Carbon sequestration in dryland ecosystems
4
2004
... 受不合理土地利用和气候变化的共同影响,土地荒漠化已经成为干旱半干旱区最为严重的生态环境问题[1-3].这种广泛发生的土地退化使土壤有机碳(Soil organic carbon, SOC)大量流失,导致土壤质量、生态系统生产力和多样性显著下降,对人类健康和生产活动构成了严重威胁[4-6].植被恢复重建是荒漠化逆转和促进SOC积累的有效方法[1-2,5].从全球尺度来看,适宜的生态恢复措施在25~50 a内的碳固存速率为0.9~1.9 Pg·a-1,相当于当前大气二氧化碳含量的30%[1-2,7-8].因此,植被恢复过程中土壤碳固存潜力成为近年来提升土壤质量和应对气候变化研究的焦点[1-2,9-11]. ...
... [1-2,5].从全球尺度来看,适宜的生态恢复措施在25~50 a内的碳固存速率为0.9~1.9 Pg·a-1,相当于当前大气二氧化碳含量的30%[1-2,7-8].因此,植被恢复过程中土壤碳固存潜力成为近年来提升土壤质量和应对气候变化研究的焦点[1-2,9-11]. ...
... [1-2,7-8].因此,植被恢复过程中土壤碳固存潜力成为近年来提升土壤质量和应对气候变化研究的焦点[1-2,9-11]. ...
... [1-2,9-11]. ...
Recovery of soil carbon and nitrogen stocks following afforestation with xerophytic shrubs in the Tengger Desert,North China
15
2022
... 受不合理土地利用和气候变化的共同影响,土地荒漠化已经成为干旱半干旱区最为严重的生态环境问题[1-3].这种广泛发生的土地退化使土壤有机碳(Soil organic carbon, SOC)大量流失,导致土壤质量、生态系统生产力和多样性显著下降,对人类健康和生产活动构成了严重威胁[4-6].植被恢复重建是荒漠化逆转和促进SOC积累的有效方法[1-2,5].从全球尺度来看,适宜的生态恢复措施在25~50 a内的碳固存速率为0.9~1.9 Pg·a-1,相当于当前大气二氧化碳含量的30%[1-2,7-8].因此,植被恢复过程中土壤碳固存潜力成为近年来提升土壤质量和应对气候变化研究的焦点[1-2,9-11]. ...
... -2,7-8].因此,植被恢复过程中土壤碳固存潜力成为近年来提升土壤质量和应对气候变化研究的焦点[1-2,9-11]. ...
... -2,9-11]. ...
... 值得注意的是,SOC对土壤质量和肥力以及缓解气候变化贡献的有效性取决于其组成和稳定性,而大量研究表明,将SOC库视为均一的整体进行研究无法揭示相关问题[12].SOC通常由稳定性和周转速率不同的多个功能库组成[11-13],将SOC分为颗粒有机碳(Particulate organic carbon, POC, 粒径53 μm)和矿质结合有机碳(Mineral-associated organic carbon, MAOC, 粒径53 μm)成为近年来揭示其形成和稳定性的有效方法 [11-14].POC主要由植物来源的结构聚合物组成,能够通过生化抗性和团聚体保护在土壤中存在数年至数十年[10,15-16];而MAOC主要由微生物分解和POC解聚而产生的简单生物分子组成,通过物理化学机制吸附在矿物表面,可将其与生物和酶分离,从而在土壤中存在数百年至上千年[12,17].因而,MAOC是碳长期封存的关键指标,而POC可以预测SOC短期分解过程[18-19].因此,明确不同组分随植被恢复的变化及其影响因素对于准确评价SOC稳定性至关重要.然而,尽管近年来沙区植被恢复引发的SOC变化得到了广泛证实[2,9-11],但已有研究主要集中在SOC储量、周转过程随恢复过程的变化及其与环境因子的关系等方面[2,10,20-22],而对POC和MAOC分布及其对植被恢复响应的研究较为缺乏.因此,对SOC形成和稳定性及其机制的认识仍存在很大的不确定性[10,12],从而限制了对植被恢复的长期碳固存效应及其对全球碳循环贡献的准确评价[12-13]. ...
... [2,10,20-22],而对POC和MAOC分布及其对植被恢复响应的研究较为缺乏.因此,对SOC形成和稳定性及其机制的认识仍存在很大的不确定性[10,12],从而限制了对植被恢复的长期碳固存效应及其对全球碳循环贡献的准确评价[12-13]. ...
... 植被恢复过程中生物和非生物因素的协同变化强烈影响着SOC及其组分的稳定性.大量研究表明,伴随着植被恢复,植被组成、土壤理化性质及微生物群落组成发生了显著变化[2,9,23-24].这可能会最终影响碳氮在不同组分中的分配,并进一步影响其稳定性[11,25].SOC不同组分在来源、形成过程及保护机制等方面的差异使其对植被恢复引发的生物和非生物因子变化的响应各不相同[20].C∶N较小的凋落物的输入、土壤pH值的减小、活性矿质颗粒较为丰富的土壤以及植物多样性的恢复可使植物来源的有机物有效地转化为微生物产物,从而促进MAOC的形成 [26];而不利于微生物活动的环境条件(如温度、水分和土壤类型等)通常会阻碍植物残体的分解,进而导致POC的积累[12].因此,植被恢复过程中SOC的变化可能是多种因素共同作用的结果,而明确不同组分的驱动因子是揭示SOC稳定性维持机制的前提[12-15]. ...
... 自20世纪50年代开始,种植旱生灌木成为中国北方沙区实现荒漠化逆转的有效途径[2,27].特别是为了保护公路、铁路和水利设施等大型基础设施的安全,分不同阶段在西北干旱沙区种植了大量旱生灌木,形成了建植于不同年限的人工植被区,为研究植被恢复对SOC及其组分稳定性的影响提供了天然的研究平台.本文以甘肃省古浪县鸣沙咀不同年代人工植被区为研究对象,采用空间代替时间的方法,基于土壤有机碳组成和环境要素分析,拟回答以下问题:①SOC及其组分随植被恢复年限的变化规律;②植被恢复对SOC及其组成的影响及其机理.在此基础上,揭示沙区植被恢复对SOC组成及稳定性的影响及其机理. ...
... 2023年7月,分别选择在1982年(41 a,恢复年限,下同)、1991年(32 a)、2003年(20 a)和2011年(12 a)建立的人工植被区及流动沙丘(对照,0 a)作为研究样地.在每个样地中设置10 m×10 m的样方各3个(间距大于50 m),用于灌木植物调查.由于人工固沙植被区地形起伏明显且不同地形生物和非生物因子差异显著,因此,各年代植被区的样方选择设置于地势相对平坦的丘间低地,以减少地形造成的数据波动,从而提高数据的准确性和可比性.在每个10 m×10 m的样方中随机布设3个1 m×1 m小样方,用于草本植物调查.植物群落调查包括物种组成、植被盖度和地上及地下部分生物量.灌木和草本地上部分生物量采用收割法进行测定,在每个10 m×10 m的样方中,每种灌木选择1个标准株并采集其地上部分,装入信封;在每个1 m×1 m的小样方中,将草本地上部分齐地面收割,装入信封;随后收集小样方地面凋落物,装入信封.根系生物量用根钻进行采集,在每个10 m×10 m样方中的灌丛间隙随机选取10个点,用直径为5 cm的根钻钻取0~10 cm层土壤,将其过2 mm筛以分离出根系,将根系装入信封.根据已有研究,沙区植被恢复对土壤的影响集中在表层0~10 cm土层,草本植物根系的主要分布范围也在该土层[2,10,27],因此,根系生物量的采集点均设在草本植物分布的灌丛间斑块中.在每个10 m×10 m的样方中,用直径为5 cm的土钻采集0~10 cm层土壤,在每个样方中随机选取10个点进行采样,将同一样方中10个点的样品充分混合形成一个混合样,每个年代植被区获得3个混合样品(3个重复).为避免灌丛斑块下“沃岛效应”造成的数据波动,土壤样品的采集点均设在草本植物分布的灌丛间斑块.将所有土壤样品带回实验室并分为两份,一份在-20 ℃条件下保存,用于土壤微生物群落、酶活性和微生物量碳(Microbial Biomass Carbon,MBC)的测定,另一份室温条件下风干后过2 mm筛,用于土壤理化性质及POC和MAOC的测定.将生物量和凋落物样本带回实验室,在60 ℃条件下烘干,称重. ...
... 本研究中,固沙植被建立41 a后0~10 cm土层SOC含量比流沙增加9.59倍(表1,P0.01),表明种植旱生灌木是促进旱地SOC固存和土壤质量提升的有效措施.通常情况下,SOC的动态变化是由土壤有机质(Soil organic matter, SOM)输入和通过微生物分解的SOM输出过程共同决定的,当生态系统SOM输入大于输出时即表现为净SOC累积[2,24,31].一方面,固沙植被建立使植物来源SOM输入显著增大,其中地下部分生物量和凋落物分别解释了SOC变异的60%和90%(图3),而SOC与地上部分相关性不显著,这与Li等[2]的研究结果一致.这是因为随着植被演替,以灌木为优势的群落逐渐被以草本为优势的群落所取代,导致地上部分生物量呈减小趋势[11,24].另一方面,伴随着生态恢复,植被盖度显著增大(表1),沙丘表面逐渐趋于稳定,减少了风蚀引发的凋落物流失[2,32].且植物冠层能有效减小风速,增加了生态系统对大气降尘的捕获,而降尘中丰富的SOC和细颗粒物质提供了额外的碳源[2,33].此外,植被恢复促进了土壤理化性质的改善和土壤肥力的提升(表1),从而有利于植物生长以及SOM输入的增加[2,10].值得注意的是,固沙植被的建立促进了生物土壤结皮的拓殖和定居,腾格里沙漠东南缘的研究表明,生物土壤结皮的年固碳速率可达11.36~26.75 g·m-1[34],这也是表层土壤SOC的主要贡献者. ...
... [2]的研究结果一致.这是因为随着植被演替,以灌木为优势的群落逐渐被以草本为优势的群落所取代,导致地上部分生物量呈减小趋势[11,24].另一方面,伴随着生态恢复,植被盖度显著增大(表1),沙丘表面逐渐趋于稳定,减少了风蚀引发的凋落物流失[2,32].且植物冠层能有效减小风速,增加了生态系统对大气降尘的捕获,而降尘中丰富的SOC和细颗粒物质提供了额外的碳源[2,33].此外,植被恢复促进了土壤理化性质的改善和土壤肥力的提升(表1),从而有利于植物生长以及SOM输入的增加[2,10].值得注意的是,固沙植被的建立促进了生物土壤结皮的拓殖和定居,腾格里沙漠东南缘的研究表明,生物土壤结皮的年固碳速率可达11.36~26.75 g·m-1[34],这也是表层土壤SOC的主要贡献者. ...
... [2,32].且植物冠层能有效减小风速,增加了生态系统对大气降尘的捕获,而降尘中丰富的SOC和细颗粒物质提供了额外的碳源[2,33].此外,植被恢复促进了土壤理化性质的改善和土壤肥力的提升(表1),从而有利于植物生长以及SOM输入的增加[2,10].值得注意的是,固沙植被的建立促进了生物土壤结皮的拓殖和定居,腾格里沙漠东南缘的研究表明,生物土壤结皮的年固碳速率可达11.36~26.75 g·m-1[34],这也是表层土壤SOC的主要贡献者. ...
... [2,33].此外,植被恢复促进了土壤理化性质的改善和土壤肥力的提升(表1),从而有利于植物生长以及SOM输入的增加[2,10].值得注意的是,固沙植被的建立促进了生物土壤结皮的拓殖和定居,腾格里沙漠东南缘的研究表明,生物土壤结皮的年固碳速率可达11.36~26.75 g·m-1[34],这也是表层土壤SOC的主要贡献者. ...
... [2,10].值得注意的是,固沙植被的建立促进了生物土壤结皮的拓殖和定居,腾格里沙漠东南缘的研究表明,生物土壤结皮的年固碳速率可达11.36~26.75 g·m-1[34],这也是表层土壤SOC的主要贡献者. ...
... 已有研究表明,SOC的稳定性取决于其与矿物颗粒的相互作用和微生物的可及性,而MAOC与土壤矿物颗粒形成牢固的化学键,在提高SOC稳定性和长期封存方面发挥着至关重要的作用.因此,MAOC作为受微生物分解影响最小的碳库是SOC稳定性的重要表征[15].在本研究中,MAOC和POC含量及MAOC占SOC的比例随植被恢复年限的延长而显著增大,而POC占SOC的比例则呈相反趋势(图2,P0.01).这表明沙区植被恢复不仅促进了SOC的积累,同时使SOC组成由以POC为主逐渐向以MAOC为主转变,进而改变了SOC库的稳定性.POC主要来源于植物,由颗粒有机物的结构聚合物组成[14-17].因此,固沙植被区凋落物和地下部分生物量的显著增加是POC含量增大的主要原因,分别解释了其变异的89.3%和57.9%(图3).另一方面,固沙区土壤肥力的提升,特别是氮含量的显著增加不但促进SOM输入的增加和MBC的增大,而且加强了土壤团聚体保护[35],使POC含量进一步增加.已有研究表明,植物碳的微生物转化过程是SOC形成和稳定性维持的关键影响因素[36],植被恢复不仅增加了SOM的输入,而且增加土壤孔隙度和持水性,且使微生物基质(碳和养分)的数量和质量发生了显著变化(图1),土壤微环境更有利于与有机质分解相关的微生物群落的生长和繁殖[2,24];而EC的增大可通过增加微生物代谢熵而促进微生物对底物的利用,进一步促进微生物群落的发育[37],从而促进了植物残体的分解和POC和MAOC的累积[2,35],其中微生物群落变化解释了其变异的69%~85%(图3).另一方面,SOM的增加增强了土壤颗粒的胶结作用,有利于团聚体的形成,进一步促进了POC的增加[12-14].本研究中,POC占SOC的比例随植被恢复而显著减小,这主要是由于植被恢复引发的生物和非生物因子的改变,从而促进了MAOC截存,使得SOC库由以POC为主逐渐向以MAOC为主转变. ...
... [2,35],其中微生物群落变化解释了其变异的69%~85%(图3).另一方面,SOM的增加增强了土壤颗粒的胶结作用,有利于团聚体的形成,进一步促进了POC的增加[12-14].本研究中,POC占SOC的比例随植被恢复而显著减小,这主要是由于植被恢复引发的生物和非生物因子的改变,从而促进了MAOC截存,使得SOC库由以POC为主逐渐向以MAOC为主转变. ...
Global desertification:building a science for dryland development
1
2007
... 受不合理土地利用和气候变化的共同影响,土地荒漠化已经成为干旱半干旱区最为严重的生态环境问题[1-3].这种广泛发生的土地退化使土壤有机碳(Soil organic carbon, SOC)大量流失,导致土壤质量、生态系统生产力和多样性显著下降,对人类健康和生产活动构成了严重威胁[4-6].植被恢复重建是荒漠化逆转和促进SOC积累的有效方法[1-2,5].从全球尺度来看,适宜的生态恢复措施在25~50 a内的碳固存速率为0.9~1.9 Pg·a-1,相当于当前大气二氧化碳含量的30%[1-2,7-8].因此,植被恢复过程中土壤碳固存潜力成为近年来提升土壤质量和应对气候变化研究的焦点[1-2,9-11]. ...
Accelerated dryland expansion under climate change
1
2016
... 受不合理土地利用和气候变化的共同影响,土地荒漠化已经成为干旱半干旱区最为严重的生态环境问题[1-3].这种广泛发生的土地退化使土壤有机碳(Soil organic carbon, SOC)大量流失,导致土壤质量、生态系统生产力和多样性显著下降,对人类健康和生产活动构成了严重威胁[4-6].植被恢复重建是荒漠化逆转和促进SOC积累的有效方法[1-2,5].从全球尺度来看,适宜的生态恢复措施在25~50 a内的碳固存速率为0.9~1.9 Pg·a-1,相当于当前大气二氧化碳含量的30%[1-2,7-8].因此,植被恢复过程中土壤碳固存潜力成为近年来提升土壤质量和应对气候变化研究的焦点[1-2,9-11]. ...
Biogeography of global drylands
1
2021
... 受不合理土地利用和气候变化的共同影响,土地荒漠化已经成为干旱半干旱区最为严重的生态环境问题[1-3].这种广泛发生的土地退化使土壤有机碳(Soil organic carbon, SOC)大量流失,导致土壤质量、生态系统生产力和多样性显著下降,对人类健康和生产活动构成了严重威胁[4-6].植被恢复重建是荒漠化逆转和促进SOC积累的有效方法[1-2,5].从全球尺度来看,适宜的生态恢复措施在25~50 a内的碳固存速率为0.9~1.9 Pg·a-1,相当于当前大气二氧化碳含量的30%[1-2,7-8].因此,植被恢复过程中土壤碳固存潜力成为近年来提升土壤质量和应对气候变化研究的焦点[1-2,9-11]. ...
Carbon cycling in global drylands
1
2019
... 受不合理土地利用和气候变化的共同影响,土地荒漠化已经成为干旱半干旱区最为严重的生态环境问题[1-3].这种广泛发生的土地退化使土壤有机碳(Soil organic carbon, SOC)大量流失,导致土壤质量、生态系统生产力和多样性显著下降,对人类健康和生产活动构成了严重威胁[4-6].植被恢复重建是荒漠化逆转和促进SOC积累的有效方法[1-2,5].从全球尺度来看,适宜的生态恢复措施在25~50 a内的碳固存速率为0.9~1.9 Pg·a-1,相当于当前大气二氧化碳含量的30%[1-2,7-8].因此,植被恢复过程中土壤碳固存潜力成为近年来提升土壤质量和应对气候变化研究的焦点[1-2,9-11]. ...
Potential of desertification control to sequester carbon and mitigate the greenhouse effect
1
2001
... 受不合理土地利用和气候变化的共同影响,土地荒漠化已经成为干旱半干旱区最为严重的生态环境问题[1-3].这种广泛发生的土地退化使土壤有机碳(Soil organic carbon, SOC)大量流失,导致土壤质量、生态系统生产力和多样性显著下降,对人类健康和生产活动构成了严重威胁[4-6].植被恢复重建是荒漠化逆转和促进SOC积累的有效方法[1-2,5].从全球尺度来看,适宜的生态恢复措施在25~50 a内的碳固存速率为0.9~1.9 Pg·a-1,相当于当前大气二氧化碳含量的30%[1-2,7-8].因此,植被恢复过程中土壤碳固存潜力成为近年来提升土壤质量和应对气候变化研究的焦点[1-2,9-11]. ...
1
2007
... 受不合理土地利用和气候变化的共同影响,土地荒漠化已经成为干旱半干旱区最为严重的生态环境问题[1-3].这种广泛发生的土地退化使土壤有机碳(Soil organic carbon, SOC)大量流失,导致土壤质量、生态系统生产力和多样性显著下降,对人类健康和生产活动构成了严重威胁[4-6].植被恢复重建是荒漠化逆转和促进SOC积累的有效方法[1-2,5].从全球尺度来看,适宜的生态恢复措施在25~50 a内的碳固存速率为0.9~1.9 Pg·a-1,相当于当前大气二氧化碳含量的30%[1-2,7-8].因此,植被恢复过程中土壤碳固存潜力成为近年来提升土壤质量和应对气候变化研究的焦点[1-2,9-11]. ...
Desert soil sequesters atmospheric CO2 by microbial mineral formation
3
2020
... 受不合理土地利用和气候变化的共同影响,土地荒漠化已经成为干旱半干旱区最为严重的生态环境问题[1-3].这种广泛发生的土地退化使土壤有机碳(Soil organic carbon, SOC)大量流失,导致土壤质量、生态系统生产力和多样性显著下降,对人类健康和生产活动构成了严重威胁[4-6].植被恢复重建是荒漠化逆转和促进SOC积累的有效方法[1-2,5].从全球尺度来看,适宜的生态恢复措施在25~50 a内的碳固存速率为0.9~1.9 Pg·a-1,相当于当前大气二氧化碳含量的30%[1-2,7-8].因此,植被恢复过程中土壤碳固存潜力成为近年来提升土壤质量和应对气候变化研究的焦点[1-2,9-11]. ...
... 值得注意的是,SOC对土壤质量和肥力以及缓解气候变化贡献的有效性取决于其组成和稳定性,而大量研究表明,将SOC库视为均一的整体进行研究无法揭示相关问题[12].SOC通常由稳定性和周转速率不同的多个功能库组成[11-13],将SOC分为颗粒有机碳(Particulate organic carbon, POC, 粒径53 μm)和矿质结合有机碳(Mineral-associated organic carbon, MAOC, 粒径53 μm)成为近年来揭示其形成和稳定性的有效方法 [11-14].POC主要由植物来源的结构聚合物组成,能够通过生化抗性和团聚体保护在土壤中存在数年至数十年[10,15-16];而MAOC主要由微生物分解和POC解聚而产生的简单生物分子组成,通过物理化学机制吸附在矿物表面,可将其与生物和酶分离,从而在土壤中存在数百年至上千年[12,17].因而,MAOC是碳长期封存的关键指标,而POC可以预测SOC短期分解过程[18-19].因此,明确不同组分随植被恢复的变化及其影响因素对于准确评价SOC稳定性至关重要.然而,尽管近年来沙区植被恢复引发的SOC变化得到了广泛证实[2,9-11],但已有研究主要集中在SOC储量、周转过程随恢复过程的变化及其与环境因子的关系等方面[2,10,20-22],而对POC和MAOC分布及其对植被恢复响应的研究较为缺乏.因此,对SOC形成和稳定性及其机制的认识仍存在很大的不确定性[10,12],从而限制了对植被恢复的长期碳固存效应及其对全球碳循环贡献的准确评价[12-13]. ...
... 植被恢复过程中生物和非生物因素的协同变化强烈影响着SOC及其组分的稳定性.大量研究表明,伴随着植被恢复,植被组成、土壤理化性质及微生物群落组成发生了显著变化[2,9,23-24].这可能会最终影响碳氮在不同组分中的分配,并进一步影响其稳定性[11,25].SOC不同组分在来源、形成过程及保护机制等方面的差异使其对植被恢复引发的生物和非生物因子变化的响应各不相同[20].C∶N较小的凋落物的输入、土壤pH值的减小、活性矿质颗粒较为丰富的土壤以及植物多样性的恢复可使植物来源的有机物有效地转化为微生物产物,从而促进MAOC的形成 [26];而不利于微生物活动的环境条件(如温度、水分和土壤类型等)通常会阻碍植物残体的分解,进而导致POC的积累[12].因此,植被恢复过程中SOC的变化可能是多种因素共同作用的结果,而明确不同组分的驱动因子是揭示SOC稳定性维持机制的前提[12-15]. ...
Revegetation promotes soil mineral-associated organic carbon sequestration and soil carbon stability in the Tengger Desert,Northern China
6
2023
... 值得注意的是,SOC对土壤质量和肥力以及缓解气候变化贡献的有效性取决于其组成和稳定性,而大量研究表明,将SOC库视为均一的整体进行研究无法揭示相关问题[12].SOC通常由稳定性和周转速率不同的多个功能库组成[11-13],将SOC分为颗粒有机碳(Particulate organic carbon, POC, 粒径53 μm)和矿质结合有机碳(Mineral-associated organic carbon, MAOC, 粒径53 μm)成为近年来揭示其形成和稳定性的有效方法 [11-14].POC主要由植物来源的结构聚合物组成,能够通过生化抗性和团聚体保护在土壤中存在数年至数十年[10,15-16];而MAOC主要由微生物分解和POC解聚而产生的简单生物分子组成,通过物理化学机制吸附在矿物表面,可将其与生物和酶分离,从而在土壤中存在数百年至上千年[12,17].因而,MAOC是碳长期封存的关键指标,而POC可以预测SOC短期分解过程[18-19].因此,明确不同组分随植被恢复的变化及其影响因素对于准确评价SOC稳定性至关重要.然而,尽管近年来沙区植被恢复引发的SOC变化得到了广泛证实[2,9-11],但已有研究主要集中在SOC储量、周转过程随恢复过程的变化及其与环境因子的关系等方面[2,10,20-22],而对POC和MAOC分布及其对植被恢复响应的研究较为缺乏.因此,对SOC形成和稳定性及其机制的认识仍存在很大的不确定性[10,12],从而限制了对植被恢复的长期碳固存效应及其对全球碳循环贡献的准确评价[12-13]. ...
... ,10,20-22],而对POC和MAOC分布及其对植被恢复响应的研究较为缺乏.因此,对SOC形成和稳定性及其机制的认识仍存在很大的不确定性[10,12],从而限制了对植被恢复的长期碳固存效应及其对全球碳循环贡献的准确评价[12-13]. ...
... [10,12],从而限制了对植被恢复的长期碳固存效应及其对全球碳循环贡献的准确评价[12-13]. ...
... 2023年7月,分别选择在1982年(41 a,恢复年限,下同)、1991年(32 a)、2003年(20 a)和2011年(12 a)建立的人工植被区及流动沙丘(对照,0 a)作为研究样地.在每个样地中设置10 m×10 m的样方各3个(间距大于50 m),用于灌木植物调查.由于人工固沙植被区地形起伏明显且不同地形生物和非生物因子差异显著,因此,各年代植被区的样方选择设置于地势相对平坦的丘间低地,以减少地形造成的数据波动,从而提高数据的准确性和可比性.在每个10 m×10 m的样方中随机布设3个1 m×1 m小样方,用于草本植物调查.植物群落调查包括物种组成、植被盖度和地上及地下部分生物量.灌木和草本地上部分生物量采用收割法进行测定,在每个10 m×10 m的样方中,每种灌木选择1个标准株并采集其地上部分,装入信封;在每个1 m×1 m的小样方中,将草本地上部分齐地面收割,装入信封;随后收集小样方地面凋落物,装入信封.根系生物量用根钻进行采集,在每个10 m×10 m样方中的灌丛间隙随机选取10个点,用直径为5 cm的根钻钻取0~10 cm层土壤,将其过2 mm筛以分离出根系,将根系装入信封.根据已有研究,沙区植被恢复对土壤的影响集中在表层0~10 cm土层,草本植物根系的主要分布范围也在该土层[2,10,27],因此,根系生物量的采集点均设在草本植物分布的灌丛间斑块中.在每个10 m×10 m的样方中,用直径为5 cm的土钻采集0~10 cm层土壤,在每个样方中随机选取10个点进行采样,将同一样方中10个点的样品充分混合形成一个混合样,每个年代植被区获得3个混合样品(3个重复).为避免灌丛斑块下“沃岛效应”造成的数据波动,土壤样品的采集点均设在草本植物分布的灌丛间斑块.将所有土壤样品带回实验室并分为两份,一份在-20 ℃条件下保存,用于土壤微生物群落、酶活性和微生物量碳(Microbial Biomass Carbon,MBC)的测定,另一份室温条件下风干后过2 mm筛,用于土壤理化性质及POC和MAOC的测定.将生物量和凋落物样本带回实验室,在60 ℃条件下烘干,称重. ...
... 本研究中,固沙植被建立41 a后0~10 cm土层SOC含量比流沙增加9.59倍(表1,P0.01),表明种植旱生灌木是促进旱地SOC固存和土壤质量提升的有效措施.通常情况下,SOC的动态变化是由土壤有机质(Soil organic matter, SOM)输入和通过微生物分解的SOM输出过程共同决定的,当生态系统SOM输入大于输出时即表现为净SOC累积[2,24,31].一方面,固沙植被建立使植物来源SOM输入显著增大,其中地下部分生物量和凋落物分别解释了SOC变异的60%和90%(图3),而SOC与地上部分相关性不显著,这与Li等[2]的研究结果一致.这是因为随着植被演替,以灌木为优势的群落逐渐被以草本为优势的群落所取代,导致地上部分生物量呈减小趋势[11,24].另一方面,伴随着生态恢复,植被盖度显著增大(表1),沙丘表面逐渐趋于稳定,减少了风蚀引发的凋落物流失[2,32].且植物冠层能有效减小风速,增加了生态系统对大气降尘的捕获,而降尘中丰富的SOC和细颗粒物质提供了额外的碳源[2,33].此外,植被恢复促进了土壤理化性质的改善和土壤肥力的提升(表1),从而有利于植物生长以及SOM输入的增加[2,10].值得注意的是,固沙植被的建立促进了生物土壤结皮的拓殖和定居,腾格里沙漠东南缘的研究表明,生物土壤结皮的年固碳速率可达11.36~26.75 g·m-1[34],这也是表层土壤SOC的主要贡献者. ...
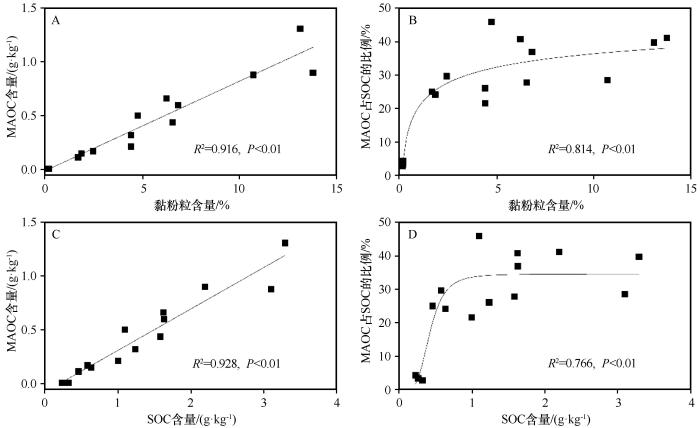
... 土壤黏粉粒是MAOC的关键影响因素,植被恢复使土壤黏粉粒含量显著增大(表1),这增加了矿物对SOM的可及性,从而促进MAOC的形成[14-15].本研究中,黏粉粒含量是与MAOC相关性最大的因子,解释了其变量的95.7%(图3).而土壤微生物对凋落物和POC的分解是形成MAOC的先决条件[15,19,38],因此,MAOC的变化与POC密切相关(图3).微生物将较粗的植物来源聚合物(POC)进行分解,形成更易溶解的化合物(微生物细胞和细胞外化合物),这些化合物与土壤矿物质结合形成MAOC[36],从而使固沙区土壤MAOC显著增加(图2).另一方面,细菌和真菌在土壤中丰度的增加,可分泌更多的黏液和多糖,进一步促进稳定性碳的形成[39].而真菌可通过其丝状菌丝体聚集矿物颗粒,促进了有机分子的吸附和固定从而有利于形成MAOC[40].放线菌多样性的显著增大可有效促进土壤腐殖质的积累和转化,这也是形成稳定有机碳的重要过程[41].值得注意的是,POC的分解过程中释放的养分和能量能完全满足微生物生长的需求,从而减少了因养分需求而对MAOC的分解,进而有利于MAOC的累积[10].伴随着植被恢复,碳分解酶BG、CBH的活性和MBC也显著增大(图1).两种酶不但可以促进植物残体分解,同时能将SOC转化为可溶性葡萄糖和其他低分子量碳化合物并为微生物提供能量和碳源,从而促进POC和MAOC的积累[42-43].而MBC的增大是MAOC形成的重要前提,可有效促进SOC与矿物质之间的相互作用,从而形成更多的MAOC[41].微生物残体是MAOC的关键组成部分,微生物多样性及生物量的增大可使微生物残体及代谢分泌物显著增加,从而有利于MAOC的累积[12-13].土壤氮含量的增加可通过土壤有机残体与氮的缩合反应、促进有机-无机复合物的形成以及丛枝菌根真菌(AMF)的丰度增大从而促进植物残体形成MAOC[35,44]. ...
Divergent changes of carbon and nitrogen in the density fractions of soil organic matter after revegetation in the Tengger Desert,North China
6
2024
... 受不合理土地利用和气候变化的共同影响,土地荒漠化已经成为干旱半干旱区最为严重的生态环境问题[1-3].这种广泛发生的土地退化使土壤有机碳(Soil organic carbon, SOC)大量流失,导致土壤质量、生态系统生产力和多样性显著下降,对人类健康和生产活动构成了严重威胁[4-6].植被恢复重建是荒漠化逆转和促进SOC积累的有效方法[1-2,5].从全球尺度来看,适宜的生态恢复措施在25~50 a内的碳固存速率为0.9~1.9 Pg·a-1,相当于当前大气二氧化碳含量的30%[1-2,7-8].因此,植被恢复过程中土壤碳固存潜力成为近年来提升土壤质量和应对气候变化研究的焦点[1-2,9-11]. ...
... 值得注意的是,SOC对土壤质量和肥力以及缓解气候变化贡献的有效性取决于其组成和稳定性,而大量研究表明,将SOC库视为均一的整体进行研究无法揭示相关问题[12].SOC通常由稳定性和周转速率不同的多个功能库组成[11-13],将SOC分为颗粒有机碳(Particulate organic carbon, POC, 粒径53 μm)和矿质结合有机碳(Mineral-associated organic carbon, MAOC, 粒径53 μm)成为近年来揭示其形成和稳定性的有效方法 [11-14].POC主要由植物来源的结构聚合物组成,能够通过生化抗性和团聚体保护在土壤中存在数年至数十年[10,15-16];而MAOC主要由微生物分解和POC解聚而产生的简单生物分子组成,通过物理化学机制吸附在矿物表面,可将其与生物和酶分离,从而在土壤中存在数百年至上千年[12,17].因而,MAOC是碳长期封存的关键指标,而POC可以预测SOC短期分解过程[18-19].因此,明确不同组分随植被恢复的变化及其影响因素对于准确评价SOC稳定性至关重要.然而,尽管近年来沙区植被恢复引发的SOC变化得到了广泛证实[2,9-11],但已有研究主要集中在SOC储量、周转过程随恢复过程的变化及其与环境因子的关系等方面[2,10,20-22],而对POC和MAOC分布及其对植被恢复响应的研究较为缺乏.因此,对SOC形成和稳定性及其机制的认识仍存在很大的不确定性[10,12],从而限制了对植被恢复的长期碳固存效应及其对全球碳循环贡献的准确评价[12-13]. ...
... [11-14].POC主要由植物来源的结构聚合物组成,能够通过生化抗性和团聚体保护在土壤中存在数年至数十年[10,15-16];而MAOC主要由微生物分解和POC解聚而产生的简单生物分子组成,通过物理化学机制吸附在矿物表面,可将其与生物和酶分离,从而在土壤中存在数百年至上千年[12,17].因而,MAOC是碳长期封存的关键指标,而POC可以预测SOC短期分解过程[18-19].因此,明确不同组分随植被恢复的变化及其影响因素对于准确评价SOC稳定性至关重要.然而,尽管近年来沙区植被恢复引发的SOC变化得到了广泛证实[2,9-11],但已有研究主要集中在SOC储量、周转过程随恢复过程的变化及其与环境因子的关系等方面[2,10,20-22],而对POC和MAOC分布及其对植被恢复响应的研究较为缺乏.因此,对SOC形成和稳定性及其机制的认识仍存在很大的不确定性[10,12],从而限制了对植被恢复的长期碳固存效应及其对全球碳循环贡献的准确评价[12-13]. ...
... -11],但已有研究主要集中在SOC储量、周转过程随恢复过程的变化及其与环境因子的关系等方面[2,10,20-22],而对POC和MAOC分布及其对植被恢复响应的研究较为缺乏.因此,对SOC形成和稳定性及其机制的认识仍存在很大的不确定性[10,12],从而限制了对植被恢复的长期碳固存效应及其对全球碳循环贡献的准确评价[12-13]. ...
... 植被恢复过程中生物和非生物因素的协同变化强烈影响着SOC及其组分的稳定性.大量研究表明,伴随着植被恢复,植被组成、土壤理化性质及微生物群落组成发生了显著变化[2,9,23-24].这可能会最终影响碳氮在不同组分中的分配,并进一步影响其稳定性[11,25].SOC不同组分在来源、形成过程及保护机制等方面的差异使其对植被恢复引发的生物和非生物因子变化的响应各不相同[20].C∶N较小的凋落物的输入、土壤pH值的减小、活性矿质颗粒较为丰富的土壤以及植物多样性的恢复可使植物来源的有机物有效地转化为微生物产物,从而促进MAOC的形成 [26];而不利于微生物活动的环境条件(如温度、水分和土壤类型等)通常会阻碍植物残体的分解,进而导致POC的积累[12].因此,植被恢复过程中SOC的变化可能是多种因素共同作用的结果,而明确不同组分的驱动因子是揭示SOC稳定性维持机制的前提[12-15]. ...
... 本研究中,固沙植被建立41 a后0~10 cm土层SOC含量比流沙增加9.59倍(表1,P0.01),表明种植旱生灌木是促进旱地SOC固存和土壤质量提升的有效措施.通常情况下,SOC的动态变化是由土壤有机质(Soil organic matter, SOM)输入和通过微生物分解的SOM输出过程共同决定的,当生态系统SOM输入大于输出时即表现为净SOC累积[2,24,31].一方面,固沙植被建立使植物来源SOM输入显著增大,其中地下部分生物量和凋落物分别解释了SOC变异的60%和90%(图3),而SOC与地上部分相关性不显著,这与Li等[2]的研究结果一致.这是因为随着植被演替,以灌木为优势的群落逐渐被以草本为优势的群落所取代,导致地上部分生物量呈减小趋势[11,24].另一方面,伴随着生态恢复,植被盖度显著增大(表1),沙丘表面逐渐趋于稳定,减少了风蚀引发的凋落物流失[2,32].且植物冠层能有效减小风速,增加了生态系统对大气降尘的捕获,而降尘中丰富的SOC和细颗粒物质提供了额外的碳源[2,33].此外,植被恢复促进了土壤理化性质的改善和土壤肥力的提升(表1),从而有利于植物生长以及SOM输入的增加[2,10].值得注意的是,固沙植被的建立促进了生物土壤结皮的拓殖和定居,腾格里沙漠东南缘的研究表明,生物土壤结皮的年固碳速率可达11.36~26.75 g·m-1[34],这也是表层土壤SOC的主要贡献者. ...
Soil carbon storage informed by particulate and mineral-associated organic matter
9
2019
... 值得注意的是,SOC对土壤质量和肥力以及缓解气候变化贡献的有效性取决于其组成和稳定性,而大量研究表明,将SOC库视为均一的整体进行研究无法揭示相关问题[12].SOC通常由稳定性和周转速率不同的多个功能库组成[11-13],将SOC分为颗粒有机碳(Particulate organic carbon, POC, 粒径53 μm)和矿质结合有机碳(Mineral-associated organic carbon, MAOC, 粒径53 μm)成为近年来揭示其形成和稳定性的有效方法 [11-14].POC主要由植物来源的结构聚合物组成,能够通过生化抗性和团聚体保护在土壤中存在数年至数十年[10,15-16];而MAOC主要由微生物分解和POC解聚而产生的简单生物分子组成,通过物理化学机制吸附在矿物表面,可将其与生物和酶分离,从而在土壤中存在数百年至上千年[12,17].因而,MAOC是碳长期封存的关键指标,而POC可以预测SOC短期分解过程[18-19].因此,明确不同组分随植被恢复的变化及其影响因素对于准确评价SOC稳定性至关重要.然而,尽管近年来沙区植被恢复引发的SOC变化得到了广泛证实[2,9-11],但已有研究主要集中在SOC储量、周转过程随恢复过程的变化及其与环境因子的关系等方面[2,10,20-22],而对POC和MAOC分布及其对植被恢复响应的研究较为缺乏.因此,对SOC形成和稳定性及其机制的认识仍存在很大的不确定性[10,12],从而限制了对植被恢复的长期碳固存效应及其对全球碳循环贡献的准确评价[12-13]. ...
... [12,17].因而,MAOC是碳长期封存的关键指标,而POC可以预测SOC短期分解过程[18-19].因此,明确不同组分随植被恢复的变化及其影响因素对于准确评价SOC稳定性至关重要.然而,尽管近年来沙区植被恢复引发的SOC变化得到了广泛证实[2,9-11],但已有研究主要集中在SOC储量、周转过程随恢复过程的变化及其与环境因子的关系等方面[2,10,20-22],而对POC和MAOC分布及其对植被恢复响应的研究较为缺乏.因此,对SOC形成和稳定性及其机制的认识仍存在很大的不确定性[10,12],从而限制了对植被恢复的长期碳固存效应及其对全球碳循环贡献的准确评价[12-13]. ...
... ,12],从而限制了对植被恢复的长期碳固存效应及其对全球碳循环贡献的准确评价[12-13]. ...
... [12-13]. ...
... 植被恢复过程中生物和非生物因素的协同变化强烈影响着SOC及其组分的稳定性.大量研究表明,伴随着植被恢复,植被组成、土壤理化性质及微生物群落组成发生了显著变化[2,9,23-24].这可能会最终影响碳氮在不同组分中的分配,并进一步影响其稳定性[11,25].SOC不同组分在来源、形成过程及保护机制等方面的差异使其对植被恢复引发的生物和非生物因子变化的响应各不相同[20].C∶N较小的凋落物的输入、土壤pH值的减小、活性矿质颗粒较为丰富的土壤以及植物多样性的恢复可使植物来源的有机物有效地转化为微生物产物,从而促进MAOC的形成 [26];而不利于微生物活动的环境条件(如温度、水分和土壤类型等)通常会阻碍植物残体的分解,进而导致POC的积累[12].因此,植被恢复过程中SOC的变化可能是多种因素共同作用的结果,而明确不同组分的驱动因子是揭示SOC稳定性维持机制的前提[12-15]. ...
... [12-15]. ...
... 已有研究表明,SOC的稳定性取决于其与矿物颗粒的相互作用和微生物的可及性,而MAOC与土壤矿物颗粒形成牢固的化学键,在提高SOC稳定性和长期封存方面发挥着至关重要的作用.因此,MAOC作为受微生物分解影响最小的碳库是SOC稳定性的重要表征[15].在本研究中,MAOC和POC含量及MAOC占SOC的比例随植被恢复年限的延长而显著增大,而POC占SOC的比例则呈相反趋势(图2,P0.01).这表明沙区植被恢复不仅促进了SOC的积累,同时使SOC组成由以POC为主逐渐向以MAOC为主转变,进而改变了SOC库的稳定性.POC主要来源于植物,由颗粒有机物的结构聚合物组成[14-17].因此,固沙植被区凋落物和地下部分生物量的显著增加是POC含量增大的主要原因,分别解释了其变异的89.3%和57.9%(图3).另一方面,固沙区土壤肥力的提升,特别是氮含量的显著增加不但促进SOM输入的增加和MBC的增大,而且加强了土壤团聚体保护[35],使POC含量进一步增加.已有研究表明,植物碳的微生物转化过程是SOC形成和稳定性维持的关键影响因素[36],植被恢复不仅增加了SOM的输入,而且增加土壤孔隙度和持水性,且使微生物基质(碳和养分)的数量和质量发生了显著变化(图1),土壤微环境更有利于与有机质分解相关的微生物群落的生长和繁殖[2,24];而EC的增大可通过增加微生物代谢熵而促进微生物对底物的利用,进一步促进微生物群落的发育[37],从而促进了植物残体的分解和POC和MAOC的累积[2,35],其中微生物群落变化解释了其变异的69%~85%(图3).另一方面,SOM的增加增强了土壤颗粒的胶结作用,有利于团聚体的形成,进一步促进了POC的增加[12-14].本研究中,POC占SOC的比例随植被恢复而显著减小,这主要是由于植被恢复引发的生物和非生物因子的改变,从而促进了MAOC截存,使得SOC库由以POC为主逐渐向以MAOC为主转变. ...
... 土壤黏粉粒是MAOC的关键影响因素,植被恢复使土壤黏粉粒含量显著增大(表1),这增加了矿物对SOM的可及性,从而促进MAOC的形成[14-15].本研究中,黏粉粒含量是与MAOC相关性最大的因子,解释了其变量的95.7%(图3).而土壤微生物对凋落物和POC的分解是形成MAOC的先决条件[15,19,38],因此,MAOC的变化与POC密切相关(图3).微生物将较粗的植物来源聚合物(POC)进行分解,形成更易溶解的化合物(微生物细胞和细胞外化合物),这些化合物与土壤矿物质结合形成MAOC[36],从而使固沙区土壤MAOC显著增加(图2).另一方面,细菌和真菌在土壤中丰度的增加,可分泌更多的黏液和多糖,进一步促进稳定性碳的形成[39].而真菌可通过其丝状菌丝体聚集矿物颗粒,促进了有机分子的吸附和固定从而有利于形成MAOC[40].放线菌多样性的显著增大可有效促进土壤腐殖质的积累和转化,这也是形成稳定有机碳的重要过程[41].值得注意的是,POC的分解过程中释放的养分和能量能完全满足微生物生长的需求,从而减少了因养分需求而对MAOC的分解,进而有利于MAOC的累积[10].伴随着植被恢复,碳分解酶BG、CBH的活性和MBC也显著增大(图1).两种酶不但可以促进植物残体分解,同时能将SOC转化为可溶性葡萄糖和其他低分子量碳化合物并为微生物提供能量和碳源,从而促进POC和MAOC的积累[42-43].而MBC的增大是MAOC形成的重要前提,可有效促进SOC与矿物质之间的相互作用,从而形成更多的MAOC[41].微生物残体是MAOC的关键组成部分,微生物多样性及生物量的增大可使微生物残体及代谢分泌物显著增加,从而有利于MAOC的累积[12-13].土壤氮含量的增加可通过土壤有机残体与氮的缩合反应、促进有机-无机复合物的形成以及丛枝菌根真菌(AMF)的丰度增大从而促进植物残体形成MAOC[35,44]. ...
... 通常情况下,60%以上的SOC储存在MAOC库中[15].而在本研究中,固沙植被建立41 a后,MAOC占表层土壤SOC的比例为36.47%,这一数值显著小于已有研究结果.一方面,沙区降水稀少,土壤含水量长期处于较低水平,因此微生物对植物残体的分解速率较低,从而更有利于POC的积累[15,17].本研究中,MAOC及其占SOC的比例随黏粉粒含量增大而显著增大,这正表明了大部分SOC处于POC库,且从POC向MAOC的转化速率处于较低水平[45].而MAOC与黏粉粒和SOC含量之间显著的线性关系表明固沙植被区土壤具有较大的MAOC封存潜力[45].另一方面,沙区土壤粗颗粒占优势的土壤类型,黏粉粒含量较小,因此,土壤矿物表面特征特别是矿物可及性低下,不利于MAOC的形成[12,14-15].真菌群落通常分解化学上难降解的SOC,而微生物群落中真菌相对于细菌的优势逐渐增大(F∶B增大)可促进MAOC的分解[46],因此真菌PLFA与MAOC呈负相关关系(图3和图5).此外,固沙区土壤pH显著减小,土壤酸化可能导致微生物多样性及生物量的增加减缓,从而降低了MAOC的增加速率[35].值得注意的是,尽管植被恢复有效地促进了SOC及其稳定性的增加,然而本研究区SOC、MAOC和POC含量仍处于较低水平,特别是MAOC占SOC比例的增加速率随植被恢复年限的延长和SOC含量的增加均呈减小趋势(图2和图4),表明沙区土壤碳储量和稳定性的恢复是一个长期过程.因此,水分限制生态系统修复是土壤生境的关键,而生态系统管理措施的优化是解决这一问题的核心. ...
Beyond bulk:density fractions explain heterogeneity in global soil carbon abundance and persistence
3
2022
... 值得注意的是,SOC对土壤质量和肥力以及缓解气候变化贡献的有效性取决于其组成和稳定性,而大量研究表明,将SOC库视为均一的整体进行研究无法揭示相关问题[12].SOC通常由稳定性和周转速率不同的多个功能库组成[11-13],将SOC分为颗粒有机碳(Particulate organic carbon, POC, 粒径53 μm)和矿质结合有机碳(Mineral-associated organic carbon, MAOC, 粒径53 μm)成为近年来揭示其形成和稳定性的有效方法 [11-14].POC主要由植物来源的结构聚合物组成,能够通过生化抗性和团聚体保护在土壤中存在数年至数十年[10,15-16];而MAOC主要由微生物分解和POC解聚而产生的简单生物分子组成,通过物理化学机制吸附在矿物表面,可将其与生物和酶分离,从而在土壤中存在数百年至上千年[12,17].因而,MAOC是碳长期封存的关键指标,而POC可以预测SOC短期分解过程[18-19].因此,明确不同组分随植被恢复的变化及其影响因素对于准确评价SOC稳定性至关重要.然而,尽管近年来沙区植被恢复引发的SOC变化得到了广泛证实[2,9-11],但已有研究主要集中在SOC储量、周转过程随恢复过程的变化及其与环境因子的关系等方面[2,10,20-22],而对POC和MAOC分布及其对植被恢复响应的研究较为缺乏.因此,对SOC形成和稳定性及其机制的认识仍存在很大的不确定性[10,12],从而限制了对植被恢复的长期碳固存效应及其对全球碳循环贡献的准确评价[12-13]. ...
... -13]. ...
... 土壤黏粉粒是MAOC的关键影响因素,植被恢复使土壤黏粉粒含量显著增大(表1),这增加了矿物对SOM的可及性,从而促进MAOC的形成[14-15].本研究中,黏粉粒含量是与MAOC相关性最大的因子,解释了其变量的95.7%(图3).而土壤微生物对凋落物和POC的分解是形成MAOC的先决条件[15,19,38],因此,MAOC的变化与POC密切相关(图3).微生物将较粗的植物来源聚合物(POC)进行分解,形成更易溶解的化合物(微生物细胞和细胞外化合物),这些化合物与土壤矿物质结合形成MAOC[36],从而使固沙区土壤MAOC显著增加(图2).另一方面,细菌和真菌在土壤中丰度的增加,可分泌更多的黏液和多糖,进一步促进稳定性碳的形成[39].而真菌可通过其丝状菌丝体聚集矿物颗粒,促进了有机分子的吸附和固定从而有利于形成MAOC[40].放线菌多样性的显著增大可有效促进土壤腐殖质的积累和转化,这也是形成稳定有机碳的重要过程[41].值得注意的是,POC的分解过程中释放的养分和能量能完全满足微生物生长的需求,从而减少了因养分需求而对MAOC的分解,进而有利于MAOC的累积[10].伴随着植被恢复,碳分解酶BG、CBH的活性和MBC也显著增大(图1).两种酶不但可以促进植物残体分解,同时能将SOC转化为可溶性葡萄糖和其他低分子量碳化合物并为微生物提供能量和碳源,从而促进POC和MAOC的积累[42-43].而MBC的增大是MAOC形成的重要前提,可有效促进SOC与矿物质之间的相互作用,从而形成更多的MAOC[41].微生物残体是MAOC的关键组成部分,微生物多样性及生物量的增大可使微生物残体及代谢分泌物显著增加,从而有利于MAOC的累积[12-13].土壤氮含量的增加可通过土壤有机残体与氮的缩合反应、促进有机-无机复合物的形成以及丛枝菌根真菌(AMF)的丰度增大从而促进植物残体形成MAOC[35,44]. ...
Global turnover of soil mineral-associated and particulate organic carbon
5
2024
... 值得注意的是,SOC对土壤质量和肥力以及缓解气候变化贡献的有效性取决于其组成和稳定性,而大量研究表明,将SOC库视为均一的整体进行研究无法揭示相关问题[12].SOC通常由稳定性和周转速率不同的多个功能库组成[11-13],将SOC分为颗粒有机碳(Particulate organic carbon, POC, 粒径53 μm)和矿质结合有机碳(Mineral-associated organic carbon, MAOC, 粒径53 μm)成为近年来揭示其形成和稳定性的有效方法 [11-14].POC主要由植物来源的结构聚合物组成,能够通过生化抗性和团聚体保护在土壤中存在数年至数十年[10,15-16];而MAOC主要由微生物分解和POC解聚而产生的简单生物分子组成,通过物理化学机制吸附在矿物表面,可将其与生物和酶分离,从而在土壤中存在数百年至上千年[12,17].因而,MAOC是碳长期封存的关键指标,而POC可以预测SOC短期分解过程[18-19].因此,明确不同组分随植被恢复的变化及其影响因素对于准确评价SOC稳定性至关重要.然而,尽管近年来沙区植被恢复引发的SOC变化得到了广泛证实[2,9-11],但已有研究主要集中在SOC储量、周转过程随恢复过程的变化及其与环境因子的关系等方面[2,10,20-22],而对POC和MAOC分布及其对植被恢复响应的研究较为缺乏.因此,对SOC形成和稳定性及其机制的认识仍存在很大的不确定性[10,12],从而限制了对植被恢复的长期碳固存效应及其对全球碳循环贡献的准确评价[12-13]. ...
... 已有研究表明,SOC的稳定性取决于其与矿物颗粒的相互作用和微生物的可及性,而MAOC与土壤矿物颗粒形成牢固的化学键,在提高SOC稳定性和长期封存方面发挥着至关重要的作用.因此,MAOC作为受微生物分解影响最小的碳库是SOC稳定性的重要表征[15].在本研究中,MAOC和POC含量及MAOC占SOC的比例随植被恢复年限的延长而显著增大,而POC占SOC的比例则呈相反趋势(图2,P0.01).这表明沙区植被恢复不仅促进了SOC的积累,同时使SOC组成由以POC为主逐渐向以MAOC为主转变,进而改变了SOC库的稳定性.POC主要来源于植物,由颗粒有机物的结构聚合物组成[14-17].因此,固沙植被区凋落物和地下部分生物量的显著增加是POC含量增大的主要原因,分别解释了其变异的89.3%和57.9%(图3).另一方面,固沙区土壤肥力的提升,特别是氮含量的显著增加不但促进SOM输入的增加和MBC的增大,而且加强了土壤团聚体保护[35],使POC含量进一步增加.已有研究表明,植物碳的微生物转化过程是SOC形成和稳定性维持的关键影响因素[36],植被恢复不仅增加了SOM的输入,而且增加土壤孔隙度和持水性,且使微生物基质(碳和养分)的数量和质量发生了显著变化(图1),土壤微环境更有利于与有机质分解相关的微生物群落的生长和繁殖[2,24];而EC的增大可通过增加微生物代谢熵而促进微生物对底物的利用,进一步促进微生物群落的发育[37],从而促进了植物残体的分解和POC和MAOC的累积[2,35],其中微生物群落变化解释了其变异的69%~85%(图3).另一方面,SOM的增加增强了土壤颗粒的胶结作用,有利于团聚体的形成,进一步促进了POC的增加[12-14].本研究中,POC占SOC的比例随植被恢复而显著减小,这主要是由于植被恢复引发的生物和非生物因子的改变,从而促进了MAOC截存,使得SOC库由以POC为主逐渐向以MAOC为主转变. ...
... -14].本研究中,POC占SOC的比例随植被恢复而显著减小,这主要是由于植被恢复引发的生物和非生物因子的改变,从而促进了MAOC截存,使得SOC库由以POC为主逐渐向以MAOC为主转变. ...
... 土壤黏粉粒是MAOC的关键影响因素,植被恢复使土壤黏粉粒含量显著增大(表1),这增加了矿物对SOM的可及性,从而促进MAOC的形成[14-15].本研究中,黏粉粒含量是与MAOC相关性最大的因子,解释了其变量的95.7%(图3).而土壤微生物对凋落物和POC的分解是形成MAOC的先决条件[15,19,38],因此,MAOC的变化与POC密切相关(图3).微生物将较粗的植物来源聚合物(POC)进行分解,形成更易溶解的化合物(微生物细胞和细胞外化合物),这些化合物与土壤矿物质结合形成MAOC[36],从而使固沙区土壤MAOC显著增加(图2).另一方面,细菌和真菌在土壤中丰度的增加,可分泌更多的黏液和多糖,进一步促进稳定性碳的形成[39].而真菌可通过其丝状菌丝体聚集矿物颗粒,促进了有机分子的吸附和固定从而有利于形成MAOC[40].放线菌多样性的显著增大可有效促进土壤腐殖质的积累和转化,这也是形成稳定有机碳的重要过程[41].值得注意的是,POC的分解过程中释放的养分和能量能完全满足微生物生长的需求,从而减少了因养分需求而对MAOC的分解,进而有利于MAOC的累积[10].伴随着植被恢复,碳分解酶BG、CBH的活性和MBC也显著增大(图1).两种酶不但可以促进植物残体分解,同时能将SOC转化为可溶性葡萄糖和其他低分子量碳化合物并为微生物提供能量和碳源,从而促进POC和MAOC的积累[42-43].而MBC的增大是MAOC形成的重要前提,可有效促进SOC与矿物质之间的相互作用,从而形成更多的MAOC[41].微生物残体是MAOC的关键组成部分,微生物多样性及生物量的增大可使微生物残体及代谢分泌物显著增加,从而有利于MAOC的累积[12-13].土壤氮含量的增加可通过土壤有机残体与氮的缩合反应、促进有机-无机复合物的形成以及丛枝菌根真菌(AMF)的丰度增大从而促进植物残体形成MAOC[35,44]. ...
... 通常情况下,60%以上的SOC储存在MAOC库中[15].而在本研究中,固沙植被建立41 a后,MAOC占表层土壤SOC的比例为36.47%,这一数值显著小于已有研究结果.一方面,沙区降水稀少,土壤含水量长期处于较低水平,因此微生物对植物残体的分解速率较低,从而更有利于POC的积累[15,17].本研究中,MAOC及其占SOC的比例随黏粉粒含量增大而显著增大,这正表明了大部分SOC处于POC库,且从POC向MAOC的转化速率处于较低水平[45].而MAOC与黏粉粒和SOC含量之间显著的线性关系表明固沙植被区土壤具有较大的MAOC封存潜力[45].另一方面,沙区土壤粗颗粒占优势的土壤类型,黏粉粒含量较小,因此,土壤矿物表面特征特别是矿物可及性低下,不利于MAOC的形成[12,14-15].真菌群落通常分解化学上难降解的SOC,而微生物群落中真菌相对于细菌的优势逐渐增大(F∶B增大)可促进MAOC的分解[46],因此真菌PLFA与MAOC呈负相关关系(图3和图5).此外,固沙区土壤pH显著减小,土壤酸化可能导致微生物多样性及生物量的增加减缓,从而降低了MAOC的增加速率[35].值得注意的是,尽管植被恢复有效地促进了SOC及其稳定性的增加,然而本研究区SOC、MAOC和POC含量仍处于较低水平,特别是MAOC占SOC比例的增加速率随植被恢复年限的延长和SOC含量的增加均呈减小趋势(图2和图4),表明沙区土壤碳储量和稳定性的恢复是一个长期过程.因此,水分限制生态系统修复是土壤生境的关键,而生态系统管理措施的优化是解决这一问题的核心. ...
Conceptualizing soil organic matter into particulate and mineral-associated forms to address global change in the 21st century
8
2020
... 值得注意的是,SOC对土壤质量和肥力以及缓解气候变化贡献的有效性取决于其组成和稳定性,而大量研究表明,将SOC库视为均一的整体进行研究无法揭示相关问题[12].SOC通常由稳定性和周转速率不同的多个功能库组成[11-13],将SOC分为颗粒有机碳(Particulate organic carbon, POC, 粒径53 μm)和矿质结合有机碳(Mineral-associated organic carbon, MAOC, 粒径53 μm)成为近年来揭示其形成和稳定性的有效方法 [11-14].POC主要由植物来源的结构聚合物组成,能够通过生化抗性和团聚体保护在土壤中存在数年至数十年[10,15-16];而MAOC主要由微生物分解和POC解聚而产生的简单生物分子组成,通过物理化学机制吸附在矿物表面,可将其与生物和酶分离,从而在土壤中存在数百年至上千年[12,17].因而,MAOC是碳长期封存的关键指标,而POC可以预测SOC短期分解过程[18-19].因此,明确不同组分随植被恢复的变化及其影响因素对于准确评价SOC稳定性至关重要.然而,尽管近年来沙区植被恢复引发的SOC变化得到了广泛证实[2,9-11],但已有研究主要集中在SOC储量、周转过程随恢复过程的变化及其与环境因子的关系等方面[2,10,20-22],而对POC和MAOC分布及其对植被恢复响应的研究较为缺乏.因此,对SOC形成和稳定性及其机制的认识仍存在很大的不确定性[10,12],从而限制了对植被恢复的长期碳固存效应及其对全球碳循环贡献的准确评价[12-13]. ...
... 植被恢复过程中生物和非生物因素的协同变化强烈影响着SOC及其组分的稳定性.大量研究表明,伴随着植被恢复,植被组成、土壤理化性质及微生物群落组成发生了显著变化[2,9,23-24].这可能会最终影响碳氮在不同组分中的分配,并进一步影响其稳定性[11,25].SOC不同组分在来源、形成过程及保护机制等方面的差异使其对植被恢复引发的生物和非生物因子变化的响应各不相同[20].C∶N较小的凋落物的输入、土壤pH值的减小、活性矿质颗粒较为丰富的土壤以及植物多样性的恢复可使植物来源的有机物有效地转化为微生物产物,从而促进MAOC的形成 [26];而不利于微生物活动的环境条件(如温度、水分和土壤类型等)通常会阻碍植物残体的分解,进而导致POC的积累[12].因此,植被恢复过程中SOC的变化可能是多种因素共同作用的结果,而明确不同组分的驱动因子是揭示SOC稳定性维持机制的前提[12-15]. ...
... 已有研究表明,SOC的稳定性取决于其与矿物颗粒的相互作用和微生物的可及性,而MAOC与土壤矿物颗粒形成牢固的化学键,在提高SOC稳定性和长期封存方面发挥着至关重要的作用.因此,MAOC作为受微生物分解影响最小的碳库是SOC稳定性的重要表征[15].在本研究中,MAOC和POC含量及MAOC占SOC的比例随植被恢复年限的延长而显著增大,而POC占SOC的比例则呈相反趋势(图2,P0.01).这表明沙区植被恢复不仅促进了SOC的积累,同时使SOC组成由以POC为主逐渐向以MAOC为主转变,进而改变了SOC库的稳定性.POC主要来源于植物,由颗粒有机物的结构聚合物组成[14-17].因此,固沙植被区凋落物和地下部分生物量的显著增加是POC含量增大的主要原因,分别解释了其变异的89.3%和57.9%(图3).另一方面,固沙区土壤肥力的提升,特别是氮含量的显著增加不但促进SOM输入的增加和MBC的增大,而且加强了土壤团聚体保护[35],使POC含量进一步增加.已有研究表明,植物碳的微生物转化过程是SOC形成和稳定性维持的关键影响因素[36],植被恢复不仅增加了SOM的输入,而且增加土壤孔隙度和持水性,且使微生物基质(碳和养分)的数量和质量发生了显著变化(图1),土壤微环境更有利于与有机质分解相关的微生物群落的生长和繁殖[2,24];而EC的增大可通过增加微生物代谢熵而促进微生物对底物的利用,进一步促进微生物群落的发育[37],从而促进了植物残体的分解和POC和MAOC的累积[2,35],其中微生物群落变化解释了其变异的69%~85%(图3).另一方面,SOM的增加增强了土壤颗粒的胶结作用,有利于团聚体的形成,进一步促进了POC的增加[12-14].本研究中,POC占SOC的比例随植被恢复而显著减小,这主要是由于植被恢复引发的生物和非生物因子的改变,从而促进了MAOC截存,使得SOC库由以POC为主逐渐向以MAOC为主转变. ...
... 土壤黏粉粒是MAOC的关键影响因素,植被恢复使土壤黏粉粒含量显著增大(表1),这增加了矿物对SOM的可及性,从而促进MAOC的形成[14-15].本研究中,黏粉粒含量是与MAOC相关性最大的因子,解释了其变量的95.7%(图3).而土壤微生物对凋落物和POC的分解是形成MAOC的先决条件[15,19,38],因此,MAOC的变化与POC密切相关(图3).微生物将较粗的植物来源聚合物(POC)进行分解,形成更易溶解的化合物(微生物细胞和细胞外化合物),这些化合物与土壤矿物质结合形成MAOC[36],从而使固沙区土壤MAOC显著增加(图2).另一方面,细菌和真菌在土壤中丰度的增加,可分泌更多的黏液和多糖,进一步促进稳定性碳的形成[39].而真菌可通过其丝状菌丝体聚集矿物颗粒,促进了有机分子的吸附和固定从而有利于形成MAOC[40].放线菌多样性的显著增大可有效促进土壤腐殖质的积累和转化,这也是形成稳定有机碳的重要过程[41].值得注意的是,POC的分解过程中释放的养分和能量能完全满足微生物生长的需求,从而减少了因养分需求而对MAOC的分解,进而有利于MAOC的累积[10].伴随着植被恢复,碳分解酶BG、CBH的活性和MBC也显著增大(图1).两种酶不但可以促进植物残体分解,同时能将SOC转化为可溶性葡萄糖和其他低分子量碳化合物并为微生物提供能量和碳源,从而促进POC和MAOC的积累[42-43].而MBC的增大是MAOC形成的重要前提,可有效促进SOC与矿物质之间的相互作用,从而形成更多的MAOC[41].微生物残体是MAOC的关键组成部分,微生物多样性及生物量的增大可使微生物残体及代谢分泌物显著增加,从而有利于MAOC的累积[12-13].土壤氮含量的增加可通过土壤有机残体与氮的缩合反应、促进有机-无机复合物的形成以及丛枝菌根真菌(AMF)的丰度增大从而促进植物残体形成MAOC[35,44]. ...
... [15,19,38],因此,MAOC的变化与POC密切相关(图3).微生物将较粗的植物来源聚合物(POC)进行分解,形成更易溶解的化合物(微生物细胞和细胞外化合物),这些化合物与土壤矿物质结合形成MAOC[36],从而使固沙区土壤MAOC显著增加(图2).另一方面,细菌和真菌在土壤中丰度的增加,可分泌更多的黏液和多糖,进一步促进稳定性碳的形成[39].而真菌可通过其丝状菌丝体聚集矿物颗粒,促进了有机分子的吸附和固定从而有利于形成MAOC[40].放线菌多样性的显著增大可有效促进土壤腐殖质的积累和转化,这也是形成稳定有机碳的重要过程[41].值得注意的是,POC的分解过程中释放的养分和能量能完全满足微生物生长的需求,从而减少了因养分需求而对MAOC的分解,进而有利于MAOC的累积[10].伴随着植被恢复,碳分解酶BG、CBH的活性和MBC也显著增大(图1).两种酶不但可以促进植物残体分解,同时能将SOC转化为可溶性葡萄糖和其他低分子量碳化合物并为微生物提供能量和碳源,从而促进POC和MAOC的积累[42-43].而MBC的增大是MAOC形成的重要前提,可有效促进SOC与矿物质之间的相互作用,从而形成更多的MAOC[41].微生物残体是MAOC的关键组成部分,微生物多样性及生物量的增大可使微生物残体及代谢分泌物显著增加,从而有利于MAOC的累积[12-13].土壤氮含量的增加可通过土壤有机残体与氮的缩合反应、促进有机-无机复合物的形成以及丛枝菌根真菌(AMF)的丰度增大从而促进植物残体形成MAOC[35,44]. ...
... 通常情况下,60%以上的SOC储存在MAOC库中[15].而在本研究中,固沙植被建立41 a后,MAOC占表层土壤SOC的比例为36.47%,这一数值显著小于已有研究结果.一方面,沙区降水稀少,土壤含水量长期处于较低水平,因此微生物对植物残体的分解速率较低,从而更有利于POC的积累[15,17].本研究中,MAOC及其占SOC的比例随黏粉粒含量增大而显著增大,这正表明了大部分SOC处于POC库,且从POC向MAOC的转化速率处于较低水平[45].而MAOC与黏粉粒和SOC含量之间显著的线性关系表明固沙植被区土壤具有较大的MAOC封存潜力[45].另一方面,沙区土壤粗颗粒占优势的土壤类型,黏粉粒含量较小,因此,土壤矿物表面特征特别是矿物可及性低下,不利于MAOC的形成[12,14-15].真菌群落通常分解化学上难降解的SOC,而微生物群落中真菌相对于细菌的优势逐渐增大(F∶B增大)可促进MAOC的分解[46],因此真菌PLFA与MAOC呈负相关关系(图3和图5).此外,固沙区土壤pH显著减小,土壤酸化可能导致微生物多样性及生物量的增加减缓,从而降低了MAOC的增加速率[35].值得注意的是,尽管植被恢复有效地促进了SOC及其稳定性的增加,然而本研究区SOC、MAOC和POC含量仍处于较低水平,特别是MAOC占SOC比例的增加速率随植被恢复年限的延长和SOC含量的增加均呈减小趋势(图2和图4),表明沙区土壤碳储量和稳定性的恢复是一个长期过程.因此,水分限制生态系统修复是土壤生境的关键,而生态系统管理措施的优化是解决这一问题的核心. ...
... [15,17].本研究中,MAOC及其占SOC的比例随黏粉粒含量增大而显著增大,这正表明了大部分SOC处于POC库,且从POC向MAOC的转化速率处于较低水平[45].而MAOC与黏粉粒和SOC含量之间显著的线性关系表明固沙植被区土壤具有较大的MAOC封存潜力[45].另一方面,沙区土壤粗颗粒占优势的土壤类型,黏粉粒含量较小,因此,土壤矿物表面特征特别是矿物可及性低下,不利于MAOC的形成[12,14-15].真菌群落通常分解化学上难降解的SOC,而微生物群落中真菌相对于细菌的优势逐渐增大(F∶B增大)可促进MAOC的分解[46],因此真菌PLFA与MAOC呈负相关关系(图3和图5).此外,固沙区土壤pH显著减小,土壤酸化可能导致微生物多样性及生物量的增加减缓,从而降低了MAOC的增加速率[35].值得注意的是,尽管植被恢复有效地促进了SOC及其稳定性的增加,然而本研究区SOC、MAOC和POC含量仍处于较低水平,特别是MAOC占SOC比例的增加速率随植被恢复年限的延长和SOC含量的增加均呈减小趋势(图2和图4),表明沙区土壤碳储量和稳定性的恢复是一个长期过程.因此,水分限制生态系统修复是土壤生境的关键,而生态系统管理措施的优化是解决这一问题的核心. ...
... -15].真菌群落通常分解化学上难降解的SOC,而微生物群落中真菌相对于细菌的优势逐渐增大(F∶B增大)可促进MAOC的分解[46],因此真菌PLFA与MAOC呈负相关关系(图3和图5).此外,固沙区土壤pH显著减小,土壤酸化可能导致微生物多样性及生物量的增加减缓,从而降低了MAOC的增加速率[35].值得注意的是,尽管植被恢复有效地促进了SOC及其稳定性的增加,然而本研究区SOC、MAOC和POC含量仍处于较低水平,特别是MAOC占SOC比例的增加速率随植被恢复年限的延长和SOC含量的增加均呈减小趋势(图2和图4),表明沙区土壤碳储量和稳定性的恢复是一个长期过程.因此,水分限制生态系统修复是土壤生境的关键,而生态系统管理措施的优化是解决这一问题的核心. ...
SOM fractionation methods:relevance to functional pools and to stabilization mechanisms
1
2007
... 值得注意的是,SOC对土壤质量和肥力以及缓解气候变化贡献的有效性取决于其组成和稳定性,而大量研究表明,将SOC库视为均一的整体进行研究无法揭示相关问题[12].SOC通常由稳定性和周转速率不同的多个功能库组成[11-13],将SOC分为颗粒有机碳(Particulate organic carbon, POC, 粒径53 μm)和矿质结合有机碳(Mineral-associated organic carbon, MAOC, 粒径53 μm)成为近年来揭示其形成和稳定性的有效方法 [11-14].POC主要由植物来源的结构聚合物组成,能够通过生化抗性和团聚体保护在土壤中存在数年至数十年[10,15-16];而MAOC主要由微生物分解和POC解聚而产生的简单生物分子组成,通过物理化学机制吸附在矿物表面,可将其与生物和酶分离,从而在土壤中存在数百年至上千年[12,17].因而,MAOC是碳长期封存的关键指标,而POC可以预测SOC短期分解过程[18-19].因此,明确不同组分随植被恢复的变化及其影响因素对于准确评价SOC稳定性至关重要.然而,尽管近年来沙区植被恢复引发的SOC变化得到了广泛证实[2,9-11],但已有研究主要集中在SOC储量、周转过程随恢复过程的变化及其与环境因子的关系等方面[2,10,20-22],而对POC和MAOC分布及其对植被恢复响应的研究较为缺乏.因此,对SOC形成和稳定性及其机制的认识仍存在很大的不确定性[10,12],从而限制了对植被恢复的长期碳固存效应及其对全球碳循环贡献的准确评价[12-13]. ...
Dynamic interactions at the mineral-organic matter interface
3
2021
... 值得注意的是,SOC对土壤质量和肥力以及缓解气候变化贡献的有效性取决于其组成和稳定性,而大量研究表明,将SOC库视为均一的整体进行研究无法揭示相关问题[12].SOC通常由稳定性和周转速率不同的多个功能库组成[11-13],将SOC分为颗粒有机碳(Particulate organic carbon, POC, 粒径53 μm)和矿质结合有机碳(Mineral-associated organic carbon, MAOC, 粒径53 μm)成为近年来揭示其形成和稳定性的有效方法 [11-14].POC主要由植物来源的结构聚合物组成,能够通过生化抗性和团聚体保护在土壤中存在数年至数十年[10,15-16];而MAOC主要由微生物分解和POC解聚而产生的简单生物分子组成,通过物理化学机制吸附在矿物表面,可将其与生物和酶分离,从而在土壤中存在数百年至上千年[12,17].因而,MAOC是碳长期封存的关键指标,而POC可以预测SOC短期分解过程[18-19].因此,明确不同组分随植被恢复的变化及其影响因素对于准确评价SOC稳定性至关重要.然而,尽管近年来沙区植被恢复引发的SOC变化得到了广泛证实[2,9-11],但已有研究主要集中在SOC储量、周转过程随恢复过程的变化及其与环境因子的关系等方面[2,10,20-22],而对POC和MAOC分布及其对植被恢复响应的研究较为缺乏.因此,对SOC形成和稳定性及其机制的认识仍存在很大的不确定性[10,12],从而限制了对植被恢复的长期碳固存效应及其对全球碳循环贡献的准确评价[12-13]. ...
... 已有研究表明,SOC的稳定性取决于其与矿物颗粒的相互作用和微生物的可及性,而MAOC与土壤矿物颗粒形成牢固的化学键,在提高SOC稳定性和长期封存方面发挥着至关重要的作用.因此,MAOC作为受微生物分解影响最小的碳库是SOC稳定性的重要表征[15].在本研究中,MAOC和POC含量及MAOC占SOC的比例随植被恢复年限的延长而显著增大,而POC占SOC的比例则呈相反趋势(图2,P0.01).这表明沙区植被恢复不仅促进了SOC的积累,同时使SOC组成由以POC为主逐渐向以MAOC为主转变,进而改变了SOC库的稳定性.POC主要来源于植物,由颗粒有机物的结构聚合物组成[14-17].因此,固沙植被区凋落物和地下部分生物量的显著增加是POC含量增大的主要原因,分别解释了其变异的89.3%和57.9%(图3).另一方面,固沙区土壤肥力的提升,特别是氮含量的显著增加不但促进SOM输入的增加和MBC的增大,而且加强了土壤团聚体保护[35],使POC含量进一步增加.已有研究表明,植物碳的微生物转化过程是SOC形成和稳定性维持的关键影响因素[36],植被恢复不仅增加了SOM的输入,而且增加土壤孔隙度和持水性,且使微生物基质(碳和养分)的数量和质量发生了显著变化(图1),土壤微环境更有利于与有机质分解相关的微生物群落的生长和繁殖[2,24];而EC的增大可通过增加微生物代谢熵而促进微生物对底物的利用,进一步促进微生物群落的发育[37],从而促进了植物残体的分解和POC和MAOC的累积[2,35],其中微生物群落变化解释了其变异的69%~85%(图3).另一方面,SOM的增加增强了土壤颗粒的胶结作用,有利于团聚体的形成,进一步促进了POC的增加[12-14].本研究中,POC占SOC的比例随植被恢复而显著减小,这主要是由于植被恢复引发的生物和非生物因子的改变,从而促进了MAOC截存,使得SOC库由以POC为主逐渐向以MAOC为主转变. ...
... 通常情况下,60%以上的SOC储存在MAOC库中[15].而在本研究中,固沙植被建立41 a后,MAOC占表层土壤SOC的比例为36.47%,这一数值显著小于已有研究结果.一方面,沙区降水稀少,土壤含水量长期处于较低水平,因此微生物对植物残体的分解速率较低,从而更有利于POC的积累[15,17].本研究中,MAOC及其占SOC的比例随黏粉粒含量增大而显著增大,这正表明了大部分SOC处于POC库,且从POC向MAOC的转化速率处于较低水平[45].而MAOC与黏粉粒和SOC含量之间显著的线性关系表明固沙植被区土壤具有较大的MAOC封存潜力[45].另一方面,沙区土壤粗颗粒占优势的土壤类型,黏粉粒含量较小,因此,土壤矿物表面特征特别是矿物可及性低下,不利于MAOC的形成[12,14-15].真菌群落通常分解化学上难降解的SOC,而微生物群落中真菌相对于细菌的优势逐渐增大(F∶B增大)可促进MAOC的分解[46],因此真菌PLFA与MAOC呈负相关关系(图3和图5).此外,固沙区土壤pH显著减小,土壤酸化可能导致微生物多样性及生物量的增加减缓,从而降低了MAOC的增加速率[35].值得注意的是,尽管植被恢复有效地促进了SOC及其稳定性的增加,然而本研究区SOC、MAOC和POC含量仍处于较低水平,特别是MAOC占SOC比例的增加速率随植被恢复年限的延长和SOC含量的增加均呈减小趋势(图2和图4),表明沙区土壤碳储量和稳定性的恢复是一个长期过程.因此,水分限制生态系统修复是土壤生境的关键,而生态系统管理措施的优化是解决这一问题的核心. ...
Soil organic matter pools and their associations with carbon mineralization kinetics
1
2000
... 值得注意的是,SOC对土壤质量和肥力以及缓解气候变化贡献的有效性取决于其组成和稳定性,而大量研究表明,将SOC库视为均一的整体进行研究无法揭示相关问题[12].SOC通常由稳定性和周转速率不同的多个功能库组成[11-13],将SOC分为颗粒有机碳(Particulate organic carbon, POC, 粒径53 μm)和矿质结合有机碳(Mineral-associated organic carbon, MAOC, 粒径53 μm)成为近年来揭示其形成和稳定性的有效方法 [11-14].POC主要由植物来源的结构聚合物组成,能够通过生化抗性和团聚体保护在土壤中存在数年至数十年[10,15-16];而MAOC主要由微生物分解和POC解聚而产生的简单生物分子组成,通过物理化学机制吸附在矿物表面,可将其与生物和酶分离,从而在土壤中存在数百年至上千年[12,17].因而,MAOC是碳长期封存的关键指标,而POC可以预测SOC短期分解过程[18-19].因此,明确不同组分随植被恢复的变化及其影响因素对于准确评价SOC稳定性至关重要.然而,尽管近年来沙区植被恢复引发的SOC变化得到了广泛证实[2,9-11],但已有研究主要集中在SOC储量、周转过程随恢复过程的变化及其与环境因子的关系等方面[2,10,20-22],而对POC和MAOC分布及其对植被恢复响应的研究较为缺乏.因此,对SOC形成和稳定性及其机制的认识仍存在很大的不确定性[10,12],从而限制了对植被恢复的长期碳固存效应及其对全球碳循环贡献的准确评价[12-13]. ...
Microbial formation of stable soil carbon is more efficient from belowground than aboveground input
2
2019
... 值得注意的是,SOC对土壤质量和肥力以及缓解气候变化贡献的有效性取决于其组成和稳定性,而大量研究表明,将SOC库视为均一的整体进行研究无法揭示相关问题[12].SOC通常由稳定性和周转速率不同的多个功能库组成[11-13],将SOC分为颗粒有机碳(Particulate organic carbon, POC, 粒径53 μm)和矿质结合有机碳(Mineral-associated organic carbon, MAOC, 粒径53 μm)成为近年来揭示其形成和稳定性的有效方法 [11-14].POC主要由植物来源的结构聚合物组成,能够通过生化抗性和团聚体保护在土壤中存在数年至数十年[10,15-16];而MAOC主要由微生物分解和POC解聚而产生的简单生物分子组成,通过物理化学机制吸附在矿物表面,可将其与生物和酶分离,从而在土壤中存在数百年至上千年[12,17].因而,MAOC是碳长期封存的关键指标,而POC可以预测SOC短期分解过程[18-19].因此,明确不同组分随植被恢复的变化及其影响因素对于准确评价SOC稳定性至关重要.然而,尽管近年来沙区植被恢复引发的SOC变化得到了广泛证实[2,9-11],但已有研究主要集中在SOC储量、周转过程随恢复过程的变化及其与环境因子的关系等方面[2,10,20-22],而对POC和MAOC分布及其对植被恢复响应的研究较为缺乏.因此,对SOC形成和稳定性及其机制的认识仍存在很大的不确定性[10,12],从而限制了对植被恢复的长期碳固存效应及其对全球碳循环贡献的准确评价[12-13]. ...
... 土壤黏粉粒是MAOC的关键影响因素,植被恢复使土壤黏粉粒含量显著增大(表1),这增加了矿物对SOM的可及性,从而促进MAOC的形成[14-15].本研究中,黏粉粒含量是与MAOC相关性最大的因子,解释了其变量的95.7%(图3).而土壤微生物对凋落物和POC的分解是形成MAOC的先决条件[15,19,38],因此,MAOC的变化与POC密切相关(图3).微生物将较粗的植物来源聚合物(POC)进行分解,形成更易溶解的化合物(微生物细胞和细胞外化合物),这些化合物与土壤矿物质结合形成MAOC[36],从而使固沙区土壤MAOC显著增加(图2).另一方面,细菌和真菌在土壤中丰度的增加,可分泌更多的黏液和多糖,进一步促进稳定性碳的形成[39].而真菌可通过其丝状菌丝体聚集矿物颗粒,促进了有机分子的吸附和固定从而有利于形成MAOC[40].放线菌多样性的显著增大可有效促进土壤腐殖质的积累和转化,这也是形成稳定有机碳的重要过程[41].值得注意的是,POC的分解过程中释放的养分和能量能完全满足微生物生长的需求,从而减少了因养分需求而对MAOC的分解,进而有利于MAOC的累积[10].伴随着植被恢复,碳分解酶BG、CBH的活性和MBC也显著增大(图1).两种酶不但可以促进植物残体分解,同时能将SOC转化为可溶性葡萄糖和其他低分子量碳化合物并为微生物提供能量和碳源,从而促进POC和MAOC的积累[42-43].而MBC的增大是MAOC形成的重要前提,可有效促进SOC与矿物质之间的相互作用,从而形成更多的MAOC[41].微生物残体是MAOC的关键组成部分,微生物多样性及生物量的增大可使微生物残体及代谢分泌物显著增加,从而有利于MAOC的累积[12-13].土壤氮含量的增加可通过土壤有机残体与氮的缩合反应、促进有机-无机复合物的形成以及丛枝菌根真菌(AMF)的丰度增大从而促进植物残体形成MAOC[35,44]. ...
退化和恢复过程驱动的荒漠草地生态系统有机碳密度变化
2
2022
... 值得注意的是,SOC对土壤质量和肥力以及缓解气候变化贡献的有效性取决于其组成和稳定性,而大量研究表明,将SOC库视为均一的整体进行研究无法揭示相关问题[12].SOC通常由稳定性和周转速率不同的多个功能库组成[11-13],将SOC分为颗粒有机碳(Particulate organic carbon, POC, 粒径53 μm)和矿质结合有机碳(Mineral-associated organic carbon, MAOC, 粒径53 μm)成为近年来揭示其形成和稳定性的有效方法 [11-14].POC主要由植物来源的结构聚合物组成,能够通过生化抗性和团聚体保护在土壤中存在数年至数十年[10,15-16];而MAOC主要由微生物分解和POC解聚而产生的简单生物分子组成,通过物理化学机制吸附在矿物表面,可将其与生物和酶分离,从而在土壤中存在数百年至上千年[12,17].因而,MAOC是碳长期封存的关键指标,而POC可以预测SOC短期分解过程[18-19].因此,明确不同组分随植被恢复的变化及其影响因素对于准确评价SOC稳定性至关重要.然而,尽管近年来沙区植被恢复引发的SOC变化得到了广泛证实[2,9-11],但已有研究主要集中在SOC储量、周转过程随恢复过程的变化及其与环境因子的关系等方面[2,10,20-22],而对POC和MAOC分布及其对植被恢复响应的研究较为缺乏.因此,对SOC形成和稳定性及其机制的认识仍存在很大的不确定性[10,12],从而限制了对植被恢复的长期碳固存效应及其对全球碳循环贡献的准确评价[12-13]. ...
... 植被恢复过程中生物和非生物因素的协同变化强烈影响着SOC及其组分的稳定性.大量研究表明,伴随着植被恢复,植被组成、土壤理化性质及微生物群落组成发生了显著变化[2,9,23-24].这可能会最终影响碳氮在不同组分中的分配,并进一步影响其稳定性[11,25].SOC不同组分在来源、形成过程及保护机制等方面的差异使其对植被恢复引发的生物和非生物因子变化的响应各不相同[20].C∶N较小的凋落物的输入、土壤pH值的减小、活性矿质颗粒较为丰富的土壤以及植物多样性的恢复可使植物来源的有机物有效地转化为微生物产物,从而促进MAOC的形成 [26];而不利于微生物活动的环境条件(如温度、水分和土壤类型等)通常会阻碍植物残体的分解,进而导致POC的积累[12].因此,植被恢复过程中SOC的变化可能是多种因素共同作用的结果,而明确不同组分的驱动因子是揭示SOC稳定性维持机制的前提[12-15]. ...
中国北方干旱半干旱区沙漠化治理对植被格局的影响
0
2023
Effects of anthropogenic revegetation on the water and carbon cycles of a desert steppe ecosystem
1
2021
... 值得注意的是,SOC对土壤质量和肥力以及缓解气候变化贡献的有效性取决于其组成和稳定性,而大量研究表明,将SOC库视为均一的整体进行研究无法揭示相关问题[12].SOC通常由稳定性和周转速率不同的多个功能库组成[11-13],将SOC分为颗粒有机碳(Particulate organic carbon, POC, 粒径53 μm)和矿质结合有机碳(Mineral-associated organic carbon, MAOC, 粒径53 μm)成为近年来揭示其形成和稳定性的有效方法 [11-14].POC主要由植物来源的结构聚合物组成,能够通过生化抗性和团聚体保护在土壤中存在数年至数十年[10,15-16];而MAOC主要由微生物分解和POC解聚而产生的简单生物分子组成,通过物理化学机制吸附在矿物表面,可将其与生物和酶分离,从而在土壤中存在数百年至上千年[12,17].因而,MAOC是碳长期封存的关键指标,而POC可以预测SOC短期分解过程[18-19].因此,明确不同组分随植被恢复的变化及其影响因素对于准确评价SOC稳定性至关重要.然而,尽管近年来沙区植被恢复引发的SOC变化得到了广泛证实[2,9-11],但已有研究主要集中在SOC储量、周转过程随恢复过程的变化及其与环境因子的关系等方面[2,10,20-22],而对POC和MAOC分布及其对植被恢复响应的研究较为缺乏.因此,对SOC形成和稳定性及其机制的认识仍存在很大的不确定性[10,12],从而限制了对植被恢复的长期碳固存效应及其对全球碳循环贡献的准确评价[12-13]. ...
腾格里沙漠东南缘植被恢复过程中土壤微生物量及酶活性
1
2019
... 植被恢复过程中生物和非生物因素的协同变化强烈影响着SOC及其组分的稳定性.大量研究表明,伴随着植被恢复,植被组成、土壤理化性质及微生物群落组成发生了显著变化[2,9,23-24].这可能会最终影响碳氮在不同组分中的分配,并进一步影响其稳定性[11,25].SOC不同组分在来源、形成过程及保护机制等方面的差异使其对植被恢复引发的生物和非生物因子变化的响应各不相同[20].C∶N较小的凋落物的输入、土壤pH值的减小、活性矿质颗粒较为丰富的土壤以及植物多样性的恢复可使植物来源的有机物有效地转化为微生物产物,从而促进MAOC的形成 [26];而不利于微生物活动的环境条件(如温度、水分和土壤类型等)通常会阻碍植物残体的分解,进而导致POC的积累[12].因此,植被恢复过程中SOC的变化可能是多种因素共同作用的结果,而明确不同组分的驱动因子是揭示SOC稳定性维持机制的前提[12-15]. ...
Carbon sequestration of sand-fixing plantation of Haloxylon ammodendron in Shiyang River Basin:storage,rate and potential
5
2021
... 植被恢复过程中生物和非生物因素的协同变化强烈影响着SOC及其组分的稳定性.大量研究表明,伴随着植被恢复,植被组成、土壤理化性质及微生物群落组成发生了显著变化[2,9,23-24].这可能会最终影响碳氮在不同组分中的分配,并进一步影响其稳定性[11,25].SOC不同组分在来源、形成过程及保护机制等方面的差异使其对植被恢复引发的生物和非生物因子变化的响应各不相同[20].C∶N较小的凋落物的输入、土壤pH值的减小、活性矿质颗粒较为丰富的土壤以及植物多样性的恢复可使植物来源的有机物有效地转化为微生物产物,从而促进MAOC的形成 [26];而不利于微生物活动的环境条件(如温度、水分和土壤类型等)通常会阻碍植物残体的分解,进而导致POC的积累[12].因此,植被恢复过程中SOC的变化可能是多种因素共同作用的结果,而明确不同组分的驱动因子是揭示SOC稳定性维持机制的前提[12-15]. ...
... 研究区位于古浪县北部沙区的鸣沙咀(37°32′— 37°35′N,103°38′—103°43′E,海拔1 730~1 810 m).该区属温带干旱大陆性气候,多年平均气温6 ℃,极端最低和最高气温分别为-27.3 ℃和36.6 ℃.多年平均降水量为175 mm,主要集中在7—9月;多年平均蒸发量3 038.5 mm;地下水位40~80 m.土壤类型为风沙土,植被由旱生灌木白刺(Nitraria tangutorum)、沙蒿(Artemisia aphaerocephala)、油蒿(Artemisia ordosica)和中亚紫菀木(Asterothamnus centralis-asiaticus),多年生草本短花针茅(Stipa breviflora)和沙竹(Psammochloa villosa)等,以及一年生草本画眉草(Eragrostis pilosa)、虫实(Corispermum declinatum)、雾滨藜(Grubovia dasyphylla)和砂蓝刺头(Echinops gmelini)等组成,盖度低且分布不均[24,27]. ...
... 本研究中,固沙植被建立41 a后0~10 cm土层SOC含量比流沙增加9.59倍(表1,P0.01),表明种植旱生灌木是促进旱地SOC固存和土壤质量提升的有效措施.通常情况下,SOC的动态变化是由土壤有机质(Soil organic matter, SOM)输入和通过微生物分解的SOM输出过程共同决定的,当生态系统SOM输入大于输出时即表现为净SOC累积[2,24,31].一方面,固沙植被建立使植物来源SOM输入显著增大,其中地下部分生物量和凋落物分别解释了SOC变异的60%和90%(图3),而SOC与地上部分相关性不显著,这与Li等[2]的研究结果一致.这是因为随着植被演替,以灌木为优势的群落逐渐被以草本为优势的群落所取代,导致地上部分生物量呈减小趋势[11,24].另一方面,伴随着生态恢复,植被盖度显著增大(表1),沙丘表面逐渐趋于稳定,减少了风蚀引发的凋落物流失[2,32].且植物冠层能有效减小风速,增加了生态系统对大气降尘的捕获,而降尘中丰富的SOC和细颗粒物质提供了额外的碳源[2,33].此外,植被恢复促进了土壤理化性质的改善和土壤肥力的提升(表1),从而有利于植物生长以及SOM输入的增加[2,10].值得注意的是,固沙植被的建立促进了生物土壤结皮的拓殖和定居,腾格里沙漠东南缘的研究表明,生物土壤结皮的年固碳速率可达11.36~26.75 g·m-1[34],这也是表层土壤SOC的主要贡献者. ...
... ,24].另一方面,伴随着生态恢复,植被盖度显著增大(表1),沙丘表面逐渐趋于稳定,减少了风蚀引发的凋落物流失[2,32].且植物冠层能有效减小风速,增加了生态系统对大气降尘的捕获,而降尘中丰富的SOC和细颗粒物质提供了额外的碳源[2,33].此外,植被恢复促进了土壤理化性质的改善和土壤肥力的提升(表1),从而有利于植物生长以及SOM输入的增加[2,10].值得注意的是,固沙植被的建立促进了生物土壤结皮的拓殖和定居,腾格里沙漠东南缘的研究表明,生物土壤结皮的年固碳速率可达11.36~26.75 g·m-1[34],这也是表层土壤SOC的主要贡献者. ...
... 已有研究表明,SOC的稳定性取决于其与矿物颗粒的相互作用和微生物的可及性,而MAOC与土壤矿物颗粒形成牢固的化学键,在提高SOC稳定性和长期封存方面发挥着至关重要的作用.因此,MAOC作为受微生物分解影响最小的碳库是SOC稳定性的重要表征[15].在本研究中,MAOC和POC含量及MAOC占SOC的比例随植被恢复年限的延长而显著增大,而POC占SOC的比例则呈相反趋势(图2,P0.01).这表明沙区植被恢复不仅促进了SOC的积累,同时使SOC组成由以POC为主逐渐向以MAOC为主转变,进而改变了SOC库的稳定性.POC主要来源于植物,由颗粒有机物的结构聚合物组成[14-17].因此,固沙植被区凋落物和地下部分生物量的显著增加是POC含量增大的主要原因,分别解释了其变异的89.3%和57.9%(图3).另一方面,固沙区土壤肥力的提升,特别是氮含量的显著增加不但促进SOM输入的增加和MBC的增大,而且加强了土壤团聚体保护[35],使POC含量进一步增加.已有研究表明,植物碳的微生物转化过程是SOC形成和稳定性维持的关键影响因素[36],植被恢复不仅增加了SOM的输入,而且增加土壤孔隙度和持水性,且使微生物基质(碳和养分)的数量和质量发生了显著变化(图1),土壤微环境更有利于与有机质分解相关的微生物群落的生长和繁殖[2,24];而EC的增大可通过增加微生物代谢熵而促进微生物对底物的利用,进一步促进微生物群落的发育[37],从而促进了植物残体的分解和POC和MAOC的累积[2,35],其中微生物群落变化解释了其变异的69%~85%(图3).另一方面,SOM的增加增强了土壤颗粒的胶结作用,有利于团聚体的形成,进一步促进了POC的增加[12-14].本研究中,POC占SOC的比例随植被恢复而显著减小,这主要是由于植被恢复引发的生物和非生物因子的改变,从而促进了MAOC截存,使得SOC库由以POC为主逐渐向以MAOC为主转变. ...
Controls on labile and stabilized soil organic matter during long-term ecosystem development
1
2022
... 植被恢复过程中生物和非生物因素的协同变化强烈影响着SOC及其组分的稳定性.大量研究表明,伴随着植被恢复,植被组成、土壤理化性质及微生物群落组成发生了显著变化[2,9,23-24].这可能会最终影响碳氮在不同组分中的分配,并进一步影响其稳定性[11,25].SOC不同组分在来源、形成过程及保护机制等方面的差异使其对植被恢复引发的生物和非生物因子变化的响应各不相同[20].C∶N较小的凋落物的输入、土壤pH值的减小、活性矿质颗粒较为丰富的土壤以及植物多样性的恢复可使植物来源的有机物有效地转化为微生物产物,从而促进MAOC的形成 [26];而不利于微生物活动的环境条件(如温度、水分和土壤类型等)通常会阻碍植物残体的分解,进而导致POC的积累[12].因此,植被恢复过程中SOC的变化可能是多种因素共同作用的结果,而明确不同组分的驱动因子是揭示SOC稳定性维持机制的前提[12-15]. ...
Unlocking complex soil systems as carbon sinks:multi-pool management as the key
1
2023
... 植被恢复过程中生物和非生物因素的协同变化强烈影响着SOC及其组分的稳定性.大量研究表明,伴随着植被恢复,植被组成、土壤理化性质及微生物群落组成发生了显著变化[2,9,23-24].这可能会最终影响碳氮在不同组分中的分配,并进一步影响其稳定性[11,25].SOC不同组分在来源、形成过程及保护机制等方面的差异使其对植被恢复引发的生物和非生物因子变化的响应各不相同[20].C∶N较小的凋落物的输入、土壤pH值的减小、活性矿质颗粒较为丰富的土壤以及植物多样性的恢复可使植物来源的有机物有效地转化为微生物产物,从而促进MAOC的形成 [26];而不利于微生物活动的环境条件(如温度、水分和土壤类型等)通常会阻碍植物残体的分解,进而导致POC的积累[12].因此,植被恢复过程中SOC的变化可能是多种因素共同作用的结果,而明确不同组分的驱动因子是揭示SOC稳定性维持机制的前提[12-15]. ...
腾格里沙漠南缘土壤-植被系统恢复及其驱动机制
3
2009
... 自20世纪50年代开始,种植旱生灌木成为中国北方沙区实现荒漠化逆转的有效途径[2,27].特别是为了保护公路、铁路和水利设施等大型基础设施的安全,分不同阶段在西北干旱沙区种植了大量旱生灌木,形成了建植于不同年限的人工植被区,为研究植被恢复对SOC及其组分稳定性的影响提供了天然的研究平台.本文以甘肃省古浪县鸣沙咀不同年代人工植被区为研究对象,采用空间代替时间的方法,基于土壤有机碳组成和环境要素分析,拟回答以下问题:①SOC及其组分随植被恢复年限的变化规律;②植被恢复对SOC及其组成的影响及其机理.在此基础上,揭示沙区植被恢复对SOC组成及稳定性的影响及其机理. ...
... 研究区位于古浪县北部沙区的鸣沙咀(37°32′— 37°35′N,103°38′—103°43′E,海拔1 730~1 810 m).该区属温带干旱大陆性气候,多年平均气温6 ℃,极端最低和最高气温分别为-27.3 ℃和36.6 ℃.多年平均降水量为175 mm,主要集中在7—9月;多年平均蒸发量3 038.5 mm;地下水位40~80 m.土壤类型为风沙土,植被由旱生灌木白刺(Nitraria tangutorum)、沙蒿(Artemisia aphaerocephala)、油蒿(Artemisia ordosica)和中亚紫菀木(Asterothamnus centralis-asiaticus),多年生草本短花针茅(Stipa breviflora)和沙竹(Psammochloa villosa)等,以及一年生草本画眉草(Eragrostis pilosa)、虫实(Corispermum declinatum)、雾滨藜(Grubovia dasyphylla)和砂蓝刺头(Echinops gmelini)等组成,盖度低且分布不均[24,27]. ...
... 2023年7月,分别选择在1982年(41 a,恢复年限,下同)、1991年(32 a)、2003年(20 a)和2011年(12 a)建立的人工植被区及流动沙丘(对照,0 a)作为研究样地.在每个样地中设置10 m×10 m的样方各3个(间距大于50 m),用于灌木植物调查.由于人工固沙植被区地形起伏明显且不同地形生物和非生物因子差异显著,因此,各年代植被区的样方选择设置于地势相对平坦的丘间低地,以减少地形造成的数据波动,从而提高数据的准确性和可比性.在每个10 m×10 m的样方中随机布设3个1 m×1 m小样方,用于草本植物调查.植物群落调查包括物种组成、植被盖度和地上及地下部分生物量.灌木和草本地上部分生物量采用收割法进行测定,在每个10 m×10 m的样方中,每种灌木选择1个标准株并采集其地上部分,装入信封;在每个1 m×1 m的小样方中,将草本地上部分齐地面收割,装入信封;随后收集小样方地面凋落物,装入信封.根系生物量用根钻进行采集,在每个10 m×10 m样方中的灌丛间隙随机选取10个点,用直径为5 cm的根钻钻取0~10 cm层土壤,将其过2 mm筛以分离出根系,将根系装入信封.根据已有研究,沙区植被恢复对土壤的影响集中在表层0~10 cm土层,草本植物根系的主要分布范围也在该土层[2,10,27],因此,根系生物量的采集点均设在草本植物分布的灌丛间斑块中.在每个10 m×10 m的样方中,用直径为5 cm的土钻采集0~10 cm层土壤,在每个样方中随机选取10个点进行采样,将同一样方中10个点的样品充分混合形成一个混合样,每个年代植被区获得3个混合样品(3个重复).为避免灌丛斑块下“沃岛效应”造成的数据波动,土壤样品的采集点均设在草本植物分布的灌丛间斑块.将所有土壤样品带回实验室并分为两份,一份在-20 ℃条件下保存,用于土壤微生物群落、酶活性和微生物量碳(Microbial Biomass Carbon,MBC)的测定,另一份室温条件下风干后过2 mm筛,用于土壤理化性质及POC和MAOC的测定.将生物量和凋落物样本带回实验室,在60 ℃条件下烘干,称重. ...
Shifts in the structure of soil microbial communities in limed forests as revealed by phospholipid fatty acid analysis
1
1993
... SOC含量用总有机碳分析仪(Vario EL III, Elementar, DE)进行分析,全氮含量采用半微量凯氏定氮法进行测定.pH值采用5∶1水土比浸提后用酸度计测定,电导率(EC)采用2.5∶1水土比浸提后用便携式电导仪测定.土壤微生物群落组成用磷脂脂肪酸(Phospholipid fatty acid, PLFA)法进行分析[28],β-1, 4-葡萄糖苷酶(BG)和纤维二糖水解酶(CBH)活性采用微孔板荧光法进行测定[29],MBC采用氯仿熏蒸-萃取法进行测定[30]. ...
The effects of long term nitrogen deposition on extracellular enzyme activity in an Acer saccharum forest soil
1
2002
... SOC含量用总有机碳分析仪(Vario EL III, Elementar, DE)进行分析,全氮含量采用半微量凯氏定氮法进行测定.pH值采用5∶1水土比浸提后用酸度计测定,电导率(EC)采用2.5∶1水土比浸提后用便携式电导仪测定.土壤微生物群落组成用磷脂脂肪酸(Phospholipid fatty acid, PLFA)法进行分析[28],β-1, 4-葡萄糖苷酶(BG)和纤维二糖水解酶(CBH)活性采用微孔板荧光法进行测定[29],MBC采用氯仿熏蒸-萃取法进行测定[30]. ...
An extraction method for measuring soil microbial biomass C
1
1987
... SOC含量用总有机碳分析仪(Vario EL III, Elementar, DE)进行分析,全氮含量采用半微量凯氏定氮法进行测定.pH值采用5∶1水土比浸提后用酸度计测定,电导率(EC)采用2.5∶1水土比浸提后用便携式电导仪测定.土壤微生物群落组成用磷脂脂肪酸(Phospholipid fatty acid, PLFA)法进行分析[28],β-1, 4-葡萄糖苷酶(BG)和纤维二糖水解酶(CBH)活性采用微孔板荧光法进行测定[29],MBC采用氯仿熏蒸-萃取法进行测定[30]. ...
Carbon sequestration in semi-arid rangelands:comparison of Pinus ponderosa plantations and grazing exclusion in NW Patagonia
1
2006
... 本研究中,固沙植被建立41 a后0~10 cm土层SOC含量比流沙增加9.59倍(表1,P0.01),表明种植旱生灌木是促进旱地SOC固存和土壤质量提升的有效措施.通常情况下,SOC的动态变化是由土壤有机质(Soil organic matter, SOM)输入和通过微生物分解的SOM输出过程共同决定的,当生态系统SOM输入大于输出时即表现为净SOC累积[2,24,31].一方面,固沙植被建立使植物来源SOM输入显著增大,其中地下部分生物量和凋落物分别解释了SOC变异的60%和90%(图3),而SOC与地上部分相关性不显著,这与Li等[2]的研究结果一致.这是因为随着植被演替,以灌木为优势的群落逐渐被以草本为优势的群落所取代,导致地上部分生物量呈减小趋势[11,24].另一方面,伴随着生态恢复,植被盖度显著增大(表1),沙丘表面逐渐趋于稳定,减少了风蚀引发的凋落物流失[2,32].且植物冠层能有效减小风速,增加了生态系统对大气降尘的捕获,而降尘中丰富的SOC和细颗粒物质提供了额外的碳源[2,33].此外,植被恢复促进了土壤理化性质的改善和土壤肥力的提升(表1),从而有利于植物生长以及SOM输入的增加[2,10].值得注意的是,固沙植被的建立促进了生物土壤结皮的拓殖和定居,腾格里沙漠东南缘的研究表明,生物土壤结皮的年固碳速率可达11.36~26.75 g·m-1[34],这也是表层土壤SOC的主要贡献者. ...
Recovery of topsoil physicochemical properties in revegetated sites in the sand-burial ecosystems of the Tengger Desert,Northern China
1
2007
... 本研究中,固沙植被建立41 a后0~10 cm土层SOC含量比流沙增加9.59倍(表1,P0.01),表明种植旱生灌木是促进旱地SOC固存和土壤质量提升的有效措施.通常情况下,SOC的动态变化是由土壤有机质(Soil organic matter, SOM)输入和通过微生物分解的SOM输出过程共同决定的,当生态系统SOM输入大于输出时即表现为净SOC累积[2,24,31].一方面,固沙植被建立使植物来源SOM输入显著增大,其中地下部分生物量和凋落物分别解释了SOC变异的60%和90%(图3),而SOC与地上部分相关性不显著,这与Li等[2]的研究结果一致.这是因为随着植被演替,以灌木为优势的群落逐渐被以草本为优势的群落所取代,导致地上部分生物量呈减小趋势[11,24].另一方面,伴随着生态恢复,植被盖度显著增大(表1),沙丘表面逐渐趋于稳定,减少了风蚀引发的凋落物流失[2,32].且植物冠层能有效减小风速,增加了生态系统对大气降尘的捕获,而降尘中丰富的SOC和细颗粒物质提供了额外的碳源[2,33].此外,植被恢复促进了土壤理化性质的改善和土壤肥力的提升(表1),从而有利于植物生长以及SOM输入的增加[2,10].值得注意的是,固沙植被的建立促进了生物土壤结皮的拓殖和定居,腾格里沙漠东南缘的研究表明,生物土壤结皮的年固碳速率可达11.36~26.75 g·m-1[34],这也是表层土壤SOC的主要贡献者. ...
策勒绿洲近地表大气降尘粒度指示的分选特征及其意义
1
2022
... 本研究中,固沙植被建立41 a后0~10 cm土层SOC含量比流沙增加9.59倍(表1,P0.01),表明种植旱生灌木是促进旱地SOC固存和土壤质量提升的有效措施.通常情况下,SOC的动态变化是由土壤有机质(Soil organic matter, SOM)输入和通过微生物分解的SOM输出过程共同决定的,当生态系统SOM输入大于输出时即表现为净SOC累积[2,24,31].一方面,固沙植被建立使植物来源SOM输入显著增大,其中地下部分生物量和凋落物分别解释了SOC变异的60%和90%(图3),而SOC与地上部分相关性不显著,这与Li等[2]的研究结果一致.这是因为随着植被演替,以灌木为优势的群落逐渐被以草本为优势的群落所取代,导致地上部分生物量呈减小趋势[11,24].另一方面,伴随着生态恢复,植被盖度显著增大(表1),沙丘表面逐渐趋于稳定,减少了风蚀引发的凋落物流失[2,32].且植物冠层能有效减小风速,增加了生态系统对大气降尘的捕获,而降尘中丰富的SOC和细颗粒物质提供了额外的碳源[2,33].此外,植被恢复促进了土壤理化性质的改善和土壤肥力的提升(表1),从而有利于植物生长以及SOM输入的增加[2,10].值得注意的是,固沙植被的建立促进了生物土壤结皮的拓殖和定居,腾格里沙漠东南缘的研究表明,生物土壤结皮的年固碳速率可达11.36~26.75 g·m-1[34],这也是表层土壤SOC的主要贡献者. ...
Sand barriers of straw checkerboards for habitat restoration in extremely arid desert regions
1
2006
... 本研究中,固沙植被建立41 a后0~10 cm土层SOC含量比流沙增加9.59倍(表1,P0.01),表明种植旱生灌木是促进旱地SOC固存和土壤质量提升的有效措施.通常情况下,SOC的动态变化是由土壤有机质(Soil organic matter, SOM)输入和通过微生物分解的SOM输出过程共同决定的,当生态系统SOM输入大于输出时即表现为净SOC累积[2,24,31].一方面,固沙植被建立使植物来源SOM输入显著增大,其中地下部分生物量和凋落物分别解释了SOC变异的60%和90%(图3),而SOC与地上部分相关性不显著,这与Li等[2]的研究结果一致.这是因为随着植被演替,以灌木为优势的群落逐渐被以草本为优势的群落所取代,导致地上部分生物量呈减小趋势[11,24].另一方面,伴随着生态恢复,植被盖度显著增大(表1),沙丘表面逐渐趋于稳定,减少了风蚀引发的凋落物流失[2,32].且植物冠层能有效减小风速,增加了生态系统对大气降尘的捕获,而降尘中丰富的SOC和细颗粒物质提供了额外的碳源[2,33].此外,植被恢复促进了土壤理化性质的改善和土壤肥力的提升(表1),从而有利于植物生长以及SOM输入的增加[2,10].值得注意的是,固沙植被的建立促进了生物土壤结皮的拓殖和定居,腾格里沙漠东南缘的研究表明,生物土壤结皮的年固碳速率可达11.36~26.75 g·m-1[34],这也是表层土壤SOC的主要贡献者. ...
Changes in soil particulate and mineral-associated organic carbon concentrations under nitrogen addition in China:a meta-analysis
4
2023
... 已有研究表明,SOC的稳定性取决于其与矿物颗粒的相互作用和微生物的可及性,而MAOC与土壤矿物颗粒形成牢固的化学键,在提高SOC稳定性和长期封存方面发挥着至关重要的作用.因此,MAOC作为受微生物分解影响最小的碳库是SOC稳定性的重要表征[15].在本研究中,MAOC和POC含量及MAOC占SOC的比例随植被恢复年限的延长而显著增大,而POC占SOC的比例则呈相反趋势(图2,P0.01).这表明沙区植被恢复不仅促进了SOC的积累,同时使SOC组成由以POC为主逐渐向以MAOC为主转变,进而改变了SOC库的稳定性.POC主要来源于植物,由颗粒有机物的结构聚合物组成[14-17].因此,固沙植被区凋落物和地下部分生物量的显著增加是POC含量增大的主要原因,分别解释了其变异的89.3%和57.9%(图3).另一方面,固沙区土壤肥力的提升,特别是氮含量的显著增加不但促进SOM输入的增加和MBC的增大,而且加强了土壤团聚体保护[35],使POC含量进一步增加.已有研究表明,植物碳的微生物转化过程是SOC形成和稳定性维持的关键影响因素[36],植被恢复不仅增加了SOM的输入,而且增加土壤孔隙度和持水性,且使微生物基质(碳和养分)的数量和质量发生了显著变化(图1),土壤微环境更有利于与有机质分解相关的微生物群落的生长和繁殖[2,24];而EC的增大可通过增加微生物代谢熵而促进微生物对底物的利用,进一步促进微生物群落的发育[37],从而促进了植物残体的分解和POC和MAOC的累积[2,35],其中微生物群落变化解释了其变异的69%~85%(图3).另一方面,SOM的增加增强了土壤颗粒的胶结作用,有利于团聚体的形成,进一步促进了POC的增加[12-14].本研究中,POC占SOC的比例随植被恢复而显著减小,这主要是由于植被恢复引发的生物和非生物因子的改变,从而促进了MAOC截存,使得SOC库由以POC为主逐渐向以MAOC为主转变. ...
... ,35],其中微生物群落变化解释了其变异的69%~85%(图3).另一方面,SOM的增加增强了土壤颗粒的胶结作用,有利于团聚体的形成,进一步促进了POC的增加[12-14].本研究中,POC占SOC的比例随植被恢复而显著减小,这主要是由于植被恢复引发的生物和非生物因子的改变,从而促进了MAOC截存,使得SOC库由以POC为主逐渐向以MAOC为主转变. ...
... 土壤黏粉粒是MAOC的关键影响因素,植被恢复使土壤黏粉粒含量显著增大(表1),这增加了矿物对SOM的可及性,从而促进MAOC的形成[14-15].本研究中,黏粉粒含量是与MAOC相关性最大的因子,解释了其变量的95.7%(图3).而土壤微生物对凋落物和POC的分解是形成MAOC的先决条件[15,19,38],因此,MAOC的变化与POC密切相关(图3).微生物将较粗的植物来源聚合物(POC)进行分解,形成更易溶解的化合物(微生物细胞和细胞外化合物),这些化合物与土壤矿物质结合形成MAOC[36],从而使固沙区土壤MAOC显著增加(图2).另一方面,细菌和真菌在土壤中丰度的增加,可分泌更多的黏液和多糖,进一步促进稳定性碳的形成[39].而真菌可通过其丝状菌丝体聚集矿物颗粒,促进了有机分子的吸附和固定从而有利于形成MAOC[40].放线菌多样性的显著增大可有效促进土壤腐殖质的积累和转化,这也是形成稳定有机碳的重要过程[41].值得注意的是,POC的分解过程中释放的养分和能量能完全满足微生物生长的需求,从而减少了因养分需求而对MAOC的分解,进而有利于MAOC的累积[10].伴随着植被恢复,碳分解酶BG、CBH的活性和MBC也显著增大(图1).两种酶不但可以促进植物残体分解,同时能将SOC转化为可溶性葡萄糖和其他低分子量碳化合物并为微生物提供能量和碳源,从而促进POC和MAOC的积累[42-43].而MBC的增大是MAOC形成的重要前提,可有效促进SOC与矿物质之间的相互作用,从而形成更多的MAOC[41].微生物残体是MAOC的关键组成部分,微生物多样性及生物量的增大可使微生物残体及代谢分泌物显著增加,从而有利于MAOC的累积[12-13].土壤氮含量的增加可通过土壤有机残体与氮的缩合反应、促进有机-无机复合物的形成以及丛枝菌根真菌(AMF)的丰度增大从而促进植物残体形成MAOC[35,44]. ...
... 通常情况下,60%以上的SOC储存在MAOC库中[15].而在本研究中,固沙植被建立41 a后,MAOC占表层土壤SOC的比例为36.47%,这一数值显著小于已有研究结果.一方面,沙区降水稀少,土壤含水量长期处于较低水平,因此微生物对植物残体的分解速率较低,从而更有利于POC的积累[15,17].本研究中,MAOC及其占SOC的比例随黏粉粒含量增大而显著增大,这正表明了大部分SOC处于POC库,且从POC向MAOC的转化速率处于较低水平[45].而MAOC与黏粉粒和SOC含量之间显著的线性关系表明固沙植被区土壤具有较大的MAOC封存潜力[45].另一方面,沙区土壤粗颗粒占优势的土壤类型,黏粉粒含量较小,因此,土壤矿物表面特征特别是矿物可及性低下,不利于MAOC的形成[12,14-15].真菌群落通常分解化学上难降解的SOC,而微生物群落中真菌相对于细菌的优势逐渐增大(F∶B增大)可促进MAOC的分解[46],因此真菌PLFA与MAOC呈负相关关系(图3和图5).此外,固沙区土壤pH显著减小,土壤酸化可能导致微生物多样性及生物量的增加减缓,从而降低了MAOC的增加速率[35].值得注意的是,尽管植被恢复有效地促进了SOC及其稳定性的增加,然而本研究区SOC、MAOC和POC含量仍处于较低水平,特别是MAOC占SOC比例的增加速率随植被恢复年限的延长和SOC含量的增加均呈减小趋势(图2和图4),表明沙区土壤碳储量和稳定性的恢复是一个长期过程.因此,水分限制生态系统修复是土壤生境的关键,而生态系统管理措施的优化是解决这一问题的核心. ...
Changes in plant inputs alter soil carbon and microbial communities in forest ecosystems
2
2022
... 已有研究表明,SOC的稳定性取决于其与矿物颗粒的相互作用和微生物的可及性,而MAOC与土壤矿物颗粒形成牢固的化学键,在提高SOC稳定性和长期封存方面发挥着至关重要的作用.因此,MAOC作为受微生物分解影响最小的碳库是SOC稳定性的重要表征[15].在本研究中,MAOC和POC含量及MAOC占SOC的比例随植被恢复年限的延长而显著增大,而POC占SOC的比例则呈相反趋势(图2,P0.01).这表明沙区植被恢复不仅促进了SOC的积累,同时使SOC组成由以POC为主逐渐向以MAOC为主转变,进而改变了SOC库的稳定性.POC主要来源于植物,由颗粒有机物的结构聚合物组成[14-17].因此,固沙植被区凋落物和地下部分生物量的显著增加是POC含量增大的主要原因,分别解释了其变异的89.3%和57.9%(图3).另一方面,固沙区土壤肥力的提升,特别是氮含量的显著增加不但促进SOM输入的增加和MBC的增大,而且加强了土壤团聚体保护[35],使POC含量进一步增加.已有研究表明,植物碳的微生物转化过程是SOC形成和稳定性维持的关键影响因素[36],植被恢复不仅增加了SOM的输入,而且增加土壤孔隙度和持水性,且使微生物基质(碳和养分)的数量和质量发生了显著变化(图1),土壤微环境更有利于与有机质分解相关的微生物群落的生长和繁殖[2,24];而EC的增大可通过增加微生物代谢熵而促进微生物对底物的利用,进一步促进微生物群落的发育[37],从而促进了植物残体的分解和POC和MAOC的累积[2,35],其中微生物群落变化解释了其变异的69%~85%(图3).另一方面,SOM的增加增强了土壤颗粒的胶结作用,有利于团聚体的形成,进一步促进了POC的增加[12-14].本研究中,POC占SOC的比例随植被恢复而显著减小,这主要是由于植被恢复引发的生物和非生物因子的改变,从而促进了MAOC截存,使得SOC库由以POC为主逐渐向以MAOC为主转变. ...
... 土壤黏粉粒是MAOC的关键影响因素,植被恢复使土壤黏粉粒含量显著增大(表1),这增加了矿物对SOM的可及性,从而促进MAOC的形成[14-15].本研究中,黏粉粒含量是与MAOC相关性最大的因子,解释了其变量的95.7%(图3).而土壤微生物对凋落物和POC的分解是形成MAOC的先决条件[15,19,38],因此,MAOC的变化与POC密切相关(图3).微生物将较粗的植物来源聚合物(POC)进行分解,形成更易溶解的化合物(微生物细胞和细胞外化合物),这些化合物与土壤矿物质结合形成MAOC[36],从而使固沙区土壤MAOC显著增加(图2).另一方面,细菌和真菌在土壤中丰度的增加,可分泌更多的黏液和多糖,进一步促进稳定性碳的形成[39].而真菌可通过其丝状菌丝体聚集矿物颗粒,促进了有机分子的吸附和固定从而有利于形成MAOC[40].放线菌多样性的显著增大可有效促进土壤腐殖质的积累和转化,这也是形成稳定有机碳的重要过程[41].值得注意的是,POC的分解过程中释放的养分和能量能完全满足微生物生长的需求,从而减少了因养分需求而对MAOC的分解,进而有利于MAOC的累积[10].伴随着植被恢复,碳分解酶BG、CBH的活性和MBC也显著增大(图1).两种酶不但可以促进植物残体分解,同时能将SOC转化为可溶性葡萄糖和其他低分子量碳化合物并为微生物提供能量和碳源,从而促进POC和MAOC的积累[42-43].而MBC的增大是MAOC形成的重要前提,可有效促进SOC与矿物质之间的相互作用,从而形成更多的MAOC[41].微生物残体是MAOC的关键组成部分,微生物多样性及生物量的增大可使微生物残体及代谢分泌物显著增加,从而有利于MAOC的累积[12-13].土壤氮含量的增加可通过土壤有机残体与氮的缩合反应、促进有机-无机复合物的形成以及丛枝菌根真菌(AMF)的丰度增大从而促进植物残体形成MAOC[35,44]. ...
Soil property and microbial activity in natural salt affected soils in an alternating wet-dry tropical climate
1
2012
... 已有研究表明,SOC的稳定性取决于其与矿物颗粒的相互作用和微生物的可及性,而MAOC与土壤矿物颗粒形成牢固的化学键,在提高SOC稳定性和长期封存方面发挥着至关重要的作用.因此,MAOC作为受微生物分解影响最小的碳库是SOC稳定性的重要表征[15].在本研究中,MAOC和POC含量及MAOC占SOC的比例随植被恢复年限的延长而显著增大,而POC占SOC的比例则呈相反趋势(图2,P0.01).这表明沙区植被恢复不仅促进了SOC的积累,同时使SOC组成由以POC为主逐渐向以MAOC为主转变,进而改变了SOC库的稳定性.POC主要来源于植物,由颗粒有机物的结构聚合物组成[14-17].因此,固沙植被区凋落物和地下部分生物量的显著增加是POC含量增大的主要原因,分别解释了其变异的89.3%和57.9%(图3).另一方面,固沙区土壤肥力的提升,特别是氮含量的显著增加不但促进SOM输入的增加和MBC的增大,而且加强了土壤团聚体保护[35],使POC含量进一步增加.已有研究表明,植物碳的微生物转化过程是SOC形成和稳定性维持的关键影响因素[36],植被恢复不仅增加了SOM的输入,而且增加土壤孔隙度和持水性,且使微生物基质(碳和养分)的数量和质量发生了显著变化(图1),土壤微环境更有利于与有机质分解相关的微生物群落的生长和繁殖[2,24];而EC的增大可通过增加微生物代谢熵而促进微生物对底物的利用,进一步促进微生物群落的发育[37],从而促进了植物残体的分解和POC和MAOC的累积[2,35],其中微生物群落变化解释了其变异的69%~85%(图3).另一方面,SOM的增加增强了土壤颗粒的胶结作用,有利于团聚体的形成,进一步促进了POC的增加[12-14].本研究中,POC占SOC的比例随植被恢复而显著减小,这主要是由于植被恢复引发的生物和非生物因子的改变,从而促进了MAOC截存,使得SOC库由以POC为主逐渐向以MAOC为主转变. ...
Biological mechanisms may contribute to soil carbon saturation patterns
1
2021
... 土壤黏粉粒是MAOC的关键影响因素,植被恢复使土壤黏粉粒含量显著增大(表1),这增加了矿物对SOM的可及性,从而促进MAOC的形成[14-15].本研究中,黏粉粒含量是与MAOC相关性最大的因子,解释了其变量的95.7%(图3).而土壤微生物对凋落物和POC的分解是形成MAOC的先决条件[15,19,38],因此,MAOC的变化与POC密切相关(图3).微生物将较粗的植物来源聚合物(POC)进行分解,形成更易溶解的化合物(微生物细胞和细胞外化合物),这些化合物与土壤矿物质结合形成MAOC[36],从而使固沙区土壤MAOC显著增加(图2).另一方面,细菌和真菌在土壤中丰度的增加,可分泌更多的黏液和多糖,进一步促进稳定性碳的形成[39].而真菌可通过其丝状菌丝体聚集矿物颗粒,促进了有机分子的吸附和固定从而有利于形成MAOC[40].放线菌多样性的显著增大可有效促进土壤腐殖质的积累和转化,这也是形成稳定有机碳的重要过程[41].值得注意的是,POC的分解过程中释放的养分和能量能完全满足微生物生长的需求,从而减少了因养分需求而对MAOC的分解,进而有利于MAOC的累积[10].伴随着植被恢复,碳分解酶BG、CBH的活性和MBC也显著增大(图1).两种酶不但可以促进植物残体分解,同时能将SOC转化为可溶性葡萄糖和其他低分子量碳化合物并为微生物提供能量和碳源,从而促进POC和MAOC的积累[42-43].而MBC的增大是MAOC形成的重要前提,可有效促进SOC与矿物质之间的相互作用,从而形成更多的MAOC[41].微生物残体是MAOC的关键组成部分,微生物多样性及生物量的增大可使微生物残体及代谢分泌物显著增加,从而有利于MAOC的累积[12-13].土壤氮含量的增加可通过土壤有机残体与氮的缩合反应、促进有机-无机复合物的形成以及丛枝菌根真菌(AMF)的丰度增大从而促进植物残体形成MAOC[35,44]. ...
Chemical constituents and biochemistry
1
2000
... 土壤黏粉粒是MAOC的关键影响因素,植被恢复使土壤黏粉粒含量显著增大(表1),这增加了矿物对SOM的可及性,从而促进MAOC的形成[14-15].本研究中,黏粉粒含量是与MAOC相关性最大的因子,解释了其变量的95.7%(图3).而土壤微生物对凋落物和POC的分解是形成MAOC的先决条件[15,19,38],因此,MAOC的变化与POC密切相关(图3).微生物将较粗的植物来源聚合物(POC)进行分解,形成更易溶解的化合物(微生物细胞和细胞外化合物),这些化合物与土壤矿物质结合形成MAOC[36],从而使固沙区土壤MAOC显著增加(图2).另一方面,细菌和真菌在土壤中丰度的增加,可分泌更多的黏液和多糖,进一步促进稳定性碳的形成[39].而真菌可通过其丝状菌丝体聚集矿物颗粒,促进了有机分子的吸附和固定从而有利于形成MAOC[40].放线菌多样性的显著增大可有效促进土壤腐殖质的积累和转化,这也是形成稳定有机碳的重要过程[41].值得注意的是,POC的分解过程中释放的养分和能量能完全满足微生物生长的需求,从而减少了因养分需求而对MAOC的分解,进而有利于MAOC的累积[10].伴随着植被恢复,碳分解酶BG、CBH的活性和MBC也显著增大(图1).两种酶不但可以促进植物残体分解,同时能将SOC转化为可溶性葡萄糖和其他低分子量碳化合物并为微生物提供能量和碳源,从而促进POC和MAOC的积累[42-43].而MBC的增大是MAOC形成的重要前提,可有效促进SOC与矿物质之间的相互作用,从而形成更多的MAOC[41].微生物残体是MAOC的关键组成部分,微生物多样性及生物量的增大可使微生物残体及代谢分泌物显著增加,从而有利于MAOC的累积[12-13].土壤氮含量的增加可通过土壤有机残体与氮的缩合反应、促进有机-无机复合物的形成以及丛枝菌根真菌(AMF)的丰度增大从而促进植物残体形成MAOC[35,44]. ...
Diversity of free-living and lichenized fungal communities in biological soil crusts of the Sultanate of Oman and their role in improving soil properties
1
2013
... 土壤黏粉粒是MAOC的关键影响因素,植被恢复使土壤黏粉粒含量显著增大(表1),这增加了矿物对SOM的可及性,从而促进MAOC的形成[14-15].本研究中,黏粉粒含量是与MAOC相关性最大的因子,解释了其变量的95.7%(图3).而土壤微生物对凋落物和POC的分解是形成MAOC的先决条件[15,19,38],因此,MAOC的变化与POC密切相关(图3).微生物将较粗的植物来源聚合物(POC)进行分解,形成更易溶解的化合物(微生物细胞和细胞外化合物),这些化合物与土壤矿物质结合形成MAOC[36],从而使固沙区土壤MAOC显著增加(图2).另一方面,细菌和真菌在土壤中丰度的增加,可分泌更多的黏液和多糖,进一步促进稳定性碳的形成[39].而真菌可通过其丝状菌丝体聚集矿物颗粒,促进了有机分子的吸附和固定从而有利于形成MAOC[40].放线菌多样性的显著增大可有效促进土壤腐殖质的积累和转化,这也是形成稳定有机碳的重要过程[41].值得注意的是,POC的分解过程中释放的养分和能量能完全满足微生物生长的需求,从而减少了因养分需求而对MAOC的分解,进而有利于MAOC的累积[10].伴随着植被恢复,碳分解酶BG、CBH的活性和MBC也显著增大(图1).两种酶不但可以促进植物残体分解,同时能将SOC转化为可溶性葡萄糖和其他低分子量碳化合物并为微生物提供能量和碳源,从而促进POC和MAOC的积累[42-43].而MBC的增大是MAOC形成的重要前提,可有效促进SOC与矿物质之间的相互作用,从而形成更多的MAOC[41].微生物残体是MAOC的关键组成部分,微生物多样性及生物量的增大可使微生物残体及代谢分泌物显著增加,从而有利于MAOC的累积[12-13].土壤氮含量的增加可通过土壤有机残体与氮的缩合反应、促进有机-无机复合物的形成以及丛枝菌根真菌(AMF)的丰度增大从而促进植物残体形成MAOC[35,44]. ...
Fertilizer reduction and biochar amendment promote soil mineral-associated organic carbon,bacterial activity,and enzyme activity in a jasmine garden in southeast China
2
2024
... 土壤黏粉粒是MAOC的关键影响因素,植被恢复使土壤黏粉粒含量显著增大(表1),这增加了矿物对SOM的可及性,从而促进MAOC的形成[14-15].本研究中,黏粉粒含量是与MAOC相关性最大的因子,解释了其变量的95.7%(图3).而土壤微生物对凋落物和POC的分解是形成MAOC的先决条件[15,19,38],因此,MAOC的变化与POC密切相关(图3).微生物将较粗的植物来源聚合物(POC)进行分解,形成更易溶解的化合物(微生物细胞和细胞外化合物),这些化合物与土壤矿物质结合形成MAOC[36],从而使固沙区土壤MAOC显著增加(图2).另一方面,细菌和真菌在土壤中丰度的增加,可分泌更多的黏液和多糖,进一步促进稳定性碳的形成[39].而真菌可通过其丝状菌丝体聚集矿物颗粒,促进了有机分子的吸附和固定从而有利于形成MAOC[40].放线菌多样性的显著增大可有效促进土壤腐殖质的积累和转化,这也是形成稳定有机碳的重要过程[41].值得注意的是,POC的分解过程中释放的养分和能量能完全满足微生物生长的需求,从而减少了因养分需求而对MAOC的分解,进而有利于MAOC的累积[10].伴随着植被恢复,碳分解酶BG、CBH的活性和MBC也显著增大(图1).两种酶不但可以促进植物残体分解,同时能将SOC转化为可溶性葡萄糖和其他低分子量碳化合物并为微生物提供能量和碳源,从而促进POC和MAOC的积累[42-43].而MBC的增大是MAOC形成的重要前提,可有效促进SOC与矿物质之间的相互作用,从而形成更多的MAOC[41].微生物残体是MAOC的关键组成部分,微生物多样性及生物量的增大可使微生物残体及代谢分泌物显著增加,从而有利于MAOC的累积[12-13].土壤氮含量的增加可通过土壤有机残体与氮的缩合反应、促进有机-无机复合物的形成以及丛枝菌根真菌(AMF)的丰度增大从而促进植物残体形成MAOC[35,44]. ...
... [41].微生物残体是MAOC的关键组成部分,微生物多样性及生物量的增大可使微生物残体及代谢分泌物显著增加,从而有利于MAOC的累积[12-13].土壤氮含量的增加可通过土壤有机残体与氮的缩合反应、促进有机-无机复合物的形成以及丛枝菌根真菌(AMF)的丰度增大从而促进植物残体形成MAOC[35,44]. ...
Effects of wood char and activated carbon on the hydrolysis of cellobiose by β-glucosidase from Aspergillus niger
1
2011
... 土壤黏粉粒是MAOC的关键影响因素,植被恢复使土壤黏粉粒含量显著增大(表1),这增加了矿物对SOM的可及性,从而促进MAOC的形成[14-15].本研究中,黏粉粒含量是与MAOC相关性最大的因子,解释了其变量的95.7%(图3).而土壤微生物对凋落物和POC的分解是形成MAOC的先决条件[15,19,38],因此,MAOC的变化与POC密切相关(图3).微生物将较粗的植物来源聚合物(POC)进行分解,形成更易溶解的化合物(微生物细胞和细胞外化合物),这些化合物与土壤矿物质结合形成MAOC[36],从而使固沙区土壤MAOC显著增加(图2).另一方面,细菌和真菌在土壤中丰度的增加,可分泌更多的黏液和多糖,进一步促进稳定性碳的形成[39].而真菌可通过其丝状菌丝体聚集矿物颗粒,促进了有机分子的吸附和固定从而有利于形成MAOC[40].放线菌多样性的显著增大可有效促进土壤腐殖质的积累和转化,这也是形成稳定有机碳的重要过程[41].值得注意的是,POC的分解过程中释放的养分和能量能完全满足微生物生长的需求,从而减少了因养分需求而对MAOC的分解,进而有利于MAOC的累积[10].伴随着植被恢复,碳分解酶BG、CBH的活性和MBC也显著增大(图1).两种酶不但可以促进植物残体分解,同时能将SOC转化为可溶性葡萄糖和其他低分子量碳化合物并为微生物提供能量和碳源,从而促进POC和MAOC的积累[42-43].而MBC的增大是MAOC形成的重要前提,可有效促进SOC与矿物质之间的相互作用,从而形成更多的MAOC[41].微生物残体是MAOC的关键组成部分,微生物多样性及生物量的增大可使微生物残体及代谢分泌物显著增加,从而有利于MAOC的累积[12-13].土壤氮含量的增加可通过土壤有机残体与氮的缩合反应、促进有机-无机复合物的形成以及丛枝菌根真菌(AMF)的丰度增大从而促进植物残体形成MAOC[35,44]. ...
Enzyme activities and litter decomposition in agricultural soils in northern,central,and southern Germany
1
2007
... 土壤黏粉粒是MAOC的关键影响因素,植被恢复使土壤黏粉粒含量显著增大(表1),这增加了矿物对SOM的可及性,从而促进MAOC的形成[14-15].本研究中,黏粉粒含量是与MAOC相关性最大的因子,解释了其变量的95.7%(图3).而土壤微生物对凋落物和POC的分解是形成MAOC的先决条件[15,19,38],因此,MAOC的变化与POC密切相关(图3).微生物将较粗的植物来源聚合物(POC)进行分解,形成更易溶解的化合物(微生物细胞和细胞外化合物),这些化合物与土壤矿物质结合形成MAOC[36],从而使固沙区土壤MAOC显著增加(图2).另一方面,细菌和真菌在土壤中丰度的增加,可分泌更多的黏液和多糖,进一步促进稳定性碳的形成[39].而真菌可通过其丝状菌丝体聚集矿物颗粒,促进了有机分子的吸附和固定从而有利于形成MAOC[40].放线菌多样性的显著增大可有效促进土壤腐殖质的积累和转化,这也是形成稳定有机碳的重要过程[41].值得注意的是,POC的分解过程中释放的养分和能量能完全满足微生物生长的需求,从而减少了因养分需求而对MAOC的分解,进而有利于MAOC的累积[10].伴随着植被恢复,碳分解酶BG、CBH的活性和MBC也显著增大(图1).两种酶不但可以促进植物残体分解,同时能将SOC转化为可溶性葡萄糖和其他低分子量碳化合物并为微生物提供能量和碳源,从而促进POC和MAOC的积累[42-43].而MBC的增大是MAOC形成的重要前提,可有效促进SOC与矿物质之间的相互作用,从而形成更多的MAOC[41].微生物残体是MAOC的关键组成部分,微生物多样性及生物量的增大可使微生物残体及代谢分泌物显著增加,从而有利于MAOC的累积[12-13].土壤氮含量的增加可通过土壤有机残体与氮的缩合反应、促进有机-无机复合物的形成以及丛枝菌根真菌(AMF)的丰度增大从而促进植物残体形成MAOC[35,44]. ...
Role of mineral-nitrogen in residue decomposition and stable soil organic matter formation
1
2005
... 土壤黏粉粒是MAOC的关键影响因素,植被恢复使土壤黏粉粒含量显著增大(表1),这增加了矿物对SOM的可及性,从而促进MAOC的形成[14-15].本研究中,黏粉粒含量是与MAOC相关性最大的因子,解释了其变量的95.7%(图3).而土壤微生物对凋落物和POC的分解是形成MAOC的先决条件[15,19,38],因此,MAOC的变化与POC密切相关(图3).微生物将较粗的植物来源聚合物(POC)进行分解,形成更易溶解的化合物(微生物细胞和细胞外化合物),这些化合物与土壤矿物质结合形成MAOC[36],从而使固沙区土壤MAOC显著增加(图2).另一方面,细菌和真菌在土壤中丰度的增加,可分泌更多的黏液和多糖,进一步促进稳定性碳的形成[39].而真菌可通过其丝状菌丝体聚集矿物颗粒,促进了有机分子的吸附和固定从而有利于形成MAOC[40].放线菌多样性的显著增大可有效促进土壤腐殖质的积累和转化,这也是形成稳定有机碳的重要过程[41].值得注意的是,POC的分解过程中释放的养分和能量能完全满足微生物生长的需求,从而减少了因养分需求而对MAOC的分解,进而有利于MAOC的累积[10].伴随着植被恢复,碳分解酶BG、CBH的活性和MBC也显著增大(图1).两种酶不但可以促进植物残体分解,同时能将SOC转化为可溶性葡萄糖和其他低分子量碳化合物并为微生物提供能量和碳源,从而促进POC和MAOC的积累[42-43].而MBC的增大是MAOC形成的重要前提,可有效促进SOC与矿物质之间的相互作用,从而形成更多的MAOC[41].微生物残体是MAOC的关键组成部分,微生物多样性及生物量的增大可使微生物残体及代谢分泌物显著增加,从而有利于MAOC的累积[12-13].土壤氮含量的增加可通过土壤有机残体与氮的缩合反应、促进有机-无机复合物的形成以及丛枝菌根真菌(AMF)的丰度增大从而促进植物残体形成MAOC[35,44]. ...
No detectable upper limit of mineral-associated organic carbon in temperate agricultural soils
2
2023
... 通常情况下,60%以上的SOC储存在MAOC库中[15].而在本研究中,固沙植被建立41 a后,MAOC占表层土壤SOC的比例为36.47%,这一数值显著小于已有研究结果.一方面,沙区降水稀少,土壤含水量长期处于较低水平,因此微生物对植物残体的分解速率较低,从而更有利于POC的积累[15,17].本研究中,MAOC及其占SOC的比例随黏粉粒含量增大而显著增大,这正表明了大部分SOC处于POC库,且从POC向MAOC的转化速率处于较低水平[45].而MAOC与黏粉粒和SOC含量之间显著的线性关系表明固沙植被区土壤具有较大的MAOC封存潜力[45].另一方面,沙区土壤粗颗粒占优势的土壤类型,黏粉粒含量较小,因此,土壤矿物表面特征特别是矿物可及性低下,不利于MAOC的形成[12,14-15].真菌群落通常分解化学上难降解的SOC,而微生物群落中真菌相对于细菌的优势逐渐增大(F∶B增大)可促进MAOC的分解[46],因此真菌PLFA与MAOC呈负相关关系(图3和图5).此外,固沙区土壤pH显著减小,土壤酸化可能导致微生物多样性及生物量的增加减缓,从而降低了MAOC的增加速率[35].值得注意的是,尽管植被恢复有效地促进了SOC及其稳定性的增加,然而本研究区SOC、MAOC和POC含量仍处于较低水平,特别是MAOC占SOC比例的增加速率随植被恢复年限的延长和SOC含量的增加均呈减小趋势(图2和图4),表明沙区土壤碳储量和稳定性的恢复是一个长期过程.因此,水分限制生态系统修复是土壤生境的关键,而生态系统管理措施的优化是解决这一问题的核心. ...
... [45].另一方面,沙区土壤粗颗粒占优势的土壤类型,黏粉粒含量较小,因此,土壤矿物表面特征特别是矿物可及性低下,不利于MAOC的形成[12,14-15].真菌群落通常分解化学上难降解的SOC,而微生物群落中真菌相对于细菌的优势逐渐增大(F∶B增大)可促进MAOC的分解[46],因此真菌PLFA与MAOC呈负相关关系(图3和图5).此外,固沙区土壤pH显著减小,土壤酸化可能导致微生物多样性及生物量的增加减缓,从而降低了MAOC的增加速率[35].值得注意的是,尽管植被恢复有效地促进了SOC及其稳定性的增加,然而本研究区SOC、MAOC和POC含量仍处于较低水平,特别是MAOC占SOC比例的增加速率随植被恢复年限的延长和SOC含量的增加均呈减小趋势(图2和图4),表明沙区土壤碳储量和稳定性的恢复是一个长期过程.因此,水分限制生态系统修复是土壤生境的关键,而生态系统管理措施的优化是解决这一问题的核心. ...
Carbon and nitrogen mineralization from light-and heavy-fraction additions to soil
1
2000
... 通常情况下,60%以上的SOC储存在MAOC库中[15].而在本研究中,固沙植被建立41 a后,MAOC占表层土壤SOC的比例为36.47%,这一数值显著小于已有研究结果.一方面,沙区降水稀少,土壤含水量长期处于较低水平,因此微生物对植物残体的分解速率较低,从而更有利于POC的积累[15,17].本研究中,MAOC及其占SOC的比例随黏粉粒含量增大而显著增大,这正表明了大部分SOC处于POC库,且从POC向MAOC的转化速率处于较低水平[45].而MAOC与黏粉粒和SOC含量之间显著的线性关系表明固沙植被区土壤具有较大的MAOC封存潜力[45].另一方面,沙区土壤粗颗粒占优势的土壤类型,黏粉粒含量较小,因此,土壤矿物表面特征特别是矿物可及性低下,不利于MAOC的形成[12,14-15].真菌群落通常分解化学上难降解的SOC,而微生物群落中真菌相对于细菌的优势逐渐增大(F∶B增大)可促进MAOC的分解[46],因此真菌PLFA与MAOC呈负相关关系(图3和图5).此外,固沙区土壤pH显著减小,土壤酸化可能导致微生物多样性及生物量的增加减缓,从而降低了MAOC的增加速率[35].值得注意的是,尽管植被恢复有效地促进了SOC及其稳定性的增加,然而本研究区SOC、MAOC和POC含量仍处于较低水平,特别是MAOC占SOC比例的增加速率随植被恢复年限的延长和SOC含量的增加均呈减小趋势(图2和图4),表明沙区土壤碳储量和稳定性的恢复是一个长期过程.因此,水分限制生态系统修复是土壤生境的关键,而生态系统管理措施的优化是解决这一问题的核心. ...








 甘公网安备 62010202000688号
甘公网安备 62010202000688号
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}