Climate Change and Land:An IPCC Special Report on Climate Change
1
... 联合国政府间气候变化专门委员会(IPCC)特别评估报告指出,与1850—1900年相比,2006—2015年全球陆地表面平均气温上升了1.53 ℃,未来几十年气温上升速率可能更大[1].高海拔地区增温幅度显著高于北半球同纬度地区[2].青藏高原平均海拔大于4 000 m,对中国乃至整个亚洲的生态环境具有重要的意义,也是气候变化敏感区和生态脆弱带[3-4],青藏高原高寒草地生态系统对温度升高的响应更为敏感而迅速[5].高寒草甸作为青藏高原草地主要类型,草场面积约为7×105 km2,约占青藏高原可利用草地面积的50%,是中国畜牧业重要生产基地[6].气温上升将对高寒草甸生态系统产生重要影响,进而影响其载畜能力.因此,在气候变暖背景下,制定合理的高寒草甸载畜量是维系中国草地和畜牧业可持续发展的重要举措. ...
Extinction risk from climate change
1
2004
... 联合国政府间气候变化专门委员会(IPCC)特别评估报告指出,与1850—1900年相比,2006—2015年全球陆地表面平均气温上升了1.53 ℃,未来几十年气温上升速率可能更大[1].高海拔地区增温幅度显著高于北半球同纬度地区[2].青藏高原平均海拔大于4 000 m,对中国乃至整个亚洲的生态环境具有重要的意义,也是气候变化敏感区和生态脆弱带[3-4],青藏高原高寒草地生态系统对温度升高的响应更为敏感而迅速[5].高寒草甸作为青藏高原草地主要类型,草场面积约为7×105 km2,约占青藏高原可利用草地面积的50%,是中国畜牧业重要生产基地[6].气温上升将对高寒草甸生态系统产生重要影响,进而影响其载畜能力.因此,在气候变暖背景下,制定合理的高寒草甸载畜量是维系中国草地和畜牧业可持续发展的重要举措. ...
Recent glacial retreat and its impact on hydrological processes on the Tibetan Plateau,China,and surrounding regions
1
2007
... 联合国政府间气候变化专门委员会(IPCC)特别评估报告指出,与1850—1900年相比,2006—2015年全球陆地表面平均气温上升了1.53 ℃,未来几十年气温上升速率可能更大[1].高海拔地区增温幅度显著高于北半球同纬度地区[2].青藏高原平均海拔大于4 000 m,对中国乃至整个亚洲的生态环境具有重要的意义,也是气候变化敏感区和生态脆弱带[3-4],青藏高原高寒草地生态系统对温度升高的响应更为敏感而迅速[5].高寒草甸作为青藏高原草地主要类型,草场面积约为7×105 km2,约占青藏高原可利用草地面积的50%,是中国畜牧业重要生产基地[6].气温上升将对高寒草甸生态系统产生重要影响,进而影响其载畜能力.因此,在气候变暖背景下,制定合理的高寒草甸载畜量是维系中国草地和畜牧业可持续发展的重要举措. ...
Rangeland degradation on the Qinghai-Tibetan Plateau:a review of the evidence of its magnitude and causes
1
2010
... 联合国政府间气候变化专门委员会(IPCC)特别评估报告指出,与1850—1900年相比,2006—2015年全球陆地表面平均气温上升了1.53 ℃,未来几十年气温上升速率可能更大[1].高海拔地区增温幅度显著高于北半球同纬度地区[2].青藏高原平均海拔大于4 000 m,对中国乃至整个亚洲的生态环境具有重要的意义,也是气候变化敏感区和生态脆弱带[3-4],青藏高原高寒草地生态系统对温度升高的响应更为敏感而迅速[5].高寒草甸作为青藏高原草地主要类型,草场面积约为7×105 km2,约占青藏高原可利用草地面积的50%,是中国畜牧业重要生产基地[6].气温上升将对高寒草甸生态系统产生重要影响,进而影响其载畜能力.因此,在气候变暖背景下,制定合理的高寒草甸载畜量是维系中国草地和畜牧业可持续发展的重要举措. ...
Responses of tundra soil microbial communities to half a decade of experimental warming at two critical depths
1
2019
... 联合国政府间气候变化专门委员会(IPCC)特别评估报告指出,与1850—1900年相比,2006—2015年全球陆地表面平均气温上升了1.53 ℃,未来几十年气温上升速率可能更大[1].高海拔地区增温幅度显著高于北半球同纬度地区[2].青藏高原平均海拔大于4 000 m,对中国乃至整个亚洲的生态环境具有重要的意义,也是气候变化敏感区和生态脆弱带[3-4],青藏高原高寒草地生态系统对温度升高的响应更为敏感而迅速[5].高寒草甸作为青藏高原草地主要类型,草场面积约为7×105 km2,约占青藏高原可利用草地面积的50%,是中国畜牧业重要生产基地[6].气温上升将对高寒草甸生态系统产生重要影响,进而影响其载畜能力.因此,在气候变暖背景下,制定合理的高寒草甸载畜量是维系中国草地和畜牧业可持续发展的重要举措. ...
Comparison of ecosystem characteristics between degraded and intact alpine meadow in the Qinghai-Tibetan Plateau,China
1
2014
... 联合国政府间气候变化专门委员会(IPCC)特别评估报告指出,与1850—1900年相比,2006—2015年全球陆地表面平均气温上升了1.53 ℃,未来几十年气温上升速率可能更大[1].高海拔地区增温幅度显著高于北半球同纬度地区[2].青藏高原平均海拔大于4 000 m,对中国乃至整个亚洲的生态环境具有重要的意义,也是气候变化敏感区和生态脆弱带[3-4],青藏高原高寒草地生态系统对温度升高的响应更为敏感而迅速[5].高寒草甸作为青藏高原草地主要类型,草场面积约为7×105 km2,约占青藏高原可利用草地面积的50%,是中国畜牧业重要生产基地[6].气温上升将对高寒草甸生态系统产生重要影响,进而影响其载畜能力.因此,在气候变暖背景下,制定合理的高寒草甸载畜量是维系中国草地和畜牧业可持续发展的重要举措. ...
基于MODIS NPP数据的青海湖流域产草量与载畜量估算研究
2
2019
... 草地载畜量,是指在可持续发展的条件下,草地所能承载的最大牲畜数量[7].草地载畜量可分为数量载畜量和营养载畜量.数量载畜量由牧草产量决定,而营养载畜量由牧草产量和品质决定并和群落组成密切相关[8].温度升高在一定程度上增加了植物所需的热量,缓解了低温对高寒草甸的限制作用,植被生物量增加,可能导致草地数量载畜量增加[9-10].此外,增温也可能使土壤表面蒸发加强,水分散失,进而加剧干旱作用,造成植被生物量减少,使草地数量载畜量降低[11-12].温度升高可能会引起青藏高原牧草营养品质发生改变,导致牧草粗蛋白和粗脂肪含量降低,酸性洗涤纤维和中性洗涤纤维含量增加,牧草消化率降低,从而不利于反刍动物对牧草的消化利用,进而降低营养载畜量[13].可见,气候变暖可通过影响草地生物量和牧草品质,进而影响草地载畜量. ...
... 目前对高寒草地载畜量的安排主要依据牧草产量变化制定理论载畜量.王琪等[7]基于MODIS NPP产品数据估算青海湖流域的理论载畜量为8.11×105羊单位;王福成等[14]基于NDVI数据估算2019年玉树州高寒草甸理论载畜量为6.64×106羊单位;乔郭亮等[15]基于MODIS NPP产品数据估算2000—2018年天山中段高海拔草地暖季载畜量为2.43×105~3.19×105羊单位.尽管这些研究对于合理规划放牧生产活动具有重要的参考价值,但仅仅根据牧草产量制定草场放牧量缺乏科学决策的严谨性.若仅考虑牧草产量而忽略品质变化对载畜量的影响,在制定气候变暖后的草场载畜量时,可能会对放牧量产生负面影响,导致过度放牧和草场退化;或因实际放牧规模过小,导致牧民的收入受到不利影响.因此,本研究在青藏高原长江源北麓河流域选择典型高寒草甸进行模拟增温控制实验,探讨模拟增温对高寒草甸土壤温湿度、地上生物量、群落组成、牧草品质和载畜量的影响,以期为长江源区高寒草甸确定适宜的草场载畜量提供科学依据. ...
三江源区嵩草草地枯草期牧草营养价值评定及载畜量研究
5
2013
... 草地载畜量,是指在可持续发展的条件下,草地所能承载的最大牲畜数量[7].草地载畜量可分为数量载畜量和营养载畜量.数量载畜量由牧草产量决定,而营养载畜量由牧草产量和品质决定并和群落组成密切相关[8].温度升高在一定程度上增加了植物所需的热量,缓解了低温对高寒草甸的限制作用,植被生物量增加,可能导致草地数量载畜量增加[9-10].此外,增温也可能使土壤表面蒸发加强,水分散失,进而加剧干旱作用,造成植被生物量减少,使草地数量载畜量降低[11-12].温度升高可能会引起青藏高原牧草营养品质发生改变,导致牧草粗蛋白和粗脂肪含量降低,酸性洗涤纤维和中性洗涤纤维含量增加,牧草消化率降低,从而不利于反刍动物对牧草的消化利用,进而降低营养载畜量[13].可见,气候变暖可通过影响草地生物量和牧草品质,进而影响草地载畜量. ...
... 根据《天然草地合理载畜量的计算》(NY/T635—2002),每个标准羊单位按每天采食1.8 kg标准干草计算.根据NRC (National research council) 标准,标准羊单位需要粗蛋白0.0539 kg·d-1,代谢能8.38 MJ·d-1,暖季牲畜的牧草利用率为50%,牧草暖季和冷季粗蛋白的利用效率分别为62.25%和31.64%[19].根据研究区放牧实际,放牧天数生长季中前期为5—7月(92 d),生长季中后期为8—10月(92 d).其中代谢能需要量需通过牦牛饲喂试验得到,在本研究中对产气量进行计算时,主要基于三江源区牦牛产气量相关研究,取其平均值作为本研究中的产气量(表1).代谢能需要量=0.1456×24 h产气量+0.07675%×粗蛋白+0.1642%×粗脂肪+1.198(R= 0.97,n=139)[8]. ...
... 24 h in vitro gas production of natural pasture in grassland of Sanjiangyuan region
Table 1| 作者 | 年份 | 24 h产气量/mL | 地点 |
|---|
| 孙鹏飞等[19] | 2014 | 42.94 | 称多县 |
| 郝力壮等[8] | 2013 | 47.26 | 玉树县 |
| 张胜权[20] | 2013 | 51.07 | 玉树县 |
| 郝力壮等[21] | 2018 | 43.52 | 河南蒙古族自治县 |
| 本研究 | 2017, 2018 | 46.20 | 治多县 |
<strong>1.6</strong> 数据分析采用双因素方差法(Two-way ANOVA)分析增温和月份及其交互作用对群落和不同功能群地上生物量、酸性洗涤纤维、粗蛋白、粗脂肪和载畜量的影响,分析增温和土壤深度及其交互作用对土壤温度和土壤水分的影响.采用SPSS23.0软件进行方差分析,采用Origin 2021对图形绘制.图中误差线均为标准误. ...
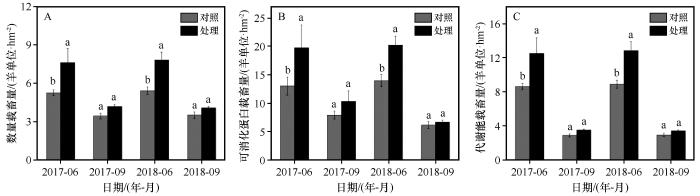
... 草地质量是由群落养分和产量所决定,主要包括牧草产量和牧草品质两个方面[29].草地质量的高低直接决定了草地的载畜能力.通常认为,当草地的实际放牧量超过理论载畜量时,就会造成过牧.在本研究中,模拟增温在生长季前期显著增加了高寒草甸草地数量载畜量,可消化蛋白载畜量和代谢能载畜量,在生长季后期无显著影响.这主要是因为增温显著增加了高寒草甸产草量(图2),虽然群落牧草粗蛋白和粗脂肪有所降低(图5),但产草量增加的促进作用大于品质降低的抑制作用,导致草地载畜量增加.青藏高原昼夜温差大、日照强的特点也促进了牧草中有机物的积累[18],因此生长季前期牧草中粗蛋白和能量含量较高.在生长季前期,模拟增温处理下可消化蛋白载畜量最高,代谢能载畜量次之,数量载畜量最低(图6).当营养充足的情况下,制定草场载畜量时,牧草产量则成为了首要考虑的因素.如按照营养载畜量安排放牧,则必然会导致过牧,草地发生退化.这与称多县高寒草甸典型嵩草草地研究结果一致[19].因此,在气温上升背景下,长江源区高寒草甸生长季前期以数量载畜量安排放牧为宜.在生长季后期,模拟增温处理下可消化蛋白载畜量最高,数量载畜量次之,代谢能载畜量最低(图6).牧草经过长时间生长发育,营养含量逐渐降低[8],牧草品质下降,同时牧草产量较低,因此代谢能载畜量低于数量载畜量.根据反刍家畜的消化生理特点(能氮平衡制约牲畜生长季后期对能量的利用)及生态学中营养容纳量和Liebig最小因子定律[8,35].在气温上升背景下,长江源区高寒草甸生长季后期应以代谢能载畜量安排放牧为宜. ...
... [8,35].在气温上升背景下,长江源区高寒草甸生长季后期应以代谢能载畜量安排放牧为宜. ...
Productivity and quality of alpine grassland vary with soil water availability under experimental warming
1
2018
... 草地载畜量,是指在可持续发展的条件下,草地所能承载的最大牲畜数量[7].草地载畜量可分为数量载畜量和营养载畜量.数量载畜量由牧草产量决定,而营养载畜量由牧草产量和品质决定并和群落组成密切相关[8].温度升高在一定程度上增加了植物所需的热量,缓解了低温对高寒草甸的限制作用,植被生物量增加,可能导致草地数量载畜量增加[9-10].此外,增温也可能使土壤表面蒸发加强,水分散失,进而加剧干旱作用,造成植被生物量减少,使草地数量载畜量降低[11-12].温度升高可能会引起青藏高原牧草营养品质发生改变,导致牧草粗蛋白和粗脂肪含量降低,酸性洗涤纤维和中性洗涤纤维含量增加,牧草消化率降低,从而不利于反刍动物对牧草的消化利用,进而降低营养载畜量[13].可见,气候变暖可通过影响草地生物量和牧草品质,进而影响草地载畜量. ...
Global assessment of experimental climate warming on tundra vegetation:heterogeneity over space and time
1
2012
... 草地载畜量,是指在可持续发展的条件下,草地所能承载的最大牲畜数量[7].草地载畜量可分为数量载畜量和营养载畜量.数量载畜量由牧草产量决定,而营养载畜量由牧草产量和品质决定并和群落组成密切相关[8].温度升高在一定程度上增加了植物所需的热量,缓解了低温对高寒草甸的限制作用,植被生物量增加,可能导致草地数量载畜量增加[9-10].此外,增温也可能使土壤表面蒸发加强,水分散失,进而加剧干旱作用,造成植被生物量减少,使草地数量载畜量降低[11-12].温度升高可能会引起青藏高原牧草营养品质发生改变,导致牧草粗蛋白和粗脂肪含量降低,酸性洗涤纤维和中性洗涤纤维含量增加,牧草消化率降低,从而不利于反刍动物对牧草的消化利用,进而降低营养载畜量[13].可见,气候变暖可通过影响草地生物量和牧草品质,进而影响草地载畜量. ...
Combined effects of climate warming and plant diversity loss on above-and below-ground grassland productivity
1
2007
... 草地载畜量,是指在可持续发展的条件下,草地所能承载的最大牲畜数量[7].草地载畜量可分为数量载畜量和营养载畜量.数量载畜量由牧草产量决定,而营养载畜量由牧草产量和品质决定并和群落组成密切相关[8].温度升高在一定程度上增加了植物所需的热量,缓解了低温对高寒草甸的限制作用,植被生物量增加,可能导致草地数量载畜量增加[9-10].此外,增温也可能使土壤表面蒸发加强,水分散失,进而加剧干旱作用,造成植被生物量减少,使草地数量载畜量降低[11-12].温度升高可能会引起青藏高原牧草营养品质发生改变,导致牧草粗蛋白和粗脂肪含量降低,酸性洗涤纤维和中性洗涤纤维含量增加,牧草消化率降低,从而不利于反刍动物对牧草的消化利用,进而降低营养载畜量[13].可见,气候变暖可通过影响草地生物量和牧草品质,进而影响草地载畜量. ...
Differential response of alpine steppe and alpine meadow to climate warming in the Central Qinghai-Tibetan Plateau
1
2016
... 草地载畜量,是指在可持续发展的条件下,草地所能承载的最大牲畜数量[7].草地载畜量可分为数量载畜量和营养载畜量.数量载畜量由牧草产量决定,而营养载畜量由牧草产量和品质决定并和群落组成密切相关[8].温度升高在一定程度上增加了植物所需的热量,缓解了低温对高寒草甸的限制作用,植被生物量增加,可能导致草地数量载畜量增加[9-10].此外,增温也可能使土壤表面蒸发加强,水分散失,进而加剧干旱作用,造成植被生物量减少,使草地数量载畜量降低[11-12].温度升高可能会引起青藏高原牧草营养品质发生改变,导致牧草粗蛋白和粗脂肪含量降低,酸性洗涤纤维和中性洗涤纤维含量增加,牧草消化率降低,从而不利于反刍动物对牧草的消化利用,进而降低营养载畜量[13].可见,气候变暖可通过影响草地生物量和牧草品质,进而影响草地载畜量. ...
高山植物对全球气候变暖的响应研究进展
1
2011
... 草地载畜量,是指在可持续发展的条件下,草地所能承载的最大牲畜数量[7].草地载畜量可分为数量载畜量和营养载畜量.数量载畜量由牧草产量决定,而营养载畜量由牧草产量和品质决定并和群落组成密切相关[8].温度升高在一定程度上增加了植物所需的热量,缓解了低温对高寒草甸的限制作用,植被生物量增加,可能导致草地数量载畜量增加[9-10].此外,增温也可能使土壤表面蒸发加强,水分散失,进而加剧干旱作用,造成植被生物量减少,使草地数量载畜量降低[11-12].温度升高可能会引起青藏高原牧草营养品质发生改变,导致牧草粗蛋白和粗脂肪含量降低,酸性洗涤纤维和中性洗涤纤维含量增加,牧草消化率降低,从而不利于反刍动物对牧草的消化利用,进而降低营养载畜量[13].可见,气候变暖可通过影响草地生物量和牧草品质,进而影响草地载畜量. ...
基于SPOT NDVI的2010-2018年青海省草地资源动态监测研究
1
2023
... 目前对高寒草地载畜量的安排主要依据牧草产量变化制定理论载畜量.王琪等[7]基于MODIS NPP产品数据估算青海湖流域的理论载畜量为8.11×105羊单位;王福成等[14]基于NDVI数据估算2019年玉树州高寒草甸理论载畜量为6.64×106羊单位;乔郭亮等[15]基于MODIS NPP产品数据估算2000—2018年天山中段高海拔草地暖季载畜量为2.43×105~3.19×105羊单位.尽管这些研究对于合理规划放牧生产活动具有重要的参考价值,但仅仅根据牧草产量制定草场放牧量缺乏科学决策的严谨性.若仅考虑牧草产量而忽略品质变化对载畜量的影响,在制定气候变暖后的草场载畜量时,可能会对放牧量产生负面影响,导致过度放牧和草场退化;或因实际放牧规模过小,导致牧民的收入受到不利影响.因此,本研究在青藏高原长江源北麓河流域选择典型高寒草甸进行模拟增温控制实验,探讨模拟增温对高寒草甸土壤温湿度、地上生物量、群落组成、牧草品质和载畜量的影响,以期为长江源区高寒草甸确定适宜的草场载畜量提供科学依据. ...
2000-2018年天山中段高海拔草地暖季承载力
1
2021
... 目前对高寒草地载畜量的安排主要依据牧草产量变化制定理论载畜量.王琪等[7]基于MODIS NPP产品数据估算青海湖流域的理论载畜量为8.11×105羊单位;王福成等[14]基于NDVI数据估算2019年玉树州高寒草甸理论载畜量为6.64×106羊单位;乔郭亮等[15]基于MODIS NPP产品数据估算2000—2018年天山中段高海拔草地暖季载畜量为2.43×105~3.19×105羊单位.尽管这些研究对于合理规划放牧生产活动具有重要的参考价值,但仅仅根据牧草产量制定草场放牧量缺乏科学决策的严谨性.若仅考虑牧草产量而忽略品质变化对载畜量的影响,在制定气候变暖后的草场载畜量时,可能会对放牧量产生负面影响,导致过度放牧和草场退化;或因实际放牧规模过小,导致牧民的收入受到不利影响.因此,本研究在青藏高原长江源北麓河流域选择典型高寒草甸进行模拟增温控制实验,探讨模拟增温对高寒草甸土壤温湿度、地上生物量、群落组成、牧草品质和载畜量的影响,以期为长江源区高寒草甸确定适宜的草场载畜量提供科学依据. ...
高寒草甸水热过程及其对草地退化和气候变暖的响应与反馈
2
2015
... 研究区位于青藏高原腹地长江源头一级支流北麓河流域内、风火山北麓、青藏高原北麓河冻土工程与环境综合观测研究站(简称北麓河试验站)附近(34°49′N、92°56′E,海拔4 600~4 700 m).研究区属于亚寒带半干旱气候,具有典型的内陆高原气候特征.全年仅有冷暖二季之别,年平均气温-3.8 ℃,年均降水量290 mm,超过90%的降水发生在生长季(5—9月),年平均空气相对湿度为57%[16].年均风速大,可达4.3 m·s-1,年均潜在蒸散量为1 316.9 mm,年日照时数2 600~3 000 h[16].研究区原生高寒草甸以莎草科植物为主,试验样地常见物种主要有高山嵩草(Kobresia pygmaea)、矮生嵩草(Kobresia humilis)、线叶嵩草(Kobresia capillifolia)、青藏薹草(Carex moorcroftii)、冷地早熟禾(Poa crymophila)、沙生风毛菊(Saussurea arenaria)、矮火绒草(Leontopodium nanum)等.植被高度5~10 cm. ...
... [16].研究区原生高寒草甸以莎草科植物为主,试验样地常见物种主要有高山嵩草(Kobresia pygmaea)、矮生嵩草(Kobresia humilis)、线叶嵩草(Kobresia capillifolia)、青藏薹草(Carex moorcroftii)、冷地早熟禾(Poa crymophila)、沙生风毛菊(Saussurea arenaria)、矮火绒草(Leontopodium nanum)等.植被高度5~10 cm. ...
1
2002
... 牧草品质主要测定了牧草中的粗蛋白、粗脂肪和酸性洗涤纤维含量.所有的分析过程均在兰州大学草地农业生态系统国家重点实验室完成.酸性洗涤纤维含量测定采用Van soest纤维分析法;粗脂肪含量测定采用索氏提取法(GB6433-94);粗蛋白含量测定采用凯氏微量定氮法(GB6432-94)[17]. ...
玛多县高山嵩草草地天然牧草营养评定与载畜量研究
2
2011
... 载畜量分为数量载畜量、可消化蛋白载畜量、代谢能载畜量[18]. ...
... 草地质量是由群落养分和产量所决定,主要包括牧草产量和牧草品质两个方面[29].草地质量的高低直接决定了草地的载畜能力.通常认为,当草地的实际放牧量超过理论载畜量时,就会造成过牧.在本研究中,模拟增温在生长季前期显著增加了高寒草甸草地数量载畜量,可消化蛋白载畜量和代谢能载畜量,在生长季后期无显著影响.这主要是因为增温显著增加了高寒草甸产草量(图2),虽然群落牧草粗蛋白和粗脂肪有所降低(图5),但产草量增加的促进作用大于品质降低的抑制作用,导致草地载畜量增加.青藏高原昼夜温差大、日照强的特点也促进了牧草中有机物的积累[18],因此生长季前期牧草中粗蛋白和能量含量较高.在生长季前期,模拟增温处理下可消化蛋白载畜量最高,代谢能载畜量次之,数量载畜量最低(图6).当营养充足的情况下,制定草场载畜量时,牧草产量则成为了首要考虑的因素.如按照营养载畜量安排放牧,则必然会导致过牧,草地发生退化.这与称多县高寒草甸典型嵩草草地研究结果一致[19].因此,在气温上升背景下,长江源区高寒草甸生长季前期以数量载畜量安排放牧为宜.在生长季后期,模拟增温处理下可消化蛋白载畜量最高,数量载畜量次之,代谢能载畜量最低(图6).牧草经过长时间生长发育,营养含量逐渐降低[8],牧草品质下降,同时牧草产量较低,因此代谢能载畜量低于数量载畜量.根据反刍家畜的消化生理特点(能氮平衡制约牲畜生长季后期对能量的利用)及生态学中营养容纳量和Liebig最小因子定律[8,35].在气温上升背景下,长江源区高寒草甸生长季后期应以代谢能载畜量安排放牧为宜. ...
三江源区不同季节放牧草场天然牧草营养价值评定及载畜量研究
3
2015
... 根据《天然草地合理载畜量的计算》(NY/T635—2002),每个标准羊单位按每天采食1.8 kg标准干草计算.根据NRC (National research council) 标准,标准羊单位需要粗蛋白0.0539 kg·d-1,代谢能8.38 MJ·d-1,暖季牲畜的牧草利用率为50%,牧草暖季和冷季粗蛋白的利用效率分别为62.25%和31.64%[19].根据研究区放牧实际,放牧天数生长季中前期为5—7月(92 d),生长季中后期为8—10月(92 d).其中代谢能需要量需通过牦牛饲喂试验得到,在本研究中对产气量进行计算时,主要基于三江源区牦牛产气量相关研究,取其平均值作为本研究中的产气量(表1).代谢能需要量=0.1456×24 h产气量+0.07675%×粗蛋白+0.1642%×粗脂肪+1.198(R= 0.97,n=139)[8]. ...
... 24 h in vitro gas production of natural pasture in grassland of Sanjiangyuan region
Table 1| 作者 | 年份 | 24 h产气量/mL | 地点 |
|---|
| 孙鹏飞等[19] | 2014 | 42.94 | 称多县 |
| 郝力壮等[8] | 2013 | 47.26 | 玉树县 |
| 张胜权[20] | 2013 | 51.07 | 玉树县 |
| 郝力壮等[21] | 2018 | 43.52 | 河南蒙古族自治县 |
| 本研究 | 2017, 2018 | 46.20 | 治多县 |
<strong>1.6</strong> 数据分析采用双因素方差法(Two-way ANOVA)分析增温和月份及其交互作用对群落和不同功能群地上生物量、酸性洗涤纤维、粗蛋白、粗脂肪和载畜量的影响,分析增温和土壤深度及其交互作用对土壤温度和土壤水分的影响.采用SPSS23.0软件进行方差分析,采用Origin 2021对图形绘制.图中误差线均为标准误. ...
... 草地质量是由群落养分和产量所决定,主要包括牧草产量和牧草品质两个方面[29].草地质量的高低直接决定了草地的载畜能力.通常认为,当草地的实际放牧量超过理论载畜量时,就会造成过牧.在本研究中,模拟增温在生长季前期显著增加了高寒草甸草地数量载畜量,可消化蛋白载畜量和代谢能载畜量,在生长季后期无显著影响.这主要是因为增温显著增加了高寒草甸产草量(图2),虽然群落牧草粗蛋白和粗脂肪有所降低(图5),但产草量增加的促进作用大于品质降低的抑制作用,导致草地载畜量增加.青藏高原昼夜温差大、日照强的特点也促进了牧草中有机物的积累[18],因此生长季前期牧草中粗蛋白和能量含量较高.在生长季前期,模拟增温处理下可消化蛋白载畜量最高,代谢能载畜量次之,数量载畜量最低(图6).当营养充足的情况下,制定草场载畜量时,牧草产量则成为了首要考虑的因素.如按照营养载畜量安排放牧,则必然会导致过牧,草地发生退化.这与称多县高寒草甸典型嵩草草地研究结果一致[19].因此,在气温上升背景下,长江源区高寒草甸生长季前期以数量载畜量安排放牧为宜.在生长季后期,模拟增温处理下可消化蛋白载畜量最高,数量载畜量次之,代谢能载畜量最低(图6).牧草经过长时间生长发育,营养含量逐渐降低[8],牧草品质下降,同时牧草产量较低,因此代谢能载畜量低于数量载畜量.根据反刍家畜的消化生理特点(能氮平衡制约牲畜生长季后期对能量的利用)及生态学中营养容纳量和Liebig最小因子定律[8,35].在气温上升背景下,长江源区高寒草甸生长季后期应以代谢能载畜量安排放牧为宜. ...
三江源地区草甸草场13种典型牧草营养价值评定研究
1
2013
... 24 h in vitro gas production of natural pasture in grassland of Sanjiangyuan region
Table 1| 作者 | 年份 | 24 h产气量/mL | 地点 |
|---|
| 孙鹏飞等[19] | 2014 | 42.94 | 称多县 |
| 郝力壮等[8] | 2013 | 47.26 | 玉树县 |
| 张胜权[20] | 2013 | 51.07 | 玉树县 |
| 郝力壮等[21] | 2018 | 43.52 | 河南蒙古族自治县 |
| 本研究 | 2017, 2018 | 46.20 | 治多县 |
<strong>1.6</strong> 数据分析采用双因素方差法(Two-way ANOVA)分析增温和月份及其交互作用对群落和不同功能群地上生物量、酸性洗涤纤维、粗蛋白、粗脂肪和载畜量的影响,分析增温和土壤深度及其交互作用对土壤温度和土壤水分的影响.采用SPSS23.0软件进行方差分析,采用Origin 2021对图形绘制.图中误差线均为标准误. ...
三江源区河南县天然草场草畜营养平衡关系研究
1
2018
... 24 h in vitro gas production of natural pasture in grassland of Sanjiangyuan region
Table 1| 作者 | 年份 | 24 h产气量/mL | 地点 |
|---|
| 孙鹏飞等[19] | 2014 | 42.94 | 称多县 |
| 郝力壮等[8] | 2013 | 47.26 | 玉树县 |
| 张胜权[20] | 2013 | 51.07 | 玉树县 |
| 郝力壮等[21] | 2018 | 43.52 | 河南蒙古族自治县 |
| 本研究 | 2017, 2018 | 46.20 | 治多县 |
<strong>1.6</strong> 数据分析采用双因素方差法(Two-way ANOVA)分析增温和月份及其交互作用对群落和不同功能群地上生物量、酸性洗涤纤维、粗蛋白、粗脂肪和载畜量的影响,分析增温和土壤深度及其交互作用对土壤温度和土壤水分的影响.采用SPSS23.0软件进行方差分析,采用Origin 2021对图形绘制.图中误差线均为标准误. ...
Phenological changes offset the warming effects on biomass production in an alpine meadow on the Qinghai-Tibetan Plateau
1
2021
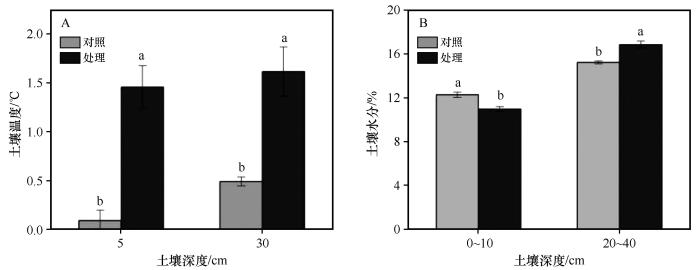
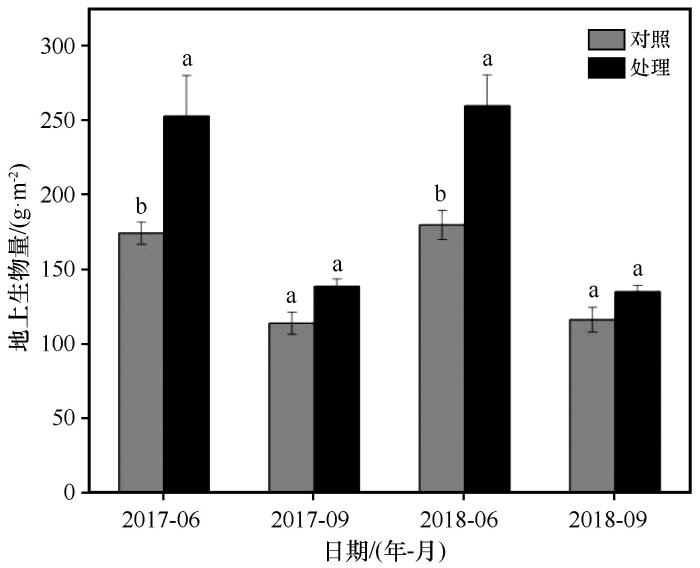
... 植物的生理活动和生化反应,都必须在一定的温度条件下才能进行.温度可引起环境中其他因子如湿度、降水、风、水中氧的溶解度等的变化,而环境诸因子的综合作用,又能影响植物的生长发育[22].在高寒草甸生态系统中,低温和较短的生长季通常被认为是影响植物生长最重要的限制因素[23].温度升高在一定程度上满足了植物生长发育对热量的需求,有利于高寒草甸地上生物量的增加[24].本研究中,模拟增温在生长季前期(6月)显著增加了地上生物量,但在生长季后期(9月)无显著影响.这可能是因为在生长前期,植物的生长较快,在这一时期,植物对热量的需求较大,增温满足了植物生长对热量的需求[24],导致地上生物量显著增加,这与在风火山地区的研究结果一致[25].而在生长季后期,植物生长速率放缓,植物的生长周期即将完成,此时对热量的需求较小[24],导致地上生物量无显著变化.此外,在先前的研究中发现,增温增强了冻土活性层融化过程,冻土活动层在春季提前解冻[26],可能导致植物生长季提前,进而使地上生物量增加.多年冻土上覆活动层在春季提前解冻也可能是导致增温后深层土壤水分增加的重要因素. ...
Global patterns of terrestrial nitrogen and phosphorus limitation
1
2020
... 植物的生理活动和生化反应,都必须在一定的温度条件下才能进行.温度可引起环境中其他因子如湿度、降水、风、水中氧的溶解度等的变化,而环境诸因子的综合作用,又能影响植物的生长发育[22].在高寒草甸生态系统中,低温和较短的生长季通常被认为是影响植物生长最重要的限制因素[23].温度升高在一定程度上满足了植物生长发育对热量的需求,有利于高寒草甸地上生物量的增加[24].本研究中,模拟增温在生长季前期(6月)显著增加了地上生物量,但在生长季后期(9月)无显著影响.这可能是因为在生长前期,植物的生长较快,在这一时期,植物对热量的需求较大,增温满足了植物生长对热量的需求[24],导致地上生物量显著增加,这与在风火山地区的研究结果一致[25].而在生长季后期,植物生长速率放缓,植物的生长周期即将完成,此时对热量的需求较小[24],导致地上生物量无显著变化.此外,在先前的研究中发现,增温增强了冻土活性层融化过程,冻土活动层在春季提前解冻[26],可能导致植物生长季提前,进而使地上生物量增加.多年冻土上覆活动层在春季提前解冻也可能是导致增温后深层土壤水分增加的重要因素. ...
Winter and spring warming result in delayed spring phenology on the Tibetan Plateau
3
2010
... 植物的生理活动和生化反应,都必须在一定的温度条件下才能进行.温度可引起环境中其他因子如湿度、降水、风、水中氧的溶解度等的变化,而环境诸因子的综合作用,又能影响植物的生长发育[22].在高寒草甸生态系统中,低温和较短的生长季通常被认为是影响植物生长最重要的限制因素[23].温度升高在一定程度上满足了植物生长发育对热量的需求,有利于高寒草甸地上生物量的增加[24].本研究中,模拟增温在生长季前期(6月)显著增加了地上生物量,但在生长季后期(9月)无显著影响.这可能是因为在生长前期,植物的生长较快,在这一时期,植物对热量的需求较大,增温满足了植物生长对热量的需求[24],导致地上生物量显著增加,这与在风火山地区的研究结果一致[25].而在生长季后期,植物生长速率放缓,植物的生长周期即将完成,此时对热量的需求较小[24],导致地上生物量无显著变化.此外,在先前的研究中发现,增温增强了冻土活性层融化过程,冻土活动层在春季提前解冻[26],可能导致植物生长季提前,进而使地上生物量增加.多年冻土上覆活动层在春季提前解冻也可能是导致增温后深层土壤水分增加的重要因素. ...
... [24],导致地上生物量显著增加,这与在风火山地区的研究结果一致[25].而在生长季后期,植物生长速率放缓,植物的生长周期即将完成,此时对热量的需求较小[24],导致地上生物量无显著变化.此外,在先前的研究中发现,增温增强了冻土活性层融化过程,冻土活动层在春季提前解冻[26],可能导致植物生长季提前,进而使地上生物量增加.多年冻土上覆活动层在春季提前解冻也可能是导致增温后深层土壤水分增加的重要因素. ...
... [24],导致地上生物量无显著变化.此外,在先前的研究中发现,增温增强了冻土活性层融化过程,冻土活动层在春季提前解冻[26],可能导致植物生长季提前,进而使地上生物量增加.多年冻土上覆活动层在春季提前解冻也可能是导致增温后深层土壤水分增加的重要因素. ...
Plant production,and carbon and nitrogen source pools,are strongly intensified by experimental warming in alpine ecosystems in the Qinghai-Tibet Plateau
2
2011
... 植物的生理活动和生化反应,都必须在一定的温度条件下才能进行.温度可引起环境中其他因子如湿度、降水、风、水中氧的溶解度等的变化,而环境诸因子的综合作用,又能影响植物的生长发育[22].在高寒草甸生态系统中,低温和较短的生长季通常被认为是影响植物生长最重要的限制因素[23].温度升高在一定程度上满足了植物生长发育对热量的需求,有利于高寒草甸地上生物量的增加[24].本研究中,模拟增温在生长季前期(6月)显著增加了地上生物量,但在生长季后期(9月)无显著影响.这可能是因为在生长前期,植物的生长较快,在这一时期,植物对热量的需求较大,增温满足了植物生长对热量的需求[24],导致地上生物量显著增加,这与在风火山地区的研究结果一致[25].而在生长季后期,植物生长速率放缓,植物的生长周期即将完成,此时对热量的需求较小[24],导致地上生物量无显著变化.此外,在先前的研究中发现,增温增强了冻土活性层融化过程,冻土活动层在春季提前解冻[26],可能导致植物生长季提前,进而使地上生物量增加.多年冻土上覆活动层在春季提前解冻也可能是导致增温后深层土壤水分增加的重要因素. ...
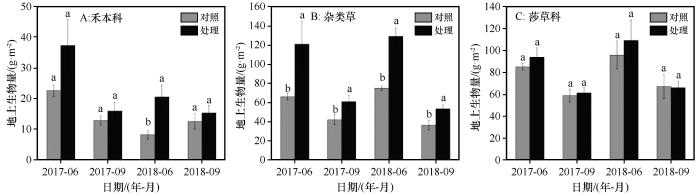
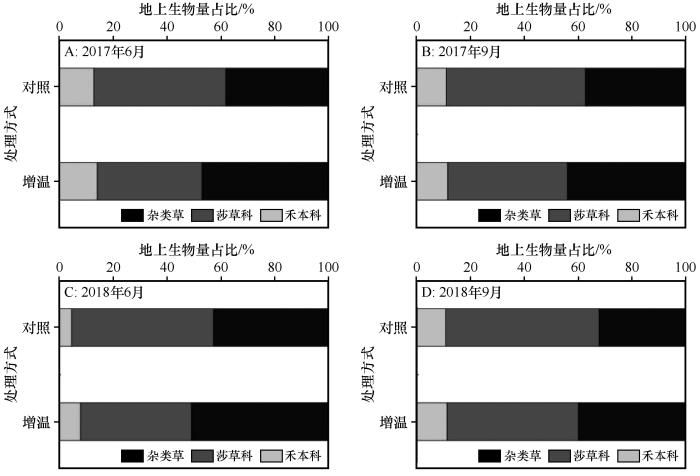
... 在全球变暖背景下,相对于植物群落而言,不同物种对温度升高的敏感性各异,总有一些物种比另一些物种对温度升高的响应更为敏感,引起群落优势种和群落组成改变[25].在本研究中,增温后,尽管莎草科植物地上生物量无显著变化(图3),但其群落占比降低(图4);杂类草和禾本科植物地上生物量和占群落比重增加,这与在海北高寒草甸[27]的研究结果一致.莎草科植物为浅根系植物,其根系主要分布在0~10 cm层,且呈须根系,以莎草科植物为建群种的高寒草甸通常以降低其生物量在群落中的比重来适应增温和水分胁迫的环境[28].杂类草和禾本科植物主要是深根系,其根系相对莎草科植物较深[29].增温后表层土壤温度显著增加(图1),加剧了土壤水分蒸发,表层逐渐干旱化,但深层土壤水分增加.杂类草和禾本科植物可利用表层和深层土壤水分和养分以满足自身的生长.此外,气候变暖可能引起土壤有机质含量增加,同时加速土壤有机质的分解速率和无机营养成分释放速率[30].和莎草科植物相比,杂类草植物对养分获取能力强且更加灵活,如其可通过根系分泌低分子量有机酸活化土壤钙磷[31],从而满足其对磷的需求,这也反映了杂类草在面对气候变暖的环境下具有更强的竞争能力. ...
Influence of experimental warming on heat and water fluxes of alpine meadows in the Qinghai-Tibet Plateau
1
2014
... 植物的生理活动和生化反应,都必须在一定的温度条件下才能进行.温度可引起环境中其他因子如湿度、降水、风、水中氧的溶解度等的变化,而环境诸因子的综合作用,又能影响植物的生长发育[22].在高寒草甸生态系统中,低温和较短的生长季通常被认为是影响植物生长最重要的限制因素[23].温度升高在一定程度上满足了植物生长发育对热量的需求,有利于高寒草甸地上生物量的增加[24].本研究中,模拟增温在生长季前期(6月)显著增加了地上生物量,但在生长季后期(9月)无显著影响.这可能是因为在生长前期,植物的生长较快,在这一时期,植物对热量的需求较大,增温满足了植物生长对热量的需求[24],导致地上生物量显著增加,这与在风火山地区的研究结果一致[25].而在生长季后期,植物生长速率放缓,植物的生长周期即将完成,此时对热量的需求较小[24],导致地上生物量无显著变化.此外,在先前的研究中发现,增温增强了冻土活性层融化过程,冻土活动层在春季提前解冻[26],可能导致植物生长季提前,进而使地上生物量增加.多年冻土上覆活动层在春季提前解冻也可能是导致增温后深层土壤水分增加的重要因素. ...
Warming changed the relationship between species diversity and primary productivity of alpine meadow on the Tibetan Plateau
1
2022
... 在全球变暖背景下,相对于植物群落而言,不同物种对温度升高的敏感性各异,总有一些物种比另一些物种对温度升高的响应更为敏感,引起群落优势种和群落组成改变[25].在本研究中,增温后,尽管莎草科植物地上生物量无显著变化(图3),但其群落占比降低(图4);杂类草和禾本科植物地上生物量和占群落比重增加,这与在海北高寒草甸[27]的研究结果一致.莎草科植物为浅根系植物,其根系主要分布在0~10 cm层,且呈须根系,以莎草科植物为建群种的高寒草甸通常以降低其生物量在群落中的比重来适应增温和水分胁迫的环境[28].杂类草和禾本科植物主要是深根系,其根系相对莎草科植物较深[29].增温后表层土壤温度显著增加(图1),加剧了土壤水分蒸发,表层逐渐干旱化,但深层土壤水分增加.杂类草和禾本科植物可利用表层和深层土壤水分和养分以满足自身的生长.此外,气候变暖可能引起土壤有机质含量增加,同时加速土壤有机质的分解速率和无机营养成分释放速率[30].和莎草科植物相比,杂类草植物对养分获取能力强且更加灵活,如其可通过根系分泌低分子量有机酸活化土壤钙磷[31],从而满足其对磷的需求,这也反映了杂类草在面对气候变暖的环境下具有更强的竞争能力. ...
Grassland vegetation changes and nocturnal global warming
1
1999
... 在全球变暖背景下,相对于植物群落而言,不同物种对温度升高的敏感性各异,总有一些物种比另一些物种对温度升高的响应更为敏感,引起群落优势种和群落组成改变[25].在本研究中,增温后,尽管莎草科植物地上生物量无显著变化(图3),但其群落占比降低(图4);杂类草和禾本科植物地上生物量和占群落比重增加,这与在海北高寒草甸[27]的研究结果一致.莎草科植物为浅根系植物,其根系主要分布在0~10 cm层,且呈须根系,以莎草科植物为建群种的高寒草甸通常以降低其生物量在群落中的比重来适应增温和水分胁迫的环境[28].杂类草和禾本科植物主要是深根系,其根系相对莎草科植物较深[29].增温后表层土壤温度显著增加(图1),加剧了土壤水分蒸发,表层逐渐干旱化,但深层土壤水分增加.杂类草和禾本科植物可利用表层和深层土壤水分和养分以满足自身的生长.此外,气候变暖可能引起土壤有机质含量增加,同时加速土壤有机质的分解速率和无机营养成分释放速率[30].和莎草科植物相比,杂类草植物对养分获取能力强且更加灵活,如其可通过根系分泌低分子量有机酸活化土壤钙磷[31],从而满足其对磷的需求,这也反映了杂类草在面对气候变暖的环境下具有更强的竞争能力. ...
Experimentally simulating warmer and wetter climate additively improves rangeland quality on the Tibetan Plateau
2
2018
... 在全球变暖背景下,相对于植物群落而言,不同物种对温度升高的敏感性各异,总有一些物种比另一些物种对温度升高的响应更为敏感,引起群落优势种和群落组成改变[25].在本研究中,增温后,尽管莎草科植物地上生物量无显著变化(图3),但其群落占比降低(图4);杂类草和禾本科植物地上生物量和占群落比重增加,这与在海北高寒草甸[27]的研究结果一致.莎草科植物为浅根系植物,其根系主要分布在0~10 cm层,且呈须根系,以莎草科植物为建群种的高寒草甸通常以降低其生物量在群落中的比重来适应增温和水分胁迫的环境[28].杂类草和禾本科植物主要是深根系,其根系相对莎草科植物较深[29].增温后表层土壤温度显著增加(图1),加剧了土壤水分蒸发,表层逐渐干旱化,但深层土壤水分增加.杂类草和禾本科植物可利用表层和深层土壤水分和养分以满足自身的生长.此外,气候变暖可能引起土壤有机质含量增加,同时加速土壤有机质的分解速率和无机营养成分释放速率[30].和莎草科植物相比,杂类草植物对养分获取能力强且更加灵活,如其可通过根系分泌低分子量有机酸活化土壤钙磷[31],从而满足其对磷的需求,这也反映了杂类草在面对气候变暖的环境下具有更强的竞争能力. ...
... 草地质量是由群落养分和产量所决定,主要包括牧草产量和牧草品质两个方面[29].草地质量的高低直接决定了草地的载畜能力.通常认为,当草地的实际放牧量超过理论载畜量时,就会造成过牧.在本研究中,模拟增温在生长季前期显著增加了高寒草甸草地数量载畜量,可消化蛋白载畜量和代谢能载畜量,在生长季后期无显著影响.这主要是因为增温显著增加了高寒草甸产草量(图2),虽然群落牧草粗蛋白和粗脂肪有所降低(图5),但产草量增加的促进作用大于品质降低的抑制作用,导致草地载畜量增加.青藏高原昼夜温差大、日照强的特点也促进了牧草中有机物的积累[18],因此生长季前期牧草中粗蛋白和能量含量较高.在生长季前期,模拟增温处理下可消化蛋白载畜量最高,代谢能载畜量次之,数量载畜量最低(图6).当营养充足的情况下,制定草场载畜量时,牧草产量则成为了首要考虑的因素.如按照营养载畜量安排放牧,则必然会导致过牧,草地发生退化.这与称多县高寒草甸典型嵩草草地研究结果一致[19].因此,在气温上升背景下,长江源区高寒草甸生长季前期以数量载畜量安排放牧为宜.在生长季后期,模拟增温处理下可消化蛋白载畜量最高,数量载畜量次之,代谢能载畜量最低(图6).牧草经过长时间生长发育,营养含量逐渐降低[8],牧草品质下降,同时牧草产量较低,因此代谢能载畜量低于数量载畜量.根据反刍家畜的消化生理特点(能氮平衡制约牲畜生长季后期对能量的利用)及生态学中营养容纳量和Liebig最小因子定律[8,35].在气温上升背景下,长江源区高寒草甸生长季后期应以代谢能载畜量安排放牧为宜. ...
Warming and grazing enhance litter decomposition and nutrient release independent of litter quality in an alpine meadow
1
2022
... 在全球变暖背景下,相对于植物群落而言,不同物种对温度升高的敏感性各异,总有一些物种比另一些物种对温度升高的响应更为敏感,引起群落优势种和群落组成改变[25].在本研究中,增温后,尽管莎草科植物地上生物量无显著变化(图3),但其群落占比降低(图4);杂类草和禾本科植物地上生物量和占群落比重增加,这与在海北高寒草甸[27]的研究结果一致.莎草科植物为浅根系植物,其根系主要分布在0~10 cm层,且呈须根系,以莎草科植物为建群种的高寒草甸通常以降低其生物量在群落中的比重来适应增温和水分胁迫的环境[28].杂类草和禾本科植物主要是深根系,其根系相对莎草科植物较深[29].增温后表层土壤温度显著增加(图1),加剧了土壤水分蒸发,表层逐渐干旱化,但深层土壤水分增加.杂类草和禾本科植物可利用表层和深层土壤水分和养分以满足自身的生长.此外,气候变暖可能引起土壤有机质含量增加,同时加速土壤有机质的分解速率和无机营养成分释放速率[30].和莎草科植物相比,杂类草植物对养分获取能力强且更加灵活,如其可通过根系分泌低分子量有机酸活化土壤钙磷[31],从而满足其对磷的需求,这也反映了杂类草在面对气候变暖的环境下具有更强的竞争能力. ...
Mobilization of soil phosphate after 8 years of warming is linked to plant phosphorus-acquisition strategies in an alpine meadow on the Qinghai-Tibetan Plateau
1
2021
... 在全球变暖背景下,相对于植物群落而言,不同物种对温度升高的敏感性各异,总有一些物种比另一些物种对温度升高的响应更为敏感,引起群落优势种和群落组成改变[25].在本研究中,增温后,尽管莎草科植物地上生物量无显著变化(图3),但其群落占比降低(图4);杂类草和禾本科植物地上生物量和占群落比重增加,这与在海北高寒草甸[27]的研究结果一致.莎草科植物为浅根系植物,其根系主要分布在0~10 cm层,且呈须根系,以莎草科植物为建群种的高寒草甸通常以降低其生物量在群落中的比重来适应增温和水分胁迫的环境[28].杂类草和禾本科植物主要是深根系,其根系相对莎草科植物较深[29].增温后表层土壤温度显著增加(图1),加剧了土壤水分蒸发,表层逐渐干旱化,但深层土壤水分增加.杂类草和禾本科植物可利用表层和深层土壤水分和养分以满足自身的生长.此外,气候变暖可能引起土壤有机质含量增加,同时加速土壤有机质的分解速率和无机营养成分释放速率[30].和莎草科植物相比,杂类草植物对养分获取能力强且更加灵活,如其可通过根系分泌低分子量有机酸活化土壤钙磷[31],从而满足其对磷的需求,这也反映了杂类草在面对气候变暖的环境下具有更强的竞争能力. ...
Effect of long-term experimental warming on the nutritional quality of alpine meadows in the northern Tibet
1
2020
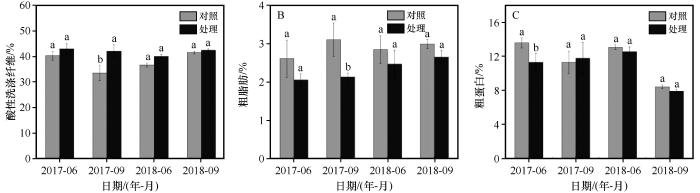
... 从家畜营养角度出发,牧草中粗蛋白和粗脂肪含量越高,品质越好;而纤维含量越高,品质越差[32].牧草品质在一定程度上也决定了畜产品的产量和品质,其可通过影响牲畜对牧草的利用效率、能量摄入以及养分获取,进而影响草地的载畜能力[33].增温后,植物群落粗蛋白和粗脂肪含量有所降低,酸性洗涤纤维含量有所增加(图5),这与在海北站矮嵩草草甸[34]的研究结果一致.这意味着增温后牧草品质降低,不利于反刍动物对牧草的消化利用,这可能和杂类草和禾本科植物粗蛋白和粗脂肪含量降低有关(表4).当产草量增加时,可能会对营养元素产生“稀释”效应,导致牧草中粗蛋白和粗脂肪含量下降,粗纤维含量增加[33].先前研究也表明牧草粗蛋白和粗脂肪含量和土壤温度呈负相关,而酸性洗涤纤维含量和土壤温度呈正相关[33].本研究中土壤温度增加可能是导致植物中粗蛋白含量降低、酸性洗涤纤维含量增加的重要因素. ...
中国草地的产草量和牧草品质:格局及其与环境因子之间的关系
3
2013
... 从家畜营养角度出发,牧草中粗蛋白和粗脂肪含量越高,品质越好;而纤维含量越高,品质越差[32].牧草品质在一定程度上也决定了畜产品的产量和品质,其可通过影响牲畜对牧草的利用效率、能量摄入以及养分获取,进而影响草地的载畜能力[33].增温后,植物群落粗蛋白和粗脂肪含量有所降低,酸性洗涤纤维含量有所增加(图5),这与在海北站矮嵩草草甸[34]的研究结果一致.这意味着增温后牧草品质降低,不利于反刍动物对牧草的消化利用,这可能和杂类草和禾本科植物粗蛋白和粗脂肪含量降低有关(表4).当产草量增加时,可能会对营养元素产生“稀释”效应,导致牧草中粗蛋白和粗脂肪含量下降,粗纤维含量增加[33].先前研究也表明牧草粗蛋白和粗脂肪含量和土壤温度呈负相关,而酸性洗涤纤维含量和土壤温度呈正相关[33].本研究中土壤温度增加可能是导致植物中粗蛋白含量降低、酸性洗涤纤维含量增加的重要因素. ...
... [33].先前研究也表明牧草粗蛋白和粗脂肪含量和土壤温度呈负相关,而酸性洗涤纤维含量和土壤温度呈正相关[33].本研究中土壤温度增加可能是导致植物中粗蛋白含量降低、酸性洗涤纤维含量增加的重要因素. ...
... [33].本研究中土壤温度增加可能是导致植物中粗蛋白含量降低、酸性洗涤纤维含量增加的重要因素. ...
高寒草甸植物对长期增温的响应
1
2017
... 从家畜营养角度出发,牧草中粗蛋白和粗脂肪含量越高,品质越好;而纤维含量越高,品质越差[32].牧草品质在一定程度上也决定了畜产品的产量和品质,其可通过影响牲畜对牧草的利用效率、能量摄入以及养分获取,进而影响草地的载畜能力[33].增温后,植物群落粗蛋白和粗脂肪含量有所降低,酸性洗涤纤维含量有所增加(图5),这与在海北站矮嵩草草甸[34]的研究结果一致.这意味着增温后牧草品质降低,不利于反刍动物对牧草的消化利用,这可能和杂类草和禾本科植物粗蛋白和粗脂肪含量降低有关(表4).当产草量增加时,可能会对营养元素产生“稀释”效应,导致牧草中粗蛋白和粗脂肪含量下降,粗纤维含量增加[33].先前研究也表明牧草粗蛋白和粗脂肪含量和土壤温度呈负相关,而酸性洗涤纤维含量和土壤温度呈正相关[33].本研究中土壤温度增加可能是导致植物中粗蛋白含量降低、酸性洗涤纤维含量增加的重要因素. ...
Estimates of habitat carrying capacity incorporating explicit nutritional constraints
1
1985
... 草地质量是由群落养分和产量所决定,主要包括牧草产量和牧草品质两个方面[29].草地质量的高低直接决定了草地的载畜能力.通常认为,当草地的实际放牧量超过理论载畜量时,就会造成过牧.在本研究中,模拟增温在生长季前期显著增加了高寒草甸草地数量载畜量,可消化蛋白载畜量和代谢能载畜量,在生长季后期无显著影响.这主要是因为增温显著增加了高寒草甸产草量(图2),虽然群落牧草粗蛋白和粗脂肪有所降低(图5),但产草量增加的促进作用大于品质降低的抑制作用,导致草地载畜量增加.青藏高原昼夜温差大、日照强的特点也促进了牧草中有机物的积累[18],因此生长季前期牧草中粗蛋白和能量含量较高.在生长季前期,模拟增温处理下可消化蛋白载畜量最高,代谢能载畜量次之,数量载畜量最低(图6).当营养充足的情况下,制定草场载畜量时,牧草产量则成为了首要考虑的因素.如按照营养载畜量安排放牧,则必然会导致过牧,草地发生退化.这与称多县高寒草甸典型嵩草草地研究结果一致[19].因此,在气温上升背景下,长江源区高寒草甸生长季前期以数量载畜量安排放牧为宜.在生长季后期,模拟增温处理下可消化蛋白载畜量最高,数量载畜量次之,代谢能载畜量最低(图6).牧草经过长时间生长发育,营养含量逐渐降低[8],牧草品质下降,同时牧草产量较低,因此代谢能载畜量低于数量载畜量.根据反刍家畜的消化生理特点(能氮平衡制约牲畜生长季后期对能量的利用)及生态学中营养容纳量和Liebig最小因子定律[8,35].在气温上升背景下,长江源区高寒草甸生长季后期应以代谢能载畜量安排放牧为宜. ...




 甘公网安备 62010202000688号
甘公网安备 62010202000688号
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}