Long‐term ecosystem effects of sand‐binding vegetation in the Tengger Desert,northern China
1
2004
... 在干旱和半干旱地区,植被恢复被认为是控制荒漠化、促进生态恢复的有效方式[1].为遏制荒漠化,中国政府自20世纪50年代起实施了一系列生态建设项目[2].其中,“三北”防护林体系建设工程在水土保持、防风固沙以及调节气候等方面发挥了重要的环境修复作用[3].沙漠灌木对极端环境有强大适应性,并在改变地表风向、提高土壤肥力等方面发挥着积极作用,经常被用作旱区生态恢复的先锋物种[4].然而,近年来全球气候变化导致干旱频发,引发了大面积木本植物枯死,对生态系统格局和过程产生了严重影响[5].受水资源短缺和气候变化的影响,内蒙古中部地区、新疆西北地区以及陕北地区出现了冠层枯萎甚至死亡等退化现象[6].因此,研究荒漠植物在干旱胁迫环境下的生理、形态和解剖结构响应与适应机制,有助于了解植物在不断变化环境中的耐旱性和适应性,为预测荒漠植物对全球气候变化的应对能力提供理论依据,对荒漠植物保护和指导植被恢复实践具有重要意义. ...
Assessment on forest carbon sequestration in the Three-North Shelterbelt Program region,China
1
2019
... 在干旱和半干旱地区,植被恢复被认为是控制荒漠化、促进生态恢复的有效方式[1].为遏制荒漠化,中国政府自20世纪50年代起实施了一系列生态建设项目[2].其中,“三北”防护林体系建设工程在水土保持、防风固沙以及调节气候等方面发挥了重要的环境修复作用[3].沙漠灌木对极端环境有强大适应性,并在改变地表风向、提高土壤肥力等方面发挥着积极作用,经常被用作旱区生态恢复的先锋物种[4].然而,近年来全球气候变化导致干旱频发,引发了大面积木本植物枯死,对生态系统格局和过程产生了严重影响[5].受水资源短缺和气候变化的影响,内蒙古中部地区、新疆西北地区以及陕北地区出现了冠层枯萎甚至死亡等退化现象[6].因此,研究荒漠植物在干旱胁迫环境下的生理、形态和解剖结构响应与适应机制,有助于了解植物在不断变化环境中的耐旱性和适应性,为预测荒漠植物对全球气候变化的应对能力提供理论依据,对荒漠植物保护和指导植被恢复实践具有重要意义. ...
Assessment of future potential carbon sequestration and water consumption in the construction area of the Three-North Shelterbelt Programme in China
1
2021
... 在干旱和半干旱地区,植被恢复被认为是控制荒漠化、促进生态恢复的有效方式[1].为遏制荒漠化,中国政府自20世纪50年代起实施了一系列生态建设项目[2].其中,“三北”防护林体系建设工程在水土保持、防风固沙以及调节气候等方面发挥了重要的环境修复作用[3].沙漠灌木对极端环境有强大适应性,并在改变地表风向、提高土壤肥力等方面发挥着积极作用,经常被用作旱区生态恢复的先锋物种[4].然而,近年来全球气候变化导致干旱频发,引发了大面积木本植物枯死,对生态系统格局和过程产生了严重影响[5].受水资源短缺和气候变化的影响,内蒙古中部地区、新疆西北地区以及陕北地区出现了冠层枯萎甚至死亡等退化现象[6].因此,研究荒漠植物在干旱胁迫环境下的生理、形态和解剖结构响应与适应机制,有助于了解植物在不断变化环境中的耐旱性和适应性,为预测荒漠植物对全球气候变化的应对能力提供理论依据,对荒漠植物保护和指导植被恢复实践具有重要意义. ...
Responses of different herb life-history groups to a dominant shrub species along a dune stabilization gradient
1
2019
... 在干旱和半干旱地区,植被恢复被认为是控制荒漠化、促进生态恢复的有效方式[1].为遏制荒漠化,中国政府自20世纪50年代起实施了一系列生态建设项目[2].其中,“三北”防护林体系建设工程在水土保持、防风固沙以及调节气候等方面发挥了重要的环境修复作用[3].沙漠灌木对极端环境有强大适应性,并在改变地表风向、提高土壤肥力等方面发挥着积极作用,经常被用作旱区生态恢复的先锋物种[4].然而,近年来全球气候变化导致干旱频发,引发了大面积木本植物枯死,对生态系统格局和过程产生了严重影响[5].受水资源短缺和气候变化的影响,内蒙古中部地区、新疆西北地区以及陕北地区出现了冠层枯萎甚至死亡等退化现象[6].因此,研究荒漠植物在干旱胁迫环境下的生理、形态和解剖结构响应与适应机制,有助于了解植物在不断变化环境中的耐旱性和适应性,为预测荒漠植物对全球气候变化的应对能力提供理论依据,对荒漠植物保护和指导植被恢复实践具有重要意义. ...
Darcy's law predicts widespread forest mortality under climate warming
1
2015
... 在干旱和半干旱地区,植被恢复被认为是控制荒漠化、促进生态恢复的有效方式[1].为遏制荒漠化,中国政府自20世纪50年代起实施了一系列生态建设项目[2].其中,“三北”防护林体系建设工程在水土保持、防风固沙以及调节气候等方面发挥了重要的环境修复作用[3].沙漠灌木对极端环境有强大适应性,并在改变地表风向、提高土壤肥力等方面发挥着积极作用,经常被用作旱区生态恢复的先锋物种[4].然而,近年来全球气候变化导致干旱频发,引发了大面积木本植物枯死,对生态系统格局和过程产生了严重影响[5].受水资源短缺和气候变化的影响,内蒙古中部地区、新疆西北地区以及陕北地区出现了冠层枯萎甚至死亡等退化现象[6].因此,研究荒漠植物在干旱胁迫环境下的生理、形态和解剖结构响应与适应机制,有助于了解植物在不断变化环境中的耐旱性和适应性,为预测荒漠植物对全球气候变化的应对能力提供理论依据,对荒漠植物保护和指导植被恢复实践具有重要意义. ...
Detecting forest degradation in the three-north forest shelterbelt in China from multi-scale satellite images
1
2021
... 在干旱和半干旱地区,植被恢复被认为是控制荒漠化、促进生态恢复的有效方式[1].为遏制荒漠化,中国政府自20世纪50年代起实施了一系列生态建设项目[2].其中,“三北”防护林体系建设工程在水土保持、防风固沙以及调节气候等方面发挥了重要的环境修复作用[3].沙漠灌木对极端环境有强大适应性,并在改变地表风向、提高土壤肥力等方面发挥着积极作用,经常被用作旱区生态恢复的先锋物种[4].然而,近年来全球气候变化导致干旱频发,引发了大面积木本植物枯死,对生态系统格局和过程产生了严重影响[5].受水资源短缺和气候变化的影响,内蒙古中部地区、新疆西北地区以及陕北地区出现了冠层枯萎甚至死亡等退化现象[6].因此,研究荒漠植物在干旱胁迫环境下的生理、形态和解剖结构响应与适应机制,有助于了解植物在不断变化环境中的耐旱性和适应性,为预测荒漠植物对全球气候变化的应对能力提供理论依据,对荒漠植物保护和指导植被恢复实践具有重要意义. ...
植物水分生理研究中的压力-容积测定技术几个关键问题
1
2014
... 在干旱胁迫环境下,植物通过调整其形态、结构特征及各器官的生长速度来应对不利条件.叶片作为植物进行光合作用与水气交换的重要器官,其结构的变化能够反映植物对逆境的适应能力.PV曲线水分参数通过压力-容积(Pressure-Volume, PV)技术获得,反映植物组织在干旱条件下维持细胞膨压的能力,与植物的耐旱性密切相关[7].此外,叶片的解剖特征作为研究植物内部结构和细胞组织的重要手段,体现了植物在逆境条件下的适应能力.这些解剖特征通常具有较强的可塑性,能够适应不断变化的环境[8-9].C3植物和C4植物在CO2固定途径、生理结构和光合效率等方面存在差异.这两类植物适应温度和水分的能力也不同.相较于C3植物,C4植物具有更高的光能利用效率和较低的光呼吸速率,在高温和高光照条件下具有更高的生长和光合作用效率[10].水分特征与光合作用过程之间存在着协同与权衡关系[11-12].目前关于沙生灌木的研究较少[13]. ...
干旱胁迫对绢毛委陵菜叶片解剖结构和生理指标的影响
1
2012
... 在干旱胁迫环境下,植物通过调整其形态、结构特征及各器官的生长速度来应对不利条件.叶片作为植物进行光合作用与水气交换的重要器官,其结构的变化能够反映植物对逆境的适应能力.PV曲线水分参数通过压力-容积(Pressure-Volume, PV)技术获得,反映植物组织在干旱条件下维持细胞膨压的能力,与植物的耐旱性密切相关[7].此外,叶片的解剖特征作为研究植物内部结构和细胞组织的重要手段,体现了植物在逆境条件下的适应能力.这些解剖特征通常具有较强的可塑性,能够适应不断变化的环境[8-9].C3植物和C4植物在CO2固定途径、生理结构和光合效率等方面存在差异.这两类植物适应温度和水分的能力也不同.相较于C3植物,C4植物具有更高的光能利用效率和较低的光呼吸速率,在高温和高光照条件下具有更高的生长和光合作用效率[10].水分特征与光合作用过程之间存在着协同与权衡关系[11-12].目前关于沙生灌木的研究较少[13]. ...
盐胁迫对无芒雀麦幼苗叶片形态及解剖结构的影响
1
2022
... 在干旱胁迫环境下,植物通过调整其形态、结构特征及各器官的生长速度来应对不利条件.叶片作为植物进行光合作用与水气交换的重要器官,其结构的变化能够反映植物对逆境的适应能力.PV曲线水分参数通过压力-容积(Pressure-Volume, PV)技术获得,反映植物组织在干旱条件下维持细胞膨压的能力,与植物的耐旱性密切相关[7].此外,叶片的解剖特征作为研究植物内部结构和细胞组织的重要手段,体现了植物在逆境条件下的适应能力.这些解剖特征通常具有较强的可塑性,能够适应不断变化的环境[8-9].C3植物和C4植物在CO2固定途径、生理结构和光合效率等方面存在差异.这两类植物适应温度和水分的能力也不同.相较于C3植物,C4植物具有更高的光能利用效率和较低的光呼吸速率,在高温和高光照条件下具有更高的生长和光合作用效率[10].水分特征与光合作用过程之间存在着协同与权衡关系[11-12].目前关于沙生灌木的研究较少[13]. ...
C3 and C4 photosynthesis
1
2002
... 在干旱胁迫环境下,植物通过调整其形态、结构特征及各器官的生长速度来应对不利条件.叶片作为植物进行光合作用与水气交换的重要器官,其结构的变化能够反映植物对逆境的适应能力.PV曲线水分参数通过压力-容积(Pressure-Volume, PV)技术获得,反映植物组织在干旱条件下维持细胞膨压的能力,与植物的耐旱性密切相关[7].此外,叶片的解剖特征作为研究植物内部结构和细胞组织的重要手段,体现了植物在逆境条件下的适应能力.这些解剖特征通常具有较强的可塑性,能够适应不断变化的环境[8-9].C3植物和C4植物在CO2固定途径、生理结构和光合效率等方面存在差异.这两类植物适应温度和水分的能力也不同.相较于C3植物,C4植物具有更高的光能利用效率和较低的光呼吸速率,在高温和高光照条件下具有更高的生长和光合作用效率[10].水分特征与光合作用过程之间存在着协同与权衡关系[11-12].目前关于沙生灌木的研究较少[13]. ...
Hydraulic and photosynthetic co-ordination in seasonally dry tropical forest trees
1
2002
... 在干旱胁迫环境下,植物通过调整其形态、结构特征及各器官的生长速度来应对不利条件.叶片作为植物进行光合作用与水气交换的重要器官,其结构的变化能够反映植物对逆境的适应能力.PV曲线水分参数通过压力-容积(Pressure-Volume, PV)技术获得,反映植物组织在干旱条件下维持细胞膨压的能力,与植物的耐旱性密切相关[7].此外,叶片的解剖特征作为研究植物内部结构和细胞组织的重要手段,体现了植物在逆境条件下的适应能力.这些解剖特征通常具有较强的可塑性,能够适应不断变化的环境[8-9].C3植物和C4植物在CO2固定途径、生理结构和光合效率等方面存在差异.这两类植物适应温度和水分的能力也不同.相较于C3植物,C4植物具有更高的光能利用效率和较低的光呼吸速率,在高温和高光照条件下具有更高的生长和光合作用效率[10].水分特征与光合作用过程之间存在着协同与权衡关系[11-12].目前关于沙生灌木的研究较少[13]. ...
Photosynthetic pathway influences xylem structure and function in Flaveria (Asteraceae)
1
2008
... 在干旱胁迫环境下,植物通过调整其形态、结构特征及各器官的生长速度来应对不利条件.叶片作为植物进行光合作用与水气交换的重要器官,其结构的变化能够反映植物对逆境的适应能力.PV曲线水分参数通过压力-容积(Pressure-Volume, PV)技术获得,反映植物组织在干旱条件下维持细胞膨压的能力,与植物的耐旱性密切相关[7].此外,叶片的解剖特征作为研究植物内部结构和细胞组织的重要手段,体现了植物在逆境条件下的适应能力.这些解剖特征通常具有较强的可塑性,能够适应不断变化的环境[8-9].C3植物和C4植物在CO2固定途径、生理结构和光合效率等方面存在差异.这两类植物适应温度和水分的能力也不同.相较于C3植物,C4植物具有更高的光能利用效率和较低的光呼吸速率,在高温和高光照条件下具有更高的生长和光合作用效率[10].水分特征与光合作用过程之间存在着协同与权衡关系[11-12].目前关于沙生灌木的研究较少[13]. ...
Life history is a key factor explaining functional trait diversity among subtropical grasses,and its influence differs between C3 and C4 species
1
2019
... 在干旱胁迫环境下,植物通过调整其形态、结构特征及各器官的生长速度来应对不利条件.叶片作为植物进行光合作用与水气交换的重要器官,其结构的变化能够反映植物对逆境的适应能力.PV曲线水分参数通过压力-容积(Pressure-Volume, PV)技术获得,反映植物组织在干旱条件下维持细胞膨压的能力,与植物的耐旱性密切相关[7].此外,叶片的解剖特征作为研究植物内部结构和细胞组织的重要手段,体现了植物在逆境条件下的适应能力.这些解剖特征通常具有较强的可塑性,能够适应不断变化的环境[8-9].C3植物和C4植物在CO2固定途径、生理结构和光合效率等方面存在差异.这两类植物适应温度和水分的能力也不同.相较于C3植物,C4植物具有更高的光能利用效率和较低的光呼吸速率,在高温和高光照条件下具有更高的生长和光合作用效率[10].水分特征与光合作用过程之间存在着协同与权衡关系[11-12].目前关于沙生灌木的研究较少[13]. ...
荒漠植物红砂根系对干旱胁迫的生理响应
1
2015
... 荒漠植物形态和生理特征的变化是长期适应干旱环境的结果.众多学者围绕荒漠植物在干旱胁迫下的形态和生理等方面进行了一系列研究[14-15],然而,关于不同光合作用途径的沙漠灌木如何通过水分适应策略和解剖特征适应干旱环境的机制仍不清楚.本研究以乌兰布和沙漠10种植物为研究对象,并依据光合作用途径的不同,将植物划分为C3和C4植物.通过测定叶片的PV曲线水分参数、解剖和结构特征,将这些参数相结合,探究植物水分调节、生理适应性以及解剖结构与功能之间的关系,为深入理解荒漠植物的生理机制、适应性和环境响应提供理论参考. ...
干旱胁迫对疏叶骆驼刺幼苗生长和生理的影响
1
2023
... 荒漠植物形态和生理特征的变化是长期适应干旱环境的结果.众多学者围绕荒漠植物在干旱胁迫下的形态和生理等方面进行了一系列研究[14-15],然而,关于不同光合作用途径的沙漠灌木如何通过水分适应策略和解剖特征适应干旱环境的机制仍不清楚.本研究以乌兰布和沙漠10种植物为研究对象,并依据光合作用途径的不同,将植物划分为C3和C4植物.通过测定叶片的PV曲线水分参数、解剖和结构特征,将这些参数相结合,探究植物水分调节、生理适应性以及解剖结构与功能之间的关系,为深入理解荒漠植物的生理机制、适应性和环境响应提供理论参考. ...
Water relations of drought hardy shrubs:osmotic potential and stomatal reactivity
1
1980
... 黎明前从每种植物中收集了4根10 cm长的阳生带叶小枝,放入装有湿滤纸的塑封袋中,再放入冷藏箱中带回实验室,置于蒸馏水中并在避光条件下水化30~120 min,直到完全饱和.PV曲线采用自然蒸腾台架干燥法获得[16-17].在一天中(通常在7~10 h内)反复测量嫩枝水势(Ψ)和重量,直到-1/Ψ与叶片水质量损失的图形成线性关系.PV曲线是以叶片水分饱和亏缺(1-RWC)为x轴,叶片水势的倒数(1/Ψ)为y轴建立关系图[18],并推导PV性状:失膨点水势(Ψtlp)、饱和含水时的渗透势(Ψ0)和质壁分离时的相对含水量(RWCtlp).叶水容(Cleaf)参考Tyree等[19]的方法得出.细胞弹性模量(ε)被估计为膨压损失前PV曲线初始部分上膨压与RWC之间回归的斜率[20]. ...
Measurement of plant water status by the pressure chamber technique
1
1988
... 黎明前从每种植物中收集了4根10 cm长的阳生带叶小枝,放入装有湿滤纸的塑封袋中,再放入冷藏箱中带回实验室,置于蒸馏水中并在避光条件下水化30~120 min,直到完全饱和.PV曲线采用自然蒸腾台架干燥法获得[16-17].在一天中(通常在7~10 h内)反复测量嫩枝水势(Ψ)和重量,直到-1/Ψ与叶片水质量损失的图形成线性关系.PV曲线是以叶片水分饱和亏缺(1-RWC)为x轴,叶片水势的倒数(1/Ψ)为y轴建立关系图[18],并推导PV性状:失膨点水势(Ψtlp)、饱和含水时的渗透势(Ψ0)和质壁分离时的相对含水量(RWCtlp).叶水容(Cleaf)参考Tyree等[19]的方法得出.细胞弹性模量(ε)被估计为膨压损失前PV曲线初始部分上膨压与RWC之间回归的斜率[20]. ...
Dynamics of leaf water relations components in co-occurring iso-and anisohydric conifer species
1
2014
... 黎明前从每种植物中收集了4根10 cm长的阳生带叶小枝,放入装有湿滤纸的塑封袋中,再放入冷藏箱中带回实验室,置于蒸馏水中并在避光条件下水化30~120 min,直到完全饱和.PV曲线采用自然蒸腾台架干燥法获得[16-17].在一天中(通常在7~10 h内)反复测量嫩枝水势(Ψ)和重量,直到-1/Ψ与叶片水质量损失的图形成线性关系.PV曲线是以叶片水分饱和亏缺(1-RWC)为x轴,叶片水势的倒数(1/Ψ)为y轴建立关系图[18],并推导PV性状:失膨点水势(Ψtlp)、饱和含水时的渗透势(Ψ0)和质壁分离时的相对含水量(RWCtlp).叶水容(Cleaf)参考Tyree等[19]的方法得出.细胞弹性模量(ε)被估计为膨压损失前PV曲线初始部分上膨压与RWC之间回归的斜率[20]. ...
The measurement of the turgor pressure and the water relations of plants by the pressure-bomb technique
1
1972
... 黎明前从每种植物中收集了4根10 cm长的阳生带叶小枝,放入装有湿滤纸的塑封袋中,再放入冷藏箱中带回实验室,置于蒸馏水中并在避光条件下水化30~120 min,直到完全饱和.PV曲线采用自然蒸腾台架干燥法获得[16-17].在一天中(通常在7~10 h内)反复测量嫩枝水势(Ψ)和重量,直到-1/Ψ与叶片水质量损失的图形成线性关系.PV曲线是以叶片水分饱和亏缺(1-RWC)为x轴,叶片水势的倒数(1/Ψ)为y轴建立关系图[18],并推导PV性状:失膨点水势(Ψtlp)、饱和含水时的渗透势(Ψ0)和质壁分离时的相对含水量(RWCtlp).叶水容(Cleaf)参考Tyree等[19]的方法得出.细胞弹性模量(ε)被估计为膨压损失前PV曲线初始部分上膨压与RWC之间回归的斜率[20]. ...
The determinants of leaf turgor loss point and prediction of drought tolerance of species and biomes:a global meta‐analysis
1
2012
... 黎明前从每种植物中收集了4根10 cm长的阳生带叶小枝,放入装有湿滤纸的塑封袋中,再放入冷藏箱中带回实验室,置于蒸馏水中并在避光条件下水化30~120 min,直到完全饱和.PV曲线采用自然蒸腾台架干燥法获得[16-17].在一天中(通常在7~10 h内)反复测量嫩枝水势(Ψ)和重量,直到-1/Ψ与叶片水质量损失的图形成线性关系.PV曲线是以叶片水分饱和亏缺(1-RWC)为x轴,叶片水势的倒数(1/Ψ)为y轴建立关系图[18],并推导PV性状:失膨点水势(Ψtlp)、饱和含水时的渗透势(Ψ0)和质壁分离时的相对含水量(RWCtlp).叶水容(Cleaf)参考Tyree等[19]的方法得出.细胞弹性模量(ε)被估计为膨压损失前PV曲线初始部分上膨压与RWC之间回归的斜率[20]. ...
压力室 (PRESSURE CHAMBER) 在植物水分状况研究中的应用
1
1984
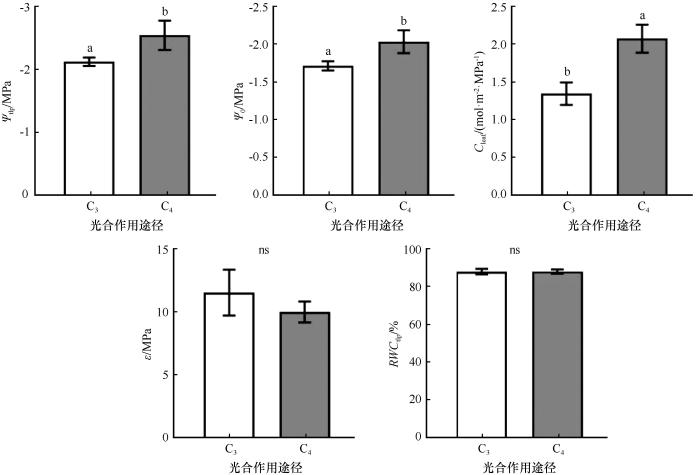
... PV技术作为研究树木耐旱性的重要手段之一,已广泛应用于植物水分生理研究[21-23].通过PV曲线求得的水分参数,对了解植物组织内部水分状况及渗透调节具有重要意义.其中,Ψtlp反映了植物维持最低膨压的极限渗透势,在耐旱性较强的植物中通常表现为较低的值(更负),被认为是衡量植物耐旱性的关键指标[24-26],在水分为主要限制因子的干旱、半干旱地区有重要意义.本研究中,Ψtlp和Ψ0的变化范围分别为-3.44~-1.52 MPa和-2.66~-1.33 MPa.Bartlett等[24]对72项研究中的317种植物进行了Meta分析,结果显示,全球半荒漠地区植物的Ψtlp和Ψ0的变化范围分别为-1.25~-4.25 MPa和-0.96~-2.78 MPa,本研究中Ψtlp和Ψ0值均在此范围内.斯琴巴特尔等[27]利用PV技术研究了荒漠区植物蒙古扁桃的水分生理特性,结果表明其Ψtlp为-3.11 MPa;李向义等[23]对沙漠-绿洲过渡带的4种植物进行水分关系研究,发现Ψtlp的变化范围为-1.50~-2.41 MPa;王乐乐等[28]通过对东北温带森林中20种生长于半湿润区的乔木进行PV曲线性状测定,结果显示Ψtlp平均值为-1.2 MPa.可以看出,从荒漠、沙漠-绿洲过渡带到半湿润区,Ψtlp随着水分梯度的增加而增加(绝对值更小).在干旱区,Ψtlp通常被认为具有较低值[29].Zhu等[25]汇编了中国9个木本植物群落中389种木本植物的Ψtlp数据集,发现沙漠灌木林带的Ψtlp为-1.52~-2.71 MPa,本研究中除梭梭、霸王外,其余植物Ψtlp值均在该范围内.梭梭与霸王具有更负的Ψtlp,表明其耐旱能力较强于其他树种.Xu等[30]研究了梭梭对水分条件变化的适应性,认为梭梭具有高效的形态调节和强大的气孔控制能力,有助于其适应环境的变化.石松利等[31]对霸王水分关系参数的研究表明,霸王主要通过增加细胞质浓度的渗透调节来维持膨压,显示出较强的耐旱性. ...
PV技术在油松侧柏苗木抗旱特性研究中的应用
0
1989
沙漠-绿洲过渡带四种多年生植物水分关系特征
2
2004
... PV技术作为研究树木耐旱性的重要手段之一,已广泛应用于植物水分生理研究[21-23].通过PV曲线求得的水分参数,对了解植物组织内部水分状况及渗透调节具有重要意义.其中,Ψtlp反映了植物维持最低膨压的极限渗透势,在耐旱性较强的植物中通常表现为较低的值(更负),被认为是衡量植物耐旱性的关键指标[24-26],在水分为主要限制因子的干旱、半干旱地区有重要意义.本研究中,Ψtlp和Ψ0的变化范围分别为-3.44~-1.52 MPa和-2.66~-1.33 MPa.Bartlett等[24]对72项研究中的317种植物进行了Meta分析,结果显示,全球半荒漠地区植物的Ψtlp和Ψ0的变化范围分别为-1.25~-4.25 MPa和-0.96~-2.78 MPa,本研究中Ψtlp和Ψ0值均在此范围内.斯琴巴特尔等[27]利用PV技术研究了荒漠区植物蒙古扁桃的水分生理特性,结果表明其Ψtlp为-3.11 MPa;李向义等[23]对沙漠-绿洲过渡带的4种植物进行水分关系研究,发现Ψtlp的变化范围为-1.50~-2.41 MPa;王乐乐等[28]通过对东北温带森林中20种生长于半湿润区的乔木进行PV曲线性状测定,结果显示Ψtlp平均值为-1.2 MPa.可以看出,从荒漠、沙漠-绿洲过渡带到半湿润区,Ψtlp随着水分梯度的增加而增加(绝对值更小).在干旱区,Ψtlp通常被认为具有较低值[29].Zhu等[25]汇编了中国9个木本植物群落中389种木本植物的Ψtlp数据集,发现沙漠灌木林带的Ψtlp为-1.52~-2.71 MPa,本研究中除梭梭、霸王外,其余植物Ψtlp值均在该范围内.梭梭与霸王具有更负的Ψtlp,表明其耐旱能力较强于其他树种.Xu等[30]研究了梭梭对水分条件变化的适应性,认为梭梭具有高效的形态调节和强大的气孔控制能力,有助于其适应环境的变化.石松利等[31]对霸王水分关系参数的研究表明,霸王主要通过增加细胞质浓度的渗透调节来维持膨压,显示出较强的耐旱性. ...
... [23]对沙漠-绿洲过渡带的4种植物进行水分关系研究,发现Ψtlp的变化范围为-1.50~-2.41 MPa;王乐乐等[28]通过对东北温带森林中20种生长于半湿润区的乔木进行PV曲线性状测定,结果显示Ψtlp平均值为-1.2 MPa.可以看出,从荒漠、沙漠-绿洲过渡带到半湿润区,Ψtlp随着水分梯度的增加而增加(绝对值更小).在干旱区,Ψtlp通常被认为具有较低值[29].Zhu等[25]汇编了中国9个木本植物群落中389种木本植物的Ψtlp数据集,发现沙漠灌木林带的Ψtlp为-1.52~-2.71 MPa,本研究中除梭梭、霸王外,其余植物Ψtlp值均在该范围内.梭梭与霸王具有更负的Ψtlp,表明其耐旱能力较强于其他树种.Xu等[30]研究了梭梭对水分条件变化的适应性,认为梭梭具有高效的形态调节和强大的气孔控制能力,有助于其适应环境的变化.石松利等[31]对霸王水分关系参数的研究表明,霸王主要通过增加细胞质浓度的渗透调节来维持膨压,显示出较强的耐旱性. ...
Global analysis of plasticity in turgor loss point,a key drought tolerance trait
2
2014
... PV技术作为研究树木耐旱性的重要手段之一,已广泛应用于植物水分生理研究[21-23].通过PV曲线求得的水分参数,对了解植物组织内部水分状况及渗透调节具有重要意义.其中,Ψtlp反映了植物维持最低膨压的极限渗透势,在耐旱性较强的植物中通常表现为较低的值(更负),被认为是衡量植物耐旱性的关键指标[24-26],在水分为主要限制因子的干旱、半干旱地区有重要意义.本研究中,Ψtlp和Ψ0的变化范围分别为-3.44~-1.52 MPa和-2.66~-1.33 MPa.Bartlett等[24]对72项研究中的317种植物进行了Meta分析,结果显示,全球半荒漠地区植物的Ψtlp和Ψ0的变化范围分别为-1.25~-4.25 MPa和-0.96~-2.78 MPa,本研究中Ψtlp和Ψ0值均在此范围内.斯琴巴特尔等[27]利用PV技术研究了荒漠区植物蒙古扁桃的水分生理特性,结果表明其Ψtlp为-3.11 MPa;李向义等[23]对沙漠-绿洲过渡带的4种植物进行水分关系研究,发现Ψtlp的变化范围为-1.50~-2.41 MPa;王乐乐等[28]通过对东北温带森林中20种生长于半湿润区的乔木进行PV曲线性状测定,结果显示Ψtlp平均值为-1.2 MPa.可以看出,从荒漠、沙漠-绿洲过渡带到半湿润区,Ψtlp随着水分梯度的增加而增加(绝对值更小).在干旱区,Ψtlp通常被认为具有较低值[29].Zhu等[25]汇编了中国9个木本植物群落中389种木本植物的Ψtlp数据集,发现沙漠灌木林带的Ψtlp为-1.52~-2.71 MPa,本研究中除梭梭、霸王外,其余植物Ψtlp值均在该范围内.梭梭与霸王具有更负的Ψtlp,表明其耐旱能力较强于其他树种.Xu等[30]研究了梭梭对水分条件变化的适应性,认为梭梭具有高效的形态调节和强大的气孔控制能力,有助于其适应环境的变化.石松利等[31]对霸王水分关系参数的研究表明,霸王主要通过增加细胞质浓度的渗透调节来维持膨压,显示出较强的耐旱性. ...
... [24]对72项研究中的317种植物进行了Meta分析,结果显示,全球半荒漠地区植物的Ψtlp和Ψ0的变化范围分别为-1.25~-4.25 MPa和-0.96~-2.78 MPa,本研究中Ψtlp和Ψ0值均在此范围内.斯琴巴特尔等[27]利用PV技术研究了荒漠区植物蒙古扁桃的水分生理特性,结果表明其Ψtlp为-3.11 MPa;李向义等[23]对沙漠-绿洲过渡带的4种植物进行水分关系研究,发现Ψtlp的变化范围为-1.50~-2.41 MPa;王乐乐等[28]通过对东北温带森林中20种生长于半湿润区的乔木进行PV曲线性状测定,结果显示Ψtlp平均值为-1.2 MPa.可以看出,从荒漠、沙漠-绿洲过渡带到半湿润区,Ψtlp随着水分梯度的增加而增加(绝对值更小).在干旱区,Ψtlp通常被认为具有较低值[29].Zhu等[25]汇编了中国9个木本植物群落中389种木本植物的Ψtlp数据集,发现沙漠灌木林带的Ψtlp为-1.52~-2.71 MPa,本研究中除梭梭、霸王外,其余植物Ψtlp值均在该范围内.梭梭与霸王具有更负的Ψtlp,表明其耐旱能力较强于其他树种.Xu等[30]研究了梭梭对水分条件变化的适应性,认为梭梭具有高效的形态调节和强大的气孔控制能力,有助于其适应环境的变化.石松利等[31]对霸王水分关系参数的研究表明,霸王主要通过增加细胞质浓度的渗透调节来维持膨压,显示出较强的耐旱性. ...
Leaf turgor loss point is correlated with drought tolerance and leaf carbon economics traits
1
2018
... PV技术作为研究树木耐旱性的重要手段之一,已广泛应用于植物水分生理研究[21-23].通过PV曲线求得的水分参数,对了解植物组织内部水分状况及渗透调节具有重要意义.其中,Ψtlp反映了植物维持最低膨压的极限渗透势,在耐旱性较强的植物中通常表现为较低的值(更负),被认为是衡量植物耐旱性的关键指标[24-26],在水分为主要限制因子的干旱、半干旱地区有重要意义.本研究中,Ψtlp和Ψ0的变化范围分别为-3.44~-1.52 MPa和-2.66~-1.33 MPa.Bartlett等[24]对72项研究中的317种植物进行了Meta分析,结果显示,全球半荒漠地区植物的Ψtlp和Ψ0的变化范围分别为-1.25~-4.25 MPa和-0.96~-2.78 MPa,本研究中Ψtlp和Ψ0值均在此范围内.斯琴巴特尔等[27]利用PV技术研究了荒漠区植物蒙古扁桃的水分生理特性,结果表明其Ψtlp为-3.11 MPa;李向义等[23]对沙漠-绿洲过渡带的4种植物进行水分关系研究,发现Ψtlp的变化范围为-1.50~-2.41 MPa;王乐乐等[28]通过对东北温带森林中20种生长于半湿润区的乔木进行PV曲线性状测定,结果显示Ψtlp平均值为-1.2 MPa.可以看出,从荒漠、沙漠-绿洲过渡带到半湿润区,Ψtlp随着水分梯度的增加而增加(绝对值更小).在干旱区,Ψtlp通常被认为具有较低值[29].Zhu等[25]汇编了中国9个木本植物群落中389种木本植物的Ψtlp数据集,发现沙漠灌木林带的Ψtlp为-1.52~-2.71 MPa,本研究中除梭梭、霸王外,其余植物Ψtlp值均在该范围内.梭梭与霸王具有更负的Ψtlp,表明其耐旱能力较强于其他树种.Xu等[30]研究了梭梭对水分条件变化的适应性,认为梭梭具有高效的形态调节和强大的气孔控制能力,有助于其适应环境的变化.石松利等[31]对霸王水分关系参数的研究表明,霸王主要通过增加细胞质浓度的渗透调节来维持膨压,显示出较强的耐旱性. ...
Leaf turgor loss as a predictor of plant drought response strategies
1
2018
... PV技术作为研究树木耐旱性的重要手段之一,已广泛应用于植物水分生理研究[21-23].通过PV曲线求得的水分参数,对了解植物组织内部水分状况及渗透调节具有重要意义.其中,Ψtlp反映了植物维持最低膨压的极限渗透势,在耐旱性较强的植物中通常表现为较低的值(更负),被认为是衡量植物耐旱性的关键指标[24-26],在水分为主要限制因子的干旱、半干旱地区有重要意义.本研究中,Ψtlp和Ψ0的变化范围分别为-3.44~-1.52 MPa和-2.66~-1.33 MPa.Bartlett等[24]对72项研究中的317种植物进行了Meta分析,结果显示,全球半荒漠地区植物的Ψtlp和Ψ0的变化范围分别为-1.25~-4.25 MPa和-0.96~-2.78 MPa,本研究中Ψtlp和Ψ0值均在此范围内.斯琴巴特尔等[27]利用PV技术研究了荒漠区植物蒙古扁桃的水分生理特性,结果表明其Ψtlp为-3.11 MPa;李向义等[23]对沙漠-绿洲过渡带的4种植物进行水分关系研究,发现Ψtlp的变化范围为-1.50~-2.41 MPa;王乐乐等[28]通过对东北温带森林中20种生长于半湿润区的乔木进行PV曲线性状测定,结果显示Ψtlp平均值为-1.2 MPa.可以看出,从荒漠、沙漠-绿洲过渡带到半湿润区,Ψtlp随着水分梯度的增加而增加(绝对值更小).在干旱区,Ψtlp通常被认为具有较低值[29].Zhu等[25]汇编了中国9个木本植物群落中389种木本植物的Ψtlp数据集,发现沙漠灌木林带的Ψtlp为-1.52~-2.71 MPa,本研究中除梭梭、霸王外,其余植物Ψtlp值均在该范围内.梭梭与霸王具有更负的Ψtlp,表明其耐旱能力较强于其他树种.Xu等[30]研究了梭梭对水分条件变化的适应性,认为梭梭具有高效的形态调节和强大的气孔控制能力,有助于其适应环境的变化.石松利等[31]对霸王水分关系参数的研究表明,霸王主要通过增加细胞质浓度的渗透调节来维持膨压,显示出较强的耐旱性. ...
荒漠植物蒙古扁桃水分生理特征
1
2007
... PV技术作为研究树木耐旱性的重要手段之一,已广泛应用于植物水分生理研究[21-23].通过PV曲线求得的水分参数,对了解植物组织内部水分状况及渗透调节具有重要意义.其中,Ψtlp反映了植物维持最低膨压的极限渗透势,在耐旱性较强的植物中通常表现为较低的值(更负),被认为是衡量植物耐旱性的关键指标[24-26],在水分为主要限制因子的干旱、半干旱地区有重要意义.本研究中,Ψtlp和Ψ0的变化范围分别为-3.44~-1.52 MPa和-2.66~-1.33 MPa.Bartlett等[24]对72项研究中的317种植物进行了Meta分析,结果显示,全球半荒漠地区植物的Ψtlp和Ψ0的变化范围分别为-1.25~-4.25 MPa和-0.96~-2.78 MPa,本研究中Ψtlp和Ψ0值均在此范围内.斯琴巴特尔等[27]利用PV技术研究了荒漠区植物蒙古扁桃的水分生理特性,结果表明其Ψtlp为-3.11 MPa;李向义等[23]对沙漠-绿洲过渡带的4种植物进行水分关系研究,发现Ψtlp的变化范围为-1.50~-2.41 MPa;王乐乐等[28]通过对东北温带森林中20种生长于半湿润区的乔木进行PV曲线性状测定,结果显示Ψtlp平均值为-1.2 MPa.可以看出,从荒漠、沙漠-绿洲过渡带到半湿润区,Ψtlp随着水分梯度的增加而增加(绝对值更小).在干旱区,Ψtlp通常被认为具有较低值[29].Zhu等[25]汇编了中国9个木本植物群落中389种木本植物的Ψtlp数据集,发现沙漠灌木林带的Ψtlp为-1.52~-2.71 MPa,本研究中除梭梭、霸王外,其余植物Ψtlp值均在该范围内.梭梭与霸王具有更负的Ψtlp,表明其耐旱能力较强于其他树种.Xu等[30]研究了梭梭对水分条件变化的适应性,认为梭梭具有高效的形态调节和强大的气孔控制能力,有助于其适应环境的变化.石松利等[31]对霸王水分关系参数的研究表明,霸王主要通过增加细胞质浓度的渗透调节来维持膨压,显示出较强的耐旱性. ...
东北温带森林20 种乔木树种叶片干旱容忍性特征
1
2022
... PV技术作为研究树木耐旱性的重要手段之一,已广泛应用于植物水分生理研究[21-23].通过PV曲线求得的水分参数,对了解植物组织内部水分状况及渗透调节具有重要意义.其中,Ψtlp反映了植物维持最低膨压的极限渗透势,在耐旱性较强的植物中通常表现为较低的值(更负),被认为是衡量植物耐旱性的关键指标[24-26],在水分为主要限制因子的干旱、半干旱地区有重要意义.本研究中,Ψtlp和Ψ0的变化范围分别为-3.44~-1.52 MPa和-2.66~-1.33 MPa.Bartlett等[24]对72项研究中的317种植物进行了Meta分析,结果显示,全球半荒漠地区植物的Ψtlp和Ψ0的变化范围分别为-1.25~-4.25 MPa和-0.96~-2.78 MPa,本研究中Ψtlp和Ψ0值均在此范围内.斯琴巴特尔等[27]利用PV技术研究了荒漠区植物蒙古扁桃的水分生理特性,结果表明其Ψtlp为-3.11 MPa;李向义等[23]对沙漠-绿洲过渡带的4种植物进行水分关系研究,发现Ψtlp的变化范围为-1.50~-2.41 MPa;王乐乐等[28]通过对东北温带森林中20种生长于半湿润区的乔木进行PV曲线性状测定,结果显示Ψtlp平均值为-1.2 MPa.可以看出,从荒漠、沙漠-绿洲过渡带到半湿润区,Ψtlp随着水分梯度的增加而增加(绝对值更小).在干旱区,Ψtlp通常被认为具有较低值[29].Zhu等[25]汇编了中国9个木本植物群落中389种木本植物的Ψtlp数据集,发现沙漠灌木林带的Ψtlp为-1.52~-2.71 MPa,本研究中除梭梭、霸王外,其余植物Ψtlp值均在该范围内.梭梭与霸王具有更负的Ψtlp,表明其耐旱能力较强于其他树种.Xu等[30]研究了梭梭对水分条件变化的适应性,认为梭梭具有高效的形态调节和强大的气孔控制能力,有助于其适应环境的变化.石松利等[31]对霸王水分关系参数的研究表明,霸王主要通过增加细胞质浓度的渗透调节来维持膨压,显示出较强的耐旱性. ...
Adjustments and coordination of hydraulic,leaf and stem traits along a water availability gradient
1
2019
... PV技术作为研究树木耐旱性的重要手段之一,已广泛应用于植物水分生理研究[21-23].通过PV曲线求得的水分参数,对了解植物组织内部水分状况及渗透调节具有重要意义.其中,Ψtlp反映了植物维持最低膨压的极限渗透势,在耐旱性较强的植物中通常表现为较低的值(更负),被认为是衡量植物耐旱性的关键指标[24-26],在水分为主要限制因子的干旱、半干旱地区有重要意义.本研究中,Ψtlp和Ψ0的变化范围分别为-3.44~-1.52 MPa和-2.66~-1.33 MPa.Bartlett等[24]对72项研究中的317种植物进行了Meta分析,结果显示,全球半荒漠地区植物的Ψtlp和Ψ0的变化范围分别为-1.25~-4.25 MPa和-0.96~-2.78 MPa,本研究中Ψtlp和Ψ0值均在此范围内.斯琴巴特尔等[27]利用PV技术研究了荒漠区植物蒙古扁桃的水分生理特性,结果表明其Ψtlp为-3.11 MPa;李向义等[23]对沙漠-绿洲过渡带的4种植物进行水分关系研究,发现Ψtlp的变化范围为-1.50~-2.41 MPa;王乐乐等[28]通过对东北温带森林中20种生长于半湿润区的乔木进行PV曲线性状测定,结果显示Ψtlp平均值为-1.2 MPa.可以看出,从荒漠、沙漠-绿洲过渡带到半湿润区,Ψtlp随着水分梯度的增加而增加(绝对值更小).在干旱区,Ψtlp通常被认为具有较低值[29].Zhu等[25]汇编了中国9个木本植物群落中389种木本植物的Ψtlp数据集,发现沙漠灌木林带的Ψtlp为-1.52~-2.71 MPa,本研究中除梭梭、霸王外,其余植物Ψtlp值均在该范围内.梭梭与霸王具有更负的Ψtlp,表明其耐旱能力较强于其他树种.Xu等[30]研究了梭梭对水分条件变化的适应性,认为梭梭具有高效的形态调节和强大的气孔控制能力,有助于其适应环境的变化.石松利等[31]对霸王水分关系参数的研究表明,霸王主要通过增加细胞质浓度的渗透调节来维持膨压,显示出较强的耐旱性. ...
Ecophysiological response and morphological adjustment of two Central Asian desert shrubs towards variation in summer precipitation
1
2007
... PV技术作为研究树木耐旱性的重要手段之一,已广泛应用于植物水分生理研究[21-23].通过PV曲线求得的水分参数,对了解植物组织内部水分状况及渗透调节具有重要意义.其中,Ψtlp反映了植物维持最低膨压的极限渗透势,在耐旱性较强的植物中通常表现为较低的值(更负),被认为是衡量植物耐旱性的关键指标[24-26],在水分为主要限制因子的干旱、半干旱地区有重要意义.本研究中,Ψtlp和Ψ0的变化范围分别为-3.44~-1.52 MPa和-2.66~-1.33 MPa.Bartlett等[24]对72项研究中的317种植物进行了Meta分析,结果显示,全球半荒漠地区植物的Ψtlp和Ψ0的变化范围分别为-1.25~-4.25 MPa和-0.96~-2.78 MPa,本研究中Ψtlp和Ψ0值均在此范围内.斯琴巴特尔等[27]利用PV技术研究了荒漠区植物蒙古扁桃的水分生理特性,结果表明其Ψtlp为-3.11 MPa;李向义等[23]对沙漠-绿洲过渡带的4种植物进行水分关系研究,发现Ψtlp的变化范围为-1.50~-2.41 MPa;王乐乐等[28]通过对东北温带森林中20种生长于半湿润区的乔木进行PV曲线性状测定,结果显示Ψtlp平均值为-1.2 MPa.可以看出,从荒漠、沙漠-绿洲过渡带到半湿润区,Ψtlp随着水分梯度的增加而增加(绝对值更小).在干旱区,Ψtlp通常被认为具有较低值[29].Zhu等[25]汇编了中国9个木本植物群落中389种木本植物的Ψtlp数据集,发现沙漠灌木林带的Ψtlp为-1.52~-2.71 MPa,本研究中除梭梭、霸王外,其余植物Ψtlp值均在该范围内.梭梭与霸王具有更负的Ψtlp,表明其耐旱能力较强于其他树种.Xu等[30]研究了梭梭对水分条件变化的适应性,认为梭梭具有高效的形态调节和强大的气孔控制能力,有助于其适应环境的变化.石松利等[31]对霸王水分关系参数的研究表明,霸王主要通过增加细胞质浓度的渗透调节来维持膨压,显示出较强的耐旱性. ...
濒危种四合木与其近缘种霸王水分关系参数和光合特性的比较
1
... PV技术作为研究树木耐旱性的重要手段之一,已广泛应用于植物水分生理研究[21-23].通过PV曲线求得的水分参数,对了解植物组织内部水分状况及渗透调节具有重要意义.其中,Ψtlp反映了植物维持最低膨压的极限渗透势,在耐旱性较强的植物中通常表现为较低的值(更负),被认为是衡量植物耐旱性的关键指标[24-26],在水分为主要限制因子的干旱、半干旱地区有重要意义.本研究中,Ψtlp和Ψ0的变化范围分别为-3.44~-1.52 MPa和-2.66~-1.33 MPa.Bartlett等[24]对72项研究中的317种植物进行了Meta分析,结果显示,全球半荒漠地区植物的Ψtlp和Ψ0的变化范围分别为-1.25~-4.25 MPa和-0.96~-2.78 MPa,本研究中Ψtlp和Ψ0值均在此范围内.斯琴巴特尔等[27]利用PV技术研究了荒漠区植物蒙古扁桃的水分生理特性,结果表明其Ψtlp为-3.11 MPa;李向义等[23]对沙漠-绿洲过渡带的4种植物进行水分关系研究,发现Ψtlp的变化范围为-1.50~-2.41 MPa;王乐乐等[28]通过对东北温带森林中20种生长于半湿润区的乔木进行PV曲线性状测定,结果显示Ψtlp平均值为-1.2 MPa.可以看出,从荒漠、沙漠-绿洲过渡带到半湿润区,Ψtlp随着水分梯度的增加而增加(绝对值更小).在干旱区,Ψtlp通常被认为具有较低值[29].Zhu等[25]汇编了中国9个木本植物群落中389种木本植物的Ψtlp数据集,发现沙漠灌木林带的Ψtlp为-1.52~-2.71 MPa,本研究中除梭梭、霸王外,其余植物Ψtlp值均在该范围内.梭梭与霸王具有更负的Ψtlp,表明其耐旱能力较强于其他树种.Xu等[30]研究了梭梭对水分条件变化的适应性,认为梭梭具有高效的形态调节和强大的气孔控制能力,有助于其适应环境的变化.石松利等[31]对霸王水分关系参数的研究表明,霸王主要通过增加细胞质浓度的渗透调节来维持膨压,显示出较强的耐旱性. ...
植物叶片形态解剖结构对环境变化的响应与适应
2
2005
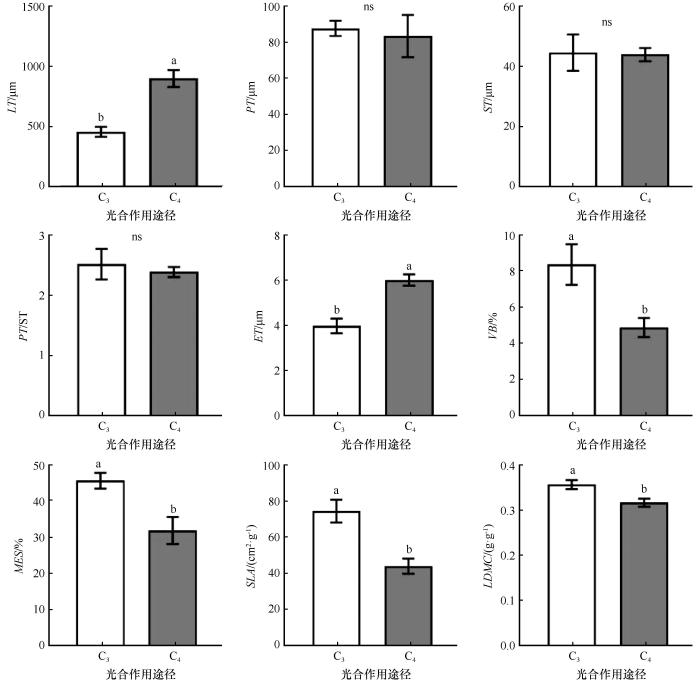
... 叶片是植物进化过程中对环境变化敏感且具有高度可塑性的器官[32],其解剖结构特征对植物的生理功能及其在特定环境下的适应性有重要影响.沙漠木本植物通过长期适应干旱环境,形成了一系列的旱生结构特征.例如,荒漠植物的叶片通常具有较厚的角质层、下陷的气孔、发达的栅栏组织和贮水组织、退化的海绵组织以及增强的机械组织等特征[33-34].结合本研究对叶片解剖结构的观察结果,我们发现荒漠植物的栅栏组织发达,没有或只有少数植物有海绵组织,栅栏组织的层数通常为2~3层.这些特征有助于增加叶绿体对光照和CO2的吸收面积,从而提高光合作用效率;同时,较厚的角质层和发达的贮水组织有助于植物在干旱条件下进行水分调节.此外,梭梭和沙拐枣等植物的叶片退化为同化枝[35-36],同化枝代替叶片进行光合和蒸腾作用,这是荒漠植物适应干旱环境形成的独特旱生结构.这些叶片特征有助于植物减少水分蒸散、提高光合作用效率,增强自身对环境变化的适应性,从而有利于在荒漠中生存和繁衍.Zhang等[37]的研究结果显示,湿润地区藤本植物、攀缘灌木和乔木的叶片平均厚度分别为206.03、192.15 μm和15.62 μm.相较之下,本研究中荒漠植物叶片平均厚度为632.63 μm,较厚的叶片或同化枝直径的增加有利于植物更有效地利用水分.此外,本研究中角质层平均厚度为4.80 μm,较厚的角质层能够减少水分蒸散[35,38],有利于植物维持体内水分平衡. ...
... 通过测量和计算叶片中叶肉组织、维管束和贮水组织的百分比,可以定量评估叶片的解剖结构特征,并提供关于植物生理功能和适应性线索[39].这些数据在植物生态学、生理学以及与环境适应性相关的研究中具有重要意义.在荒漠植物中,植物的叶肉组织向着提高光合效率的方向发展[32,40].栅栏组织形成了紧密排列的细胞,一方面形成有效的屏障,减少水分的蒸散和流失,另一方面可避免强光对叶肉细胞的灼伤,并能够有效利用衍射光进行光合作用[41-42].维管束组织在水分和养分的输送、维持植物结构的机械支持、代谢活动和生长调控等方面发挥着重要的作用,有利于植物适应干旱和恶劣的环境条件,保证了植物正常的生长和代谢.研究表明,具有贮水组织的荒漠植物通常具有较高的抗旱能力[43].贮水组织储存了大量的水分,使植物在干旱条件下能够更长时间维持正常的生理功能.当植物受到干旱胁迫时,贮水组织可以向栅栏组织等提供水分,帮助叶肉细胞维持一定的渗透压,从而维持正常的生理功能[44].本研究中,多数植物的贮水组织所占比例较大,其中梭梭贮水组织比例高达78.47%.由此可见,在干旱的荒漠环境中,贮水组织在植物叶片的渗透调节中发挥着重要作用.黄振英等[34]对30种新疆沙生植物营养器官的解剖学研究表明,旱生植物叶片的结构向着栅栏组织与贮水组织发达、叶片退化等方向演化,这与本研究的结果一致. ...
沙芥幼苗叶片解剖结构和光合作用对干旱胁迫的响应
1
2013
... 叶片是植物进化过程中对环境变化敏感且具有高度可塑性的器官[32],其解剖结构特征对植物的生理功能及其在特定环境下的适应性有重要影响.沙漠木本植物通过长期适应干旱环境,形成了一系列的旱生结构特征.例如,荒漠植物的叶片通常具有较厚的角质层、下陷的气孔、发达的栅栏组织和贮水组织、退化的海绵组织以及增强的机械组织等特征[33-34].结合本研究对叶片解剖结构的观察结果,我们发现荒漠植物的栅栏组织发达,没有或只有少数植物有海绵组织,栅栏组织的层数通常为2~3层.这些特征有助于增加叶绿体对光照和CO2的吸收面积,从而提高光合作用效率;同时,较厚的角质层和发达的贮水组织有助于植物在干旱条件下进行水分调节.此外,梭梭和沙拐枣等植物的叶片退化为同化枝[35-36],同化枝代替叶片进行光合和蒸腾作用,这是荒漠植物适应干旱环境形成的独特旱生结构.这些叶片特征有助于植物减少水分蒸散、提高光合作用效率,增强自身对环境变化的适应性,从而有利于在荒漠中生存和繁衍.Zhang等[37]的研究结果显示,湿润地区藤本植物、攀缘灌木和乔木的叶片平均厚度分别为206.03、192.15 μm和15.62 μm.相较之下,本研究中荒漠植物叶片平均厚度为632.63 μm,较厚的叶片或同化枝直径的增加有利于植物更有效地利用水分.此外,本研究中角质层平均厚度为4.80 μm,较厚的角质层能够减少水分蒸散[35,38],有利于植物维持体内水分平衡. ...
30种新疆沙生植物的结构及其对沙漠环境的适应
2
1997
... 叶片是植物进化过程中对环境变化敏感且具有高度可塑性的器官[32],其解剖结构特征对植物的生理功能及其在特定环境下的适应性有重要影响.沙漠木本植物通过长期适应干旱环境,形成了一系列的旱生结构特征.例如,荒漠植物的叶片通常具有较厚的角质层、下陷的气孔、发达的栅栏组织和贮水组织、退化的海绵组织以及增强的机械组织等特征[33-34].结合本研究对叶片解剖结构的观察结果,我们发现荒漠植物的栅栏组织发达,没有或只有少数植物有海绵组织,栅栏组织的层数通常为2~3层.这些特征有助于增加叶绿体对光照和CO2的吸收面积,从而提高光合作用效率;同时,较厚的角质层和发达的贮水组织有助于植物在干旱条件下进行水分调节.此外,梭梭和沙拐枣等植物的叶片退化为同化枝[35-36],同化枝代替叶片进行光合和蒸腾作用,这是荒漠植物适应干旱环境形成的独特旱生结构.这些叶片特征有助于植物减少水分蒸散、提高光合作用效率,增强自身对环境变化的适应性,从而有利于在荒漠中生存和繁衍.Zhang等[37]的研究结果显示,湿润地区藤本植物、攀缘灌木和乔木的叶片平均厚度分别为206.03、192.15 μm和15.62 μm.相较之下,本研究中荒漠植物叶片平均厚度为632.63 μm,较厚的叶片或同化枝直径的增加有利于植物更有效地利用水分.此外,本研究中角质层平均厚度为4.80 μm,较厚的角质层能够减少水分蒸散[35,38],有利于植物维持体内水分平衡. ...
... 通过测量和计算叶片中叶肉组织、维管束和贮水组织的百分比,可以定量评估叶片的解剖结构特征,并提供关于植物生理功能和适应性线索[39].这些数据在植物生态学、生理学以及与环境适应性相关的研究中具有重要意义.在荒漠植物中,植物的叶肉组织向着提高光合效率的方向发展[32,40].栅栏组织形成了紧密排列的细胞,一方面形成有效的屏障,减少水分的蒸散和流失,另一方面可避免强光对叶肉细胞的灼伤,并能够有效利用衍射光进行光合作用[41-42].维管束组织在水分和养分的输送、维持植物结构的机械支持、代谢活动和生长调控等方面发挥着重要的作用,有利于植物适应干旱和恶劣的环境条件,保证了植物正常的生长和代谢.研究表明,具有贮水组织的荒漠植物通常具有较高的抗旱能力[43].贮水组织储存了大量的水分,使植物在干旱条件下能够更长时间维持正常的生理功能.当植物受到干旱胁迫时,贮水组织可以向栅栏组织等提供水分,帮助叶肉细胞维持一定的渗透压,从而维持正常的生理功能[44].本研究中,多数植物的贮水组织所占比例较大,其中梭梭贮水组织比例高达78.47%.由此可见,在干旱的荒漠环境中,贮水组织在植物叶片的渗透调节中发挥着重要作用.黄振英等[34]对30种新疆沙生植物营养器官的解剖学研究表明,旱生植物叶片的结构向着栅栏组织与贮水组织发达、叶片退化等方向演化,这与本研究的结果一致. ...
中国干旱半干旱区荒漠植物叶片 (或同化枝) 表皮微形态特征
2
2016
... 叶片是植物进化过程中对环境变化敏感且具有高度可塑性的器官[32],其解剖结构特征对植物的生理功能及其在特定环境下的适应性有重要影响.沙漠木本植物通过长期适应干旱环境,形成了一系列的旱生结构特征.例如,荒漠植物的叶片通常具有较厚的角质层、下陷的气孔、发达的栅栏组织和贮水组织、退化的海绵组织以及增强的机械组织等特征[33-34].结合本研究对叶片解剖结构的观察结果,我们发现荒漠植物的栅栏组织发达,没有或只有少数植物有海绵组织,栅栏组织的层数通常为2~3层.这些特征有助于增加叶绿体对光照和CO2的吸收面积,从而提高光合作用效率;同时,较厚的角质层和发达的贮水组织有助于植物在干旱条件下进行水分调节.此外,梭梭和沙拐枣等植物的叶片退化为同化枝[35-36],同化枝代替叶片进行光合和蒸腾作用,这是荒漠植物适应干旱环境形成的独特旱生结构.这些叶片特征有助于植物减少水分蒸散、提高光合作用效率,增强自身对环境变化的适应性,从而有利于在荒漠中生存和繁衍.Zhang等[37]的研究结果显示,湿润地区藤本植物、攀缘灌木和乔木的叶片平均厚度分别为206.03、192.15 μm和15.62 μm.相较之下,本研究中荒漠植物叶片平均厚度为632.63 μm,较厚的叶片或同化枝直径的增加有利于植物更有效地利用水分.此外,本研究中角质层平均厚度为4.80 μm,较厚的角质层能够减少水分蒸散[35,38],有利于植物维持体内水分平衡. ...
... [35,38],有利于植物维持体内水分平衡. ...
Multi-level physiological and morphological adjustment of haloxylon ammodendron related to groundwater drawdown in a desert ecosystem
1
2022
... 叶片是植物进化过程中对环境变化敏感且具有高度可塑性的器官[32],其解剖结构特征对植物的生理功能及其在特定环境下的适应性有重要影响.沙漠木本植物通过长期适应干旱环境,形成了一系列的旱生结构特征.例如,荒漠植物的叶片通常具有较厚的角质层、下陷的气孔、发达的栅栏组织和贮水组织、退化的海绵组织以及增强的机械组织等特征[33-34].结合本研究对叶片解剖结构的观察结果,我们发现荒漠植物的栅栏组织发达,没有或只有少数植物有海绵组织,栅栏组织的层数通常为2~3层.这些特征有助于增加叶绿体对光照和CO2的吸收面积,从而提高光合作用效率;同时,较厚的角质层和发达的贮水组织有助于植物在干旱条件下进行水分调节.此外,梭梭和沙拐枣等植物的叶片退化为同化枝[35-36],同化枝代替叶片进行光合和蒸腾作用,这是荒漠植物适应干旱环境形成的独特旱生结构.这些叶片特征有助于植物减少水分蒸散、提高光合作用效率,增强自身对环境变化的适应性,从而有利于在荒漠中生存和繁衍.Zhang等[37]的研究结果显示,湿润地区藤本植物、攀缘灌木和乔木的叶片平均厚度分别为206.03、192.15 μm和15.62 μm.相较之下,本研究中荒漠植物叶片平均厚度为632.63 μm,较厚的叶片或同化枝直径的增加有利于植物更有效地利用水分.此外,本研究中角质层平均厚度为4.80 μm,较厚的角质层能够减少水分蒸散[35,38],有利于植物维持体内水分平衡. ...
Differentiation in stem and leaf traits among sympatric lianas,scandent shrubs and trees in a subalpine cold temperate forest
1
2021
... 叶片是植物进化过程中对环境变化敏感且具有高度可塑性的器官[32],其解剖结构特征对植物的生理功能及其在特定环境下的适应性有重要影响.沙漠木本植物通过长期适应干旱环境,形成了一系列的旱生结构特征.例如,荒漠植物的叶片通常具有较厚的角质层、下陷的气孔、发达的栅栏组织和贮水组织、退化的海绵组织以及增强的机械组织等特征[33-34].结合本研究对叶片解剖结构的观察结果,我们发现荒漠植物的栅栏组织发达,没有或只有少数植物有海绵组织,栅栏组织的层数通常为2~3层.这些特征有助于增加叶绿体对光照和CO2的吸收面积,从而提高光合作用效率;同时,较厚的角质层和发达的贮水组织有助于植物在干旱条件下进行水分调节.此外,梭梭和沙拐枣等植物的叶片退化为同化枝[35-36],同化枝代替叶片进行光合和蒸腾作用,这是荒漠植物适应干旱环境形成的独特旱生结构.这些叶片特征有助于植物减少水分蒸散、提高光合作用效率,增强自身对环境变化的适应性,从而有利于在荒漠中生存和繁衍.Zhang等[37]的研究结果显示,湿润地区藤本植物、攀缘灌木和乔木的叶片平均厚度分别为206.03、192.15 μm和15.62 μm.相较之下,本研究中荒漠植物叶片平均厚度为632.63 μm,较厚的叶片或同化枝直径的增加有利于植物更有效地利用水分.此外,本研究中角质层平均厚度为4.80 μm,较厚的角质层能够减少水分蒸散[35,38],有利于植物维持体内水分平衡. ...
荒漠区植物光合器官解剖结构对水分利用效率的指示作用
1
2013
... 叶片是植物进化过程中对环境变化敏感且具有高度可塑性的器官[32],其解剖结构特征对植物的生理功能及其在特定环境下的适应性有重要影响.沙漠木本植物通过长期适应干旱环境,形成了一系列的旱生结构特征.例如,荒漠植物的叶片通常具有较厚的角质层、下陷的气孔、发达的栅栏组织和贮水组织、退化的海绵组织以及增强的机械组织等特征[33-34].结合本研究对叶片解剖结构的观察结果,我们发现荒漠植物的栅栏组织发达,没有或只有少数植物有海绵组织,栅栏组织的层数通常为2~3层.这些特征有助于增加叶绿体对光照和CO2的吸收面积,从而提高光合作用效率;同时,较厚的角质层和发达的贮水组织有助于植物在干旱条件下进行水分调节.此外,梭梭和沙拐枣等植物的叶片退化为同化枝[35-36],同化枝代替叶片进行光合和蒸腾作用,这是荒漠植物适应干旱环境形成的独特旱生结构.这些叶片特征有助于植物减少水分蒸散、提高光合作用效率,增强自身对环境变化的适应性,从而有利于在荒漠中生存和繁衍.Zhang等[37]的研究结果显示,湿润地区藤本植物、攀缘灌木和乔木的叶片平均厚度分别为206.03、192.15 μm和15.62 μm.相较之下,本研究中荒漠植物叶片平均厚度为632.63 μm,较厚的叶片或同化枝直径的增加有利于植物更有效地利用水分.此外,本研究中角质层平均厚度为4.80 μm,较厚的角质层能够减少水分蒸散[35,38],有利于植物维持体内水分平衡. ...
Quantitative mesophyll parameters rather than whole-leaf traits predict response of C3 steppe plants to aridity
1
2018
... 通过测量和计算叶片中叶肉组织、维管束和贮水组织的百分比,可以定量评估叶片的解剖结构特征,并提供关于植物生理功能和适应性线索[39].这些数据在植物生态学、生理学以及与环境适应性相关的研究中具有重要意义.在荒漠植物中,植物的叶肉组织向着提高光合效率的方向发展[32,40].栅栏组织形成了紧密排列的细胞,一方面形成有效的屏障,减少水分的蒸散和流失,另一方面可避免强光对叶肉细胞的灼伤,并能够有效利用衍射光进行光合作用[41-42].维管束组织在水分和养分的输送、维持植物结构的机械支持、代谢活动和生长调控等方面发挥着重要的作用,有利于植物适应干旱和恶劣的环境条件,保证了植物正常的生长和代谢.研究表明,具有贮水组织的荒漠植物通常具有较高的抗旱能力[43].贮水组织储存了大量的水分,使植物在干旱条件下能够更长时间维持正常的生理功能.当植物受到干旱胁迫时,贮水组织可以向栅栏组织等提供水分,帮助叶肉细胞维持一定的渗透压,从而维持正常的生理功能[44].本研究中,多数植物的贮水组织所占比例较大,其中梭梭贮水组织比例高达78.47%.由此可见,在干旱的荒漠环境中,贮水组织在植物叶片的渗透调节中发挥着重要作用.黄振英等[34]对30种新疆沙生植物营养器官的解剖学研究表明,旱生植物叶片的结构向着栅栏组织与贮水组织发达、叶片退化等方向演化,这与本研究的结果一致. ...
库布齐沙漠几种沙生灌木叶解剖结构耐旱特征研究
1
2010
... 通过测量和计算叶片中叶肉组织、维管束和贮水组织的百分比,可以定量评估叶片的解剖结构特征,并提供关于植物生理功能和适应性线索[39].这些数据在植物生态学、生理学以及与环境适应性相关的研究中具有重要意义.在荒漠植物中,植物的叶肉组织向着提高光合效率的方向发展[32,40].栅栏组织形成了紧密排列的细胞,一方面形成有效的屏障,减少水分的蒸散和流失,另一方面可避免强光对叶肉细胞的灼伤,并能够有效利用衍射光进行光合作用[41-42].维管束组织在水分和养分的输送、维持植物结构的机械支持、代谢活动和生长调控等方面发挥着重要的作用,有利于植物适应干旱和恶劣的环境条件,保证了植物正常的生长和代谢.研究表明,具有贮水组织的荒漠植物通常具有较高的抗旱能力[43].贮水组织储存了大量的水分,使植物在干旱条件下能够更长时间维持正常的生理功能.当植物受到干旱胁迫时,贮水组织可以向栅栏组织等提供水分,帮助叶肉细胞维持一定的渗透压,从而维持正常的生理功能[44].本研究中,多数植物的贮水组织所占比例较大,其中梭梭贮水组织比例高达78.47%.由此可见,在干旱的荒漠环境中,贮水组织在植物叶片的渗透调节中发挥着重要作用.黄振英等[34]对30种新疆沙生植物营养器官的解剖学研究表明,旱生植物叶片的结构向着栅栏组织与贮水组织发达、叶片退化等方向演化,这与本研究的结果一致. ...
胡杨披针形叶与宽卵形叶的渗透调节能力的差异
1
2004
... 通过测量和计算叶片中叶肉组织、维管束和贮水组织的百分比,可以定量评估叶片的解剖结构特征,并提供关于植物生理功能和适应性线索[39].这些数据在植物生态学、生理学以及与环境适应性相关的研究中具有重要意义.在荒漠植物中,植物的叶肉组织向着提高光合效率的方向发展[32,40].栅栏组织形成了紧密排列的细胞,一方面形成有效的屏障,减少水分的蒸散和流失,另一方面可避免强光对叶肉细胞的灼伤,并能够有效利用衍射光进行光合作用[41-42].维管束组织在水分和养分的输送、维持植物结构的机械支持、代谢活动和生长调控等方面发挥着重要的作用,有利于植物适应干旱和恶劣的环境条件,保证了植物正常的生长和代谢.研究表明,具有贮水组织的荒漠植物通常具有较高的抗旱能力[43].贮水组织储存了大量的水分,使植物在干旱条件下能够更长时间维持正常的生理功能.当植物受到干旱胁迫时,贮水组织可以向栅栏组织等提供水分,帮助叶肉细胞维持一定的渗透压,从而维持正常的生理功能[44].本研究中,多数植物的贮水组织所占比例较大,其中梭梭贮水组织比例高达78.47%.由此可见,在干旱的荒漠环境中,贮水组织在植物叶片的渗透调节中发挥着重要作用.黄振英等[34]对30种新疆沙生植物营养器官的解剖学研究表明,旱生植物叶片的结构向着栅栏组织与贮水组织发达、叶片退化等方向演化,这与本研究的结果一致. ...
罗布泊地区几种旱生植物茎、叶结构的初步研究
1
1984
... 通过测量和计算叶片中叶肉组织、维管束和贮水组织的百分比,可以定量评估叶片的解剖结构特征,并提供关于植物生理功能和适应性线索[39].这些数据在植物生态学、生理学以及与环境适应性相关的研究中具有重要意义.在荒漠植物中,植物的叶肉组织向着提高光合效率的方向发展[32,40].栅栏组织形成了紧密排列的细胞,一方面形成有效的屏障,减少水分的蒸散和流失,另一方面可避免强光对叶肉细胞的灼伤,并能够有效利用衍射光进行光合作用[41-42].维管束组织在水分和养分的输送、维持植物结构的机械支持、代谢活动和生长调控等方面发挥着重要的作用,有利于植物适应干旱和恶劣的环境条件,保证了植物正常的生长和代谢.研究表明,具有贮水组织的荒漠植物通常具有较高的抗旱能力[43].贮水组织储存了大量的水分,使植物在干旱条件下能够更长时间维持正常的生理功能.当植物受到干旱胁迫时,贮水组织可以向栅栏组织等提供水分,帮助叶肉细胞维持一定的渗透压,从而维持正常的生理功能[44].本研究中,多数植物的贮水组织所占比例较大,其中梭梭贮水组织比例高达78.47%.由此可见,在干旱的荒漠环境中,贮水组织在植物叶片的渗透调节中发挥着重要作用.黄振英等[34]对30种新疆沙生植物营养器官的解剖学研究表明,旱生植物叶片的结构向着栅栏组织与贮水组织发达、叶片退化等方向演化,这与本研究的结果一致. ...
不同梭梭种群同化枝的解剖结构特征及其与生态因子的关系分析
1
2016
... 通过测量和计算叶片中叶肉组织、维管束和贮水组织的百分比,可以定量评估叶片的解剖结构特征,并提供关于植物生理功能和适应性线索[39].这些数据在植物生态学、生理学以及与环境适应性相关的研究中具有重要意义.在荒漠植物中,植物的叶肉组织向着提高光合效率的方向发展[32,40].栅栏组织形成了紧密排列的细胞,一方面形成有效的屏障,减少水分的蒸散和流失,另一方面可避免强光对叶肉细胞的灼伤,并能够有效利用衍射光进行光合作用[41-42].维管束组织在水分和养分的输送、维持植物结构的机械支持、代谢活动和生长调控等方面发挥着重要的作用,有利于植物适应干旱和恶劣的环境条件,保证了植物正常的生长和代谢.研究表明,具有贮水组织的荒漠植物通常具有较高的抗旱能力[43].贮水组织储存了大量的水分,使植物在干旱条件下能够更长时间维持正常的生理功能.当植物受到干旱胁迫时,贮水组织可以向栅栏组织等提供水分,帮助叶肉细胞维持一定的渗透压,从而维持正常的生理功能[44].本研究中,多数植物的贮水组织所占比例较大,其中梭梭贮水组织比例高达78.47%.由此可见,在干旱的荒漠环境中,贮水组织在植物叶片的渗透调节中发挥着重要作用.黄振英等[34]对30种新疆沙生植物营养器官的解剖学研究表明,旱生植物叶片的结构向着栅栏组织与贮水组织发达、叶片退化等方向演化,这与本研究的结果一致. ...
几种沙生植物水分代谢问题探讨
1
2011
... 通过测量和计算叶片中叶肉组织、维管束和贮水组织的百分比,可以定量评估叶片的解剖结构特征,并提供关于植物生理功能和适应性线索[39].这些数据在植物生态学、生理学以及与环境适应性相关的研究中具有重要意义.在荒漠植物中,植物的叶肉组织向着提高光合效率的方向发展[32,40].栅栏组织形成了紧密排列的细胞,一方面形成有效的屏障,减少水分的蒸散和流失,另一方面可避免强光对叶肉细胞的灼伤,并能够有效利用衍射光进行光合作用[41-42].维管束组织在水分和养分的输送、维持植物结构的机械支持、代谢活动和生长调控等方面发挥着重要的作用,有利于植物适应干旱和恶劣的环境条件,保证了植物正常的生长和代谢.研究表明,具有贮水组织的荒漠植物通常具有较高的抗旱能力[43].贮水组织储存了大量的水分,使植物在干旱条件下能够更长时间维持正常的生理功能.当植物受到干旱胁迫时,贮水组织可以向栅栏组织等提供水分,帮助叶肉细胞维持一定的渗透压,从而维持正常的生理功能[44].本研究中,多数植物的贮水组织所占比例较大,其中梭梭贮水组织比例高达78.47%.由此可见,在干旱的荒漠环境中,贮水组织在植物叶片的渗透调节中发挥着重要作用.黄振英等[34]对30种新疆沙生植物营养器官的解剖学研究表明,旱生植物叶片的结构向着栅栏组织与贮水组织发达、叶片退化等方向演化,这与本研究的结果一致. ...
C3和C4植物光合途径的适应性变化和进化
1
2009
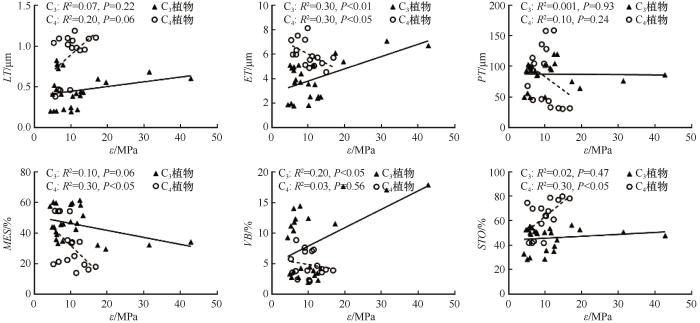
... C3和C4植物在光合作用过程中固定二氧化碳的方式存在显著差异.C3植物通过卡尔文循环直接固定二氧化碳,而C4植物则在进入卡尔文循环之前,先将二氧化碳固定为四碳化合物.由于光合作用途径不同,C3和C4植物的水分利用效率和耐旱性也有所差异.本研究中,C3植物的Ψtlp和Ψ0均显著高于C4植物,表明C4植物耐旱性优于C3植物,这与C4植物通常生存于更为严酷的高温干旱环境的特性一致[45].Jacob等[46]研究发现,C3草本的Ψtlp比C4草本更负,这与本研究结果相反.这种差异可能与Ψtlp在不同生活型植物中的溶质积累方式有关.通过对比解剖结构,我们发现C4植物LT和ET显著大于C3植物.一方面,较厚的叶片可以储存充足的水分,维持稳定的水分供应[47].另一方面,较厚的角质层被认为是防止排气孔水分损失的有效屏障[48].厚叶片通过增加叶绿素等成分的体积和氮含量,从而提高植物在强光照条件下的光合能力.相较于薄叶,厚叶更能有效保护植物免受阳光损害[49-50],这可能是C4植物能够在干旱生境中生存的一种机制,这与C4植物在高温、强光和低水分条件下能保持高的光合速率的特性[51]一致. ...
High safety margins to drought‐induced hydraulic failure found in five pasture grasses
1
2022
... C3和C4植物在光合作用过程中固定二氧化碳的方式存在显著差异.C3植物通过卡尔文循环直接固定二氧化碳,而C4植物则在进入卡尔文循环之前,先将二氧化碳固定为四碳化合物.由于光合作用途径不同,C3和C4植物的水分利用效率和耐旱性也有所差异.本研究中,C3植物的Ψtlp和Ψ0均显著高于C4植物,表明C4植物耐旱性优于C3植物,这与C4植物通常生存于更为严酷的高温干旱环境的特性一致[45].Jacob等[46]研究发现,C3草本的Ψtlp比C4草本更负,这与本研究结果相反.这种差异可能与Ψtlp在不同生活型植物中的溶质积累方式有关.通过对比解剖结构,我们发现C4植物LT和ET显著大于C3植物.一方面,较厚的叶片可以储存充足的水分,维持稳定的水分供应[47].另一方面,较厚的角质层被认为是防止排气孔水分损失的有效屏障[48].厚叶片通过增加叶绿素等成分的体积和氮含量,从而提高植物在强光照条件下的光合能力.相较于薄叶,厚叶更能有效保护植物免受阳光损害[49-50],这可能是C4植物能够在干旱生境中生存的一种机制,这与C4植物在高温、强光和低水分条件下能保持高的光合速率的特性[51]一致. ...
Quantifying succulence:a rapid,physiologically meaningful metric of plant water storage
1
2012
... C3和C4植物在光合作用过程中固定二氧化碳的方式存在显著差异.C3植物通过卡尔文循环直接固定二氧化碳,而C4植物则在进入卡尔文循环之前,先将二氧化碳固定为四碳化合物.由于光合作用途径不同,C3和C4植物的水分利用效率和耐旱性也有所差异.本研究中,C3植物的Ψtlp和Ψ0均显著高于C4植物,表明C4植物耐旱性优于C3植物,这与C4植物通常生存于更为严酷的高温干旱环境的特性一致[45].Jacob等[46]研究发现,C3草本的Ψtlp比C4草本更负,这与本研究结果相反.这种差异可能与Ψtlp在不同生活型植物中的溶质积累方式有关.通过对比解剖结构,我们发现C4植物LT和ET显著大于C3植物.一方面,较厚的叶片可以储存充足的水分,维持稳定的水分供应[47].另一方面,较厚的角质层被认为是防止排气孔水分损失的有效屏障[48].厚叶片通过增加叶绿素等成分的体积和氮含量,从而提高植物在强光照条件下的光合能力.相较于薄叶,厚叶更能有效保护植物免受阳光损害[49-50],这可能是C4植物能够在干旱生境中生存的一种机制,这与C4植物在高温、强光和低水分条件下能保持高的光合速率的特性[51]一致. ...
塔里木沙漠公路防护林植物沙拐枣气体交换特性对干旱胁迫的响应
1
2007
... C3和C4植物在光合作用过程中固定二氧化碳的方式存在显著差异.C3植物通过卡尔文循环直接固定二氧化碳,而C4植物则在进入卡尔文循环之前,先将二氧化碳固定为四碳化合物.由于光合作用途径不同,C3和C4植物的水分利用效率和耐旱性也有所差异.本研究中,C3植物的Ψtlp和Ψ0均显著高于C4植物,表明C4植物耐旱性优于C3植物,这与C4植物通常生存于更为严酷的高温干旱环境的特性一致[45].Jacob等[46]研究发现,C3草本的Ψtlp比C4草本更负,这与本研究结果相反.这种差异可能与Ψtlp在不同生活型植物中的溶质积累方式有关.通过对比解剖结构,我们发现C4植物LT和ET显著大于C3植物.一方面,较厚的叶片可以储存充足的水分,维持稳定的水分供应[47].另一方面,较厚的角质层被认为是防止排气孔水分损失的有效屏障[48].厚叶片通过增加叶绿素等成分的体积和氮含量,从而提高植物在强光照条件下的光合能力.相较于薄叶,厚叶更能有效保护植物免受阳光损害[49-50],这可能是C4植物能够在干旱生境中生存的一种机制,这与C4植物在高温、强光和低水分条件下能保持高的光合速率的特性[51]一致. ...
Global‐scale climatic controls of leaf dry mass per area,density,and thickness in trees and shrubs
2
2001
... C3和C4植物在光合作用过程中固定二氧化碳的方式存在显著差异.C3植物通过卡尔文循环直接固定二氧化碳,而C4植物则在进入卡尔文循环之前,先将二氧化碳固定为四碳化合物.由于光合作用途径不同,C3和C4植物的水分利用效率和耐旱性也有所差异.本研究中,C3植物的Ψtlp和Ψ0均显著高于C4植物,表明C4植物耐旱性优于C3植物,这与C4植物通常生存于更为严酷的高温干旱环境的特性一致[45].Jacob等[46]研究发现,C3草本的Ψtlp比C4草本更负,这与本研究结果相反.这种差异可能与Ψtlp在不同生活型植物中的溶质积累方式有关.通过对比解剖结构,我们发现C4植物LT和ET显著大于C3植物.一方面,较厚的叶片可以储存充足的水分,维持稳定的水分供应[47].另一方面,较厚的角质层被认为是防止排气孔水分损失的有效屏障[48].厚叶片通过增加叶绿素等成分的体积和氮含量,从而提高植物在强光照条件下的光合能力.相较于薄叶,厚叶更能有效保护植物免受阳光损害[49-50],这可能是C4植物能够在干旱生境中生存的一种机制,这与C4植物在高温、强光和低水分条件下能保持高的光合速率的特性[51]一致. ...
... C3植物的SLA和LDMC均高于C4植物,这与Akram等[52]的研究结果一致.C3植物的高SLA表明其具有较高的叶片内阻力和较低的角质层导水率[53].C3植物具有较高LDMC,意味着C3植物叶细胞之间存在较小的细胞间隙,且叶肉细胞对气体扩散具有较高的阻力,进而减少叶片蒸腾,这可能是适应干旱的一种策略.C4植物的高LT表明其可能具有较低的LDMC [49]. ...
Variation in leaf morphological,stomatal,and anatomical traits and their relationships in temperate and subtropical forests
1
2019
... C3和C4植物在光合作用过程中固定二氧化碳的方式存在显著差异.C3植物通过卡尔文循环直接固定二氧化碳,而C4植物则在进入卡尔文循环之前,先将二氧化碳固定为四碳化合物.由于光合作用途径不同,C3和C4植物的水分利用效率和耐旱性也有所差异.本研究中,C3植物的Ψtlp和Ψ0均显著高于C4植物,表明C4植物耐旱性优于C3植物,这与C4植物通常生存于更为严酷的高温干旱环境的特性一致[45].Jacob等[46]研究发现,C3草本的Ψtlp比C4草本更负,这与本研究结果相反.这种差异可能与Ψtlp在不同生活型植物中的溶质积累方式有关.通过对比解剖结构,我们发现C4植物LT和ET显著大于C3植物.一方面,较厚的叶片可以储存充足的水分,维持稳定的水分供应[47].另一方面,较厚的角质层被认为是防止排气孔水分损失的有效屏障[48].厚叶片通过增加叶绿素等成分的体积和氮含量,从而提高植物在强光照条件下的光合能力.相较于薄叶,厚叶更能有效保护植物免受阳光损害[49-50],这可能是C4植物能够在干旱生境中生存的一种机制,这与C4植物在高温、强光和低水分条件下能保持高的光合速率的特性[51]一致. ...
High hopes for C4 plants
1
1994
... C3和C4植物在光合作用过程中固定二氧化碳的方式存在显著差异.C3植物通过卡尔文循环直接固定二氧化碳,而C4植物则在进入卡尔文循环之前,先将二氧化碳固定为四碳化合物.由于光合作用途径不同,C3和C4植物的水分利用效率和耐旱性也有所差异.本研究中,C3植物的Ψtlp和Ψ0均显著高于C4植物,表明C4植物耐旱性优于C3植物,这与C4植物通常生存于更为严酷的高温干旱环境的特性一致[45].Jacob等[46]研究发现,C3草本的Ψtlp比C4草本更负,这与本研究结果相反.这种差异可能与Ψtlp在不同生活型植物中的溶质积累方式有关.通过对比解剖结构,我们发现C4植物LT和ET显著大于C3植物.一方面,较厚的叶片可以储存充足的水分,维持稳定的水分供应[47].另一方面,较厚的角质层被认为是防止排气孔水分损失的有效屏障[48].厚叶片通过增加叶绿素等成分的体积和氮含量,从而提高植物在强光照条件下的光合能力.相较于薄叶,厚叶更能有效保护植物免受阳光损害[49-50],这可能是C4植物能够在干旱生境中生存的一种机制,这与C4植物在高温、强光和低水分条件下能保持高的光合速率的特性[51]一致. ...
Phylogenetic independence in the variations in leaf functional traits among different plant life forms in an arid environment
1
2022
... C3植物的SLA和LDMC均高于C4植物,这与Akram等[52]的研究结果一致.C3植物的高SLA表明其具有较高的叶片内阻力和较低的角质层导水率[53].C3植物具有较高LDMC,意味着C3植物叶细胞之间存在较小的细胞间隙,且叶肉细胞对气体扩散具有较高的阻力,进而减少叶片蒸腾,这可能是适应干旱的一种策略.C4植物的高LT表明其可能具有较低的LDMC [49]. ...
How do leaf veins influence the worldwide leaf economic spectrum?Review and synthesis
1
2013
... C3植物的SLA和LDMC均高于C4植物,这与Akram等[52]的研究结果一致.C3植物的高SLA表明其具有较高的叶片内阻力和较低的角质层导水率[53].C3植物具有较高LDMC,意味着C3植物叶细胞之间存在较小的细胞间隙,且叶肉细胞对气体扩散具有较高的阻力,进而减少叶片蒸腾,这可能是适应干旱的一种策略.C4植物的高LT表明其可能具有较低的LDMC [49]. ...
Assessing the generality of global leaf trait relationships
1
2005
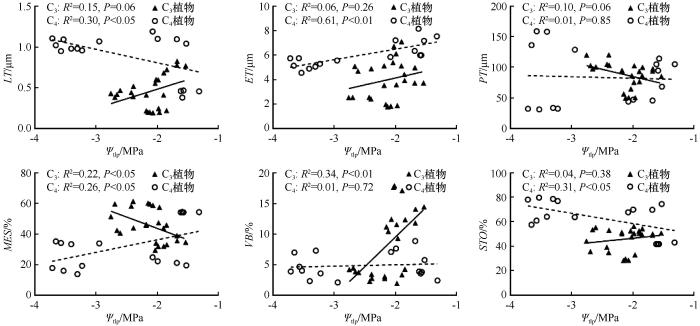
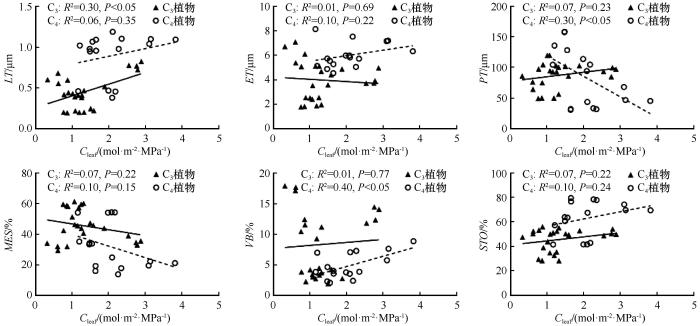
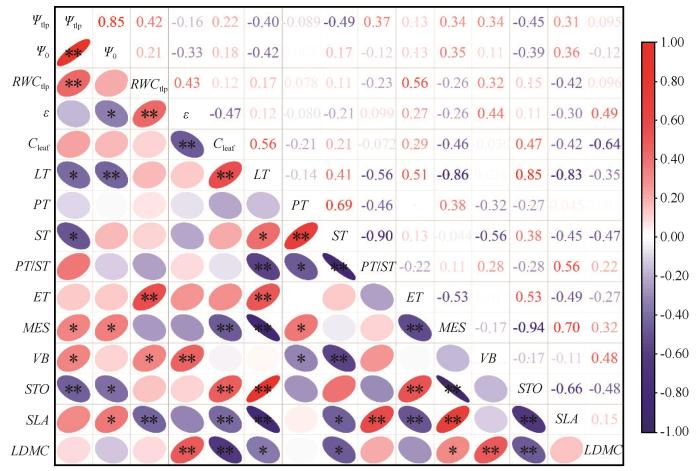
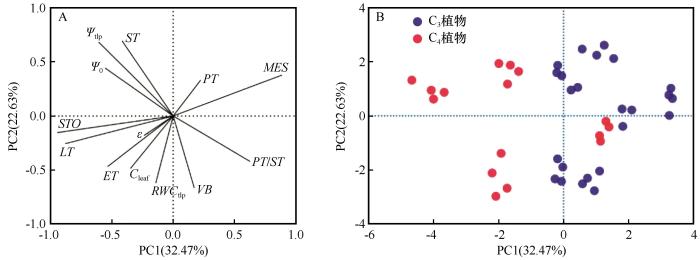
... PV曲线参数与叶片解剖学之间的关系为理解植物如何在干旱胁迫下调节水分提供了新的见解.对于两种不同光合作用途径的植物,PV性状与解剖结构间的关系较为复杂.有些性状之间关系一致,如ET与Ψtlp、MES与ε.然而,有些性状之间关系呈相反关系,如MES与Ψtlp,这可能反映出不同植物在生态策略轴上处于适应或竞争的位置[54].相关性分析结果显示,Ψtlp与叶片解剖性状之间存在显著相关性.Ψtlp与LT和STO呈显著负相关,表明随着叶片厚度和贮水组织的增加,膨压丧失点的水势变得更负.Ψtlp与MES、VB呈显著正相关,表明随着叶肉细胞和维管束比例的减少,植物忍耐干旱的能力增强.Nguyen 等[55-56]的研究强调了叶片解剖学在解释PV曲线中的重要性,突出了叶片解剖结构在决定Ψtlp、Ψ0以及ε和Cleaf等参数中的作用.通过探究不同光合作用途径植物的PV曲线参数与叶片解剖学特征之间的关系,我们不仅加深了对沙生植物在干旱胁迫下生理适应机制的理解,也为未来在不同生态环境下探索植物适应策略提供了新的视角. ...
Leaf water storage increases with salinity and aridity in the mangrove Avicennia marina:integration of leaf structure,osmotic adjustment and access to multiple water sources
1
2017
... PV曲线参数与叶片解剖学之间的关系为理解植物如何在干旱胁迫下调节水分提供了新的见解.对于两种不同光合作用途径的植物,PV性状与解剖结构间的关系较为复杂.有些性状之间关系一致,如ET与Ψtlp、MES与ε.然而,有些性状之间关系呈相反关系,如MES与Ψtlp,这可能反映出不同植物在生态策略轴上处于适应或竞争的位置[54].相关性分析结果显示,Ψtlp与叶片解剖性状之间存在显著相关性.Ψtlp与LT和STO呈显著负相关,表明随着叶片厚度和贮水组织的增加,膨压丧失点的水势变得更负.Ψtlp与MES、VB呈显著正相关,表明随着叶肉细胞和维管束比例的减少,植物忍耐干旱的能力增强.Nguyen 等[55-56]的研究强调了叶片解剖学在解释PV曲线中的重要性,突出了叶片解剖结构在决定Ψtlp、Ψ0以及ε和Cleaf等参数中的作用.通过探究不同光合作用途径植物的PV曲线参数与叶片解剖学特征之间的关系,我们不仅加深了对沙生植物在干旱胁迫下生理适应机制的理解,也为未来在不同生态环境下探索植物适应策略提供了新的视角. ...
Plumbing the depths:extracellular water storage in specialized leaf structures and its functional expression in a three‐domain pressure-volume relationship
1
2017
... PV曲线参数与叶片解剖学之间的关系为理解植物如何在干旱胁迫下调节水分提供了新的见解.对于两种不同光合作用途径的植物,PV性状与解剖结构间的关系较为复杂.有些性状之间关系一致,如ET与Ψtlp、MES与ε.然而,有些性状之间关系呈相反关系,如MES与Ψtlp,这可能反映出不同植物在生态策略轴上处于适应或竞争的位置[54].相关性分析结果显示,Ψtlp与叶片解剖性状之间存在显著相关性.Ψtlp与LT和STO呈显著负相关,表明随着叶片厚度和贮水组织的增加,膨压丧失点的水势变得更负.Ψtlp与MES、VB呈显著正相关,表明随着叶肉细胞和维管束比例的减少,植物忍耐干旱的能力增强.Nguyen 等[55-56]的研究强调了叶片解剖学在解释PV曲线中的重要性,突出了叶片解剖结构在决定Ψtlp、Ψ0以及ε和Cleaf等参数中的作用.通过探究不同光合作用途径植物的PV曲线参数与叶片解剖学特征之间的关系,我们不仅加深了对沙生植物在干旱胁迫下生理适应机制的理解,也为未来在不同生态环境下探索植物适应策略提供了新的视角. ...




 甘公网安备 62010202000688号
甘公网安备 62010202000688号
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}