Assessing vulnerability to climate change in dryland livelihood systems:conceptual challenges and interdisciplinary solutions
1
2011
... 干旱半干旱区的生态环境极其脆弱,对全球气候变化高度敏感[1].近几十年来,人口增长和快速城镇化对全国生态系统造成了巨大压力[2],导致生态服务功能显著退化[3].尽管如此,人类活动通过改变土地利用,能够直接影响旱地生态系统的植被覆盖率,并对土壤有机碳产生了重要影响[4].土壤有机碳的快速转变又会导致植被结构和组成的改变[5].此外,全球气候变化引发的长期干旱和短期强降雨,对植被生长产生了不同程度的影响.长期干旱通常导致生物生产力难以逆转的下降,而强降雨虽可能缓解短期生长压力[6],但也可能引发土壤水分剧烈波动,影响植物稳定生存[7].与此同时,气温的快速波动加剧了这些问题,导致植物生长季节和物种分布的变化.尽管某些植物能够适应温度的变化,但剧烈的气候变动可能超出许多物种的适应极限[8].因此,植被对人口和气候的急剧变化可能做出强烈反应,其反应形式可能是非线性或曲线型,甚至具有突发性.这使得研究植被突变尤为重要,以便深入探讨和验证这些变化的机制. ...
城镇化进程中的生态环境问题及对策研究
1
2014
... 干旱半干旱区的生态环境极其脆弱,对全球气候变化高度敏感[1].近几十年来,人口增长和快速城镇化对全国生态系统造成了巨大压力[2],导致生态服务功能显著退化[3].尽管如此,人类活动通过改变土地利用,能够直接影响旱地生态系统的植被覆盖率,并对土壤有机碳产生了重要影响[4].土壤有机碳的快速转变又会导致植被结构和组成的改变[5].此外,全球气候变化引发的长期干旱和短期强降雨,对植被生长产生了不同程度的影响.长期干旱通常导致生物生产力难以逆转的下降,而强降雨虽可能缓解短期生长压力[6],但也可能引发土壤水分剧烈波动,影响植物稳定生存[7].与此同时,气温的快速波动加剧了这些问题,导致植物生长季节和物种分布的变化.尽管某些植物能够适应温度的变化,但剧烈的气候变动可能超出许多物种的适应极限[8].因此,植被对人口和气候的急剧变化可能做出强烈反应,其反应形式可能是非线性或曲线型,甚至具有突发性.这使得研究植被突变尤为重要,以便深入探讨和验证这些变化的机制. ...
甘肃省河西地区水源涵养功能与人类活动强度演变
1
2024
... 干旱半干旱区的生态环境极其脆弱,对全球气候变化高度敏感[1].近几十年来,人口增长和快速城镇化对全国生态系统造成了巨大压力[2],导致生态服务功能显著退化[3].尽管如此,人类活动通过改变土地利用,能够直接影响旱地生态系统的植被覆盖率,并对土壤有机碳产生了重要影响[4].土壤有机碳的快速转变又会导致植被结构和组成的改变[5].此外,全球气候变化引发的长期干旱和短期强降雨,对植被生长产生了不同程度的影响.长期干旱通常导致生物生产力难以逆转的下降,而强降雨虽可能缓解短期生长压力[6],但也可能引发土壤水分剧烈波动,影响植物稳定生存[7].与此同时,气温的快速波动加剧了这些问题,导致植物生长季节和物种分布的变化.尽管某些植物能够适应温度的变化,但剧烈的气候变动可能超出许多物种的适应极限[8].因此,植被对人口和气候的急剧变化可能做出强烈反应,其反应形式可能是非线性或曲线型,甚至具有突发性.这使得研究植被突变尤为重要,以便深入探讨和验证这些变化的机制. ...
黄土高原4种典型植被对土壤活性有机碳及土壤碳库的影响
1
... 干旱半干旱区的生态环境极其脆弱,对全球气候变化高度敏感[1].近几十年来,人口增长和快速城镇化对全国生态系统造成了巨大压力[2],导致生态服务功能显著退化[3].尽管如此,人类活动通过改变土地利用,能够直接影响旱地生态系统的植被覆盖率,并对土壤有机碳产生了重要影响[4].土壤有机碳的快速转变又会导致植被结构和组成的改变[5].此外,全球气候变化引发的长期干旱和短期强降雨,对植被生长产生了不同程度的影响.长期干旱通常导致生物生产力难以逆转的下降,而强降雨虽可能缓解短期生长压力[6],但也可能引发土壤水分剧烈波动,影响植物稳定生存[7].与此同时,气温的快速波动加剧了这些问题,导致植物生长季节和物种分布的变化.尽管某些植物能够适应温度的变化,但剧烈的气候变动可能超出许多物种的适应极限[8].因此,植被对人口和气候的急剧变化可能做出强烈反应,其反应形式可能是非线性或曲线型,甚至具有突发性.这使得研究植被突变尤为重要,以便深入探讨和验证这些变化的机制. ...
Global consequences of land use
1
2005
... 干旱半干旱区的生态环境极其脆弱,对全球气候变化高度敏感[1].近几十年来,人口增长和快速城镇化对全国生态系统造成了巨大压力[2],导致生态服务功能显著退化[3].尽管如此,人类活动通过改变土地利用,能够直接影响旱地生态系统的植被覆盖率,并对土壤有机碳产生了重要影响[4].土壤有机碳的快速转变又会导致植被结构和组成的改变[5].此外,全球气候变化引发的长期干旱和短期强降雨,对植被生长产生了不同程度的影响.长期干旱通常导致生物生产力难以逆转的下降,而强降雨虽可能缓解短期生长压力[6],但也可能引发土壤水分剧烈波动,影响植物稳定生存[7].与此同时,气温的快速波动加剧了这些问题,导致植物生长季节和物种分布的变化.尽管某些植物能够适应温度的变化,但剧烈的气候变动可能超出许多物种的适应极限[8].因此,植被对人口和气候的急剧变化可能做出强烈反应,其反应形式可能是非线性或曲线型,甚至具有突发性.这使得研究植被突变尤为重要,以便深入探讨和验证这些变化的机制. ...
Consequences of more extreme precipitation regimes for terrestrial ecosystems
1
2008
... 干旱半干旱区的生态环境极其脆弱,对全球气候变化高度敏感[1].近几十年来,人口增长和快速城镇化对全国生态系统造成了巨大压力[2],导致生态服务功能显著退化[3].尽管如此,人类活动通过改变土地利用,能够直接影响旱地生态系统的植被覆盖率,并对土壤有机碳产生了重要影响[4].土壤有机碳的快速转变又会导致植被结构和组成的改变[5].此外,全球气候变化引发的长期干旱和短期强降雨,对植被生长产生了不同程度的影响.长期干旱通常导致生物生产力难以逆转的下降,而强降雨虽可能缓解短期生长压力[6],但也可能引发土壤水分剧烈波动,影响植物稳定生存[7].与此同时,气温的快速波动加剧了这些问题,导致植物生长季节和物种分布的变化.尽管某些植物能够适应温度的变化,但剧烈的气候变动可能超出许多物种的适应极限[8].因此,植被对人口和气候的急剧变化可能做出强烈反应,其反应形式可能是非线性或曲线型,甚至具有突发性.这使得研究植被突变尤为重要,以便深入探讨和验证这些变化的机制. ...
Climate extremes and the carbon cycle
1
2013
... 干旱半干旱区的生态环境极其脆弱,对全球气候变化高度敏感[1].近几十年来,人口增长和快速城镇化对全国生态系统造成了巨大压力[2],导致生态服务功能显著退化[3].尽管如此,人类活动通过改变土地利用,能够直接影响旱地生态系统的植被覆盖率,并对土壤有机碳产生了重要影响[4].土壤有机碳的快速转变又会导致植被结构和组成的改变[5].此外,全球气候变化引发的长期干旱和短期强降雨,对植被生长产生了不同程度的影响.长期干旱通常导致生物生产力难以逆转的下降,而强降雨虽可能缓解短期生长压力[6],但也可能引发土壤水分剧烈波动,影响植物稳定生存[7].与此同时,气温的快速波动加剧了这些问题,导致植物生长季节和物种分布的变化.尽管某些植物能够适应温度的变化,但剧烈的气候变动可能超出许多物种的适应极限[8].因此,植被对人口和气候的急剧变化可能做出强烈反应,其反应形式可能是非线性或曲线型,甚至具有突发性.这使得研究植被突变尤为重要,以便深入探讨和验证这些变化的机制. ...
Running to stand still:adaptation and the response of plants to rapid climate change
1
2005
... 干旱半干旱区的生态环境极其脆弱,对全球气候变化高度敏感[1].近几十年来,人口增长和快速城镇化对全国生态系统造成了巨大压力[2],导致生态服务功能显著退化[3].尽管如此,人类活动通过改变土地利用,能够直接影响旱地生态系统的植被覆盖率,并对土壤有机碳产生了重要影响[4].土壤有机碳的快速转变又会导致植被结构和组成的改变[5].此外,全球气候变化引发的长期干旱和短期强降雨,对植被生长产生了不同程度的影响.长期干旱通常导致生物生产力难以逆转的下降,而强降雨虽可能缓解短期生长压力[6],但也可能引发土壤水分剧烈波动,影响植物稳定生存[7].与此同时,气温的快速波动加剧了这些问题,导致植物生长季节和物种分布的变化.尽管某些植物能够适应温度的变化,但剧烈的气候变动可能超出许多物种的适应极限[8].因此,植被对人口和气候的急剧变化可能做出强烈反应,其反应形式可能是非线性或曲线型,甚至具有突发性.这使得研究植被突变尤为重要,以便深入探讨和验证这些变化的机制. ...
Prevalence and drivers of abrupt vegetation shifts in global drylands
3
2022
... 全球范围内,约四分之一的旱地植被经历了显著的突变,同时揭示了气候变异性和人类活动在植被突变中扮演的关键角色[9].中国黄土高原29%的区域发生了突变,温度上升和人口增长对植被突变有显著贡献[10].这表明干旱区植被突变的驱动机制在全球和区域尺度并不一致,因此,西北干旱半干旱区植被突变是一个较为迫切的科学问题.同时,干旱梯度下的植被突变特征和驱动机制尚未得到充分探讨. ...
... 使用R语言“chngpt”包中的阶跃模型(Step)定义植被的突变,并用“glm” 的formula参数定义非突变.在多模型轨迹诊断方法中,使用Akaike信息准则(Akaike Information Criterion, AIC)评估模型对数据的拟合程度和模型复杂度,通常选择AIC最小的模型.同时,本研究采用自举方法降低轨迹分类的不确定性,将每个回归迭代100次[9].此外,为了排除极端异常情况,忽略了发生在轨迹开始或结束3年内的突变点[9]. ...
... [9]. ...
Distinguishing trajectories and drivers of vegetated ecosystems in China's Loess Plateau
1
2024
... 全球范围内,约四分之一的旱地植被经历了显著的突变,同时揭示了气候变异性和人类活动在植被突变中扮演的关键角色[9].中国黄土高原29%的区域发生了突变,温度上升和人口增长对植被突变有显著贡献[10].这表明干旱区植被突变的驱动机制在全球和区域尺度并不一致,因此,西北干旱半干旱区植被突变是一个较为迫切的科学问题.同时,干旱梯度下的植被突变特征和驱动机制尚未得到充分探讨. ...
Approximating XGBoost with an interpretable decision tree
1
2021
... 在识别植被突变时,传统的单一突变检测方法通常难以全面捕捉植被变化的复杂性.为此,本研究采用了基于阈值回归模型的多模型轨迹诊断方法.尽管机器学习方法在植被突变分析中应用广泛,但其解释能力有限[11].相比之下,偏最小二乘结构方程模型(Partial Least Squares Structural Equation Modeling, PLS-SEM)可以提供更高的解释性,能够深入探讨多变量间的因果关系,并揭示单一或多因素共同驱动植被突变的机制[12].在植被突变研究中,增强型植被指数(Enhanced Vegetation Index, EVI)相比归一化差异植被指数(Normalized Difference Vegetation Index, NDVI)和核归一化差异植被指数(Kernel Normalized Difference Vegetation Index, kNDVI)具有明显优势,能够有效减少大气散射和土壤背景的干扰[13].因此,本研究结合多模型轨迹诊断方法和PLS-SEM,利用EVI数据,揭示西北干旱半干旱区及不同干旱梯度下的植被突变特征及其驱动因素. ...
1
2022
... 在识别植被突变时,传统的单一突变检测方法通常难以全面捕捉植被变化的复杂性.为此,本研究采用了基于阈值回归模型的多模型轨迹诊断方法.尽管机器学习方法在植被突变分析中应用广泛,但其解释能力有限[11].相比之下,偏最小二乘结构方程模型(Partial Least Squares Structural Equation Modeling, PLS-SEM)可以提供更高的解释性,能够深入探讨多变量间的因果关系,并揭示单一或多因素共同驱动植被突变的机制[12].在植被突变研究中,增强型植被指数(Enhanced Vegetation Index, EVI)相比归一化差异植被指数(Normalized Difference Vegetation Index, NDVI)和核归一化差异植被指数(Kernel Normalized Difference Vegetation Index, kNDVI)具有明显优势,能够有效减少大气散射和土壤背景的干扰[13].因此,本研究结合多模型轨迹诊断方法和PLS-SEM,利用EVI数据,揭示西北干旱半干旱区及不同干旱梯度下的植被突变特征及其驱动因素. ...
Overview of the radiometric and biophysical performance of the MODIS vegetation indices
2
2002
... 在识别植被突变时,传统的单一突变检测方法通常难以全面捕捉植被变化的复杂性.为此,本研究采用了基于阈值回归模型的多模型轨迹诊断方法.尽管机器学习方法在植被突变分析中应用广泛,但其解释能力有限[11].相比之下,偏最小二乘结构方程模型(Partial Least Squares Structural Equation Modeling, PLS-SEM)可以提供更高的解释性,能够深入探讨多变量间的因果关系,并揭示单一或多因素共同驱动植被突变的机制[12].在植被突变研究中,增强型植被指数(Enhanced Vegetation Index, EVI)相比归一化差异植被指数(Normalized Difference Vegetation Index, NDVI)和核归一化差异植被指数(Kernel Normalized Difference Vegetation Index, kNDVI)具有明显优势,能够有效减少大气散射和土壤背景的干扰[13].因此,本研究结合多模型轨迹诊断方法和PLS-SEM,利用EVI数据,揭示西北干旱半干旱区及不同干旱梯度下的植被突变特征及其驱动因素. ...
... 增强型植被指数(Enhanced Vegetation Index,EVI)是反映旱区植被覆盖程度的重要指标[13].源于MOD13A3数据集的逐月EVI栅格数据(https://www.earthdata.nasa.gov)[18].该数据集从2000年2月至2020年12月,分辨率为1 km,通过最大值合成法将月数据合成为年栅格数据. ...
Spatiotemporal changes of land desertification sensitivity in northwest China from 2000 to 2017
1
2021
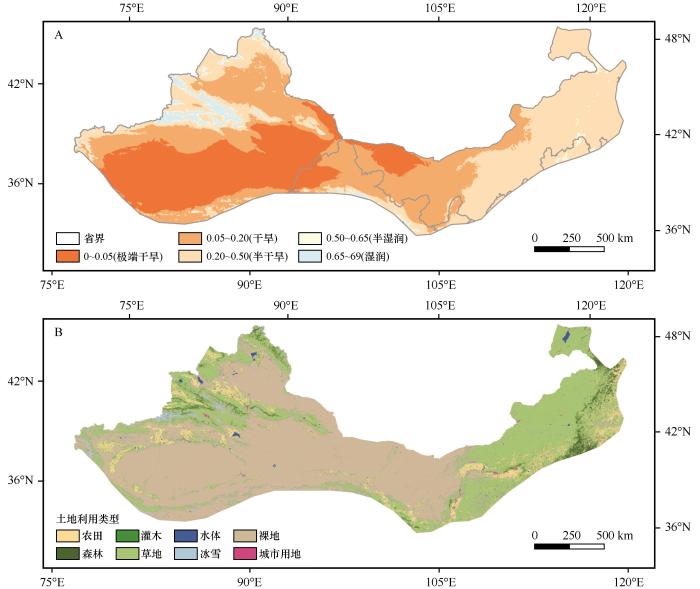
... 西北干旱半干旱区位于32°—47°N、65°—130°E.该地区主要为温带大陆性气候,降水稀少,平均年降水量为200~400 mm,自东向西逐渐减少;年均温为5~15 ℃,该区蒸发作用强烈,平均年蒸发量800~3 200 mm,显著高于降雨量,导致水资源严重短缺[14],自东向西表现为显著的干旱梯度分布.依据联合国环境规划署(UNEP)的气候分类办法[15],采用干旱指数(AI=Pre/PET,其中Pre代表年降水量,PET代表年潜在蒸散发量)可将该区域细分为极端干旱、干旱、半干旱和半湿润4个亚区(图1A)[16].受气候条件制约,区域植被覆盖度普遍较低.2020年土地利用数据显示,草地(33.53%)和裸地(52.89%)是主要土地利用类型,其次为农田(6.95%)、森林(2.39%)、灌木(0.90%)、水体及冰雪用地(2.95%)和城市用地(0.39%,图1B).该区域的经济活动主要依赖于农业和畜牧业[17]. ...
Version 3 of the global aridity index and potential evapotranspiration database
2
2022
... 西北干旱半干旱区位于32°—47°N、65°—130°E.该地区主要为温带大陆性气候,降水稀少,平均年降水量为200~400 mm,自东向西逐渐减少;年均温为5~15 ℃,该区蒸发作用强烈,平均年蒸发量800~3 200 mm,显著高于降雨量,导致水资源严重短缺[14],自东向西表现为显著的干旱梯度分布.依据联合国环境规划署(UNEP)的气候分类办法[15],采用干旱指数(AI=Pre/PET,其中Pre代表年降水量,PET代表年潜在蒸散发量)可将该区域细分为极端干旱、干旱、半干旱和半湿润4个亚区(图1A)[16].受气候条件制约,区域植被覆盖度普遍较低.2020年土地利用数据显示,草地(33.53%)和裸地(52.89%)是主要土地利用类型,其次为农田(6.95%)、森林(2.39%)、灌木(0.90%)、水体及冰雪用地(2.95%)和城市用地(0.39%,图1B).该区域的经济活动主要依赖于农业和畜牧业[17]. ...
... 干燥度指数(Aridity Index,AI)定义为年降水量(Precipitation,P)与年潜在蒸散发量(Potential Evapotranspiration,PET)之比[15].中国逐年干燥度指数来源于国家青藏高原科学数据中心的中国1 km干燥度数据集(1901—2022)[21]. ...
农业干旱业务化监测研究进展与展望
1
2023
... 西北干旱半干旱区位于32°—47°N、65°—130°E.该地区主要为温带大陆性气候,降水稀少,平均年降水量为200~400 mm,自东向西逐渐减少;年均温为5~15 ℃,该区蒸发作用强烈,平均年蒸发量800~3 200 mm,显著高于降雨量,导致水资源严重短缺[14],自东向西表现为显著的干旱梯度分布.依据联合国环境规划署(UNEP)的气候分类办法[15],采用干旱指数(AI=Pre/PET,其中Pre代表年降水量,PET代表年潜在蒸散发量)可将该区域细分为极端干旱、干旱、半干旱和半湿润4个亚区(图1A)[16].受气候条件制约,区域植被覆盖度普遍较低.2020年土地利用数据显示,草地(33.53%)和裸地(52.89%)是主要土地利用类型,其次为农田(6.95%)、森林(2.39%)、灌木(0.90%)、水体及冰雪用地(2.95%)和城市用地(0.39%,图1B).该区域的经济活动主要依赖于农业和畜牧业[17]. ...
1
2019
... 西北干旱半干旱区位于32°—47°N、65°—130°E.该地区主要为温带大陆性气候,降水稀少,平均年降水量为200~400 mm,自东向西逐渐减少;年均温为5~15 ℃,该区蒸发作用强烈,平均年蒸发量800~3 200 mm,显著高于降雨量,导致水资源严重短缺[14],自东向西表现为显著的干旱梯度分布.依据联合国环境规划署(UNEP)的气候分类办法[15],采用干旱指数(AI=Pre/PET,其中Pre代表年降水量,PET代表年潜在蒸散发量)可将该区域细分为极端干旱、干旱、半干旱和半湿润4个亚区(图1A)[16].受气候条件制约,区域植被覆盖度普遍较低.2020年土地利用数据显示,草地(33.53%)和裸地(52.89%)是主要土地利用类型,其次为农田(6.95%)、森林(2.39%)、灌木(0.90%)、水体及冰雪用地(2.95%)和城市用地(0.39%,图1B).该区域的经济活动主要依赖于农业和畜牧业[17]. ...
MODIS/Terra Vegetation Indices Monthly L3 Global 1km SIN Grid V006,NASA EOSDIS Land Processes DAAC
1
2015
... 增强型植被指数(Enhanced Vegetation Index,EVI)是反映旱区植被覆盖程度的重要指标[13].源于MOD13A3数据集的逐月EVI栅格数据(https://www.earthdata.nasa.gov)[18].该数据集从2000年2月至2020年12月,分辨率为1 km,通过最大值合成法将月数据合成为年栅格数据. ...
中国1 km分辨率逐月平均气温数据集(1901-2023)
1
2024
... 降水数据和温度数据来源于国家青藏高原科学数据中心的中国1 km分辨率数据集(https://data.tpdc.ac.cn/zh-hans/data)(1901—2021)[19-20]. ...
中国1 km分辨率逐月降水量数据集(1901-2023)
1
2024
... 降水数据和温度数据来源于国家青藏高原科学数据中心的中国1 km分辨率数据集(https://data.tpdc.ac.cn/zh-hans/data)(1901—2021)[19-20]. ...
中国1 km逐年干燥度数据集(1901-2023)
1
2024
... 干燥度指数(Aridity Index,AI)定义为年降水量(Precipitation,P)与年潜在蒸散发量(Potential Evapotranspiration,PET)之比[15].中国逐年干燥度指数来源于国家青藏高原科学数据中心的中国1 km干燥度数据集(1901—2022)[21]. ...
A global record of annual terrestrial human footprint dataset from 2000 to 2018
1
2022
... 人口足迹是一个综合性指标,用于衡量人类对自然资源的消耗和对环境的影响.源于全球陆地人口足迹年度数据集(2000—2020年)(https://www.x-mol.com/groups/li_xuecao/news/48145)[22],该数据反映了人类在8个方面对环境的压力,包括建筑环境、人口密度、夜间灯光、农田、牧场、公路、铁路、通航水道等变量[23],空间分辨率为1 km. ...
The human footprint and the last of the wild:the human footprint is a global map of human influence on the land surface,which suggests that human beings are stewards of nature,whether we like it or not
1
2002
... 人口足迹是一个综合性指标,用于衡量人类对自然资源的消耗和对环境的影响.源于全球陆地人口足迹年度数据集(2000—2020年)(https://www.x-mol.com/groups/li_xuecao/news/48145)[22],该数据反映了人类在8个方面对环境的压力,包括建筑环境、人口密度、夜间灯光、农田、牧场、公路、铁路、通航水道等变量[23],空间分辨率为1 km. ...
The 30?m annual land cover dataset and its dynamics in China from 1990 to 2019
1
2021
... 土地利用数据基于中国年度土地覆盖数据集(2000—2020年),分辨率为30 m,分类精度约为80%[24],采用优势聚类法将数据重采样至1 km分辨率. ...
Catastrophic shifts in ecosystems
1
2001
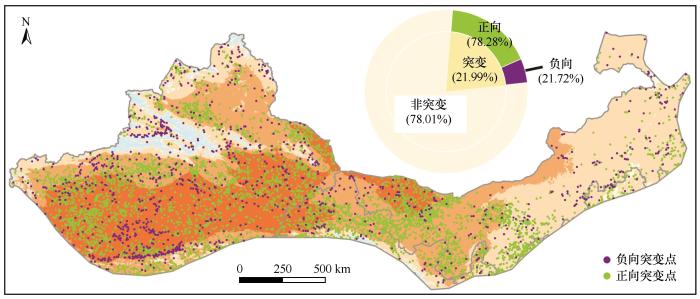
... 植被突变通常由于环境压力或者环境长期持续的变化达到了某个阈值,与非突变的渐进式响应不同,突变具有不连续性和阈值效应[25].在研究区随机选择25 000个样点,并逐点提取2000—2020年的植被指数序列.为保证样点的空间分布均衡,设定了最小距离阈值,避免样点在区域内聚集.其中时间序列完整的样点为24 164个,去除湿润区后最终有效的样点为24 079个.通过多模型轨迹诊断方法中的阶跃模型将植被指数的变化分为突变和非突变从而识别出突变区域[26].根据阶跃模型的阈值参数,将植被突变分为正向突变(阈值为正数)和负向突变(阈值为负数). ...
A new look at the statistical model identification
2
1974
... 植被突变通常由于环境压力或者环境长期持续的变化达到了某个阈值,与非突变的渐进式响应不同,突变具有不连续性和阈值效应[25].在研究区随机选择25 000个样点,并逐点提取2000—2020年的植被指数序列.为保证样点的空间分布均衡,设定了最小距离阈值,避免样点在区域内聚集.其中时间序列完整的样点为24 164个,去除湿润区后最终有效的样点为24 079个.通过多模型轨迹诊断方法中的阶跃模型将植被指数的变化分为突变和非突变从而识别出突变区域[26].根据阶跃模型的阈值参数,将植被突变分为正向突变(阈值为正数)和负向突变(阈值为负数). ...
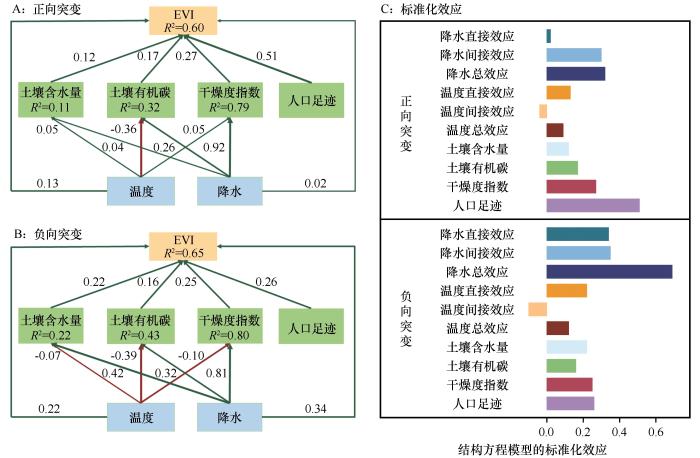
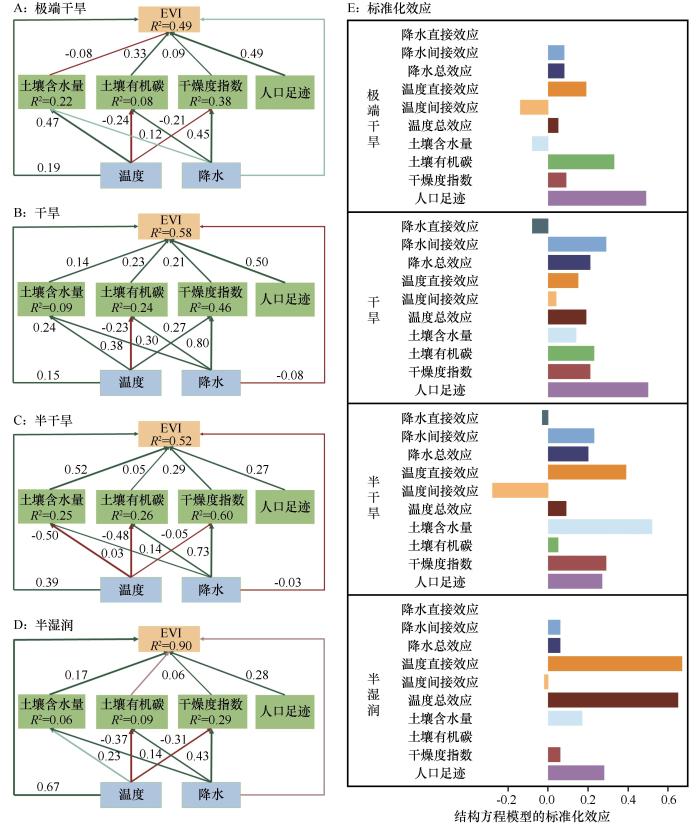
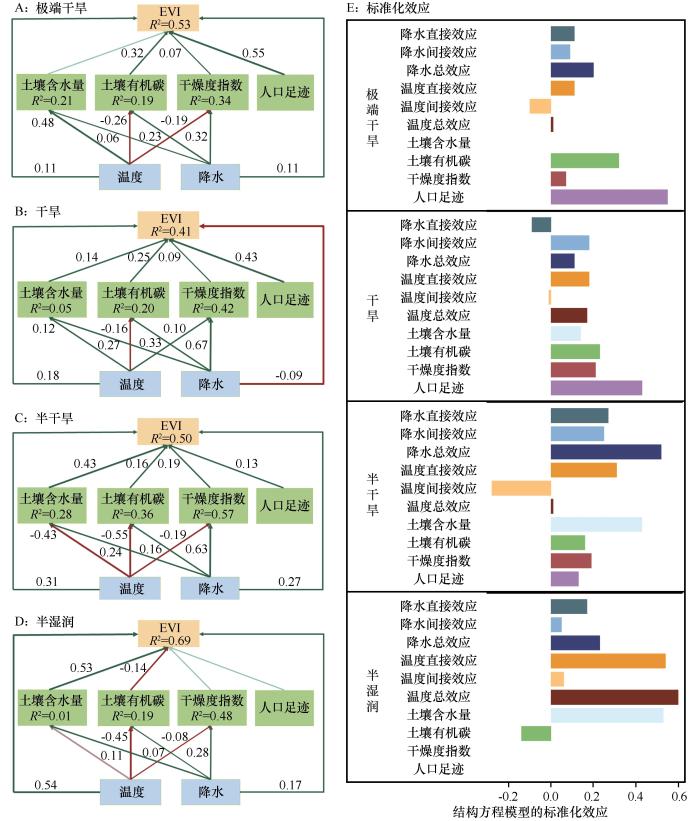
... 本研究采用PLS-SEM分析植被生态系统中多变量之间的因果关系,以及直接和间接关系,并计算其直接、间接和总效应[27].基于当前全球旱地突变的研究[26],本研究选择了土壤因素(如土壤含水量和有机碳)、气候因素(包括降水、温度和干燥度指数),以及人口足迹等6个因子作为植被突变的影响因子.在此基础上,构建了一个PLS-SEM模型框架,并将时间作为随机效应纳入其中[28]. ...
Vegetation greening intensified soil drying in some semi-arid and arid areas of the world
1
2020
... 本研究采用PLS-SEM分析植被生态系统中多变量之间的因果关系,以及直接和间接关系,并计算其直接、间接和总效应[27].基于当前全球旱地突变的研究[26],本研究选择了土壤因素(如土壤含水量和有机碳)、气候因素(包括降水、温度和干燥度指数),以及人口足迹等6个因子作为植被突变的影响因子.在此基础上,构建了一个PLS-SEM模型框架,并将时间作为随机效应纳入其中[28]. ...
Functional traits explain the consistent resistance of biodiversity to plant invasion under nitrogen enrichment
1
2021
... 本研究采用PLS-SEM分析植被生态系统中多变量之间的因果关系,以及直接和间接关系,并计算其直接、间接和总效应[27].基于当前全球旱地突变的研究[26],本研究选择了土壤因素(如土壤含水量和有机碳)、气候因素(包括降水、温度和干燥度指数),以及人口足迹等6个因子作为植被突变的影响因子.在此基础上,构建了一个PLS-SEM模型框架,并将时间作为随机效应纳入其中[28]. ...
Linear and nonlinear characteristics of long-term NDVI using trend analysis:a case study of Lancang-Mekong River Basin
1
2022
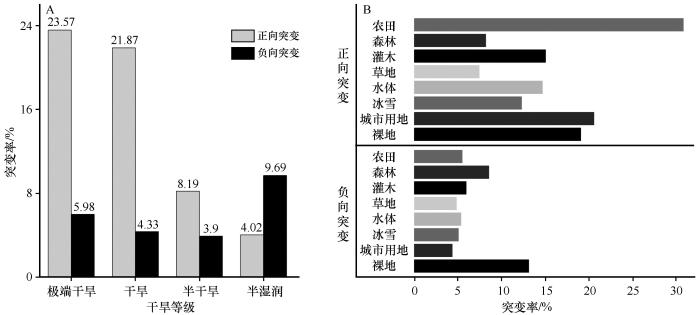
... 自2000年以来,全球旱地生态系统中植被突变现象越来越普遍[29].在西北干旱半干旱区,约21.99%的植被经历了突变,这一比例与全球旱地的突变水平(23.60%)基本持平,说明了西北干旱半干旱区与全球可能经历着相同的情景.该区域植被突变的主要影响因子是人类活动与水分可利用性,这与李卓忆等[30]所指出的植被变化影响因子一致.正向突变则约占突变的3/4,虽然突变可能暗示生态系统存在某种不稳定性,但也反映了整体生态环境的改善.正向突变多发生在人类活动较密集的区域,主要土地利用类型为耕地和草地,这与人类在该地区的水资源分配[31]和草原养育工程[32]有关.然而,耕地对水资源的优先使用可能会占用生态用水,加剧植被的水分压力[33],导致旱地生态系统发生负向突变,因此,科学的水资源管理对该地区的生态环境保护至关重要.负向突变只占1/4,但识别出负向突变的生态退化区,并采取相应的措施缓解非常关键.半湿润区的负向突变率最高,这可能是快速的增温导致的,增温导致蒸散发增加限制植物生长[34].在干旱亚类交界区域也分布着负向突变点,主要原因是降水不稳定以及人类的扰动[35]. ...
中国北方干旱半干旱区植被对气候变化和人类活动的响应
1
2024
... 自2000年以来,全球旱地生态系统中植被突变现象越来越普遍[29].在西北干旱半干旱区,约21.99%的植被经历了突变,这一比例与全球旱地的突变水平(23.60%)基本持平,说明了西北干旱半干旱区与全球可能经历着相同的情景.该区域植被突变的主要影响因子是人类活动与水分可利用性,这与李卓忆等[30]所指出的植被变化影响因子一致.正向突变则约占突变的3/4,虽然突变可能暗示生态系统存在某种不稳定性,但也反映了整体生态环境的改善.正向突变多发生在人类活动较密集的区域,主要土地利用类型为耕地和草地,这与人类在该地区的水资源分配[31]和草原养育工程[32]有关.然而,耕地对水资源的优先使用可能会占用生态用水,加剧植被的水分压力[33],导致旱地生态系统发生负向突变,因此,科学的水资源管理对该地区的生态环境保护至关重要.负向突变只占1/4,但识别出负向突变的生态退化区,并采取相应的措施缓解非常关键.半湿润区的负向突变率最高,这可能是快速的增温导致的,增温导致蒸散发增加限制植物生长[34].在干旱亚类交界区域也分布着负向突变点,主要原因是降水不稳定以及人类的扰动[35]. ...
西北地区水资源合理开发利用与生态环境保护研究
1
2001
... 自2000年以来,全球旱地生态系统中植被突变现象越来越普遍[29].在西北干旱半干旱区,约21.99%的植被经历了突变,这一比例与全球旱地的突变水平(23.60%)基本持平,说明了西北干旱半干旱区与全球可能经历着相同的情景.该区域植被突变的主要影响因子是人类活动与水分可利用性,这与李卓忆等[30]所指出的植被变化影响因子一致.正向突变则约占突变的3/4,虽然突变可能暗示生态系统存在某种不稳定性,但也反映了整体生态环境的改善.正向突变多发生在人类活动较密集的区域,主要土地利用类型为耕地和草地,这与人类在该地区的水资源分配[31]和草原养育工程[32]有关.然而,耕地对水资源的优先使用可能会占用生态用水,加剧植被的水分压力[33],导致旱地生态系统发生负向突变,因此,科学的水资源管理对该地区的生态环境保护至关重要.负向突变只占1/4,但识别出负向突变的生态退化区,并采取相应的措施缓解非常关键.半湿润区的负向突变率最高,这可能是快速的增温导致的,增温导致蒸散发增加限制植物生长[34].在干旱亚类交界区域也分布着负向突变点,主要原因是降水不稳定以及人类的扰动[35]. ...
Grassland ecological management and utilization for sustainability
1
2024
... 自2000年以来,全球旱地生态系统中植被突变现象越来越普遍[29].在西北干旱半干旱区,约21.99%的植被经历了突变,这一比例与全球旱地的突变水平(23.60%)基本持平,说明了西北干旱半干旱区与全球可能经历着相同的情景.该区域植被突变的主要影响因子是人类活动与水分可利用性,这与李卓忆等[30]所指出的植被变化影响因子一致.正向突变则约占突变的3/4,虽然突变可能暗示生态系统存在某种不稳定性,但也反映了整体生态环境的改善.正向突变多发生在人类活动较密集的区域,主要土地利用类型为耕地和草地,这与人类在该地区的水资源分配[31]和草原养育工程[32]有关.然而,耕地对水资源的优先使用可能会占用生态用水,加剧植被的水分压力[33],导致旱地生态系统发生负向突变,因此,科学的水资源管理对该地区的生态环境保护至关重要.负向突变只占1/4,但识别出负向突变的生态退化区,并采取相应的措施缓解非常关键.半湿润区的负向突变率最高,这可能是快速的增温导致的,增温导致蒸散发增加限制植物生长[34].在干旱亚类交界区域也分布着负向突变点,主要原因是降水不稳定以及人类的扰动[35]. ...
Global analysis of urban surface water supply vulnerability
1
2014
... 自2000年以来,全球旱地生态系统中植被突变现象越来越普遍[29].在西北干旱半干旱区,约21.99%的植被经历了突变,这一比例与全球旱地的突变水平(23.60%)基本持平,说明了西北干旱半干旱区与全球可能经历着相同的情景.该区域植被突变的主要影响因子是人类活动与水分可利用性,这与李卓忆等[30]所指出的植被变化影响因子一致.正向突变则约占突变的3/4,虽然突变可能暗示生态系统存在某种不稳定性,但也反映了整体生态环境的改善.正向突变多发生在人类活动较密集的区域,主要土地利用类型为耕地和草地,这与人类在该地区的水资源分配[31]和草原养育工程[32]有关.然而,耕地对水资源的优先使用可能会占用生态用水,加剧植被的水分压力[33],导致旱地生态系统发生负向突变,因此,科学的水资源管理对该地区的生态环境保护至关重要.负向突变只占1/4,但识别出负向突变的生态退化区,并采取相应的措施缓解非常关键.半湿润区的负向突变率最高,这可能是快速的增温导致的,增温导致蒸散发增加限制植物生长[34].在干旱亚类交界区域也分布着负向突变点,主要原因是降水不稳定以及人类的扰动[35]. ...
The impacts of climate change on water resources and agriculture in China
1
2010
... 自2000年以来,全球旱地生态系统中植被突变现象越来越普遍[29].在西北干旱半干旱区,约21.99%的植被经历了突变,这一比例与全球旱地的突变水平(23.60%)基本持平,说明了西北干旱半干旱区与全球可能经历着相同的情景.该区域植被突变的主要影响因子是人类活动与水分可利用性,这与李卓忆等[30]所指出的植被变化影响因子一致.正向突变则约占突变的3/4,虽然突变可能暗示生态系统存在某种不稳定性,但也反映了整体生态环境的改善.正向突变多发生在人类活动较密集的区域,主要土地利用类型为耕地和草地,这与人类在该地区的水资源分配[31]和草原养育工程[32]有关.然而,耕地对水资源的优先使用可能会占用生态用水,加剧植被的水分压力[33],导致旱地生态系统发生负向突变,因此,科学的水资源管理对该地区的生态环境保护至关重要.负向突变只占1/4,但识别出负向突变的生态退化区,并采取相应的措施缓解非常关键.半湿润区的负向突变率最高,这可能是快速的增温导致的,增温导致蒸散发增加限制植物生长[34].在干旱亚类交界区域也分布着负向突变点,主要原因是降水不稳定以及人类的扰动[35]. ...
Accelerated dryland expansion under climate change
1
2016
... 自2000年以来,全球旱地生态系统中植被突变现象越来越普遍[29].在西北干旱半干旱区,约21.99%的植被经历了突变,这一比例与全球旱地的突变水平(23.60%)基本持平,说明了西北干旱半干旱区与全球可能经历着相同的情景.该区域植被突变的主要影响因子是人类活动与水分可利用性,这与李卓忆等[30]所指出的植被变化影响因子一致.正向突变则约占突变的3/4,虽然突变可能暗示生态系统存在某种不稳定性,但也反映了整体生态环境的改善.正向突变多发生在人类活动较密集的区域,主要土地利用类型为耕地和草地,这与人类在该地区的水资源分配[31]和草原养育工程[32]有关.然而,耕地对水资源的优先使用可能会占用生态用水,加剧植被的水分压力[33],导致旱地生态系统发生负向突变,因此,科学的水资源管理对该地区的生态环境保护至关重要.负向突变只占1/4,但识别出负向突变的生态退化区,并采取相应的措施缓解非常关键.半湿润区的负向突变率最高,这可能是快速的增温导致的,增温导致蒸散发增加限制植物生长[34].在干旱亚类交界区域也分布着负向突变点,主要原因是降水不稳定以及人类的扰动[35]. ...
Drought-induced reduction in global terrestrial net primary production from 2000 through 2009
1
2010
... 4个干旱亚区植被突变的主要驱动因子各不相同,随着干旱梯度增加,主要因子分别是温度、降水和人类活动,这反映了植被突变机制的复杂性.半湿润区主要分布在东北部,该地对水分变化较为敏感,增温引起蒸散发增加导致水分胁迫加剧,限制植被生长[36].半干旱区植被主要受降水的影响[37],该地降水量减少且不稳定[38],加剧了对降水的依赖性.干旱和极端干旱区域由于气候环境恶劣,自然植被稀少且对干旱条件具有较强的适应性[39],植被的变化主要受人类活动控制[40],如放牧、草地围栏养育以及灌溉耕地等.人口足迹和降水是全区范围内植被突变的主要驱动因子,但随着干旱梯度的增加,人口足迹的贡献度显著增加,降水的贡献呈先增加后减少趋势.因此,在全球变暖背景下,增温对半湿润区部分地区的植被可能存在负向突变的影响,而半干旱区需要采取应对水分胁迫的措施,在干旱与极端干旱区域需要科学合理安排人类活动的项目与强度,促进可持续发展. ...
半干旱区植被覆盖时空变化特征及其对气候变化的响应:以锡林郭勒盟为例
1
2024
... 4个干旱亚区植被突变的主要驱动因子各不相同,随着干旱梯度增加,主要因子分别是温度、降水和人类活动,这反映了植被突变机制的复杂性.半湿润区主要分布在东北部,该地对水分变化较为敏感,增温引起蒸散发增加导致水分胁迫加剧,限制植被生长[36].半干旱区植被主要受降水的影响[37],该地降水量减少且不稳定[38],加剧了对降水的依赖性.干旱和极端干旱区域由于气候环境恶劣,自然植被稀少且对干旱条件具有较强的适应性[39],植被的变化主要受人类活动控制[40],如放牧、草地围栏养育以及灌溉耕地等.人口足迹和降水是全区范围内植被突变的主要驱动因子,但随着干旱梯度的增加,人口足迹的贡献度显著增加,降水的贡献呈先增加后减少趋势.因此,在全球变暖背景下,增温对半湿润区部分地区的植被可能存在负向突变的影响,而半干旱区需要采取应对水分胁迫的措施,在干旱与极端干旱区域需要科学合理安排人类活动的项目与强度,促进可持续发展. ...
我国干旱半干旱区气候变化特征及其对干湿波动的影响
1
2023
... 4个干旱亚区植被突变的主要驱动因子各不相同,随着干旱梯度增加,主要因子分别是温度、降水和人类活动,这反映了植被突变机制的复杂性.半湿润区主要分布在东北部,该地对水分变化较为敏感,增温引起蒸散发增加导致水分胁迫加剧,限制植被生长[36].半干旱区植被主要受降水的影响[37],该地降水量减少且不稳定[38],加剧了对降水的依赖性.干旱和极端干旱区域由于气候环境恶劣,自然植被稀少且对干旱条件具有较强的适应性[39],植被的变化主要受人类活动控制[40],如放牧、草地围栏养育以及灌溉耕地等.人口足迹和降水是全区范围内植被突变的主要驱动因子,但随着干旱梯度的增加,人口足迹的贡献度显著增加,降水的贡献呈先增加后减少趋势.因此,在全球变暖背景下,增温对半湿润区部分地区的植被可能存在负向突变的影响,而半干旱区需要采取应对水分胁迫的措施,在干旱与极端干旱区域需要科学合理安排人类活动的项目与强度,促进可持续发展. ...
植物对干旱胁迫的生理生态响应及其研究进展
1
2009
... 4个干旱亚区植被突变的主要驱动因子各不相同,随着干旱梯度增加,主要因子分别是温度、降水和人类活动,这反映了植被突变机制的复杂性.半湿润区主要分布在东北部,该地对水分变化较为敏感,增温引起蒸散发增加导致水分胁迫加剧,限制植被生长[36].半干旱区植被主要受降水的影响[37],该地降水量减少且不稳定[38],加剧了对降水的依赖性.干旱和极端干旱区域由于气候环境恶劣,自然植被稀少且对干旱条件具有较强的适应性[39],植被的变化主要受人类活动控制[40],如放牧、草地围栏养育以及灌溉耕地等.人口足迹和降水是全区范围内植被突变的主要驱动因子,但随着干旱梯度的增加,人口足迹的贡献度显著增加,降水的贡献呈先增加后减少趋势.因此,在全球变暖背景下,增温对半湿润区部分地区的植被可能存在负向突变的影响,而半干旱区需要采取应对水分胁迫的措施,在干旱与极端干旱区域需要科学合理安排人类活动的项目与强度,促进可持续发展. ...
Spatiotemporal characteristics in ecosystem service value and its interaction with human activities in Xinjiang,China
1
2020
... 4个干旱亚区植被突变的主要驱动因子各不相同,随着干旱梯度增加,主要因子分别是温度、降水和人类活动,这反映了植被突变机制的复杂性.半湿润区主要分布在东北部,该地对水分变化较为敏感,增温引起蒸散发增加导致水分胁迫加剧,限制植被生长[36].半干旱区植被主要受降水的影响[37],该地降水量减少且不稳定[38],加剧了对降水的依赖性.干旱和极端干旱区域由于气候环境恶劣,自然植被稀少且对干旱条件具有较强的适应性[39],植被的变化主要受人类活动控制[40],如放牧、草地围栏养育以及灌溉耕地等.人口足迹和降水是全区范围内植被突变的主要驱动因子,但随着干旱梯度的增加,人口足迹的贡献度显著增加,降水的贡献呈先增加后减少趋势.因此,在全球变暖背景下,增温对半湿润区部分地区的植被可能存在负向突变的影响,而半干旱区需要采取应对水分胁迫的措施,在干旱与极端干旱区域需要科学合理安排人类活动的项目与强度,促进可持续发展. ...




 甘公网安备 62010202000688号
甘公网安备 62010202000688号
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}