2
2019
... 树木生长指一定时期内树木各调查因子所发生的变化,如断面积生长、直径生长、树高生长、生物量生长等[1].影响树木生长的因素很多,如邻域个体间的相互作用、树木的大小和年龄等生物因素,分析影响树木生长的因素有助于准确评估和预测森林的生长、采取适当的管理措施来改善森林生长状况[1-2]. ...
... [1-2]. ...
Competition and site weakly explain tree growth variability in undisturbed Central African moist forests
2
2023
... 树木生长指一定时期内树木各调查因子所发生的变化,如断面积生长、直径生长、树高生长、生物量生长等[1].影响树木生长的因素很多,如邻域个体间的相互作用、树木的大小和年龄等生物因素,分析影响树木生长的因素有助于准确评估和预测森林的生长、采取适当的管理措施来改善森林生长状况[1-2]. ...
... 竞争通常表现为树木个体间对光、水、矿物质元素等资源的争夺,从而影响树木的生长[2-3].康剑等[4]研究了竞争和气候对西伯利亚五针松树木径向生长的影响,发现竞争相对于气候具有更大的作用.而用来描述竞争强度的指标包括与距离无关的非完备型竞争指数和与距离有关的完备型竞争指数两类[5],很多研究表明竞争与树木生长的关系受其所选择的竞争指数类型的影响.Kahriman等[6]选取了9种非完备型和9种完备型的竞争指数,基于固定效应模型和混合效应模型的分析,发现在不包含竞争指数或以不同竞争指数作为解释变量之一时,模型评价指标R2、RMSE、AIC等存在明显不同,并发现非完备型竞争指标的加入显著提升了模型预测直径生长的解释率.然而,Ledermann[7]的研究却显示两类竞争指数与树木生长的关系没有显著差异.因而,没有一种竞争指数能够准确描述所有条件下林木竞争状况而表征其对树木生长的影响[8]. ...
竞争和气候及其交互作用对杉木人工林胸径生长的影响
1
2021
... 竞争通常表现为树木个体间对光、水、矿物质元素等资源的争夺,从而影响树木的生长[2-3].康剑等[4]研究了竞争和气候对西伯利亚五针松树木径向生长的影响,发现竞争相对于气候具有更大的作用.而用来描述竞争强度的指标包括与距离无关的非完备型竞争指数和与距离有关的完备型竞争指数两类[5],很多研究表明竞争与树木生长的关系受其所选择的竞争指数类型的影响.Kahriman等[6]选取了9种非完备型和9种完备型的竞争指数,基于固定效应模型和混合效应模型的分析,发现在不包含竞争指数或以不同竞争指数作为解释变量之一时,模型评价指标R2、RMSE、AIC等存在明显不同,并发现非完备型竞争指标的加入显著提升了模型预测直径生长的解释率.然而,Ledermann[7]的研究却显示两类竞争指数与树木生长的关系没有显著差异.因而,没有一种竞争指数能够准确描述所有条件下林木竞争状况而表征其对树木生长的影响[8]. ...
竞争和气候对新疆阿尔泰山西伯利亚五针松树木径向生长的影响
2
2020
... 竞争通常表现为树木个体间对光、水、矿物质元素等资源的争夺,从而影响树木的生长[2-3].康剑等[4]研究了竞争和气候对西伯利亚五针松树木径向生长的影响,发现竞争相对于气候具有更大的作用.而用来描述竞争强度的指标包括与距离无关的非完备型竞争指数和与距离有关的完备型竞争指数两类[5],很多研究表明竞争与树木生长的关系受其所选择的竞争指数类型的影响.Kahriman等[6]选取了9种非完备型和9种完备型的竞争指数,基于固定效应模型和混合效应模型的分析,发现在不包含竞争指数或以不同竞争指数作为解释变量之一时,模型评价指标R2、RMSE、AIC等存在明显不同,并发现非完备型竞争指标的加入显著提升了模型预测直径生长的解释率.然而,Ledermann[7]的研究却显示两类竞争指数与树木生长的关系没有显著差异.因而,没有一种竞争指数能够准确描述所有条件下林木竞争状况而表征其对树木生长的影响[8]. ...
... 不同树木个体间的竞争,通过影响对光照、水分、养分等资源的获取而影响着树木的生长,是制约树木生长的重要因素[70-71].有学者在研究不同竞争强度下欧洲赤松(Pinus sylvestris L.)树木径向生长对气候的响应时发现,树木径向生长和竞争之间呈负指数关系,竞争强度的增加使树木径向生长速率放缓[72].中国亚热带地区马尾松(Pinus massoniana)的生长量也随竞争指数增加而降低[73].刘耀凤等[71]、窦啸文等[74]和康剑等[4]在研究中也发现竞争对树木生长的类似效应.但上述研究大都只是直接选取了常用的竞争指数来描述林木个体间相互作用,这样或许不能准确地代表林木间实际竞争强度,产生有偏估计.Britton等[75]发现当不能正确描述林木生长和林木大小间关系(绝对生长量和生长率)时,不考虑竞争指数的类型,直接选用一个常用的竞争指数可能会夸大断面积生长过程中竞争的作用.基于此,为确定最优竞争指数,本研究考虑了28种基于距离的竞争指数和18种与距离无关的竞争指数,分析其对沙地樟子松树木生长的影响.结果表明,树木生长和不同竞争指数的相关性存在较大差异,即林木生长对不同竞争指数表征的竞争强度的响应存在较大差异,进一步证明了考虑竞争指数类型的必要性. ...
基于综合竞争指数和数量化理论模型的红松人工林立地质量评价
1
2021
... 竞争通常表现为树木个体间对光、水、矿物质元素等资源的争夺,从而影响树木的生长[2-3].康剑等[4]研究了竞争和气候对西伯利亚五针松树木径向生长的影响,发现竞争相对于气候具有更大的作用.而用来描述竞争强度的指标包括与距离无关的非完备型竞争指数和与距离有关的完备型竞争指数两类[5],很多研究表明竞争与树木生长的关系受其所选择的竞争指数类型的影响.Kahriman等[6]选取了9种非完备型和9种完备型的竞争指数,基于固定效应模型和混合效应模型的分析,发现在不包含竞争指数或以不同竞争指数作为解释变量之一时,模型评价指标R2、RMSE、AIC等存在明显不同,并发现非完备型竞争指标的加入显著提升了模型预测直径生长的解释率.然而,Ledermann[7]的研究却显示两类竞争指数与树木生长的关系没有显著差异.因而,没有一种竞争指数能够准确描述所有条件下林木竞争状况而表征其对树木生长的影响[8]. ...
A novel approach to selecting a competition index:the effect of competition on individual-tree diameter growth of Calabrian pine
1
2018
... 竞争通常表现为树木个体间对光、水、矿物质元素等资源的争夺,从而影响树木的生长[2-3].康剑等[4]研究了竞争和气候对西伯利亚五针松树木径向生长的影响,发现竞争相对于气候具有更大的作用.而用来描述竞争强度的指标包括与距离无关的非完备型竞争指数和与距离有关的完备型竞争指数两类[5],很多研究表明竞争与树木生长的关系受其所选择的竞争指数类型的影响.Kahriman等[6]选取了9种非完备型和9种完备型的竞争指数,基于固定效应模型和混合效应模型的分析,发现在不包含竞争指数或以不同竞争指数作为解释变量之一时,模型评价指标R2、RMSE、AIC等存在明显不同,并发现非完备型竞争指标的加入显著提升了模型预测直径生长的解释率.然而,Ledermann[7]的研究却显示两类竞争指数与树木生长的关系没有显著差异.因而,没有一种竞争指数能够准确描述所有条件下林木竞争状况而表征其对树木生长的影响[8]. ...
Evaluating the performance of semi-distance-independent competition indices in predicting the basal area growth of individual trees
1
2010
... 竞争通常表现为树木个体间对光、水、矿物质元素等资源的争夺,从而影响树木的生长[2-3].康剑等[4]研究了竞争和气候对西伯利亚五针松树木径向生长的影响,发现竞争相对于气候具有更大的作用.而用来描述竞争强度的指标包括与距离无关的非完备型竞争指数和与距离有关的完备型竞争指数两类[5],很多研究表明竞争与树木生长的关系受其所选择的竞争指数类型的影响.Kahriman等[6]选取了9种非完备型和9种完备型的竞争指数,基于固定效应模型和混合效应模型的分析,发现在不包含竞争指数或以不同竞争指数作为解释变量之一时,模型评价指标R2、RMSE、AIC等存在明显不同,并发现非完备型竞争指标的加入显著提升了模型预测直径生长的解释率.然而,Ledermann[7]的研究却显示两类竞争指数与树木生长的关系没有显著差异.因而,没有一种竞争指数能够准确描述所有条件下林木竞争状况而表征其对树木生长的影响[8]. ...
Evaluating tree competition indices as predictors of basal area increment in western Montana forests
3
2011
... 竞争通常表现为树木个体间对光、水、矿物质元素等资源的争夺,从而影响树木的生长[2-3].康剑等[4]研究了竞争和气候对西伯利亚五针松树木径向生长的影响,发现竞争相对于气候具有更大的作用.而用来描述竞争强度的指标包括与距离无关的非完备型竞争指数和与距离有关的完备型竞争指数两类[5],很多研究表明竞争与树木生长的关系受其所选择的竞争指数类型的影响.Kahriman等[6]选取了9种非完备型和9种完备型的竞争指数,基于固定效应模型和混合效应模型的分析,发现在不包含竞争指数或以不同竞争指数作为解释变量之一时,模型评价指标R2、RMSE、AIC等存在明显不同,并发现非完备型竞争指标的加入显著提升了模型预测直径生长的解释率.然而,Ledermann[7]的研究却显示两类竞争指数与树木生长的关系没有显著差异.因而,没有一种竞争指数能够准确描述所有条件下林木竞争状况而表征其对树木生长的影响[8]. ...
... 通常用一些数学公式量化个体间的竞争强度,称之为竞争指数,代表每棵树木受周围个体影响的大小.为比较不同竞争指数的相对大小,本研究将部分竞争指数取倒数调整数值大小,竞争指数越小表示目标木遭受周围竞争木的压力越小,反之越大.研究通过文献收集了不同学者提出的竞争指数,计算过程如下:首先确定对象木和竞争木;然后根据不同的竞争指数公式计算竞争指数[25].对象木所承受的竞争压力主要源于周围一定距离范围内的竞争木,超出此范围的竞争压力逐渐减弱或消失[26].根据前期研究,10 m是确定沙地樟子松林木竞争最适宜的范围[18].同时,为避免边缘效应,本研究选择样地中心30 m×30 m区域内胸径≥5 cm的170株树木为对象木,每个对象木半径10 m范围内树木为竞争木.本文选取了46种单木竞争指数[8,27-49],其中包括28种基于距离的竞争指数和18种与距离无关的竞争指数,具体如表2所列. ...
... Individual tree competition index
Table 2| 竞争指数 | 文献 | 竞争指数 | 文献 | 竞争指数 | 文献 |
|---|
| [27] | | [29] | | [44] |
| [28] | | [29] | | [45] |
| [29] | | [29] | | [46] |
| [29] | | [8] | | [46] |
| [30] | | [31] | | [46] |
| [29] | | [36] | | [47] |
| [31] | | [36] | | [48] |
| [32] | | [37] | | [48] |
| [33] | | [38] | | [48] |
| [34] | | [39] | | [48] |
| [35] | | [40] | | [49] |
| [35] | | [41] | | [48] |
| [35] | | [42] | | [49] |
| [35] | | [43] | | [36] |
| [35] | | [43] | | |
| [35] | | [43] | | |
注:CI1~CI28为基于距离的竞争指数,CI29~CI46是与距离无关的竞争指数;i代表目标木,j代表竞争木;n为目标木i所在结构单元内竞争木数量;distij 为竞争木j和目标木i间的距离;di 为目标木胸径,dj 为竞争木胸径,为结构单元内树木平均胸径,q为样地内树木胸径均方根;gi 、gmax、分别为目标木胸高断面积、结构单元内树木胸高断面积的最大值、平均值;hj 、hi 、分别为竞争木、目标木的树高、样地内树高平均值;CRi 为目标木冠幅,CRj 为竞争木冠幅,为结构单元内树木平均冠幅,CLi 是目标木冠长;aj 、ai 分别是竞争木和目标木的冠幅面积,Oij 、OLij 分别是目标木和竞争木的树冠重叠面积、重叠部分最宽处长度;CrownVi 是目标木树冠体积,CSAi 是目标木树冠表面积. ...
天目山常绿阔叶林空间结构与地形因子的关系
1
2021
... 林分结构综合反映了树木生长、竞争、更新等林分发育过程和人类经营活动的影响[9].林分结构多样性可衡量不同结构属性的相对复杂程度与均匀程度,包含林分物种多样性、林木大小分化多样性、林木空间分布多样性等,常通过Simpson指数、Shannon-Wiener指数、偏度、基尼系数、大小比数、林木大小分异指数、变异系数等多样性指数来表征[10].不同的林分结构对树木生长的影响存在着较大差异.李新建等[11]利用22个林分结构指标和3个林分生长量指标分析了栎类天然林林分结构对生长的影响,发现并不是所有结构指标与林分生长显著相关,且影响途径不一致,例如混交度、林分密度、物种组成间接影响蓄积生长量.Lin等[12]发现森林树木生长对林分结构的响应在不同树种间存在差异,且与林木研究时间段内初始大小密切相关.因此,充分考虑林分结构的差异性,对于准确评估森林生长动态具有重要的科学意义. ...
林分结构多样性研究进展
3
2020
... 林分结构综合反映了树木生长、竞争、更新等林分发育过程和人类经营活动的影响[9].林分结构多样性可衡量不同结构属性的相对复杂程度与均匀程度,包含林分物种多样性、林木大小分化多样性、林木空间分布多样性等,常通过Simpson指数、Shannon-Wiener指数、偏度、基尼系数、大小比数、林木大小分异指数、变异系数等多样性指数来表征[10].不同的林分结构对树木生长的影响存在着较大差异.李新建等[11]利用22个林分结构指标和3个林分生长量指标分析了栎类天然林林分结构对生长的影响,发现并不是所有结构指标与林分生长显著相关,且影响途径不一致,例如混交度、林分密度、物种组成间接影响蓄积生长量.Lin等[12]发现森林树木生长对林分结构的响应在不同树种间存在差异,且与林木研究时间段内初始大小密切相关.因此,充分考虑林分结构的差异性,对于准确评估森林生长动态具有重要的科学意义. ...
... 作为描述林分生长变异性的重要指标,林分结构在很大程度上影响着森林生长的时空动态,而森林经营很大程度上通过调控林分的结构实现[50-51].林分结构多样性的表征方式多样,如树种多样性、树木大小多样性、林木分布格局多样性等,空间和非空间分析法是量化林分结构多样性指标常用的两种方法[52].研究对象为沙地樟子松纯林,选取香农-威纳指数[53]、辛普森指数[54]、大小比数[55]、大小分异指数[56]、基尼系数[57]、变异系数[57]、偏度[10]、峰度[58]、Clark-Evans指数[59]等9种林分结构多样性指数来描述对象木对应结构单元的林分结构状况(表3).在计算林分结构多样性指数时,树木胸径按3 cm的径阶距划分. ...
... Stand structure diversity indices
Table 3| 名称 | 公式 | 意义 | 文献 |
|---|
| 香农-威纳指数 | | SW越大,树木大小差异越大 | [53] |
| 辛普森指数 | | S数大,代表树木大小分化程度高 | [54] |
| 大小比数 | | NCI越大,表示相邻木相比于目标木优势越大 | [55] |
| 大小分异指数 | | DFI越大,代表树木大小差异越大 | [56] |
| 基尼系数 | | GC越大,代表树木大小分布越不平衡 | [57] |
| 变异系数 | | CV越小,代表林木大小差异越小 | [57] |
| 偏度 | | SK越大,代表树木大小分布偏态程度越高 | [10] |
| 峰度 | | Kg越大,树木胸径最大值越大,分布越陡峭 | [58] |
| Clark-Evans指数 | | 判断林木种群空间分布格局(聚集、随机、均匀)的指数 | [59] |
注:n为树木i所在结构单元内竞争木数量;di 为目标木胸径,dj 为竞争木胸径,为结构单元内树木平均胸径,σd 为结构单元内林木胸径标准差;gi 为目标木胸高断面积;S为径阶数目;pm 为第m个径阶林木株数占相应结构单元林木总株数的比例.Vij 为目标木i比它的第j株相邻木小时,Vij =1;树木i比它的第j株相邻木大时,Vij =0.ri 为树木i与其最近一棵树的距离;A为结构单元样圆的面积.CE<1时为聚集分布,CE=1时为随机分布,CE>1时为均匀分布. ...
湖南栎类天然林林分结构对林分生长的影响研究
1
2022
... 林分结构综合反映了树木生长、竞争、更新等林分发育过程和人类经营活动的影响[9].林分结构多样性可衡量不同结构属性的相对复杂程度与均匀程度,包含林分物种多样性、林木大小分化多样性、林木空间分布多样性等,常通过Simpson指数、Shannon-Wiener指数、偏度、基尼系数、大小比数、林木大小分异指数、变异系数等多样性指数来表征[10].不同的林分结构对树木生长的影响存在着较大差异.李新建等[11]利用22个林分结构指标和3个林分生长量指标分析了栎类天然林林分结构对生长的影响,发现并不是所有结构指标与林分生长显著相关,且影响途径不一致,例如混交度、林分密度、物种组成间接影响蓄积生长量.Lin等[12]发现森林树木生长对林分结构的响应在不同树种间存在差异,且与林木研究时间段内初始大小密切相关.因此,充分考虑林分结构的差异性,对于准确评估森林生长动态具有重要的科学意义. ...
Effects of local neighbourhood structure on radial growth of Picea crassifolia Kom.and Betula platyphylla Suk.plantations in the loess alpine region,China
3
2021
... 林分结构综合反映了树木生长、竞争、更新等林分发育过程和人类经营活动的影响[9].林分结构多样性可衡量不同结构属性的相对复杂程度与均匀程度,包含林分物种多样性、林木大小分化多样性、林木空间分布多样性等,常通过Simpson指数、Shannon-Wiener指数、偏度、基尼系数、大小比数、林木大小分异指数、变异系数等多样性指数来表征[10].不同的林分结构对树木生长的影响存在着较大差异.李新建等[11]利用22个林分结构指标和3个林分生长量指标分析了栎类天然林林分结构对生长的影响,发现并不是所有结构指标与林分生长显著相关,且影响途径不一致,例如混交度、林分密度、物种组成间接影响蓄积生长量.Lin等[12]发现森林树木生长对林分结构的响应在不同树种间存在差异,且与林木研究时间段内初始大小密切相关.因此,充分考虑林分结构的差异性,对于准确评估森林生长动态具有重要的科学意义. ...
... 森林结构体现为树木个体及其种类、大小、分布等属性的连接方式,它决定着森林功能,是树木生长等生态过程的主要驱动因素.在一定范围内,树种多样性及大小差异越大代表结构多样性越高[52,65]. 很多研究表明结构越复杂的森林具有越高的生产力[66-69].Peter等[66]发现,树种丰富度与森林生产力之间存在明显的正反馈.而Elizabeth等[67]也发现结构多样性是预测森林生产力的可靠指标,在一定程度上说明结构复杂性对树木生长的影响不可忽略.较高的结构多样性,也有利于减缓干旱对树木生长的不利影响.有研究显示,全球气候暖干化会导致森林生长衰退,具有高物种多样性的森林因干旱而导致的树木生长损失更少,结构复杂性不是单独发挥作用的[68].然而,对于天然纯林来说,虽然树种单一,但结构多样性对树木生长也具有较大的影响.例如,Sha等[12]对青海云杉(Picea crassifolia Kom.)的研究表明树木生长受树木大小分异指数的显著影响.我们的研究也表明,树木大小比数(NCI)、树木大小分异指数(DFI)以及Clark-Evans指数(CE)也是影响天然沙地樟子松纯林树木生长的重要因素.樟子松树木与其周围相邻木的大小比数及分异指数的值越大,可能表明樟子松树木受周围相邻木的竞争压力更大,进而导致树木生长量降低.而Clark-Evans指数表征着树木个体在空间上的分布状况,本研究表明樟子松树木生长随Clark-Evans指数的增加而显著降低,这可能与本研究中树木的分布格局对竞争的影响有关.我们发现胸径较大的樟子松树木多呈聚集分布状态,而胸径较小的树木更多呈均匀分布,这或许是导致这一结果的重要原因.然而,林分结构复杂性对于树木生长的影响并不是固定的.Zeller等[69]发现,随着森林演替的进行,结构复杂性对树木生长的影响由早期的抑制逐渐转变为促进作用.未来的研究,应结合树木年轮学等的方法,关注更长时间尺度上林分结构复杂性及其对树木生长影响的变化. ...
... 树木生长往往同时受到很多因素,诸如气候、土壤、地形、林分结构等的综合影响.例如窦啸文等[74]发现天目山常绿阔叶林树木径向生长同时受到胸径、冠幅和竞争的共同作用.然而,这些因素在决定树木生长中的相对重要性却各不相同.韩大校等[76]在黑龙江凉水自然保护区阔叶红松林的研究中发现, 大径级树木的径向生长主要受地形因素的影响,而小径级树木的径向生长主要受到竞争的影响.也有研究认为树木的竞争作用,而非气候是限制树木生长的最主要因素[73].我们研究发现,当同时考虑林木本身属性、林分结构、单木竞争时,竞争与结构多样性显著影响了沙地樟子松树木径向生长.竞争是影响樟子松树木生长的最重要因素,其解释了约三分之一的变异.而结构多样性因素,即树木大小分异指数解释了樟子松生长约12%的变异性,而树木自身的特性,包括树木的冠幅和高径比共同解释了树木生长约四分之一的变异性,这与前人研究[17-18,69,74]基本一致.同时,我们也发现,林分结构多样性与竞争间存在交互作用[12,19],共同影响了樟子松树木的生长,气候同时也影响了树木生长动态[77],未来的研究应重视不同因子的交互作用对树木生长的影响.另外,本研究的样本量相对较小,导致研究结果可能存在一定局限性,未来研究需要从更长时间尺度、更大地理范围开展研究,以增强结果的普适性与可靠性. ...
2
2018
... 樟子松(Pinussylvestris var. mongolica)作为中国内蒙古呼伦贝尔沙地重要的建群种,在防风固沙、水源涵养、固碳释氧等方面发挥着非常重要的生态服务功能[13].目前,中国在呼伦贝尔沙地樟子松树木生长固碳[14]、火干扰与林分更新[15-16]、生长对气候的响应[17-20]等方面已经开展了大量的研究,但这些研究多强调气候对树木生长的影响.近年来,生态学家逐渐重视林分结构和竞争等生物学因素对树木生长的影响[17-18,21],然而对林分结构差异和树间竞争如何影响树木生长的认识仍然较弱.为此,本研究以呼伦贝尔沙地樟子松天然林为研究对象,基于野外调查和树木年轮数据,分析了46种竞争指数 (其中包含28种基于距离的竞争指数和18种与距离无关的竞争指数)和9种林分结构指数对树木生长的影响,并揭示了竞争和林分结构对树木生长的重要性,以期为沙地樟子松天然林的可持续管理与科学经营提供科学参考. ...
... 研究区位于内蒙古自治区呼伦贝尔市鄂温克旗呼伦贝尔沙地生态系统定位研究站(49°04′46.30″N、120°28′12.79″E),属于中温带半干旱大陆性气候,冬季漫长寒冷,干燥多风,夏季温和短促,降水较集中,春秋两季气候变化剧烈,昼夜温差大,降水少,多大风.年均气温-2 ℃左右,多年平均降水量332 mm,发生于7—8月,多年平均蒸发量1 174 mm,无霜期90~110 d[22-23].该区地下水储量比较丰富,但土壤持水性不佳,有机质含量较低,非常贫瘠[13,18].当地主要森林类型为沙地樟子松纯林,并伴生有白桦(Betula platyphylla)、榛子(Corylus heterophylla)等树种,以及冰草(Agropyron cristatum)、羊草(Leymus chinensis)、线叶菊(Filifolium sibiricum)等草本物种[24].土壤为砂质土壤. ...
呼伦贝尔沙地樟子松人工林乔木层固碳速率及其对气象因子的响应
1
2020
... 樟子松(Pinussylvestris var. mongolica)作为中国内蒙古呼伦贝尔沙地重要的建群种,在防风固沙、水源涵养、固碳释氧等方面发挥着非常重要的生态服务功能[13].目前,中国在呼伦贝尔沙地樟子松树木生长固碳[14]、火干扰与林分更新[15-16]、生长对气候的响应[17-20]等方面已经开展了大量的研究,但这些研究多强调气候对树木生长的影响.近年来,生态学家逐渐重视林分结构和竞争等生物学因素对树木生长的影响[17-18,21],然而对林分结构差异和树间竞争如何影响树木生长的认识仍然较弱.为此,本研究以呼伦贝尔沙地樟子松天然林为研究对象,基于野外调查和树木年轮数据,分析了46种竞争指数 (其中包含28种基于距离的竞争指数和18种与距离无关的竞争指数)和9种林分结构指数对树木生长的影响,并揭示了竞争和林分结构对树木生长的重要性,以期为沙地樟子松天然林的可持续管理与科学经营提供科学参考. ...
不同强度林火干扰对红花尔基樟子松天然林更新的影响
1
2015
... 樟子松(Pinussylvestris var. mongolica)作为中国内蒙古呼伦贝尔沙地重要的建群种,在防风固沙、水源涵养、固碳释氧等方面发挥着非常重要的生态服务功能[13].目前,中国在呼伦贝尔沙地樟子松树木生长固碳[14]、火干扰与林分更新[15-16]、生长对气候的响应[17-20]等方面已经开展了大量的研究,但这些研究多强调气候对树木生长的影响.近年来,生态学家逐渐重视林分结构和竞争等生物学因素对树木生长的影响[17-18,21],然而对林分结构差异和树间竞争如何影响树木生长的认识仍然较弱.为此,本研究以呼伦贝尔沙地樟子松天然林为研究对象,基于野外调查和树木年轮数据,分析了46种竞争指数 (其中包含28种基于距离的竞争指数和18种与距离无关的竞争指数)和9种林分结构指数对树木生长的影响,并揭示了竞争和林分结构对树木生长的重要性,以期为沙地樟子松天然林的可持续管理与科学经营提供科学参考. ...
地表火干扰下呼伦贝尔沙地樟子松林乔木层不同组分空间格局的变化
1
2009
... 樟子松(Pinussylvestris var. mongolica)作为中国内蒙古呼伦贝尔沙地重要的建群种,在防风固沙、水源涵养、固碳释氧等方面发挥着非常重要的生态服务功能[13].目前,中国在呼伦贝尔沙地樟子松树木生长固碳[14]、火干扰与林分更新[15-16]、生长对气候的响应[17-20]等方面已经开展了大量的研究,但这些研究多强调气候对树木生长的影响.近年来,生态学家逐渐重视林分结构和竞争等生物学因素对树木生长的影响[17-18,21],然而对林分结构差异和树间竞争如何影响树木生长的认识仍然较弱.为此,本研究以呼伦贝尔沙地樟子松天然林为研究对象,基于野外调查和树木年轮数据,分析了46种竞争指数 (其中包含28种基于距离的竞争指数和18种与距离无关的竞争指数)和9种林分结构指数对树木生长的影响,并揭示了竞争和林分结构对树木生长的重要性,以期为沙地樟子松天然林的可持续管理与科学经营提供科学参考. ...
沙地樟子松天然林树木竞争对其生长及气候敏感性影响
3
2019
... 樟子松(Pinussylvestris var. mongolica)作为中国内蒙古呼伦贝尔沙地重要的建群种,在防风固沙、水源涵养、固碳释氧等方面发挥着非常重要的生态服务功能[13].目前,中国在呼伦贝尔沙地樟子松树木生长固碳[14]、火干扰与林分更新[15-16]、生长对气候的响应[17-20]等方面已经开展了大量的研究,但这些研究多强调气候对树木生长的影响.近年来,生态学家逐渐重视林分结构和竞争等生物学因素对树木生长的影响[17-18,21],然而对林分结构差异和树间竞争如何影响树木生长的认识仍然较弱.为此,本研究以呼伦贝尔沙地樟子松天然林为研究对象,基于野外调查和树木年轮数据,分析了46种竞争指数 (其中包含28种基于距离的竞争指数和18种与距离无关的竞争指数)和9种林分结构指数对树木生长的影响,并揭示了竞争和林分结构对树木生长的重要性,以期为沙地樟子松天然林的可持续管理与科学经营提供科学参考. ...
... [17-18,21],然而对林分结构差异和树间竞争如何影响树木生长的认识仍然较弱.为此,本研究以呼伦贝尔沙地樟子松天然林为研究对象,基于野外调查和树木年轮数据,分析了46种竞争指数 (其中包含28种基于距离的竞争指数和18种与距离无关的竞争指数)和9种林分结构指数对树木生长的影响,并揭示了竞争和林分结构对树木生长的重要性,以期为沙地樟子松天然林的可持续管理与科学经营提供科学参考. ...
... 树木生长往往同时受到很多因素,诸如气候、土壤、地形、林分结构等的综合影响.例如窦啸文等[74]发现天目山常绿阔叶林树木径向生长同时受到胸径、冠幅和竞争的共同作用.然而,这些因素在决定树木生长中的相对重要性却各不相同.韩大校等[76]在黑龙江凉水自然保护区阔叶红松林的研究中发现, 大径级树木的径向生长主要受地形因素的影响,而小径级树木的径向生长主要受到竞争的影响.也有研究认为树木的竞争作用,而非气候是限制树木生长的最主要因素[73].我们研究发现,当同时考虑林木本身属性、林分结构、单木竞争时,竞争与结构多样性显著影响了沙地樟子松树木径向生长.竞争是影响樟子松树木生长的最重要因素,其解释了约三分之一的变异.而结构多样性因素,即树木大小分异指数解释了樟子松生长约12%的变异性,而树木自身的特性,包括树木的冠幅和高径比共同解释了树木生长约四分之一的变异性,这与前人研究[17-18,69,74]基本一致.同时,我们也发现,林分结构多样性与竞争间存在交互作用[12,19],共同影响了樟子松树木的生长,气候同时也影响了树木生长动态[77],未来的研究应重视不同因子的交互作用对树木生长的影响.另外,本研究的样本量相对较小,导致研究结果可能存在一定局限性,未来研究需要从更长时间尺度、更大地理范围开展研究,以增强结果的普适性与可靠性. ...
沙地樟子松天然林林分结构及气候对其生长的影响
4
2019
... 樟子松(Pinussylvestris var. mongolica)作为中国内蒙古呼伦贝尔沙地重要的建群种,在防风固沙、水源涵养、固碳释氧等方面发挥着非常重要的生态服务功能[13].目前,中国在呼伦贝尔沙地樟子松树木生长固碳[14]、火干扰与林分更新[15-16]、生长对气候的响应[17-20]等方面已经开展了大量的研究,但这些研究多强调气候对树木生长的影响.近年来,生态学家逐渐重视林分结构和竞争等生物学因素对树木生长的影响[17-18,21],然而对林分结构差异和树间竞争如何影响树木生长的认识仍然较弱.为此,本研究以呼伦贝尔沙地樟子松天然林为研究对象,基于野外调查和树木年轮数据,分析了46种竞争指数 (其中包含28种基于距离的竞争指数和18种与距离无关的竞争指数)和9种林分结构指数对树木生长的影响,并揭示了竞争和林分结构对树木生长的重要性,以期为沙地樟子松天然林的可持续管理与科学经营提供科学参考. ...
... 研究区位于内蒙古自治区呼伦贝尔市鄂温克旗呼伦贝尔沙地生态系统定位研究站(49°04′46.30″N、120°28′12.79″E),属于中温带半干旱大陆性气候,冬季漫长寒冷,干燥多风,夏季温和短促,降水较集中,春秋两季气候变化剧烈,昼夜温差大,降水少,多大风.年均气温-2 ℃左右,多年平均降水量332 mm,发生于7—8月,多年平均蒸发量1 174 mm,无霜期90~110 d[22-23].该区地下水储量比较丰富,但土壤持水性不佳,有机质含量较低,非常贫瘠[13,18].当地主要森林类型为沙地樟子松纯林,并伴生有白桦(Betula platyphylla)、榛子(Corylus heterophylla)等树种,以及冰草(Agropyron cristatum)、羊草(Leymus chinensis)、线叶菊(Filifolium sibiricum)等草本物种[24].土壤为砂质土壤. ...
... 通常用一些数学公式量化个体间的竞争强度,称之为竞争指数,代表每棵树木受周围个体影响的大小.为比较不同竞争指数的相对大小,本研究将部分竞争指数取倒数调整数值大小,竞争指数越小表示目标木遭受周围竞争木的压力越小,反之越大.研究通过文献收集了不同学者提出的竞争指数,计算过程如下:首先确定对象木和竞争木;然后根据不同的竞争指数公式计算竞争指数[25].对象木所承受的竞争压力主要源于周围一定距离范围内的竞争木,超出此范围的竞争压力逐渐减弱或消失[26].根据前期研究,10 m是确定沙地樟子松林木竞争最适宜的范围[18].同时,为避免边缘效应,本研究选择样地中心30 m×30 m区域内胸径≥5 cm的170株树木为对象木,每个对象木半径10 m范围内树木为竞争木.本文选取了46种单木竞争指数[8,27-49],其中包括28种基于距离的竞争指数和18种与距离无关的竞争指数,具体如表2所列. ...
... 树木生长往往同时受到很多因素,诸如气候、土壤、地形、林分结构等的综合影响.例如窦啸文等[74]发现天目山常绿阔叶林树木径向生长同时受到胸径、冠幅和竞争的共同作用.然而,这些因素在决定树木生长中的相对重要性却各不相同.韩大校等[76]在黑龙江凉水自然保护区阔叶红松林的研究中发现, 大径级树木的径向生长主要受地形因素的影响,而小径级树木的径向生长主要受到竞争的影响.也有研究认为树木的竞争作用,而非气候是限制树木生长的最主要因素[73].我们研究发现,当同时考虑林木本身属性、林分结构、单木竞争时,竞争与结构多样性显著影响了沙地樟子松树木径向生长.竞争是影响樟子松树木生长的最重要因素,其解释了约三分之一的变异.而结构多样性因素,即树木大小分异指数解释了樟子松生长约12%的变异性,而树木自身的特性,包括树木的冠幅和高径比共同解释了树木生长约四分之一的变异性,这与前人研究[17-18,69,74]基本一致.同时,我们也发现,林分结构多样性与竞争间存在交互作用[12,19],共同影响了樟子松树木的生长,气候同时也影响了树木生长动态[77],未来的研究应重视不同因子的交互作用对树木生长的影响.另外,本研究的样本量相对较小,导致研究结果可能存在一定局限性,未来研究需要从更长时间尺度、更大地理范围开展研究,以增强结果的普适性与可靠性. ...
Effect of climate and competition on radial growth of Pinus sylvestris var.mongolica forest in Hulunbuir Sandy Land of Inner Mongolia,China
1
2023
... 树木生长往往同时受到很多因素,诸如气候、土壤、地形、林分结构等的综合影响.例如窦啸文等[74]发现天目山常绿阔叶林树木径向生长同时受到胸径、冠幅和竞争的共同作用.然而,这些因素在决定树木生长中的相对重要性却各不相同.韩大校等[76]在黑龙江凉水自然保护区阔叶红松林的研究中发现, 大径级树木的径向生长主要受地形因素的影响,而小径级树木的径向生长主要受到竞争的影响.也有研究认为树木的竞争作用,而非气候是限制树木生长的最主要因素[73].我们研究发现,当同时考虑林木本身属性、林分结构、单木竞争时,竞争与结构多样性显著影响了沙地樟子松树木径向生长.竞争是影响樟子松树木生长的最重要因素,其解释了约三分之一的变异.而结构多样性因素,即树木大小分异指数解释了樟子松生长约12%的变异性,而树木自身的特性,包括树木的冠幅和高径比共同解释了树木生长约四分之一的变异性,这与前人研究[17-18,69,74]基本一致.同时,我们也发现,林分结构多样性与竞争间存在交互作用[12,19],共同影响了樟子松树木的生长,气候同时也影响了树木生长动态[77],未来的研究应重视不同因子的交互作用对树木生长的影响.另外,本研究的样本量相对较小,导致研究结果可能存在一定局限性,未来研究需要从更长时间尺度、更大地理范围开展研究,以增强结果的普适性与可靠性. ...
呼伦贝尔沙地樟子松生产力及其对气候因子的响应
1
2022
... 樟子松(Pinussylvestris var. mongolica)作为中国内蒙古呼伦贝尔沙地重要的建群种,在防风固沙、水源涵养、固碳释氧等方面发挥着非常重要的生态服务功能[13].目前,中国在呼伦贝尔沙地樟子松树木生长固碳[14]、火干扰与林分更新[15-16]、生长对气候的响应[17-20]等方面已经开展了大量的研究,但这些研究多强调气候对树木生长的影响.近年来,生态学家逐渐重视林分结构和竞争等生物学因素对树木生长的影响[17-18,21],然而对林分结构差异和树间竞争如何影响树木生长的认识仍然较弱.为此,本研究以呼伦贝尔沙地樟子松天然林为研究对象,基于野外调查和树木年轮数据,分析了46种竞争指数 (其中包含28种基于距离的竞争指数和18种与距离无关的竞争指数)和9种林分结构指数对树木生长的影响,并揭示了竞争和林分结构对树木生长的重要性,以期为沙地樟子松天然林的可持续管理与科学经营提供科学参考. ...
沙地樟子松天然纯林的结构特征
1
2016
... 樟子松(Pinussylvestris var. mongolica)作为中国内蒙古呼伦贝尔沙地重要的建群种,在防风固沙、水源涵养、固碳释氧等方面发挥着非常重要的生态服务功能[13].目前,中国在呼伦贝尔沙地樟子松树木生长固碳[14]、火干扰与林分更新[15-16]、生长对气候的响应[17-20]等方面已经开展了大量的研究,但这些研究多强调气候对树木生长的影响.近年来,生态学家逐渐重视林分结构和竞争等生物学因素对树木生长的影响[17-18,21],然而对林分结构差异和树间竞争如何影响树木生长的认识仍然较弱.为此,本研究以呼伦贝尔沙地樟子松天然林为研究对象,基于野外调查和树木年轮数据,分析了46种竞争指数 (其中包含28种基于距离的竞争指数和18种与距离无关的竞争指数)和9种林分结构指数对树木生长的影响,并揭示了竞争和林分结构对树木生长的重要性,以期为沙地樟子松天然林的可持续管理与科学经营提供科学参考. ...
1
1963
... 研究区位于内蒙古自治区呼伦贝尔市鄂温克旗呼伦贝尔沙地生态系统定位研究站(49°04′46.30″N、120°28′12.79″E),属于中温带半干旱大陆性气候,冬季漫长寒冷,干燥多风,夏季温和短促,降水较集中,春秋两季气候变化剧烈,昼夜温差大,降水少,多大风.年均气温-2 ℃左右,多年平均降水量332 mm,发生于7—8月,多年平均蒸发量1 174 mm,无霜期90~110 d[22-23].该区地下水储量比较丰富,但土壤持水性不佳,有机质含量较低,非常贫瘠[13,18].当地主要森林类型为沙地樟子松纯林,并伴生有白桦(Betula platyphylla)、榛子(Corylus heterophylla)等树种,以及冰草(Agropyron cristatum)、羊草(Leymus chinensis)、线叶菊(Filifolium sibiricum)等草本物种[24].土壤为砂质土壤. ...
红花尔基沙地樟子松种群优势度增长动态及自疏规律的研究
1
1999
... 研究区位于内蒙古自治区呼伦贝尔市鄂温克旗呼伦贝尔沙地生态系统定位研究站(49°04′46.30″N、120°28′12.79″E),属于中温带半干旱大陆性气候,冬季漫长寒冷,干燥多风,夏季温和短促,降水较集中,春秋两季气候变化剧烈,昼夜温差大,降水少,多大风.年均气温-2 ℃左右,多年平均降水量332 mm,发生于7—8月,多年平均蒸发量1 174 mm,无霜期90~110 d[22-23].该区地下水储量比较丰富,但土壤持水性不佳,有机质含量较低,非常贫瘠[13,18].当地主要森林类型为沙地樟子松纯林,并伴生有白桦(Betula platyphylla)、榛子(Corylus heterophylla)等树种,以及冰草(Agropyron cristatum)、羊草(Leymus chinensis)、线叶菊(Filifolium sibiricum)等草本物种[24].土壤为砂质土壤. ...
1
2005
... 研究区位于内蒙古自治区呼伦贝尔市鄂温克旗呼伦贝尔沙地生态系统定位研究站(49°04′46.30″N、120°28′12.79″E),属于中温带半干旱大陆性气候,冬季漫长寒冷,干燥多风,夏季温和短促,降水较集中,春秋两季气候变化剧烈,昼夜温差大,降水少,多大风.年均气温-2 ℃左右,多年平均降水量332 mm,发生于7—8月,多年平均蒸发量1 174 mm,无霜期90~110 d[22-23].该区地下水储量比较丰富,但土壤持水性不佳,有机质含量较低,非常贫瘠[13,18].当地主要森林类型为沙地樟子松纯林,并伴生有白桦(Betula platyphylla)、榛子(Corylus heterophylla)等树种,以及冰草(Agropyron cristatum)、羊草(Leymus chinensis)、线叶菊(Filifolium sibiricum)等草本物种[24].土壤为砂质土壤. ...
单木竞争的研究概述
1
2021
... 通常用一些数学公式量化个体间的竞争强度,称之为竞争指数,代表每棵树木受周围个体影响的大小.为比较不同竞争指数的相对大小,本研究将部分竞争指数取倒数调整数值大小,竞争指数越小表示目标木遭受周围竞争木的压力越小,反之越大.研究通过文献收集了不同学者提出的竞争指数,计算过程如下:首先确定对象木和竞争木;然后根据不同的竞争指数公式计算竞争指数[25].对象木所承受的竞争压力主要源于周围一定距离范围内的竞争木,超出此范围的竞争压力逐渐减弱或消失[26].根据前期研究,10 m是确定沙地樟子松林木竞争最适宜的范围[18].同时,为避免边缘效应,本研究选择样地中心30 m×30 m区域内胸径≥5 cm的170株树木为对象木,每个对象木半径10 m范围内树木为竞争木.本文选取了46种单木竞争指数[8,27-49],其中包括28种基于距离的竞争指数和18种与距离无关的竞争指数,具体如表2所列. ...
沙地樟子松天然林南缘分布区林木竞争、空间格局及其更新特征
1
2019
... 通常用一些数学公式量化个体间的竞争强度,称之为竞争指数,代表每棵树木受周围个体影响的大小.为比较不同竞争指数的相对大小,本研究将部分竞争指数取倒数调整数值大小,竞争指数越小表示目标木遭受周围竞争木的压力越小,反之越大.研究通过文献收集了不同学者提出的竞争指数,计算过程如下:首先确定对象木和竞争木;然后根据不同的竞争指数公式计算竞争指数[25].对象木所承受的竞争压力主要源于周围一定距离范围内的竞争木,超出此范围的竞争压力逐渐减弱或消失[26].根据前期研究,10 m是确定沙地樟子松林木竞争最适宜的范围[18].同时,为避免边缘效应,本研究选择样地中心30 m×30 m区域内胸径≥5 cm的170株树木为对象木,每个对象木半径10 m范围内树木为竞争木.本文选取了46种单木竞争指数[8,27-49],其中包括28种基于距离的竞争指数和18种与距离无关的竞争指数,具体如表2所列. ...
A simulation model for managing jack-pine stands
2
1974
... 通常用一些数学公式量化个体间的竞争强度,称之为竞争指数,代表每棵树木受周围个体影响的大小.为比较不同竞争指数的相对大小,本研究将部分竞争指数取倒数调整数值大小,竞争指数越小表示目标木遭受周围竞争木的压力越小,反之越大.研究通过文献收集了不同学者提出的竞争指数,计算过程如下:首先确定对象木和竞争木;然后根据不同的竞争指数公式计算竞争指数[25].对象木所承受的竞争压力主要源于周围一定距离范围内的竞争木,超出此范围的竞争压力逐渐减弱或消失[26].根据前期研究,10 m是确定沙地樟子松林木竞争最适宜的范围[18].同时,为避免边缘效应,本研究选择样地中心30 m×30 m区域内胸径≥5 cm的170株树木为对象木,每个对象木半径10 m范围内树木为竞争木.本文选取了46种单木竞争指数[8,27-49],其中包括28种基于距离的竞争指数和18种与距离无关的竞争指数,具体如表2所列. ...
... Individual tree competition index
Table 2| 竞争指数 | 文献 | 竞争指数 | 文献 | 竞争指数 | 文献 |
|---|
| [27] | | [29] | | [44] |
| [28] | | [29] | | [45] |
| [29] | | [29] | | [46] |
| [29] | | [8] | | [46] |
| [30] | | [31] | | [46] |
| [29] | | [36] | | [47] |
| [31] | | [36] | | [48] |
| [32] | | [37] | | [48] |
| [33] | | [38] | | [48] |
| [34] | | [39] | | [48] |
| [35] | | [40] | | [49] |
| [35] | | [41] | | [48] |
| [35] | | [42] | | [49] |
| [35] | | [43] | | [36] |
| [35] | | [43] | | |
| [35] | | [43] | | |
注:CI1~CI28为基于距离的竞争指数,CI29~CI46是与距离无关的竞争指数;i代表目标木,j代表竞争木;n为目标木i所在结构单元内竞争木数量;distij 为竞争木j和目标木i间的距离;di 为目标木胸径,dj 为竞争木胸径,为结构单元内树木平均胸径,q为样地内树木胸径均方根;gi 、gmax、分别为目标木胸高断面积、结构单元内树木胸高断面积的最大值、平均值;hj 、hi 、分别为竞争木、目标木的树高、样地内树高平均值;CRi 为目标木冠幅,CRj 为竞争木冠幅,为结构单元内树木平均冠幅,CLi 是目标木冠长;aj 、ai 分别是竞争木和目标木的冠幅面积,Oij 、OLij 分别是目标木和竞争木的树冠重叠面积、重叠部分最宽处长度;CrownVi 是目标木树冠体积,CSAi 是目标木树冠表面积. ...
A comparison of competition measures and growth models for predicting plantation red pine diameter and height growth
1
1984
... Individual tree competition index
Table 2| 竞争指数 | 文献 | 竞争指数 | 文献 | 竞争指数 | 文献 |
|---|
| [27] | | [29] | | [44] |
| [28] | | [29] | | [45] |
| [29] | | [29] | | [46] |
| [29] | | [8] | | [46] |
| [30] | | [31] | | [46] |
| [29] | | [36] | | [47] |
| [31] | | [36] | | [48] |
| [32] | | [37] | | [48] |
| [33] | | [38] | | [48] |
| [34] | | [39] | | [48] |
| [35] | | [40] | | [49] |
| [35] | | [41] | | [48] |
| [35] | | [42] | | [49] |
| [35] | | [43] | | [36] |
| [35] | | [43] | | |
| [35] | | [43] | | |
注:CI1~CI28为基于距离的竞争指数,CI29~CI46是与距离无关的竞争指数;i代表目标木,j代表竞争木;n为目标木i所在结构单元内竞争木数量;distij 为竞争木j和目标木i间的距离;di 为目标木胸径,dj 为竞争木胸径,为结构单元内树木平均胸径,q为样地内树木胸径均方根;gi 、gmax、分别为目标木胸高断面积、结构单元内树木胸高断面积的最大值、平均值;hj 、hi 、分别为竞争木、目标木的树高、样地内树高平均值;CRi 为目标木冠幅,CRj 为竞争木冠幅,为结构单元内树木平均冠幅,CLi 是目标木冠长;aj 、ai 分别是竞争木和目标木的冠幅面积,Oij 、OLij 分别是目标木和竞争木的树冠重叠面积、重叠部分最宽处长度;CrownVi 是目标木树冠体积,CSAi 是目标木树冠表面积. ...
Structure and asymmetry of tree crowns in relation to local competition in a natural mature Scots pine forest
6
1997
... Individual tree competition index
Table 2| 竞争指数 | 文献 | 竞争指数 | 文献 | 竞争指数 | 文献 |
|---|
| [27] | | [29] | | [44] |
| [28] | | [29] | | [45] |
| [29] | | [29] | | [46] |
| [29] | | [8] | | [46] |
| [30] | | [31] | | [46] |
| [29] | | [36] | | [47] |
| [31] | | [36] | | [48] |
| [32] | | [37] | | [48] |
| [33] | | [38] | | [48] |
| [34] | | [39] | | [48] |
| [35] | | [40] | | [49] |
| [35] | | [41] | | [48] |
| [35] | | [42] | | [49] |
| [35] | | [43] | | [36] |
| [35] | | [43] | | |
| [35] | | [43] | | |
注:CI1~CI28为基于距离的竞争指数,CI29~CI46是与距离无关的竞争指数;i代表目标木,j代表竞争木;n为目标木i所在结构单元内竞争木数量;distij 为竞争木j和目标木i间的距离;di 为目标木胸径,dj 为竞争木胸径,为结构单元内树木平均胸径,q为样地内树木胸径均方根;gi 、gmax、分别为目标木胸高断面积、结构单元内树木胸高断面积的最大值、平均值;hj 、hi 、分别为竞争木、目标木的树高、样地内树高平均值;CRi 为目标木冠幅,CRj 为竞争木冠幅,为结构单元内树木平均冠幅,CLi 是目标木冠长;aj 、ai 分别是竞争木和目标木的冠幅面积,Oij 、OLij 分别是目标木和竞争木的树冠重叠面积、重叠部分最宽处长度;CrownVi 是目标木树冠体积,CSAi 是目标木树冠表面积. ...
... [
29]
| [45] | | [29] | | [29] | | [46] |
| [29] | | [8] | | [46] |
| [30] | | [31] | | [46] |
| [29] | | [36] | | [47] |
| [31] | | [36] | | [48] |
| [32] | | [37] | | [48] |
| [33] | | [38] | | [48] |
| [34] | | [39] | | [48] |
| [35] | | [40] | | [49] |
| [35] | | [41] | | [48] |
| [35] | | [42] | | [49] |
| [35] | | [43] | | [36] |
| [35] | | [43] | | |
| [35] | | [43] | | |
注:CI1~CI28为基于距离的竞争指数,CI29~CI46是与距离无关的竞争指数;i代表目标木,j代表竞争木;n为目标木i所在结构单元内竞争木数量;distij 为竞争木j和目标木i间的距离;di 为目标木胸径,dj 为竞争木胸径,为结构单元内树木平均胸径,q为样地内树木胸径均方根;gi 、gmax、分别为目标木胸高断面积、结构单元内树木胸高断面积的最大值、平均值;hj 、hi 、分别为竞争木、目标木的树高、样地内树高平均值;CRi 为目标木冠幅,CRj 为竞争木冠幅,为结构单元内树木平均冠幅,CLi 是目标木冠长;aj 、ai 分别是竞争木和目标木的冠幅面积,Oij 、OLij 分别是目标木和竞争木的树冠重叠面积、重叠部分最宽处长度;CrownVi 是目标木树冠体积,CSAi 是目标木树冠表面积. ...
... [
29]
| [29] | | [46] | | [29] | | [8] | | [46] |
| [30] | | [31] | | [46] |
| [29] | | [36] | | [47] |
| [31] | | [36] | | [48] |
| [32] | | [37] | | [48] |
| [33] | | [38] | | [48] |
| [34] | | [39] | | [48] |
| [35] | | [40] | | [49] |
| [35] | | [41] | | [48] |
| [35] | | [42] | | [49] |
| [35] | | [43] | | [36] |
| [35] | | [43] | | |
| [35] | | [43] | | |
注:CI1~CI28为基于距离的竞争指数,CI29~CI46是与距离无关的竞争指数;i代表目标木,j代表竞争木;n为目标木i所在结构单元内竞争木数量;distij 为竞争木j和目标木i间的距离;di 为目标木胸径,dj 为竞争木胸径,为结构单元内树木平均胸径,q为样地内树木胸径均方根;gi 、gmax、分别为目标木胸高断面积、结构单元内树木胸高断面积的最大值、平均值;hj 、hi 、分别为竞争木、目标木的树高、样地内树高平均值;CRi 为目标木冠幅,CRj 为竞争木冠幅,为结构单元内树木平均冠幅,CLi 是目标木冠长;aj 、ai 分别是竞争木和目标木的冠幅面积,Oij 、OLij 分别是目标木和竞争木的树冠重叠面积、重叠部分最宽处长度;CrownVi 是目标木树冠体积,CSAi 是目标木树冠表面积. ...
... [
29]
| [46] | | [29] | | [8] | | [46] |
| [30] | | [31] | | [46] |
| [29] | | [36] | | [47] |
| [31] | | [36] | | [48] |
| [32] | | [37] | | [48] |
| [33] | | [38] | | [48] |
| [34] | | [39] | | [48] |
| [35] | | [40] | | [49] |
| [35] | | [41] | | [48] |
| [35] | | [42] | | [49] |
| [35] | | [43] | | [36] |
| [35] | | [43] | | |
| [35] | | [43] | | |
注:CI1~CI28为基于距离的竞争指数,CI29~CI46是与距离无关的竞争指数;i代表目标木,j代表竞争木;n为目标木i所在结构单元内竞争木数量;distij 为竞争木j和目标木i间的距离;di 为目标木胸径,dj 为竞争木胸径,为结构单元内树木平均胸径,q为样地内树木胸径均方根;gi 、gmax、分别为目标木胸高断面积、结构单元内树木胸高断面积的最大值、平均值;hj 、hi 、分别为竞争木、目标木的树高、样地内树高平均值;CRi 为目标木冠幅,CRj 为竞争木冠幅,为结构单元内树木平均冠幅,CLi 是目标木冠长;aj 、ai 分别是竞争木和目标木的冠幅面积,Oij 、OLij 分别是目标木和竞争木的树冠重叠面积、重叠部分最宽处长度;CrownVi 是目标木树冠体积,CSAi 是目标木树冠表面积. ...
... [
29]
| [8] | | [46] | | [30] | | [31] | | [46] |
| [29] | | [36] | | [47] |
| [31] | | [36] | | [48] |
| [32] | | [37] | | [48] |
| [33] | | [38] | | [48] |
| [34] | | [39] | | [48] |
| [35] | | [40] | | [49] |
| [35] | | [41] | | [48] |
| [35] | | [42] | | [49] |
| [35] | | [43] | | [36] |
| [35] | | [43] | | |
| [35] | | [43] | | |
注:CI1~CI28为基于距离的竞争指数,CI29~CI46是与距离无关的竞争指数;i代表目标木,j代表竞争木;n为目标木i所在结构单元内竞争木数量;distij 为竞争木j和目标木i间的距离;di 为目标木胸径,dj 为竞争木胸径,为结构单元内树木平均胸径,q为样地内树木胸径均方根;gi 、gmax、分别为目标木胸高断面积、结构单元内树木胸高断面积的最大值、平均值;hj 、hi 、分别为竞争木、目标木的树高、样地内树高平均值;CRi 为目标木冠幅,CRj 为竞争木冠幅,为结构单元内树木平均冠幅,CLi 是目标木冠长;aj 、ai 分别是竞争木和目标木的冠幅面积,Oij 、OLij 分别是目标木和竞争木的树冠重叠面积、重叠部分最宽处长度;CrownVi 是目标木树冠体积,CSAi 是目标木树冠表面积. ...
... [
29]
| [36] | | [47] | | [31] | | [36] | | [48] |
| [32] | | [37] | | [48] |
| [33] | | [38] | | [48] |
| [34] | | [39] | | [48] |
| [35] | | [40] | | [49] |
| [35] | | [41] | | [48] |
| [35] | | [42] | | [49] |
| [35] | | [43] | | [36] |
| [35] | | [43] | | |
| [35] | | [43] | | |
注:CI1~CI28为基于距离的竞争指数,CI29~CI46是与距离无关的竞争指数;i代表目标木,j代表竞争木;n为目标木i所在结构单元内竞争木数量;distij 为竞争木j和目标木i间的距离;di 为目标木胸径,dj 为竞争木胸径,为结构单元内树木平均胸径,q为样地内树木胸径均方根;gi 、gmax、分别为目标木胸高断面积、结构单元内树木胸高断面积的最大值、平均值;hj 、hi 、分别为竞争木、目标木的树高、样地内树高平均值;CRi 为目标木冠幅,CRj 为竞争木冠幅,为结构单元内树木平均冠幅,CLi 是目标木冠长;aj 、ai 分别是竞争木和目标木的冠幅面积,Oij 、OLij 分别是目标木和竞争木的树冠重叠面积、重叠部分最宽处长度;CrownVi 是目标木树冠体积,CSAi 是目标木树冠表面积. ...
A new index representing individual tree competitive status
1
1973
... Individual tree competition index
Table 2| 竞争指数 | 文献 | 竞争指数 | 文献 | 竞争指数 | 文献 |
|---|
| [27] | | [29] | | [44] |
| [28] | | [29] | | [45] |
| [29] | | [29] | | [46] |
| [29] | | [8] | | [46] |
| [30] | | [31] | | [46] |
| [29] | | [36] | | [47] |
| [31] | | [36] | | [48] |
| [32] | | [37] | | [48] |
| [33] | | [38] | | [48] |
| [34] | | [39] | | [48] |
| [35] | | [40] | | [49] |
| [35] | | [41] | | [48] |
| [35] | | [42] | | [49] |
| [35] | | [43] | | [36] |
| [35] | | [43] | | |
| [35] | | [43] | | |
注:CI1~CI28为基于距离的竞争指数,CI29~CI46是与距离无关的竞争指数;i代表目标木,j代表竞争木;n为目标木i所在结构单元内竞争木数量;distij 为竞争木j和目标木i间的距离;di 为目标木胸径,dj 为竞争木胸径,为结构单元内树木平均胸径,q为样地内树木胸径均方根;gi 、gmax、分别为目标木胸高断面积、结构单元内树木胸高断面积的最大值、平均值;hj 、hi 、分别为竞争木、目标木的树高、样地内树高平均值;CRi 为目标木冠幅,CRj 为竞争木冠幅,为结构单元内树木平均冠幅,CLi 是目标木冠长;aj 、ai 分别是竞争木和目标木的冠幅面积,Oij 、OLij 分别是目标木和竞争木的树冠重叠面积、重叠部分最宽处长度;CrownVi 是目标木树冠体积,CSAi 是目标木树冠表面积. ...
Competition indices and the prediction of radial growth in Scots pine
2
1987
... Individual tree competition index
Table 2| 竞争指数 | 文献 | 竞争指数 | 文献 | 竞争指数 | 文献 |
|---|
| [27] | | [29] | | [44] |
| [28] | | [29] | | [45] |
| [29] | | [29] | | [46] |
| [29] | | [8] | | [46] |
| [30] | | [31] | | [46] |
| [29] | | [36] | | [47] |
| [31] | | [36] | | [48] |
| [32] | | [37] | | [48] |
| [33] | | [38] | | [48] |
| [34] | | [39] | | [48] |
| [35] | | [40] | | [49] |
| [35] | | [41] | | [48] |
| [35] | | [42] | | [49] |
| [35] | | [43] | | [36] |
| [35] | | [43] | | |
| [35] | | [43] | | |
注:CI1~CI28为基于距离的竞争指数,CI29~CI46是与距离无关的竞争指数;i代表目标木,j代表竞争木;n为目标木i所在结构单元内竞争木数量;distij 为竞争木j和目标木i间的距离;di 为目标木胸径,dj 为竞争木胸径,为结构单元内树木平均胸径,q为样地内树木胸径均方根;gi 、gmax、分别为目标木胸高断面积、结构单元内树木胸高断面积的最大值、平均值;hj 、hi 、分别为竞争木、目标木的树高、样地内树高平均值;CRi 为目标木冠幅,CRj 为竞争木冠幅,为结构单元内树木平均冠幅,CLi 是目标木冠长;aj 、ai 分别是竞争木和目标木的冠幅面积,Oij 、OLij 分别是目标木和竞争木的树冠重叠面积、重叠部分最宽处长度;CrownVi 是目标木树冠体积,CSAi 是目标木树冠表面积. ...
... [
31]
| [36] | | [48] | | [32] | | [37] | | [48] |
| [33] | | [38] | | [48] |
| [34] | | [39] | | [48] |
| [35] | | [40] | | [49] |
| [35] | | [41] | | [48] |
| [35] | | [42] | | [49] |
| [35] | | [43] | | [36] |
| [35] | | [43] | | |
| [35] | | [43] | | |
注:CI1~CI28为基于距离的竞争指数,CI29~CI46是与距离无关的竞争指数;i代表目标木,j代表竞争木;n为目标木i所在结构单元内竞争木数量;distij 为竞争木j和目标木i间的距离;di 为目标木胸径,dj 为竞争木胸径,为结构单元内树木平均胸径,q为样地内树木胸径均方根;gi 、gmax、分别为目标木胸高断面积、结构单元内树木胸高断面积的最大值、平均值;hj 、hi 、分别为竞争木、目标木的树高、样地内树高平均值;CRi 为目标木冠幅,CRj 为竞争木冠幅,为结构单元内树木平均冠幅,CLi 是目标木冠长;aj 、ai 分别是竞争木和目标木的冠幅面积,Oij 、OLij 分别是目标木和竞争木的树冠重叠面积、重叠部分最宽处长度;CrownVi 是目标木树冠体积,CSAi 是目标木树冠表面积. ...
基于引力模型的林木竞争分析
1
2022
... Individual tree competition index
Table 2| 竞争指数 | 文献 | 竞争指数 | 文献 | 竞争指数 | 文献 |
|---|
| [27] | | [29] | | [44] |
| [28] | | [29] | | [45] |
| [29] | | [29] | | [46] |
| [29] | | [8] | | [46] |
| [30] | | [31] | | [46] |
| [29] | | [36] | | [47] |
| [31] | | [36] | | [48] |
| [32] | | [37] | | [48] |
| [33] | | [38] | | [48] |
| [34] | | [39] | | [48] |
| [35] | | [40] | | [49] |
| [35] | | [41] | | [48] |
| [35] | | [42] | | [49] |
| [35] | | [43] | | [36] |
| [35] | | [43] | | |
| [35] | | [43] | | |
注:CI1~CI28为基于距离的竞争指数,CI29~CI46是与距离无关的竞争指数;i代表目标木,j代表竞争木;n为目标木i所在结构单元内竞争木数量;distij 为竞争木j和目标木i间的距离;di 为目标木胸径,dj 为竞争木胸径,为结构单元内树木平均胸径,q为样地内树木胸径均方根;gi 、gmax、分别为目标木胸高断面积、结构单元内树木胸高断面积的最大值、平均值;hj 、hi 、分别为竞争木、目标木的树高、样地内树高平均值;CRi 为目标木冠幅,CRj 为竞争木冠幅,为结构单元内树木平均冠幅,CLi 是目标木冠长;aj 、ai 分别是竞争木和目标木的冠幅面积,Oij 、OLij 分别是目标木和竞争木的树冠重叠面积、重叠部分最宽处长度;CrownVi 是目标木树冠体积,CSAi 是目标木树冠表面积. ...
Growth and spacing in an even-aged stand of Douglas fir
1
1951
... Individual tree competition index
Table 2| 竞争指数 | 文献 | 竞争指数 | 文献 | 竞争指数 | 文献 |
|---|
| [27] | | [29] | | [44] |
| [28] | | [29] | | [45] |
| [29] | | [29] | | [46] |
| [29] | | [8] | | [46] |
| [30] | | [31] | | [46] |
| [29] | | [36] | | [47] |
| [31] | | [36] | | [48] |
| [32] | | [37] | | [48] |
| [33] | | [38] | | [48] |
| [34] | | [39] | | [48] |
| [35] | | [40] | | [49] |
| [35] | | [41] | | [48] |
| [35] | | [42] | | [49] |
| [35] | | [43] | | [36] |
| [35] | | [43] | | |
| [35] | | [43] | | |
注:CI1~CI28为基于距离的竞争指数,CI29~CI46是与距离无关的竞争指数;i代表目标木,j代表竞争木;n为目标木i所在结构单元内竞争木数量;distij 为竞争木j和目标木i间的距离;di 为目标木胸径,dj 为竞争木胸径,为结构单元内树木平均胸径,q为样地内树木胸径均方根;gi 、gmax、分别为目标木胸高断面积、结构单元内树木胸高断面积的最大值、平均值;hj 、hi 、分别为竞争木、目标木的树高、样地内树高平均值;CRi 为目标木冠幅,CRj 为竞争木冠幅,为结构单元内树木平均冠幅,CLi 是目标木冠长;aj 、ai 分别是竞争木和目标木的冠幅面积,Oij 、OLij 分别是目标木和竞争木的树冠重叠面积、重叠部分最宽处长度;CrownVi 是目标木树冠体积,CSAi 是目标木树冠表面积. ...
Influencia de la competencia en el crecimiento en sección en Pinus radiata D.Don
1
2003
... Individual tree competition index
Table 2| 竞争指数 | 文献 | 竞争指数 | 文献 | 竞争指数 | 文献 |
|---|
| [27] | | [29] | | [44] |
| [28] | | [29] | | [45] |
| [29] | | [29] | | [46] |
| [29] | | [8] | | [46] |
| [30] | | [31] | | [46] |
| [29] | | [36] | | [47] |
| [31] | | [36] | | [48] |
| [32] | | [37] | | [48] |
| [33] | | [38] | | [48] |
| [34] | | [39] | | [48] |
| [35] | | [40] | | [49] |
| [35] | | [41] | | [48] |
| [35] | | [42] | | [49] |
| [35] | | [43] | | [36] |
| [35] | | [43] | | |
| [35] | | [43] | | |
注:CI1~CI28为基于距离的竞争指数,CI29~CI46是与距离无关的竞争指数;i代表目标木,j代表竞争木;n为目标木i所在结构单元内竞争木数量;distij 为竞争木j和目标木i间的距离;di 为目标木胸径,dj 为竞争木胸径,为结构单元内树木平均胸径,q为样地内树木胸径均方根;gi 、gmax、分别为目标木胸高断面积、结构单元内树木胸高断面积的最大值、平均值;hj 、hi 、分别为竞争木、目标木的树高、样地内树高平均值;CRi 为目标木冠幅,CRj 为竞争木冠幅,为结构单元内树木平均冠幅,CLi 是目标木冠长;aj 、ai 分别是竞争木和目标木的冠幅面积,Oij 、OLij 分别是目标木和竞争木的树冠重叠面积、重叠部分最宽处长度;CrownVi 是目标木树冠体积,CSAi 是目标木树冠表面积. ...
Analysis of intraspecific competition in two subalpine Norway spruce(Picea abies(L.) Karst.) stands in Paneveggio(Trento,Italy)
6
2008
... Individual tree competition index
Table 2| 竞争指数 | 文献 | 竞争指数 | 文献 | 竞争指数 | 文献 |
|---|
| [27] | | [29] | | [44] |
| [28] | | [29] | | [45] |
| [29] | | [29] | | [46] |
| [29] | | [8] | | [46] |
| [30] | | [31] | | [46] |
| [29] | | [36] | | [47] |
| [31] | | [36] | | [48] |
| [32] | | [37] | | [48] |
| [33] | | [38] | | [48] |
| [34] | | [39] | | [48] |
| [35] | | [40] | | [49] |
| [35] | | [41] | | [48] |
| [35] | | [42] | | [49] |
| [35] | | [43] | | [36] |
| [35] | | [43] | | |
| [35] | | [43] | | |
注:CI1~CI28为基于距离的竞争指数,CI29~CI46是与距离无关的竞争指数;i代表目标木,j代表竞争木;n为目标木i所在结构单元内竞争木数量;distij 为竞争木j和目标木i间的距离;di 为目标木胸径,dj 为竞争木胸径,为结构单元内树木平均胸径,q为样地内树木胸径均方根;gi 、gmax、分别为目标木胸高断面积、结构单元内树木胸高断面积的最大值、平均值;hj 、hi 、分别为竞争木、目标木的树高、样地内树高平均值;CRi 为目标木冠幅,CRj 为竞争木冠幅,为结构单元内树木平均冠幅,CLi 是目标木冠长;aj 、ai 分别是竞争木和目标木的冠幅面积,Oij 、OLij 分别是目标木和竞争木的树冠重叠面积、重叠部分最宽处长度;CrownVi 是目标木树冠体积,CSAi 是目标木树冠表面积. ...
... [
35]
| [41] | | [48] | | [35] | | [42] | | [49] |
| [35] | | [43] | | [36] |
| [35] | | [43] | | |
| [35] | | [43] | | |
注:CI1~CI28为基于距离的竞争指数,CI29~CI46是与距离无关的竞争指数;i代表目标木,j代表竞争木;n为目标木i所在结构单元内竞争木数量;distij 为竞争木j和目标木i间的距离;di 为目标木胸径,dj 为竞争木胸径,为结构单元内树木平均胸径,q为样地内树木胸径均方根;gi 、gmax、分别为目标木胸高断面积、结构单元内树木胸高断面积的最大值、平均值;hj 、hi 、分别为竞争木、目标木的树高、样地内树高平均值;CRi 为目标木冠幅,CRj 为竞争木冠幅,为结构单元内树木平均冠幅,CLi 是目标木冠长;aj 、ai 分别是竞争木和目标木的冠幅面积,Oij 、OLij 分别是目标木和竞争木的树冠重叠面积、重叠部分最宽处长度;CrownVi 是目标木树冠体积,CSAi 是目标木树冠表面积. ...
... [
35]
| [42] | | [49] | | [35] | | [43] | | [36] |
| [35] | | [43] | | |
| [35] | | [43] | | |
注:CI1~CI28为基于距离的竞争指数,CI29~CI46是与距离无关的竞争指数;i代表目标木,j代表竞争木;n为目标木i所在结构单元内竞争木数量;distij 为竞争木j和目标木i间的距离;di 为目标木胸径,dj 为竞争木胸径,为结构单元内树木平均胸径,q为样地内树木胸径均方根;gi 、gmax、分别为目标木胸高断面积、结构单元内树木胸高断面积的最大值、平均值;hj 、hi 、分别为竞争木、目标木的树高、样地内树高平均值;CRi 为目标木冠幅,CRj 为竞争木冠幅,为结构单元内树木平均冠幅,CLi 是目标木冠长;aj 、ai 分别是竞争木和目标木的冠幅面积,Oij 、OLij 分别是目标木和竞争木的树冠重叠面积、重叠部分最宽处长度;CrownVi 是目标木树冠体积,CSAi 是目标木树冠表面积. ...
... [
35]
| [43] | | [36] | | [35] | | [43] | | |
| [35] | | [43] | | |
注:CI1~CI28为基于距离的竞争指数,CI29~CI46是与距离无关的竞争指数;i代表目标木,j代表竞争木;n为目标木i所在结构单元内竞争木数量;distij 为竞争木j和目标木i间的距离;di 为目标木胸径,dj 为竞争木胸径,为结构单元内树木平均胸径,q为样地内树木胸径均方根;gi 、gmax、分别为目标木胸高断面积、结构单元内树木胸高断面积的最大值、平均值;hj 、hi 、分别为竞争木、目标木的树高、样地内树高平均值;CRi 为目标木冠幅,CRj 为竞争木冠幅,为结构单元内树木平均冠幅,CLi 是目标木冠长;aj 、ai 分别是竞争木和目标木的冠幅面积,Oij 、OLij 分别是目标木和竞争木的树冠重叠面积、重叠部分最宽处长度;CrownVi 是目标木树冠体积,CSAi 是目标木树冠表面积. ...
... [
35]
| [43] | | | | [35] | | [43] | | |
注:CI1~CI28为基于距离的竞争指数,CI29~CI46是与距离无关的竞争指数;i代表目标木,j代表竞争木;n为目标木i所在结构单元内竞争木数量;distij 为竞争木j和目标木i间的距离;di 为目标木胸径,dj 为竞争木胸径,为结构单元内树木平均胸径,q为样地内树木胸径均方根;gi 、gmax、分别为目标木胸高断面积、结构单元内树木胸高断面积的最大值、平均值;hj 、hi 、分别为竞争木、目标木的树高、样地内树高平均值;CRi 为目标木冠幅,CRj 为竞争木冠幅,为结构单元内树木平均冠幅,CLi 是目标木冠长;aj 、ai 分别是竞争木和目标木的冠幅面积,Oij 、OLij 分别是目标木和竞争木的树冠重叠面积、重叠部分最宽处长度;CrownVi 是目标木树冠体积,CSAi 是目标木树冠表面积. ...
... [
35]
| [43] | | | 注:CI1~CI28为基于距离的竞争指数,CI29~CI46是与距离无关的竞争指数;i代表目标木,j代表竞争木;n为目标木i所在结构单元内竞争木数量;distij 为竞争木j和目标木i间的距离;di 为目标木胸径,dj 为竞争木胸径,为结构单元内树木平均胸径,q为样地内树木胸径均方根;gi 、gmax、分别为目标木胸高断面积、结构单元内树木胸高断面积的最大值、平均值;hj 、hi 、分别为竞争木、目标木的树高、样地内树高平均值;CRi 为目标木冠幅,CRj 为竞争木冠幅,为结构单元内树木平均冠幅,CLi 是目标木冠长;aj 、ai 分别是竞争木和目标木的冠幅面积,Oij 、OLij 分别是目标木和竞争木的树冠重叠面积、重叠部分最宽处长度;CrownVi 是目标木树冠体积,CSAi 是目标木树冠表面积. ...
Competition indices and their relationship with Basal Area Increment of Araucaria
3
2018
... Individual tree competition index
Table 2| 竞争指数 | 文献 | 竞争指数 | 文献 | 竞争指数 | 文献 |
|---|
| [27] | | [29] | | [44] |
| [28] | | [29] | | [45] |
| [29] | | [29] | | [46] |
| [29] | | [8] | | [46] |
| [30] | | [31] | | [46] |
| [29] | | [36] | | [47] |
| [31] | | [36] | | [48] |
| [32] | | [37] | | [48] |
| [33] | | [38] | | [48] |
| [34] | | [39] | | [48] |
| [35] | | [40] | | [49] |
| [35] | | [41] | | [48] |
| [35] | | [42] | | [49] |
| [35] | | [43] | | [36] |
| [35] | | [43] | | |
| [35] | | [43] | | |
注:CI1~CI28为基于距离的竞争指数,CI29~CI46是与距离无关的竞争指数;i代表目标木,j代表竞争木;n为目标木i所在结构单元内竞争木数量;distij 为竞争木j和目标木i间的距离;di 为目标木胸径,dj 为竞争木胸径,为结构单元内树木平均胸径,q为样地内树木胸径均方根;gi 、gmax、分别为目标木胸高断面积、结构单元内树木胸高断面积的最大值、平均值;hj 、hi 、分别为竞争木、目标木的树高、样地内树高平均值;CRi 为目标木冠幅,CRj 为竞争木冠幅,为结构单元内树木平均冠幅,CLi 是目标木冠长;aj 、ai 分别是竞争木和目标木的冠幅面积,Oij 、OLij 分别是目标木和竞争木的树冠重叠面积、重叠部分最宽处长度;CrownVi 是目标木树冠体积,CSAi 是目标木树冠表面积. ...
... [
36]
| [48] | | [32] | | [37] | | [48] |
| [33] | | [38] | | [48] |
| [34] | | [39] | | [48] |
| [35] | | [40] | | [49] |
| [35] | | [41] | | [48] |
| [35] | | [42] | | [49] |
| [35] | | [43] | | [36] |
| [35] | | [43] | | |
| [35] | | [43] | | |
注:CI1~CI28为基于距离的竞争指数,CI29~CI46是与距离无关的竞争指数;i代表目标木,j代表竞争木;n为目标木i所在结构单元内竞争木数量;distij 为竞争木j和目标木i间的距离;di 为目标木胸径,dj 为竞争木胸径,为结构单元内树木平均胸径,q为样地内树木胸径均方根;gi 、gmax、分别为目标木胸高断面积、结构单元内树木胸高断面积的最大值、平均值;hj 、hi 、分别为竞争木、目标木的树高、样地内树高平均值;CRi 为目标木冠幅,CRj 为竞争木冠幅,为结构单元内树木平均冠幅,CLi 是目标木冠长;aj 、ai 分别是竞争木和目标木的冠幅面积,Oij 、OLij 分别是目标木和竞争木的树冠重叠面积、重叠部分最宽处长度;CrownVi 是目标木树冠体积,CSAi 是目标木树冠表面积. ...
... [
36]
| [35] | | [43] | | |
| [35] | | [43] | | |
注:CI1~CI28为基于距离的竞争指数,CI29~CI46是与距离无关的竞争指数;i代表目标木,j代表竞争木;n为目标木i所在结构单元内竞争木数量;distij 为竞争木j和目标木i间的距离;di 为目标木胸径,dj 为竞争木胸径,为结构单元内树木平均胸径,q为样地内树木胸径均方根;gi 、gmax、分别为目标木胸高断面积、结构单元内树木胸高断面积的最大值、平均值;hj 、hi 、分别为竞争木、目标木的树高、样地内树高平均值;CRi 为目标木冠幅,CRj 为竞争木冠幅,为结构单元内树木平均冠幅,CLi 是目标木冠长;aj 、ai 分别是竞争木和目标木的冠幅面积,Oij 、OLij 分别是目标木和竞争木的树冠重叠面积、重叠部分最宽处长度;CrownVi 是目标木树冠体积,CSAi 是目标木树冠表面积. ...
Competition quotient:a new measure for the competition affecting individual forest trees
1
1969
... Individual tree competition index
Table 2| 竞争指数 | 文献 | 竞争指数 | 文献 | 竞争指数 | 文献 |
|---|
| [27] | | [29] | | [44] |
| [28] | | [29] | | [45] |
| [29] | | [29] | | [46] |
| [29] | | [8] | | [46] |
| [30] | | [31] | | [46] |
| [29] | | [36] | | [47] |
| [31] | | [36] | | [48] |
| [32] | | [37] | | [48] |
| [33] | | [38] | | [48] |
| [34] | | [39] | | [48] |
| [35] | | [40] | | [49] |
| [35] | | [41] | | [48] |
| [35] | | [42] | | [49] |
| [35] | | [43] | | [36] |
| [35] | | [43] | | |
| [35] | | [43] | | |
注:CI1~CI28为基于距离的竞争指数,CI29~CI46是与距离无关的竞争指数;i代表目标木,j代表竞争木;n为目标木i所在结构单元内竞争木数量;distij 为竞争木j和目标木i间的距离;di 为目标木胸径,dj 为竞争木胸径,为结构单元内树木平均胸径,q为样地内树木胸径均方根;gi 、gmax、分别为目标木胸高断面积、结构单元内树木胸高断面积的最大值、平均值;hj 、hi 、分别为竞争木、目标木的树高、样地内树高平均值;CRi 为目标木冠幅,CRj 为竞争木冠幅,为结构单元内树木平均冠幅,CLi 是目标木冠长;aj 、ai 分别是竞争木和目标木的冠幅面积,Oij 、OLij 分别是目标木和竞争木的树冠重叠面积、重叠部分最宽处长度;CrownVi 是目标木树冠体积,CSAi 是目标木树冠表面积. ...
A new competition model for individual trees
1
1971
... Individual tree competition index
Table 2| 竞争指数 | 文献 | 竞争指数 | 文献 | 竞争指数 | 文献 |
|---|
| [27] | | [29] | | [44] |
| [28] | | [29] | | [45] |
| [29] | | [29] | | [46] |
| [29] | | [8] | | [46] |
| [30] | | [31] | | [46] |
| [29] | | [36] | | [47] |
| [31] | | [36] | | [48] |
| [32] | | [37] | | [48] |
| [33] | | [38] | | [48] |
| [34] | | [39] | | [48] |
| [35] | | [40] | | [49] |
| [35] | | [41] | | [48] |
| [35] | | [42] | | [49] |
| [35] | | [43] | | [36] |
| [35] | | [43] | | |
| [35] | | [43] | | |
注:CI1~CI28为基于距离的竞争指数,CI29~CI46是与距离无关的竞争指数;i代表目标木,j代表竞争木;n为目标木i所在结构单元内竞争木数量;distij 为竞争木j和目标木i间的距离;di 为目标木胸径,dj 为竞争木胸径,为结构单元内树木平均胸径,q为样地内树木胸径均方根;gi 、gmax、分别为目标木胸高断面积、结构单元内树木胸高断面积的最大值、平均值;hj 、hi 、分别为竞争木、目标木的树高、样地内树高平均值;CRi 为目标木冠幅,CRj 为竞争木冠幅,为结构单元内树木平均冠幅,CLi 是目标木冠长;aj 、ai 分别是竞争木和目标木的冠幅面积,Oij 、OLij 分别是目标木和竞争木的树冠重叠面积、重叠部分最宽处长度;CrownVi 是目标木树冠体积,CSAi 是目标木树冠表面积. ...
Growth model for simulation of a Calabrian pine(Pinus brutia Ten.) tree
1
1977
... Individual tree competition index
Table 2| 竞争指数 | 文献 | 竞争指数 | 文献 | 竞争指数 | 文献 |
|---|
| [27] | | [29] | | [44] |
| [28] | | [29] | | [45] |
| [29] | | [29] | | [46] |
| [29] | | [8] | | [46] |
| [30] | | [31] | | [46] |
| [29] | | [36] | | [47] |
| [31] | | [36] | | [48] |
| [32] | | [37] | | [48] |
| [33] | | [38] | | [48] |
| [34] | | [39] | | [48] |
| [35] | | [40] | | [49] |
| [35] | | [41] | | [48] |
| [35] | | [42] | | [49] |
| [35] | | [43] | | [36] |
| [35] | | [43] | | |
| [35] | | [43] | | |
注:CI1~CI28为基于距离的竞争指数,CI29~CI46是与距离无关的竞争指数;i代表目标木,j代表竞争木;n为目标木i所在结构单元内竞争木数量;distij 为竞争木j和目标木i间的距离;di 为目标木胸径,dj 为竞争木胸径,为结构单元内树木平均胸径,q为样地内树木胸径均方根;gi 、gmax、分别为目标木胸高断面积、结构单元内树木胸高断面积的最大值、平均值;hj 、hi 、分别为竞争木、目标木的树高、样地内树高平均值;CRi 为目标木冠幅,CRj 为竞争木冠幅,为结构单元内树木平均冠幅,CLi 是目标木冠长;aj 、ai 分别是竞争木和目标木的冠幅面积,Oij 、OLij 分别是目标木和竞争木的树冠重叠面积、重叠部分最宽处长度;CrownVi 是目标木树冠体积,CSAi 是目标木树冠表面积. ...
Trials with program FOREST:growth and reproduction simulation for mixed species even and or uneven-aged forest stands
1
1974
... Individual tree competition index
Table 2| 竞争指数 | 文献 | 竞争指数 | 文献 | 竞争指数 | 文献 |
|---|
| [27] | | [29] | | [44] |
| [28] | | [29] | | [45] |
| [29] | | [29] | | [46] |
| [29] | | [8] | | [46] |
| [30] | | [31] | | [46] |
| [29] | | [36] | | [47] |
| [31] | | [36] | | [48] |
| [32] | | [37] | | [48] |
| [33] | | [38] | | [48] |
| [34] | | [39] | | [48] |
| [35] | | [40] | | [49] |
| [35] | | [41] | | [48] |
| [35] | | [42] | | [49] |
| [35] | | [43] | | [36] |
| [35] | | [43] | | |
| [35] | | [43] | | |
注:CI1~CI28为基于距离的竞争指数,CI29~CI46是与距离无关的竞争指数;i代表目标木,j代表竞争木;n为目标木i所在结构单元内竞争木数量;distij 为竞争木j和目标木i间的距离;di 为目标木胸径,dj 为竞争木胸径,为结构单元内树木平均胸径,q为样地内树木胸径均方根;gi 、gmax、分别为目标木胸高断面积、结构单元内树木胸高断面积的最大值、平均值;hj 、hi 、分别为竞争木、目标木的树高、样地内树高平均值;CRi 为目标木冠幅,CRj 为竞争木冠幅,为结构单元内树木平均冠幅,CLi 是目标木冠长;aj 、ai 分别是竞争木和目标木的冠幅面积,Oij 、OLij 分别是目标木和竞争木的树冠重叠面积、重叠部分最宽处长度;CrownVi 是目标木树冠体积,CSAi 是目标木树冠表面积. ...
Evaluation of some competition indexes for the prediction of diameter increment in planted white spruce
1
1978
... Individual tree competition index
Table 2| 竞争指数 | 文献 | 竞争指数 | 文献 | 竞争指数 | 文献 |
|---|
| [27] | | [29] | | [44] |
| [28] | | [29] | | [45] |
| [29] | | [29] | | [46] |
| [29] | | [8] | | [46] |
| [30] | | [31] | | [46] |
| [29] | | [36] | | [47] |
| [31] | | [36] | | [48] |
| [32] | | [37] | | [48] |
| [33] | | [38] | | [48] |
| [34] | | [39] | | [48] |
| [35] | | [40] | | [49] |
| [35] | | [41] | | [48] |
| [35] | | [42] | | [49] |
| [35] | | [43] | | [36] |
| [35] | | [43] | | |
| [35] | | [43] | | |
注:CI1~CI28为基于距离的竞争指数,CI29~CI46是与距离无关的竞争指数;i代表目标木,j代表竞争木;n为目标木i所在结构单元内竞争木数量;distij 为竞争木j和目标木i间的距离;di 为目标木胸径,dj 为竞争木胸径,为结构单元内树木平均胸径,q为样地内树木胸径均方根;gi 、gmax、分别为目标木胸高断面积、结构单元内树木胸高断面积的最大值、平均值;hj 、hi 、分别为竞争木、目标木的树高、样地内树高平均值;CRi 为目标木冠幅,CRj 为竞争木冠幅,为结构单元内树木平均冠幅,CLi 是目标木冠长;aj 、ai 分别是竞争木和目标木的冠幅面积,Oij 、OLij 分别是目标木和竞争木的树冠重叠面积、重叠部分最宽处长度;CrownVi 是目标木树冠体积,CSAi 是目标木树冠表面积. ...
A basal area ratio predictor of loblolly pine plantation mortality
1
1979
... Individual tree competition index
Table 2| 竞争指数 | 文献 | 竞争指数 | 文献 | 竞争指数 | 文献 |
|---|
| [27] | | [29] | | [44] |
| [28] | | [29] | | [45] |
| [29] | | [29] | | [46] |
| [29] | | [8] | | [46] |
| [30] | | [31] | | [46] |
| [29] | | [36] | | [47] |
| [31] | | [36] | | [48] |
| [32] | | [37] | | [48] |
| [33] | | [38] | | [48] |
| [34] | | [39] | | [48] |
| [35] | | [40] | | [49] |
| [35] | | [41] | | [48] |
| [35] | | [42] | | [49] |
| [35] | | [43] | | [36] |
| [35] | | [43] | | |
| [35] | | [43] | | |
注:CI1~CI28为基于距离的竞争指数,CI29~CI46是与距离无关的竞争指数;i代表目标木,j代表竞争木;n为目标木i所在结构单元内竞争木数量;distij 为竞争木j和目标木i间的距离;di 为目标木胸径,dj 为竞争木胸径,为结构单元内树木平均胸径,q为样地内树木胸径均方根;gi 、gmax、分别为目标木胸高断面积、结构单元内树木胸高断面积的最大值、平均值;hj 、hi 、分别为竞争木、目标木的树高、样地内树高平均值;CRi 为目标木冠幅,CRj 为竞争木冠幅,为结构单元内树木平均冠幅,CLi 是目标木冠长;aj 、ai 分别是竞争木和目标木的冠幅面积,Oij 、OLij 分别是目标木和竞争木的树冠重叠面积、重叠部分最宽处长度;CrownVi 是目标木树冠体积,CSAi 是目标木树冠表面积. ...
A comparison of competition measures for predicting growth of loblolly pine trees
3
1986
... Individual tree competition index
Table 2| 竞争指数 | 文献 | 竞争指数 | 文献 | 竞争指数 | 文献 |
|---|
| [27] | | [29] | | [44] |
| [28] | | [29] | | [45] |
| [29] | | [29] | | [46] |
| [29] | | [8] | | [46] |
| [30] | | [31] | | [46] |
| [29] | | [36] | | [47] |
| [31] | | [36] | | [48] |
| [32] | | [37] | | [48] |
| [33] | | [38] | | [48] |
| [34] | | [39] | | [48] |
| [35] | | [40] | | [49] |
| [35] | | [41] | | [48] |
| [35] | | [42] | | [49] |
| [35] | | [43] | | [36] |
| [35] | | [43] | | |
| [35] | | [43] | | |
注:CI1~CI28为基于距离的竞争指数,CI29~CI46是与距离无关的竞争指数;i代表目标木,j代表竞争木;n为目标木i所在结构单元内竞争木数量;distij 为竞争木j和目标木i间的距离;di 为目标木胸径,dj 为竞争木胸径,为结构单元内树木平均胸径,q为样地内树木胸径均方根;gi 、gmax、分别为目标木胸高断面积、结构单元内树木胸高断面积的最大值、平均值;hj 、hi 、分别为竞争木、目标木的树高、样地内树高平均值;CRi 为目标木冠幅,CRj 为竞争木冠幅,为结构单元内树木平均冠幅,CLi 是目标木冠长;aj 、ai 分别是竞争木和目标木的冠幅面积,Oij 、OLij 分别是目标木和竞争木的树冠重叠面积、重叠部分最宽处长度;CrownVi 是目标木树冠体积,CSAi 是目标木树冠表面积. ...
... [
43]
| | | [35] | | [43] | | |
注:CI1~CI28为基于距离的竞争指数,CI29~CI46是与距离无关的竞争指数;i代表目标木,j代表竞争木;n为目标木i所在结构单元内竞争木数量;distij 为竞争木j和目标木i间的距离;di 为目标木胸径,dj 为竞争木胸径,为结构单元内树木平均胸径,q为样地内树木胸径均方根;gi 、gmax、分别为目标木胸高断面积、结构单元内树木胸高断面积的最大值、平均值;hj 、hi 、分别为竞争木、目标木的树高、样地内树高平均值;CRi 为目标木冠幅,CRj 为竞争木冠幅,为结构单元内树木平均冠幅,CLi 是目标木冠长;aj 、ai 分别是竞争木和目标木的冠幅面积,Oij 、OLij 分别是目标木和竞争木的树冠重叠面积、重叠部分最宽处长度;CrownVi 是目标木树冠体积,CSAi 是目标木树冠表面积. ...
... [
43]
| | 注:CI1~CI28为基于距离的竞争指数,CI29~CI46是与距离无关的竞争指数;i代表目标木,j代表竞争木;n为目标木i所在结构单元内竞争木数量;distij 为竞争木j和目标木i间的距离;di 为目标木胸径,dj 为竞争木胸径,为结构单元内树木平均胸径,q为样地内树木胸径均方根;gi 、gmax、分别为目标木胸高断面积、结构单元内树木胸高断面积的最大值、平均值;hj 、hi 、分别为竞争木、目标木的树高、样地内树高平均值;CRi 为目标木冠幅,CRj 为竞争木冠幅,为结构单元内树木平均冠幅,CLi 是目标木冠长;aj 、ai 分别是竞争木和目标木的冠幅面积,Oij 、OLij 分别是目标木和竞争木的树冠重叠面积、重叠部分最宽处长度;CrownVi 是目标木树冠体积,CSAi 是目标木树冠表面积. ...
Prognosis model for stand development
1
1973
... Individual tree competition index
Table 2| 竞争指数 | 文献 | 竞争指数 | 文献 | 竞争指数 | 文献 |
|---|
| [27] | | [29] | | [44] |
| [28] | | [29] | | [45] |
| [29] | | [29] | | [46] |
| [29] | | [8] | | [46] |
| [30] | | [31] | | [46] |
| [29] | | [36] | | [47] |
| [31] | | [36] | | [48] |
| [32] | | [37] | | [48] |
| [33] | | [38] | | [48] |
| [34] | | [39] | | [48] |
| [35] | | [40] | | [49] |
| [35] | | [41] | | [48] |
| [35] | | [42] | | [49] |
| [35] | | [43] | | [36] |
| [35] | | [43] | | |
| [35] | | [43] | | |
注:CI1~CI28为基于距离的竞争指数,CI29~CI46是与距离无关的竞争指数;i代表目标木,j代表竞争木;n为目标木i所在结构单元内竞争木数量;distij 为竞争木j和目标木i间的距离;di 为目标木胸径,dj 为竞争木胸径,为结构单元内树木平均胸径,q为样地内树木胸径均方根;gi 、gmax、分别为目标木胸高断面积、结构单元内树木胸高断面积的最大值、平均值;hj 、hi 、分别为竞争木、目标木的树高、样地内树高平均值;CRi 为目标木冠幅,CRj 为竞争木冠幅,为结构单元内树木平均冠幅,CLi 是目标木冠长;aj 、ai 分别是竞争木和目标木的冠幅面积,Oij 、OLij 分别是目标木和竞争木的树冠重叠面积、重叠部分最宽处长度;CrownVi 是目标木树冠体积,CSAi 是目标木树冠表面积. ...
User's guide to the stand prognosis model
1
1982
... Individual tree competition index
Table 2| 竞争指数 | 文献 | 竞争指数 | 文献 | 竞争指数 | 文献 |
|---|
| [27] | | [29] | | [44] |
| [28] | | [29] | | [45] |
| [29] | | [29] | | [46] |
| [29] | | [8] | | [46] |
| [30] | | [31] | | [46] |
| [29] | | [36] | | [47] |
| [31] | | [36] | | [48] |
| [32] | | [37] | | [48] |
| [33] | | [38] | | [48] |
| [34] | | [39] | | [48] |
| [35] | | [40] | | [49] |
| [35] | | [41] | | [48] |
| [35] | | [42] | | [49] |
| [35] | | [43] | | [36] |
| [35] | | [43] | | |
| [35] | | [43] | | |
注:CI1~CI28为基于距离的竞争指数,CI29~CI46是与距离无关的竞争指数;i代表目标木,j代表竞争木;n为目标木i所在结构单元内竞争木数量;distij 为竞争木j和目标木i间的距离;di 为目标木胸径,dj 为竞争木胸径,为结构单元内树木平均胸径,q为样地内树木胸径均方根;gi 、gmax、分别为目标木胸高断面积、结构单元内树木胸高断面积的最大值、平均值;hj 、hi 、分别为竞争木、目标木的树高、样地内树高平均值;CRi 为目标木冠幅,CRj 为竞争木冠幅,为结构单元内树木平均冠幅,CLi 是目标木冠长;aj 、ai 分别是竞争木和目标木的冠幅面积,Oij 、OLij 分别是目标木和竞争木的树冠重叠面积、重叠部分最宽处长度;CrownVi 是目标木树冠体积,CSAi 是目标木树冠表面积. ...
Distance-dependent competition measures for predicting growth of individual trees
3
1989
... Individual tree competition index
Table 2| 竞争指数 | 文献 | 竞争指数 | 文献 | 竞争指数 | 文献 |
|---|
| [27] | | [29] | | [44] |
| [28] | | [29] | | [45] |
| [29] | | [29] | | [46] |
| [29] | | [8] | | [46] |
| [30] | | [31] | | [46] |
| [29] | | [36] | | [47] |
| [31] | | [36] | | [48] |
| [32] | | [37] | | [48] |
| [33] | | [38] | | [48] |
| [34] | | [39] | | [48] |
| [35] | | [40] | | [49] |
| [35] | | [41] | | [48] |
| [35] | | [42] | | [49] |
| [35] | | [43] | | [36] |
| [35] | | [43] | | |
| [35] | | [43] | | |
注:CI1~CI28为基于距离的竞争指数,CI29~CI46是与距离无关的竞争指数;i代表目标木,j代表竞争木;n为目标木i所在结构单元内竞争木数量;distij 为竞争木j和目标木i间的距离;di 为目标木胸径,dj 为竞争木胸径,为结构单元内树木平均胸径,q为样地内树木胸径均方根;gi 、gmax、分别为目标木胸高断面积、结构单元内树木胸高断面积的最大值、平均值;hj 、hi 、分别为竞争木、目标木的树高、样地内树高平均值;CRi 为目标木冠幅,CRj 为竞争木冠幅,为结构单元内树木平均冠幅,CLi 是目标木冠长;aj 、ai 分别是竞争木和目标木的冠幅面积,Oij 、OLij 分别是目标木和竞争木的树冠重叠面积、重叠部分最宽处长度;CrownVi 是目标木树冠体积,CSAi 是目标木树冠表面积. ...
... [
46]
| [30] | | [31] | | [46] |
| [29] | | [36] | | [47] |
| [31] | | [36] | | [48] |
| [32] | | [37] | | [48] |
| [33] | | [38] | | [48] |
| [34] | | [39] | | [48] |
| [35] | | [40] | | [49] |
| [35] | | [41] | | [48] |
| [35] | | [42] | | [49] |
| [35] | | [43] | | [36] |
| [35] | | [43] | | |
| [35] | | [43] | | |
注:CI1~CI28为基于距离的竞争指数,CI29~CI46是与距离无关的竞争指数;i代表目标木,j代表竞争木;n为目标木i所在结构单元内竞争木数量;distij 为竞争木j和目标木i间的距离;di 为目标木胸径,dj 为竞争木胸径,为结构单元内树木平均胸径,q为样地内树木胸径均方根;gi 、gmax、分别为目标木胸高断面积、结构单元内树木胸高断面积的最大值、平均值;hj 、hi 、分别为竞争木、目标木的树高、样地内树高平均值;CRi 为目标木冠幅,CRj 为竞争木冠幅,为结构单元内树木平均冠幅,CLi 是目标木冠长;aj 、ai 分别是竞争木和目标木的冠幅面积,Oij 、OLij 分别是目标木和竞争木的树冠重叠面积、重叠部分最宽处长度;CrownVi 是目标木树冠体积,CSAi 是目标木树冠表面积. ...
... [
46]
| [29] | | [36] | | [47] |
| [31] | | [36] | | [48] |
| [32] | | [37] | | [48] |
| [33] | | [38] | | [48] |
| [34] | | [39] | | [48] |
| [35] | | [40] | | [49] |
| [35] | | [41] | | [48] |
| [35] | | [42] | | [49] |
| [35] | | [43] | | [36] |
| [35] | | [43] | | |
| [35] | | [43] | | |
注:CI1~CI28为基于距离的竞争指数,CI29~CI46是与距离无关的竞争指数;i代表目标木,j代表竞争木;n为目标木i所在结构单元内竞争木数量;distij 为竞争木j和目标木i间的距离;di 为目标木胸径,dj 为竞争木胸径,为结构单元内树木平均胸径,q为样地内树木胸径均方根;gi 、gmax、分别为目标木胸高断面积、结构单元内树木胸高断面积的最大值、平均值;hj 、hi 、分别为竞争木、目标木的树高、样地内树高平均值;CRi 为目标木冠幅,CRj 为竞争木冠幅,为结构单元内树木平均冠幅,CLi 是目标木冠长;aj 、ai 分别是竞争木和目标木的冠幅面积,Oij 、OLij 分别是目标木和竞争木的树冠重叠面积、重叠部分最宽处长度;CrownVi 是目标木树冠体积,CSAi 是目标木树冠表面积. ...
Individual competition indices for conifer plantations
1
1989
... Individual tree competition index
Table 2| 竞争指数 | 文献 | 竞争指数 | 文献 | 竞争指数 | 文献 |
|---|
| [27] | | [29] | | [44] |
| [28] | | [29] | | [45] |
| [29] | | [29] | | [46] |
| [29] | | [8] | | [46] |
| [30] | | [31] | | [46] |
| [29] | | [36] | | [47] |
| [31] | | [36] | | [48] |
| [32] | | [37] | | [48] |
| [33] | | [38] | | [48] |
| [34] | | [39] | | [48] |
| [35] | | [40] | | [49] |
| [35] | | [41] | | [48] |
| [35] | | [42] | | [49] |
| [35] | | [43] | | [36] |
| [35] | | [43] | | |
| [35] | | [43] | | |
注:CI1~CI28为基于距离的竞争指数,CI29~CI46是与距离无关的竞争指数;i代表目标木,j代表竞争木;n为目标木i所在结构单元内竞争木数量;distij 为竞争木j和目标木i间的距离;di 为目标木胸径,dj 为竞争木胸径,为结构单元内树木平均胸径,q为样地内树木胸径均方根;gi 、gmax、分别为目标木胸高断面积、结构单元内树木胸高断面积的最大值、平均值;hj 、hi 、分别为竞争木、目标木的树高、样地内树高平均值;CRi 为目标木冠幅,CRj 为竞争木冠幅,为结构单元内树木平均冠幅,CLi 是目标木冠长;aj 、ai 分别是竞争木和目标木的冠幅面积,Oij 、OLij 分别是目标木和竞争木的树冠重叠面积、重叠部分最宽处长度;CrownVi 是目标木树冠体积,CSAi 是目标木树冠表面积. ...
林分空间结构指标研究进展
5
2016
... Individual tree competition index
Table 2| 竞争指数 | 文献 | 竞争指数 | 文献 | 竞争指数 | 文献 |
|---|
| [27] | | [29] | | [44] |
| [28] | | [29] | | [45] |
| [29] | | [29] | | [46] |
| [29] | | [8] | | [46] |
| [30] | | [31] | | [46] |
| [29] | | [36] | | [47] |
| [31] | | [36] | | [48] |
| [32] | | [37] | | [48] |
| [33] | | [38] | | [48] |
| [34] | | [39] | | [48] |
| [35] | | [40] | | [49] |
| [35] | | [41] | | [48] |
| [35] | | [42] | | [49] |
| [35] | | [43] | | [36] |
| [35] | | [43] | | |
| [35] | | [43] | | |
注:CI1~CI28为基于距离的竞争指数,CI29~CI46是与距离无关的竞争指数;i代表目标木,j代表竞争木;n为目标木i所在结构单元内竞争木数量;distij 为竞争木j和目标木i间的距离;di 为目标木胸径,dj 为竞争木胸径,为结构单元内树木平均胸径,q为样地内树木胸径均方根;gi 、gmax、分别为目标木胸高断面积、结构单元内树木胸高断面积的最大值、平均值;hj 、hi 、分别为竞争木、目标木的树高、样地内树高平均值;CRi 为目标木冠幅,CRj 为竞争木冠幅,为结构单元内树木平均冠幅,CLi 是目标木冠长;aj 、ai 分别是竞争木和目标木的冠幅面积,Oij 、OLij 分别是目标木和竞争木的树冠重叠面积、重叠部分最宽处长度;CrownVi 是目标木树冠体积,CSAi 是目标木树冠表面积. ...
... [
48]
| [33] | | [38] | | [48] |
| [34] | | [39] | | [48] |
| [35] | | [40] | | [49] |
| [35] | | [41] | | [48] |
| [35] | | [42] | | [49] |
| [35] | | [43] | | [36] |
| [35] | | [43] | | |
| [35] | | [43] | | |
注:CI1~CI28为基于距离的竞争指数,CI29~CI46是与距离无关的竞争指数;i代表目标木,j代表竞争木;n为目标木i所在结构单元内竞争木数量;distij 为竞争木j和目标木i间的距离;di 为目标木胸径,dj 为竞争木胸径,为结构单元内树木平均胸径,q为样地内树木胸径均方根;gi 、gmax、分别为目标木胸高断面积、结构单元内树木胸高断面积的最大值、平均值;hj 、hi 、分别为竞争木、目标木的树高、样地内树高平均值;CRi 为目标木冠幅,CRj 为竞争木冠幅,为结构单元内树木平均冠幅,CLi 是目标木冠长;aj 、ai 分别是竞争木和目标木的冠幅面积,Oij 、OLij 分别是目标木和竞争木的树冠重叠面积、重叠部分最宽处长度;CrownVi 是目标木树冠体积,CSAi 是目标木树冠表面积. ...
... [
48]
| [34] | | [39] | | [48] |
| [35] | | [40] | | [49] |
| [35] | | [41] | | [48] |
| [35] | | [42] | | [49] |
| [35] | | [43] | | [36] |
| [35] | | [43] | | |
| [35] | | [43] | | |
注:CI1~CI28为基于距离的竞争指数,CI29~CI46是与距离无关的竞争指数;i代表目标木,j代表竞争木;n为目标木i所在结构单元内竞争木数量;distij 为竞争木j和目标木i间的距离;di 为目标木胸径,dj 为竞争木胸径,为结构单元内树木平均胸径,q为样地内树木胸径均方根;gi 、gmax、分别为目标木胸高断面积、结构单元内树木胸高断面积的最大值、平均值;hj 、hi 、分别为竞争木、目标木的树高、样地内树高平均值;CRi 为目标木冠幅,CRj 为竞争木冠幅,为结构单元内树木平均冠幅,CLi 是目标木冠长;aj 、ai 分别是竞争木和目标木的冠幅面积,Oij 、OLij 分别是目标木和竞争木的树冠重叠面积、重叠部分最宽处长度;CrownVi 是目标木树冠体积,CSAi 是目标木树冠表面积. ...
... [
48]
| [35] | | [40] | | [49] |
| [35] | | [41] | | [48] |
| [35] | | [42] | | [49] |
| [35] | | [43] | | [36] |
| [35] | | [43] | | |
| [35] | | [43] | | |
注:CI1~CI28为基于距离的竞争指数,CI29~CI46是与距离无关的竞争指数;i代表目标木,j代表竞争木;n为目标木i所在结构单元内竞争木数量;distij 为竞争木j和目标木i间的距离;di 为目标木胸径,dj 为竞争木胸径,为结构单元内树木平均胸径,q为样地内树木胸径均方根;gi 、gmax、分别为目标木胸高断面积、结构单元内树木胸高断面积的最大值、平均值;hj 、hi 、分别为竞争木、目标木的树高、样地内树高平均值;CRi 为目标木冠幅,CRj 为竞争木冠幅,为结构单元内树木平均冠幅,CLi 是目标木冠长;aj 、ai 分别是竞争木和目标木的冠幅面积,Oij 、OLij 分别是目标木和竞争木的树冠重叠面积、重叠部分最宽处长度;CrownVi 是目标木树冠体积,CSAi 是目标木树冠表面积. ...
... [
48]
| [35] | | [42] | | [49] |
| [35] | | [43] | | [36] |
| [35] | | [43] | | |
| [35] | | [43] | | |
注:CI1~CI28为基于距离的竞争指数,CI29~CI46是与距离无关的竞争指数;i代表目标木,j代表竞争木;n为目标木i所在结构单元内竞争木数量;distij 为竞争木j和目标木i间的距离;di 为目标木胸径,dj 为竞争木胸径,为结构单元内树木平均胸径,q为样地内树木胸径均方根;gi 、gmax、分别为目标木胸高断面积、结构单元内树木胸高断面积的最大值、平均值;hj 、hi 、分别为竞争木、目标木的树高、样地内树高平均值;CRi 为目标木冠幅,CRj 为竞争木冠幅,为结构单元内树木平均冠幅,CLi 是目标木冠长;aj 、ai 分别是竞争木和目标木的冠幅面积,Oij 、OLij 分别是目标木和竞争木的树冠重叠面积、重叠部分最宽处长度;CrownVi 是目标木树冠体积,CSAi 是目标木树冠表面积. ...
生长空间竞争指数及其在油松、侧柏种内竞争中的应用研究
3
2006
... 通常用一些数学公式量化个体间的竞争强度,称之为竞争指数,代表每棵树木受周围个体影响的大小.为比较不同竞争指数的相对大小,本研究将部分竞争指数取倒数调整数值大小,竞争指数越小表示目标木遭受周围竞争木的压力越小,反之越大.研究通过文献收集了不同学者提出的竞争指数,计算过程如下:首先确定对象木和竞争木;然后根据不同的竞争指数公式计算竞争指数[25].对象木所承受的竞争压力主要源于周围一定距离范围内的竞争木,超出此范围的竞争压力逐渐减弱或消失[26].根据前期研究,10 m是确定沙地樟子松林木竞争最适宜的范围[18].同时,为避免边缘效应,本研究选择样地中心30 m×30 m区域内胸径≥5 cm的170株树木为对象木,每个对象木半径10 m范围内树木为竞争木.本文选取了46种单木竞争指数[8,27-49],其中包括28种基于距离的竞争指数和18种与距离无关的竞争指数,具体如表2所列. ...
... Individual tree competition index
Table 2| 竞争指数 | 文献 | 竞争指数 | 文献 | 竞争指数 | 文献 |
|---|
| [27] | | [29] | | [44] |
| [28] | | [29] | | [45] |
| [29] | | [29] | | [46] |
| [29] | | [8] | | [46] |
| [30] | | [31] | | [46] |
| [29] | | [36] | | [47] |
| [31] | | [36] | | [48] |
| [32] | | [37] | | [48] |
| [33] | | [38] | | [48] |
| [34] | | [39] | | [48] |
| [35] | | [40] | | [49] |
| [35] | | [41] | | [48] |
| [35] | | [42] | | [49] |
| [35] | | [43] | | [36] |
| [35] | | [43] | | |
| [35] | | [43] | | |
注:CI1~CI28为基于距离的竞争指数,CI29~CI46是与距离无关的竞争指数;i代表目标木,j代表竞争木;n为目标木i所在结构单元内竞争木数量;distij 为竞争木j和目标木i间的距离;di 为目标木胸径,dj 为竞争木胸径,为结构单元内树木平均胸径,q为样地内树木胸径均方根;gi 、gmax、分别为目标木胸高断面积、结构单元内树木胸高断面积的最大值、平均值;hj 、hi 、分别为竞争木、目标木的树高、样地内树高平均值;CRi 为目标木冠幅,CRj 为竞争木冠幅,为结构单元内树木平均冠幅,CLi 是目标木冠长;aj 、ai 分别是竞争木和目标木的冠幅面积,Oij 、OLij 分别是目标木和竞争木的树冠重叠面积、重叠部分最宽处长度;CrownVi 是目标木树冠体积,CSAi 是目标木树冠表面积. ...
... [
49]
| [35] | | [43] | | [36] |
| [35] | | [43] | | |
| [35] | | [43] | | |
注:CI1~CI28为基于距离的竞争指数,CI29~CI46是与距离无关的竞争指数;i代表目标木,j代表竞争木;n为目标木i所在结构单元内竞争木数量;distij 为竞争木j和目标木i间的距离;di 为目标木胸径,dj 为竞争木胸径,为结构单元内树木平均胸径,q为样地内树木胸径均方根;gi 、gmax、分别为目标木胸高断面积、结构单元内树木胸高断面积的最大值、平均值;hj 、hi 、分别为竞争木、目标木的树高、样地内树高平均值;CRi 为目标木冠幅,CRj 为竞争木冠幅,为结构单元内树木平均冠幅,CLi 是目标木冠长;aj 、ai 分别是竞争木和目标木的冠幅面积,Oij 、OLij 分别是目标木和竞争木的树冠重叠面积、重叠部分最宽处长度;CrownVi 是目标木树冠体积,CSAi 是目标木树冠表面积. ...
湖北木林子自然保护区天然林不同恢复阶段林分结构特征
1
2023
... 作为描述林分生长变异性的重要指标,林分结构在很大程度上影响着森林生长的时空动态,而森林经营很大程度上通过调控林分的结构实现[50-51].林分结构多样性的表征方式多样,如树种多样性、树木大小多样性、林木分布格局多样性等,空间和非空间分析法是量化林分结构多样性指标常用的两种方法[52].研究对象为沙地樟子松纯林,选取香农-威纳指数[53]、辛普森指数[54]、大小比数[55]、大小分异指数[56]、基尼系数[57]、变异系数[57]、偏度[10]、峰度[58]、Clark-Evans指数[59]等9种林分结构多样性指数来描述对象木对应结构单元的林分结构状况(表3).在计算林分结构多样性指数时,树木胸径按3 cm的径阶距划分. ...
六盘山华北落叶松人工林生物量对立地条件和林分结构的响应与模拟
1
2020
... 作为描述林分生长变异性的重要指标,林分结构在很大程度上影响着森林生长的时空动态,而森林经营很大程度上通过调控林分的结构实现[50-51].林分结构多样性的表征方式多样,如树种多样性、树木大小多样性、林木分布格局多样性等,空间和非空间分析法是量化林分结构多样性指标常用的两种方法[52].研究对象为沙地樟子松纯林,选取香农-威纳指数[53]、辛普森指数[54]、大小比数[55]、大小分异指数[56]、基尼系数[57]、变异系数[57]、偏度[10]、峰度[58]、Clark-Evans指数[59]等9种林分结构多样性指数来描述对象木对应结构单元的林分结构状况(表3).在计算林分结构多样性指数时,树木胸径按3 cm的径阶距划分. ...
林分空间结构参数二元分布的研究
2
2014
... 作为描述林分生长变异性的重要指标,林分结构在很大程度上影响着森林生长的时空动态,而森林经营很大程度上通过调控林分的结构实现[50-51].林分结构多样性的表征方式多样,如树种多样性、树木大小多样性、林木分布格局多样性等,空间和非空间分析法是量化林分结构多样性指标常用的两种方法[52].研究对象为沙地樟子松纯林,选取香农-威纳指数[53]、辛普森指数[54]、大小比数[55]、大小分异指数[56]、基尼系数[57]、变异系数[57]、偏度[10]、峰度[58]、Clark-Evans指数[59]等9种林分结构多样性指数来描述对象木对应结构单元的林分结构状况(表3).在计算林分结构多样性指数时,树木胸径按3 cm的径阶距划分. ...
... 森林结构体现为树木个体及其种类、大小、分布等属性的连接方式,它决定着森林功能,是树木生长等生态过程的主要驱动因素.在一定范围内,树种多样性及大小差异越大代表结构多样性越高[52,65]. 很多研究表明结构越复杂的森林具有越高的生产力[66-69].Peter等[66]发现,树种丰富度与森林生产力之间存在明显的正反馈.而Elizabeth等[67]也发现结构多样性是预测森林生产力的可靠指标,在一定程度上说明结构复杂性对树木生长的影响不可忽略.较高的结构多样性,也有利于减缓干旱对树木生长的不利影响.有研究显示,全球气候暖干化会导致森林生长衰退,具有高物种多样性的森林因干旱而导致的树木生长损失更少,结构复杂性不是单独发挥作用的[68].然而,对于天然纯林来说,虽然树种单一,但结构多样性对树木生长也具有较大的影响.例如,Sha等[12]对青海云杉(Picea crassifolia Kom.)的研究表明树木生长受树木大小分异指数的显著影响.我们的研究也表明,树木大小比数(NCI)、树木大小分异指数(DFI)以及Clark-Evans指数(CE)也是影响天然沙地樟子松纯林树木生长的重要因素.樟子松树木与其周围相邻木的大小比数及分异指数的值越大,可能表明樟子松树木受周围相邻木的竞争压力更大,进而导致树木生长量降低.而Clark-Evans指数表征着树木个体在空间上的分布状况,本研究表明樟子松树木生长随Clark-Evans指数的增加而显著降低,这可能与本研究中树木的分布格局对竞争的影响有关.我们发现胸径较大的樟子松树木多呈聚集分布状态,而胸径较小的树木更多呈均匀分布,这或许是导致这一结果的重要原因.然而,林分结构复杂性对于树木生长的影响并不是固定的.Zeller等[69]发现,随着森林演替的进行,结构复杂性对树木生长的影响由早期的抑制逐渐转变为促进作用.未来的研究,应结合树木年轮学等的方法,关注更长时间尺度上林分结构复杂性及其对树木生长影响的变化. ...
A mathematical theory of communication
2
1948
... 作为描述林分生长变异性的重要指标,林分结构在很大程度上影响着森林生长的时空动态,而森林经营很大程度上通过调控林分的结构实现[50-51].林分结构多样性的表征方式多样,如树种多样性、树木大小多样性、林木分布格局多样性等,空间和非空间分析法是量化林分结构多样性指标常用的两种方法[52].研究对象为沙地樟子松纯林,选取香农-威纳指数[53]、辛普森指数[54]、大小比数[55]、大小分异指数[56]、基尼系数[57]、变异系数[57]、偏度[10]、峰度[58]、Clark-Evans指数[59]等9种林分结构多样性指数来描述对象木对应结构单元的林分结构状况(表3).在计算林分结构多样性指数时,树木胸径按3 cm的径阶距划分. ...
... Stand structure diversity indices
Table 3| 名称 | 公式 | 意义 | 文献 |
|---|
| 香农-威纳指数 | | SW越大,树木大小差异越大 | [53] |
| 辛普森指数 | | S数大,代表树木大小分化程度高 | [54] |
| 大小比数 | | NCI越大,表示相邻木相比于目标木优势越大 | [55] |
| 大小分异指数 | | DFI越大,代表树木大小差异越大 | [56] |
| 基尼系数 | | GC越大,代表树木大小分布越不平衡 | [57] |
| 变异系数 | | CV越小,代表林木大小差异越小 | [57] |
| 偏度 | | SK越大,代表树木大小分布偏态程度越高 | [10] |
| 峰度 | | Kg越大,树木胸径最大值越大,分布越陡峭 | [58] |
| Clark-Evans指数 | | 判断林木种群空间分布格局(聚集、随机、均匀)的指数 | [59] |
注:n为树木i所在结构单元内竞争木数量;di 为目标木胸径,dj 为竞争木胸径,为结构单元内树木平均胸径,σd 为结构单元内林木胸径标准差;gi 为目标木胸高断面积;S为径阶数目;pm 为第m个径阶林木株数占相应结构单元林木总株数的比例.Vij 为目标木i比它的第j株相邻木小时,Vij =1;树木i比它的第j株相邻木大时,Vij =0.ri 为树木i与其最近一棵树的距离;A为结构单元样圆的面积.CE<1时为聚集分布,CE=1时为随机分布,CE>1时为均匀分布. ...
Measurement of diversity
2
1949
... 作为描述林分生长变异性的重要指标,林分结构在很大程度上影响着森林生长的时空动态,而森林经营很大程度上通过调控林分的结构实现[50-51].林分结构多样性的表征方式多样,如树种多样性、树木大小多样性、林木分布格局多样性等,空间和非空间分析法是量化林分结构多样性指标常用的两种方法[52].研究对象为沙地樟子松纯林,选取香农-威纳指数[53]、辛普森指数[54]、大小比数[55]、大小分异指数[56]、基尼系数[57]、变异系数[57]、偏度[10]、峰度[58]、Clark-Evans指数[59]等9种林分结构多样性指数来描述对象木对应结构单元的林分结构状况(表3).在计算林分结构多样性指数时,树木胸径按3 cm的径阶距划分. ...
... Stand structure diversity indices
Table 3| 名称 | 公式 | 意义 | 文献 |
|---|
| 香农-威纳指数 | | SW越大,树木大小差异越大 | [53] |
| 辛普森指数 | | S数大,代表树木大小分化程度高 | [54] |
| 大小比数 | | NCI越大,表示相邻木相比于目标木优势越大 | [55] |
| 大小分异指数 | | DFI越大,代表树木大小差异越大 | [56] |
| 基尼系数 | | GC越大,代表树木大小分布越不平衡 | [57] |
| 变异系数 | | CV越小,代表林木大小差异越小 | [57] |
| 偏度 | | SK越大,代表树木大小分布偏态程度越高 | [10] |
| 峰度 | | Kg越大,树木胸径最大值越大,分布越陡峭 | [58] |
| Clark-Evans指数 | | 判断林木种群空间分布格局(聚集、随机、均匀)的指数 | [59] |
注:n为树木i所在结构单元内竞争木数量;di 为目标木胸径,dj 为竞争木胸径,为结构单元内树木平均胸径,σd 为结构单元内林木胸径标准差;gi 为目标木胸高断面积;S为径阶数目;pm 为第m个径阶林木株数占相应结构单元林木总株数的比例.Vij 为目标木i比它的第j株相邻木小时,Vij =1;树木i比它的第j株相邻木大时,Vij =0.ri 为树木i与其最近一棵树的距离;A为结构单元样圆的面积.CE<1时为聚集分布,CE=1时为随机分布,CE>1时为均匀分布. ...
一个新的林分空间结构参数-大小比数
2
1999
... 作为描述林分生长变异性的重要指标,林分结构在很大程度上影响着森林生长的时空动态,而森林经营很大程度上通过调控林分的结构实现[50-51].林分结构多样性的表征方式多样,如树种多样性、树木大小多样性、林木分布格局多样性等,空间和非空间分析法是量化林分结构多样性指标常用的两种方法[52].研究对象为沙地樟子松纯林,选取香农-威纳指数[53]、辛普森指数[54]、大小比数[55]、大小分异指数[56]、基尼系数[57]、变异系数[57]、偏度[10]、峰度[58]、Clark-Evans指数[59]等9种林分结构多样性指数来描述对象木对应结构单元的林分结构状况(表3).在计算林分结构多样性指数时,树木胸径按3 cm的径阶距划分. ...
... Stand structure diversity indices
Table 3| 名称 | 公式 | 意义 | 文献 |
|---|
| 香农-威纳指数 | | SW越大,树木大小差异越大 | [53] |
| 辛普森指数 | | S数大,代表树木大小分化程度高 | [54] |
| 大小比数 | | NCI越大,表示相邻木相比于目标木优势越大 | [55] |
| 大小分异指数 | | DFI越大,代表树木大小差异越大 | [56] |
| 基尼系数 | | GC越大,代表树木大小分布越不平衡 | [57] |
| 变异系数 | | CV越小,代表林木大小差异越小 | [57] |
| 偏度 | | SK越大,代表树木大小分布偏态程度越高 | [10] |
| 峰度 | | Kg越大,树木胸径最大值越大,分布越陡峭 | [58] |
| Clark-Evans指数 | | 判断林木种群空间分布格局(聚集、随机、均匀)的指数 | [59] |
注:n为树木i所在结构单元内竞争木数量;di 为目标木胸径,dj 为竞争木胸径,为结构单元内树木平均胸径,σd 为结构单元内林木胸径标准差;gi 为目标木胸高断面积;S为径阶数目;pm 为第m个径阶林木株数占相应结构单元林木总株数的比例.Vij 为目标木i比它的第j株相邻木小时,Vij =1;树木i比它的第j株相邻木大时,Vij =0.ri 为树木i与其最近一棵树的距离;A为结构单元样圆的面积.CE<1时为聚集分布,CE=1时为随机分布,CE>1时为均匀分布. ...
Approaches to quantifying forest structures
2
2002
... 作为描述林分生长变异性的重要指标,林分结构在很大程度上影响着森林生长的时空动态,而森林经营很大程度上通过调控林分的结构实现[50-51].林分结构多样性的表征方式多样,如树种多样性、树木大小多样性、林木分布格局多样性等,空间和非空间分析法是量化林分结构多样性指标常用的两种方法[52].研究对象为沙地樟子松纯林,选取香农-威纳指数[53]、辛普森指数[54]、大小比数[55]、大小分异指数[56]、基尼系数[57]、变异系数[57]、偏度[10]、峰度[58]、Clark-Evans指数[59]等9种林分结构多样性指数来描述对象木对应结构单元的林分结构状况(表3).在计算林分结构多样性指数时,树木胸径按3 cm的径阶距划分. ...
... Stand structure diversity indices
Table 3| 名称 | 公式 | 意义 | 文献 |
|---|
| 香农-威纳指数 | | SW越大,树木大小差异越大 | [53] |
| 辛普森指数 | | S数大,代表树木大小分化程度高 | [54] |
| 大小比数 | | NCI越大,表示相邻木相比于目标木优势越大 | [55] |
| 大小分异指数 | | DFI越大,代表树木大小差异越大 | [56] |
| 基尼系数 | | GC越大,代表树木大小分布越不平衡 | [57] |
| 变异系数 | | CV越小,代表林木大小差异越小 | [57] |
| 偏度 | | SK越大,代表树木大小分布偏态程度越高 | [10] |
| 峰度 | | Kg越大,树木胸径最大值越大,分布越陡峭 | [58] |
| Clark-Evans指数 | | 判断林木种群空间分布格局(聚集、随机、均匀)的指数 | [59] |
注:n为树木i所在结构单元内竞争木数量;di 为目标木胸径,dj 为竞争木胸径,为结构单元内树木平均胸径,σd 为结构单元内林木胸径标准差;gi 为目标木胸高断面积;S为径阶数目;pm 为第m个径阶林木株数占相应结构单元林木总株数的比例.Vij 为目标木i比它的第j株相邻木小时,Vij =1;树木i比它的第j株相邻木大时,Vij =0.ri 为树木i与其最近一棵树的距离;A为结构单元样圆的面积.CE<1时为聚集分布,CE=1时为随机分布,CE>1时为均匀分布. ...
吉林蛟河阔叶红松林木本植物物种多样性及群落结构与生产力的关系
4
2017
... 作为描述林分生长变异性的重要指标,林分结构在很大程度上影响着森林生长的时空动态,而森林经营很大程度上通过调控林分的结构实现[50-51].林分结构多样性的表征方式多样,如树种多样性、树木大小多样性、林木分布格局多样性等,空间和非空间分析法是量化林分结构多样性指标常用的两种方法[52].研究对象为沙地樟子松纯林,选取香农-威纳指数[53]、辛普森指数[54]、大小比数[55]、大小分异指数[56]、基尼系数[57]、变异系数[57]、偏度[10]、峰度[58]、Clark-Evans指数[59]等9种林分结构多样性指数来描述对象木对应结构单元的林分结构状况(表3).在计算林分结构多样性指数时,树木胸径按3 cm的径阶距划分. ...
... [57]、偏度[10]、峰度[58]、Clark-Evans指数[59]等9种林分结构多样性指数来描述对象木对应结构单元的林分结构状况(表3).在计算林分结构多样性指数时,树木胸径按3 cm的径阶距划分. ...
... Stand structure diversity indices
Table 3| 名称 | 公式 | 意义 | 文献 |
|---|
| 香农-威纳指数 | | SW越大,树木大小差异越大 | [53] |
| 辛普森指数 | | S数大,代表树木大小分化程度高 | [54] |
| 大小比数 | | NCI越大,表示相邻木相比于目标木优势越大 | [55] |
| 大小分异指数 | | DFI越大,代表树木大小差异越大 | [56] |
| 基尼系数 | | GC越大,代表树木大小分布越不平衡 | [57] |
| 变异系数 | | CV越小,代表林木大小差异越小 | [57] |
| 偏度 | | SK越大,代表树木大小分布偏态程度越高 | [10] |
| 峰度 | | Kg越大,树木胸径最大值越大,分布越陡峭 | [58] |
| Clark-Evans指数 | | 判断林木种群空间分布格局(聚集、随机、均匀)的指数 | [59] |
注:n为树木i所在结构单元内竞争木数量;di 为目标木胸径,dj 为竞争木胸径,为结构单元内树木平均胸径,σd 为结构单元内林木胸径标准差;gi 为目标木胸高断面积;S为径阶数目;pm 为第m个径阶林木株数占相应结构单元林木总株数的比例.Vij 为目标木i比它的第j株相邻木小时,Vij =1;树木i比它的第j株相邻木大时,Vij =0.ri 为树木i与其最近一棵树的距离;A为结构单元样圆的面积.CE<1时为聚集分布,CE=1时为随机分布,CE>1时为均匀分布. ...
... [
57]
| 偏度 | | SK越大,代表树木大小分布偏态程度越高 | [10] |
| 峰度 | | Kg越大,树木胸径最大值越大,分布越陡峭 | [58] |
| Clark-Evans指数 | | 判断林木种群空间分布格局(聚集、随机、均匀)的指数 | [59] |
注:n为树木i所在结构单元内竞争木数量;di 为目标木胸径,dj 为竞争木胸径,为结构单元内树木平均胸径,σd 为结构单元内林木胸径标准差;gi 为目标木胸高断面积;S为径阶数目;pm 为第m个径阶林木株数占相应结构单元林木总株数的比例.Vij 为目标木i比它的第j株相邻木小时,Vij =1;树木i比它的第j株相邻木大时,Vij =0.ri 为树木i与其最近一棵树的距离;A为结构单元样圆的面积.CE<1时为聚集分布,CE=1时为随机分布,CE>1时为均匀分布. ...
2
2019
... 作为描述林分生长变异性的重要指标,林分结构在很大程度上影响着森林生长的时空动态,而森林经营很大程度上通过调控林分的结构实现[50-51].林分结构多样性的表征方式多样,如树种多样性、树木大小多样性、林木分布格局多样性等,空间和非空间分析法是量化林分结构多样性指标常用的两种方法[52].研究对象为沙地樟子松纯林,选取香农-威纳指数[53]、辛普森指数[54]、大小比数[55]、大小分异指数[56]、基尼系数[57]、变异系数[57]、偏度[10]、峰度[58]、Clark-Evans指数[59]等9种林分结构多样性指数来描述对象木对应结构单元的林分结构状况(表3).在计算林分结构多样性指数时,树木胸径按3 cm的径阶距划分. ...
... Stand structure diversity indices
Table 3| 名称 | 公式 | 意义 | 文献 |
|---|
| 香农-威纳指数 | | SW越大,树木大小差异越大 | [53] |
| 辛普森指数 | | S数大,代表树木大小分化程度高 | [54] |
| 大小比数 | | NCI越大,表示相邻木相比于目标木优势越大 | [55] |
| 大小分异指数 | | DFI越大,代表树木大小差异越大 | [56] |
| 基尼系数 | | GC越大,代表树木大小分布越不平衡 | [57] |
| 变异系数 | | CV越小,代表林木大小差异越小 | [57] |
| 偏度 | | SK越大,代表树木大小分布偏态程度越高 | [10] |
| 峰度 | | Kg越大,树木胸径最大值越大,分布越陡峭 | [58] |
| Clark-Evans指数 | | 判断林木种群空间分布格局(聚集、随机、均匀)的指数 | [59] |
注:n为树木i所在结构单元内竞争木数量;di 为目标木胸径,dj 为竞争木胸径,为结构单元内树木平均胸径,σd 为结构单元内林木胸径标准差;gi 为目标木胸高断面积;S为径阶数目;pm 为第m个径阶林木株数占相应结构单元林木总株数的比例.Vij 为目标木i比它的第j株相邻木小时,Vij =1;树木i比它的第j株相邻木大时,Vij =0.ri 为树木i与其最近一棵树的距离;A为结构单元样圆的面积.CE<1时为聚集分布,CE=1时为随机分布,CE>1时为均匀分布. ...
Distance to nearest neighbor as measure of spatial relationships in populations
2
1954
... 作为描述林分生长变异性的重要指标,林分结构在很大程度上影响着森林生长的时空动态,而森林经营很大程度上通过调控林分的结构实现[50-51].林分结构多样性的表征方式多样,如树种多样性、树木大小多样性、林木分布格局多样性等,空间和非空间分析法是量化林分结构多样性指标常用的两种方法[52].研究对象为沙地樟子松纯林,选取香农-威纳指数[53]、辛普森指数[54]、大小比数[55]、大小分异指数[56]、基尼系数[57]、变异系数[57]、偏度[10]、峰度[58]、Clark-Evans指数[59]等9种林分结构多样性指数来描述对象木对应结构单元的林分结构状况(表3).在计算林分结构多样性指数时,树木胸径按3 cm的径阶距划分. ...
... Stand structure diversity indices
Table 3| 名称 | 公式 | 意义 | 文献 |
|---|
| 香农-威纳指数 | | SW越大,树木大小差异越大 | [53] |
| 辛普森指数 | | S数大,代表树木大小分化程度高 | [54] |
| 大小比数 | | NCI越大,表示相邻木相比于目标木优势越大 | [55] |
| 大小分异指数 | | DFI越大,代表树木大小差异越大 | [56] |
| 基尼系数 | | GC越大,代表树木大小分布越不平衡 | [57] |
| 变异系数 | | CV越小,代表林木大小差异越小 | [57] |
| 偏度 | | SK越大,代表树木大小分布偏态程度越高 | [10] |
| 峰度 | | Kg越大,树木胸径最大值越大,分布越陡峭 | [58] |
| Clark-Evans指数 | | 判断林木种群空间分布格局(聚集、随机、均匀)的指数 | [59] |
注:n为树木i所在结构单元内竞争木数量;di 为目标木胸径,dj 为竞争木胸径,为结构单元内树木平均胸径,σd 为结构单元内林木胸径标准差;gi 为目标木胸高断面积;S为径阶数目;pm 为第m个径阶林木株数占相应结构单元林木总株数的比例.Vij 为目标木i比它的第j株相邻木小时,Vij =1;树木i比它的第j株相邻木大时,Vij =0.ri 为树木i与其最近一棵树的距离;A为结构单元样圆的面积.CE<1时为聚集分布,CE=1时为随机分布,CE>1时为均匀分布. ...
Analysis of tree rings and fire scars to establish fire history
1
1983
... 野外采集的树芯样本,用白乳胶固定在树芯槽上,在室温下自然风干,避免阳光暴晒,而后用600目的砂纸打磨,直至显微镜下可以清晰辨别年轮的边界.使用Lintab 6年轮分析系统(Rintech, Heidelberg, 德国)进行年轮宽度的测量,测量精度0.01 mm,利用COFECHA程序[60]检验矫正测量结果.用dplR包[61]处理矫正后年轮宽度测量结果,得到每年胸高断面积增长量序列.为排除气候因素对树木生长的影响,本研究采用5年(2011—2016年)累积胸高断面积生长量来表征树木的生长量: ...
A dendrochronology program library in R(dplR)
1
2008
... 野外采集的树芯样本,用白乳胶固定在树芯槽上,在室温下自然风干,避免阳光暴晒,而后用600目的砂纸打磨,直至显微镜下可以清晰辨别年轮的边界.使用Lintab 6年轮分析系统(Rintech, Heidelberg, 德国)进行年轮宽度的测量,测量精度0.01 mm,利用COFECHA程序[60]检验矫正测量结果.用dplR包[61]处理矫正后年轮宽度测量结果,得到每年胸高断面积增长量序列.为排除气候因素对树木生长的影响,本研究采用5年(2011—2016年)累积胸高断面积生长量来表征树木的生长量: ...
Generalizing hierarchical and variation partitioning in multiple regression and canonical analyses using the rdacca.hp R package
1
2022
... 式中:a为常数项;是对应的第n个自变量;为第n个自变量的回归系数.本研究中,选取与BAI相关性最强的基于距离的竞争指数和与距离无关的竞争指数分别建立回归模型,为消除自变量之间的共线性问题,纳入模型的自变量方差膨胀因子小于10.基于R2、调整R2、RSE、AIC等指标确定最优模型.利用R语言rdacca.hp包[62]估算不同因子解释率,利用UpSetVP包[63]绘制不同因子的相对重要性排序. ...
UpSetVP:An Alternative Visualization of VPA and HP in Canonical Analysis
1
... 式中:a为常数项;是对应的第n个自变量;为第n个自变量的回归系数.本研究中,选取与BAI相关性最强的基于距离的竞争指数和与距离无关的竞争指数分别建立回归模型,为消除自变量之间的共线性问题,纳入模型的自变量方差膨胀因子小于10.基于R2、调整R2、RSE、AIC等指标确定最优模型.利用R语言rdacca.hp包[62]估算不同因子解释率,利用UpSetVP包[63]绘制不同因子的相对重要性排序. ...
R程序包“rdacca.hp”在生态学数据分析中的应用:案例与进展
1
2023
... 为揭示林分结构、竞争和树木本身特性对樟子松树木生长的相对贡献,通过变差分解和层次分割进行分析.结果表明,竞争是解释树木生长变异性的最重要变量,解释了31.98%的变异,其中15.5%由其单独解释;其次为冠幅,解释了14.97%的变异;树木大小分异指数解释了12.30%的变异,高径比解释了11.51%的变异(图5).不同因素之间的交互作用也是影响树木生长的关键因素,竞争、冠幅、大小分异指数和高径比共4个因素的交互作用解释了24.83%的生长变异,这说明不同变量之间还存在较强的相互作用,不能仅通过边际效应来估计不同解释变量的相对重要性[64]. ...
Methods of forest structure research:a review
1
2019
... 森林结构体现为树木个体及其种类、大小、分布等属性的连接方式,它决定着森林功能,是树木生长等生态过程的主要驱动因素.在一定范围内,树种多样性及大小差异越大代表结构多样性越高[52,65]. 很多研究表明结构越复杂的森林具有越高的生产力[66-69].Peter等[66]发现,树种丰富度与森林生产力之间存在明显的正反馈.而Elizabeth等[67]也发现结构多样性是预测森林生产力的可靠指标,在一定程度上说明结构复杂性对树木生长的影响不可忽略.较高的结构多样性,也有利于减缓干旱对树木生长的不利影响.有研究显示,全球气候暖干化会导致森林生长衰退,具有高物种多样性的森林因干旱而导致的树木生长损失更少,结构复杂性不是单独发挥作用的[68].然而,对于天然纯林来说,虽然树种单一,但结构多样性对树木生长也具有较大的影响.例如,Sha等[12]对青海云杉(Picea crassifolia Kom.)的研究表明树木生长受树木大小分异指数的显著影响.我们的研究也表明,树木大小比数(NCI)、树木大小分异指数(DFI)以及Clark-Evans指数(CE)也是影响天然沙地樟子松纯林树木生长的重要因素.樟子松树木与其周围相邻木的大小比数及分异指数的值越大,可能表明樟子松树木受周围相邻木的竞争压力更大,进而导致树木生长量降低.而Clark-Evans指数表征着树木个体在空间上的分布状况,本研究表明樟子松树木生长随Clark-Evans指数的增加而显著降低,这可能与本研究中树木的分布格局对竞争的影响有关.我们发现胸径较大的樟子松树木多呈聚集分布状态,而胸径较小的树木更多呈均匀分布,这或许是导致这一结果的重要原因.然而,林分结构复杂性对于树木生长的影响并不是固定的.Zeller等[69]发现,随着森林演替的进行,结构复杂性对树木生长的影响由早期的抑制逐渐转变为促进作用.未来的研究,应结合树木年轮学等的方法,关注更长时间尺度上林分结构复杂性及其对树木生长影响的变化. ...
Tree diversity effects on productivity depend on mycorrhizae and life strategies in a temperate forest experiment
2
2022
... 森林结构体现为树木个体及其种类、大小、分布等属性的连接方式,它决定着森林功能,是树木生长等生态过程的主要驱动因素.在一定范围内,树种多样性及大小差异越大代表结构多样性越高[52,65]. 很多研究表明结构越复杂的森林具有越高的生产力[66-69].Peter等[66]发现,树种丰富度与森林生产力之间存在明显的正反馈.而Elizabeth等[67]也发现结构多样性是预测森林生产力的可靠指标,在一定程度上说明结构复杂性对树木生长的影响不可忽略.较高的结构多样性,也有利于减缓干旱对树木生长的不利影响.有研究显示,全球气候暖干化会导致森林生长衰退,具有高物种多样性的森林因干旱而导致的树木生长损失更少,结构复杂性不是单独发挥作用的[68].然而,对于天然纯林来说,虽然树种单一,但结构多样性对树木生长也具有较大的影响.例如,Sha等[12]对青海云杉(Picea crassifolia Kom.)的研究表明树木生长受树木大小分异指数的显著影响.我们的研究也表明,树木大小比数(NCI)、树木大小分异指数(DFI)以及Clark-Evans指数(CE)也是影响天然沙地樟子松纯林树木生长的重要因素.樟子松树木与其周围相邻木的大小比数及分异指数的值越大,可能表明樟子松树木受周围相邻木的竞争压力更大,进而导致树木生长量降低.而Clark-Evans指数表征着树木个体在空间上的分布状况,本研究表明樟子松树木生长随Clark-Evans指数的增加而显著降低,这可能与本研究中树木的分布格局对竞争的影响有关.我们发现胸径较大的樟子松树木多呈聚集分布状态,而胸径较小的树木更多呈均匀分布,这或许是导致这一结果的重要原因.然而,林分结构复杂性对于树木生长的影响并不是固定的.Zeller等[69]发现,随着森林演替的进行,结构复杂性对树木生长的影响由早期的抑制逐渐转变为促进作用.未来的研究,应结合树木年轮学等的方法,关注更长时间尺度上林分结构复杂性及其对树木生长影响的变化. ...
... [66]发现,树种丰富度与森林生产力之间存在明显的正反馈.而Elizabeth等[67]也发现结构多样性是预测森林生产力的可靠指标,在一定程度上说明结构复杂性对树木生长的影响不可忽略.较高的结构多样性,也有利于减缓干旱对树木生长的不利影响.有研究显示,全球气候暖干化会导致森林生长衰退,具有高物种多样性的森林因干旱而导致的树木生长损失更少,结构复杂性不是单独发挥作用的[68].然而,对于天然纯林来说,虽然树种单一,但结构多样性对树木生长也具有较大的影响.例如,Sha等[12]对青海云杉(Picea crassifolia Kom.)的研究表明树木生长受树木大小分异指数的显著影响.我们的研究也表明,树木大小比数(NCI)、树木大小分异指数(DFI)以及Clark-Evans指数(CE)也是影响天然沙地樟子松纯林树木生长的重要因素.樟子松树木与其周围相邻木的大小比数及分异指数的值越大,可能表明樟子松树木受周围相邻木的竞争压力更大,进而导致树木生长量降低.而Clark-Evans指数表征着树木个体在空间上的分布状况,本研究表明樟子松树木生长随Clark-Evans指数的增加而显著降低,这可能与本研究中树木的分布格局对竞争的影响有关.我们发现胸径较大的樟子松树木多呈聚集分布状态,而胸径较小的树木更多呈均匀分布,这或许是导致这一结果的重要原因.然而,林分结构复杂性对于树木生长的影响并不是固定的.Zeller等[69]发现,随着森林演替的进行,结构复杂性对树木生长的影响由早期的抑制逐渐转变为促进作用.未来的研究,应结合树木年轮学等的方法,关注更长时间尺度上林分结构复杂性及其对树木生长影响的变化. ...
Structural diversity as a reliable and novel predictor for ecosystem productivity
1
2023
... 森林结构体现为树木个体及其种类、大小、分布等属性的连接方式,它决定着森林功能,是树木生长等生态过程的主要驱动因素.在一定范围内,树种多样性及大小差异越大代表结构多样性越高[52,65]. 很多研究表明结构越复杂的森林具有越高的生产力[66-69].Peter等[66]发现,树种丰富度与森林生产力之间存在明显的正反馈.而Elizabeth等[67]也发现结构多样性是预测森林生产力的可靠指标,在一定程度上说明结构复杂性对树木生长的影响不可忽略.较高的结构多样性,也有利于减缓干旱对树木生长的不利影响.有研究显示,全球气候暖干化会导致森林生长衰退,具有高物种多样性的森林因干旱而导致的树木生长损失更少,结构复杂性不是单独发挥作用的[68].然而,对于天然纯林来说,虽然树种单一,但结构多样性对树木生长也具有较大的影响.例如,Sha等[12]对青海云杉(Picea crassifolia Kom.)的研究表明树木生长受树木大小分异指数的显著影响.我们的研究也表明,树木大小比数(NCI)、树木大小分异指数(DFI)以及Clark-Evans指数(CE)也是影响天然沙地樟子松纯林树木生长的重要因素.樟子松树木与其周围相邻木的大小比数及分异指数的值越大,可能表明樟子松树木受周围相邻木的竞争压力更大,进而导致树木生长量降低.而Clark-Evans指数表征着树木个体在空间上的分布状况,本研究表明樟子松树木生长随Clark-Evans指数的增加而显著降低,这可能与本研究中树木的分布格局对竞争的影响有关.我们发现胸径较大的樟子松树木多呈聚集分布状态,而胸径较小的树木更多呈均匀分布,这或许是导致这一结果的重要原因.然而,林分结构复杂性对于树木生长的影响并不是固定的.Zeller等[69]发现,随着森林演替的进行,结构复杂性对树木生长的影响由早期的抑制逐渐转变为促进作用.未来的研究,应结合树木年轮学等的方法,关注更长时间尺度上林分结构复杂性及其对树木生长影响的变化. ...
Tree diversity and identity modulate the growth response of thermophilous deciduous forests to climate warming
1
2022
... 森林结构体现为树木个体及其种类、大小、分布等属性的连接方式,它决定着森林功能,是树木生长等生态过程的主要驱动因素.在一定范围内,树种多样性及大小差异越大代表结构多样性越高[52,65]. 很多研究表明结构越复杂的森林具有越高的生产力[66-69].Peter等[66]发现,树种丰富度与森林生产力之间存在明显的正反馈.而Elizabeth等[67]也发现结构多样性是预测森林生产力的可靠指标,在一定程度上说明结构复杂性对树木生长的影响不可忽略.较高的结构多样性,也有利于减缓干旱对树木生长的不利影响.有研究显示,全球气候暖干化会导致森林生长衰退,具有高物种多样性的森林因干旱而导致的树木生长损失更少,结构复杂性不是单独发挥作用的[68].然而,对于天然纯林来说,虽然树种单一,但结构多样性对树木生长也具有较大的影响.例如,Sha等[12]对青海云杉(Picea crassifolia Kom.)的研究表明树木生长受树木大小分异指数的显著影响.我们的研究也表明,树木大小比数(NCI)、树木大小分异指数(DFI)以及Clark-Evans指数(CE)也是影响天然沙地樟子松纯林树木生长的重要因素.樟子松树木与其周围相邻木的大小比数及分异指数的值越大,可能表明樟子松树木受周围相邻木的竞争压力更大,进而导致树木生长量降低.而Clark-Evans指数表征着树木个体在空间上的分布状况,本研究表明樟子松树木生长随Clark-Evans指数的增加而显著降低,这可能与本研究中树木的分布格局对竞争的影响有关.我们发现胸径较大的樟子松树木多呈聚集分布状态,而胸径较小的树木更多呈均匀分布,这或许是导致这一结果的重要原因.然而,林分结构复杂性对于树木生长的影响并不是固定的.Zeller等[69]发现,随着森林演替的进行,结构复杂性对树木生长的影响由早期的抑制逐渐转变为促进作用.未来的研究,应结合树木年轮学等的方法,关注更长时间尺度上林分结构复杂性及其对树木生长影响的变化. ...
Effect of forest structure on stand productivity in Central European forests depends on developmental stage and tree species diversity
3
2019
... 森林结构体现为树木个体及其种类、大小、分布等属性的连接方式,它决定着森林功能,是树木生长等生态过程的主要驱动因素.在一定范围内,树种多样性及大小差异越大代表结构多样性越高[52,65]. 很多研究表明结构越复杂的森林具有越高的生产力[66-69].Peter等[66]发现,树种丰富度与森林生产力之间存在明显的正反馈.而Elizabeth等[67]也发现结构多样性是预测森林生产力的可靠指标,在一定程度上说明结构复杂性对树木生长的影响不可忽略.较高的结构多样性,也有利于减缓干旱对树木生长的不利影响.有研究显示,全球气候暖干化会导致森林生长衰退,具有高物种多样性的森林因干旱而导致的树木生长损失更少,结构复杂性不是单独发挥作用的[68].然而,对于天然纯林来说,虽然树种单一,但结构多样性对树木生长也具有较大的影响.例如,Sha等[12]对青海云杉(Picea crassifolia Kom.)的研究表明树木生长受树木大小分异指数的显著影响.我们的研究也表明,树木大小比数(NCI)、树木大小分异指数(DFI)以及Clark-Evans指数(CE)也是影响天然沙地樟子松纯林树木生长的重要因素.樟子松树木与其周围相邻木的大小比数及分异指数的值越大,可能表明樟子松树木受周围相邻木的竞争压力更大,进而导致树木生长量降低.而Clark-Evans指数表征着树木个体在空间上的分布状况,本研究表明樟子松树木生长随Clark-Evans指数的增加而显著降低,这可能与本研究中树木的分布格局对竞争的影响有关.我们发现胸径较大的樟子松树木多呈聚集分布状态,而胸径较小的树木更多呈均匀分布,这或许是导致这一结果的重要原因.然而,林分结构复杂性对于树木生长的影响并不是固定的.Zeller等[69]发现,随着森林演替的进行,结构复杂性对树木生长的影响由早期的抑制逐渐转变为促进作用.未来的研究,应结合树木年轮学等的方法,关注更长时间尺度上林分结构复杂性及其对树木生长影响的变化. ...
... [69]发现,随着森林演替的进行,结构复杂性对树木生长的影响由早期的抑制逐渐转变为促进作用.未来的研究,应结合树木年轮学等的方法,关注更长时间尺度上林分结构复杂性及其对树木生长影响的变化. ...
... 树木生长往往同时受到很多因素,诸如气候、土壤、地形、林分结构等的综合影响.例如窦啸文等[74]发现天目山常绿阔叶林树木径向生长同时受到胸径、冠幅和竞争的共同作用.然而,这些因素在决定树木生长中的相对重要性却各不相同.韩大校等[76]在黑龙江凉水自然保护区阔叶红松林的研究中发现, 大径级树木的径向生长主要受地形因素的影响,而小径级树木的径向生长主要受到竞争的影响.也有研究认为树木的竞争作用,而非气候是限制树木生长的最主要因素[73].我们研究发现,当同时考虑林木本身属性、林分结构、单木竞争时,竞争与结构多样性显著影响了沙地樟子松树木径向生长.竞争是影响樟子松树木生长的最重要因素,其解释了约三分之一的变异.而结构多样性因素,即树木大小分异指数解释了樟子松生长约12%的变异性,而树木自身的特性,包括树木的冠幅和高径比共同解释了树木生长约四分之一的变异性,这与前人研究[17-18,69,74]基本一致.同时,我们也发现,林分结构多样性与竞争间存在交互作用[12,19],共同影响了樟子松树木的生长,气候同时也影响了树木生长动态[77],未来的研究应重视不同因子的交互作用对树木生长的影响.另外,本研究的样本量相对较小,导致研究结果可能存在一定局限性,未来研究需要从更长时间尺度、更大地理范围开展研究,以增强结果的普适性与可靠性. ...
不同竞争强度下的沙地樟子松天然林树木径向生长及其气候响应
1
2019
... 不同树木个体间的竞争,通过影响对光照、水分、养分等资源的获取而影响着树木的生长,是制约树木生长的重要因素[70-71].有学者在研究不同竞争强度下欧洲赤松(Pinus sylvestris L.)树木径向生长对气候的响应时发现,树木径向生长和竞争之间呈负指数关系,竞争强度的增加使树木径向生长速率放缓[72].中国亚热带地区马尾松(Pinus massoniana)的生长量也随竞争指数增加而降低[73].刘耀凤等[71]、窦啸文等[74]和康剑等[4]在研究中也发现竞争对树木生长的类似效应.但上述研究大都只是直接选取了常用的竞争指数来描述林木个体间相互作用,这样或许不能准确地代表林木间实际竞争强度,产生有偏估计.Britton等[75]发现当不能正确描述林木生长和林木大小间关系(绝对生长量和生长率)时,不考虑竞争指数的类型,直接选用一个常用的竞争指数可能会夸大断面积生长过程中竞争的作用.基于此,为确定最优竞争指数,本研究考虑了28种基于距离的竞争指数和18种与距离无关的竞争指数,分析其对沙地樟子松树木生长的影响.结果表明,树木生长和不同竞争指数的相关性存在较大差异,即林木生长对不同竞争指数表征的竞争强度的响应存在较大差异,进一步证明了考虑竞争指数类型的必要性. ...
竞争对天然针阔混交林树木直径生长的影响
2
2023
... 不同树木个体间的竞争,通过影响对光照、水分、养分等资源的获取而影响着树木的生长,是制约树木生长的重要因素[70-71].有学者在研究不同竞争强度下欧洲赤松(Pinus sylvestris L.)树木径向生长对气候的响应时发现,树木径向生长和竞争之间呈负指数关系,竞争强度的增加使树木径向生长速率放缓[72].中国亚热带地区马尾松(Pinus massoniana)的生长量也随竞争指数增加而降低[73].刘耀凤等[71]、窦啸文等[74]和康剑等[4]在研究中也发现竞争对树木生长的类似效应.但上述研究大都只是直接选取了常用的竞争指数来描述林木个体间相互作用,这样或许不能准确地代表林木间实际竞争强度,产生有偏估计.Britton等[75]发现当不能正确描述林木生长和林木大小间关系(绝对生长量和生长率)时,不考虑竞争指数的类型,直接选用一个常用的竞争指数可能会夸大断面积生长过程中竞争的作用.基于此,为确定最优竞争指数,本研究考虑了28种基于距离的竞争指数和18种与距离无关的竞争指数,分析其对沙地樟子松树木生长的影响.结果表明,树木生长和不同竞争指数的相关性存在较大差异,即林木生长对不同竞争指数表征的竞争强度的响应存在较大差异,进一步证明了考虑竞争指数类型的必要性. ...
... [71]、窦啸文等[74]和康剑等[4]在研究中也发现竞争对树木生长的类似效应.但上述研究大都只是直接选取了常用的竞争指数来描述林木个体间相互作用,这样或许不能准确地代表林木间实际竞争强度,产生有偏估计.Britton等[75]发现当不能正确描述林木生长和林木大小间关系(绝对生长量和生长率)时,不考虑竞争指数的类型,直接选用一个常用的竞争指数可能会夸大断面积生长过程中竞争的作用.基于此,为确定最优竞争指数,本研究考虑了28种基于距离的竞争指数和18种与距离无关的竞争指数,分析其对沙地樟子松树木生长的影响.结果表明,树木生长和不同竞争指数的相关性存在较大差异,即林木生长对不同竞争指数表征的竞争强度的响应存在较大差异,进一步证明了考虑竞争指数类型的必要性. ...
Disentangling the effects of competition and climate on individual tree growth:a retrospective and dynamic approach in Scots pine
1
2015
... 不同树木个体间的竞争,通过影响对光照、水分、养分等资源的获取而影响着树木的生长,是制约树木生长的重要因素[70-71].有学者在研究不同竞争强度下欧洲赤松(Pinus sylvestris L.)树木径向生长对气候的响应时发现,树木径向生长和竞争之间呈负指数关系,竞争强度的增加使树木径向生长速率放缓[72].中国亚热带地区马尾松(Pinus massoniana)的生长量也随竞争指数增加而降低[73].刘耀凤等[71]、窦啸文等[74]和康剑等[4]在研究中也发现竞争对树木生长的类似效应.但上述研究大都只是直接选取了常用的竞争指数来描述林木个体间相互作用,这样或许不能准确地代表林木间实际竞争强度,产生有偏估计.Britton等[75]发现当不能正确描述林木生长和林木大小间关系(绝对生长量和生长率)时,不考虑竞争指数的类型,直接选用一个常用的竞争指数可能会夸大断面积生长过程中竞争的作用.基于此,为确定最优竞争指数,本研究考虑了28种基于距离的竞争指数和18种与距离无关的竞争指数,分析其对沙地樟子松树木生长的影响.结果表明,树木生长和不同竞争指数的相关性存在较大差异,即林木生长对不同竞争指数表征的竞争强度的响应存在较大差异,进一步证明了考虑竞争指数类型的必要性. ...
Contributions of competition and climate on radial growth of Pinus massoniana in subtropics of China
2
2019
... 不同树木个体间的竞争,通过影响对光照、水分、养分等资源的获取而影响着树木的生长,是制约树木生长的重要因素[70-71].有学者在研究不同竞争强度下欧洲赤松(Pinus sylvestris L.)树木径向生长对气候的响应时发现,树木径向生长和竞争之间呈负指数关系,竞争强度的增加使树木径向生长速率放缓[72].中国亚热带地区马尾松(Pinus massoniana)的生长量也随竞争指数增加而降低[73].刘耀凤等[71]、窦啸文等[74]和康剑等[4]在研究中也发现竞争对树木生长的类似效应.但上述研究大都只是直接选取了常用的竞争指数来描述林木个体间相互作用,这样或许不能准确地代表林木间实际竞争强度,产生有偏估计.Britton等[75]发现当不能正确描述林木生长和林木大小间关系(绝对生长量和生长率)时,不考虑竞争指数的类型,直接选用一个常用的竞争指数可能会夸大断面积生长过程中竞争的作用.基于此,为确定最优竞争指数,本研究考虑了28种基于距离的竞争指数和18种与距离无关的竞争指数,分析其对沙地樟子松树木生长的影响.结果表明,树木生长和不同竞争指数的相关性存在较大差异,即林木生长对不同竞争指数表征的竞争强度的响应存在较大差异,进一步证明了考虑竞争指数类型的必要性. ...
... 树木生长往往同时受到很多因素,诸如气候、土壤、地形、林分结构等的综合影响.例如窦啸文等[74]发现天目山常绿阔叶林树木径向生长同时受到胸径、冠幅和竞争的共同作用.然而,这些因素在决定树木生长中的相对重要性却各不相同.韩大校等[76]在黑龙江凉水自然保护区阔叶红松林的研究中发现, 大径级树木的径向生长主要受地形因素的影响,而小径级树木的径向生长主要受到竞争的影响.也有研究认为树木的竞争作用,而非气候是限制树木生长的最主要因素[73].我们研究发现,当同时考虑林木本身属性、林分结构、单木竞争时,竞争与结构多样性显著影响了沙地樟子松树木径向生长.竞争是影响樟子松树木生长的最重要因素,其解释了约三分之一的变异.而结构多样性因素,即树木大小分异指数解释了樟子松生长约12%的变异性,而树木自身的特性,包括树木的冠幅和高径比共同解释了树木生长约四分之一的变异性,这与前人研究[17-18,69,74]基本一致.同时,我们也发现,林分结构多样性与竞争间存在交互作用[12,19],共同影响了樟子松树木的生长,气候同时也影响了树木生长动态[77],未来的研究应重视不同因子的交互作用对树木生长的影响.另外,本研究的样本量相对较小,导致研究结果可能存在一定局限性,未来研究需要从更长时间尺度、更大地理范围开展研究,以增强结果的普适性与可靠性. ...
天目山常绿阔叶林胸高断面积生长量影响因子研究
3
2023
... 不同树木个体间的竞争,通过影响对光照、水分、养分等资源的获取而影响着树木的生长,是制约树木生长的重要因素[70-71].有学者在研究不同竞争强度下欧洲赤松(Pinus sylvestris L.)树木径向生长对气候的响应时发现,树木径向生长和竞争之间呈负指数关系,竞争强度的增加使树木径向生长速率放缓[72].中国亚热带地区马尾松(Pinus massoniana)的生长量也随竞争指数增加而降低[73].刘耀凤等[71]、窦啸文等[74]和康剑等[4]在研究中也发现竞争对树木生长的类似效应.但上述研究大都只是直接选取了常用的竞争指数来描述林木个体间相互作用,这样或许不能准确地代表林木间实际竞争强度,产生有偏估计.Britton等[75]发现当不能正确描述林木生长和林木大小间关系(绝对生长量和生长率)时,不考虑竞争指数的类型,直接选用一个常用的竞争指数可能会夸大断面积生长过程中竞争的作用.基于此,为确定最优竞争指数,本研究考虑了28种基于距离的竞争指数和18种与距离无关的竞争指数,分析其对沙地樟子松树木生长的影响.结果表明,树木生长和不同竞争指数的相关性存在较大差异,即林木生长对不同竞争指数表征的竞争强度的响应存在较大差异,进一步证明了考虑竞争指数类型的必要性. ...
... 树木生长往往同时受到很多因素,诸如气候、土壤、地形、林分结构等的综合影响.例如窦啸文等[74]发现天目山常绿阔叶林树木径向生长同时受到胸径、冠幅和竞争的共同作用.然而,这些因素在决定树木生长中的相对重要性却各不相同.韩大校等[76]在黑龙江凉水自然保护区阔叶红松林的研究中发现, 大径级树木的径向生长主要受地形因素的影响,而小径级树木的径向生长主要受到竞争的影响.也有研究认为树木的竞争作用,而非气候是限制树木生长的最主要因素[73].我们研究发现,当同时考虑林木本身属性、林分结构、单木竞争时,竞争与结构多样性显著影响了沙地樟子松树木径向生长.竞争是影响樟子松树木生长的最重要因素,其解释了约三分之一的变异.而结构多样性因素,即树木大小分异指数解释了樟子松生长约12%的变异性,而树木自身的特性,包括树木的冠幅和高径比共同解释了树木生长约四分之一的变异性,这与前人研究[17-18,69,74]基本一致.同时,我们也发现,林分结构多样性与竞争间存在交互作用[12,19],共同影响了樟子松树木的生长,气候同时也影响了树木生长动态[77],未来的研究应重视不同因子的交互作用对树木生长的影响.另外,本研究的样本量相对较小,导致研究结果可能存在一定局限性,未来研究需要从更长时间尺度、更大地理范围开展研究,以增强结果的普适性与可靠性. ...
... ,74]基本一致.同时,我们也发现,林分结构多样性与竞争间存在交互作用[12,19],共同影响了樟子松树木的生长,气候同时也影响了树木生长动态[77],未来的研究应重视不同因子的交互作用对树木生长的影响.另外,本研究的样本量相对较小,导致研究结果可能存在一定局限性,未来研究需要从更长时间尺度、更大地理范围开展研究,以增强结果的普适性与可靠性. ...
Quantifying neighbour effects on tree growth:Are common 'competition' indices biased?
1
2023
... 不同树木个体间的竞争,通过影响对光照、水分、养分等资源的获取而影响着树木的生长,是制约树木生长的重要因素[70-71].有学者在研究不同竞争强度下欧洲赤松(Pinus sylvestris L.)树木径向生长对气候的响应时发现,树木径向生长和竞争之间呈负指数关系,竞争强度的增加使树木径向生长速率放缓[72].中国亚热带地区马尾松(Pinus massoniana)的生长量也随竞争指数增加而降低[73].刘耀凤等[71]、窦啸文等[74]和康剑等[4]在研究中也发现竞争对树木生长的类似效应.但上述研究大都只是直接选取了常用的竞争指数来描述林木个体间相互作用,这样或许不能准确地代表林木间实际竞争强度,产生有偏估计.Britton等[75]发现当不能正确描述林木生长和林木大小间关系(绝对生长量和生长率)时,不考虑竞争指数的类型,直接选用一个常用的竞争指数可能会夸大断面积生长过程中竞争的作用.基于此,为确定最优竞争指数,本研究考虑了28种基于距离的竞争指数和18种与距离无关的竞争指数,分析其对沙地樟子松树木生长的影响.结果表明,树木生长和不同竞争指数的相关性存在较大差异,即林木生长对不同竞争指数表征的竞争强度的响应存在较大差异,进一步证明了考虑竞争指数类型的必要性. ...
地形和竞争对典型阔叶红松林不同生长阶段树木胸径生长的影响
1
2017
... 树木生长往往同时受到很多因素,诸如气候、土壤、地形、林分结构等的综合影响.例如窦啸文等[74]发现天目山常绿阔叶林树木径向生长同时受到胸径、冠幅和竞争的共同作用.然而,这些因素在决定树木生长中的相对重要性却各不相同.韩大校等[76]在黑龙江凉水自然保护区阔叶红松林的研究中发现, 大径级树木的径向生长主要受地形因素的影响,而小径级树木的径向生长主要受到竞争的影响.也有研究认为树木的竞争作用,而非气候是限制树木生长的最主要因素[73].我们研究发现,当同时考虑林木本身属性、林分结构、单木竞争时,竞争与结构多样性显著影响了沙地樟子松树木径向生长.竞争是影响樟子松树木生长的最重要因素,其解释了约三分之一的变异.而结构多样性因素,即树木大小分异指数解释了樟子松生长约12%的变异性,而树木自身的特性,包括树木的冠幅和高径比共同解释了树木生长约四分之一的变异性,这与前人研究[17-18,69,74]基本一致.同时,我们也发现,林分结构多样性与竞争间存在交互作用[12,19],共同影响了樟子松树木的生长,气候同时也影响了树木生长动态[77],未来的研究应重视不同因子的交互作用对树木生长的影响.另外,本研究的样本量相对较小,导致研究结果可能存在一定局限性,未来研究需要从更长时间尺度、更大地理范围开展研究,以增强结果的普适性与可靠性. ...
沙地樟子松(Pinus sylvestris var.mongolica)物候特征及其对气候的响应
1
2022
... 树木生长往往同时受到很多因素,诸如气候、土壤、地形、林分结构等的综合影响.例如窦啸文等[74]发现天目山常绿阔叶林树木径向生长同时受到胸径、冠幅和竞争的共同作用.然而,这些因素在决定树木生长中的相对重要性却各不相同.韩大校等[76]在黑龙江凉水自然保护区阔叶红松林的研究中发现, 大径级树木的径向生长主要受地形因素的影响,而小径级树木的径向生长主要受到竞争的影响.也有研究认为树木的竞争作用,而非气候是限制树木生长的最主要因素[73].我们研究发现,当同时考虑林木本身属性、林分结构、单木竞争时,竞争与结构多样性显著影响了沙地樟子松树木径向生长.竞争是影响樟子松树木生长的最重要因素,其解释了约三分之一的变异.而结构多样性因素,即树木大小分异指数解释了樟子松生长约12%的变异性,而树木自身的特性,包括树木的冠幅和高径比共同解释了树木生长约四分之一的变异性,这与前人研究[17-18,69,74]基本一致.同时,我们也发现,林分结构多样性与竞争间存在交互作用[12,19],共同影响了樟子松树木的生长,气候同时也影响了树木生长动态[77],未来的研究应重视不同因子的交互作用对树木生长的影响.另外,本研究的样本量相对较小,导致研究结果可能存在一定局限性,未来研究需要从更长时间尺度、更大地理范围开展研究,以增强结果的普适性与可靠性. ...
结构化森林经营研究进展
1
2018
... 从结构决定功能的角度出发,将林分尺度与单株尺度相结合来提升森林的生态功能是开展森林管理的重要方向,准确了解当前林分结构和竞争状况是提升森林生态功能的必然要求[78].同时,气候变化背景下,林分结构与竞争也是评估退化森林生态系统不可忽视的重要组分[79].未来,在沙地樟子松林分经营管理中,管理者应重视林分结构和密度调整,确定适宜的间伐或择伐强度,优化林分结构和竞争强度,增加树木大小分异性,从而有效提高林分生产力,增强林分的生长固碳功能与稳定性. ...
退化森林生态系统评价指标体系研究进展
1
2018
... 从结构决定功能的角度出发,将林分尺度与单株尺度相结合来提升森林的生态功能是开展森林管理的重要方向,准确了解当前林分结构和竞争状况是提升森林生态功能的必然要求[78].同时,气候变化背景下,林分结构与竞争也是评估退化森林生态系统不可忽视的重要组分[79].未来,在沙地樟子松林分经营管理中,管理者应重视林分结构和密度调整,确定适宜的间伐或择伐强度,优化林分结构和竞争强度,增加树木大小分异性,从而有效提高林分生产力,增强林分的生长固碳功能与稳定性. ...



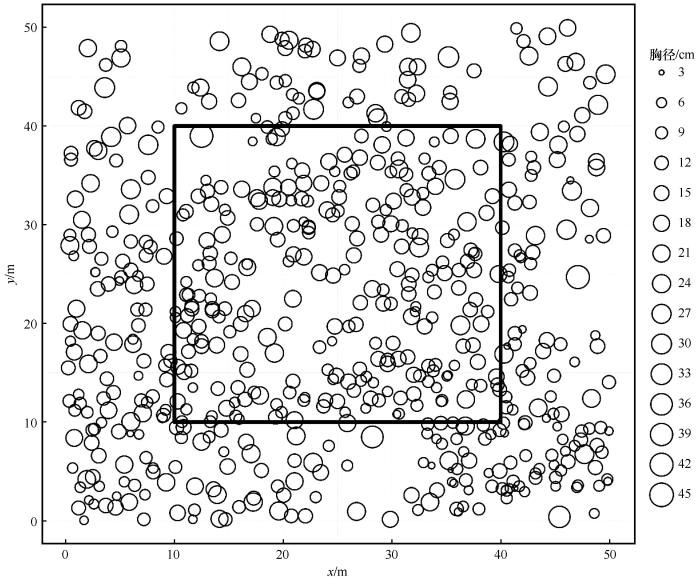
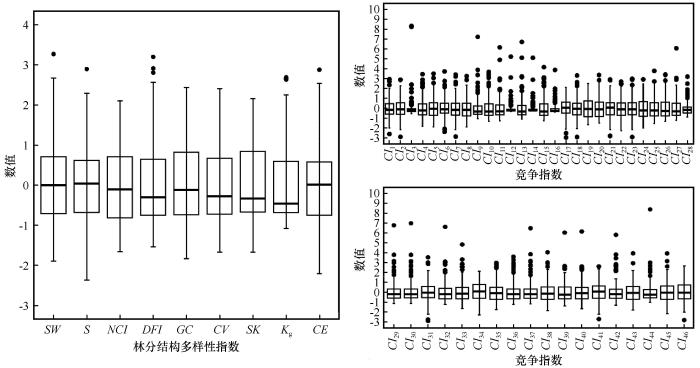
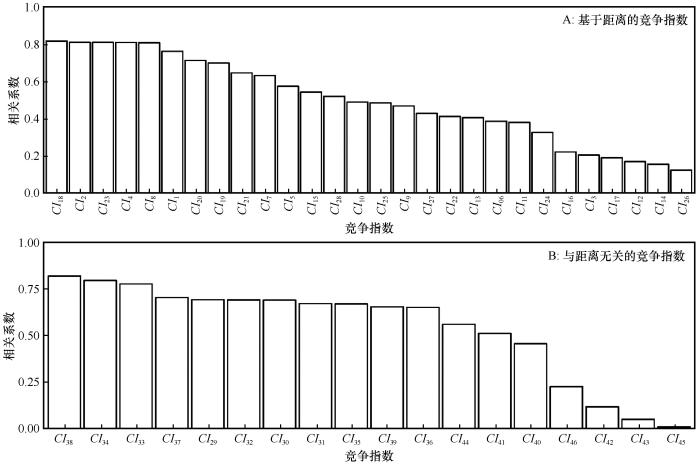
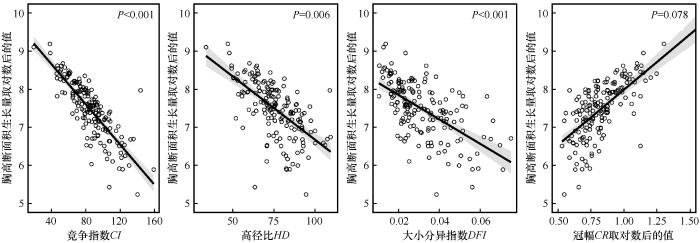
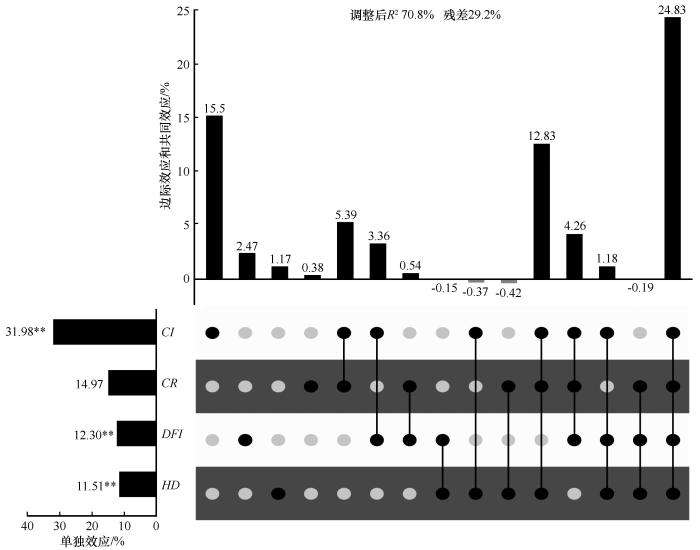

 甘公网安备 62010202000688号
甘公网安备 62010202000688号
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}