中国草地生态系统固碳现状、速率和潜力研究
1
2018
... 草地是全球陆地生态系统的重要组成部分,占中国国土总面积的41.7%,是重要的畜牧产品生产基地,在调节温室气体循环利用、维持生态系统碳收支平衡等方面发挥着重要的作用[1-2].气候变化引起的全球降水格局变化存在空间和时空差异,因此有必要分地区探讨降水格局对草地生态系统的影响[3].降水格局变化是全球气候变化的重要内容,包括降水量的变化、降水季节分布的变化以及降水间隔的变化.降水波动,指年度和季节降水量、强度和频率在一定时期内增加或减少,降水波动对草地植物群落地上生物量的影响程度因降水强度和持续时间而产生差异[4].土壤水分胁迫可导致植物生长受到抑制造成植物死亡,植被组成和多样性发生改变的同时地上生物量降低[5-6].草地植物群落地上生物量可以衡量植被动态和生态系统功能的变化[7],是表征草地生态系统恢复进程最直观的指标[8],因此在气候变化背景下,降水格局变化会改变物种间关系,影响草地植物群落组成、结构和地上生物量的同时最终改变生态系统结构与功能[3,9]. ...
草地和荒漠生态系统服务功能的形成与调控机制
1
2014
... 草地是全球陆地生态系统的重要组成部分,占中国国土总面积的41.7%,是重要的畜牧产品生产基地,在调节温室气体循环利用、维持生态系统碳收支平衡等方面发挥着重要的作用[1-2].气候变化引起的全球降水格局变化存在空间和时空差异,因此有必要分地区探讨降水格局对草地生态系统的影响[3].降水格局变化是全球气候变化的重要内容,包括降水量的变化、降水季节分布的变化以及降水间隔的变化.降水波动,指年度和季节降水量、强度和频率在一定时期内增加或减少,降水波动对草地植物群落地上生物量的影响程度因降水强度和持续时间而产生差异[4].土壤水分胁迫可导致植物生长受到抑制造成植物死亡,植被组成和多样性发生改变的同时地上生物量降低[5-6].草地植物群落地上生物量可以衡量植被动态和生态系统功能的变化[7],是表征草地生态系统恢复进程最直观的指标[8],因此在气候变化背景下,降水格局变化会改变物种间关系,影响草地植物群落组成、结构和地上生物量的同时最终改变生态系统结构与功能[3,9]. ...
Toward a better integration of biological data from precipitation manipulation experiments into Earth system models
2
2014
... 草地是全球陆地生态系统的重要组成部分,占中国国土总面积的41.7%,是重要的畜牧产品生产基地,在调节温室气体循环利用、维持生态系统碳收支平衡等方面发挥着重要的作用[1-2].气候变化引起的全球降水格局变化存在空间和时空差异,因此有必要分地区探讨降水格局对草地生态系统的影响[3].降水格局变化是全球气候变化的重要内容,包括降水量的变化、降水季节分布的变化以及降水间隔的变化.降水波动,指年度和季节降水量、强度和频率在一定时期内增加或减少,降水波动对草地植物群落地上生物量的影响程度因降水强度和持续时间而产生差异[4].土壤水分胁迫可导致植物生长受到抑制造成植物死亡,植被组成和多样性发生改变的同时地上生物量降低[5-6].草地植物群落地上生物量可以衡量植被动态和生态系统功能的变化[7],是表征草地生态系统恢复进程最直观的指标[8],因此在气候变化背景下,降水格局变化会改变物种间关系,影响草地植物群落组成、结构和地上生物量的同时最终改变生态系统结构与功能[3,9]. ...
... [3,9]. ...
Assessing the response of terrestrial ecosystems to potential changes in precipitation
1
2003
... 草地是全球陆地生态系统的重要组成部分,占中国国土总面积的41.7%,是重要的畜牧产品生产基地,在调节温室气体循环利用、维持生态系统碳收支平衡等方面发挥着重要的作用[1-2].气候变化引起的全球降水格局变化存在空间和时空差异,因此有必要分地区探讨降水格局对草地生态系统的影响[3].降水格局变化是全球气候变化的重要内容,包括降水量的变化、降水季节分布的变化以及降水间隔的变化.降水波动,指年度和季节降水量、强度和频率在一定时期内增加或减少,降水波动对草地植物群落地上生物量的影响程度因降水强度和持续时间而产生差异[4].土壤水分胁迫可导致植物生长受到抑制造成植物死亡,植被组成和多样性发生改变的同时地上生物量降低[5-6].草地植物群落地上生物量可以衡量植被动态和生态系统功能的变化[7],是表征草地生态系统恢复进程最直观的指标[8],因此在气候变化背景下,降水格局变化会改变物种间关系,影响草地植物群落组成、结构和地上生物量的同时最终改变生态系统结构与功能[3,9]. ...
Experimental droughts with rainout shelters:a methodological review
1
2018
... 草地是全球陆地生态系统的重要组成部分,占中国国土总面积的41.7%,是重要的畜牧产品生产基地,在调节温室气体循环利用、维持生态系统碳收支平衡等方面发挥着重要的作用[1-2].气候变化引起的全球降水格局变化存在空间和时空差异,因此有必要分地区探讨降水格局对草地生态系统的影响[3].降水格局变化是全球气候变化的重要内容,包括降水量的变化、降水季节分布的变化以及降水间隔的变化.降水波动,指年度和季节降水量、强度和频率在一定时期内增加或减少,降水波动对草地植物群落地上生物量的影响程度因降水强度和持续时间而产生差异[4].土壤水分胁迫可导致植物生长受到抑制造成植物死亡,植被组成和多样性发生改变的同时地上生物量降低[5-6].草地植物群落地上生物量可以衡量植被动态和生态系统功能的变化[7],是表征草地生态系统恢复进程最直观的指标[8],因此在气候变化背景下,降水格局变化会改变物种间关系,影响草地植物群落组成、结构和地上生物量的同时最终改变生态系统结构与功能[3,9]. ...
The response of tropical rainforests to drought:lessons from recent research and future prospects
1
2016
... 草地是全球陆地生态系统的重要组成部分,占中国国土总面积的41.7%,是重要的畜牧产品生产基地,在调节温室气体循环利用、维持生态系统碳收支平衡等方面发挥着重要的作用[1-2].气候变化引起的全球降水格局变化存在空间和时空差异,因此有必要分地区探讨降水格局对草地生态系统的影响[3].降水格局变化是全球气候变化的重要内容,包括降水量的变化、降水季节分布的变化以及降水间隔的变化.降水波动,指年度和季节降水量、强度和频率在一定时期内增加或减少,降水波动对草地植物群落地上生物量的影响程度因降水强度和持续时间而产生差异[4].土壤水分胁迫可导致植物生长受到抑制造成植物死亡,植被组成和多样性发生改变的同时地上生物量降低[5-6].草地植物群落地上生物量可以衡量植被动态和生态系统功能的变化[7],是表征草地生态系统恢复进程最直观的指标[8],因此在气候变化背景下,降水格局变化会改变物种间关系,影响草地植物群落组成、结构和地上生物量的同时最终改变生态系统结构与功能[3,9]. ...
半干旱沙地生境变化对植物地上生物量及其碳、氮储量的影响
1
2014
... 草地是全球陆地生态系统的重要组成部分,占中国国土总面积的41.7%,是重要的畜牧产品生产基地,在调节温室气体循环利用、维持生态系统碳收支平衡等方面发挥着重要的作用[1-2].气候变化引起的全球降水格局变化存在空间和时空差异,因此有必要分地区探讨降水格局对草地生态系统的影响[3].降水格局变化是全球气候变化的重要内容,包括降水量的变化、降水季节分布的变化以及降水间隔的变化.降水波动,指年度和季节降水量、强度和频率在一定时期内增加或减少,降水波动对草地植物群落地上生物量的影响程度因降水强度和持续时间而产生差异[4].土壤水分胁迫可导致植物生长受到抑制造成植物死亡,植被组成和多样性发生改变的同时地上生物量降低[5-6].草地植物群落地上生物量可以衡量植被动态和生态系统功能的变化[7],是表征草地生态系统恢复进程最直观的指标[8],因此在气候变化背景下,降水格局变化会改变物种间关系,影响草地植物群落组成、结构和地上生物量的同时最终改变生态系统结构与功能[3,9]. ...
短期降雨改变对荒漠草原植物群落特征的影响
1
2018
... 草地是全球陆地生态系统的重要组成部分,占中国国土总面积的41.7%,是重要的畜牧产品生产基地,在调节温室气体循环利用、维持生态系统碳收支平衡等方面发挥着重要的作用[1-2].气候变化引起的全球降水格局变化存在空间和时空差异,因此有必要分地区探讨降水格局对草地生态系统的影响[3].降水格局变化是全球气候变化的重要内容,包括降水量的变化、降水季节分布的变化以及降水间隔的变化.降水波动,指年度和季节降水量、强度和频率在一定时期内增加或减少,降水波动对草地植物群落地上生物量的影响程度因降水强度和持续时间而产生差异[4].土壤水分胁迫可导致植物生长受到抑制造成植物死亡,植被组成和多样性发生改变的同时地上生物量降低[5-6].草地植物群落地上生物量可以衡量植被动态和生态系统功能的变化[7],是表征草地生态系统恢复进程最直观的指标[8],因此在气候变化背景下,降水格局变化会改变物种间关系,影响草地植物群落组成、结构和地上生物量的同时最终改变生态系统结构与功能[3,9]. ...
荒漠草原沙生针茅(Stipa glareosa)群落物种多样性和地上生物量对降雨量的响应
2
2019
... 草地是全球陆地生态系统的重要组成部分,占中国国土总面积的41.7%,是重要的畜牧产品生产基地,在调节温室气体循环利用、维持生态系统碳收支平衡等方面发挥着重要的作用[1-2].气候变化引起的全球降水格局变化存在空间和时空差异,因此有必要分地区探讨降水格局对草地生态系统的影响[3].降水格局变化是全球气候变化的重要内容,包括降水量的变化、降水季节分布的变化以及降水间隔的变化.降水波动,指年度和季节降水量、强度和频率在一定时期内增加或减少,降水波动对草地植物群落地上生物量的影响程度因降水强度和持续时间而产生差异[4].土壤水分胁迫可导致植物生长受到抑制造成植物死亡,植被组成和多样性发生改变的同时地上生物量降低[5-6].草地植物群落地上生物量可以衡量植被动态和生态系统功能的变化[7],是表征草地生态系统恢复进程最直观的指标[8],因此在气候变化背景下,降水格局变化会改变物种间关系,影响草地植物群落组成、结构和地上生物量的同时最终改变生态系统结构与功能[3,9]. ...
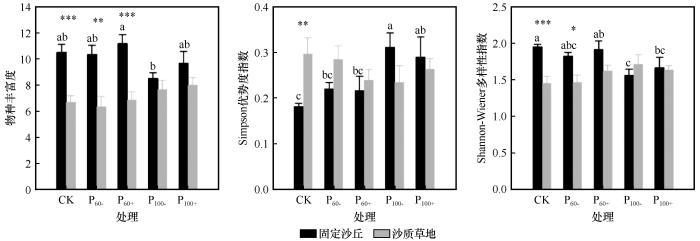
... 在固定沙丘中,干旱降低Shannon-Wiener多样性指数的同时提高了Simpson优势度指数和地上生物量,这可能是因为5月和6月的极端干旱加剧了种间竞争[46],限制了竞争力较低草种的发芽,缺乏耐旱机制的植物将会减少,在生长发育早期就个体消亡[28,47],而具有耐旱和避旱机制的植物不仅不会受干旱条件的影响,甚至会生长得更好,因此地上生物量升高.这与以往在干旱、半干旱草原区开展的研究结果一致,即极端干旱或降水大幅波动会对草地植物群落结构和生物多样性产生影响,甚至会使整个生态系统的结构和功能发生改变,主要体现在植物群落优势种的更替上[21,48].在沙质草地中,生长季降水格局变化对物种丰富度、Simpson优势度指数和Shannon-Wiener多样性指数的影响不显著,主要原因可能是沙质草地中多年生白草和糙隐子草的优势度较高,对生长季降水格局变化有一定的缓冲能力[49],这也与多年生植物群落物种组成及结构的稳定性优于一二年生植物群落的研究结果相一致[50-51].干旱处理下,沙质草地的植被盖度和植物密度显著降低,这主要是因为长期干旱通过影响土壤水分进而抑制土壤潜在种子库的萌发和对植物群落生长产生滞后影响效应[9,52]. ...
干旱半干旱区退化草地土壤水分变化及其对降雨时间格局的响应
1
2020
... 由于特殊的气候条件、地理位置等因素,干旱、半干旱地区的草地生态系统对降水格局变化高度敏感[10].降水量对草地植物群落地上生物量的影响结果不一致:在内蒙古典型草原和荒漠草原,降水增加显著提高了植物群落地上生物量[11-12];而在科尔沁沙地和黄土高原草原荒漠带,降水增加显著降低了植物群落地上生物量[13],主要原因可能是降水量对植物群落地上生物量的影响存在阈值范围,当降水量超出植物生长所适宜的水分阈值时植物群落地上生物量反而会下降[14].1—7月降水量对内蒙古草地地上生物量的影响更为显著[15],夏季降水量对北方温带草原植被的生长影响最为显著[16];7—8月增加降水30%对北方温带草原的地上生物量有显著的提高作用[17];减少降水频次的同时增加降水强度更有利于土壤较高含水量的维持,有助于草地地上生物量的提高[18].非生长季的降水仅有部分存留在土壤中保证植物在生长季生长发育所需,即使在生长季,植物各个生长期对水分的需求也不尽相同[19].因此,对于干旱、半干旱区的草原,生长季内的平均单次降水量和降水次数在调控草地植被生长过程方面比年降水量发挥着更为重要的作用[20-21]. ...
降水变化对内蒙古典型草原地上生物量的影响
1
... 由于特殊的气候条件、地理位置等因素,干旱、半干旱地区的草地生态系统对降水格局变化高度敏感[10].降水量对草地植物群落地上生物量的影响结果不一致:在内蒙古典型草原和荒漠草原,降水增加显著提高了植物群落地上生物量[11-12];而在科尔沁沙地和黄土高原草原荒漠带,降水增加显著降低了植物群落地上生物量[13],主要原因可能是降水量对植物群落地上生物量的影响存在阈值范围,当降水量超出植物生长所适宜的水分阈值时植物群落地上生物量反而会下降[14].1—7月降水量对内蒙古草地地上生物量的影响更为显著[15],夏季降水量对北方温带草原植被的生长影响最为显著[16];7—8月增加降水30%对北方温带草原的地上生物量有显著的提高作用[17];减少降水频次的同时增加降水强度更有利于土壤较高含水量的维持,有助于草地地上生物量的提高[18].非生长季的降水仅有部分存留在土壤中保证植物在生长季生长发育所需,即使在生长季,植物各个生长期对水分的需求也不尽相同[19].因此,对于干旱、半干旱区的草原,生长季内的平均单次降水量和降水次数在调控草地植被生长过程方面比年降水量发挥着更为重要的作用[20-21]. ...
Direct and indirect effects of precipitation change and nutrients addition on desert grassland productivity in Inner Mongolia,northern China
1
2022
... 由于特殊的气候条件、地理位置等因素,干旱、半干旱地区的草地生态系统对降水格局变化高度敏感[10].降水量对草地植物群落地上生物量的影响结果不一致:在内蒙古典型草原和荒漠草原,降水增加显著提高了植物群落地上生物量[11-12];而在科尔沁沙地和黄土高原草原荒漠带,降水增加显著降低了植物群落地上生物量[13],主要原因可能是降水量对植物群落地上生物量的影响存在阈值范围,当降水量超出植物生长所适宜的水分阈值时植物群落地上生物量反而会下降[14].1—7月降水量对内蒙古草地地上生物量的影响更为显著[15],夏季降水量对北方温带草原植被的生长影响最为显著[16];7—8月增加降水30%对北方温带草原的地上生物量有显著的提高作用[17];减少降水频次的同时增加降水强度更有利于土壤较高含水量的维持,有助于草地地上生物量的提高[18].非生长季的降水仅有部分存留在土壤中保证植物在生长季生长发育所需,即使在生长季,植物各个生长期对水分的需求也不尽相同[19].因此,对于干旱、半干旱区的草原,生长季内的平均单次降水量和降水次数在调控草地植被生长过程方面比年降水量发挥着更为重要的作用[20-21]. ...
黄土高原 4 种植被带草本群落特征及其对降水变化的响应
1
2019
... 由于特殊的气候条件、地理位置等因素,干旱、半干旱地区的草地生态系统对降水格局变化高度敏感[10].降水量对草地植物群落地上生物量的影响结果不一致:在内蒙古典型草原和荒漠草原,降水增加显著提高了植物群落地上生物量[11-12];而在科尔沁沙地和黄土高原草原荒漠带,降水增加显著降低了植物群落地上生物量[13],主要原因可能是降水量对植物群落地上生物量的影响存在阈值范围,当降水量超出植物生长所适宜的水分阈值时植物群落地上生物量反而会下降[14].1—7月降水量对内蒙古草地地上生物量的影响更为显著[15],夏季降水量对北方温带草原植被的生长影响最为显著[16];7—8月增加降水30%对北方温带草原的地上生物量有显著的提高作用[17];减少降水频次的同时增加降水强度更有利于土壤较高含水量的维持,有助于草地地上生物量的提高[18].非生长季的降水仅有部分存留在土壤中保证植物在生长季生长发育所需,即使在生长季,植物各个生长期对水分的需求也不尽相同[19].因此,对于干旱、半干旱区的草原,生长季内的平均单次降水量和降水次数在调控草地植被生长过程方面比年降水量发挥着更为重要的作用[20-21]. ...
模拟短期降雨量变化对准噶尔荒漠植物群落的影响
1
2019
... 由于特殊的气候条件、地理位置等因素,干旱、半干旱地区的草地生态系统对降水格局变化高度敏感[10].降水量对草地植物群落地上生物量的影响结果不一致:在内蒙古典型草原和荒漠草原,降水增加显著提高了植物群落地上生物量[11-12];而在科尔沁沙地和黄土高原草原荒漠带,降水增加显著降低了植物群落地上生物量[13],主要原因可能是降水量对植物群落地上生物量的影响存在阈值范围,当降水量超出植物生长所适宜的水分阈值时植物群落地上生物量反而会下降[14].1—7月降水量对内蒙古草地地上生物量的影响更为显著[15],夏季降水量对北方温带草原植被的生长影响最为显著[16];7—8月增加降水30%对北方温带草原的地上生物量有显著的提高作用[17];减少降水频次的同时增加降水强度更有利于土壤较高含水量的维持,有助于草地地上生物量的提高[18].非生长季的降水仅有部分存留在土壤中保证植物在生长季生长发育所需,即使在生长季,植物各个生长期对水分的需求也不尽相同[19].因此,对于干旱、半干旱区的草原,生长季内的平均单次降水量和降水次数在调控草地植被生长过程方面比年降水量发挥着更为重要的作用[20-21]. ...
Ecosystem stability and compensatory effects in the Inner Mongolia grassland
1
2004
... 由于特殊的气候条件、地理位置等因素,干旱、半干旱地区的草地生态系统对降水格局变化高度敏感[10].降水量对草地植物群落地上生物量的影响结果不一致:在内蒙古典型草原和荒漠草原,降水增加显著提高了植物群落地上生物量[11-12];而在科尔沁沙地和黄土高原草原荒漠带,降水增加显著降低了植物群落地上生物量[13],主要原因可能是降水量对植物群落地上生物量的影响存在阈值范围,当降水量超出植物生长所适宜的水分阈值时植物群落地上生物量反而会下降[14].1—7月降水量对内蒙古草地地上生物量的影响更为显著[15],夏季降水量对北方温带草原植被的生长影响最为显著[16];7—8月增加降水30%对北方温带草原的地上生物量有显著的提高作用[17];减少降水频次的同时增加降水强度更有利于土壤较高含水量的维持,有助于草地地上生物量的提高[18].非生长季的降水仅有部分存留在土壤中保证植物在生长季生长发育所需,即使在生长季,植物各个生长期对水分的需求也不尽相同[19].因此,对于干旱、半干旱区的草原,生长季内的平均单次降水量和降水次数在调控草地植被生长过程方面比年降水量发挥着更为重要的作用[20-21]. ...
气候变化对中国北方温带草原植被的影响
1
2006
... 由于特殊的气候条件、地理位置等因素,干旱、半干旱地区的草地生态系统对降水格局变化高度敏感[10].降水量对草地植物群落地上生物量的影响结果不一致:在内蒙古典型草原和荒漠草原,降水增加显著提高了植物群落地上生物量[11-12];而在科尔沁沙地和黄土高原草原荒漠带,降水增加显著降低了植物群落地上生物量[13],主要原因可能是降水量对植物群落地上生物量的影响存在阈值范围,当降水量超出植物生长所适宜的水分阈值时植物群落地上生物量反而会下降[14].1—7月降水量对内蒙古草地地上生物量的影响更为显著[15],夏季降水量对北方温带草原植被的生长影响最为显著[16];7—8月增加降水30%对北方温带草原的地上生物量有显著的提高作用[17];减少降水频次的同时增加降水强度更有利于土壤较高含水量的维持,有助于草地地上生物量的提高[18].非生长季的降水仅有部分存留在土壤中保证植物在生长季生长发育所需,即使在生长季,植物各个生长期对水分的需求也不尽相同[19].因此,对于干旱、半干旱区的草原,生长季内的平均单次降水量和降水次数在调控草地植被生长过程方面比年降水量发挥着更为重要的作用[20-21]. ...
Water-mediated responses of ecosystem carbon fluxes to climatic change in a temperate steppe
1
2010
... 由于特殊的气候条件、地理位置等因素,干旱、半干旱地区的草地生态系统对降水格局变化高度敏感[10].降水量对草地植物群落地上生物量的影响结果不一致:在内蒙古典型草原和荒漠草原,降水增加显著提高了植物群落地上生物量[11-12];而在科尔沁沙地和黄土高原草原荒漠带,降水增加显著降低了植物群落地上生物量[13],主要原因可能是降水量对植物群落地上生物量的影响存在阈值范围,当降水量超出植物生长所适宜的水分阈值时植物群落地上生物量反而会下降[14].1—7月降水量对内蒙古草地地上生物量的影响更为显著[15],夏季降水量对北方温带草原植被的生长影响最为显著[16];7—8月增加降水30%对北方温带草原的地上生物量有显著的提高作用[17];减少降水频次的同时增加降水强度更有利于土壤较高含水量的维持,有助于草地地上生物量的提高[18].非生长季的降水仅有部分存留在土壤中保证植物在生长季生长发育所需,即使在生长季,植物各个生长期对水分的需求也不尽相同[19].因此,对于干旱、半干旱区的草原,生长季内的平均单次降水量和降水次数在调控草地植被生长过程方面比年降水量发挥着更为重要的作用[20-21]. ...
Variation among biomes in temporal dynamics of aboveground primary production
1
2001
... 由于特殊的气候条件、地理位置等因素,干旱、半干旱地区的草地生态系统对降水格局变化高度敏感[10].降水量对草地植物群落地上生物量的影响结果不一致:在内蒙古典型草原和荒漠草原,降水增加显著提高了植物群落地上生物量[11-12];而在科尔沁沙地和黄土高原草原荒漠带,降水增加显著降低了植物群落地上生物量[13],主要原因可能是降水量对植物群落地上生物量的影响存在阈值范围,当降水量超出植物生长所适宜的水分阈值时植物群落地上生物量反而会下降[14].1—7月降水量对内蒙古草地地上生物量的影响更为显著[15],夏季降水量对北方温带草原植被的生长影响最为显著[16];7—8月增加降水30%对北方温带草原的地上生物量有显著的提高作用[17];减少降水频次的同时增加降水强度更有利于土壤较高含水量的维持,有助于草地地上生物量的提高[18].非生长季的降水仅有部分存留在土壤中保证植物在生长季生长发育所需,即使在生长季,植物各个生长期对水分的需求也不尽相同[19].因此,对于干旱、半干旱区的草原,生长季内的平均单次降水量和降水次数在调控草地植被生长过程方面比年降水量发挥着更为重要的作用[20-21]. ...
中国北方草地生产力研究进展
1
2022
... 由于特殊的气候条件、地理位置等因素,干旱、半干旱地区的草地生态系统对降水格局变化高度敏感[10].降水量对草地植物群落地上生物量的影响结果不一致:在内蒙古典型草原和荒漠草原,降水增加显著提高了植物群落地上生物量[11-12];而在科尔沁沙地和黄土高原草原荒漠带,降水增加显著降低了植物群落地上生物量[13],主要原因可能是降水量对植物群落地上生物量的影响存在阈值范围,当降水量超出植物生长所适宜的水分阈值时植物群落地上生物量反而会下降[14].1—7月降水量对内蒙古草地地上生物量的影响更为显著[15],夏季降水量对北方温带草原植被的生长影响最为显著[16];7—8月增加降水30%对北方温带草原的地上生物量有显著的提高作用[17];减少降水频次的同时增加降水强度更有利于土壤较高含水量的维持,有助于草地地上生物量的提高[18].非生长季的降水仅有部分存留在土壤中保证植物在生长季生长发育所需,即使在生长季,植物各个生长期对水分的需求也不尽相同[19].因此,对于干旱、半干旱区的草原,生长季内的平均单次降水量和降水次数在调控草地植被生长过程方面比年降水量发挥着更为重要的作用[20-21]. ...
Seasonal not annual rainfall determines grassland biomass response to carbon dioxide
1
2014
... 由于特殊的气候条件、地理位置等因素,干旱、半干旱地区的草地生态系统对降水格局变化高度敏感[10].降水量对草地植物群落地上生物量的影响结果不一致:在内蒙古典型草原和荒漠草原,降水增加显著提高了植物群落地上生物量[11-12];而在科尔沁沙地和黄土高原草原荒漠带,降水增加显著降低了植物群落地上生物量[13],主要原因可能是降水量对植物群落地上生物量的影响存在阈值范围,当降水量超出植物生长所适宜的水分阈值时植物群落地上生物量反而会下降[14].1—7月降水量对内蒙古草地地上生物量的影响更为显著[15],夏季降水量对北方温带草原植被的生长影响最为显著[16];7—8月增加降水30%对北方温带草原的地上生物量有显著的提高作用[17];减少降水频次的同时增加降水强度更有利于土壤较高含水量的维持,有助于草地地上生物量的提高[18].非生长季的降水仅有部分存留在土壤中保证植物在生长季生长发育所需,即使在生长季,植物各个生长期对水分的需求也不尽相同[19].因此,对于干旱、半干旱区的草原,生长季内的平均单次降水量和降水次数在调控草地植被生长过程方面比年降水量发挥着更为重要的作用[20-21]. ...
科尔沁固定沙地植被特征对降雨变化的响应
3
2014
... 由于特殊的气候条件、地理位置等因素,干旱、半干旱地区的草地生态系统对降水格局变化高度敏感[10].降水量对草地植物群落地上生物量的影响结果不一致:在内蒙古典型草原和荒漠草原,降水增加显著提高了植物群落地上生物量[11-12];而在科尔沁沙地和黄土高原草原荒漠带,降水增加显著降低了植物群落地上生物量[13],主要原因可能是降水量对植物群落地上生物量的影响存在阈值范围,当降水量超出植物生长所适宜的水分阈值时植物群落地上生物量反而会下降[14].1—7月降水量对内蒙古草地地上生物量的影响更为显著[15],夏季降水量对北方温带草原植被的生长影响最为显著[16];7—8月增加降水30%对北方温带草原的地上生物量有显著的提高作用[17];减少降水频次的同时增加降水强度更有利于土壤较高含水量的维持,有助于草地地上生物量的提高[18].非生长季的降水仅有部分存留在土壤中保证植物在生长季生长发育所需,即使在生长季,植物各个生长期对水分的需求也不尽相同[19].因此,对于干旱、半干旱区的草原,生长季内的平均单次降水量和降水次数在调控草地植被生长过程方面比年降水量发挥着更为重要的作用[20-21]. ...
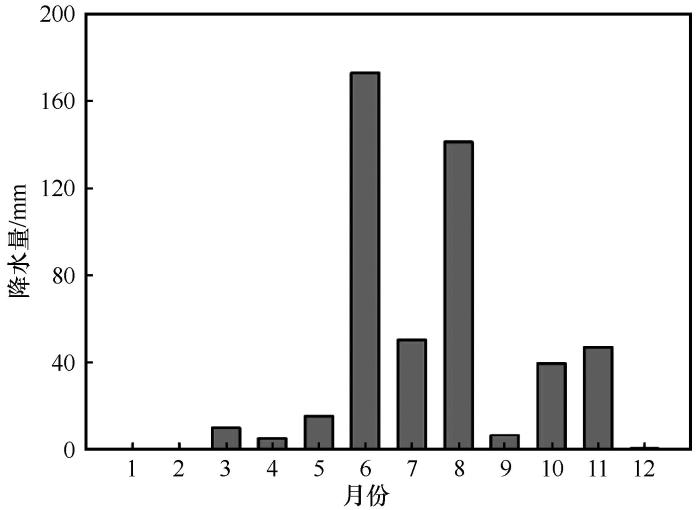
... 该地区2001—2016年降水量波动变化且略有升高,表现为最大降水量比平均降水量大64.8%,而最小降水量比平均降水量小56.2%.极端干旱与极端降水事件发生的频率增大,尤其是生长季前期(5—6月)降水间隔较长、有效降水较少(单次降水量<5 mm),因此,我们设置了5个水平的降水处理:自然降水(CK)、5—9月减水60%(P60-)、5—9月增水60%(P60+)、5—6月减水100%(P100-)和5—6月增水100%(P100+).随机选择固定沙丘和沙质草地两种生境类型作为研究样地,每种生境各建立12个大小为8 m×8 m的样地,其中每个生境的6个样地内设置两套增减水装置(图1A,P60-和P60+;图1B,P100-和P100+;间距大于1 m),另6个作为对照处理.每套增减水装置主要由不锈钢支架、遮雨板、集雨槽、增雨管组成:不锈钢支架规格为长260 cm、宽250 cm,远离集雨槽的一端高为140 cm,而靠近集雨槽的一端高为100 cm,增减水装置的原理、所用材料及规格均与张腊梅等[21]的大体一致.将厚5 mm、长1 050 cm、宽105 cm的铁皮及塑料布埋入减水区及增水区的四周,铁皮和塑料布漏出地面约5 cm,从而排除地表径流、地下水分迁移等对实验造成的影响. ...
... 在固定沙丘中,干旱降低Shannon-Wiener多样性指数的同时提高了Simpson优势度指数和地上生物量,这可能是因为5月和6月的极端干旱加剧了种间竞争[46],限制了竞争力较低草种的发芽,缺乏耐旱机制的植物将会减少,在生长发育早期就个体消亡[28,47],而具有耐旱和避旱机制的植物不仅不会受干旱条件的影响,甚至会生长得更好,因此地上生物量升高.这与以往在干旱、半干旱草原区开展的研究结果一致,即极端干旱或降水大幅波动会对草地植物群落结构和生物多样性产生影响,甚至会使整个生态系统的结构和功能发生改变,主要体现在植物群落优势种的更替上[21,48].在沙质草地中,生长季降水格局变化对物种丰富度、Simpson优势度指数和Shannon-Wiener多样性指数的影响不显著,主要原因可能是沙质草地中多年生白草和糙隐子草的优势度较高,对生长季降水格局变化有一定的缓冲能力[49],这也与多年生植物群落物种组成及结构的稳定性优于一二年生植物群落的研究结果相一致[50-51].干旱处理下,沙质草地的植被盖度和植物密度显著降低,这主要是因为长期干旱通过影响土壤水分进而抑制土壤潜在种子库的萌发和对植物群落生长产生滞后影响效应[9,52]. ...
盐渍化草地根际土壤理化性质对降水改变和氮添加的响应
1
2022
... 土壤理化性质均会对草地植物的生长造成影响,生长季降水格局变化可通过影响土壤水分、植物根际效应和土壤微生物特性来影响土壤微团聚体大小、吸附性能、亲水性以及pH值,最终影响草地地上生物量[22-23].植物通过性状权衡将获取的资源转变为地上生物量,进而影响生态系统功能[24].使用单一性状不能提供足够的信息来预测地上生物量,因为每个性状都代表一个物种有限的信息集[25],例如株高、比叶面积、叶干物质含量和叶片碳氮含量是影响植物生长速率和植物组织质量结构支撑的主要性状,与地上生物量相关联[26].由此,生长季降水格局变化对生态系统功能的影响实际上是通过功能性状调节地上生物量所实现的.因此,在生长季模拟提前和推迟降水峰值和改变降水分配格局情境下,通过模型模拟方式研究草地植被特征、功能多样性、土壤理化性质和地上生物量关系的影响及调控机制,不仅有助于预测未来干旱、半干旱草地生态系统功能的变化趋势,而且对科学管理草地、遏制草地退化和促进草地生态系统可持续发展具有重要的科学指导意义. ...
长期氮水添加对温带草原土壤化学性质和微生物学特性的短期遗留效应
2
2023
... 土壤理化性质均会对草地植物的生长造成影响,生长季降水格局变化可通过影响土壤水分、植物根际效应和土壤微生物特性来影响土壤微团聚体大小、吸附性能、亲水性以及pH值,最终影响草地地上生物量[22-23].植物通过性状权衡将获取的资源转变为地上生物量,进而影响生态系统功能[24].使用单一性状不能提供足够的信息来预测地上生物量,因为每个性状都代表一个物种有限的信息集[25],例如株高、比叶面积、叶干物质含量和叶片碳氮含量是影响植物生长速率和植物组织质量结构支撑的主要性状,与地上生物量相关联[26].由此,生长季降水格局变化对生态系统功能的影响实际上是通过功能性状调节地上生物量所实现的.因此,在生长季模拟提前和推迟降水峰值和改变降水分配格局情境下,通过模型模拟方式研究草地植被特征、功能多样性、土壤理化性质和地上生物量关系的影响及调控机制,不仅有助于预测未来干旱、半干旱草地生态系统功能的变化趋势,而且对科学管理草地、遏制草地退化和促进草地生态系统可持续发展具有重要的科学指导意义. ...
... 在植被调查的同时,用直径为3 cm的土钻在每个调查样方内,以“五点取样法”钻取0~10 cm的土壤样品,过2 mm标准筛后取部分新鲜土壤样品装入小铝盒,用于测定土壤质量含水量,剩余土壤样品进行自然风干处理.部分风干土样研磨后用于测定土壤全碳、全氮含量,另一部分风干土样用于测定土壤pH、电导率、机械组成,用容重钻(环刀体积100 cm
3)钻取0~10 cm土壤样品,用于测定土壤容重
[34-35].采用元素分析仪(Costech ECS4010)测定土壤全碳、全氮含量,采用105 ℃烘干恒重法测定土壤质量含水量,pH计测定土壤酸碱度(水土比2.5∶1),电导率仪(水土比5∶1)测定土壤电导率,湿筛法测定土壤机械组成(粗砂>0.5 mm,中砂0.25~0.5 mm,细砂0.10~0.25 mm,极细砂0.05~0.10 mm,黏粉粒< 0.05 mm),环刀内的土壤样品105 ℃烘干至恒重即为土壤容重.
<strong>1.4</strong> 数据分析<strong>1.4.1</strong> 物种多样性<sup>[<xref ref-type="bibr" rid="R23">23</xref>,<xref ref-type="bibr" rid="R36">36</xref>]</sup>物种重要值=(相对盖度+相对高度+相对密度+相对地上生物量)/4 ...
Predicting changes in community composition and ecosystem functioning from plant traits:revisiting the Holy Grail
1
2002
... 土壤理化性质均会对草地植物的生长造成影响,生长季降水格局变化可通过影响土壤水分、植物根际效应和土壤微生物特性来影响土壤微团聚体大小、吸附性能、亲水性以及pH值,最终影响草地地上生物量[22-23].植物通过性状权衡将获取的资源转变为地上生物量,进而影响生态系统功能[24].使用单一性状不能提供足够的信息来预测地上生物量,因为每个性状都代表一个物种有限的信息集[25],例如株高、比叶面积、叶干物质含量和叶片碳氮含量是影响植物生长速率和植物组织质量结构支撑的主要性状,与地上生物量相关联[26].由此,生长季降水格局变化对生态系统功能的影响实际上是通过功能性状调节地上生物量所实现的.因此,在生长季模拟提前和推迟降水峰值和改变降水分配格局情境下,通过模型模拟方式研究草地植被特征、功能多样性、土壤理化性质和地上生物量关系的影响及调控机制,不仅有助于预测未来干旱、半干旱草地生态系统功能的变化趋势,而且对科学管理草地、遏制草地退化和促进草地生态系统可持续发展具有重要的科学指导意义. ...
Different clades and traits yield similar grassland functional responses
1
2017
... 土壤理化性质均会对草地植物的生长造成影响,生长季降水格局变化可通过影响土壤水分、植物根际效应和土壤微生物特性来影响土壤微团聚体大小、吸附性能、亲水性以及pH值,最终影响草地地上生物量[22-23].植物通过性状权衡将获取的资源转变为地上生物量,进而影响生态系统功能[24].使用单一性状不能提供足够的信息来预测地上生物量,因为每个性状都代表一个物种有限的信息集[25],例如株高、比叶面积、叶干物质含量和叶片碳氮含量是影响植物生长速率和植物组织质量结构支撑的主要性状,与地上生物量相关联[26].由此,生长季降水格局变化对生态系统功能的影响实际上是通过功能性状调节地上生物量所实现的.因此,在生长季模拟提前和推迟降水峰值和改变降水分配格局情境下,通过模型模拟方式研究草地植被特征、功能多样性、土壤理化性质和地上生物量关系的影响及调控机制,不仅有助于预测未来干旱、半干旱草地生态系统功能的变化趋势,而且对科学管理草地、遏制草地退化和促进草地生态系统可持续发展具有重要的科学指导意义. ...
Functional diversity supports the physiological tolerance hypothesis for plant species richness along climatic gradients
2
2014
... 土壤理化性质均会对草地植物的生长造成影响,生长季降水格局变化可通过影响土壤水分、植物根际效应和土壤微生物特性来影响土壤微团聚体大小、吸附性能、亲水性以及pH值,最终影响草地地上生物量[22-23].植物通过性状权衡将获取的资源转变为地上生物量,进而影响生态系统功能[24].使用单一性状不能提供足够的信息来预测地上生物量,因为每个性状都代表一个物种有限的信息集[25],例如株高、比叶面积、叶干物质含量和叶片碳氮含量是影响植物生长速率和植物组织质量结构支撑的主要性状,与地上生物量相关联[26].由此,生长季降水格局变化对生态系统功能的影响实际上是通过功能性状调节地上生物量所实现的.因此,在生长季模拟提前和推迟降水峰值和改变降水分配格局情境下,通过模型模拟方式研究草地植被特征、功能多样性、土壤理化性质和地上生物量关系的影响及调控机制,不仅有助于预测未来干旱、半干旱草地生态系统功能的变化趋势,而且对科学管理草地、遏制草地退化和促进草地生态系统可持续发展具有重要的科学指导意义. ...
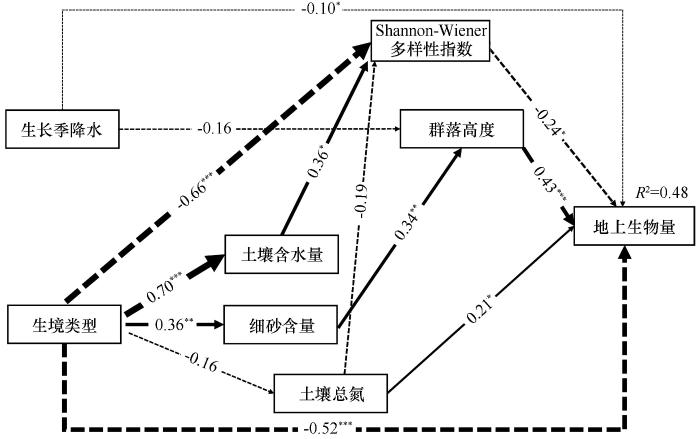
... 本研究中描述生境类型、生长季降水格局变化、植被特征和土壤理化性质对地上生物量影响效应路径的结构方程模型表明,植物群落高度对地上生物量具有显著的正向影响,这与以往的研究结果一致,即株高是影响植物生长速率和植物组织质量结构支撑的主要性状,与地上生物量的正相关联作用最为明显[26].本研究中生长季降水格局变化对地上生物量产生了微弱的负向影响,主要原因是干旱处理下根冠较为发达、单株生物量较高的蒺藜和尖头叶藜等植物的优势度大幅度升高,而增水处理增加了糙隐子草和狗尾草等单株生物量较小植物的优势度,因此干旱或增水处理下地上生物量的提高或降低主要是由于物种更替所导致. ...
生态工程对科尔沁沙地主要生态服务功能的影响
1
2021
... 科尔沁沙地原是科尔沁草原的主体部分,原生景观是疏林草原,较早的农业开发和近百年农牧交错带在这里大规模北侵,造成沙漠化急剧发展,成为中国现代沙漠化严重地区,也是中国沙漠化演变与防治研究的典型区域[27].近年来随着围栏封育措施的实施,该地区的土地沙漠化趋势得到遏制并发生逆转,植被和土壤都呈现一定程度的恢复趋势[28-29],相继出现了流动沙丘、半固定沙丘、固定沙丘和沙质草地交错分布的地貌类型.固定沙丘和沙质草地是反映科尔沁沙地沙漠化进程的关键景观特征,其中固定沙丘植物群落稳定性较高,而沙质草地生态环境脆弱,对人为干扰及气候变化反应敏感,可塑性极小,在未来降水格局的背景下,对这两种景观进行研究可以为沙漠化或逆沙漠化进程提供基础资料[28,30].因此,本文通过在野外设置模拟控制增减水实验,研究分析固定沙丘和沙质草地生长季降水格局变化下植被特征、植物群落功能性状、土壤理化性质的变化规律和对地上生物量影响及调控机制,旨在了解科尔沁沙地自然恢复过程中植被的演替方向,为阐释逆转土地沙化过程和植被恢复提供一定的理论依据和实践支持. ...
科尔沁沙地植被恢复过程中群落组成及多样性演变特征
3
2022
... 科尔沁沙地原是科尔沁草原的主体部分,原生景观是疏林草原,较早的农业开发和近百年农牧交错带在这里大规模北侵,造成沙漠化急剧发展,成为中国现代沙漠化严重地区,也是中国沙漠化演变与防治研究的典型区域[27].近年来随着围栏封育措施的实施,该地区的土地沙漠化趋势得到遏制并发生逆转,植被和土壤都呈现一定程度的恢复趋势[28-29],相继出现了流动沙丘、半固定沙丘、固定沙丘和沙质草地交错分布的地貌类型.固定沙丘和沙质草地是反映科尔沁沙地沙漠化进程的关键景观特征,其中固定沙丘植物群落稳定性较高,而沙质草地生态环境脆弱,对人为干扰及气候变化反应敏感,可塑性极小,在未来降水格局的背景下,对这两种景观进行研究可以为沙漠化或逆沙漠化进程提供基础资料[28,30].因此,本文通过在野外设置模拟控制增减水实验,研究分析固定沙丘和沙质草地生长季降水格局变化下植被特征、植物群落功能性状、土壤理化性质的变化规律和对地上生物量影响及调控机制,旨在了解科尔沁沙地自然恢复过程中植被的演替方向,为阐释逆转土地沙化过程和植被恢复提供一定的理论依据和实践支持. ...
... [28,30].因此,本文通过在野外设置模拟控制增减水实验,研究分析固定沙丘和沙质草地生长季降水格局变化下植被特征、植物群落功能性状、土壤理化性质的变化规律和对地上生物量影响及调控机制,旨在了解科尔沁沙地自然恢复过程中植被的演替方向,为阐释逆转土地沙化过程和植被恢复提供一定的理论依据和实践支持. ...
... 在固定沙丘中,干旱降低Shannon-Wiener多样性指数的同时提高了Simpson优势度指数和地上生物量,这可能是因为5月和6月的极端干旱加剧了种间竞争[46],限制了竞争力较低草种的发芽,缺乏耐旱机制的植物将会减少,在生长发育早期就个体消亡[28,47],而具有耐旱和避旱机制的植物不仅不会受干旱条件的影响,甚至会生长得更好,因此地上生物量升高.这与以往在干旱、半干旱草原区开展的研究结果一致,即极端干旱或降水大幅波动会对草地植物群落结构和生物多样性产生影响,甚至会使整个生态系统的结构和功能发生改变,主要体现在植物群落优势种的更替上[21,48].在沙质草地中,生长季降水格局变化对物种丰富度、Simpson优势度指数和Shannon-Wiener多样性指数的影响不显著,主要原因可能是沙质草地中多年生白草和糙隐子草的优势度较高,对生长季降水格局变化有一定的缓冲能力[49],这也与多年生植物群落物种组成及结构的稳定性优于一二年生植物群落的研究结果相一致[50-51].干旱处理下,沙质草地的植被盖度和植物密度显著降低,这主要是因为长期干旱通过影响土壤水分进而抑制土壤潜在种子库的萌发和对植物群落生长产生滞后影响效应[9,52]. ...
科尔沁沙地封育过程中植被特征的动态变化
2
2018
... 科尔沁沙地原是科尔沁草原的主体部分,原生景观是疏林草原,较早的农业开发和近百年农牧交错带在这里大规模北侵,造成沙漠化急剧发展,成为中国现代沙漠化严重地区,也是中国沙漠化演变与防治研究的典型区域[27].近年来随着围栏封育措施的实施,该地区的土地沙漠化趋势得到遏制并发生逆转,植被和土壤都呈现一定程度的恢复趋势[28-29],相继出现了流动沙丘、半固定沙丘、固定沙丘和沙质草地交错分布的地貌类型.固定沙丘和沙质草地是反映科尔沁沙地沙漠化进程的关键景观特征,其中固定沙丘植物群落稳定性较高,而沙质草地生态环境脆弱,对人为干扰及气候变化反应敏感,可塑性极小,在未来降水格局的背景下,对这两种景观进行研究可以为沙漠化或逆沙漠化进程提供基础资料[28,30].因此,本文通过在野外设置模拟控制增减水实验,研究分析固定沙丘和沙质草地生长季降水格局变化下植被特征、植物群落功能性状、土壤理化性质的变化规律和对地上生物量影响及调控机制,旨在了解科尔沁沙地自然恢复过程中植被的演替方向,为阐释逆转土地沙化过程和植被恢复提供一定的理论依据和实践支持. ...
... 2022年8月在每个试验样地内建立1个1 m×1 m的样方进行实地调查,分物种估测植被盖度,用卷尺测量各物种的自然垂直高度(即为植物高度),采用齐地刈割的方法分物种获取地上植物,同时记录每种植物的株数(即为植物密度),样方内的物种个数即为物种丰富度[29,31].以上所得地上植物样品65 ℃烘干至恒重,从而得到地上生物量. ...
科尔沁沙质草地不同恢复年限草本层群落结构及其与土壤理化性质的关系
1
2022
... 科尔沁沙地原是科尔沁草原的主体部分,原生景观是疏林草原,较早的农业开发和近百年农牧交错带在这里大规模北侵,造成沙漠化急剧发展,成为中国现代沙漠化严重地区,也是中国沙漠化演变与防治研究的典型区域[27].近年来随着围栏封育措施的实施,该地区的土地沙漠化趋势得到遏制并发生逆转,植被和土壤都呈现一定程度的恢复趋势[28-29],相继出现了流动沙丘、半固定沙丘、固定沙丘和沙质草地交错分布的地貌类型.固定沙丘和沙质草地是反映科尔沁沙地沙漠化进程的关键景观特征,其中固定沙丘植物群落稳定性较高,而沙质草地生态环境脆弱,对人为干扰及气候变化反应敏感,可塑性极小,在未来降水格局的背景下,对这两种景观进行研究可以为沙漠化或逆沙漠化进程提供基础资料[28,30].因此,本文通过在野外设置模拟控制增减水实验,研究分析固定沙丘和沙质草地生长季降水格局变化下植被特征、植物群落功能性状、土壤理化性质的变化规律和对地上生物量影响及调控机制,旨在了解科尔沁沙地自然恢复过程中植被的演替方向,为阐释逆转土地沙化过程和植被恢复提供一定的理论依据和实践支持. ...
毛乌素荒漠草原植被特征对降水变化的响应
1
2022
... 2022年8月在每个试验样地内建立1个1 m×1 m的样方进行实地调查,分物种估测植被盖度,用卷尺测量各物种的自然垂直高度(即为植物高度),采用齐地刈割的方法分物种获取地上植物,同时记录每种植物的株数(即为植物密度),样方内的物种个数即为物种丰富度[29,31].以上所得地上植物样品65 ℃烘干至恒重,从而得到地上生物量. ...
New handbook for standardized measurement of plant functional traits worldwide
1
2013
... 根据植物功能性状取样和测定的国际标准方法[32-33],测定植物高度(Height)、叶片厚度(LT)、比叶面积(SLA)、叶干物质含量(LDMC)、叶片碳含量(LCC)和叶片氮含量(LNC). ...
A handbook of protocols for standardized and easy measurement of plant functional traits worldwide
1
2003
... 根据植物功能性状取样和测定的国际标准方法[32-33],测定植物高度(Height)、叶片厚度(LT)、比叶面积(SLA)、叶干物质含量(LDMC)、叶片碳含量(LCC)和叶片氮含量(LNC). ...
Effects of grazing exclusion on carbon sequestration and the associated vegetation and soil characteristics at a semi-arid desertified sandy site in Inner Mongolia,northern China
1
2012
... 在植被调查的同时,用直径为3 cm的土钻在每个调查样方内,以“五点取样法”钻取0~10 cm的土壤样品,过2 mm标准筛后取部分新鲜土壤样品装入小铝盒,用于测定土壤质量含水量,剩余土壤样品进行自然风干处理.部分风干土样研磨后用于测定土壤全碳、全氮含量,另一部分风干土样用于测定土壤pH、电导率、机械组成,用容重钻(环刀体积100 cm3)钻取0~10 cm土壤样品,用于测定土壤容重[34-35].采用元素分析仪(Costech ECS4010)测定土壤全碳、全氮含量,采用105 ℃烘干恒重法测定土壤质量含水量,pH计测定土壤酸碱度(水土比2.5∶1),电导率仪(水土比5∶1)测定土壤电导率,湿筛法测定土壤机械组成(粗砂>0.5 mm,中砂0.25~0.5 mm,细砂0.10~0.25 mm,极细砂0.05~0.10 mm,黏粉粒< 0.05 mm),环刀内的土壤样品105 ℃烘干至恒重即为土壤容重. ...
Soil net nitrogen transformation rates are co-determined by multiple factors during the landscape evolution in Horqin Sandy Land
1
2021
... 在植被调查的同时,用直径为3 cm的土钻在每个调查样方内,以“五点取样法”钻取0~10 cm的土壤样品,过2 mm标准筛后取部分新鲜土壤样品装入小铝盒,用于测定土壤质量含水量,剩余土壤样品进行自然风干处理.部分风干土样研磨后用于测定土壤全碳、全氮含量,另一部分风干土样用于测定土壤pH、电导率、机械组成,用容重钻(环刀体积100 cm3)钻取0~10 cm土壤样品,用于测定土壤容重[34-35].采用元素分析仪(Costech ECS4010)测定土壤全碳、全氮含量,采用105 ℃烘干恒重法测定土壤质量含水量,pH计测定土壤酸碱度(水土比2.5∶1),电导率仪(水土比5∶1)测定土壤电导率,湿筛法测定土壤机械组成(粗砂>0.5 mm,中砂0.25~0.5 mm,细砂0.10~0.25 mm,极细砂0.05~0.10 mm,黏粉粒< 0.05 mm),环刀内的土壤样品105 ℃烘干至恒重即为土壤容重. ...
放牧强度对科尔沁沙地沙质草地植被的影响
1
2016
... 在植被调查的同时,用直径为3 cm的土钻在每个调查样方内,以“五点取样法”钻取0~10 cm的土壤样品,过2 mm标准筛后取部分新鲜土壤样品装入小铝盒,用于测定土壤质量含水量,剩余土壤样品进行自然风干处理.部分风干土样研磨后用于测定土壤全碳、全氮含量,另一部分风干土样用于测定土壤pH、电导率、机械组成,用容重钻(环刀体积100 cm
3)钻取0~10 cm土壤样品,用于测定土壤容重
[34-35].采用元素分析仪(Costech ECS4010)测定土壤全碳、全氮含量,采用105 ℃烘干恒重法测定土壤质量含水量,pH计测定土壤酸碱度(水土比2.5∶1),电导率仪(水土比5∶1)测定土壤电导率,湿筛法测定土壤机械组成(粗砂>0.5 mm,中砂0.25~0.5 mm,细砂0.10~0.25 mm,极细砂0.05~0.10 mm,黏粉粒< 0.05 mm),环刀内的土壤样品105 ℃烘干至恒重即为土壤容重.
<strong>1.4</strong> 数据分析<strong>1.4.1</strong> 物种多样性<sup>[<xref ref-type="bibr" rid="R23">23</xref>,<xref ref-type="bibr" rid="R36">36</xref>]</sup>物种重要值=(相对盖度+相对高度+相对密度+相对地上生物量)/4 ...
植物群落功能多样性计算方法
1
2011
... 式中:
Ni 为种
i的相对重要值;
S为植物群落物种数.
<strong>1.4.2</strong> 植物群落功能性状<sup>[<xref ref-type="bibr" rid="R37">37</xref>]</sup>植物群落功能性状(CWM),即就某一性状而言,群落内所有该性状的值与其相对地上生物量积的总和,表示植物群落内某一功能性状的均值. ...
半干旱沙地草本植物群落特征对短期降水变化的响应
1
2020
... 物种丰富度和生态优势度能全面衡量物种多样性,反映植物群落物种组成和结构水平[38-39].植物群落生活型结构是植物群落结构的重要组成部分,不同生活型植物的生态适应能力以重要值作为指标,且重要值的大小是确定优势和亚优势植物的唯一指标[40-41].本研究中固定沙丘和沙质草地在生长季降水格局变化下优势物种生活型发生较大幅度变化,表现为干旱处理下一年生植物的重要值显著增加而多年生植物的重要值显著降低,尤其是虎尾草和蒺藜等一二年生植物的重要值显著提高,主要原因为虎尾草和蒺藜属于C4植物,这些植物在干旱、炎热生境中的水分竞争力更强,对环境水分高度敏感且能够顺应气候波动,在干旱胁迫时期能通过调节生理、生长和繁殖策略,快速完成生活史[42-43].多年生植物根系发达且深厚,该区域土壤颗粒较大且持水力较低[44],干旱处理下土壤含水量降低时多年生植物的根系无法从深层土壤获得足够的水分,因此重要值显著降低[45]. ...
科尔沁沙地草甸草场退化的原因与植物多样性变化
1
2006
... 物种丰富度和生态优势度能全面衡量物种多样性,反映植物群落物种组成和结构水平[38-39].植物群落生活型结构是植物群落结构的重要组成部分,不同生活型植物的生态适应能力以重要值作为指标,且重要值的大小是确定优势和亚优势植物的唯一指标[40-41].本研究中固定沙丘和沙质草地在生长季降水格局变化下优势物种生活型发生较大幅度变化,表现为干旱处理下一年生植物的重要值显著增加而多年生植物的重要值显著降低,尤其是虎尾草和蒺藜等一二年生植物的重要值显著提高,主要原因为虎尾草和蒺藜属于C4植物,这些植物在干旱、炎热生境中的水分竞争力更强,对环境水分高度敏感且能够顺应气候波动,在干旱胁迫时期能通过调节生理、生长和繁殖策略,快速完成生活史[42-43].多年生植物根系发达且深厚,该区域土壤颗粒较大且持水力较低[44],干旱处理下土壤含水量降低时多年生植物的根系无法从深层土壤获得足够的水分,因此重要值显著降低[45]. ...
黄土丘陵区天然群落的植物组成、植物多样性及其与环境因子的关系
1
2012
... 物种丰富度和生态优势度能全面衡量物种多样性,反映植物群落物种组成和结构水平[38-39].植物群落生活型结构是植物群落结构的重要组成部分,不同生活型植物的生态适应能力以重要值作为指标,且重要值的大小是确定优势和亚优势植物的唯一指标[40-41].本研究中固定沙丘和沙质草地在生长季降水格局变化下优势物种生活型发生较大幅度变化,表现为干旱处理下一年生植物的重要值显著增加而多年生植物的重要值显著降低,尤其是虎尾草和蒺藜等一二年生植物的重要值显著提高,主要原因为虎尾草和蒺藜属于C4植物,这些植物在干旱、炎热生境中的水分竞争力更强,对环境水分高度敏感且能够顺应气候波动,在干旱胁迫时期能通过调节生理、生长和繁殖策略,快速完成生活史[42-43].多年生植物根系发达且深厚,该区域土壤颗粒较大且持水力较低[44],干旱处理下土壤含水量降低时多年生植物的根系无法从深层土壤获得足够的水分,因此重要值显著降低[45]. ...
宁夏荒漠草原不同群落生物多样性与生物量关系及影响因子分析
1
2015
... 物种丰富度和生态优势度能全面衡量物种多样性,反映植物群落物种组成和结构水平[38-39].植物群落生活型结构是植物群落结构的重要组成部分,不同生活型植物的生态适应能力以重要值作为指标,且重要值的大小是确定优势和亚优势植物的唯一指标[40-41].本研究中固定沙丘和沙质草地在生长季降水格局变化下优势物种生活型发生较大幅度变化,表现为干旱处理下一年生植物的重要值显著增加而多年生植物的重要值显著降低,尤其是虎尾草和蒺藜等一二年生植物的重要值显著提高,主要原因为虎尾草和蒺藜属于C4植物,这些植物在干旱、炎热生境中的水分竞争力更强,对环境水分高度敏感且能够顺应气候波动,在干旱胁迫时期能通过调节生理、生长和繁殖策略,快速完成生活史[42-43].多年生植物根系发达且深厚,该区域土壤颗粒较大且持水力较低[44],干旱处理下土壤含水量降低时多年生植物的根系无法从深层土壤获得足够的水分,因此重要值显著降低[45]. ...
阿拉善荒漠区一年生植物层片物种多样性及其分布特征
1
2003
... 物种丰富度和生态优势度能全面衡量物种多样性,反映植物群落物种组成和结构水平[38-39].植物群落生活型结构是植物群落结构的重要组成部分,不同生活型植物的生态适应能力以重要值作为指标,且重要值的大小是确定优势和亚优势植物的唯一指标[40-41].本研究中固定沙丘和沙质草地在生长季降水格局变化下优势物种生活型发生较大幅度变化,表现为干旱处理下一年生植物的重要值显著增加而多年生植物的重要值显著降低,尤其是虎尾草和蒺藜等一二年生植物的重要值显著提高,主要原因为虎尾草和蒺藜属于C4植物,这些植物在干旱、炎热生境中的水分竞争力更强,对环境水分高度敏感且能够顺应气候波动,在干旱胁迫时期能通过调节生理、生长和繁殖策略,快速完成生活史[42-43].多年生植物根系发达且深厚,该区域土壤颗粒较大且持水力较低[44],干旱处理下土壤含水量降低时多年生植物的根系无法从深层土壤获得足够的水分,因此重要值显著降低[45]. ...
一年生植物层片物种多样性及其分布特征
1
2010
... 物种丰富度和生态优势度能全面衡量物种多样性,反映植物群落物种组成和结构水平[38-39].植物群落生活型结构是植物群落结构的重要组成部分,不同生活型植物的生态适应能力以重要值作为指标,且重要值的大小是确定优势和亚优势植物的唯一指标[40-41].本研究中固定沙丘和沙质草地在生长季降水格局变化下优势物种生活型发生较大幅度变化,表现为干旱处理下一年生植物的重要值显著增加而多年生植物的重要值显著降低,尤其是虎尾草和蒺藜等一二年生植物的重要值显著提高,主要原因为虎尾草和蒺藜属于C4植物,这些植物在干旱、炎热生境中的水分竞争力更强,对环境水分高度敏感且能够顺应气候波动,在干旱胁迫时期能通过调节生理、生长和繁殖策略,快速完成生活史[42-43].多年生植物根系发达且深厚,该区域土壤颗粒较大且持水力较低[44],干旱处理下土壤含水量降低时多年生植物的根系无法从深层土壤获得足够的水分,因此重要值显著降低[45]. ...
Plant responses to warming and increased precipitation in three categories of dune stabilization in northeastern China
1
2017
... 物种丰富度和生态优势度能全面衡量物种多样性,反映植物群落物种组成和结构水平[38-39].植物群落生活型结构是植物群落结构的重要组成部分,不同生活型植物的生态适应能力以重要值作为指标,且重要值的大小是确定优势和亚优势植物的唯一指标[40-41].本研究中固定沙丘和沙质草地在生长季降水格局变化下优势物种生活型发生较大幅度变化,表现为干旱处理下一年生植物的重要值显著增加而多年生植物的重要值显著降低,尤其是虎尾草和蒺藜等一二年生植物的重要值显著提高,主要原因为虎尾草和蒺藜属于C4植物,这些植物在干旱、炎热生境中的水分竞争力更强,对环境水分高度敏感且能够顺应气候波动,在干旱胁迫时期能通过调节生理、生长和繁殖策略,快速完成生活史[42-43].多年生植物根系发达且深厚,该区域土壤颗粒较大且持水力较低[44],干旱处理下土壤含水量降低时多年生植物的根系无法从深层土壤获得足够的水分,因此重要值显著降低[45]. ...
Above-and belowground biomass in relation to environmental factors in temperate grasslands,Inner Mongolia
1
2008
... 物种丰富度和生态优势度能全面衡量物种多样性,反映植物群落物种组成和结构水平[38-39].植物群落生活型结构是植物群落结构的重要组成部分,不同生活型植物的生态适应能力以重要值作为指标,且重要值的大小是确定优势和亚优势植物的唯一指标[40-41].本研究中固定沙丘和沙质草地在生长季降水格局变化下优势物种生活型发生较大幅度变化,表现为干旱处理下一年生植物的重要值显著增加而多年生植物的重要值显著降低,尤其是虎尾草和蒺藜等一二年生植物的重要值显著提高,主要原因为虎尾草和蒺藜属于C4植物,这些植物在干旱、炎热生境中的水分竞争力更强,对环境水分高度敏感且能够顺应气候波动,在干旱胁迫时期能通过调节生理、生长和繁殖策略,快速完成生活史[42-43].多年生植物根系发达且深厚,该区域土壤颗粒较大且持水力较低[44],干旱处理下土壤含水量降低时多年生植物的根系无法从深层土壤获得足够的水分,因此重要值显著降低[45]. ...
科尔沁草地植被结构与功能关系及其对人类活动和气候变化的响应
1
2019
... 在固定沙丘中,干旱降低Shannon-Wiener多样性指数的同时提高了Simpson优势度指数和地上生物量,这可能是因为5月和6月的极端干旱加剧了种间竞争[46],限制了竞争力较低草种的发芽,缺乏耐旱机制的植物将会减少,在生长发育早期就个体消亡[28,47],而具有耐旱和避旱机制的植物不仅不会受干旱条件的影响,甚至会生长得更好,因此地上生物量升高.这与以往在干旱、半干旱草原区开展的研究结果一致,即极端干旱或降水大幅波动会对草地植物群落结构和生物多样性产生影响,甚至会使整个生态系统的结构和功能发生改变,主要体现在植物群落优势种的更替上[21,48].在沙质草地中,生长季降水格局变化对物种丰富度、Simpson优势度指数和Shannon-Wiener多样性指数的影响不显著,主要原因可能是沙质草地中多年生白草和糙隐子草的优势度较高,对生长季降水格局变化有一定的缓冲能力[49],这也与多年生植物群落物种组成及结构的稳定性优于一二年生植物群落的研究结果相一致[50-51].干旱处理下,沙质草地的植被盖度和植物密度显著降低,这主要是因为长期干旱通过影响土壤水分进而抑制土壤潜在种子库的萌发和对植物群落生长产生滞后影响效应[9,52]. ...
Effects of rainfall manipulation and nitrogen addition on plant biomass allocation in a semiarid sandy grassland
1
2020
... 在固定沙丘中,干旱降低Shannon-Wiener多样性指数的同时提高了Simpson优势度指数和地上生物量,这可能是因为5月和6月的极端干旱加剧了种间竞争[46],限制了竞争力较低草种的发芽,缺乏耐旱机制的植物将会减少,在生长发育早期就个体消亡[28,47],而具有耐旱和避旱机制的植物不仅不会受干旱条件的影响,甚至会生长得更好,因此地上生物量升高.这与以往在干旱、半干旱草原区开展的研究结果一致,即极端干旱或降水大幅波动会对草地植物群落结构和生物多样性产生影响,甚至会使整个生态系统的结构和功能发生改变,主要体现在植物群落优势种的更替上[21,48].在沙质草地中,生长季降水格局变化对物种丰富度、Simpson优势度指数和Shannon-Wiener多样性指数的影响不显著,主要原因可能是沙质草地中多年生白草和糙隐子草的优势度较高,对生长季降水格局变化有一定的缓冲能力[49],这也与多年生植物群落物种组成及结构的稳定性优于一二年生植物群落的研究结果相一致[50-51].干旱处理下,沙质草地的植被盖度和植物密度显著降低,这主要是因为长期干旱通过影响土壤水分进而抑制土壤潜在种子库的萌发和对植物群落生长产生滞后影响效应[9,52]. ...
Drought effect on plant nitrogen andphosphorus:a meta-analysis
1
2014
... 在固定沙丘中,干旱降低Shannon-Wiener多样性指数的同时提高了Simpson优势度指数和地上生物量,这可能是因为5月和6月的极端干旱加剧了种间竞争[46],限制了竞争力较低草种的发芽,缺乏耐旱机制的植物将会减少,在生长发育早期就个体消亡[28,47],而具有耐旱和避旱机制的植物不仅不会受干旱条件的影响,甚至会生长得更好,因此地上生物量升高.这与以往在干旱、半干旱草原区开展的研究结果一致,即极端干旱或降水大幅波动会对草地植物群落结构和生物多样性产生影响,甚至会使整个生态系统的结构和功能发生改变,主要体现在植物群落优势种的更替上[21,48].在沙质草地中,生长季降水格局变化对物种丰富度、Simpson优势度指数和Shannon-Wiener多样性指数的影响不显著,主要原因可能是沙质草地中多年生白草和糙隐子草的优势度较高,对生长季降水格局变化有一定的缓冲能力[49],这也与多年生植物群落物种组成及结构的稳定性优于一二年生植物群落的研究结果相一致[50-51].干旱处理下,沙质草地的植被盖度和植物密度显著降低,这主要是因为长期干旱通过影响土壤水分进而抑制土壤潜在种子库的萌发和对植物群落生长产生滞后影响效应[9,52]. ...
Root and shoot competition:a meta-analysis
1
2013
... 在固定沙丘中,干旱降低Shannon-Wiener多样性指数的同时提高了Simpson优势度指数和地上生物量,这可能是因为5月和6月的极端干旱加剧了种间竞争[46],限制了竞争力较低草种的发芽,缺乏耐旱机制的植物将会减少,在生长发育早期就个体消亡[28,47],而具有耐旱和避旱机制的植物不仅不会受干旱条件的影响,甚至会生长得更好,因此地上生物量升高.这与以往在干旱、半干旱草原区开展的研究结果一致,即极端干旱或降水大幅波动会对草地植物群落结构和生物多样性产生影响,甚至会使整个生态系统的结构和功能发生改变,主要体现在植物群落优势种的更替上[21,48].在沙质草地中,生长季降水格局变化对物种丰富度、Simpson优势度指数和Shannon-Wiener多样性指数的影响不显著,主要原因可能是沙质草地中多年生白草和糙隐子草的优势度较高,对生长季降水格局变化有一定的缓冲能力[49],这也与多年生植物群落物种组成及结构的稳定性优于一二年生植物群落的研究结果相一致[50-51].干旱处理下,沙质草地的植被盖度和植物密度显著降低,这主要是因为长期干旱通过影响土壤水分进而抑制土壤潜在种子库的萌发和对植物群落生长产生滞后影响效应[9,52]. ...
乌兰布和沙漠绿洲东缘植被群落结构及物种多样性研究
1
2016
... 在固定沙丘中,干旱降低Shannon-Wiener多样性指数的同时提高了Simpson优势度指数和地上生物量,这可能是因为5月和6月的极端干旱加剧了种间竞争[46],限制了竞争力较低草种的发芽,缺乏耐旱机制的植物将会减少,在生长发育早期就个体消亡[28,47],而具有耐旱和避旱机制的植物不仅不会受干旱条件的影响,甚至会生长得更好,因此地上生物量升高.这与以往在干旱、半干旱草原区开展的研究结果一致,即极端干旱或降水大幅波动会对草地植物群落结构和生物多样性产生影响,甚至会使整个生态系统的结构和功能发生改变,主要体现在植物群落优势种的更替上[21,48].在沙质草地中,生长季降水格局变化对物种丰富度、Simpson优势度指数和Shannon-Wiener多样性指数的影响不显著,主要原因可能是沙质草地中多年生白草和糙隐子草的优势度较高,对生长季降水格局变化有一定的缓冲能力[49],这也与多年生植物群落物种组成及结构的稳定性优于一二年生植物群落的研究结果相一致[50-51].干旱处理下,沙质草地的植被盖度和植物密度显著降低,这主要是因为长期干旱通过影响土壤水分进而抑制土壤潜在种子库的萌发和对植物群落生长产生滞后影响效应[9,52]. ...
青藏高原高寒草地生物多样性与生态系统功能的关系
1
2018
... 在固定沙丘中,干旱降低Shannon-Wiener多样性指数的同时提高了Simpson优势度指数和地上生物量,这可能是因为5月和6月的极端干旱加剧了种间竞争[46],限制了竞争力较低草种的发芽,缺乏耐旱机制的植物将会减少,在生长发育早期就个体消亡[28,47],而具有耐旱和避旱机制的植物不仅不会受干旱条件的影响,甚至会生长得更好,因此地上生物量升高.这与以往在干旱、半干旱草原区开展的研究结果一致,即极端干旱或降水大幅波动会对草地植物群落结构和生物多样性产生影响,甚至会使整个生态系统的结构和功能发生改变,主要体现在植物群落优势种的更替上[21,48].在沙质草地中,生长季降水格局变化对物种丰富度、Simpson优势度指数和Shannon-Wiener多样性指数的影响不显著,主要原因可能是沙质草地中多年生白草和糙隐子草的优势度较高,对生长季降水格局变化有一定的缓冲能力[49],这也与多年生植物群落物种组成及结构的稳定性优于一二年生植物群落的研究结果相一致[50-51].干旱处理下,沙质草地的植被盖度和植物密度显著降低,这主要是因为长期干旱通过影响土壤水分进而抑制土壤潜在种子库的萌发和对植物群落生长产生滞后影响效应[9,52]. ...
模拟增雨条件下沙质草地地表植被和节肢动物群落变化特征
1
2014
... 在固定沙丘中,干旱降低Shannon-Wiener多样性指数的同时提高了Simpson优势度指数和地上生物量,这可能是因为5月和6月的极端干旱加剧了种间竞争[46],限制了竞争力较低草种的发芽,缺乏耐旱机制的植物将会减少,在生长发育早期就个体消亡[28,47],而具有耐旱和避旱机制的植物不仅不会受干旱条件的影响,甚至会生长得更好,因此地上生物量升高.这与以往在干旱、半干旱草原区开展的研究结果一致,即极端干旱或降水大幅波动会对草地植物群落结构和生物多样性产生影响,甚至会使整个生态系统的结构和功能发生改变,主要体现在植物群落优势种的更替上[21,48].在沙质草地中,生长季降水格局变化对物种丰富度、Simpson优势度指数和Shannon-Wiener多样性指数的影响不显著,主要原因可能是沙质草地中多年生白草和糙隐子草的优势度较高,对生长季降水格局变化有一定的缓冲能力[49],这也与多年生植物群落物种组成及结构的稳定性优于一二年生植物群落的研究结果相一致[50-51].干旱处理下,沙质草地的植被盖度和植物密度显著降低,这主要是因为长期干旱通过影响土壤水分进而抑制土壤潜在种子库的萌发和对植物群落生长产生滞后影响效应[9,52]. ...
不同土壤水分条件下焕镛木幼苗的生理生态特性
1
2004
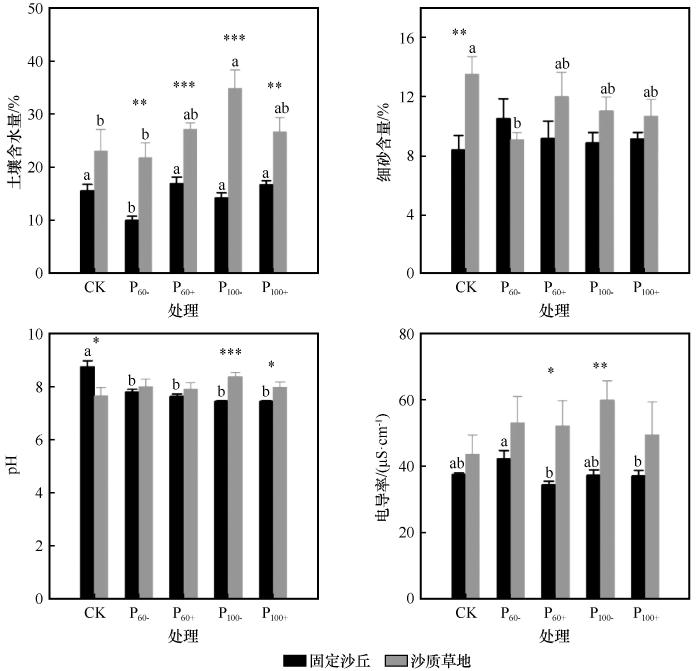
... 研究表明干旱、半干旱区植物群落功能性状在生长季降水格局变化时会发生较大幅度改变,如增加降水会提高植物群落的高度和比叶面积[53],而干旱胁迫会提高植物叶干物质含量和叶片氮含量[54-55],主要原因是草地植物叶片渗透水含量、叶绿素含量均会随着湿润到干旱环境的变化而升高,将光合作用合成的干物质更多投入到叶片构建,增大叶肉密度的同时降低比叶面积,从而提高对干旱环境的抗性[56].然而在本研究中,生长季降水格局变化对科尔沁沙地草地植物群落功能性状的影响不显著,主要原因一方面可能为降水格局变化形成的水分梯度没有达到对植物群落功能性状影响的阈值[57],另一方面可能为降水、干旱时长和频率对植物功能性状的影响具有“累加效应”,大量长期气候变化试验结果也表明植物群落功能性状的显著变化也是在持续干旱的最后几年才逐渐突显[58].此外,在沙质草地中生长季前期极端干旱显著提高了土壤含水量,主要原因是生长季前期减少降雨抑制植物种子萌发和生长,生长季后期的植被盖度、丰富度、生物量等指标都相对较低,8月取样时,因植物个体较少、植被耗水低,反而使土壤含水量提高. ...
科尔沁沙地52种植物叶片性状变异特征研究
1
2010
... 研究表明干旱、半干旱区植物群落功能性状在生长季降水格局变化时会发生较大幅度改变,如增加降水会提高植物群落的高度和比叶面积[53],而干旱胁迫会提高植物叶干物质含量和叶片氮含量[54-55],主要原因是草地植物叶片渗透水含量、叶绿素含量均会随着湿润到干旱环境的变化而升高,将光合作用合成的干物质更多投入到叶片构建,增大叶肉密度的同时降低比叶面积,从而提高对干旱环境的抗性[56].然而在本研究中,生长季降水格局变化对科尔沁沙地草地植物群落功能性状的影响不显著,主要原因一方面可能为降水格局变化形成的水分梯度没有达到对植物群落功能性状影响的阈值[57],另一方面可能为降水、干旱时长和频率对植物功能性状的影响具有“累加效应”,大量长期气候变化试验结果也表明植物群落功能性状的显著变化也是在持续干旱的最后几年才逐渐突显[58].此外,在沙质草地中生长季前期极端干旱显著提高了土壤含水量,主要原因是生长季前期减少降雨抑制植物种子萌发和生长,生长季后期的植被盖度、丰富度、生物量等指标都相对较低,8月取样时,因植物个体较少、植被耗水低,反而使土壤含水量提高. ...
植物叶片功能性状及其环境适应研究
1
2017
... 研究表明干旱、半干旱区植物群落功能性状在生长季降水格局变化时会发生较大幅度改变,如增加降水会提高植物群落的高度和比叶面积[53],而干旱胁迫会提高植物叶干物质含量和叶片氮含量[54-55],主要原因是草地植物叶片渗透水含量、叶绿素含量均会随着湿润到干旱环境的变化而升高,将光合作用合成的干物质更多投入到叶片构建,增大叶肉密度的同时降低比叶面积,从而提高对干旱环境的抗性[56].然而在本研究中,生长季降水格局变化对科尔沁沙地草地植物群落功能性状的影响不显著,主要原因一方面可能为降水格局变化形成的水分梯度没有达到对植物群落功能性状影响的阈值[57],另一方面可能为降水、干旱时长和频率对植物功能性状的影响具有“累加效应”,大量长期气候变化试验结果也表明植物群落功能性状的显著变化也是在持续干旱的最后几年才逐渐突显[58].此外,在沙质草地中生长季前期极端干旱显著提高了土壤含水量,主要原因是生长季前期减少降雨抑制植物种子萌发和生长,生长季后期的植被盖度、丰富度、生物量等指标都相对较低,8月取样时,因植物个体较少、植被耗水低,反而使土壤含水量提高. ...
降雨极端化对喀斯特草本植物功能性状的影响
1
2022
... 研究表明干旱、半干旱区植物群落功能性状在生长季降水格局变化时会发生较大幅度改变,如增加降水会提高植物群落的高度和比叶面积[53],而干旱胁迫会提高植物叶干物质含量和叶片氮含量[54-55],主要原因是草地植物叶片渗透水含量、叶绿素含量均会随着湿润到干旱环境的变化而升高,将光合作用合成的干物质更多投入到叶片构建,增大叶肉密度的同时降低比叶面积,从而提高对干旱环境的抗性[56].然而在本研究中,生长季降水格局变化对科尔沁沙地草地植物群落功能性状的影响不显著,主要原因一方面可能为降水格局变化形成的水分梯度没有达到对植物群落功能性状影响的阈值[57],另一方面可能为降水、干旱时长和频率对植物功能性状的影响具有“累加效应”,大量长期气候变化试验结果也表明植物群落功能性状的显著变化也是在持续干旱的最后几年才逐渐突显[58].此外,在沙质草地中生长季前期极端干旱显著提高了土壤含水量,主要原因是生长季前期减少降雨抑制植物种子萌发和生长,生长季后期的植被盖度、丰富度、生物量等指标都相对较低,8月取样时,因植物个体较少、植被耗水低,反而使土壤含水量提高. ...
沙质草地植物功能性状对放牧、增水、氮添加及其耦合效应的响应机制
1
2021
... 研究表明干旱、半干旱区植物群落功能性状在生长季降水格局变化时会发生较大幅度改变,如增加降水会提高植物群落的高度和比叶面积[53],而干旱胁迫会提高植物叶干物质含量和叶片氮含量[54-55],主要原因是草地植物叶片渗透水含量、叶绿素含量均会随着湿润到干旱环境的变化而升高,将光合作用合成的干物质更多投入到叶片构建,增大叶肉密度的同时降低比叶面积,从而提高对干旱环境的抗性[56].然而在本研究中,生长季降水格局变化对科尔沁沙地草地植物群落功能性状的影响不显著,主要原因一方面可能为降水格局变化形成的水分梯度没有达到对植物群落功能性状影响的阈值[57],另一方面可能为降水、干旱时长和频率对植物功能性状的影响具有“累加效应”,大量长期气候变化试验结果也表明植物群落功能性状的显著变化也是在持续干旱的最后几年才逐渐突显[58].此外,在沙质草地中生长季前期极端干旱显著提高了土壤含水量,主要原因是生长季前期减少降雨抑制植物种子萌发和生长,生长季后期的植被盖度、丰富度、生物量等指标都相对较低,8月取样时,因植物个体较少、植被耗水低,反而使土壤含水量提高. ...
Shifts in plant functional composition following long-term drought in grasslands
1
2019
... 研究表明干旱、半干旱区植物群落功能性状在生长季降水格局变化时会发生较大幅度改变,如增加降水会提高植物群落的高度和比叶面积[53],而干旱胁迫会提高植物叶干物质含量和叶片氮含量[54-55],主要原因是草地植物叶片渗透水含量、叶绿素含量均会随着湿润到干旱环境的变化而升高,将光合作用合成的干物质更多投入到叶片构建,增大叶肉密度的同时降低比叶面积,从而提高对干旱环境的抗性[56].然而在本研究中,生长季降水格局变化对科尔沁沙地草地植物群落功能性状的影响不显著,主要原因一方面可能为降水格局变化形成的水分梯度没有达到对植物群落功能性状影响的阈值[57],另一方面可能为降水、干旱时长和频率对植物功能性状的影响具有“累加效应”,大量长期气候变化试验结果也表明植物群落功能性状的显著变化也是在持续干旱的最后几年才逐渐突显[58].此外,在沙质草地中生长季前期极端干旱显著提高了土壤含水量,主要原因是生长季前期减少降雨抑制植物种子萌发和生长,生长季后期的植被盖度、丰富度、生物量等指标都相对较低,8月取样时,因植物个体较少、植被耗水低,反而使土壤含水量提高. ...
草地生态系统中土壤氮素矿化影响因素的研究进展
1
2004
... 土壤是植物生长发育的基础,为植物生长提供必需的水分和养分,且不同生境中土壤理化性质存在较大差异[59].本研究中土壤含水量对Shannon-Wiener多样性指数具有正向影响,而Shannon-Wiener多样性指数对于地上生物量具有负向影响,这主要是因为土壤含水量升高刺激了休眠种子萌发,促使水分竞争力弱的植物大量生长,这些植物的生长占用了光照和养分资源,从一定程度上抑制了单株生物量较高的优势植物的生长,从而降低了地上生物量.细砂含量影响植物群落高度的同时进一步对地上生物量产生正向影响,这也与以往的研究结果一致,表明土壤中细颗粒物质能够提高土壤有机质和养分含量进而促进地上生物量的积累,该结论进一步明确了土壤颗粒组成对养分固持起着关键性作用[60]. ...
退化沙质草地恢复过程土壤颗粒组成变化对土壤-植被系统稳定性的影响
1
2009
... 土壤是植物生长发育的基础,为植物生长提供必需的水分和养分,且不同生境中土壤理化性质存在较大差异[59].本研究中土壤含水量对Shannon-Wiener多样性指数具有正向影响,而Shannon-Wiener多样性指数对于地上生物量具有负向影响,这主要是因为土壤含水量升高刺激了休眠种子萌发,促使水分竞争力弱的植物大量生长,这些植物的生长占用了光照和养分资源,从一定程度上抑制了单株生物量较高的优势植物的生长,从而降低了地上生物量.细砂含量影响植物群落高度的同时进一步对地上生物量产生正向影响,这也与以往的研究结果一致,表明土壤中细颗粒物质能够提高土壤有机质和养分含量进而促进地上生物量的积累,该结论进一步明确了土壤颗粒组成对养分固持起着关键性作用[60]. ...







 甘公网安备 62010202000688号
甘公网安备 62010202000688号
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}