排放[16,19-21],而高频率小降水事件更能刺激土壤呼吸而促进碳排放[21].一项最新的研究表明,增温和降雨频率对干旱半干旱区土壤呼吸没有显著交互作用[21],但增温也往往会因加剧干旱效应而抑制土壤呼吸,从而导致碳排放减少[22].荒漠约占全球陆地面积的41%[23],因此,增温与不同降水量及其互作效应如何影响不同类型荒漠生物结皮的碳交换关键过程关系到该区域乃至全球的碳平衡,但有关这方面的试验研究还比较欠缺,导致难以准确地预测未来全球变化背景下荒漠生态系统的碳收支与平衡. ...
... ,19-21],而高频率小降水事件更能刺激土壤呼吸而促进碳排放[21].一项最新的研究表明,增温和降雨频率对干旱半干旱区土壤呼吸没有显著交互作用[21],但增温也往往会因加剧干旱效应而抑制土壤呼吸,从而导致碳排放减少[22].荒漠约占全球陆地面积的41%[23],因此,增温与不同降水量及其互作效应如何影响不同类型荒漠生物结皮的碳交换关键过程关系到该区域乃至全球的碳平衡,但有关这方面的试验研究还比较欠缺,导致难以准确地预测未来全球变化背景下荒漠生态系统的碳收支与平衡. ...
Evolution of soil respiration depends on biological soil crusts across a 50-year chronosequence of desert revegetation
0
2016
模拟增温和改变降雨频率对干旱半干旱区土壤呼吸的影响
4
2021
... 荒漠生物结皮的固碳量与碳排放量和结皮类型、降水事件和温度密切相关[16-18].通常情况下,处于演替后期的藓结皮年固碳量明显高于早期的藻结皮[16].目前,许多学者研究了模拟增温、不同降水量、降水频率等单一环境因子对荒漠生物结皮总初级生产力、生态系统呼吸或净碳交换量的影响[19-21].大多研究表明降水增加会同时促进生物结皮的碳固定和排放[16,19-21],而高频率小降水事件更能刺激土壤呼吸而促进碳排放[21].一项最新的研究表明,增温和降雨频率对干旱半干旱区土壤呼吸没有显著交互作用[21],但增温也往往会因加剧干旱效应而抑制土壤呼吸,从而导致碳排放减少[22].荒漠约占全球陆地面积的41%[23],因此,增温与不同降水量及其互作效应如何影响不同类型荒漠生物结皮的碳交换关键过程关系到该区域乃至全球的碳平衡,但有关这方面的试验研究还比较欠缺,导致难以准确地预测未来全球变化背景下荒漠生态系统的碳收支与平衡. ...
... -21],而高频率小降水事件更能刺激土壤呼吸而促进碳排放[21].一项最新的研究表明,增温和降雨频率对干旱半干旱区土壤呼吸没有显著交互作用[21],但增温也往往会因加剧干旱效应而抑制土壤呼吸,从而导致碳排放减少[22].荒漠约占全球陆地面积的41%[23],因此,增温与不同降水量及其互作效应如何影响不同类型荒漠生物结皮的碳交换关键过程关系到该区域乃至全球的碳平衡,但有关这方面的试验研究还比较欠缺,导致难以准确地预测未来全球变化背景下荒漠生态系统的碳收支与平衡. ...
... [21].一项最新的研究表明,增温和降雨频率对干旱半干旱区土壤呼吸没有显著交互作用[21],但增温也往往会因加剧干旱效应而抑制土壤呼吸,从而导致碳排放减少[22].荒漠约占全球陆地面积的41%[23],因此,增温与不同降水量及其互作效应如何影响不同类型荒漠生物结皮的碳交换关键过程关系到该区域乃至全球的碳平衡,但有关这方面的试验研究还比较欠缺,导致难以准确地预测未来全球变化背景下荒漠生态系统的碳收支与平衡. ...
... [21],但增温也往往会因加剧干旱效应而抑制土壤呼吸,从而导致碳排放减少[22].荒漠约占全球陆地面积的41%[23],因此,增温与不同降水量及其互作效应如何影响不同类型荒漠生物结皮的碳交换关键过程关系到该区域乃至全球的碳平衡,但有关这方面的试验研究还比较欠缺,导致难以准确地预测未来全球变化背景下荒漠生态系统的碳收支与平衡. ...
Effects of warming and rainfall pulses on soil respiration in a biological soil crust-dominated desert ecosystem
1
2021
... 荒漠生物结皮的固碳量与碳排放量和结皮类型、降水事件和温度密切相关[16-18].通常情况下,处于演替后期的藓结皮年固碳量明显高于早期的藻结皮[16].目前,许多学者研究了模拟增温、不同降水量、降水频率等单一环境因子对荒漠生物结皮总初级生产力、生态系统呼吸或净碳交换量的影响[19-21].大多研究表明降水增加会同时促进生物结皮的碳固定和排放[16,19-21],而高频率小降水事件更能刺激土壤呼吸而促进碳排放[21].一项最新的研究表明,增温和降雨频率对干旱半干旱区土壤呼吸没有显著交互作用[21],但增温也往往会因加剧干旱效应而抑制土壤呼吸,从而导致碳排放减少[22].荒漠约占全球陆地面积的41%[23],因此,增温与不同降水量及其互作效应如何影响不同类型荒漠生物结皮的碳交换关键过程关系到该区域乃至全球的碳平衡,但有关这方面的试验研究还比较欠缺,导致难以准确地预测未来全球变化背景下荒漠生态系统的碳收支与平衡. ...
Aridity-driven shift in biodiversity-soil multifunctionality relationships
1
2021
... 荒漠生物结皮的固碳量与碳排放量和结皮类型、降水事件和温度密切相关[16-18].通常情况下,处于演替后期的藓结皮年固碳量明显高于早期的藻结皮[16].目前,许多学者研究了模拟增温、不同降水量、降水频率等单一环境因子对荒漠生物结皮总初级生产力、生态系统呼吸或净碳交换量的影响[19-21].大多研究表明降水增加会同时促进生物结皮的碳固定和排放[16,19-21],而高频率小降水事件更能刺激土壤呼吸而促进碳排放[21].一项最新的研究表明,增温和降雨频率对干旱半干旱区土壤呼吸没有显著交互作用[21],但增温也往往会因加剧干旱效应而抑制土壤呼吸,从而导致碳排放减少[22].荒漠约占全球陆地面积的41%[23],因此,增温与不同降水量及其互作效应如何影响不同类型荒漠生物结皮的碳交换关键过程关系到该区域乃至全球的碳平衡,但有关这方面的试验研究还比较欠缺,导致难以准确地预测未来全球变化背景下荒漠生态系统的碳收支与平衡. ...
半荒漠藻结皮中藻类的种类组成和分布
1
2000
... 研究区位于宁夏回族自治区中国科学院沙坡头沙漠研究试验站(简称沙坡头站)以西30 km的一碗泉天然植被区(37°29′N,104°25′E).该地区位于腾格里沙漠东南缘,属于草原化荒漠和荒漠化草原的过渡带,海拔1 339 m,年平均气温10.6 ℃,多年平均降水量为186 mm,降水主要在5—9月,年蒸发量约为2 900 mm.土壤基质为松散贫瘠的风沙土.生态系统类型为沙生灌木和半灌木与生物结皮镶嵌分布的典型荒漠,主要优势灌木和半灌木为柠条(Caraganakorshinskii)、油蒿(Artemisia ordosica)和狭叶锦鸡儿(Caragana stenophylla),优势草本植物包括刺沙蓬(Salsola ruthenica)、小画眉草(Eragrostis minor)和茵陈蒿(Artemisia capillaris).生物结皮以藓结皮、藻结皮和地衣混生为主,其盖度超过80%,优势种为真藓(Bryum argenteum)、尖叶对齿藓(Didymodon constrictus)和土生对齿藓(Didymodon vinealis)等;藻结皮优势种包括具鞘微鞘藻(Microcoleus vaginatus)、泥褐席藻(Phormidium luridum)、隐鞘鞘丝藻(Lyngbya cryptovaginatus)等;地衣优势种有坚韧胶衣(Collema tenax)、石果衣(Endocarpon pusillum)、藓生双缘衣(Diploschistes muscorum)等[24-25]. ...
The effects of sand stabilization and revegetation on cryptogam species diversity and soil fertility in the Tengger Desert,Northern China
1
2003
... 研究区位于宁夏回族自治区中国科学院沙坡头沙漠研究试验站(简称沙坡头站)以西30 km的一碗泉天然植被区(37°29′N,104°25′E).该地区位于腾格里沙漠东南缘,属于草原化荒漠和荒漠化草原的过渡带,海拔1 339 m,年平均气温10.6 ℃,多年平均降水量为186 mm,降水主要在5—9月,年蒸发量约为2 900 mm.土壤基质为松散贫瘠的风沙土.生态系统类型为沙生灌木和半灌木与生物结皮镶嵌分布的典型荒漠,主要优势灌木和半灌木为柠条(Caraganakorshinskii)、油蒿(Artemisia ordosica)和狭叶锦鸡儿(Caragana stenophylla),优势草本植物包括刺沙蓬(Salsola ruthenica)、小画眉草(Eragrostis minor)和茵陈蒿(Artemisia capillaris).生物结皮以藓结皮、藻结皮和地衣混生为主,其盖度超过80%,优势种为真藓(Bryum argenteum)、尖叶对齿藓(Didymodon constrictus)和土生对齿藓(Didymodon vinealis)等;藻结皮优势种包括具鞘微鞘藻(Microcoleus vaginatus)、泥褐席藻(Phormidium luridum)、隐鞘鞘丝藻(Lyngbya cryptovaginatus)等;地衣优势种有坚韧胶衣(Collema tenax)、石果衣(Endocarpon pusillum)、藓生双缘衣(Diploschistes muscorum)等[24-25]. ...
Temperature sensitivity of soil carbon decompositi on and feedbacks to climate change
1
2006
... 利用Li-6400(Li-cor,Inc.,美国)连接自制的透明叶室监测生物结皮的净生态系统碳交换量(NEE).然后,用遮光布将叶室完全遮严避光,测定其生态系统呼吸(ER).测定时间为09:00—12:00,每24 h测定1次,连续监测1个月.总初级生产力(GPP)为ER与NEE之差,即GPP=ER-NEE.用GPP、ER和NEE与温度间的指数方程系数计算其温度敏感性(Q10)[26]. ...
Biological soil crusts:structure,function,and management
1
2003
... 生物结皮是干旱半干旱陆地生态系统碳循环的重要组成部分[27].荒漠地区的生物结皮通常由早期的蓝藻结皮逐渐向地衣结皮和藓结皮演替[28],并伴随着土壤养分水平、隐花植物和微生物的多样性及生物量的增加[12,29].我们的研究结果表明,增温与不增温条件下藓结皮与藻结皮的GPP无显著差异,这与毛乌素沙地藓结皮光合速率是藻结皮2倍的研究报道并不一致[30].这可能是试验期内的高温导致结皮层水分迅速蒸发,从而缩短了其有效光合时间.藓结皮有更高的隐花植物、土壤微生物生物量和更为丰富的碳源[31],自养[14]和异养呼吸[29]均高于藻结皮,特别是在中、高降水强度下,藓结皮比藻结皮有更高的呼吸碳排放(图3),其更高的碳分解相关功能基因也为此提供了佐证[12]. ...
1
2012
... 生物结皮是干旱半干旱陆地生态系统碳循环的重要组成部分[27].荒漠地区的生物结皮通常由早期的蓝藻结皮逐渐向地衣结皮和藓结皮演替[28],并伴随着土壤养分水平、隐花植物和微生物的多样性及生物量的增加[12,29].我们的研究结果表明,增温与不增温条件下藓结皮与藻结皮的GPP无显著差异,这与毛乌素沙地藓结皮光合速率是藻结皮2倍的研究报道并不一致[30].这可能是试验期内的高温导致结皮层水分迅速蒸发,从而缩短了其有效光合时间.藓结皮有更高的隐花植物、土壤微生物生物量和更为丰富的碳源[31],自养[14]和异养呼吸[29]均高于藻结皮,特别是在中、高降水强度下,藓结皮比藻结皮有更高的呼吸碳排放(图3),其更高的碳分解相关功能基因也为此提供了佐证[12]. ...
The seasonal and successional variations of carbon release from biological soil crust-covered soil
2
2016
... 生物结皮是干旱半干旱陆地生态系统碳循环的重要组成部分[27].荒漠地区的生物结皮通常由早期的蓝藻结皮逐渐向地衣结皮和藓结皮演替[28],并伴随着土壤养分水平、隐花植物和微生物的多样性及生物量的增加[12,29].我们的研究结果表明,增温与不增温条件下藓结皮与藻结皮的GPP无显著差异,这与毛乌素沙地藓结皮光合速率是藻结皮2倍的研究报道并不一致[30].这可能是试验期内的高温导致结皮层水分迅速蒸发,从而缩短了其有效光合时间.藓结皮有更高的隐花植物、土壤微生物生物量和更为丰富的碳源[31],自养[14]和异养呼吸[29]均高于藻结皮,特别是在中、高降水强度下,藓结皮比藻结皮有更高的呼吸碳排放(图3),其更高的碳分解相关功能基因也为此提供了佐证[12]. ...
... [29]均高于藻结皮,特别是在中、高降水强度下,藓结皮比藻结皮有更高的呼吸碳排放(图3),其更高的碳分解相关功能基因也为此提供了佐证[12]. ...
毛乌素沙地生物结皮的光合及土壤CO2通量特征研究
1
2019
... 生物结皮是干旱半干旱陆地生态系统碳循环的重要组成部分[27].荒漠地区的生物结皮通常由早期的蓝藻结皮逐渐向地衣结皮和藓结皮演替[28],并伴随着土壤养分水平、隐花植物和微生物的多样性及生物量的增加[12,29].我们的研究结果表明,增温与不增温条件下藓结皮与藻结皮的GPP无显著差异,这与毛乌素沙地藓结皮光合速率是藻结皮2倍的研究报道并不一致[30].这可能是试验期内的高温导致结皮层水分迅速蒸发,从而缩短了其有效光合时间.藓结皮有更高的隐花植物、土壤微生物生物量和更为丰富的碳源[31],自养[14]和异养呼吸[29]均高于藻结皮,特别是在中、高降水强度下,藓结皮比藻结皮有更高的呼吸碳排放(图3),其更高的碳分解相关功能基因也为此提供了佐证[12]. ...
腾格里沙漠东南缘生物结皮土壤呼吸对水热因子变化的响应
1
2017
... 生物结皮是干旱半干旱陆地生态系统碳循环的重要组成部分[27].荒漠地区的生物结皮通常由早期的蓝藻结皮逐渐向地衣结皮和藓结皮演替[28],并伴随着土壤养分水平、隐花植物和微生物的多样性及生物量的增加[12,29].我们的研究结果表明,增温与不增温条件下藓结皮与藻结皮的GPP无显著差异,这与毛乌素沙地藓结皮光合速率是藻结皮2倍的研究报道并不一致[30].这可能是试验期内的高温导致结皮层水分迅速蒸发,从而缩短了其有效光合时间.藓结皮有更高的隐花植物、土壤微生物生物量和更为丰富的碳源[31],自养[14]和异养呼吸[29]均高于藻结皮,特别是在中、高降水强度下,藓结皮比藻结皮有更高的呼吸碳排放(图3),其更高的碳分解相关功能基因也为此提供了佐证[12]. ...
The mechanism of soil nitrogen transformation under different biocrusts to warming and reduced precipitation:from microbial functional genes to enzyme activity
1
2020
... 除了藓结皮NEE外,增温对两类生物结皮的GPP、ER和NEE没有显著影响(表1).增温加速了土壤水分的散失,缩短了其有效光合时间,由于藻结皮层更薄,其持水力不及藓结皮,水分散失更加强烈(图1).增温条件下藓结皮的GPP显著降低(图2),表明藓结皮作为演替后期的结皮类型,未来的全球变暖将可能更容易限制其光合固碳潜力.相比而言,藻结皮恢复光合作用所需的降水量更少,增温也会刺激其光合效率,从而促进其光合固碳有所增加(图3).生物结皮的ER来源于隐花植物的自养呼吸和异养呼吸.虽然增温往往会通过刺激土壤酶的活性而加快有机质的分解[32],从而促进土壤微生物呼吸致使碳排放增加,但荒漠地区有机质的分解速率和土壤养分有效性还受水分的强烈限制[33].增温不仅促使隐花植物更快地进入休眠状态,限制其自养呼吸的碳排放,还降低了土壤水分的有效性[34],从而抑制了土壤微生物的活性.因此,增温对生物结皮呼吸碳排放的净效应取决于其对呼吸的刺激作用和水分有效性的限制作用之间的相互折中,但在大多数情况下(除13 mm藻结皮),增温促使呼吸碳排放增加,表明未来全球气候变暖可能会通过刺激荒漠生物结皮-土壤生态系统呼吸而引发碳排放. ...
Effect of warming and grazing on litter mass loss and temperature sensitivity of litter and dung mass loss on the Tibetan plateau
1
2010
... 除了藓结皮NEE外,增温对两类生物结皮的GPP、ER和NEE没有显著影响(表1).增温加速了土壤水分的散失,缩短了其有效光合时间,由于藻结皮层更薄,其持水力不及藓结皮,水分散失更加强烈(图1).增温条件下藓结皮的GPP显著降低(图2),表明藓结皮作为演替后期的结皮类型,未来的全球变暖将可能更容易限制其光合固碳潜力.相比而言,藻结皮恢复光合作用所需的降水量更少,增温也会刺激其光合效率,从而促进其光合固碳有所增加(图3).生物结皮的ER来源于隐花植物的自养呼吸和异养呼吸.虽然增温往往会通过刺激土壤酶的活性而加快有机质的分解[32],从而促进土壤微生物呼吸致使碳排放增加,但荒漠地区有机质的分解速率和土壤养分有效性还受水分的强烈限制[33].增温不仅促使隐花植物更快地进入休眠状态,限制其自养呼吸的碳排放,还降低了土壤水分的有效性[34],从而抑制了土壤微生物的活性.因此,增温对生物结皮呼吸碳排放的净效应取决于其对呼吸的刺激作用和水分有效性的限制作用之间的相互折中,但在大多数情况下(除13 mm藻结皮),增温促使呼吸碳排放增加,表明未来全球气候变暖可能会通过刺激荒漠生物结皮-土壤生态系统呼吸而引发碳排放. ...
Warming and increased precipitation frequency on the Colorado Plateau:implications for biological soil crusts and soil processes
1
2012
... 除了藓结皮NEE外,增温对两类生物结皮的GPP、ER和NEE没有显著影响(表1).增温加速了土壤水分的散失,缩短了其有效光合时间,由于藻结皮层更薄,其持水力不及藓结皮,水分散失更加强烈(图1).增温条件下藓结皮的GPP显著降低(图2),表明藓结皮作为演替后期的结皮类型,未来的全球变暖将可能更容易限制其光合固碳潜力.相比而言,藻结皮恢复光合作用所需的降水量更少,增温也会刺激其光合效率,从而促进其光合固碳有所增加(图3).生物结皮的ER来源于隐花植物的自养呼吸和异养呼吸.虽然增温往往会通过刺激土壤酶的活性而加快有机质的分解[32],从而促进土壤微生物呼吸致使碳排放增加,但荒漠地区有机质的分解速率和土壤养分有效性还受水分的强烈限制[33].增温不仅促使隐花植物更快地进入休眠状态,限制其自养呼吸的碳排放,还降低了土壤水分的有效性[34],从而抑制了土壤微生物的活性.因此,增温对生物结皮呼吸碳排放的净效应取决于其对呼吸的刺激作用和水分有效性的限制作用之间的相互折中,但在大多数情况下(除13 mm藻结皮),增温促使呼吸碳排放增加,表明未来全球气候变暖可能会通过刺激荒漠生物结皮-土壤生态系统呼吸而引发碳排放. ...
Biological soil crusts as an organizing principle in drylands
2
2016
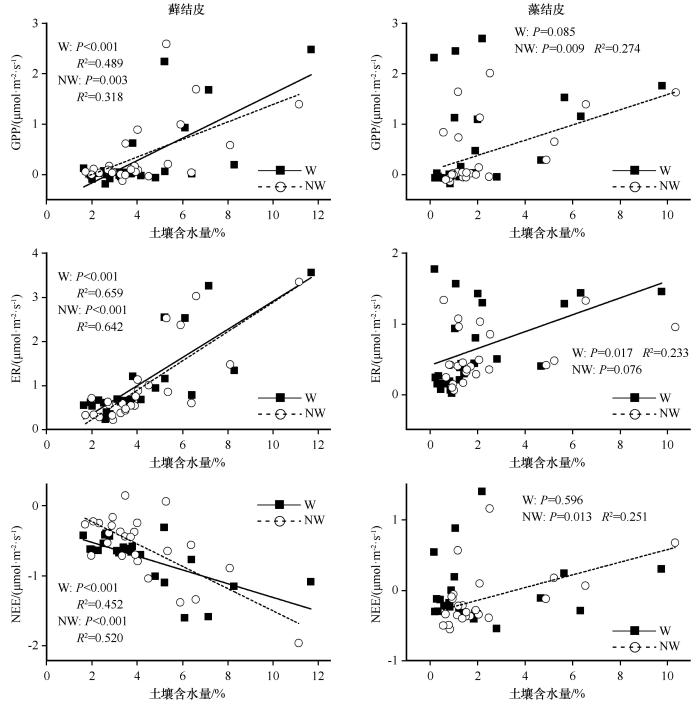
... 生物结皮属于典型的变水植物,在水分低于一定的阈值时处于代谢不活跃的休眠状态,而在降水后迅速恢复生理活性[35-37].我们发现,降水量极显著(P<0.001)影响两种生物结皮的GPP、ER和NEE,而增温和降水量之间无交互作用(表1).无论降水强度如何,生物结皮-土壤系统GPP和ER均在模拟降水后达到最大值,表现为光合固碳和呼吸碳排放的脉冲现象(图2),这主要源于恢复活性的隐花植物的光合固碳和自养呼吸以及异养呼吸的迅速增加[35,37-39].两类生物结皮的累积GPP和ER随着降水量的增加而线性增加,表明降水量越大,生物结皮的光合和呼吸作用的持续时间就越长.尽管降水会引发生物结皮较强的光合和呼吸作用,但在炎热的夏季,土壤水分会快速散失,水分可利用性的下降也致使GPP和ER迅速下降(图2),这些结果表明水分有效性对荒漠生物结皮的碳固定和排放具有决定性作用. ...
... [35,37-39].两类生物结皮的累积GPP和ER随着降水量的增加而线性增加,表明降水量越大,生物结皮的光合和呼吸作用的持续时间就越长.尽管降水会引发生物结皮较强的光合和呼吸作用,但在炎热的夏季,土壤水分会快速散失,水分可利用性的下降也致使GPP和ER迅速下降(图2),这些结果表明水分有效性对荒漠生物结皮的碳固定和排放具有决定性作用. ...
Observations of net soil exchange of CO2 in a dryland show experimental warming increases carbon losses in biocrust soils
0
2015
Photosynthesis and water relations of lichen soil crusts:field measurements in the coastal fog zone of the Namib Desert
2
1994
... 生物结皮属于典型的变水植物,在水分低于一定的阈值时处于代谢不活跃的休眠状态,而在降水后迅速恢复生理活性[35-37].我们发现,降水量极显著(P<0.001)影响两种生物结皮的GPP、ER和NEE,而增温和降水量之间无交互作用(表1).无论降水强度如何,生物结皮-土壤系统GPP和ER均在模拟降水后达到最大值,表现为光合固碳和呼吸碳排放的脉冲现象(图2),这主要源于恢复活性的隐花植物的光合固碳和自养呼吸以及异养呼吸的迅速增加[35,37-39].两类生物结皮的累积GPP和ER随着降水量的增加而线性增加,表明降水量越大,生物结皮的光合和呼吸作用的持续时间就越长.尽管降水会引发生物结皮较强的光合和呼吸作用,但在炎热的夏季,土壤水分会快速散失,水分可利用性的下降也致使GPP和ER迅速下降(图2),这些结果表明水分有效性对荒漠生物结皮的碳固定和排放具有决定性作用. ...
... ,37-39].两类生物结皮的累积GPP和ER随着降水量的增加而线性增加,表明降水量越大,生物结皮的光合和呼吸作用的持续时间就越长.尽管降水会引发生物结皮较强的光合和呼吸作用,但在炎热的夏季,土壤水分会快速散失,水分可利用性的下降也致使GPP和ER迅速下降(图2),这些结果表明水分有效性对荒漠生物结皮的碳固定和排放具有决定性作用. ...
Pathways regulating decreased soil respiration with warming in a biocrust-dominated dryland
0
2018
Soil respiration at five sites along the Kalahari Transect:effects of temperature,precipitation pulses and biological soil crust cover
1
2011
... 生物结皮属于典型的变水植物,在水分低于一定的阈值时处于代谢不活跃的休眠状态,而在降水后迅速恢复生理活性[35-37].我们发现,降水量极显著(P<0.001)影响两种生物结皮的GPP、ER和NEE,而增温和降水量之间无交互作用(表1).无论降水强度如何,生物结皮-土壤系统GPP和ER均在模拟降水后达到最大值,表现为光合固碳和呼吸碳排放的脉冲现象(图2),这主要源于恢复活性的隐花植物的光合固碳和自养呼吸以及异养呼吸的迅速增加[35,37-39].两类生物结皮的累积GPP和ER随着降水量的增加而线性增加,表明降水量越大,生物结皮的光合和呼吸作用的持续时间就越长.尽管降水会引发生物结皮较强的光合和呼吸作用,但在炎热的夏季,土壤水分会快速散失,水分可利用性的下降也致使GPP和ER迅速下降(图2),这些结果表明水分有效性对荒漠生物结皮的碳固定和排放具有决定性作用. ...
Temperature sensitivity of soil carbon decomposition and feedbacks to climate change
1
2006
... 温度敏感性能指示生物结皮碳交换关键过程响应全球气候变化的敏感程度,目前已被广泛应用于预测各类生态系统的碳收支研究中[40-41].我们的试验结果发现,不增温条件下藻结皮ER的温度敏感性(Q10=0.21)明显低于藓结皮(Q10=0.37),且增温降低了藓结皮ER和NEE的温度敏感性,这一结果支持了已有的研究发现[42-43],表明藓结皮的光合固碳和呼吸碳排放对温度的适应性[44].一方面,增温引起的水分有效性降低可能会通过抑制土壤微生物活性而间接降低自养和异养呼吸,导致ER对增温的响应逐渐减弱.另一方面,除温度和水分外,呼吸作用还受土壤微生物活性、酶活性、呼吸底物等诸多因子的影响.我们推测,土壤中呼吸底物的消耗极有可能是藓结皮ER温度敏感性下降的主要原因.值得注意的是,由于两类生物结皮的GPP和增温条件下藻结皮的ER与温度之间不存在指数关系,难以判定增温对其温度敏感性的直接影响,这可能与试验时间相对短暂有关.因此,有关这方面的研究还需要更多长期试验数据来进一步验证. ...
Increased precipitation offsets the negative effect of warming on plant biomass and ecosystem respiration in a Tibetan alpine steppe
1
2019
... 温度敏感性能指示生物结皮碳交换关键过程响应全球气候变化的敏感程度,目前已被广泛应用于预测各类生态系统的碳收支研究中[40-41].我们的试验结果发现,不增温条件下藻结皮ER的温度敏感性(Q10=0.21)明显低于藓结皮(Q10=0.37),且增温降低了藓结皮ER和NEE的温度敏感性,这一结果支持了已有的研究发现[42-43],表明藓结皮的光合固碳和呼吸碳排放对温度的适应性[44].一方面,增温引起的水分有效性降低可能会通过抑制土壤微生物活性而间接降低自养和异养呼吸,导致ER对增温的响应逐渐减弱.另一方面,除温度和水分外,呼吸作用还受土壤微生物活性、酶活性、呼吸底物等诸多因子的影响.我们推测,土壤中呼吸底物的消耗极有可能是藓结皮ER温度敏感性下降的主要原因.值得注意的是,由于两类生物结皮的GPP和增温条件下藻结皮的ER与温度之间不存在指数关系,难以判定增温对其温度敏感性的直接影响,这可能与试验时间相对短暂有关.因此,有关这方面的研究还需要更多长期试验数据来进一步验证. ...
不同土地利用方式下土壤呼吸及其温度敏感性
1
2007
... 温度敏感性能指示生物结皮碳交换关键过程响应全球气候变化的敏感程度,目前已被广泛应用于预测各类生态系统的碳收支研究中[40-41].我们的试验结果发现,不增温条件下藻结皮ER的温度敏感性(Q10=0.21)明显低于藓结皮(Q10=0.37),且增温降低了藓结皮ER和NEE的温度敏感性,这一结果支持了已有的研究发现[42-43],表明藓结皮的光合固碳和呼吸碳排放对温度的适应性[44].一方面,增温引起的水分有效性降低可能会通过抑制土壤微生物活性而间接降低自养和异养呼吸,导致ER对增温的响应逐渐减弱.另一方面,除温度和水分外,呼吸作用还受土壤微生物活性、酶活性、呼吸底物等诸多因子的影响.我们推测,土壤中呼吸底物的消耗极有可能是藓结皮ER温度敏感性下降的主要原因.值得注意的是,由于两类生物结皮的GPP和增温条件下藻结皮的ER与温度之间不存在指数关系,难以判定增温对其温度敏感性的直接影响,这可能与试验时间相对短暂有关.因此,有关这方面的研究还需要更多长期试验数据来进一步验证. ...
土壤呼吸温度敏感性的影响因素和不确定性
1
2011
... 温度敏感性能指示生物结皮碳交换关键过程响应全球气候变化的敏感程度,目前已被广泛应用于预测各类生态系统的碳收支研究中[40-41].我们的试验结果发现,不增温条件下藻结皮ER的温度敏感性(Q10=0.21)明显低于藓结皮(Q10=0.37),且增温降低了藓结皮ER和NEE的温度敏感性,这一结果支持了已有的研究发现[42-43],表明藓结皮的光合固碳和呼吸碳排放对温度的适应性[44].一方面,增温引起的水分有效性降低可能会通过抑制土壤微生物活性而间接降低自养和异养呼吸,导致ER对增温的响应逐渐减弱.另一方面,除温度和水分外,呼吸作用还受土壤微生物活性、酶活性、呼吸底物等诸多因子的影响.我们推测,土壤中呼吸底物的消耗极有可能是藓结皮ER温度敏感性下降的主要原因.值得注意的是,由于两类生物结皮的GPP和增温条件下藻结皮的ER与温度之间不存在指数关系,难以判定增温对其温度敏感性的直接影响,这可能与试验时间相对短暂有关.因此,有关这方面的研究还需要更多长期试验数据来进一步验证. ...
Acclimatization of soil respiration to warming in a tall grass prairie
1
2001
... 温度敏感性能指示生物结皮碳交换关键过程响应全球气候变化的敏感程度,目前已被广泛应用于预测各类生态系统的碳收支研究中[40-41].我们的试验结果发现,不增温条件下藻结皮ER的温度敏感性(Q10=0.21)明显低于藓结皮(Q10=0.37),且增温降低了藓结皮ER和NEE的温度敏感性,这一结果支持了已有的研究发现[42-43],表明藓结皮的光合固碳和呼吸碳排放对温度的适应性[44].一方面,增温引起的水分有效性降低可能会通过抑制土壤微生物活性而间接降低自养和异养呼吸,导致ER对增温的响应逐渐减弱.另一方面,除温度和水分外,呼吸作用还受土壤微生物活性、酶活性、呼吸底物等诸多因子的影响.我们推测,土壤中呼吸底物的消耗极有可能是藓结皮ER温度敏感性下降的主要原因.值得注意的是,由于两类生物结皮的GPP和增温条件下藻结皮的ER与温度之间不存在指数关系,难以判定增温对其温度敏感性的直接影响,这可能与试验时间相对短暂有关.因此,有关这方面的研究还需要更多长期试验数据来进一步验证. ...







 甘公网安备 62010202000688号
甘公网安备 62010202000688号
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}