


0 引言
目前,国内外关于沙区土壤水分分布特征及其与地形和植被因子的关系研究主要从不同尺度上展开。一方面,在群落尺度上,土壤水分含量影响植被类型、分布、结构和组成,而在区域尺度上,土壤水分含量影响植被格局、景观分布、竞争相互作用和演替过程[6-7]。此外,植被的结构、类型以及群落组成也会对土壤水分的分布特征有影响。土壤水分和植物之间的耦合和相互作用进一步使它们之间关系的变得复杂。另一方面,地形一直被认为是影响局部尺度植被格局的重要因素,这主要是由于海拔和气温的密切关系[8-9]。自1890年代以来,研究人员已经明确了地形可通过影响周围的非生物环境来影响当地的植被模式[10]。地形在确定不同生境的植物分布和多样性模式方面发挥着至关重要的作用,研究主要在森林、沼泽和红树林、热带雨林、草原、沙漠和稀树草原等区域[11-16]。目前关于沙区在小尺度上对土壤水分含量与地形、植被因子之间的关系研究较少,需要进一步研究沙丘内部地形和植被因子与土壤水分的关系和对土壤水分含量的影响机理。因此,本文采用样带-网格数据,运用多个模型在小尺度上对不同深度土壤水分的分布特征以及与地形、植被因子之间的关系进行探讨。
1 研究区概况
腾格里沙漠面积约4.27
2 材料与方法
2.1 试验设计
2023年10月,在腾格里沙漠红卫地区的固定沙丘上设置了1块试验样地,将样地划分为10行37列,每个小样方的大小为4×4 m(图1)。其中3行被选择为土壤水分的采样点。采用100 cm×100 cm的草本样方,调查了样地中草本植被的分布状况,包括草本盖度、草本多度、凋落物和生物量等。同时,使用RTK精确测量调查样地的每个小样方的4个顶点,实时动态定位样地中每株灌木的位置,测定了灌木盖度-灌木多度,记录其海拔和经纬度数据。随后,利用数字高程模型计算了小样方的高差、坡度和坡向[19]。样地的高差平均为2 m,范围0~6.4 m,样地的坡度0°~17.64°,样地的坡向(以正东为0°)平均为160.4°,范围0°~359°。
图1
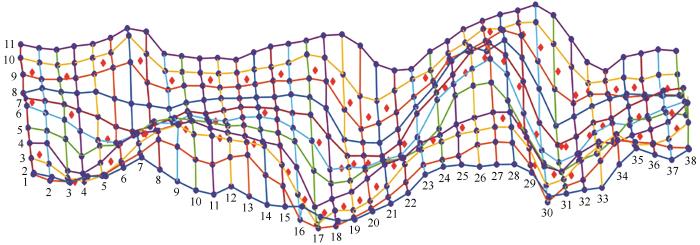
图1
试验样地及土壤水分采样点
Fig.1
Experimental sample plots and soil moisture sampling points
图1为试验样地的划分以及土壤水分采样点的三维图。从下到上、从左到右选择第3、6、9行,每行设置24个土壤水分采样点,共计72个采样点。分别在0~5、5~15、15~25、25~35、35~50 cm取样,在50~300 cm深度每隔20 cm采样,共计18层。在105 ℃下,将土壤样品烘干24 h称重。为了减少称重误差,称重精度设置为0.1%。
2.2 研究方法
2.2.1 数据标准化
由于地形和植被因子指标的量纲和数量级不同,采用归一化方法对原始数据标准化。
式中:
2.2.2 广义线性模型
广义线性模型(GLM,Generalized Linear Model)是线性回归模型的推广。GLM模型允许因变量不服从正态分布,并适用于多种模型,包括高斯模型、泊松分布下的计数数据模型,以及逻辑回归模型中的分类变量模型[20]。
式中:
2.2.3 广义加性模型
广义加性模型(Generalized Additive Model,GAM)是将线性和非线性回归结合起来的模型,是广义线性和加性模型的非参数组合,适用于各种指数分布族的因变量。该模型通过非参数化平滑回归来解决因变量与预测变量之间高度的非线性的关系。
式中:
2.2.4 随机森林
随机森林(Random Forest, RF)模型是基于机器学习的集成算法[21]。它会对初始数据集进行自助法抽样,由抽取的样本数据集构建不同的决策树模型。然后,通过决策树模型的平均值或投票结果来得出最终结果。同时,随机森林模型也可用于筛选特征变量,并输出变量的重要性度量。精度平均减少值(Increasing the Mean Square Error,IncMSE)和节点不纯度减少值(Increasing the Node Impurity, IncNodePurity)是两种常用于判断变量重要性的指标。IncNodePurity以变量作为拆分变量所引起的均方误差的平均递减的程度来衡量变量的重要性度量指标,其值越大说明该变量越重要。
3 结果与分析
3.1 土壤水分层次的划分
不同沙区的土壤水分分层边界并不明确和一致。本研究将0~300 cm最上面的土层,即受降雨影响较大的土层,定义为表层;土壤水分含量变化相对不稳定的部分定义为中层;与日蒸散和规律性降雨事件关系较弱的土层定义为深层。通过计算0~300 cm不同深度的72个采样点的平均土壤水分含量和标准误,得到了土壤水分含量的均值和标准误随着土壤深度的变化图(图2~3)。研究区固定沙丘上0~300 cm土壤水分可划分为3层:①0~40 cm:土壤水分含量的均值和标准误在该层中随着土壤深度的增加而增加,主要受降雨的影响,定义为表层土壤。②40~200 cm:土壤水分含量的标准误先增加后减小,当土壤深度在40 cm和200 cm处时,标准误最小。土壤水分含量随着土壤深度的增加而增加,表明土壤水分含量的变化在该层是不稳定的。因此,将40~200 cm定义为中层土壤。③200~300 cm:土壤水分含量在这一层中变化相对稳定。尽管土壤水分含量随着深度的增加而增加,但标准误基本保持稳定,说明该层中土壤的水分含量变化受降雨渗入的影响很小。因此,将200~300 cm的土层定义为深层土壤。
图2
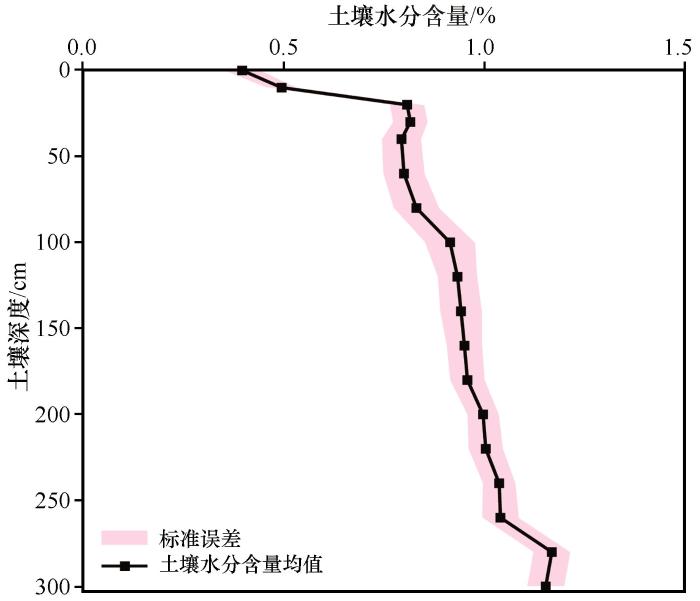
图3
3.2 不同土壤深度土壤水分含量变化特征
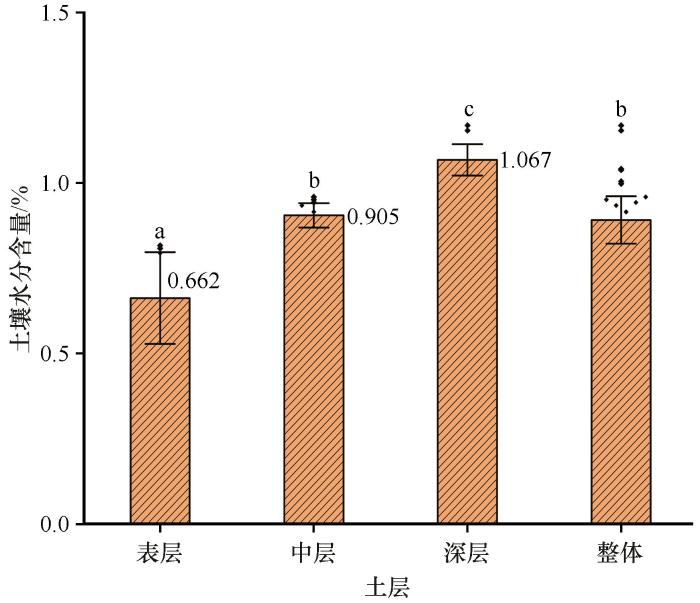
不同深度的土壤水分含量的均值深层(200~300 cm)>中层(40~200 cm)>表层(0~40 cm)。在25%~75%分位数,土壤水分含量主要集中在[1.004%,1.153%]深层、[0.831%,0.951%]中层、[0.495%,0.808%]表层和[0.816%,1.004%]整体。不同深度土壤水分含量之间差异较大。表层、中层和深层土壤水分含量之间均存在显著差异,而中层与整体的土壤水分含量之间无显著差异。
3.3 地形、植被因子对不同深度土壤水分含量的影响
为了研究不同深度土壤水分含量与地形、植被因子之间的关系,将表层、中层和深层土壤水分含量作为解释变量,地形因子(高差、坡度、坡向)和植被因子(草本盖度、草本多度、灌木盖度、灌木多度、生物量、凋落物)作为被解释变量,分别建立GLM、GAM和RF模型,并对影响不同深度土壤水分含量的因子进行显著性检验。
GLM模型结果表明(表1),地形因子高差对不同深度的土壤水分含量均有显著负向影响。植被因子灌木多度对表层土壤水分含量有显著负向影响;草本盖度和灌木多度对中层土壤水分含量有显著正向影响,而灌木盖度有显著负向影响;地形因子坡向对深层土壤水分含量有显著正向影响。
表1 基于广义线性模型的变量的显著性检验
Table 1
| 变量 | 0~40 cm | 40~200 cm | 200~300 cm | |||
|---|---|---|---|---|---|---|
| 估计值 | 估计值 | 估计值 | ||||
| 坡度 | 0.097 | 0.432 | 0.015 | 0.878 | 0.002 | 0.986 |
| 坡向 | 0.056 | 0.404 | 0.000 | 0.998 | 0.157 | 0.034 |
| 高差 | -0.291 | 0.022 | -0.379 | 0.000 | -0.351 | 0.011 |
| 灌木盖度 | -0.021 | 0.872 | -0.242 | 0.022 | 0.019 | 0.893 |
| 灌木多度 | -0.314 | 0.029 | 0.195 | 0.088 | 0.103 | 0.499 |
| 草本盖度 | 0.256 | 0.147 | 0.309 | 0.030 | -0.045 | 0.809 |
| 草本多度 | -0.143 | 0.484 | -0.025 | 0.880 | 0.211 | 0.339 |
| 凋落物 | 0.249 | 0.252 | -0.059 | 0.735 | -0.040 | 0.863 |
| 生物量 | 0.124 | 0.369 | -0.010 | 0.927 | -0.039 | 0.794 |
GAM模型的结果表明(表2),植被因子草本盖度、灌木多度、凋落物和生物量对表层土壤水分含量有显著影响,地形因子对表层土壤水分含量均没有显著影响。地形因子高差和植被因子草本盖度、灌木多度对中层土壤水分含量有显著影响。地形因子坡向和高差对深层土壤水分含量有显著影响。
表2 基于广义加性模型的变量的显著性检验
Table 2
| 变量 | 0~40 cm | 40~200 cm | 200~300 cm | |||
|---|---|---|---|---|---|---|
| edf | edf | edf | ||||
| 坡度 | 3.314 | 0.244 | 3.381 | 0.250 | 3.480 | 0.025 |
| 坡向 | 1.000 | 0.580 | 1.359 | 0.866 | 2.537 | 0.000 |
| 高差 | 1.000 | 0.197 | 1.000 | 0.006 | 1.448 | 0.000 |
| 灌木盖度 | 1.000 | 0.370 | 1.000 | 0.156 | 2.976 | 0.262 |
| 灌木多度 | 2.644 | 0.025 | 1.932 | 0.115 | 1.906 | 0.118 |
| 草本盖度 | 1.000 | 0.003 | 1.000 | 0.003 | 1.000 | 0.453 |
| 草本多度 | 1.241 | 0.052 | 1.000 | 0.224 | 1.778 | 0.301 |
| 凋落物 | 1.731 | 0.042 | 1.000 | 0.642 | 1.000 | 0.533 |
| 生物量 | 2.216 | 0.029 | 1.959 | 0.248 | 1.000 | 0.531 |
GLM和GAM模型均得出地形因子高差对中层和深层的土壤水分含量有显著的影响的结论。不同的是,GLM模型结果表明高差和灌木多度对表层土壤水分含量有显著负向影响,但GAM模型表明植被因子草本盖度、草本多度、凋落物以及生物量对表层土壤水分含量也有显著影响。GAM模型结果表明大部分的地形、植被因子仍与土壤水分含量呈现线性关系(edf≈1)。其中,地形因子坡度对不同深度的土壤水分含量均呈现出了非线性趋势(edf
表3为基于RF模型的地形、植被因子对不同深度土壤水分含量影响的相对重要性排序,表中列出了排名前6的地形、植被因子。由RF模型的结果可知,重要度排名第一的均为地形因子。与GLM和GAM模型的结果相似的是,高差对不同深度的土壤水分含量重要度均较高。植被因子中,灌木多度、灌木盖度和草本盖度对不同深度的土壤水分含量重要度也相对较高。与GLM和GAM模型不同的是,RF模型得出地形因子坡度对表层土壤水分含量影响较大,坡向对中层和深层土壤水分含量影响较大。同时,灌木多度对深层土壤水分含量影响较大;生物量对表层和中层土壤水分含量影响较大,且对表层的影响大于深层。
表3 随机森林模型下影响土壤水分含量的重要度
Table 3
| 排名 | 0~40 cm | 40~200 cm | 200~300 cm |
|---|---|---|---|
| 1 | 高差(46.01%) | 高差(36.40%) | 坡向(48.41%) |
| 2 | 灌木多度(45.33%) | 灌木多度(32.07%) | 高差(42.62%) |
| 3 | 灌木盖度(36.30%) | 草本盖度(25.90%) | 坡度(34.74%) |
| 4 | 生物量(31.25%) | 灌木盖度(24.23%) | 灌木盖度(32.27%) |
| 5 | 草本盖度(29.65%) | 凋落物(23.84%) | 生物量(30.13%) |
| 6 | 坡度(24.85%) | 生物量(22.22%) | 灌木多度(26.08 %) |
3.4 地形、植被因子对不同深度土壤水分含量影响的趋势分析
为了确定不同深度土壤水分含量随地形、植被因子的变化趋势,选取对不同深度对土壤水分含量有显著影响的地形、植被因子,运用GLM、GAM和RF模型得到各自的响应拟合曲线。
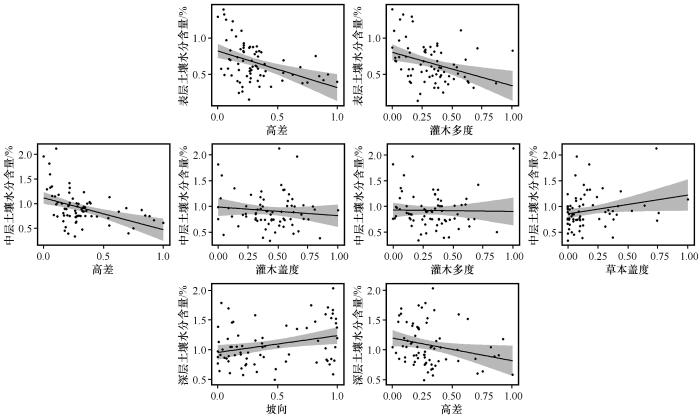
基于GLM模型的响应曲线如图4所示。表层土壤水分含量随着高差与灌木多度的增加呈现递减的趋势;中层土壤水分含量随草本盖度增加呈增加趋势,但随着高差、灌木盖度和灌木多度的增加呈下降趋势;深层土壤水分含量随着坡向的增加,呈增加趋势,但是随着高差的增加呈下降趋势。特别地,高差对不同深度土壤水分含量均有显著的负向影响。
图4
基于GAM模型的响应曲线如图5所示。结果表明,表层土壤水分含量随着草本盖度和生物量的增加呈增加趋势,但随着灌木多度的增加,表层土壤水分含量呈递减的趋势;中层土壤水分含量随着草本盖度的增加呈增加趋势,但随着高差的增加呈下降趋势;深层土壤水分含量随着坡向的增加呈增加趋势,但随着坡度和高差的增加呈下降趋势。
图5
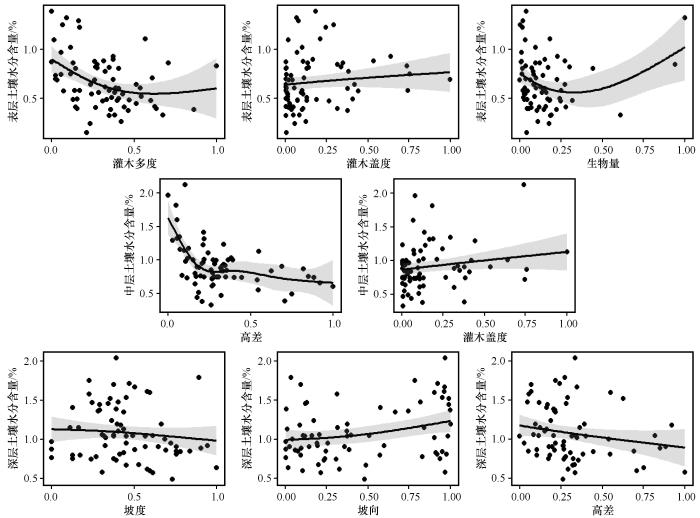
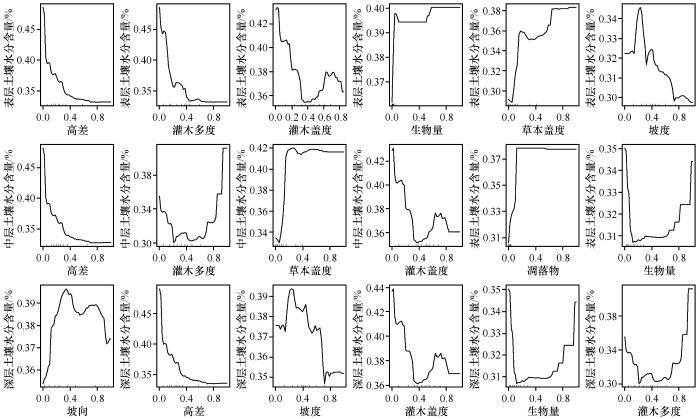
图6是基于RF模型的响应曲线。随着地形因子高差的增加,不同深度土壤水分含量均呈减少趋势;表层土壤水分含量随着灌木多度的增加呈减少趋势,但随着草本盖度和生物量的增加,表层土壤水分含量呈增加趋势;中层土壤水分含量随着草本盖度和凋落物的增加呈增加趋势,随着灌木多度和生物量的增加呈先减少后增加的趋势;对深层而言,随着生物量和灌木多度的增加,土壤水分含量呈先减少后增加的趋势。
图6
3.5 模型评价
表4为GLM、GAM和RF模型的评价结果,评价指标采用决定系数R2和均方根误差RMSE。其中,决定系数R2越接近1表明模型的拟合效果越好。RMSE用于衡量模型的预测误差,在RMSE尽量减小的情况下,R2越大说明模型的拟合效果越好。
表4 模型的评价
Table 4
| 深度 | R2 | RMSE | ||||
|---|---|---|---|---|---|---|
| GLM | GAM | RF | GLM | GAM | RF | |
| 表层 | 0.274 | 0.383 | 0.82 | 0.197 | 0.02 | 0.11 |
| 中层 | 0.34 | 0.379 | 0.902 | 0.159 | 0.017 | 0.086 |
| 深层 | 0.194 | 0.391 | 0.599 | 0.213 | 0.02 | 0.146 |
不同土层上,GLM模型的均方根误差最大,GAM模型和RF模型的均方根误差较小。R2顺序为RF>GAM>GLM,且RF模型的R2在表层和中层上都达到0.8以上。因此,3种模型中,RF模型精度最优,GLM模型精度最小。
4 讨论
4.1 不同深度土壤水分的分布特征
不同深度的土层中,表层的土壤水分含量最低(均值为0.662%),这可能是由于沙漠中降雨较少,其表层通常为干沙层,表层的土壤水分含量均值最低。同时,表层土壤水分含量在很大程度上受到降雨、蒸发的影响,导致土壤水分含量的变异增大[22]。研究区主要固沙灌木的根系长度大多为40~150 cm,40~200 cm部分的中层土壤水分大多会被灌木根系所吸收[23],土壤水分被大量利用。同时,研究区的降水以小于5 mm的降水为主,降雨入渗减少也会导致较低的中层土壤水分含量。在土壤深层,蒸发的影响相对较弱,较厚的覆盖物会增加降雨入渗与储存,植被的根系对深层的土壤水分吸收相对较少。因此,深层土壤水分含量随着土壤深度的增加而增加或保持不变。同时,深层土壤水分含量受到外界因素影响较小,因此其土壤水分含量最高且波动变异的幅度也会较小。
4.2 地形、植被因子对土壤水分含量的影响
在干旱半干旱沙区,植物生长和繁殖都需要水,土壤水资源是荒漠化防治的必要条件[27],然而,植被对沙丘上土壤水分的分布和变化起着重要的作用。植物的蒸腾作用、根系吸水等都可改变土壤水分的分布特征及变化规律。本研究表明,植被因子中灌木多度、灌木盖度与草本盖度对不同深度土壤水分含量均有显著影响。
GLM、GAM和RF模型均给出了对土壤水分含量有显著影响的地形、植被因子。随着高差的增加,不同深度土壤水分含量呈现出下降趋势。高差和坡度会影响土壤水分垂直和侧向流动,较低的位置往往有较高的土壤水分含量。陡坡也比缓坡具有更高的土壤水分含量[28]。植被因子灌木多度、灌木盖度对表层土壤水分含量有负向影响。与根系较短的草本植物不同,灌木根系消耗更多的土壤水分,越多的灌木越容易导致土壤缺水干旱[29]。凋落物对中层土壤的持水能力呈现出正向的影响,凋落物的增加会减少地表径流的流失,使得土壤的持水能力增强[30]。值得注意的是,在RF模型得到的结果中,灌木多度与对不同深度的土壤水分含量影响呈现出了先减少后增加的趋势。在固沙植被建立初期,植被覆盖度低,根系主要分布在表层与中层范围内,且生物土壤结皮层很薄,其对表层与中层的土壤水分消耗较大[31]。随着灌木多度的增加,中层、深层以及整体的土壤水分含量都呈现出增长趋势,灌木多度增加可以增加对降水的储存和入渗,使中层以下深度的土壤水分得到了一定补给并维持在稳定的范围内。
在气候条件一致的区域,低海拔或陡坡往往具有更高的土壤含水量。因此,可以在这些区域种植水分含量消耗相对较高的固沙灌木。草本植被可种植在地势平坦、土壤水分含量较低的区域。中层土壤水分含量随着草本盖度的增加呈现出增加趋势,因此可以适当增加草本植被的种植密度,以保持并增加土壤水分。对于植被恢复可以在原生植物的环境中进行改良,利用草方格固沙[32],综合考虑局部区域的环境条件,并选择合适的种植位置进行植被重建。乔木或灌木有可能会导致土壤水分加剧流失,因此可以适当地增加草本植被和消耗水分较小的灌木植被的种植密度,控制并降低水分消耗高的乔木及灌木的种植密度。
5 结论
本研究以腾格里沙漠天然植被区固定沙丘上的土壤水分含量为研究对象,采用描述性统计将0~300 cm土层划分为表层、中层和深层土壤。然后利用GLM、GAM和RF模型研究了不同深度土壤水分含量与地形、植被因子之间的关系。
根据土壤水分含量的分布特征,将0~300 cm土层分为表层(0~40 cm)、中层(40~200 cm)和深层(200~300 cm),不同深度土壤水分含量的大小顺序为深层>中层>表层,且不同深度土壤水分含量之间存在显著差异。
采用决定系数R2和均方根误差RMSE对GLM、GAM和RF模型进行评价,RF模型的精度最优。
精度最高的RF模型的结果表明,不同深度土壤水分含量均随着地形因子高差的增加而不断减少。表层和中层土壤水分含量随草本盖度增加而逐渐增加,中层和深层土壤水分含量随着灌木多度和生物量的增加均呈现出先减少后增加的趋势。
参考文献
On the origin of deep soil water infiltration in the arid sandy region of China
[J].
Variations of deep soil moisture under different vegetation types andinfluencing factors in a watershed of the Loess Plateau,China
[J].
Ecohydrological advances and applications in plant-water relations research:a review
[J].
Ecohydrology of terrestrial ecosystems
[J].
Species‐richness patterns of vascular plants along seven altitudinal transects in Norway
[J].
The patterns and causes of elevational diversity gradients
[J].
Effect of soil microtopography on seed bank distribution in the shrub-steppe
[J].
Mima mound microtopography and vegetation pattern in Kenyan savannas
[J].
Geographically comprehensive assessment of salt-meadow vegetation-elevation relations using LiDAR
[J].
Species richness in boreal swamp forests of SE Norway: the role of surface microtopography
[J].
On the role of microenvironmental heterogeneity in the ecology and diversification of neotropical rain-forest palms (Arecaceae)
[J].
Fine-scale pattern of the boundary zones in alkaline grassland communities
[J].
Generalized linear models with examples
[J].
On the relationship between mean and variance of soil moisture fields
[J].
基于Hydrus-1D模型的毛乌素沙地杨柴(Hedysarum laeve)灌木林土壤含水量模拟
[J].

 甘公网安备 62010202000688号
甘公网安备 62010202000688号
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}