


0 引言
种群是构成群落的基本单位,植物个体在水平空间内不能移动,植物种群内个体在空间内的分布状态与该种群的各种生态学过程有着非常密切的联系[4]。空间格局的形成过程如生长、死亡、竞争等常常与种群或群落的结构、动态、种内种间关系、种群与生境的关系等方面联系在一起。可以通过量化并预测斑块的特征来推理种群或者群落的形成过程以及与环境因子间的相互关系[5-7]。种群分布格局指在种群水平上种群内个体在一定范围内水平空间的分布状况,是种群生物学特性、种内和种间相互关系及环境条件综合作用的结果[8],可以反映出植物种群的生态调控能力和环境可塑性机制,是种群在群落中地位与生存能力的外在表现[3],一般分为均匀分布、聚集分布和随机分布[9]。通过测定和分析种群的分布格局,可以在一个更为细致的层次上认识群落的结构,有助于了解种群的生物学特性及与环境因子相互作用的基本规律,揭示其与空间分布相关联的生态过程。点格局分析法是利用植物个体在水平空间内的坐标数据进行计算分析的方法,目前已被广泛应用于森林[10]、荒漠[11]、草原[12]等植被空间分布及关联性研究中。刘明伟等[13]应用点格局分析法发现,神农架南坡小叶青冈种群的空间格局主要受生境过滤、种子扩散限制和种内种间竞争的影响。王丽萍等[14]应用点格局分析法发现,高黎贡常绿阔叶林10个优势种的分布格局及其关联性随尺度而变化,证明物种的空间分布具有较强的尺度依赖性。许爱云等[15]采用点格局分析中的O-ring函数统计方法发现,蒙古冰草种群中的小基径株丛主要表现为聚集分布,且个体越小聚集程度越高。但目前大部分研究主要关注单一植物种群空间格局变化,很少涉及到不同生境或不同演替阶段多种植物种群空间格局演变过程。
一年生草本植物作为荒漠生态系统中独特的生物类群[16],广泛分布于沙质、砾质以及碱性土壤中[17]。虽然其生命周期短,但繁殖能力强、种子产量高、传播范围广、适应性强[18],在提高荒漠生态系统的生产力、贡献绝大部分物种多样性和维持生态系统功能等方面发挥着重要的作用[19]。人工固沙区深根系灌木的大面积、高密度种植会引起土壤水分失衡,造成土壤干燥化和盐碱化,尤其是盐分离子在土壤表层的大量积聚[20],而一年生草本植物对土壤盐碱化相对更为敏感和脆弱[21]。一年生草本植物成为众多学者的研究对象[22]。但大部分研究仅限于一年生草本植物的生态适应模式[23]、物种多样性[24],以及分布特征[25]等,对一年生草本植物的空间格局研究有限,尤其是环境异质性下空间格局的变化。本研究以河西走廊荒漠绿洲过渡带5、10、20、30、40年梭梭(Haloxylon ammodendron)林下一年生草本植物为研究对象,通过调查梭梭林下一年生草本植物的空间分布格局及其空间特征,分析林下一年生草本植物空间分布格局及其影响因素,确定关键因子,可以为荒漠地区沙漠化治理、沙漠植物资源合理开发利用提供有力的科学依据和参考。
1 材料与方法
1.1 研究区概况
研究区位于黑河流域中游、河西走廊中段的甘肃省张掖市临泽县北部的绿洲边缘(39°20′—39°23′N、100°07′—100°08′E,海拔1 350 m)。该区域为典型的温带大陆性荒漠气候,年均气温7.6 ℃,无霜期约165天,年降水量117 mm,年蒸发量2 390 mm,年日照时数3 045 h,太阳辐射总量高达6.1×105 J·m-2。区域内降水存在明显的季节变化,夏季(7—9月)降水占全年降水总量的65%左右,冬季降雪占全年降水总量的3%;≤5 mm的小降水事件占全年总降水事件的56.1%,并形成46.7%的年降水量(1967—2008年)。区域内风沙活动强烈,集中于3—5月,年均风速3.2 m·s-1,每年风速大于17 m·s-1的天数为15天。土壤类型主要为灰棕漠土以及非地带性的风沙土、草甸土、盐土、沼泽土、灌耕土等,质地较粗,有机质含量极低。区域内地带性植被为中温带荒漠化草原植被,主要由超旱生灌木、半灌木或盐生、旱生肉质半灌木组成,通常伴有一年生草本植物,其中菊科、藜科、禾本科和怪柳科植物较常见[26]。该地区在20世纪70年代中期开始种植以梭梭为主的固沙灌木,经过40年种植固沙植被,风沙活动减弱,绿洲边缘地带植被得到了恢复,流动沙丘逐渐向固定沙丘转变,并在绿洲边缘形成了不同时间序列的梭梭人工林景观。由于流动沙丘在种植梭梭以前,土壤性状基本相同,因此可以利用空间代替时间的研究方法,对不同林龄梭梭群落的变化和土壤理化性质进行对比和动态分析[27]。
1.2 研究方法
于2023年8—10月植物生长旺盛时期,在中国科学院临泽内陆河流域研究站周边(表1),采用空间代替时间的方法,选择5、10、20、30、40年的人工梭梭林样地(图1),兼顾各类样地大小的一致性,从中分别选定地表平坦、群落外貌均匀且具有代表性的5 m×5m的典型样方。采用邻接格子法在每个样方内用1 m×1 m的样方框按从西到东,从南到北的顺序依次取样25次,以样方框的西南角为原点(0,0)、东西方向为X轴、南北方向为Y轴的一角顶点为基点,确定每株一年生草本植物在样方框内的二维坐标,并对每次放置的样方框进行拍摄,最终按照取样时样方框放置的顺序将25个1 m×1 m的图进行拼接,合并成5 m×5 m的样地图(图2),并将所有植株的位置换算成5 m×5 m样方内的信息[28]。与此同时,对样方内的一年生草本植物逐一调查,记录株高、密度、盖度。其中,株高以自然高度计测,密度以1 m2样方内一年生草本植物的个体数量计测,盖度用针刺法测定,然后将一年生草本植物地上部分全部剪下,地下部分全部根挖出,分别置于信封中,带回实验室,置于65 ℃干燥箱内烘干至恒重,称取地上生物量与地下生物量。利用GPS仪测定样方的地理坐标、海拔,用罗盘确定样方方位。
表1 样地具体地理坐标及海拔
Table 1
| 林龄/a | 地理坐标 | 海拔/m |
|---|---|---|
| 5 | 39°23′N,100°12′E | 1 400 |
| 10 | 39°25′N,100°12′E | 1 400 |
| 20 | 39°23′N,100°09′E | 1 390 |
| 30 | 39°22′N,100°08′E | 1 390 |
| 40 | 39°22′N,100°07′E | 1 390 |
图1

图1
不同林龄人工梭梭林
Fig.1
Haloxylon ammodendron plantations of different plantation ages
图2

图2
不同林龄人工梭梭林下一年生草本植物样区
Fig.2
Sample areas of annual herbaceous species of Haloxylon ammodendron plantations with different plantation ages
植被调查的同时,在每个5 m×5 m的样区中,随机抽取5个样方,采用多点混合法按梅花状采集0~20 cm土壤样品,共计25个。剔除样品中植物根系和枯落物等杂质,分别装入已编号的铝盒和自封袋中,带回实验室,用于土壤理化性状的分析。土壤水含量采用烘干法测定,土壤有机碳含量采用重铬酸钾容量法测定,土壤全氮含量凯氏定氮法测定,土壤全磷含量采用钼锑抗比色法测定。pH值采用1∶1的土水比悬液pH计测定,土壤电导率采用1∶5土水比浸提液测定,Ca2+和Na+采用1∶5水溶液-原子吸收分光光度法测定,Cl-采用1∶5水溶液-0.04 mol·L-1硝酸银溶液滴定法测定,HCO
1.3 数据处理
1.3.1 空间分布格局及其关联性分析
采用单变量成对相关函数g(r)分析不同林龄人工梭梭林下一年生草本植物的空间分布格局。g(r)由Ripley'K函数衍生而来,是将一圆环代替Ripley'K函数中的圆,并分析其中所有的个体分布,能够代表某一尺度上点的实际空间分布关系而消除了Ripley'K函数中出现的累加效应[29],能更敏感地判断每个尺度上点的实际分布偏离期望值的程度,比积累K函数更直观、更精确。采用蒙特卡罗(Monte-Carlo)拟合检验,空间尺度设置为0~0.5 m,随机拟合199次,计算上下包迹线(置信区间),得到99%的置信区间。若g(r)在包迹线上面,表示该种群为聚集分布;若g(r)值在包迹线之间,表示种群呈随机分布;若g(r)值在包迹线下面,种群呈均匀分布。采用双变量成对相关函数g12(r)研究同一林龄不同种群之间的空间关联性:若g12(r)值在包迹线上面,表明物种间为正关联;若g12(r)值在包迹线之间,表明物种间为无关联;若g12(r)值在包迹线下面,物种间则为负关联。
K函数和g函数公式如下[30]:
式中:A为样地面积;n为研究个体的总点数;r为两个体间的距离即尺度;uij 是i和j之间的距离,当uij<r,Ir (uij )=1,当uij>r,Ir (uij )=0;wij 为以点i为圆心、uij 为半径的圆面积(周长)在面积A中的比例,其为一个点(植株)可被观察到的概率,在这里引做权重,以便消除边界效应。
式中:n1为第一个种群的个体数;n2为第二个种群的个体数
当g(r)>1,聚集分布;g(r)<1,均匀分布;g(r)=1,随机分布。
当g12(r)>1时,两者正相关;当g12(r)=1时,没有相关性;当g12(r)<1时,两者呈负相关。
1.3.2 冗余分析
冗余分析是多元回归模型的延伸,可以将研究对象与环境因子排列在一定的空间内,排序轴能够反映其中的生态梯度,一次性来解释物种特征、植被分布与环境因子之间的生态关系[31]。本研究中分别选取0~2、2~5、5~10、10~20 cm土层土壤水含量、土壤全氮、土壤电导率、土壤钠离子含量、钙离子含量、碳酸氢根离子含量9个环境因子,选取3种典型一年生草本植物平均盖度、平均高度、地上生物量、地下生物量以及总生物量5个群落特征值为研究对象。在Canoco5软件中进行冗余分析时,对数据进行中心化和标准化,排序轴特征值采用蒙特罗随机模拟499次检验显著性。
本研究采用Excel 2010进行前期数据处理;方差分析由SPSS 19.0软件完成,单因子方差分析(ANOVA)不同林龄人工林对3种典型一年生植物的影响,显著性检验水平为P=0.05;作图由Origin 2017软件完成;利用Programita软件进行3种典型一年生草本植物的空间分布格局及其关联性分析;利用Canoco5软件进行一年生草本植物群落特征与土壤环境因子的冗余分析。
2 结果与分析
2.1 不同林龄梭梭林下3种典型一年生草本植物群落特征
人工固沙梭梭林下一年生草本植物的密度、高度、盖度以及地上和地下生物量、总生物量在不同林龄间存在显著差异(图3)。一年生草本植物的密度为4.2~35 株·m-2,随着林龄的增加呈先减少后显著增加的趋势,10年林龄时为最小值,林龄为40年时达到最大值;一年生草本植物平均高度随着林龄的增加而显著减少,从最初的2.93 cm降低到0.98 cm;盖度呈先增加后减少的趋势,为0.53%~10.82%,40年时为最小值;地上生物量、地下生物量、总生物量变化趋势一致,均随林龄的增加呈先减少后增加的趋势,均在林龄40年时达到最大值,分别为5.8~23.08、0.62~3.75、6.62~26.83 g·m-2。
图3
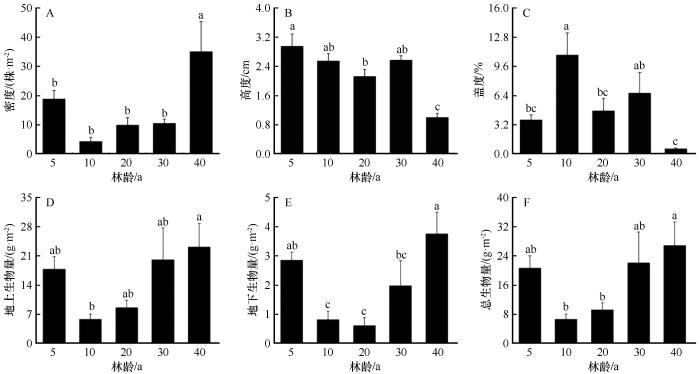
图3
不同林龄梭梭林下3种典型一年生草本植物密度、高度、盖度和生物量
Fig.3
Density, height,coverage,biomass of 3 typical annual herbaceous species of Haloxylon ammodendron plantations with different plantation ages
2.2 不同林龄人工梭梭林下3种典型一年生草本植物空间分布格局
随着人工梭梭林龄的增加,一年生草本层从林龄为5年时的沙蓬-雾冰藜群落发展成为林龄为40年时的雾冰藜-白茎盐生草群落(图4)。利用单变量函数分析不同林龄人工梭梭林下3种一年生草本植物的空间分布格局,结果在不同年限、不同种群间均有所差异(图5),主要呈随机分布或聚集分布。沙蓬随林龄的增加主要从聚集分布向随机分布转变,在人工梭梭林龄为5年时,随尺度增加呈现随机—聚集—随机—聚集—随机—聚集—随机分布,其中只有在0~5.5、21.5~23.5、34.5~37.5、45.5 cm以上呈随机分布,其余尺度均呈聚集分布;林龄为10年时,随尺度的增加呈现随机—聚集—随机—聚集—随机分布,其中在20.5~21.5、27.5~29.5 cm聚集分布,其余尺度均为随机分布;林龄为20年时,主要呈聚集分布。雾冰藜随林龄增加主要呈聚集—随机—聚集分布,在林龄为5年和40年时,分布格局相似,基本都呈聚集分布;林龄为20年时,分布格局随尺度增加在聚集和随机分布之间不断交替,其中在7.5~11.5、19.5~28.5、34.5~46.5、48.5 cm以上随机分布,其余尺度均呈聚集分布。在人工梭梭林龄为30年时,呈现随机—聚集—随机—聚集—随机分布,只有在2.5~6.5、18.5~20.5、25.5 cm尺度范围内呈聚集分布,其余尺度均呈随机分布;白茎盐生草随林龄增加主要呈局部尺度随机到大范围尺度聚集分布,在人工梭梭林龄为30年时,在0~2.5、6.5~8.5,23.5~28.5、30.5~34.4、47.5 cm以上呈随机分布,其余尺度均呈聚集分布。在林龄为40年时,主要呈聚集分布。总体来看,各林龄的人工梭梭林下一年生草本植物均出现明显的均匀分布。
图4
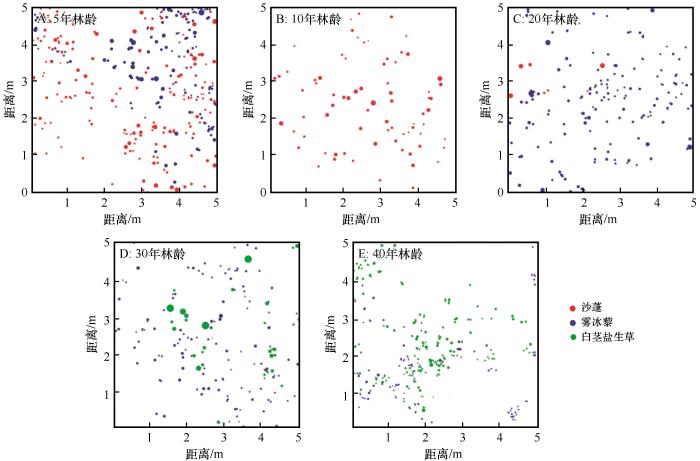
图4
不同林龄梭梭林下3种典型一年生草本植物空间分布
Fig.4
Spatial distribution of 3 typical annual herbaceous species of Haloxylon ammodendron plantations with different plantation ages
图5
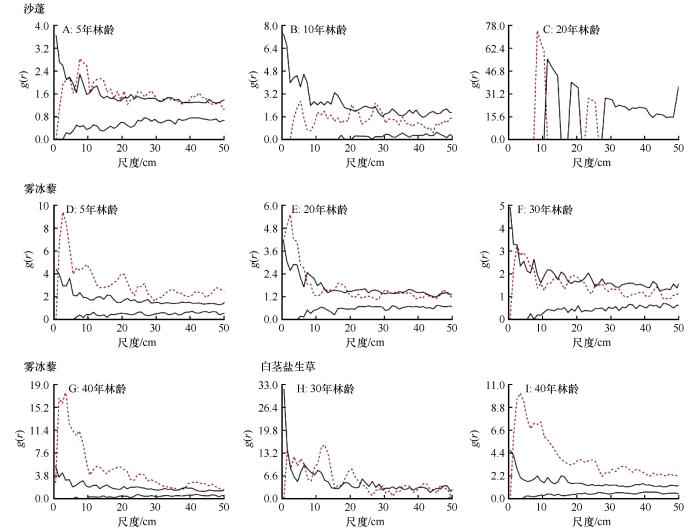
图5
不同林龄梭梭林下3种典型一年生草本植物空间分布格局分析
Fig.5
Spatial patterns of 3 typical annual herbaceous species of Haloxylon ammodendron plantations with different plantation ages
2.3 3种典型一年生草本植物之间的空间关联性
由于不同林龄人工梭梭林下出现的草本植物种群不同,我们对不同种群之间的空间关联性进行分析。在林龄为10年时,只出现了单个种群沙蓬,所以不做分析。林龄为5、20年时,只出现了沙蓬和雾冰藜,并未出现白茎盐生草,而林龄为30、40年时,只出现了雾冰藜和白茎盐生草,所以我们只分析5、20年雾冰藜与沙蓬的空间关联性以及30、40年雾冰藜与白茎盐生草的空间关联性,沙蓬和白茎盐生草之间的关联不予分析。
根据双变量分析结果,不同种类的一年生草本植物之间总体来看没有明显的空间关联性,在大部分范围内表现出相互独立(图6)。只有在局部范围表现为正关联,在所有尺度、所有林龄内均没有负关联。雾冰藜与沙蓬在林龄为5年时,在3.5、17.5~21.5、36.5~38.5 cm正关联,其余尺度均为无关联。在林龄为20年时,所有尺度均为无关联;雾冰藜与白茎盐生草在林龄为30年时,在4.5~6.5 cm正关联,其他尺度内无空间关联。林龄为40年时,除在0~4.5、8.5、12.5、31.5、36.5 cm为无关联以外,其余尺度内均表现为正关联。
图6
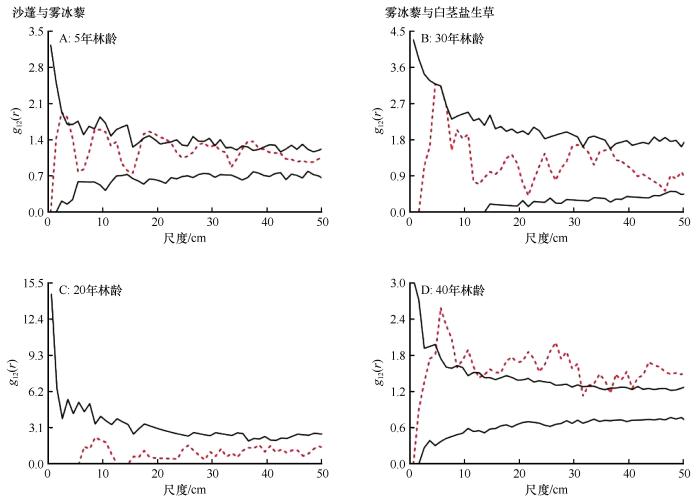
图6
不同林龄梭梭林下3种典型一年生草本植物的种间关联性分析
Fig.6
Interspecific correlation analysis of 3 typical annual herbaceous species of Haloxylon ammodendron plantations with different plantation ages
2.4 不同林龄人工梭梭林下土壤理化性质变化
0~5 cm浅层土壤水含量随人工梭梭林龄的增加而显著增加。土层深度为0~2 cm的土壤水含量从0.22%增加到0.66%,深度为2~5 cm的土壤水含量从0.49%增加到0.70%。深度为5~10 cm土壤水含量呈先增加后减少的趋势,30年生时达到最大值1.70%;深度为10~20 cm的土壤水含量为1.71%~3.90%,林龄为5年时达到最大值。土壤水含量随深度的增加而增加(图7)。
图7

图7
不同林龄梭梭林地不同土层土壤水含量
Fig.7
Soil water content of different soil layers in different plantation ages
随着人工梭梭林龄的增加,土壤有机碳、全氮、氯离子含量没有显著变化(图8)。土壤有机碳含量为1.99~3.08 g·kg-1,呈先增加后减少的趋势,在林龄为10年时达到最大值3.08 g·kg-1,土壤全氮含量为0.20~0.48 g·kg-1,随林龄的增加呈先减少后增加的趋势,林龄为10年时为最小值(0.20 g·kg-1),Cl-含量在不同林龄间无显著性变化,随林龄呈先增加后减少的趋势,含量为162.74~164.5 mg·kg-1。土壤全磷含量在不同林龄间有显著差异,为0.24~0.28 g·kg-1,随林龄的增加呈先减少后显著增加的趋势,在林龄为40年时达到最大值0.34 g·kg-1。土壤pH值为7.22~7.62,均呈碱性,随人工梭梭林龄的增加呈w型变化,林龄为40年时降低到最小值。土壤电导率和钠离子、钙离子含量随林龄的增加呈现相同的变化趋势,含量分别为36.50~274.7 μS·cm-1,18.48~262.73 mg·kg-1,2.59~34.78 mg·kg-1,均在30年前时无显著变化,30年后显著增加,并在40年生时达到最大值。碳酸氢根离子含量随林龄的增加显著增加,含量为40.75~135.3 mg·kg-1。
图8
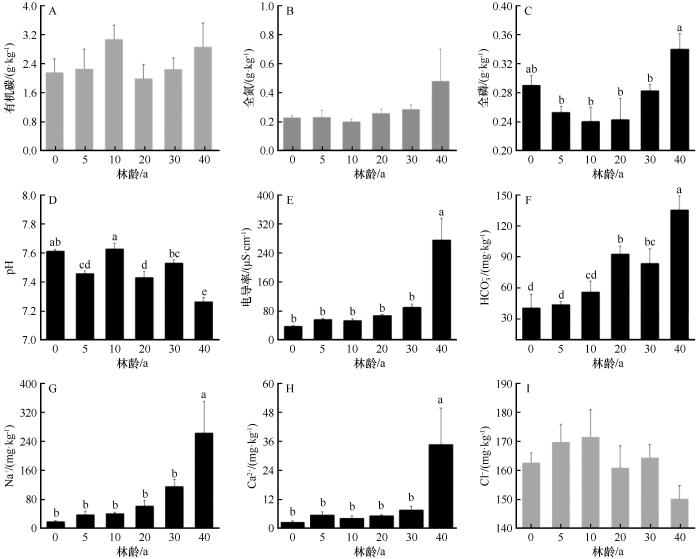
图8
不同林龄梭梭林地土壤养分、pH、电导率以及盐离子含量
Fig.8
Soil nutrients, pH, electrical conductivity, and salt ions of Haloxylon ammodendron plantations with different plantation ages
2.5 一年生草本植物群落和土壤环境因子的关系
RDA结果表明,第一轴和第二轴的特征值分别为0.6302和0.09,一年生草本植物和土壤环境因子的相关系数分别为0.9326和0.6205,累计解释了一年生草本植物群落特征的72.01%(表2)。一年生草本植物密度、地上、地下以及总生物量与0~2、2~5 cm土层土壤水含量、土壤全磷、电导率、碳酸氢根离子、钠离子、钙离子含量正相关,与pH呈负相关;一年生草本植物高度和盖度与pH呈正相关,与0~2、2~5 cm土层土壤水含量、土壤全磷、电导率、碳酸氢根离子、钠离子、钙离子含量负相关(图9)。所选的土壤环境因子对一年生草本植物群落特征的总变异解释率为73.60%,钙离子的解释率为39%,其他依次是2~5 cm土层土壤水含量(13.6%)、钠离子(7.7%)、10~20 cm土层土壤水含量(3.4%)、碳酸氢根离子(3.8%)、土壤全磷(2.9%)、电导率(1.5%)、5~10 cm土层土壤水含量(1.2%)、0~2 cm土层土壤水含量(0.4%)、pH(0.1%)。应用蒙特卡罗检验每个环境因子对群落特征影响的显著性发现,钙离子、2~5 cm土层土壤水含量、钠离子对一年生草本植物群落有显著影响,其余指标无显著影响。
表2 一年生草本植物群落特征与土壤环境因子的冗余分析(RDA)排序
Table 2
| 解释方差 | 环境因子 | 解释率/% | F | P | RDA1 | RDA2 |
|---|---|---|---|---|---|---|
| 解释方差 | 0.6302 | 0.09 | ||||
| 解释方差(累计) | 63.02 | 72.01 | ||||
| 相关系数 | 0.9326 | 0.6205 | ||||
| Ca2+ | 39 | 53.0 | 0.002 | |||
| 2~5 cm土壤水含量 | 13.6 | 18.4 | 0.008 | |||
| Na+ | 7.7 | 10.4 | 0.022 | |||
| 10~20 cm土壤水含量 | 3.4 | 4.7 | 0.202 | |||
| HCO | 3.8 | 5.2 | 0.114 | |||
| 土壤全磷 | 2.9 | 4.0 | 0.190 | |||
| EC | 1.5 | 2.0 | 0.396 | |||
| 5~10 cm土壤水含量 | 1.2 | 1.7 | 0.412 | |||
| 0~2 cm土壤水含量 | 0.4 | 0.5 | 0.806 | |||
| pH | 0.1 | 0.2 | 0.946 |
图9
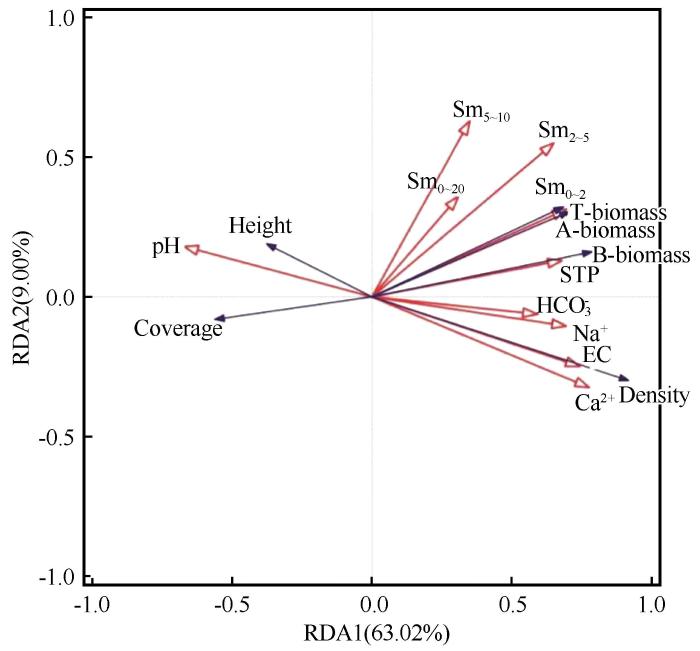
图9
一年生草本植物群落特征与土壤环境因子的冗余分析(RDA)
Fig.9
Redundancy analysis of community characteristics and soil environmental factors of annual herbaceous species
3 讨论
3.1 不同林龄人工固沙林一年生草本植物优势种的变化
随着梭梭林龄的增加,土壤的理化性状得到明显的改善。一年生草本植物群落的总密度、生物量显著增加,林下一年生草本植物由沙蓬-雾冰藜群落逐渐发展到雾冰藜-白茎盐生草群落。沙蓬的密度随着林龄的增加而减小(图10),并在20年之后不再出现,这主要与0~2、10~20 cm土壤水显著正相关(图11、12);雾冰藜的密度先增加后减少,除在林龄为10年时,其他年限均有生长,主要受到钠离子和钙离子含量的制约;白茎盐生草在30年以后出现,密度随着林龄的增加而增加,同时受到0~2 cm土壤水含量以及电导率的影响。沙蓬耐旱,耐贫瘠,生长迅速,是流动沙丘的先锋植物,对沙质土壤的适应性强;雾冰藜散生或群生于荒漠区的沙质和沙砾质土壤,有一定的耐盐性,但耐性不强;白茎盐生草耐盐性强,对生境的适应性广泛,既可以生长在沙质或沙砾质的土壤上,也可以生长在不同程度的盐土荒漠中[32]。实际上,不同林龄人工固沙林下一年生草本植物群落的演替过程是物种对其生境土壤养分和盐分的不断适应以及不同物种相互竞争和替代的过程[33]。
图10
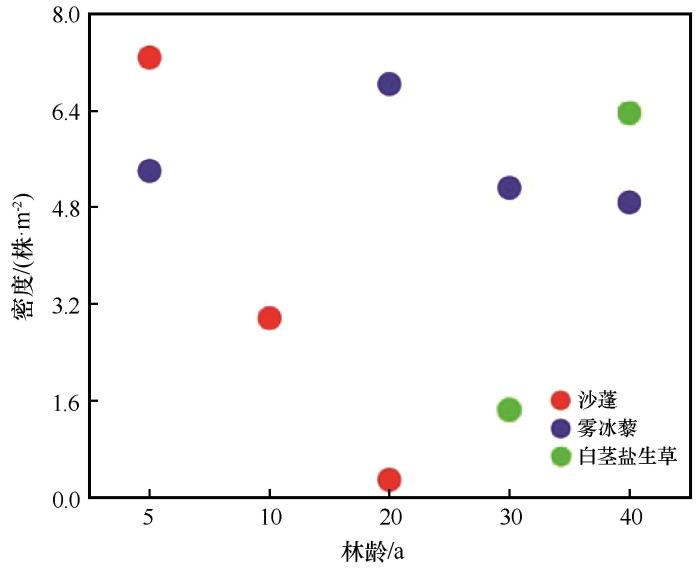
图10
不同林龄梭梭林地3种典型一年生草本植物的密度
Fig.10
Density of 3 typical annual herbaceous species of Haloxylon ammodendron plantations with different plantation ages
图11
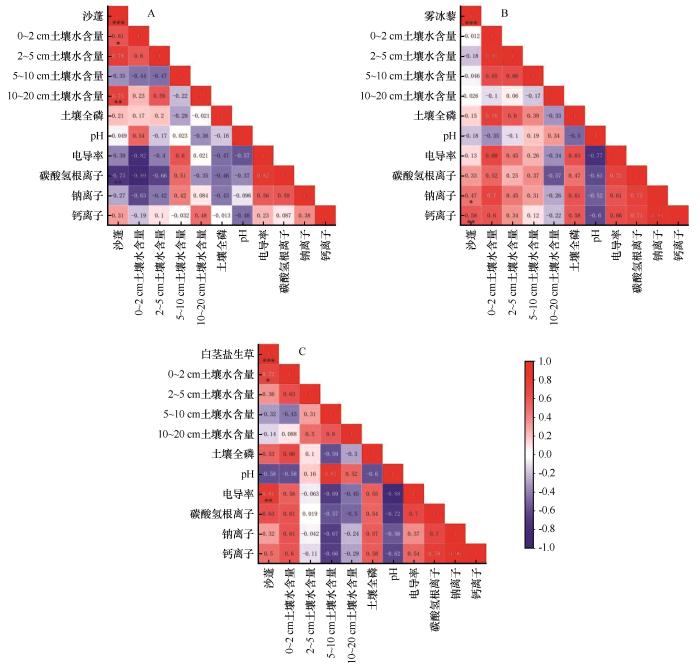
图11
3种典型一年生草本植物密度与土壤环境因子相关性系数的热图
Fig.11
Heat map of correlation coefficients between density of 3 typical annual herbaceous species and soil environmental factors
图12
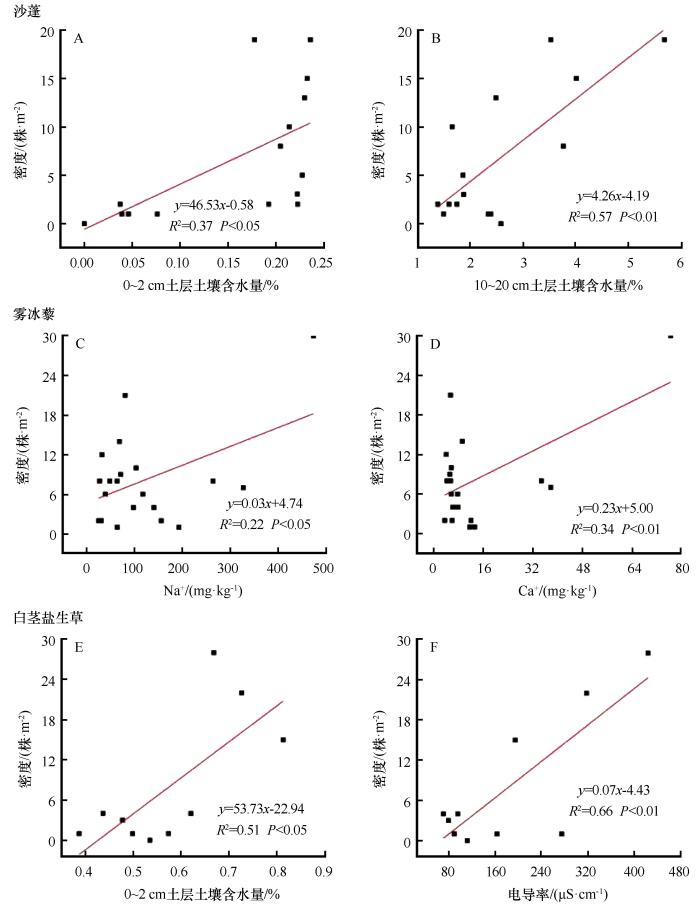
图12
3种典型一年生草本植物密度与土壤环境因子回归分析
Fig.12
Regression analysis of density of 3 typical annual herbaceous species and soil environmental factors
3.2 一年生草本植物空间格局的变化
植物种群的空间分布格局是植物自身的生物特性(扩散机制、生物相互作用、克隆繁殖生长)与生境异质性共同作用的结果[34]。种群在生境异质条件下会通过调整表型特征向有利于适合度的方向发展,改变斑块间的距离(尺度),这时的格局分布方式及格局强度是植物生态适应的重要表现[35]。集群分布和空间正关联体现了互为有利的生态学关系,均匀分布和空间负关联体现了生态学关系是相互排斥的,而随机分布和空间无关联则说明生态关系不明确[36]。空间格局与尺度相互依赖,特定尺度下存在特定的导因[37]。本研究发现,一年生草本植物在各个尺度范围内多表现为聚集分布,一是由于自然条件下,植物通过繁殖在母体周围产生新个体而呈现聚集分布[38],二是由于荒漠地区的“肥岛效应”,由于灌木的生长以及土壤养分、水分、盐分以及其他非生物因素的相互作用,其冠幅下限制性土壤资源显著富集或聚集[39],从而导致了一年生草本植物的集群分布。“肥岛效应”是荒漠地区植被在有限的资源环境中稳定生存的基础。三是由于集群分布有利于个体间相互庇护,抵御风沙等自然灾害,提高自身存活概率和种间竞争力[40]。荒漠地区水分缺乏,风沙较大,空间分布异质性程度高,生境条件严酷多变,一年生草本植物根系不发达,在严酷的环境中,聚集生长有利于稳定土壤,对抗环境的干扰[41]。在荒漠环境下,一年生草本植物随机分布,一方面主要是由于种子分布受风沙活动影响,而风沙活动作为随机因素,导致种子在风力扩散过程中随机散落。另一方面可能是由于种间竞争过程中种群自调节机理不同引起聚集分布格局向随机分布转变[42],整体上表现为小聚集、大分散,形成沙漠中最常见的簇丛状分布格局,造成这种现象的重要因素是资源的斑块性和物种特定的繁殖和扩散策略[43]。一年生草本植物的空间格局随着空间尺度的不同而变化,聚集程度随尺度的增大而减小,这种变化是种群的生存策略或适应机制[44]。在不同林龄内导致分布格局差异的主要限制因子可能是环境异质性,例如土壤理化性质等,且沙漠中资源有限,由于生境异质性可以在局部适宜的环境中增加草本植物密度,促进物种聚集,而各种因素促使有限资源分布不均匀可能会影响物种分布格局[45]。
3.3 一年生草本植物空间关联性的变化
空间关联在物种间的相互作用中发挥关键作用,这些相互作用与物种动态相关的生态过程密切相关,如生长、存活、更新和死亡等[46]。种间关联性差异与生态位重叠和生态位分化有关[47]。种间关联理论认为,正关联是由于物种间具有依赖性或对存在的环境有相同的适应而产生的,种间相互作用有利于维持群落的稳定共存。负关联则是物种间互相竞争或对环境需求的差异综合作用下的结果[48]。本研究中,沙蓬和雾冰藜的相互关系从部分尺度相互促进转变为没有明显的空间关联性。一方面说明沙蓬作为流动沙丘的先锋植物,为其他植物的萌发和生长提供了一定的生境条件,但随着植物数量的增加,相互间的关系就会减弱;另一方面说明沙蓬与雾冰藜相互间竞争较弱,相互独立,可长期共存。雾冰藜和白茎盐生草随着林龄的增加无关联尺度变小,正关联尺度变大,说明两者相互吸引,对生境的选择有一致性,并且正关联性越强,表明群落中种群之间的互补性越强,资源利用更加充分,进而增强群落稳定性。在部分尺度下存在无关联和负关联,说明雾冰藜同时存在生态位空间隔离[49]和竞争关系。
4 结论
随着人工梭梭林龄的增加,林下一年生草本植物由沙蓬-雾冰藜群落逐渐发展到雾冰藜-白茎盐生草群落。
沙蓬由最初的聚集分布向随机分布转变;雾冰藜先由最初的聚集分布转变为随机分布,然后又由随机分布转变为聚集分布;白茎盐生草由局部尺度的聚集分布转变为大范围尺度的聚集分布。沙蓬与雾冰藜之间主要呈无关联,而雾冰藜与白茎盐生草之间主要呈正关联;
不同林龄梭梭林间存在一定的空间异质性,钙离子、钠离子、2~5 cm土壤水含量是影响梭梭林下一年生草本植物群落变化的关键环境因子。
参考文献
Spatial pattern analysis in plant ecology
[J].
Understanding mechanism of spatial ecological phenomena:a preface to the special feature on “spatial statistics”
[J].
大兴安岭次生林区优势种落叶松分布格局及竞争作用
[J].
宁夏荒漠草原蒙古冰草(Agropyron mongolicum)种群小尺度空间分布及其关联性
[J].
Ecological effects of establishing a 40-year oasis protection system in a northwestern China desert
[J].
Ecological effects of desertification control and desertified land reclamation in an oasis-desert ecotone in an arid region:a case study in Hexi Corridor, Northwest China
[J].
Analyzing the spatial structure of a Sri Lankan tree species with multiple scales of clustering
[J].
Balance between facilitation and competition determines spatial patterns in a plant population
[J].
On some aspects of spatial pattern in bio-logical populations
[J].
Spatial patterns in the distribution of tropical tree species
[J].
Heterogeneity influences spatial patterns and demographics in forest stands
[J].
Spatial pattern of trees influences species productivity in a mature oak-pine mixed forest
[J].
Mechanisms of maintenance of species diversity
[J].

 甘公网安备 62010202000688号
甘公网安备 62010202000688号
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}