腾格里沙漠东南缘猪毛菜( Salsola collina
楼科尔 , 1 , 曲文杰 1 , 王磊 1 , 王兴 1 , 郜永贵 2 , 杨新国 , 1
1.宁夏大学 西北退化生态系统恢复与重建教育部重点实验室/西北土地退化与生态恢复国家重点实验室培育基地/生态环境学院,宁夏 银川 750021
2.中国铁路兰州局集团有限公司 中卫工务段中卫固沙林场,宁夏 中卫 755000
Comparison of root architecture characteristics of Salsola collina in the southeast edge of Tengger Desert
Lou Keer , 1 , Qu Wenjie 1 , Wang Lei 1 , Wang Xing 1 , Gao Yonggui 2 , Yang Xinguo , 1
1.MOE Key Laboratory of Northwest Degraded Ecosystem Restoration and Reconstruction / Cultivation Base of State Key Laboratory of Northwest Land Degradation and Ecological Restoration / College of Ecological Environment,Ningxia University,Yinchuan 750021,China
2.Zhongwei Gusha Forest Farm,Zhongwei Public Works Section of China Railway Lanzhou Bureau Group Co. ,Ltd. ,Zhongwei 755000,Ningxia,China
通讯作者: 杨新国(E-mail: xinguoyang1976@163.com )
收稿日期: 2023-11-27
修回日期: 2024-01-05
基金资助:
国家自然科学基金项目 . 32060313 宁夏重点研发计划课题 . 2021BEG03008
Received: 2023-11-27
Revised: 2024-01-05
作者简介 About authors
楼科尔(1996—),男,浙江绍兴人,硕士研究生,主要研究方向为旱区草本植物构型E-mail:keerlou@163.com
, E-mail:keerlou@163.com
摘要
根系构型反映植物对资源的吸收能力和利用情况,可揭示沙地植被更新生长与适应机制。本研究以腾格里沙漠东南缘不同地表条件下[流沙地,藻结皮地(半固定沙地),藓结皮地(固定沙地)]的猪毛菜(Salsola collina )为研究对象,采用传统挖掘法收集植物根系,基于量化的根系形态指标,利用几何拓扑学及分形理论分析了根系构型特征,探讨了地表演变对猪毛菜根系构型特征的影响及其适应机制。结果表明:从流沙地、藻结皮地到藓结皮地,猪毛菜根深宽比、根冠比、比根长和比表面积逐次显著降低,根系水平幅则显著增加;根系拓扑指数TI 分别为0.8125(流沙地)、0.7107(藻结皮地)、0.6313(藓结皮地),根系结构趋向于从鱼尾形分支结构转变为叉状分支结构;根系分形维数显著提高,但根系分形丰度显著降低。总之,伴随沙地固定程度的增加以及地表结皮覆盖条件的演变,猪毛菜根系生长投入下降,空间拓展能力退化,同时根系的整体结构进一步复杂化,根系生长对策从数量型向质量型转变,资源利用对策向高竞争力方向转变。本研究可为荒漠地区植被恢复和物种选择、配置提供参考依据。
关键词:
腾格里沙漠 猪毛菜(Salsola collina ) 根系构型 拓扑指数 分形维数
Abstract
Root architecture reflects the absorptive capacity and utilization of plant resources, and is an important aspect to reveal the mechanism of vegetation regeneration, growth and adaptation in sandy land. In this study, Salsola collina under different surface conditions (quicksand, algal crusts (semi fixed sand), moss crusts (fixed sand)) in the southeast edge of Tengger Desert was selected as the research object. The traditional mining method was used to collect plant roots. Based on the quantitative root morphology index, the geometric topology and fractal theory were used to analyze the root configuration characteristics, The effects of land surface evolution on the root architecture of Salsola collina and its adaptation mechanism were discussed. The results showed that the ratio of root depth to width, the ratio of root to shoot, the specific root length and the specific surface area of Salsola collina decreased significantly from quicksand, algae crusts to moss crusts, while the horizontal range of root system increased significantly; The root topological index TI was 0.8125 (quicksand), 0.7107 (algal crusts), 0.6313 (moss crusts), which decreased gradually, and the root structure tended to change from fishtail branching structure to fork branching structure; The fractal dimension of roots increased significantly, but the fractal abundance of roots decreased significantly. In a word, with the increase of sand fixation degree and the evolution of surface crusting and covering conditions, the investment in root growth of Salsola collina decreased, and the spatial expansion ability degenerated. At the same time, the overall structure of the root system was further complicated. The root growth strategy changed from quantity to quality, and the resource utilization strategy changed to high competitiveness. This study can provide reference for vegetation restoration, species selection and allocation in desert areas.
Keywords:
Tengger Desert Salsola collina root architecture topological index fractal dimension
本文引用格式
楼科尔, 曲文杰, 王磊, 王兴, 郜永贵, 杨新国. 腾格里沙漠东南缘猪毛菜( Salsola collina . 中国沙漠 [J], 2024, 44(4): 193-201 doi:10.7522/j.issn.1000-694X.2024.00013
Lou Keer, Qu Wenjie, Wang Lei, Wang Xing, Gao Yonggui, Yang Xinguo. Comparison of root architecture characteristics of Salsola collina in the southeast edge of Tengger Desert . Journal of Desert Research
0 引言
根系是土壤和植物的动态界面。植物依靠根系获得固定和支撑,依赖根系吸收土壤中的矿质营养和水分,并输送到地上部分供其利用[1 ] 。根系构型指植物根系在土壤中的分布和排列[2 ] 。根系构型受遗传因素控制,生境也会影响根系构型的表达[3 ] 。根系构型一定程度上反映了植物对环境的适应性及植被的退化和演替状况[4 ] 。Lynch[2 ] 认为根系构型包括根系拓扑结构、根系形态、根系空间分布和根系构筑等内容。Fitter等[5 ] 提出了根系拓扑结构的两种极端类型,即鱼尾形分支和叉状分支类型。以往的研究较多通过外形、根冠比、根重等指标反映根系生长情况,而对根系的分支和结构特征研究较少[6 -7 ] 。20世纪70年代,分形几何的诞生为根系构型研究提供了新方法[8 ] 。根系有明显的分形结构[9 ] ,运用分形理论研究根系,可以定量描述根系几何形态、分形特征及扩展范围等[10 ] 。
在干旱的荒漠地区,通过封育保护,流动沙地可以逐步演变为半固定和固定沙地。在这个过程中,一个重要的伴随演替特征就是沙面细土层的形成和生物结皮的出现[11 ] 。按照出现的早晚,生物结皮主要分为藻结皮、地衣结皮和藓结皮[12 ] 。生物结皮的出现显著改变了沙地地表覆盖条件,进而通过对土壤水文过程和种子扩散条件的改变,往往成为影响植被更新与根系生长的关键因素[13 ] 。首先,生物结皮演替过程中土壤养分的逐渐增加为维管植物的存活和生长提供了宝贵的养分条件,直接影响着维管植物种子的散布、萌发和定居[14 ] 。生物结皮发育也会导致表层土壤阻水能力增加,从而减少降水的入渗量[15 -16 ] ,加剧土壤干旱。在这个过程中,沙地植物可通过根系形态结构的改变来适应资源环境条件的变化[8 ] 。通过对物种在不同沙地地表覆盖条件下根系构型的比较研究,有助于了解沙地演变以及植被更新演替的内生机制和趋势。
一年生草本植物猪毛菜(Salsola collina ),在荒漠区和极端干旱区广泛分布。猪毛菜既能防风固沙,也是饲用植物,在干旱地区退化土地水土保持和植被恢复方面具有很大的应用潜力[17 -18 ] 。本文以腾格里沙漠东南缘的野生猪毛菜为对象,研究其在不同地表条件下的根系构型特征,比较了不同地表条件对猪毛菜根系生长以及构型的影响,揭示了猪毛菜对沙地环境演变的适应机制。
1 材料和方法
1.1 研究区概况
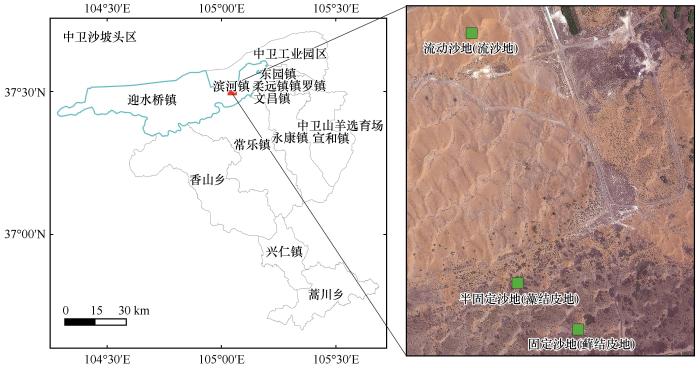
试验地点位于宁夏回族自治区中卫市,腾格里沙漠东南缘的沙坡头地区(37°27′N、104°05′E,海拔1 310~1 350 m,图1 ),该地区是荒漠和荒漠草原的过渡地带,最低气温-25.1 ℃,最高气温38.1 ℃,年均气温约9.5 ℃,年降水量约200 mm,降水主要在5—9月;全年日照约3 260 h,年蒸发量约2 400 mm,土壤稳定含水量约2.5%[19 ] 。灌木主要为柠条锦鸡儿(Caragana korshinskii )、花棒(Corethrodendron scoparium ),草本植物主要为小画眉草(Eragrostis minor )、狗尾草(Setaria viridis )、猪毛菜(Salsola collina ),雾冰藜(Grubovia dasyphylla )、虫实(Corispermum hyssopifolium )、砂蓝刺头(Echinops gmelinii )。
图1
图1
样点位置
Fig.1
Sample plots
1.2 试验方法
1.2.1 试验地的选择
选择坡向一致,坡度较缓且植物分布相对较为均匀的流动沙地、半固定沙地与固定沙地进行试验研究,每块试验地大小为20 m×20 m。其中流动沙地为格状沙丘,无结皮覆盖,植被分布稀疏,0~100 cm土壤水分含量为7.31%;半固定沙地的优势结皮为藻结皮,结皮覆盖不连续,覆盖度60%~65%,固沙年限20年左右,0~100 cm土壤水分含量为5.96%;固定沙地的优势结皮为藓结皮,结皮发育充分,覆盖度80%~100%,固沙年限60年左右,0~100 cm土壤水分含量为5.84%(表1 )。
1.2.2 样品采集与处理
选取的植株为当年生长的野生猪毛菜,对猪毛菜的地上部分指标(株高、地径、冠幅)进行调查,统计确定标准样株(表2 )。参照标准样株,在每个试验地内选择猪毛菜各10株,共选取30株植物,研究供试植株地上部带回实验室烘干称重。利用铁锹和毛刷等工具从植物根基部挖掘取样,直至无根系出现。如根系断裂,标记好位置并将根系放入自封袋中[20 ] 。取样过程中,仔细观察根系在原位的整体构型,记录根系的内部、外部连接数量,并测量根系水平幅和根系垂直幅,最后将样品放入冰盒(内径规格:长29 cm,宽23 cm,高21.5 cm)中带回实验室。清除根系表面杂质后进行图像扫描,之后设置烘箱温度为105 ℃,将根系和植株地上部分杀青120 min后,再设置烘箱温度75 ℃下烘干48 h,最后用电子天平(0.0001 g)称重,得到根系生物量和地上生物量。使用Epson Scan 2扫描仪对猪毛菜根系进行扫描,获得图像,采用根系分析系统软件Win-RHIZO PRO 2020进行分析,分析指标包含根长(RL)、根表面积(RSA)、根系分布图上边长为r 的正方形和根系所截的正方形数目Nr 。
1.3 指标测定
1.3.1 根系形态参数的测定
根据根系垂直幅、根系水平幅、根系生物量、地上生物量计算根深宽比和根冠比。
根深宽比=根系垂直幅/根系水平幅(1)
根冠比=根系生物量/地上生物量(2)
式中:根系垂直幅与根系水平幅单位为cm;生物量单位为g。
根据根长和根表面积以及根系生物量计算根系比根长(SRL)、比表面积(SRA)。
比根长=根长/生物量(3)
比表面积=表面积/生物量(4)
式中:比根长单位为cm·g-1 ;根长单位为cm;表面积单位为cm2 ;比表面积单位为cm2 ·g-1 。
1.3.2 根系拓扑参数的计算
根系拓扑结构存在两种极端模式[21 -24 ] 。根据模型计算拓扑指数TI ,拓扑指数表达式为
TI =lgA/ lgM (5)
式中:TI 为拓扑指数;A 为最长通道(内部分支最多的通道)连接数;M 为外部连接数。TI 的值为0.5~1,越接近0.5表明根系的外部分支较多,根系结构趋向于叉状分支;越接近1表明根系的外部分支较少,根系结构趋向于鱼尾形分支。
1.3.3 根系分形参数的计算
根据Win-RHIZO测定的根系分布图上边长为r 的正方形和根系所截的正方形数目Nr ,采用盒维数法[25 ] 得到根系分形特征,随着正方形边r 逐渐减小,根系所截Nr 逐渐增大,得到不同的r 上相应Nr 值后,分别以lgr 、lgNr 为横坐标和纵坐标作图,回归直线方程为lgNr =-FD lgr +lgK ,回归直线斜率的负数是所求的分形维数,lgK 为分形丰度。
1.4 数据处理
原始数据用Excel 2019进行整理,用SPSS 24.0软件进行分析,用Origin Pro 2023软件作图。显著性水平设定为α =0.05,对数据进行单因素方差分析,并利用最小显著差异法(LSD)检验不同数据组间的差异显著性。
2 结果与分析
2.1 根系水平幅、垂直幅和根深宽比
3种地表条件下猪毛菜根系水平幅和根深宽比差异显著(图2 )。藓结皮地猪毛菜根系水平幅显著大于藻结皮(P <0.05),藻结皮地猪毛菜根系水平幅显著大于流沙(P <0.05),根系水平幅4.13~10.25 cm。流沙地的根深宽比显著大于藻结皮(P <0.05),藻结皮地的深宽比显著大于流沙(P <0.05),根深宽比呈逐渐下降趋势,其比值均大于1,即植物根系水平幅小于根系垂直幅。从流沙地、藻结皮地到藓结皮地,猪毛菜根系垂直幅呈下降趋势,根系水平幅10.89~12.36 cm。
图2
图2
3种地表条件下猪毛菜根系分布范围和根深宽比
注: 不同小写字母表示差异显著(P <0.05)
Fig.2
Changes in root distribution and root depth width ratio of Salsola collina under three surface conditions
2.2 地上生物量、根系生物量和根冠比
3种地表条件下猪毛菜根系生物量和根冠比差异明显(图3 )。相对于根系生物量,地表条件对根冠比影响更明显。流沙地根冠比显著大于藻结皮地(P <0.05);藻结皮地根冠比显著大于藓结皮地(P <0.05)。藓结皮地根系生物量显著小于流沙结皮地(P <0.05),与藻结皮地无显著差异(P >0.05)。在地上生物量中,3种地表条件的差异不显著(P >0.05),但从流沙地、藻结皮地到藓结皮地,地上生物量呈现增大的趋势。
图3
图3
3种地表条件下猪毛菜地上生物量、根系生物量和根冠比
注: 不同小写字母表示差异显著(P <0.05)
Fig.3
Effects of three site conditions on aboveground biomass, root biomass and root shoot ratio of Salsola collina
2.3 比根长、比表面积
3种地表条件下比根长、比根表面积存在显著差异(表3 )。流沙地比根长最长,比藻结皮地、藓结皮地分别显著高了55%、179%;藻结皮地比根长显著高于藓结皮地(P <0.05),表明地表条件对于猪毛菜的根长度影响呈线性关系。流沙地与藻结皮地的比表面积都显著高于藓结皮地(P <0.05),且流沙地的比表面积最大,显著高于藻结皮地(P <0.05),表明猪毛菜的比根长和比表面积有相同的变化趋势,表现为流沙地>藻结皮地>藓结皮地。
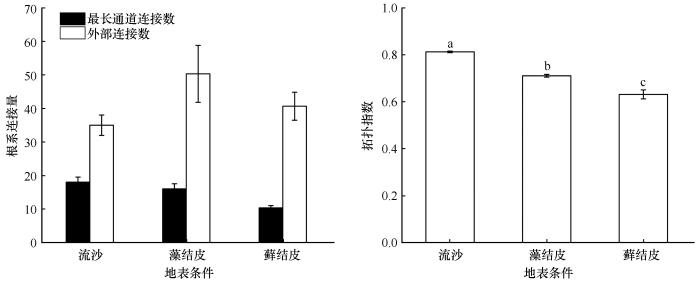
2.4 根系拓扑参数
3种地表条件下猪毛菜的根系拓扑参数差异显著(图4 )。流沙地的拓扑指数TI =0.8125,显著大于藻结皮地和藓结皮地,表明根系结构倾向于鱼尾形分支结构,根系分支结构较简单,主根明显且分支较少。藓结皮地的拓扑指数最小,TI =0.6313,显著小于流沙地和藻结皮地(P <0.05),表明根系结构相对流沙地更加复杂化,倾向于叉状分枝结构。藻结皮地的拓扑指数TI =0.7107,表明根系结构介于鱼尾形分支结构和叉状分支结构之间。
图4
图4
3种地表条件下猪毛菜根系数拓扑参数
注: 不同小写字母表示差异显著(P <0.05)
Fig.4
Changes in topological parameters of Salsola collina under three surface conditions
2.5 根系分形特征
3种地表条件下猪毛菜的根系分形维数与根系分形丰度存在明显差异(图5 )。藓结皮地的根系分形维数最大,显著大于藻结皮地和流沙地(P <0.05),而藻结皮地和流沙地的根系分形维数无明显差异(P >0.05),但从流沙地、藻结皮地到藓结皮地,根系分形维数呈现增大的趋势,表明猪毛菜的根系结构复杂化。一般植物根系的分形维数为1.0~1.9,当分形维数>1.1时,就可判定研究对象具有明显的分形特征[26 ] 。猪毛菜在不同的地表条件下根系分形维数都大于1.1,表明猪毛菜根系具有很好的分形特征。流沙地的根系分形丰度最大,比藻结皮地和藓结皮地分别大了13%和34%。藓结皮地的根系分形丰度最小,显著小于流沙地和藻结皮地。从流沙地、藻结皮地到藓结皮地,根系分形丰度呈现减小的趋势,表明猪毛菜根系拓展能力下降,在土壤中拓展的体积逐渐减少。
图5
图5
3种地表条件下猪毛菜根系分形参数
注: 不同小写字母表示差异显著(P <0.05)
Fig.5
Changes of fractal characteristics of Salsola collina roots under three surface conditions
3 讨论
3.1 根系空间分布特征
植物根系的空间分布特征受遗传因素和环境因素的共同影响。植物根系在环境作用下会发生一定的空间分布变化,表现出高度的可塑性[27 -29 ] 。生物结皮不仅能显著提高表层土壤有机质和土壤养分的含量,也可显著提高碳、氮、磷等元素相关的土壤酶活性[30 -32 ] 。同时,生物结皮的存在阻碍土壤水分入渗,具体表现为与裸地相比,降低初始入渗率、平均入渗率、稳定入渗率及非饱和导水率[33 ] 。本研究发现,从流沙地、藻结皮地到藓结皮地,猪毛菜的根系水平幅显著增加,而根系垂直幅逐渐降低,且地上部分和地下部分生物量变化不大。这可能是由于地表演变导致结皮中养分和水分的增加,使得根系分支横向生长获取表层资源,根系分支生长浅层化,同时表层有限的资源使猪毛菜首先进行地下部分的发育,地上部分变化不显著,具体表现为根系结构的复杂化以充分利用资源,并且根冠比的显著差异表明猪毛菜在生境改善的过程中减小了对根系生长的投入,根系结构的转变有可能会出现生物量下降的趋势,这与苏纪帅等[34 ] 对封育条件下的针茅研究结果相符。在根系形态方面,可能由于物种的遗传因素,根系垂直幅大于水平幅,因此深宽比>1。作为干旱荒漠区广泛分布的植物种,这种对生境的适应以及遗传的表现,是猪毛菜应对荒漠地区水分和养分胁迫的重要策略。
比根长和比表面积是根系形态结构的重要指标,决定了根系吸收水分和养分的能力,其值与养分吸收效率成正比[35 ] 。本研究发现,从流沙地、藻结皮地到藓结皮地,猪毛菜的比根长和比表面积不断减小,这可能是由于其根系水平幅的增加,根系分支浅层化,总体根长减少。同时由于根系响应生物结皮层中的养分和水分增加,减少了对根系获取沙地深处水分的资源投入,调节根的直径和根的数量来减少了比根长和比表面积。这与单立山等[36 ] 对珍珠猪毛菜的研究结果大致相符,说明在生境资源逐渐丰富的条件下,猪毛菜会调节其根系结构,同时减少比根长和比表面积来适应。
3.2 根系拓扑结构特征
在植物根系中,拓扑结构与营养物质的竞争力之间存在着密切的关系。鱼尾形分支根系通常结构简单,分布范围较大,从而增加了对养分的获取能力。相比之下,叉状分支则呈现出相反的特点。在等量的碳投入条件下,鱼尾形分支结构更有利于营养空间的占用。因此,鱼尾形分支的根系结构在干旱和贫瘠的环境中表现出更高的适应性[21 -24 ] 。本研究发现,从流沙地、藻结皮地到藓结皮地,猪毛菜的根系结构趋向于从鱼尾形分支转变成叉状分支。表明猪毛菜在养分和水分条件不足的生境中,根系结构具有分支简单、次级分支数量较少、但分布较深的特点,这种根系结构增加了获取营养物质的能力。然而随着地表生境的改善,养分和水分在地表方向集中,使得根系分支浅层化,趋向于叉状分支结构,这种根系结构增加了对有限空间内营养物质的利用能力。这表明猪毛菜的根系拓扑结构与生境中的养分存在着动态平衡,优化自身结构以得到更好的生长。
3.3 根系分形维数和分形丰度
分形维数反映了根系分支复杂程度,是植物根系发育程度和适应胁迫环境策略的重要指标 [9 ] ,分形丰度反映的是根系在生长介质中拓展程度,可定量化描述根系对空间占有能力及吸收水肥的效率[27 ] 。本研究发现流沙地猪毛菜(FD =1.2823)、藻结皮地猪毛菜(FD =1.313)和藓结皮地猪毛菜(FD =1.397)分形维数总体较小且逐渐增加。这表明在干旱胁迫条件下,猪毛菜的根系结构较简单。然而随着养分和水分条件的改善,根系增加了根系分支以及分支长度,从而提高了对养分的利用效率。根系分形丰度反映了根系的延展范围、拓展能力及对水分和养分的吸收效率 [37 ] 。本研究发现,流沙地猪毛菜(FA =3.5282)、藻结皮地猪毛菜(FA =3.1123)和藓结皮地猪毛菜(FA =2.6361)均维持较大的分形丰度。流沙地猪毛菜根系分形丰度最高,表明其根系体积大,吸收效率和空间占有能力得到进一步的提高。猪毛菜分形维数和分形丰度的特点,综合反映了其对不同地表生境的适应策略:流沙地中,由于资源比较分散并且分布较深,猪毛菜根系采取高生长量策略,空间拓展能力增加,因此具有更高的分形丰度;而在生物结皮地中,由于资源分布相对集中,并且植物之间存在根际竞争,根系采取结构优化策略,通过根系结构复杂化,提高了对局部资源利用的充分性,因此具有更大的分形维数,而藻结皮地和藓结皮地由于结皮覆盖度以及结皮组分不同,进而引起养分和水分的差异,分形维数和分形丰度出现显著变化。
4 结论
通过对腾格里沙漠东南缘不同沙地地表覆盖条件下猪毛菜根系的形态特征、拓扑结构和分形特征的比较研究,本文进一步丰富并细化了猪毛菜根系构型响应的指标体系,初步揭示了猪毛菜对沙地生境变化的适应策略。
从流沙地、藻结皮地到藓结皮地,猪毛菜根深宽比、根冠比、比根长和比表面积逐次显著降低,根系水平幅则显著增加。
猪毛菜根系拓扑结构存在显著区别。流沙地趋向鱼尾形分支结构,藓结皮地趋向于叉状分支结构,藻结皮地则介于二者之间。
猪毛菜根系分形特征存在显著差异,其中流沙地的根系分形维数最低,根系分形丰度最高。藓结皮地则相反。
总体而言,伴随沙地固定程度的增加以及地表结皮覆盖条件的演变,猪毛菜根系生长投入下降,空间拓展能力退化,同时根系的整体结构进一步复杂化,根系生长对策从数量型向质量型转变,资源利用对策向高竞争力方向转变。
参考文献
View Option
[1]
梁泉 ,廖红 ,严小龙 植物根构型的定量分析
[J].植物学通报 ,2007 (6 ):695 -702 .
[本文引用: 1]
[2]
Lynch J Root architecture and plant productivity
[J].Plant Physiology ,1995 ,109 (1 ):7 -13 .
[本文引用: 2]
[3]
单立山 ,李毅 ,张荣 ,等 降雨格局变化对白刺幼苗根系形态特征的影响
[J].生态学报 ,2017 ,37 (21 ):7324 -7332 .
[本文引用: 1]
[4]
孙栋元 ,赵成义 ,王丽娟 ,等 荒漠植物构型研究进展
[J].水土保持研究 ,2011 ,18 (5 ):281 -287 .
[本文引用: 1]
[5]
Fitter A H Stickland T R Harvey M L et al Architectural analysis of plant root systems 1.Architectural correlates of exploitation efficiency
[J].New Phytologist ,1991 ,118 (3 ):375 -382 .
[本文引用: 1]
[6]
Glimskär A Estimates of root system topology of five plant species grown at steady-state nutrition
[J].Plant and Soil ,2000 ,227 (1/2 ):249 -256 .
[本文引用: 1]
[7]
朱广龙 ,陈许斌 ,郭小倩 ,等 酸枣根系结构可塑性对自然梯度干旱生境的适应机制
[J].生态学报 ,2018 ,38 (16 ):5810 -5818 .
[本文引用: 1]
[8]
马雄忠 ,王新平 阿拉善高原2种荒漠植物根系构型及生态适应性特征
[J].生态学报 ,2020 ,40 (17 ):6001 -6008 .
[本文引用: 2]
[9]
杨培岭 ,罗远培 冬小麦根系形态的分形特征
[J].科学通报 ,1994 (20 ):1911 -1913 .
[本文引用: 2]
[10]
王义琴 ,张慧娟 ,白克智 ,等 分形几何在植物根系研究中的应用
[J].自然杂志 ,1999 (3 ):143 -146 .
[本文引用: 1]
[11]
段争虎 ,刘新民 ,屈建军 沙坡头地区土壤结皮形成机理的研究
[J].干旱区研究 ,1996 (2 ):31 -36 .
[本文引用: 1]
[12]
Lange O L Kidron E L Büdel B et al Taxonomic composition and photosynthetic characteristics of the ‘biological soil crusts' covering sand dunes in the western Negev Desert
[J].Functional Ecology ,1992 ,6 (5 ):519 -527 .
[本文引用: 1]
[13]
张元明 ,王雪芹 荒漠地表生物土壤结皮形成与演替特征概述
[J].生态学报 ,2010 ,30 (16 ):4484 -4492 .
[本文引用: 1]
[14]
Belnap J Gardner J S Soil microstructure in soils of the Colorado Plateau:the role of the cyanobacterium Microcoleus vaginatus
[J].Great Basin Naturist ,1993 ,53 :40 -47 .
[本文引用: 1]
[15]
付广军 ,廖超英 ,孙长忠 毛乌素沙地土壤结皮对水分运动的影响
[J].西北林学院学报 ,2010 ,25 (1 ):7 -10 .
[本文引用: 1]
[16]
熊好琴 ,段金跃 ,王妍 ,等 毛乌素沙地生物结皮对水分入渗和再分配的影响
[J].水土保持研究 ,2011 ,18 (4 ):82 -87 .
[本文引用: 1]
[17]
白文娟 水蚀风蚀交错带植被恢复对土壤质量的影响与植物生理生态适应性
[J].北京:中国科学院研究生院 ,2010 .
[本文引用: 1]
[18]
谭炯锐 ,查同刚 ,张泽 ,等 猪毛菜响应干旱胁迫的叶片结构、生理及转录组分析
[J].草业学报 ,2024 ,33 (1 ):75 -88 .
[本文引用: 1]
[19]
李新荣 ,肖洪浪 ,刘立超 ,等 腾格里沙漠沙坡头地区固沙植被对生物多样性恢复的长期影响
[J].中国沙漠 ,2005 ,25 (2 ):31 -39 .
[本文引用: 1]
[20]
伍红燕 ,赵倩 ,宋桂龙 ,等 平地与边坡条件下野生胡枝子根构型特征研究
[J].草业科学 ,2019 ,36 (7 ):1725 -1733 .
[本文引用: 1]
[21]
Fitter A H The topology and geometry of plant root systems:influence of watering rate on root system topology in Trifolium pratense
[J].Annals of Botany ,1986 ,58 (1 ):91 -101 .
[本文引用: 2]
[22]
Fitter A H An architectural approach to the comparative ecology of plant root systems
[J].New Phytologist ,1987 ,106 (1 ):61 -77 .
[23]
Fitter A H Stickland T R Architectural analysis of plant root systems:III.studies on plants under field conditions
[J].New Phytologist ,1992 ,121 (2 ):243 -248 .
[24]
Bouma T J Nielsen K L Hal J V et al Root system topology and diameter distribution of species from habitats differing in inundation frequency
[J].Functional Ecology ,2001 ,15 (3 ):360 -369 .
[本文引用: 2]
[25]
李雪萍 ,赵成章 ,任悦 ,等 尕海湿地不同密度条件下垂穗披碱草根系分形结构
[J].生态学报 ,2018 ,38 (4 ):1176 -1182 .
[本文引用: 1]
[26]
王浩 砒砂岩区不同生境条件下沙棘根系构型特征研究
[D].西安 :西北大学 ,2018 .
[本文引用: 1]
[27]
单立山 ,李毅 ,任伟 ,等 河西走廊中部两种荒漠植物根系构型特征
[J].应用生态学报 ,2013 ,24 (1 ):25 -31 .
[本文引用: 2]
[28]
赵艳云 ,陆昭华 ,夏江宝 ,等 黄河三角洲贝壳堤岛3种优势灌木的根系构型
[J].生态学报 ,2015 ,35 (6 ):1688 -1695 .
[29]
Meyer K M Ward D Wiegand K et al Multi-proxy evidence for competition between savanna woody species
[J].Perspectives in Plant Ecology Evolution and Systematics ,2008 ,10 (1 ):63 -72 .
[本文引用: 1]
[30]
Zhang B C Zhou X B Zhang Y M Responses of microbial activities and soil physical-chemical properties to the successional process of biological soil crusts in the Gurbantunggut Desert,Xinjiang
[J].Journal of Arid Land ,2015 ,7 (1 ):101 -109 .
[本文引用: 1]
[31]
张健 ,徐明 ,皱晓 ,等 不同土壤和植被生境下生物结皮对土壤性质的影响
[J].水土保持学报 ,2019 ,33 (5 ):323 -328 .
[32]
Yang H Y Liu C Z Liu Y M et al Impact of human trampling on biological soil crusts determined by soil microbial biomass,enzyme activities and nematode communities in a desert ecosystem
[J].European Journal of Soil Biology ,2018 ,87 :61 -71 .
[本文引用: 1]
[33]
李雨晨 ,平原 ,澹腾辉 ,等 红壤丘陵区不同盖度生物结皮对水分入渗的影响
[J].水土保持学报 ,2023 ,37 (5 ):71 -77 .
[本文引用: 1]
[34]
苏纪帅 ,赵洁 ,井光花 ,等 半干旱草地长期封育进程中针茅植物根系格局变化特征
[J].生态学报 ,2017 ,37 (19 ):6571 -6580 .
[本文引用: 1]
[35]
Bauhus J Khanna P K Menden N Aboveground and belowground interactions in mixed plantations of Eucalyptus globulus and Acacia mearnsii
[J].Canadian Journal of Forest Research ,2000 ,30 (12 ):1886 -1894 .
[本文引用: 1]
[36]
单立山 ,苏铭 ,张正中 ,等 不同生境下荒漠植物红砂-珍珠猪毛菜混生根系的垂直分布规律
[J].植物生态学报 ,2018 ,42 (4 ):475 -486 .
[本文引用: 1]
[37]
杨小林 ,张希明 ,李义玲 ,等 塔克拉玛干沙漠腹地几种植物根系分形特征
[J].干旱区地理 ,2009 ,32 (2 ):249 -254 .
[本文引用: 1]
植物根构型的定量分析
1
2007
... 根系是土壤和植物的动态界面.植物依靠根系获得固定和支撑,依赖根系吸收土壤中的矿质营养和水分,并输送到地上部分供其利用[1 ] .根系构型指植物根系在土壤中的分布和排列[2 ] .根系构型受遗传因素控制,生境也会影响根系构型的表达[3 ] .根系构型一定程度上反映了植物对环境的适应性及植被的退化和演替状况[4 ] .Lynch[2 ] 认为根系构型包括根系拓扑结构、根系形态、根系空间分布和根系构筑等内容.Fitter等[5 ] 提出了根系拓扑结构的两种极端类型,即鱼尾形分支和叉状分支类型.以往的研究较多通过外形、根冠比、根重等指标反映根系生长情况,而对根系的分支和结构特征研究较少[6 -7 ] .20世纪70年代,分形几何的诞生为根系构型研究提供了新方法[8 ] .根系有明显的分形结构[9 ] ,运用分形理论研究根系,可以定量描述根系几何形态、分形特征及扩展范围等[10 ] . ...
Root architecture and plant productivity
2
1995
... 根系是土壤和植物的动态界面.植物依靠根系获得固定和支撑,依赖根系吸收土壤中的矿质营养和水分,并输送到地上部分供其利用[1 ] .根系构型指植物根系在土壤中的分布和排列[2 ] .根系构型受遗传因素控制,生境也会影响根系构型的表达[3 ] .根系构型一定程度上反映了植物对环境的适应性及植被的退化和演替状况[4 ] .Lynch[2 ] 认为根系构型包括根系拓扑结构、根系形态、根系空间分布和根系构筑等内容.Fitter等[5 ] 提出了根系拓扑结构的两种极端类型,即鱼尾形分支和叉状分支类型.以往的研究较多通过外形、根冠比、根重等指标反映根系生长情况,而对根系的分支和结构特征研究较少[6 -7 ] .20世纪70年代,分形几何的诞生为根系构型研究提供了新方法[8 ] .根系有明显的分形结构[9 ] ,运用分形理论研究根系,可以定量描述根系几何形态、分形特征及扩展范围等[10 ] . ...
... [2 ]认为根系构型包括根系拓扑结构、根系形态、根系空间分布和根系构筑等内容.Fitter等[5 ] 提出了根系拓扑结构的两种极端类型,即鱼尾形分支和叉状分支类型.以往的研究较多通过外形、根冠比、根重等指标反映根系生长情况,而对根系的分支和结构特征研究较少[6 -7 ] .20世纪70年代,分形几何的诞生为根系构型研究提供了新方法[8 ] .根系有明显的分形结构[9 ] ,运用分形理论研究根系,可以定量描述根系几何形态、分形特征及扩展范围等[10 ] . ...
降雨格局变化对白刺幼苗根系形态特征的影响
1
2017
... 根系是土壤和植物的动态界面.植物依靠根系获得固定和支撑,依赖根系吸收土壤中的矿质营养和水分,并输送到地上部分供其利用[1 ] .根系构型指植物根系在土壤中的分布和排列[2 ] .根系构型受遗传因素控制,生境也会影响根系构型的表达[3 ] .根系构型一定程度上反映了植物对环境的适应性及植被的退化和演替状况[4 ] .Lynch[2 ] 认为根系构型包括根系拓扑结构、根系形态、根系空间分布和根系构筑等内容.Fitter等[5 ] 提出了根系拓扑结构的两种极端类型,即鱼尾形分支和叉状分支类型.以往的研究较多通过外形、根冠比、根重等指标反映根系生长情况,而对根系的分支和结构特征研究较少[6 -7 ] .20世纪70年代,分形几何的诞生为根系构型研究提供了新方法[8 ] .根系有明显的分形结构[9 ] ,运用分形理论研究根系,可以定量描述根系几何形态、分形特征及扩展范围等[10 ] . ...
荒漠植物构型研究进展
1
2011
... 根系是土壤和植物的动态界面.植物依靠根系获得固定和支撑,依赖根系吸收土壤中的矿质营养和水分,并输送到地上部分供其利用[1 ] .根系构型指植物根系在土壤中的分布和排列[2 ] .根系构型受遗传因素控制,生境也会影响根系构型的表达[3 ] .根系构型一定程度上反映了植物对环境的适应性及植被的退化和演替状况[4 ] .Lynch[2 ] 认为根系构型包括根系拓扑结构、根系形态、根系空间分布和根系构筑等内容.Fitter等[5 ] 提出了根系拓扑结构的两种极端类型,即鱼尾形分支和叉状分支类型.以往的研究较多通过外形、根冠比、根重等指标反映根系生长情况,而对根系的分支和结构特征研究较少[6 -7 ] .20世纪70年代,分形几何的诞生为根系构型研究提供了新方法[8 ] .根系有明显的分形结构[9 ] ,运用分形理论研究根系,可以定量描述根系几何形态、分形特征及扩展范围等[10 ] . ...
Architectural analysis of plant root systems 1.Architectural correlates of exploitation efficiency
1
1991
... 根系是土壤和植物的动态界面.植物依靠根系获得固定和支撑,依赖根系吸收土壤中的矿质营养和水分,并输送到地上部分供其利用[1 ] .根系构型指植物根系在土壤中的分布和排列[2 ] .根系构型受遗传因素控制,生境也会影响根系构型的表达[3 ] .根系构型一定程度上反映了植物对环境的适应性及植被的退化和演替状况[4 ] .Lynch[2 ] 认为根系构型包括根系拓扑结构、根系形态、根系空间分布和根系构筑等内容.Fitter等[5 ] 提出了根系拓扑结构的两种极端类型,即鱼尾形分支和叉状分支类型.以往的研究较多通过外形、根冠比、根重等指标反映根系生长情况,而对根系的分支和结构特征研究较少[6 -7 ] .20世纪70年代,分形几何的诞生为根系构型研究提供了新方法[8 ] .根系有明显的分形结构[9 ] ,运用分形理论研究根系,可以定量描述根系几何形态、分形特征及扩展范围等[10 ] . ...
Estimates of root system topology of five plant species grown at steady-state nutrition
1
2000
... 根系是土壤和植物的动态界面.植物依靠根系获得固定和支撑,依赖根系吸收土壤中的矿质营养和水分,并输送到地上部分供其利用[1 ] .根系构型指植物根系在土壤中的分布和排列[2 ] .根系构型受遗传因素控制,生境也会影响根系构型的表达[3 ] .根系构型一定程度上反映了植物对环境的适应性及植被的退化和演替状况[4 ] .Lynch[2 ] 认为根系构型包括根系拓扑结构、根系形态、根系空间分布和根系构筑等内容.Fitter等[5 ] 提出了根系拓扑结构的两种极端类型,即鱼尾形分支和叉状分支类型.以往的研究较多通过外形、根冠比、根重等指标反映根系生长情况,而对根系的分支和结构特征研究较少[6 -7 ] .20世纪70年代,分形几何的诞生为根系构型研究提供了新方法[8 ] .根系有明显的分形结构[9 ] ,运用分形理论研究根系,可以定量描述根系几何形态、分形特征及扩展范围等[10 ] . ...
酸枣根系结构可塑性对自然梯度干旱生境的适应机制
1
2018
... 根系是土壤和植物的动态界面.植物依靠根系获得固定和支撑,依赖根系吸收土壤中的矿质营养和水分,并输送到地上部分供其利用[1 ] .根系构型指植物根系在土壤中的分布和排列[2 ] .根系构型受遗传因素控制,生境也会影响根系构型的表达[3 ] .根系构型一定程度上反映了植物对环境的适应性及植被的退化和演替状况[4 ] .Lynch[2 ] 认为根系构型包括根系拓扑结构、根系形态、根系空间分布和根系构筑等内容.Fitter等[5 ] 提出了根系拓扑结构的两种极端类型,即鱼尾形分支和叉状分支类型.以往的研究较多通过外形、根冠比、根重等指标反映根系生长情况,而对根系的分支和结构特征研究较少[6 -7 ] .20世纪70年代,分形几何的诞生为根系构型研究提供了新方法[8 ] .根系有明显的分形结构[9 ] ,运用分形理论研究根系,可以定量描述根系几何形态、分形特征及扩展范围等[10 ] . ...
阿拉善高原2种荒漠植物根系构型及生态适应性特征
2
2020
... 根系是土壤和植物的动态界面.植物依靠根系获得固定和支撑,依赖根系吸收土壤中的矿质营养和水分,并输送到地上部分供其利用[1 ] .根系构型指植物根系在土壤中的分布和排列[2 ] .根系构型受遗传因素控制,生境也会影响根系构型的表达[3 ] .根系构型一定程度上反映了植物对环境的适应性及植被的退化和演替状况[4 ] .Lynch[2 ] 认为根系构型包括根系拓扑结构、根系形态、根系空间分布和根系构筑等内容.Fitter等[5 ] 提出了根系拓扑结构的两种极端类型,即鱼尾形分支和叉状分支类型.以往的研究较多通过外形、根冠比、根重等指标反映根系生长情况,而对根系的分支和结构特征研究较少[6 -7 ] .20世纪70年代,分形几何的诞生为根系构型研究提供了新方法[8 ] .根系有明显的分形结构[9 ] ,运用分形理论研究根系,可以定量描述根系几何形态、分形特征及扩展范围等[10 ] . ...
... 在干旱的荒漠地区,通过封育保护,流动沙地可以逐步演变为半固定和固定沙地.在这个过程中,一个重要的伴随演替特征就是沙面细土层的形成和生物结皮的出现[11 ] .按照出现的早晚,生物结皮主要分为藻结皮、地衣结皮和藓结皮[12 ] .生物结皮的出现显著改变了沙地地表覆盖条件,进而通过对土壤水文过程和种子扩散条件的改变,往往成为影响植被更新与根系生长的关键因素[13 ] .首先,生物结皮演替过程中土壤养分的逐渐增加为维管植物的存活和生长提供了宝贵的养分条件,直接影响着维管植物种子的散布、萌发和定居[14 ] .生物结皮发育也会导致表层土壤阻水能力增加,从而减少降水的入渗量[15 -16 ] ,加剧土壤干旱.在这个过程中,沙地植物可通过根系形态结构的改变来适应资源环境条件的变化[8 ] .通过对物种在不同沙地地表覆盖条件下根系构型的比较研究,有助于了解沙地演变以及植被更新演替的内生机制和趋势. ...
冬小麦根系形态的分形特征
2
1994
... 根系是土壤和植物的动态界面.植物依靠根系获得固定和支撑,依赖根系吸收土壤中的矿质营养和水分,并输送到地上部分供其利用[1 ] .根系构型指植物根系在土壤中的分布和排列[2 ] .根系构型受遗传因素控制,生境也会影响根系构型的表达[3 ] .根系构型一定程度上反映了植物对环境的适应性及植被的退化和演替状况[4 ] .Lynch[2 ] 认为根系构型包括根系拓扑结构、根系形态、根系空间分布和根系构筑等内容.Fitter等[5 ] 提出了根系拓扑结构的两种极端类型,即鱼尾形分支和叉状分支类型.以往的研究较多通过外形、根冠比、根重等指标反映根系生长情况,而对根系的分支和结构特征研究较少[6 -7 ] .20世纪70年代,分形几何的诞生为根系构型研究提供了新方法[8 ] .根系有明显的分形结构[9 ] ,运用分形理论研究根系,可以定量描述根系几何形态、分形特征及扩展范围等[10 ] . ...
... 分形维数反映了根系分支复杂程度,是植物根系发育程度和适应胁迫环境策略的重要指标 [9 ] ,分形丰度反映的是根系在生长介质中拓展程度,可定量化描述根系对空间占有能力及吸收水肥的效率[27 ] .本研究发现流沙地猪毛菜(FD =1.2823)、藻结皮地猪毛菜(FD =1.313)和藓结皮地猪毛菜(FD =1.397)分形维数总体较小且逐渐增加.这表明在干旱胁迫条件下,猪毛菜的根系结构较简单.然而随着养分和水分条件的改善,根系增加了根系分支以及分支长度,从而提高了对养分的利用效率.根系分形丰度反映了根系的延展范围、拓展能力及对水分和养分的吸收效率 [37 ] .本研究发现,流沙地猪毛菜(FA =3.5282)、藻结皮地猪毛菜(FA =3.1123)和藓结皮地猪毛菜(FA =2.6361)均维持较大的分形丰度.流沙地猪毛菜根系分形丰度最高,表明其根系体积大,吸收效率和空间占有能力得到进一步的提高.猪毛菜分形维数和分形丰度的特点,综合反映了其对不同地表生境的适应策略:流沙地中,由于资源比较分散并且分布较深,猪毛菜根系采取高生长量策略,空间拓展能力增加,因此具有更高的分形丰度;而在生物结皮地中,由于资源分布相对集中,并且植物之间存在根际竞争,根系采取结构优化策略,通过根系结构复杂化,提高了对局部资源利用的充分性,因此具有更大的分形维数,而藻结皮地和藓结皮地由于结皮覆盖度以及结皮组分不同,进而引起养分和水分的差异,分形维数和分形丰度出现显著变化. ...
分形几何在植物根系研究中的应用
1
1999
... 根系是土壤和植物的动态界面.植物依靠根系获得固定和支撑,依赖根系吸收土壤中的矿质营养和水分,并输送到地上部分供其利用[1 ] .根系构型指植物根系在土壤中的分布和排列[2 ] .根系构型受遗传因素控制,生境也会影响根系构型的表达[3 ] .根系构型一定程度上反映了植物对环境的适应性及植被的退化和演替状况[4 ] .Lynch[2 ] 认为根系构型包括根系拓扑结构、根系形态、根系空间分布和根系构筑等内容.Fitter等[5 ] 提出了根系拓扑结构的两种极端类型,即鱼尾形分支和叉状分支类型.以往的研究较多通过外形、根冠比、根重等指标反映根系生长情况,而对根系的分支和结构特征研究较少[6 -7 ] .20世纪70年代,分形几何的诞生为根系构型研究提供了新方法[8 ] .根系有明显的分形结构[9 ] ,运用分形理论研究根系,可以定量描述根系几何形态、分形特征及扩展范围等[10 ] . ...
沙坡头地区土壤结皮形成机理的研究
1
1996
... 在干旱的荒漠地区,通过封育保护,流动沙地可以逐步演变为半固定和固定沙地.在这个过程中,一个重要的伴随演替特征就是沙面细土层的形成和生物结皮的出现[11 ] .按照出现的早晚,生物结皮主要分为藻结皮、地衣结皮和藓结皮[12 ] .生物结皮的出现显著改变了沙地地表覆盖条件,进而通过对土壤水文过程和种子扩散条件的改变,往往成为影响植被更新与根系生长的关键因素[13 ] .首先,生物结皮演替过程中土壤养分的逐渐增加为维管植物的存活和生长提供了宝贵的养分条件,直接影响着维管植物种子的散布、萌发和定居[14 ] .生物结皮发育也会导致表层土壤阻水能力增加,从而减少降水的入渗量[15 -16 ] ,加剧土壤干旱.在这个过程中,沙地植物可通过根系形态结构的改变来适应资源环境条件的变化[8 ] .通过对物种在不同沙地地表覆盖条件下根系构型的比较研究,有助于了解沙地演变以及植被更新演替的内生机制和趋势. ...
Taxonomic composition and photosynthetic characteristics of the ‘biological soil crusts' covering sand dunes in the western Negev Desert
1
1992
... 在干旱的荒漠地区,通过封育保护,流动沙地可以逐步演变为半固定和固定沙地.在这个过程中,一个重要的伴随演替特征就是沙面细土层的形成和生物结皮的出现[11 ] .按照出现的早晚,生物结皮主要分为藻结皮、地衣结皮和藓结皮[12 ] .生物结皮的出现显著改变了沙地地表覆盖条件,进而通过对土壤水文过程和种子扩散条件的改变,往往成为影响植被更新与根系生长的关键因素[13 ] .首先,生物结皮演替过程中土壤养分的逐渐增加为维管植物的存活和生长提供了宝贵的养分条件,直接影响着维管植物种子的散布、萌发和定居[14 ] .生物结皮发育也会导致表层土壤阻水能力增加,从而减少降水的入渗量[15 -16 ] ,加剧土壤干旱.在这个过程中,沙地植物可通过根系形态结构的改变来适应资源环境条件的变化[8 ] .通过对物种在不同沙地地表覆盖条件下根系构型的比较研究,有助于了解沙地演变以及植被更新演替的内生机制和趋势. ...
荒漠地表生物土壤结皮形成与演替特征概述
1
2010
... 在干旱的荒漠地区,通过封育保护,流动沙地可以逐步演变为半固定和固定沙地.在这个过程中,一个重要的伴随演替特征就是沙面细土层的形成和生物结皮的出现[11 ] .按照出现的早晚,生物结皮主要分为藻结皮、地衣结皮和藓结皮[12 ] .生物结皮的出现显著改变了沙地地表覆盖条件,进而通过对土壤水文过程和种子扩散条件的改变,往往成为影响植被更新与根系生长的关键因素[13 ] .首先,生物结皮演替过程中土壤养分的逐渐增加为维管植物的存活和生长提供了宝贵的养分条件,直接影响着维管植物种子的散布、萌发和定居[14 ] .生物结皮发育也会导致表层土壤阻水能力增加,从而减少降水的入渗量[15 -16 ] ,加剧土壤干旱.在这个过程中,沙地植物可通过根系形态结构的改变来适应资源环境条件的变化[8 ] .通过对物种在不同沙地地表覆盖条件下根系构型的比较研究,有助于了解沙地演变以及植被更新演替的内生机制和趋势. ...
Soil microstructure in soils of the Colorado Plateau:the role of the cyanobacterium Microcoleus vaginatus
1
1993
... 在干旱的荒漠地区,通过封育保护,流动沙地可以逐步演变为半固定和固定沙地.在这个过程中,一个重要的伴随演替特征就是沙面细土层的形成和生物结皮的出现[11 ] .按照出现的早晚,生物结皮主要分为藻结皮、地衣结皮和藓结皮[12 ] .生物结皮的出现显著改变了沙地地表覆盖条件,进而通过对土壤水文过程和种子扩散条件的改变,往往成为影响植被更新与根系生长的关键因素[13 ] .首先,生物结皮演替过程中土壤养分的逐渐增加为维管植物的存活和生长提供了宝贵的养分条件,直接影响着维管植物种子的散布、萌发和定居[14 ] .生物结皮发育也会导致表层土壤阻水能力增加,从而减少降水的入渗量[15 -16 ] ,加剧土壤干旱.在这个过程中,沙地植物可通过根系形态结构的改变来适应资源环境条件的变化[8 ] .通过对物种在不同沙地地表覆盖条件下根系构型的比较研究,有助于了解沙地演变以及植被更新演替的内生机制和趋势. ...
毛乌素沙地土壤结皮对水分运动的影响
1
2010
... 在干旱的荒漠地区,通过封育保护,流动沙地可以逐步演变为半固定和固定沙地.在这个过程中,一个重要的伴随演替特征就是沙面细土层的形成和生物结皮的出现[11 ] .按照出现的早晚,生物结皮主要分为藻结皮、地衣结皮和藓结皮[12 ] .生物结皮的出现显著改变了沙地地表覆盖条件,进而通过对土壤水文过程和种子扩散条件的改变,往往成为影响植被更新与根系生长的关键因素[13 ] .首先,生物结皮演替过程中土壤养分的逐渐增加为维管植物的存活和生长提供了宝贵的养分条件,直接影响着维管植物种子的散布、萌发和定居[14 ] .生物结皮发育也会导致表层土壤阻水能力增加,从而减少降水的入渗量[15 -16 ] ,加剧土壤干旱.在这个过程中,沙地植物可通过根系形态结构的改变来适应资源环境条件的变化[8 ] .通过对物种在不同沙地地表覆盖条件下根系构型的比较研究,有助于了解沙地演变以及植被更新演替的内生机制和趋势. ...
毛乌素沙地生物结皮对水分入渗和再分配的影响
1
2011
... 在干旱的荒漠地区,通过封育保护,流动沙地可以逐步演变为半固定和固定沙地.在这个过程中,一个重要的伴随演替特征就是沙面细土层的形成和生物结皮的出现[11 ] .按照出现的早晚,生物结皮主要分为藻结皮、地衣结皮和藓结皮[12 ] .生物结皮的出现显著改变了沙地地表覆盖条件,进而通过对土壤水文过程和种子扩散条件的改变,往往成为影响植被更新与根系生长的关键因素[13 ] .首先,生物结皮演替过程中土壤养分的逐渐增加为维管植物的存活和生长提供了宝贵的养分条件,直接影响着维管植物种子的散布、萌发和定居[14 ] .生物结皮发育也会导致表层土壤阻水能力增加,从而减少降水的入渗量[15 -16 ] ,加剧土壤干旱.在这个过程中,沙地植物可通过根系形态结构的改变来适应资源环境条件的变化[8 ] .通过对物种在不同沙地地表覆盖条件下根系构型的比较研究,有助于了解沙地演变以及植被更新演替的内生机制和趋势. ...
水蚀风蚀交错带植被恢复对土壤质量的影响与植物生理生态适应性
1
2010
... 一年生草本植物猪毛菜(Salsola collina ),在荒漠区和极端干旱区广泛分布.猪毛菜既能防风固沙,也是饲用植物,在干旱地区退化土地水土保持和植被恢复方面具有很大的应用潜力[17 -18 ] .本文以腾格里沙漠东南缘的野生猪毛菜为对象,研究其在不同地表条件下的根系构型特征,比较了不同地表条件对猪毛菜根系生长以及构型的影响,揭示了猪毛菜对沙地环境演变的适应机制. ...
猪毛菜响应干旱胁迫的叶片结构、生理及转录组分析
1
2024
... 一年生草本植物猪毛菜(Salsola collina ),在荒漠区和极端干旱区广泛分布.猪毛菜既能防风固沙,也是饲用植物,在干旱地区退化土地水土保持和植被恢复方面具有很大的应用潜力[17 -18 ] .本文以腾格里沙漠东南缘的野生猪毛菜为对象,研究其在不同地表条件下的根系构型特征,比较了不同地表条件对猪毛菜根系生长以及构型的影响,揭示了猪毛菜对沙地环境演变的适应机制. ...
腾格里沙漠沙坡头地区固沙植被对生物多样性恢复的长期影响
1
2005
... 试验地点位于宁夏回族自治区中卫市,腾格里沙漠东南缘的沙坡头地区(37°27′N、104°05′E,海拔1 310~1 350 m,图1 ),该地区是荒漠和荒漠草原的过渡地带,最低气温-25.1 ℃,最高气温38.1 ℃,年均气温约9.5 ℃,年降水量约200 mm,降水主要在5—9月;全年日照约3 260 h,年蒸发量约2 400 mm,土壤稳定含水量约2.5%[19 ] .灌木主要为柠条锦鸡儿(Caragana korshinskii )、花棒(Corethrodendron scoparium ),草本植物主要为小画眉草(Eragrostis minor )、狗尾草(Setaria viridis )、猪毛菜(Salsola collina ),雾冰藜(Grubovia dasyphylla )、虫实(Corispermum hyssopifolium )、砂蓝刺头(Echinops gmelinii ). ...
平地与边坡条件下野生胡枝子根构型特征研究
1
2019
... 选取的植株为当年生长的野生猪毛菜,对猪毛菜的地上部分指标(株高、地径、冠幅)进行调查,统计确定标准样株(表2 ).参照标准样株,在每个试验地内选择猪毛菜各10株,共选取30株植物,研究供试植株地上部带回实验室烘干称重.利用铁锹和毛刷等工具从植物根基部挖掘取样,直至无根系出现.如根系断裂,标记好位置并将根系放入自封袋中[20 ] .取样过程中,仔细观察根系在原位的整体构型,记录根系的内部、外部连接数量,并测量根系水平幅和根系垂直幅,最后将样品放入冰盒(内径规格:长29 cm,宽23 cm,高21.5 cm)中带回实验室.清除根系表面杂质后进行图像扫描,之后设置烘箱温度为105 ℃,将根系和植株地上部分杀青120 min后,再设置烘箱温度75 ℃下烘干48 h,最后用电子天平(0.0001 g)称重,得到根系生物量和地上生物量.使用Epson Scan 2扫描仪对猪毛菜根系进行扫描,获得图像,采用根系分析系统软件Win-RHIZO PRO 2020进行分析,分析指标包含根长(RL)、根表面积(RSA)、根系分布图上边长为r 的正方形和根系所截的正方形数目Nr . ...
The topology and geometry of plant root systems:influence of watering rate on root system topology in Trifolium pratense
2
1986
... 根系拓扑结构存在两种极端模式[21 -24 ] .根据模型计算拓扑指数TI ,拓扑指数表达式为 ...
... 在植物根系中,拓扑结构与营养物质的竞争力之间存在着密切的关系.鱼尾形分支根系通常结构简单,分布范围较大,从而增加了对养分的获取能力.相比之下,叉状分支则呈现出相反的特点.在等量的碳投入条件下,鱼尾形分支结构更有利于营养空间的占用.因此,鱼尾形分支的根系结构在干旱和贫瘠的环境中表现出更高的适应性[21 -24 ] .本研究发现,从流沙地、藻结皮地到藓结皮地,猪毛菜的根系结构趋向于从鱼尾形分支转变成叉状分支.表明猪毛菜在养分和水分条件不足的生境中,根系结构具有分支简单、次级分支数量较少、但分布较深的特点,这种根系结构增加了获取营养物质的能力.然而随着地表生境的改善,养分和水分在地表方向集中,使得根系分支浅层化,趋向于叉状分支结构,这种根系结构增加了对有限空间内营养物质的利用能力.这表明猪毛菜的根系拓扑结构与生境中的养分存在着动态平衡,优化自身结构以得到更好的生长. ...
An architectural approach to the comparative ecology of plant root systems
0
1987
Architectural analysis of plant root systems:III.studies on plants under field conditions
0
1992
Root system topology and diameter distribution of species from habitats differing in inundation frequency
2
2001
... 根系拓扑结构存在两种极端模式[21 -24 ] .根据模型计算拓扑指数TI ,拓扑指数表达式为 ...
... 在植物根系中,拓扑结构与营养物质的竞争力之间存在着密切的关系.鱼尾形分支根系通常结构简单,分布范围较大,从而增加了对养分的获取能力.相比之下,叉状分支则呈现出相反的特点.在等量的碳投入条件下,鱼尾形分支结构更有利于营养空间的占用.因此,鱼尾形分支的根系结构在干旱和贫瘠的环境中表现出更高的适应性[21 -24 ] .本研究发现,从流沙地、藻结皮地到藓结皮地,猪毛菜的根系结构趋向于从鱼尾形分支转变成叉状分支.表明猪毛菜在养分和水分条件不足的生境中,根系结构具有分支简单、次级分支数量较少、但分布较深的特点,这种根系结构增加了获取营养物质的能力.然而随着地表生境的改善,养分和水分在地表方向集中,使得根系分支浅层化,趋向于叉状分支结构,这种根系结构增加了对有限空间内营养物质的利用能力.这表明猪毛菜的根系拓扑结构与生境中的养分存在着动态平衡,优化自身结构以得到更好的生长. ...
尕海湿地不同密度条件下垂穗披碱草根系分形结构
1
2018
... 根据Win-RHIZO测定的根系分布图上边长为r 的正方形和根系所截的正方形数目Nr ,采用盒维数法[25 ] 得到根系分形特征,随着正方形边r 逐渐减小,根系所截Nr 逐渐增大,得到不同的r 上相应Nr 值后,分别以lgr 、lgNr 为横坐标和纵坐标作图,回归直线方程为lgNr =-FD lgr +lgK ,回归直线斜率的负数是所求的分形维数,lgK 为分形丰度. ...
砒砂岩区不同生境条件下沙棘根系构型特征研究
1
2018
... 3种地表条件下猪毛菜的根系分形维数与根系分形丰度存在明显差异(图5 ).藓结皮地的根系分形维数最大,显著大于藻结皮地和流沙地(P <0.05),而藻结皮地和流沙地的根系分形维数无明显差异(P >0.05),但从流沙地、藻结皮地到藓结皮地,根系分形维数呈现增大的趋势,表明猪毛菜的根系结构复杂化.一般植物根系的分形维数为1.0~1.9,当分形维数>1.1时,就可判定研究对象具有明显的分形特征[26 ] .猪毛菜在不同的地表条件下根系分形维数都大于1.1,表明猪毛菜根系具有很好的分形特征.流沙地的根系分形丰度最大,比藻结皮地和藓结皮地分别大了13%和34%.藓结皮地的根系分形丰度最小,显著小于流沙地和藻结皮地.从流沙地、藻结皮地到藓结皮地,根系分形丰度呈现减小的趋势,表明猪毛菜根系拓展能力下降,在土壤中拓展的体积逐渐减少. ...
河西走廊中部两种荒漠植物根系构型特征
2
2013
... 植物根系的空间分布特征受遗传因素和环境因素的共同影响.植物根系在环境作用下会发生一定的空间分布变化,表现出高度的可塑性[27 -29 ] .生物结皮不仅能显著提高表层土壤有机质和土壤养分的含量,也可显著提高碳、氮、磷等元素相关的土壤酶活性[30 -32 ] .同时,生物结皮的存在阻碍土壤水分入渗,具体表现为与裸地相比,降低初始入渗率、平均入渗率、稳定入渗率及非饱和导水率[33 ] .本研究发现,从流沙地、藻结皮地到藓结皮地,猪毛菜的根系水平幅显著增加,而根系垂直幅逐渐降低,且地上部分和地下部分生物量变化不大.这可能是由于地表演变导致结皮中养分和水分的增加,使得根系分支横向生长获取表层资源,根系分支生长浅层化,同时表层有限的资源使猪毛菜首先进行地下部分的发育,地上部分变化不显著,具体表现为根系结构的复杂化以充分利用资源,并且根冠比的显著差异表明猪毛菜在生境改善的过程中减小了对根系生长的投入,根系结构的转变有可能会出现生物量下降的趋势,这与苏纪帅等[34 ] 对封育条件下的针茅研究结果相符.在根系形态方面,可能由于物种的遗传因素,根系垂直幅大于水平幅,因此深宽比>1.作为干旱荒漠区广泛分布的植物种,这种对生境的适应以及遗传的表现,是猪毛菜应对荒漠地区水分和养分胁迫的重要策略. ...
... 分形维数反映了根系分支复杂程度,是植物根系发育程度和适应胁迫环境策略的重要指标 [9 ] ,分形丰度反映的是根系在生长介质中拓展程度,可定量化描述根系对空间占有能力及吸收水肥的效率[27 ] .本研究发现流沙地猪毛菜(FD =1.2823)、藻结皮地猪毛菜(FD =1.313)和藓结皮地猪毛菜(FD =1.397)分形维数总体较小且逐渐增加.这表明在干旱胁迫条件下,猪毛菜的根系结构较简单.然而随着养分和水分条件的改善,根系增加了根系分支以及分支长度,从而提高了对养分的利用效率.根系分形丰度反映了根系的延展范围、拓展能力及对水分和养分的吸收效率 [37 ] .本研究发现,流沙地猪毛菜(FA =3.5282)、藻结皮地猪毛菜(FA =3.1123)和藓结皮地猪毛菜(FA =2.6361)均维持较大的分形丰度.流沙地猪毛菜根系分形丰度最高,表明其根系体积大,吸收效率和空间占有能力得到进一步的提高.猪毛菜分形维数和分形丰度的特点,综合反映了其对不同地表生境的适应策略:流沙地中,由于资源比较分散并且分布较深,猪毛菜根系采取高生长量策略,空间拓展能力增加,因此具有更高的分形丰度;而在生物结皮地中,由于资源分布相对集中,并且植物之间存在根际竞争,根系采取结构优化策略,通过根系结构复杂化,提高了对局部资源利用的充分性,因此具有更大的分形维数,而藻结皮地和藓结皮地由于结皮覆盖度以及结皮组分不同,进而引起养分和水分的差异,分形维数和分形丰度出现显著变化. ...
黄河三角洲贝壳堤岛3种优势灌木的根系构型
0
2015
Multi-proxy evidence for competition between savanna woody species
1
2008
... 植物根系的空间分布特征受遗传因素和环境因素的共同影响.植物根系在环境作用下会发生一定的空间分布变化,表现出高度的可塑性[27 -29 ] .生物结皮不仅能显著提高表层土壤有机质和土壤养分的含量,也可显著提高碳、氮、磷等元素相关的土壤酶活性[30 -32 ] .同时,生物结皮的存在阻碍土壤水分入渗,具体表现为与裸地相比,降低初始入渗率、平均入渗率、稳定入渗率及非饱和导水率[33 ] .本研究发现,从流沙地、藻结皮地到藓结皮地,猪毛菜的根系水平幅显著增加,而根系垂直幅逐渐降低,且地上部分和地下部分生物量变化不大.这可能是由于地表演变导致结皮中养分和水分的增加,使得根系分支横向生长获取表层资源,根系分支生长浅层化,同时表层有限的资源使猪毛菜首先进行地下部分的发育,地上部分变化不显著,具体表现为根系结构的复杂化以充分利用资源,并且根冠比的显著差异表明猪毛菜在生境改善的过程中减小了对根系生长的投入,根系结构的转变有可能会出现生物量下降的趋势,这与苏纪帅等[34 ] 对封育条件下的针茅研究结果相符.在根系形态方面,可能由于物种的遗传因素,根系垂直幅大于水平幅,因此深宽比>1.作为干旱荒漠区广泛分布的植物种,这种对生境的适应以及遗传的表现,是猪毛菜应对荒漠地区水分和养分胁迫的重要策略. ...
Responses of microbial activities and soil physical-chemical properties to the successional process of biological soil crusts in the Gurbantunggut Desert,Xinjiang
1
2015
... 植物根系的空间分布特征受遗传因素和环境因素的共同影响.植物根系在环境作用下会发生一定的空间分布变化,表现出高度的可塑性[27 -29 ] .生物结皮不仅能显著提高表层土壤有机质和土壤养分的含量,也可显著提高碳、氮、磷等元素相关的土壤酶活性[30 -32 ] .同时,生物结皮的存在阻碍土壤水分入渗,具体表现为与裸地相比,降低初始入渗率、平均入渗率、稳定入渗率及非饱和导水率[33 ] .本研究发现,从流沙地、藻结皮地到藓结皮地,猪毛菜的根系水平幅显著增加,而根系垂直幅逐渐降低,且地上部分和地下部分生物量变化不大.这可能是由于地表演变导致结皮中养分和水分的增加,使得根系分支横向生长获取表层资源,根系分支生长浅层化,同时表层有限的资源使猪毛菜首先进行地下部分的发育,地上部分变化不显著,具体表现为根系结构的复杂化以充分利用资源,并且根冠比的显著差异表明猪毛菜在生境改善的过程中减小了对根系生长的投入,根系结构的转变有可能会出现生物量下降的趋势,这与苏纪帅等[34 ] 对封育条件下的针茅研究结果相符.在根系形态方面,可能由于物种的遗传因素,根系垂直幅大于水平幅,因此深宽比>1.作为干旱荒漠区广泛分布的植物种,这种对生境的适应以及遗传的表现,是猪毛菜应对荒漠地区水分和养分胁迫的重要策略. ...
不同土壤和植被生境下生物结皮对土壤性质的影响
0
2019
Impact of human trampling on biological soil crusts determined by soil microbial biomass,enzyme activities and nematode communities in a desert ecosystem
1
2018
... 植物根系的空间分布特征受遗传因素和环境因素的共同影响.植物根系在环境作用下会发生一定的空间分布变化,表现出高度的可塑性[27 -29 ] .生物结皮不仅能显著提高表层土壤有机质和土壤养分的含量,也可显著提高碳、氮、磷等元素相关的土壤酶活性[30 -32 ] .同时,生物结皮的存在阻碍土壤水分入渗,具体表现为与裸地相比,降低初始入渗率、平均入渗率、稳定入渗率及非饱和导水率[33 ] .本研究发现,从流沙地、藻结皮地到藓结皮地,猪毛菜的根系水平幅显著增加,而根系垂直幅逐渐降低,且地上部分和地下部分生物量变化不大.这可能是由于地表演变导致结皮中养分和水分的增加,使得根系分支横向生长获取表层资源,根系分支生长浅层化,同时表层有限的资源使猪毛菜首先进行地下部分的发育,地上部分变化不显著,具体表现为根系结构的复杂化以充分利用资源,并且根冠比的显著差异表明猪毛菜在生境改善的过程中减小了对根系生长的投入,根系结构的转变有可能会出现生物量下降的趋势,这与苏纪帅等[34 ] 对封育条件下的针茅研究结果相符.在根系形态方面,可能由于物种的遗传因素,根系垂直幅大于水平幅,因此深宽比>1.作为干旱荒漠区广泛分布的植物种,这种对生境的适应以及遗传的表现,是猪毛菜应对荒漠地区水分和养分胁迫的重要策略. ...
红壤丘陵区不同盖度生物结皮对水分入渗的影响
1
2023
... 植物根系的空间分布特征受遗传因素和环境因素的共同影响.植物根系在环境作用下会发生一定的空间分布变化,表现出高度的可塑性[27 -29 ] .生物结皮不仅能显著提高表层土壤有机质和土壤养分的含量,也可显著提高碳、氮、磷等元素相关的土壤酶活性[30 -32 ] .同时,生物结皮的存在阻碍土壤水分入渗,具体表现为与裸地相比,降低初始入渗率、平均入渗率、稳定入渗率及非饱和导水率[33 ] .本研究发现,从流沙地、藻结皮地到藓结皮地,猪毛菜的根系水平幅显著增加,而根系垂直幅逐渐降低,且地上部分和地下部分生物量变化不大.这可能是由于地表演变导致结皮中养分和水分的增加,使得根系分支横向生长获取表层资源,根系分支生长浅层化,同时表层有限的资源使猪毛菜首先进行地下部分的发育,地上部分变化不显著,具体表现为根系结构的复杂化以充分利用资源,并且根冠比的显著差异表明猪毛菜在生境改善的过程中减小了对根系生长的投入,根系结构的转变有可能会出现生物量下降的趋势,这与苏纪帅等[34 ] 对封育条件下的针茅研究结果相符.在根系形态方面,可能由于物种的遗传因素,根系垂直幅大于水平幅,因此深宽比>1.作为干旱荒漠区广泛分布的植物种,这种对生境的适应以及遗传的表现,是猪毛菜应对荒漠地区水分和养分胁迫的重要策略. ...
半干旱草地长期封育进程中针茅植物根系格局变化特征
1
2017
... 植物根系的空间分布特征受遗传因素和环境因素的共同影响.植物根系在环境作用下会发生一定的空间分布变化,表现出高度的可塑性[27 -29 ] .生物结皮不仅能显著提高表层土壤有机质和土壤养分的含量,也可显著提高碳、氮、磷等元素相关的土壤酶活性[30 -32 ] .同时,生物结皮的存在阻碍土壤水分入渗,具体表现为与裸地相比,降低初始入渗率、平均入渗率、稳定入渗率及非饱和导水率[33 ] .本研究发现,从流沙地、藻结皮地到藓结皮地,猪毛菜的根系水平幅显著增加,而根系垂直幅逐渐降低,且地上部分和地下部分生物量变化不大.这可能是由于地表演变导致结皮中养分和水分的增加,使得根系分支横向生长获取表层资源,根系分支生长浅层化,同时表层有限的资源使猪毛菜首先进行地下部分的发育,地上部分变化不显著,具体表现为根系结构的复杂化以充分利用资源,并且根冠比的显著差异表明猪毛菜在生境改善的过程中减小了对根系生长的投入,根系结构的转变有可能会出现生物量下降的趋势,这与苏纪帅等[34 ] 对封育条件下的针茅研究结果相符.在根系形态方面,可能由于物种的遗传因素,根系垂直幅大于水平幅,因此深宽比>1.作为干旱荒漠区广泛分布的植物种,这种对生境的适应以及遗传的表现,是猪毛菜应对荒漠地区水分和养分胁迫的重要策略. ...
Aboveground and belowground interactions in mixed plantations of Eucalyptus globulus and Acacia mearnsii
1
2000
... 比根长和比表面积是根系形态结构的重要指标,决定了根系吸收水分和养分的能力,其值与养分吸收效率成正比[35 ] .本研究发现,从流沙地、藻结皮地到藓结皮地,猪毛菜的比根长和比表面积不断减小,这可能是由于其根系水平幅的增加,根系分支浅层化,总体根长减少.同时由于根系响应生物结皮层中的养分和水分增加,减少了对根系获取沙地深处水分的资源投入,调节根的直径和根的数量来减少了比根长和比表面积.这与单立山等[36 ] 对珍珠猪毛菜的研究结果大致相符,说明在生境资源逐渐丰富的条件下,猪毛菜会调节其根系结构,同时减少比根长和比表面积来适应. ...
不同生境下荒漠植物红砂-珍珠猪毛菜混生根系的垂直分布规律
1
2018
... 比根长和比表面积是根系形态结构的重要指标,决定了根系吸收水分和养分的能力,其值与养分吸收效率成正比[35 ] .本研究发现,从流沙地、藻结皮地到藓结皮地,猪毛菜的比根长和比表面积不断减小,这可能是由于其根系水平幅的增加,根系分支浅层化,总体根长减少.同时由于根系响应生物结皮层中的养分和水分增加,减少了对根系获取沙地深处水分的资源投入,调节根的直径和根的数量来减少了比根长和比表面积.这与单立山等[36 ] 对珍珠猪毛菜的研究结果大致相符,说明在生境资源逐渐丰富的条件下,猪毛菜会调节其根系结构,同时减少比根长和比表面积来适应. ...
塔克拉玛干沙漠腹地几种植物根系分形特征
1
2009
... 分形维数反映了根系分支复杂程度,是植物根系发育程度和适应胁迫环境策略的重要指标 [9 ] ,分形丰度反映的是根系在生长介质中拓展程度,可定量化描述根系对空间占有能力及吸收水肥的效率[27 ] .本研究发现流沙地猪毛菜(FD =1.2823)、藻结皮地猪毛菜(FD =1.313)和藓结皮地猪毛菜(FD =1.397)分形维数总体较小且逐渐增加.这表明在干旱胁迫条件下,猪毛菜的根系结构较简单.然而随着养分和水分条件的改善,根系增加了根系分支以及分支长度,从而提高了对养分的利用效率.根系分形丰度反映了根系的延展范围、拓展能力及对水分和养分的吸收效率 [37 ] .本研究发现,流沙地猪毛菜(FA =3.5282)、藻结皮地猪毛菜(FA =3.1123)和藓结皮地猪毛菜(FA =2.6361)均维持较大的分形丰度.流沙地猪毛菜根系分形丰度最高,表明其根系体积大,吸收效率和空间占有能力得到进一步的提高.猪毛菜分形维数和分形丰度的特点,综合反映了其对不同地表生境的适应策略:流沙地中,由于资源比较分散并且分布较深,猪毛菜根系采取高生长量策略,空间拓展能力增加,因此具有更高的分形丰度;而在生物结皮地中,由于资源分布相对集中,并且植物之间存在根际竞争,根系采取结构优化策略,通过根系结构复杂化,提高了对局部资源利用的充分性,因此具有更大的分形维数,而藻结皮地和藓结皮地由于结皮覆盖度以及结皮组分不同,进而引起养分和水分的差异,分形维数和分形丰度出现显著变化. ...







 甘公网安备 62010202000688号
甘公网安备 62010202000688号
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}