Dryland self-expansion enabled by land-atmosphere feedbacks
1
2024
... 干旱区(包括半干旱区和亚湿润偏旱区)约占全球陆地总面积的45%[1].在未来温度升高和降水格局改变的气候变化大背景下,可能会带来植被衰退、土壤退化和生态系统崩溃等问题,制约生态、经济和社会的可持续发展,因此干旱区生态恢复任务紧迫[2].水分是制约干旱区生态恢复的关键因子[3].植物水力传导主要由土壤-植物-大气连续体(Soil-Plant-Atmosphere continuum,SPAC)的水势梯度引起[4],能够调节植物生产力,影响水循环、碳循环、生物地球化学循环以及植物对干旱和气候变化的响应[5].将大气水需求和土壤水供应等非生物因子与植物根、茎、叶水分传导速率及水分传导面积等生物因子相结合,探究水分胁迫对植物不同器官水力传导的影响至关重要[6-8],有助于了解植物的水分利用策略[9]. ...
Global ecosystem thresholds driven by aridity
1
2020
... 干旱区(包括半干旱区和亚湿润偏旱区)约占全球陆地总面积的45%[1].在未来温度升高和降水格局改变的气候变化大背景下,可能会带来植被衰退、土壤退化和生态系统崩溃等问题,制约生态、经济和社会的可持续发展,因此干旱区生态恢复任务紧迫[2].水分是制约干旱区生态恢复的关键因子[3].植物水力传导主要由土壤-植物-大气连续体(Soil-Plant-Atmosphere continuum,SPAC)的水势梯度引起[4],能够调节植物生产力,影响水循环、碳循环、生物地球化学循环以及植物对干旱和气候变化的响应[5].将大气水需求和土壤水供应等非生物因子与植物根、茎、叶水分传导速率及水分传导面积等生物因子相结合,探究水分胁迫对植物不同器官水力传导的影响至关重要[6-8],有助于了解植物的水分利用策略[9]. ...
中国北方沙区水量平衡自动模拟监测系统(沙坡头蒸渗仪群)
1
2021
... 干旱区(包括半干旱区和亚湿润偏旱区)约占全球陆地总面积的45%[1].在未来温度升高和降水格局改变的气候变化大背景下,可能会带来植被衰退、土壤退化和生态系统崩溃等问题,制约生态、经济和社会的可持续发展,因此干旱区生态恢复任务紧迫[2].水分是制约干旱区生态恢复的关键因子[3].植物水力传导主要由土壤-植物-大气连续体(Soil-Plant-Atmosphere continuum,SPAC)的水势梯度引起[4],能够调节植物生产力,影响水循环、碳循环、生物地球化学循环以及植物对干旱和气候变化的响应[5].将大气水需求和土壤水供应等非生物因子与植物根、茎、叶水分传导速率及水分传导面积等生物因子相结合,探究水分胁迫对植物不同器官水力传导的影响至关重要[6-8],有助于了解植物的水分利用策略[9]. ...
Soil rather than xylem vulnerability controls stomatal response to drought
3
2020
... 干旱区(包括半干旱区和亚湿润偏旱区)约占全球陆地总面积的45%[1].在未来温度升高和降水格局改变的气候变化大背景下,可能会带来植被衰退、土壤退化和生态系统崩溃等问题,制约生态、经济和社会的可持续发展,因此干旱区生态恢复任务紧迫[2].水分是制约干旱区生态恢复的关键因子[3].植物水力传导主要由土壤-植物-大气连续体(Soil-Plant-Atmosphere continuum,SPAC)的水势梯度引起[4],能够调节植物生产力,影响水循环、碳循环、生物地球化学循环以及植物对干旱和气候变化的响应[5].将大气水需求和土壤水供应等非生物因子与植物根、茎、叶水分传导速率及水分传导面积等生物因子相结合,探究水分胁迫对植物不同器官水力传导的影响至关重要[6-8],有助于了解植物的水分利用策略[9]. ...
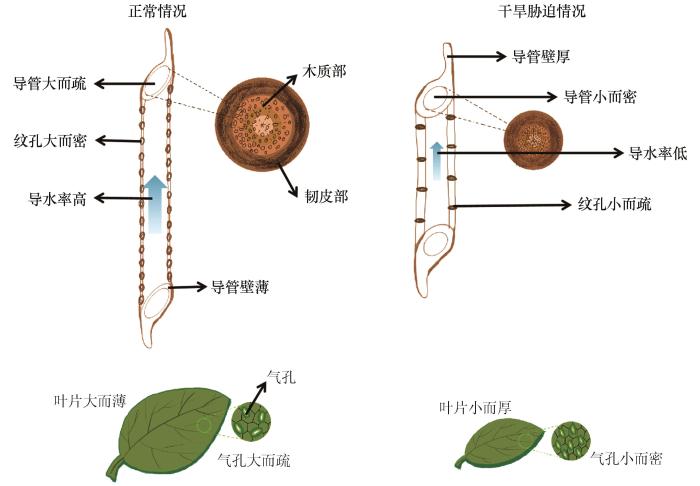
... 植物远端叶片水力阻力较大,占全植株水力阻力的30%,是SPAC水力运输的瓶颈和“安全阀”[9,19,26,47].植物叶片蒸腾约占地球表面蒸腾的70%[26],植物叶片通常通过气孔开闭适应环境.叶片气孔变化主要受两个机制调节:一是由叶表皮细胞和保卫细胞之间的水力连接引发的被动调节机制;二是激素如脱落酸的主动调节机制[4].气孔对水力调节和激素调节的敏感度与植物的等水调节和非等水调节有关[4],前者是指植物叶片气孔在干旱胁迫下迅速关闭以保证植物叶片日最小水势保持不变,在水分充足时复水快,易死于碳饥饿;后者是指植物叶片气孔在干旱胁迫下仍保持打开,降低叶片日最小水势,在水分充足时复水慢,易死于水力失败[6,11-12,48].有趣的是,Kannenberg等[6]认为非等水调节植物在水分胁迫期间并没有增加碳同化量.在自然界中没有严格的等水植物和非等水植物,通常介于两者之间[6].当土壤处于中等干旱条件下,植物通常会趋向于非等水调节;而当植物处于严重干旱条件下,植物趋向于等水调节[6,49].植物的等水和非等水程度的量化常采用水力学面积,即测量边界叶水势和1∶1线围成的多边形面积,水力学面积越小越趋向于等水调节[50];也可采用凌晨和午间水势的斜率来表示等水程度,斜率接近0表示植物偏向于等水植物,接近1或大于1表示植物偏向于非等水植物[51].陈嘉嘉[52]研究结果显示,在腾格里沙漠生态系统中,常见的固沙植物油蒿(Artemisia ordosica)、柠条锦鸡儿(Caragana korshinskii)和花棒(Hedysarum scoparium)的凌晨和午间水势斜率分别是0.33、0.92和1.22,分别属于偏等水植物、偏非等水植物和严格非等水植物.植物叶片除了通过气孔开合应对轻微水分胁迫,也可改变叶片解剖形态特征是对干旱胁迫做出的短期快速响应,如叶片气孔在干旱条件下呈小而密的分布(图2),可通过增加气孔闭合能力来减少蒸腾过程中的水分散失[44-45]. ...
... [4],前者是指植物叶片气孔在干旱胁迫下迅速关闭以保证植物叶片日最小水势保持不变,在水分充足时复水快,易死于碳饥饿;后者是指植物叶片气孔在干旱胁迫下仍保持打开,降低叶片日最小水势,在水分充足时复水慢,易死于水力失败[6,11-12,48].有趣的是,Kannenberg等[6]认为非等水调节植物在水分胁迫期间并没有增加碳同化量.在自然界中没有严格的等水植物和非等水植物,通常介于两者之间[6].当土壤处于中等干旱条件下,植物通常会趋向于非等水调节;而当植物处于严重干旱条件下,植物趋向于等水调节[6,49].植物的等水和非等水程度的量化常采用水力学面积,即测量边界叶水势和1∶1线围成的多边形面积,水力学面积越小越趋向于等水调节[50];也可采用凌晨和午间水势的斜率来表示等水程度,斜率接近0表示植物偏向于等水植物,接近1或大于1表示植物偏向于非等水植物[51].陈嘉嘉[52]研究结果显示,在腾格里沙漠生态系统中,常见的固沙植物油蒿(Artemisia ordosica)、柠条锦鸡儿(Caragana korshinskii)和花棒(Hedysarum scoparium)的凌晨和午间水势斜率分别是0.33、0.92和1.22,分别属于偏等水植物、偏非等水植物和严格非等水植物.植物叶片除了通过气孔开合应对轻微水分胁迫,也可改变叶片解剖形态特征是对干旱胁迫做出的短期快速响应,如叶片气孔在干旱条件下呈小而密的分布(图2),可通过增加气孔闭合能力来减少蒸腾过程中的水分散失[44-45]. ...
Spatial and temporal variation in plant hydraulic traits and their relevance for climate change impacts on vegetation
1
2015
... 干旱区(包括半干旱区和亚湿润偏旱区)约占全球陆地总面积的45%[1].在未来温度升高和降水格局改变的气候变化大背景下,可能会带来植被衰退、土壤退化和生态系统崩溃等问题,制约生态、经济和社会的可持续发展,因此干旱区生态恢复任务紧迫[2].水分是制约干旱区生态恢复的关键因子[3].植物水力传导主要由土壤-植物-大气连续体(Soil-Plant-Atmosphere continuum,SPAC)的水势梯度引起[4],能够调节植物生产力,影响水循环、碳循环、生物地球化学循环以及植物对干旱和气候变化的响应[5].将大气水需求和土壤水供应等非生物因子与植物根、茎、叶水分传导速率及水分传导面积等生物因子相结合,探究水分胁迫对植物不同器官水力传导的影响至关重要[6-8],有助于了解植物的水分利用策略[9]. ...
Anisohydric behavior linked to persistent hydraulic damage and delayed drought recovery across seven North American tree species
6
2019
... 干旱区(包括半干旱区和亚湿润偏旱区)约占全球陆地总面积的45%[1].在未来温度升高和降水格局改变的气候变化大背景下,可能会带来植被衰退、土壤退化和生态系统崩溃等问题,制约生态、经济和社会的可持续发展,因此干旱区生态恢复任务紧迫[2].水分是制约干旱区生态恢复的关键因子[3].植物水力传导主要由土壤-植物-大气连续体(Soil-Plant-Atmosphere continuum,SPAC)的水势梯度引起[4],能够调节植物生产力,影响水循环、碳循环、生物地球化学循环以及植物对干旱和气候变化的响应[5].将大气水需求和土壤水供应等非生物因子与植物根、茎、叶水分传导速率及水分传导面积等生物因子相结合,探究水分胁迫对植物不同器官水力传导的影响至关重要[6-8],有助于了解植物的水分利用策略[9]. ...
... 随着干旱胁迫程度的增加,植物的蒸腾张力超出一定临界值,导致其木质部导管中局部水柱断裂,产生空穴化并扩张,进而出现木质部栓塞现象[10].木质部栓塞的进一步发展会导致叶片光合速率降低和冠层枯梢,甚至整个植株因水力失败而死亡[10-11].目前关于导致植物死亡的假说主要包括水力失败、碳饥饿和生物攻击假说[11-12],且越来越多研究显示各假说之间存在交互作用[11].在目前全球气候变化大背景下,越来越多的研究都在关注植物死亡机理及过程,而关于植物如何忍耐干旱的研究较少[6].本文综述了国内外关于植物根、茎、叶不同器官在干旱胁迫条件下的形态调整、水力传导过程及其调整策略,可为揭示植物应对干旱的水力策略提供理论基础,对干旱区生态保护和植被恢复具有重要的实践意义. ...
... 植物远端叶片水力阻力较大,占全植株水力阻力的30%,是SPAC水力运输的瓶颈和“安全阀”[9,19,26,47].植物叶片蒸腾约占地球表面蒸腾的70%[26],植物叶片通常通过气孔开闭适应环境.叶片气孔变化主要受两个机制调节:一是由叶表皮细胞和保卫细胞之间的水力连接引发的被动调节机制;二是激素如脱落酸的主动调节机制[4].气孔对水力调节和激素调节的敏感度与植物的等水调节和非等水调节有关[4],前者是指植物叶片气孔在干旱胁迫下迅速关闭以保证植物叶片日最小水势保持不变,在水分充足时复水快,易死于碳饥饿;后者是指植物叶片气孔在干旱胁迫下仍保持打开,降低叶片日最小水势,在水分充足时复水慢,易死于水力失败[6,11-12,48].有趣的是,Kannenberg等[6]认为非等水调节植物在水分胁迫期间并没有增加碳同化量.在自然界中没有严格的等水植物和非等水植物,通常介于两者之间[6].当土壤处于中等干旱条件下,植物通常会趋向于非等水调节;而当植物处于严重干旱条件下,植物趋向于等水调节[6,49].植物的等水和非等水程度的量化常采用水力学面积,即测量边界叶水势和1∶1线围成的多边形面积,水力学面积越小越趋向于等水调节[50];也可采用凌晨和午间水势的斜率来表示等水程度,斜率接近0表示植物偏向于等水植物,接近1或大于1表示植物偏向于非等水植物[51].陈嘉嘉[52]研究结果显示,在腾格里沙漠生态系统中,常见的固沙植物油蒿(Artemisia ordosica)、柠条锦鸡儿(Caragana korshinskii)和花棒(Hedysarum scoparium)的凌晨和午间水势斜率分别是0.33、0.92和1.22,分别属于偏等水植物、偏非等水植物和严格非等水植物.植物叶片除了通过气孔开合应对轻微水分胁迫,也可改变叶片解剖形态特征是对干旱胁迫做出的短期快速响应,如叶片气孔在干旱条件下呈小而密的分布(图2),可通过增加气孔闭合能力来减少蒸腾过程中的水分散失[44-45]. ...
... [6]认为非等水调节植物在水分胁迫期间并没有增加碳同化量.在自然界中没有严格的等水植物和非等水植物,通常介于两者之间[6].当土壤处于中等干旱条件下,植物通常会趋向于非等水调节;而当植物处于严重干旱条件下,植物趋向于等水调节[6,49].植物的等水和非等水程度的量化常采用水力学面积,即测量边界叶水势和1∶1线围成的多边形面积,水力学面积越小越趋向于等水调节[50];也可采用凌晨和午间水势的斜率来表示等水程度,斜率接近0表示植物偏向于等水植物,接近1或大于1表示植物偏向于非等水植物[51].陈嘉嘉[52]研究结果显示,在腾格里沙漠生态系统中,常见的固沙植物油蒿(Artemisia ordosica)、柠条锦鸡儿(Caragana korshinskii)和花棒(Hedysarum scoparium)的凌晨和午间水势斜率分别是0.33、0.92和1.22,分别属于偏等水植物、偏非等水植物和严格非等水植物.植物叶片除了通过气孔开合应对轻微水分胁迫,也可改变叶片解剖形态特征是对干旱胁迫做出的短期快速响应,如叶片气孔在干旱条件下呈小而密的分布(图2),可通过增加气孔闭合能力来减少蒸腾过程中的水分散失[44-45]. ...
... [6].当土壤处于中等干旱条件下,植物通常会趋向于非等水调节;而当植物处于严重干旱条件下,植物趋向于等水调节[6,49].植物的等水和非等水程度的量化常采用水力学面积,即测量边界叶水势和1∶1线围成的多边形面积,水力学面积越小越趋向于等水调节[50];也可采用凌晨和午间水势的斜率来表示等水程度,斜率接近0表示植物偏向于等水植物,接近1或大于1表示植物偏向于非等水植物[51].陈嘉嘉[52]研究结果显示,在腾格里沙漠生态系统中,常见的固沙植物油蒿(Artemisia ordosica)、柠条锦鸡儿(Caragana korshinskii)和花棒(Hedysarum scoparium)的凌晨和午间水势斜率分别是0.33、0.92和1.22,分别属于偏等水植物、偏非等水植物和严格非等水植物.植物叶片除了通过气孔开合应对轻微水分胁迫,也可改变叶片解剖形态特征是对干旱胁迫做出的短期快速响应,如叶片气孔在干旱条件下呈小而密的分布(图2),可通过增加气孔闭合能力来减少蒸腾过程中的水分散失[44-45]. ...
... [6,49].植物的等水和非等水程度的量化常采用水力学面积,即测量边界叶水势和1∶1线围成的多边形面积,水力学面积越小越趋向于等水调节[50];也可采用凌晨和午间水势的斜率来表示等水程度,斜率接近0表示植物偏向于等水植物,接近1或大于1表示植物偏向于非等水植物[51].陈嘉嘉[52]研究结果显示,在腾格里沙漠生态系统中,常见的固沙植物油蒿(Artemisia ordosica)、柠条锦鸡儿(Caragana korshinskii)和花棒(Hedysarum scoparium)的凌晨和午间水势斜率分别是0.33、0.92和1.22,分别属于偏等水植物、偏非等水植物和严格非等水植物.植物叶片除了通过气孔开合应对轻微水分胁迫,也可改变叶片解剖形态特征是对干旱胁迫做出的短期快速响应,如叶片气孔在干旱条件下呈小而密的分布(图2),可通过增加气孔闭合能力来减少蒸腾过程中的水分散失[44-45]. ...
2
2002
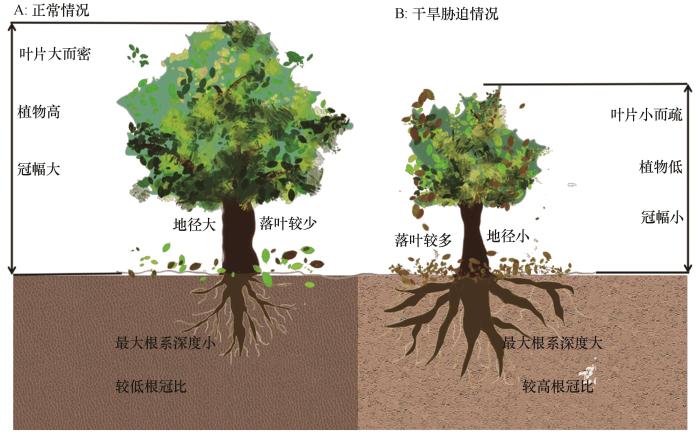
... 在目前的温度升高和极端降水事件频发的全球气候变化背景下,近10年来中国干旱区平均气温升高0.5 ℃,且降水减少4 mm,呈“暖干化”的变化趋势,气温和降水变化可能会导致干旱区植物死亡[13].量化植物水力导度和水力失败的临界水势值等水力特征有助于预测在未来气候变化背景下植物个体存活、群落组成、物种分布和生态系统功能[13].植物水力传导与植物形态特征息息相关.植物常增加最大根系分布深度和根冠比以增加根系吸水(图1)[14-16],同时降低根系直径,并增加根系表面积和长度以响应干旱胁迫[17].McDowell等[18]基于达西水力推论指出,植物可通过降低高度以适应在气候变化背景下的植物高蒸腾需求;植物茎干通常会减少单位水运输量的木质部横截面投资(地径减小)以应对干旱胁迫[13].植物叶片在干旱胁迫下较厚(高比叶重),往往具有较长的叶肉扩散路径,这增加了叶片木质部外水分移动的阻力,并降低了水力传导性[19],避免植物过多的叶片蒸腾失水导致水力失败[13];植物叶片也可通过落叶和减小叶面积以降低单位边材面积对应的叶面积来保证水平衡.在干旱胁迫下,植物碳投入成本较高的主干和主根通常会舍弃碳投入成本低、水分运输效率低和抗栓塞性低的远端叶片(即落叶)和根系以保证其水力传输(图1),即水力脆弱性分割假说[9].该假说包括水力分割假说和脆弱性分割假说.水力分割假说认为远端器官(根和叶等)水力阻力较大;脆弱性假说认为当面对水势下降时,远端器官(根和叶等)相比于基部主干等器官更脆弱且容易发生空化[7,9,20].Pivovaroff等[9]研究显示半干旱的地中海气候下灌木的叶片水力导度和茎干水力导度负相关,证实了叶片通过增加水力阻力和降低水力导度来保护茎干免受灾难性的水力失败.值得一提的是,水力脆弱性分割现象多发生在受水分限制的干旱、半干旱和季节性干旱地区,湿润地区的植物如常绿阔叶树种由于其叶片碳投入成本昂贵,茎叶的抗栓塞性较强,发生脆弱性分割的可能性较小甚至不存在[21-22]. ...
... 植物木质部主要包括薄壁细胞、木射线、纤维和导管(管胞)等,薄壁细胞具有储存糖分和水分的作用[24,40],相邻导管间的水分传输通过纹孔结构实现,不相邻导管间的水分传输基于纤维桥假说通过纤维管胞实现[41],纤维具有机械支持作用,导管(管胞)具有水分传输作用.木质部栓塞是植物应对干旱胁迫的被动结果.基于内聚力-张力理论,叶片蒸腾产生的张力大于水分子间的内聚力与水柱和管壁间的附着作用时,木质部导管中水柱断裂并产生气泡,阻碍水分向上输送[7,42-43].气泡的形成可由“气种假说”诠释[43]:①空气泡从纹孔中进入导管;②管道中疏水性区域小气泡膨胀;③木质部中水分流动时,小气泡扩张;④导管壁中小气泡与疏水区脱离.目前关于栓塞的发生主要有两种观点,一种观点指出木质部的空穴和栓塞现象是植物对干旱的响应,在自然界经常发生[43];另一种观点是栓塞只有在极端干旱胁迫下才偶尔发生,且栓塞在水分充足时会被修复[26].木质部栓塞程度随干旱程度增加而增加,常采用栓塞脆弱性曲线来表示[10].木质部抗栓塞性会随植物种类和枝条年龄而异.成年植物个体的木质部有更多的纤维组织来确保机械支撑和水力安全;幼年植物个体的木质部通常会优先形成射线薄壁组织和轴向薄壁组织来进行水分和营养存储[24].Nolf等[10]指出赤桉(Eucalyptus camaldulensis)的当年生枝条相比于老枝更容易受干旱的影响,抗栓塞性较弱;Schuldt等[42]则认为欧洲山毛榉(Fagus sylvatica)生长较慢的老枝比生长较快的新枝的栓塞脆弱性高,可能与老枝空化疲劳有关. ...
The hydraulic architecture of an arborescent monocot:ontogeny-related adjustments in vessel size and leaf area compensate for increased resistance
3
2021
... 干旱区(包括半干旱区和亚湿润偏旱区)约占全球陆地总面积的45%[1].在未来温度升高和降水格局改变的气候变化大背景下,可能会带来植被衰退、土壤退化和生态系统崩溃等问题,制约生态、经济和社会的可持续发展,因此干旱区生态恢复任务紧迫[2].水分是制约干旱区生态恢复的关键因子[3].植物水力传导主要由土壤-植物-大气连续体(Soil-Plant-Atmosphere continuum,SPAC)的水势梯度引起[4],能够调节植物生产力,影响水循环、碳循环、生物地球化学循环以及植物对干旱和气候变化的响应[5].将大气水需求和土壤水供应等非生物因子与植物根、茎、叶水分传导速率及水分传导面积等生物因子相结合,探究水分胁迫对植物不同器官水力传导的影响至关重要[6-8],有助于了解植物的水分利用策略[9]. ...
... 植物木质部可通过改变导管结构特征来应对干旱胁迫.木质部常通过减小导管直径,增加导管密度、纹孔膜厚度和木材密度,进而降低木质部导水率并提高水力安全性,且已有大量研究证实(图2)[13,42,44].He等[13]指出相比于湿润环境,在干旱环境下水力失败风险较大且维护成本较高,植物通常会降低导水率以适应环境.Schuldt等[42]发现欧洲山毛榉的木质部导管直径降低7%且纹孔膜厚度增加15%以应对干旱胁迫.导管结构特征影响木质部导水率,Hagen-Poiseuille水力传导方程显示导管导水率与导管直径的4次方成正比,因此导管直径的微小增加会带来导水率的显著提高[41].有研究采用X射线显微断层成像法观测干燥不同时间的赤桉的木质部横截面的导管栓塞情况,结果显示Hagen-Poiseuille水力传导方程模拟的理论导水率高于实际最大导水率[10],原因一是与没有考虑导管长度和导管间的纹孔结构有关[41],导管长度越短、纹孔数量越少和纹孔面积越小,则导管导水率越低[41],纹孔阻力约占整个植株总水力传导阻力的14%~84%[40];二是与导管腔的不规则形状和锥度有关[45];三是与木质部汁液有关,研究显示KCl浓度的增加可导致木质部导水率增加10%[45].枝条形态学基部导管直径通常高于形态学末端[8,40],这一形态结构有利于减小由于植物高度增长带来的水力阻力[8].通常情况下导管直径表现为主干直径>侧枝>枝条交叉点[40].植物体的枝条交叉点、根茎过渡区和节点部位的导管直径较小、导水率较小、胡伯尔值较大和抗栓塞能力较强,这些部位常被称为“水流限制区”和“安全区”[46].胡伯尔值越大,表示维持单位叶面积水分需要的木质部越多,可用于比较不同树种间的水力特征[19,40]. ...
... [8].通常情况下导管直径表现为主干直径>侧枝>枝条交叉点[40].植物体的枝条交叉点、根茎过渡区和节点部位的导管直径较小、导水率较小、胡伯尔值较大和抗栓塞能力较强,这些部位常被称为“水流限制区”和“安全区”[46].胡伯尔值越大,表示维持单位叶面积水分需要的木质部越多,可用于比较不同树种间的水力特征[19,40]. ...
Coordination of stem and leaf hydraulic conductance in southern California shrubs: a test of the hydraulic segmentation hypothesis
6
2014
... 干旱区(包括半干旱区和亚湿润偏旱区)约占全球陆地总面积的45%[1].在未来温度升高和降水格局改变的气候变化大背景下,可能会带来植被衰退、土壤退化和生态系统崩溃等问题,制约生态、经济和社会的可持续发展,因此干旱区生态恢复任务紧迫[2].水分是制约干旱区生态恢复的关键因子[3].植物水力传导主要由土壤-植物-大气连续体(Soil-Plant-Atmosphere continuum,SPAC)的水势梯度引起[4],能够调节植物生产力,影响水循环、碳循环、生物地球化学循环以及植物对干旱和气候变化的响应[5].将大气水需求和土壤水供应等非生物因子与植物根、茎、叶水分传导速率及水分传导面积等生物因子相结合,探究水分胁迫对植物不同器官水力传导的影响至关重要[6-8],有助于了解植物的水分利用策略[9]. ...
... 在目前的温度升高和极端降水事件频发的全球气候变化背景下,近10年来中国干旱区平均气温升高0.5 ℃,且降水减少4 mm,呈“暖干化”的变化趋势,气温和降水变化可能会导致干旱区植物死亡[13].量化植物水力导度和水力失败的临界水势值等水力特征有助于预测在未来气候变化背景下植物个体存活、群落组成、物种分布和生态系统功能[13].植物水力传导与植物形态特征息息相关.植物常增加最大根系分布深度和根冠比以增加根系吸水(图1)[14-16],同时降低根系直径,并增加根系表面积和长度以响应干旱胁迫[17].McDowell等[18]基于达西水力推论指出,植物可通过降低高度以适应在气候变化背景下的植物高蒸腾需求;植物茎干通常会减少单位水运输量的木质部横截面投资(地径减小)以应对干旱胁迫[13].植物叶片在干旱胁迫下较厚(高比叶重),往往具有较长的叶肉扩散路径,这增加了叶片木质部外水分移动的阻力,并降低了水力传导性[19],避免植物过多的叶片蒸腾失水导致水力失败[13];植物叶片也可通过落叶和减小叶面积以降低单位边材面积对应的叶面积来保证水平衡.在干旱胁迫下,植物碳投入成本较高的主干和主根通常会舍弃碳投入成本低、水分运输效率低和抗栓塞性低的远端叶片(即落叶)和根系以保证其水力传输(图1),即水力脆弱性分割假说[9].该假说包括水力分割假说和脆弱性分割假说.水力分割假说认为远端器官(根和叶等)水力阻力较大;脆弱性假说认为当面对水势下降时,远端器官(根和叶等)相比于基部主干等器官更脆弱且容易发生空化[7,9,20].Pivovaroff等[9]研究显示半干旱的地中海气候下灌木的叶片水力导度和茎干水力导度负相关,证实了叶片通过增加水力阻力和降低水力导度来保护茎干免受灾难性的水力失败.值得一提的是,水力脆弱性分割现象多发生在受水分限制的干旱、半干旱和季节性干旱地区,湿润地区的植物如常绿阔叶树种由于其叶片碳投入成本昂贵,茎叶的抗栓塞性较强,发生脆弱性分割的可能性较小甚至不存在[21-22]. ...
... ,9,20].Pivovaroff等[9]研究显示半干旱的地中海气候下灌木的叶片水力导度和茎干水力导度负相关,证实了叶片通过增加水力阻力和降低水力导度来保护茎干免受灾难性的水力失败.值得一提的是,水力脆弱性分割现象多发生在受水分限制的干旱、半干旱和季节性干旱地区,湿润地区的植物如常绿阔叶树种由于其叶片碳投入成本昂贵,茎叶的抗栓塞性较强,发生脆弱性分割的可能性较小甚至不存在[21-22]. ...
... [9]研究显示半干旱的地中海气候下灌木的叶片水力导度和茎干水力导度负相关,证实了叶片通过增加水力阻力和降低水力导度来保护茎干免受灾难性的水力失败.值得一提的是,水力脆弱性分割现象多发生在受水分限制的干旱、半干旱和季节性干旱地区,湿润地区的植物如常绿阔叶树种由于其叶片碳投入成本昂贵,茎叶的抗栓塞性较强,发生脆弱性分割的可能性较小甚至不存在[21-22]. ...
... 植物远端叶片水力阻力较大,占全植株水力阻力的30%,是SPAC水力运输的瓶颈和“安全阀”[9,19,26,47].植物叶片蒸腾约占地球表面蒸腾的70%[26],植物叶片通常通过气孔开闭适应环境.叶片气孔变化主要受两个机制调节:一是由叶表皮细胞和保卫细胞之间的水力连接引发的被动调节机制;二是激素如脱落酸的主动调节机制[4].气孔对水力调节和激素调节的敏感度与植物的等水调节和非等水调节有关[4],前者是指植物叶片气孔在干旱胁迫下迅速关闭以保证植物叶片日最小水势保持不变,在水分充足时复水快,易死于碳饥饿;后者是指植物叶片气孔在干旱胁迫下仍保持打开,降低叶片日最小水势,在水分充足时复水慢,易死于水力失败[6,11-12,48].有趣的是,Kannenberg等[6]认为非等水调节植物在水分胁迫期间并没有增加碳同化量.在自然界中没有严格的等水植物和非等水植物,通常介于两者之间[6].当土壤处于中等干旱条件下,植物通常会趋向于非等水调节;而当植物处于严重干旱条件下,植物趋向于等水调节[6,49].植物的等水和非等水程度的量化常采用水力学面积,即测量边界叶水势和1∶1线围成的多边形面积,水力学面积越小越趋向于等水调节[50];也可采用凌晨和午间水势的斜率来表示等水程度,斜率接近0表示植物偏向于等水植物,接近1或大于1表示植物偏向于非等水植物[51].陈嘉嘉[52]研究结果显示,在腾格里沙漠生态系统中,常见的固沙植物油蒿(Artemisia ordosica)、柠条锦鸡儿(Caragana korshinskii)和花棒(Hedysarum scoparium)的凌晨和午间水势斜率分别是0.33、0.92和1.22,分别属于偏等水植物、偏非等水植物和严格非等水植物.植物叶片除了通过气孔开合应对轻微水分胁迫,也可改变叶片解剖形态特征是对干旱胁迫做出的短期快速响应,如叶片气孔在干旱条件下呈小而密的分布(图2),可通过增加气孔闭合能力来减少蒸腾过程中的水分散失[44-45]. ...
... 植物进行形态调整,根系水力再分配,改变木质部和叶片的结构特征的同时,也会进行水力效率和水力安全之间的权衡.水力效率常采用水力导度来衡量,它是水力阻力的倒数,根据欧姆定律,水力导度可用液流速率与压力差比值表示[9],常采用、表征植物木质部的抗栓塞性,采用水力导度降低50%和88%时对应的水势(,)表征植物叶片的抗栓塞性,即植物木质部和叶片的水力安全[42],其值越低表示植物越能忍受较低水势,抗栓塞性越强[30].木质部抗栓塞性和干旱度指数呈线性增加趋势,在干旱胁迫发生时木质部能够忍受更低的水势保证水分运输的正常运行[42].“水力权衡假说”认为植物的不同器官间,存在水力传导效率和安全的高低权衡[42-43].Huo等[44]认为在干旱沙区,柠条锦鸡儿通过降低导水率并增加水力安全性来保证植物能够在干旱胁迫下生存;Zhang等[53]发现生长在干旱沙漠生态系统的植物通常具有较高的水力安全性和较低的导水率,植物的存活与植物水力安全密切相关.而目前也有研究显示植物水力效率和安全不存在权衡关系[42-43];低水力效率和低水力安全的植物可在湿润地区生存[43];也有进一步研究指出自然界中该权衡关系并不是非此即彼的,而是根据环境和植物结构特征而变化[54].水力效率和水力安全之间的权衡关系是由导管结构特征和导管间纹孔结构决定的,水力效率高的木质部通常导管直径较大、纹孔结构较多且木材密度较低,相应的水力安全性较低[55].研究显示在中国北方干旱区,灌木物种在相对干旱区根部导管直径较大并采用效率优先的导水策略;而在相对湿润区,灌木物种导管直径较小并采用安全优先的导水策略[56].尤其是在降水稀少且高温的干旱区,水分为植物生存生长的重要限制因子,植物对水力效率和水力安全的权衡关系研究需要进一步探究. ...
Visualization of xylem embolism by X-ray microtomography: a direct test against hydraulic measurements
5
2017
... 随着干旱胁迫程度的增加,植物的蒸腾张力超出一定临界值,导致其木质部导管中局部水柱断裂,产生空穴化并扩张,进而出现木质部栓塞现象[10].木质部栓塞的进一步发展会导致叶片光合速率降低和冠层枯梢,甚至整个植株因水力失败而死亡[10-11].目前关于导致植物死亡的假说主要包括水力失败、碳饥饿和生物攻击假说[11-12],且越来越多研究显示各假说之间存在交互作用[11].在目前全球气候变化大背景下,越来越多的研究都在关注植物死亡机理及过程,而关于植物如何忍耐干旱的研究较少[6].本文综述了国内外关于植物根、茎、叶不同器官在干旱胁迫条件下的形态调整、水力传导过程及其调整策略,可为揭示植物应对干旱的水力策略提供理论基础,对干旱区生态保护和植被恢复具有重要的实践意义. ...
... [10-11].目前关于导致植物死亡的假说主要包括水力失败、碳饥饿和生物攻击假说[11-12],且越来越多研究显示各假说之间存在交互作用[11].在目前全球气候变化大背景下,越来越多的研究都在关注植物死亡机理及过程,而关于植物如何忍耐干旱的研究较少[6].本文综述了国内外关于植物根、茎、叶不同器官在干旱胁迫条件下的形态调整、水力传导过程及其调整策略,可为揭示植物应对干旱的水力策略提供理论基础,对干旱区生态保护和植被恢复具有重要的实践意义. ...
... 植物木质部主要包括薄壁细胞、木射线、纤维和导管(管胞)等,薄壁细胞具有储存糖分和水分的作用[24,40],相邻导管间的水分传输通过纹孔结构实现,不相邻导管间的水分传输基于纤维桥假说通过纤维管胞实现[41],纤维具有机械支持作用,导管(管胞)具有水分传输作用.木质部栓塞是植物应对干旱胁迫的被动结果.基于内聚力-张力理论,叶片蒸腾产生的张力大于水分子间的内聚力与水柱和管壁间的附着作用时,木质部导管中水柱断裂并产生气泡,阻碍水分向上输送[7,42-43].气泡的形成可由“气种假说”诠释[43]:①空气泡从纹孔中进入导管;②管道中疏水性区域小气泡膨胀;③木质部中水分流动时,小气泡扩张;④导管壁中小气泡与疏水区脱离.目前关于栓塞的发生主要有两种观点,一种观点指出木质部的空穴和栓塞现象是植物对干旱的响应,在自然界经常发生[43];另一种观点是栓塞只有在极端干旱胁迫下才偶尔发生,且栓塞在水分充足时会被修复[26].木质部栓塞程度随干旱程度增加而增加,常采用栓塞脆弱性曲线来表示[10].木质部抗栓塞性会随植物种类和枝条年龄而异.成年植物个体的木质部有更多的纤维组织来确保机械支撑和水力安全;幼年植物个体的木质部通常会优先形成射线薄壁组织和轴向薄壁组织来进行水分和营养存储[24].Nolf等[10]指出赤桉(Eucalyptus camaldulensis)的当年生枝条相比于老枝更容易受干旱的影响,抗栓塞性较弱;Schuldt等[42]则认为欧洲山毛榉(Fagus sylvatica)生长较慢的老枝比生长较快的新枝的栓塞脆弱性高,可能与老枝空化疲劳有关. ...
... [10]指出赤桉(Eucalyptus camaldulensis)的当年生枝条相比于老枝更容易受干旱的影响,抗栓塞性较弱;Schuldt等[42]则认为欧洲山毛榉(Fagus sylvatica)生长较慢的老枝比生长较快的新枝的栓塞脆弱性高,可能与老枝空化疲劳有关. ...
... 植物木质部可通过改变导管结构特征来应对干旱胁迫.木质部常通过减小导管直径,增加导管密度、纹孔膜厚度和木材密度,进而降低木质部导水率并提高水力安全性,且已有大量研究证实(图2)[13,42,44].He等[13]指出相比于湿润环境,在干旱环境下水力失败风险较大且维护成本较高,植物通常会降低导水率以适应环境.Schuldt等[42]发现欧洲山毛榉的木质部导管直径降低7%且纹孔膜厚度增加15%以应对干旱胁迫.导管结构特征影响木质部导水率,Hagen-Poiseuille水力传导方程显示导管导水率与导管直径的4次方成正比,因此导管直径的微小增加会带来导水率的显著提高[41].有研究采用X射线显微断层成像法观测干燥不同时间的赤桉的木质部横截面的导管栓塞情况,结果显示Hagen-Poiseuille水力传导方程模拟的理论导水率高于实际最大导水率[10],原因一是与没有考虑导管长度和导管间的纹孔结构有关[41],导管长度越短、纹孔数量越少和纹孔面积越小,则导管导水率越低[41],纹孔阻力约占整个植株总水力传导阻力的14%~84%[40];二是与导管腔的不规则形状和锥度有关[45];三是与木质部汁液有关,研究显示KCl浓度的增加可导致木质部导水率增加10%[45].枝条形态学基部导管直径通常高于形态学末端[8,40],这一形态结构有利于减小由于植物高度增长带来的水力阻力[8].通常情况下导管直径表现为主干直径>侧枝>枝条交叉点[40].植物体的枝条交叉点、根茎过渡区和节点部位的导管直径较小、导水率较小、胡伯尔值较大和抗栓塞能力较强,这些部位常被称为“水流限制区”和“安全区”[46].胡伯尔值越大,表示维持单位叶面积水分需要的木质部越多,可用于比较不同树种间的水力特征[19,40]. ...
Drought-induced shoot dieback starts with massive root xylem embolism and variable depletion of nonstructural carbohydrates in seedlings of two tree species
5
2017
... 随着干旱胁迫程度的增加,植物的蒸腾张力超出一定临界值,导致其木质部导管中局部水柱断裂,产生空穴化并扩张,进而出现木质部栓塞现象[10].木质部栓塞的进一步发展会导致叶片光合速率降低和冠层枯梢,甚至整个植株因水力失败而死亡[10-11].目前关于导致植物死亡的假说主要包括水力失败、碳饥饿和生物攻击假说[11-12],且越来越多研究显示各假说之间存在交互作用[11].在目前全球气候变化大背景下,越来越多的研究都在关注植物死亡机理及过程,而关于植物如何忍耐干旱的研究较少[6].本文综述了国内外关于植物根、茎、叶不同器官在干旱胁迫条件下的形态调整、水力传导过程及其调整策略,可为揭示植物应对干旱的水力策略提供理论基础,对干旱区生态保护和植被恢复具有重要的实践意义. ...
... [11-12],且越来越多研究显示各假说之间存在交互作用[11].在目前全球气候变化大背景下,越来越多的研究都在关注植物死亡机理及过程,而关于植物如何忍耐干旱的研究较少[6].本文综述了国内外关于植物根、茎、叶不同器官在干旱胁迫条件下的形态调整、水力传导过程及其调整策略,可为揭示植物应对干旱的水力策略提供理论基础,对干旱区生态保护和植被恢复具有重要的实践意义. ...
... [11].在目前全球气候变化大背景下,越来越多的研究都在关注植物死亡机理及过程,而关于植物如何忍耐干旱的研究较少[6].本文综述了国内外关于植物根、茎、叶不同器官在干旱胁迫条件下的形态调整、水力传导过程及其调整策略,可为揭示植物应对干旱的水力策略提供理论基础,对干旱区生态保护和植被恢复具有重要的实践意义. ...
... 不同的干旱强度和时间会影响植物水分运输,在长期的低强度干旱胁迫下,植物水势由于长期处于临界水平,容易死于碳饥饿[23];在短期的严重干旱胁迫下,植物个体易因为水力失败而脱水死亡.目前关于植物水力传导的研究多针对某一过程进行研究,而针对整株植物水力传导过程及不同器官水力性状间的耦合研究较少.植物木质部水力失败是导致植物生理功能失调的主要原因[11].植物水力失败包括3个阶段:一是气孔调节阶段(水力导度降低12%以上对应的水势,>),植物可通过气孔导度调节来减少水分散失[24-25].二是叶片凋落阶段(水力导度降低值12%~88%对应水势),当水势低于时,气孔完全闭合,空气开始从纹孔结构进入导管;当水势低于(水力导度降低50%对应的水势)时,栓塞在木质部中快速扩散影响水分运输[26-27];此后,植株主要通过叶表皮和茎表皮等散失掉活组织、细胞质和细胞间隙中的水分,并通过叶片脱落来减少水分散失[24,28].三是植物死亡阶段(<),造成植物水力系统不可逆转的损伤,并导致植物死亡[24,29].为了保证植物水力系统的完整性,以下将从根系吸水、木质部导水和叶片蒸腾失水3个方面探究植物对干旱胁迫的响应(图2). ...
... 植物远端叶片水力阻力较大,占全植株水力阻力的30%,是SPAC水力运输的瓶颈和“安全阀”[9,19,26,47].植物叶片蒸腾约占地球表面蒸腾的70%[26],植物叶片通常通过气孔开闭适应环境.叶片气孔变化主要受两个机制调节:一是由叶表皮细胞和保卫细胞之间的水力连接引发的被动调节机制;二是激素如脱落酸的主动调节机制[4].气孔对水力调节和激素调节的敏感度与植物的等水调节和非等水调节有关[4],前者是指植物叶片气孔在干旱胁迫下迅速关闭以保证植物叶片日最小水势保持不变,在水分充足时复水快,易死于碳饥饿;后者是指植物叶片气孔在干旱胁迫下仍保持打开,降低叶片日最小水势,在水分充足时复水慢,易死于水力失败[6,11-12,48].有趣的是,Kannenberg等[6]认为非等水调节植物在水分胁迫期间并没有增加碳同化量.在自然界中没有严格的等水植物和非等水植物,通常介于两者之间[6].当土壤处于中等干旱条件下,植物通常会趋向于非等水调节;而当植物处于严重干旱条件下,植物趋向于等水调节[6,49].植物的等水和非等水程度的量化常采用水力学面积,即测量边界叶水势和1∶1线围成的多边形面积,水力学面积越小越趋向于等水调节[50];也可采用凌晨和午间水势的斜率来表示等水程度,斜率接近0表示植物偏向于等水植物,接近1或大于1表示植物偏向于非等水植物[51].陈嘉嘉[52]研究结果显示,在腾格里沙漠生态系统中,常见的固沙植物油蒿(Artemisia ordosica)、柠条锦鸡儿(Caragana korshinskii)和花棒(Hedysarum scoparium)的凌晨和午间水势斜率分别是0.33、0.92和1.22,分别属于偏等水植物、偏非等水植物和严格非等水植物.植物叶片除了通过气孔开合应对轻微水分胁迫,也可改变叶片解剖形态特征是对干旱胁迫做出的短期快速响应,如叶片气孔在干旱条件下呈小而密的分布(图2),可通过增加气孔闭合能力来减少蒸腾过程中的水分散失[44-45]. ...
Mechanisms of plant survival and mortality during drought: why do some plants survive while others succumb to drought
2
2008
... 随着干旱胁迫程度的增加,植物的蒸腾张力超出一定临界值,导致其木质部导管中局部水柱断裂,产生空穴化并扩张,进而出现木质部栓塞现象[10].木质部栓塞的进一步发展会导致叶片光合速率降低和冠层枯梢,甚至整个植株因水力失败而死亡[10-11].目前关于导致植物死亡的假说主要包括水力失败、碳饥饿和生物攻击假说[11-12],且越来越多研究显示各假说之间存在交互作用[11].在目前全球气候变化大背景下,越来越多的研究都在关注植物死亡机理及过程,而关于植物如何忍耐干旱的研究较少[6].本文综述了国内外关于植物根、茎、叶不同器官在干旱胁迫条件下的形态调整、水力传导过程及其调整策略,可为揭示植物应对干旱的水力策略提供理论基础,对干旱区生态保护和植被恢复具有重要的实践意义. ...
... 植物远端叶片水力阻力较大,占全植株水力阻力的30%,是SPAC水力运输的瓶颈和“安全阀”[9,19,26,47].植物叶片蒸腾约占地球表面蒸腾的70%[26],植物叶片通常通过气孔开闭适应环境.叶片气孔变化主要受两个机制调节:一是由叶表皮细胞和保卫细胞之间的水力连接引发的被动调节机制;二是激素如脱落酸的主动调节机制[4].气孔对水力调节和激素调节的敏感度与植物的等水调节和非等水调节有关[4],前者是指植物叶片气孔在干旱胁迫下迅速关闭以保证植物叶片日最小水势保持不变,在水分充足时复水快,易死于碳饥饿;后者是指植物叶片气孔在干旱胁迫下仍保持打开,降低叶片日最小水势,在水分充足时复水慢,易死于水力失败[6,11-12,48].有趣的是,Kannenberg等[6]认为非等水调节植物在水分胁迫期间并没有增加碳同化量.在自然界中没有严格的等水植物和非等水植物,通常介于两者之间[6].当土壤处于中等干旱条件下,植物通常会趋向于非等水调节;而当植物处于严重干旱条件下,植物趋向于等水调节[6,49].植物的等水和非等水程度的量化常采用水力学面积,即测量边界叶水势和1∶1线围成的多边形面积,水力学面积越小越趋向于等水调节[50];也可采用凌晨和午间水势的斜率来表示等水程度,斜率接近0表示植物偏向于等水植物,接近1或大于1表示植物偏向于非等水植物[51].陈嘉嘉[52]研究结果显示,在腾格里沙漠生态系统中,常见的固沙植物油蒿(Artemisia ordosica)、柠条锦鸡儿(Caragana korshinskii)和花棒(Hedysarum scoparium)的凌晨和午间水势斜率分别是0.33、0.92和1.22,分别属于偏等水植物、偏非等水植物和严格非等水植物.植物叶片除了通过气孔开合应对轻微水分胁迫,也可改变叶片解剖形态特征是对干旱胁迫做出的短期快速响应,如叶片气孔在干旱条件下呈小而密的分布(图2),可通过增加气孔闭合能力来减少蒸腾过程中的水分散失[44-45]. ...
Growing-season temperature and precipitation are independent drivers of global variation in xylem hydraulic conductivity
6
2020
... 在目前的温度升高和极端降水事件频发的全球气候变化背景下,近10年来中国干旱区平均气温升高0.5 ℃,且降水减少4 mm,呈“暖干化”的变化趋势,气温和降水变化可能会导致干旱区植物死亡[13].量化植物水力导度和水力失败的临界水势值等水力特征有助于预测在未来气候变化背景下植物个体存活、群落组成、物种分布和生态系统功能[13].植物水力传导与植物形态特征息息相关.植物常增加最大根系分布深度和根冠比以增加根系吸水(图1)[14-16],同时降低根系直径,并增加根系表面积和长度以响应干旱胁迫[17].McDowell等[18]基于达西水力推论指出,植物可通过降低高度以适应在气候变化背景下的植物高蒸腾需求;植物茎干通常会减少单位水运输量的木质部横截面投资(地径减小)以应对干旱胁迫[13].植物叶片在干旱胁迫下较厚(高比叶重),往往具有较长的叶肉扩散路径,这增加了叶片木质部外水分移动的阻力,并降低了水力传导性[19],避免植物过多的叶片蒸腾失水导致水力失败[13];植物叶片也可通过落叶和减小叶面积以降低单位边材面积对应的叶面积来保证水平衡.在干旱胁迫下,植物碳投入成本较高的主干和主根通常会舍弃碳投入成本低、水分运输效率低和抗栓塞性低的远端叶片(即落叶)和根系以保证其水力传输(图1),即水力脆弱性分割假说[9].该假说包括水力分割假说和脆弱性分割假说.水力分割假说认为远端器官(根和叶等)水力阻力较大;脆弱性假说认为当面对水势下降时,远端器官(根和叶等)相比于基部主干等器官更脆弱且容易发生空化[7,9,20].Pivovaroff等[9]研究显示半干旱的地中海气候下灌木的叶片水力导度和茎干水力导度负相关,证实了叶片通过增加水力阻力和降低水力导度来保护茎干免受灾难性的水力失败.值得一提的是,水力脆弱性分割现象多发生在受水分限制的干旱、半干旱和季节性干旱地区,湿润地区的植物如常绿阔叶树种由于其叶片碳投入成本昂贵,茎叶的抗栓塞性较强,发生脆弱性分割的可能性较小甚至不存在[21-22]. ...
... [13].植物水力传导与植物形态特征息息相关.植物常增加最大根系分布深度和根冠比以增加根系吸水(图1)[14-16],同时降低根系直径,并增加根系表面积和长度以响应干旱胁迫[17].McDowell等[18]基于达西水力推论指出,植物可通过降低高度以适应在气候变化背景下的植物高蒸腾需求;植物茎干通常会减少单位水运输量的木质部横截面投资(地径减小)以应对干旱胁迫[13].植物叶片在干旱胁迫下较厚(高比叶重),往往具有较长的叶肉扩散路径,这增加了叶片木质部外水分移动的阻力,并降低了水力传导性[19],避免植物过多的叶片蒸腾失水导致水力失败[13];植物叶片也可通过落叶和减小叶面积以降低单位边材面积对应的叶面积来保证水平衡.在干旱胁迫下,植物碳投入成本较高的主干和主根通常会舍弃碳投入成本低、水分运输效率低和抗栓塞性低的远端叶片(即落叶)和根系以保证其水力传输(图1),即水力脆弱性分割假说[9].该假说包括水力分割假说和脆弱性分割假说.水力分割假说认为远端器官(根和叶等)水力阻力较大;脆弱性假说认为当面对水势下降时,远端器官(根和叶等)相比于基部主干等器官更脆弱且容易发生空化[7,9,20].Pivovaroff等[9]研究显示半干旱的地中海气候下灌木的叶片水力导度和茎干水力导度负相关,证实了叶片通过增加水力阻力和降低水力导度来保护茎干免受灾难性的水力失败.值得一提的是,水力脆弱性分割现象多发生在受水分限制的干旱、半干旱和季节性干旱地区,湿润地区的植物如常绿阔叶树种由于其叶片碳投入成本昂贵,茎叶的抗栓塞性较强,发生脆弱性分割的可能性较小甚至不存在[21-22]. ...
... [13].植物叶片在干旱胁迫下较厚(高比叶重),往往具有较长的叶肉扩散路径,这增加了叶片木质部外水分移动的阻力,并降低了水力传导性[19],避免植物过多的叶片蒸腾失水导致水力失败[13];植物叶片也可通过落叶和减小叶面积以降低单位边材面积对应的叶面积来保证水平衡.在干旱胁迫下,植物碳投入成本较高的主干和主根通常会舍弃碳投入成本低、水分运输效率低和抗栓塞性低的远端叶片(即落叶)和根系以保证其水力传输(图1),即水力脆弱性分割假说[9].该假说包括水力分割假说和脆弱性分割假说.水力分割假说认为远端器官(根和叶等)水力阻力较大;脆弱性假说认为当面对水势下降时,远端器官(根和叶等)相比于基部主干等器官更脆弱且容易发生空化[7,9,20].Pivovaroff等[9]研究显示半干旱的地中海气候下灌木的叶片水力导度和茎干水力导度负相关,证实了叶片通过增加水力阻力和降低水力导度来保护茎干免受灾难性的水力失败.值得一提的是,水力脆弱性分割现象多发生在受水分限制的干旱、半干旱和季节性干旱地区,湿润地区的植物如常绿阔叶树种由于其叶片碳投入成本昂贵,茎叶的抗栓塞性较强,发生脆弱性分割的可能性较小甚至不存在[21-22]. ...
... [13];植物叶片也可通过落叶和减小叶面积以降低单位边材面积对应的叶面积来保证水平衡.在干旱胁迫下,植物碳投入成本较高的主干和主根通常会舍弃碳投入成本低、水分运输效率低和抗栓塞性低的远端叶片(即落叶)和根系以保证其水力传输(图1),即水力脆弱性分割假说[9].该假说包括水力分割假说和脆弱性分割假说.水力分割假说认为远端器官(根和叶等)水力阻力较大;脆弱性假说认为当面对水势下降时,远端器官(根和叶等)相比于基部主干等器官更脆弱且容易发生空化[7,9,20].Pivovaroff等[9]研究显示半干旱的地中海气候下灌木的叶片水力导度和茎干水力导度负相关,证实了叶片通过增加水力阻力和降低水力导度来保护茎干免受灾难性的水力失败.值得一提的是,水力脆弱性分割现象多发生在受水分限制的干旱、半干旱和季节性干旱地区,湿润地区的植物如常绿阔叶树种由于其叶片碳投入成本昂贵,茎叶的抗栓塞性较强,发生脆弱性分割的可能性较小甚至不存在[21-22]. ...
... 植物木质部可通过改变导管结构特征来应对干旱胁迫.木质部常通过减小导管直径,增加导管密度、纹孔膜厚度和木材密度,进而降低木质部导水率并提高水力安全性,且已有大量研究证实(图2)[13,42,44].He等[13]指出相比于湿润环境,在干旱环境下水力失败风险较大且维护成本较高,植物通常会降低导水率以适应环境.Schuldt等[42]发现欧洲山毛榉的木质部导管直径降低7%且纹孔膜厚度增加15%以应对干旱胁迫.导管结构特征影响木质部导水率,Hagen-Poiseuille水力传导方程显示导管导水率与导管直径的4次方成正比,因此导管直径的微小增加会带来导水率的显著提高[41].有研究采用X射线显微断层成像法观测干燥不同时间的赤桉的木质部横截面的导管栓塞情况,结果显示Hagen-Poiseuille水力传导方程模拟的理论导水率高于实际最大导水率[10],原因一是与没有考虑导管长度和导管间的纹孔结构有关[41],导管长度越短、纹孔数量越少和纹孔面积越小,则导管导水率越低[41],纹孔阻力约占整个植株总水力传导阻力的14%~84%[40];二是与导管腔的不规则形状和锥度有关[45];三是与木质部汁液有关,研究显示KCl浓度的增加可导致木质部导水率增加10%[45].枝条形态学基部导管直径通常高于形态学末端[8,40],这一形态结构有利于减小由于植物高度增长带来的水力阻力[8].通常情况下导管直径表现为主干直径>侧枝>枝条交叉点[40].植物体的枝条交叉点、根茎过渡区和节点部位的导管直径较小、导水率较小、胡伯尔值较大和抗栓塞能力较强,这些部位常被称为“水流限制区”和“安全区”[46].胡伯尔值越大,表示维持单位叶面积水分需要的木质部越多,可用于比较不同树种间的水力特征[19,40]. ...
... [13]指出相比于湿润环境,在干旱环境下水力失败风险较大且维护成本较高,植物通常会降低导水率以适应环境.Schuldt等[42]发现欧洲山毛榉的木质部导管直径降低7%且纹孔膜厚度增加15%以应对干旱胁迫.导管结构特征影响木质部导水率,Hagen-Poiseuille水力传导方程显示导管导水率与导管直径的4次方成正比,因此导管直径的微小增加会带来导水率的显著提高[41].有研究采用X射线显微断层成像法观测干燥不同时间的赤桉的木质部横截面的导管栓塞情况,结果显示Hagen-Poiseuille水力传导方程模拟的理论导水率高于实际最大导水率[10],原因一是与没有考虑导管长度和导管间的纹孔结构有关[41],导管长度越短、纹孔数量越少和纹孔面积越小,则导管导水率越低[41],纹孔阻力约占整个植株总水力传导阻力的14%~84%[40];二是与导管腔的不规则形状和锥度有关[45];三是与木质部汁液有关,研究显示KCl浓度的增加可导致木质部导水率增加10%[45].枝条形态学基部导管直径通常高于形态学末端[8,40],这一形态结构有利于减小由于植物高度增长带来的水力阻力[8].通常情况下导管直径表现为主干直径>侧枝>枝条交叉点[40].植物体的枝条交叉点、根茎过渡区和节点部位的导管直径较小、导水率较小、胡伯尔值较大和抗栓塞能力较强,这些部位常被称为“水流限制区”和“安全区”[46].胡伯尔值越大,表示维持单位叶面积水分需要的木质部越多,可用于比较不同树种间的水力特征[19,40]. ...
Rotation crop root performance and its effect on soil hydraulic properties in a clayey Utisol
1
2021
... 在目前的温度升高和极端降水事件频发的全球气候变化背景下,近10年来中国干旱区平均气温升高0.5 ℃,且降水减少4 mm,呈“暖干化”的变化趋势,气温和降水变化可能会导致干旱区植物死亡[13].量化植物水力导度和水力失败的临界水势值等水力特征有助于预测在未来气候变化背景下植物个体存活、群落组成、物种分布和生态系统功能[13].植物水力传导与植物形态特征息息相关.植物常增加最大根系分布深度和根冠比以增加根系吸水(图1)[14-16],同时降低根系直径,并增加根系表面积和长度以响应干旱胁迫[17].McDowell等[18]基于达西水力推论指出,植物可通过降低高度以适应在气候变化背景下的植物高蒸腾需求;植物茎干通常会减少单位水运输量的木质部横截面投资(地径减小)以应对干旱胁迫[13].植物叶片在干旱胁迫下较厚(高比叶重),往往具有较长的叶肉扩散路径,这增加了叶片木质部外水分移动的阻力,并降低了水力传导性[19],避免植物过多的叶片蒸腾失水导致水力失败[13];植物叶片也可通过落叶和减小叶面积以降低单位边材面积对应的叶面积来保证水平衡.在干旱胁迫下,植物碳投入成本较高的主干和主根通常会舍弃碳投入成本低、水分运输效率低和抗栓塞性低的远端叶片(即落叶)和根系以保证其水力传输(图1),即水力脆弱性分割假说[9].该假说包括水力分割假说和脆弱性分割假说.水力分割假说认为远端器官(根和叶等)水力阻力较大;脆弱性假说认为当面对水势下降时,远端器官(根和叶等)相比于基部主干等器官更脆弱且容易发生空化[7,9,20].Pivovaroff等[9]研究显示半干旱的地中海气候下灌木的叶片水力导度和茎干水力导度负相关,证实了叶片通过增加水力阻力和降低水力导度来保护茎干免受灾难性的水力失败.值得一提的是,水力脆弱性分割现象多发生在受水分限制的干旱、半干旱和季节性干旱地区,湿润地区的植物如常绿阔叶树种由于其叶片碳投入成本昂贵,茎叶的抗栓塞性较强,发生脆弱性分割的可能性较小甚至不存在[21-22]. ...
Understanding plant rooting patterns in semi-arid systems: an integrated model analysis of climate,soil type and plant biomass
2011
Spatiotemporal variation of soil water potential and its significance to water balance for a desert shrub area
5
2022
... 在目前的温度升高和极端降水事件频发的全球气候变化背景下,近10年来中国干旱区平均气温升高0.5 ℃,且降水减少4 mm,呈“暖干化”的变化趋势,气温和降水变化可能会导致干旱区植物死亡[13].量化植物水力导度和水力失败的临界水势值等水力特征有助于预测在未来气候变化背景下植物个体存活、群落组成、物种分布和生态系统功能[13].植物水力传导与植物形态特征息息相关.植物常增加最大根系分布深度和根冠比以增加根系吸水(图1)[14-16],同时降低根系直径,并增加根系表面积和长度以响应干旱胁迫[17].McDowell等[18]基于达西水力推论指出,植物可通过降低高度以适应在气候变化背景下的植物高蒸腾需求;植物茎干通常会减少单位水运输量的木质部横截面投资(地径减小)以应对干旱胁迫[13].植物叶片在干旱胁迫下较厚(高比叶重),往往具有较长的叶肉扩散路径,这增加了叶片木质部外水分移动的阻力,并降低了水力传导性[19],避免植物过多的叶片蒸腾失水导致水力失败[13];植物叶片也可通过落叶和减小叶面积以降低单位边材面积对应的叶面积来保证水平衡.在干旱胁迫下,植物碳投入成本较高的主干和主根通常会舍弃碳投入成本低、水分运输效率低和抗栓塞性低的远端叶片(即落叶)和根系以保证其水力传输(图1),即水力脆弱性分割假说[9].该假说包括水力分割假说和脆弱性分割假说.水力分割假说认为远端器官(根和叶等)水力阻力较大;脆弱性假说认为当面对水势下降时,远端器官(根和叶等)相比于基部主干等器官更脆弱且容易发生空化[7,9,20].Pivovaroff等[9]研究显示半干旱的地中海气候下灌木的叶片水力导度和茎干水力导度负相关,证实了叶片通过增加水力阻力和降低水力导度来保护茎干免受灾难性的水力失败.值得一提的是,水力脆弱性分割现象多发生在受水分限制的干旱、半干旱和季节性干旱地区,湿润地区的植物如常绿阔叶树种由于其叶片碳投入成本昂贵,茎叶的抗栓塞性较强,发生脆弱性分割的可能性较小甚至不存在[21-22]. ...
... 水力再分配是植物根系应对干旱的重要策略,能够促进植物利用土壤水分的生态位分化,改变植被分布格局[30].当土壤水势低于植物叶片水势,且根际土壤水分存在明显的梯度时,土壤剖面存在根系吸收湿润区水分运输至干旱区的现象,影响土壤水分环境,即水力再分配[16,31].水力再分配包括水力提升、水力下传和侧向再分配3个过程,分别是指植物根系对水分向上、向下和侧向的运输[16,32].水力再分配有利于土壤水分运输,缓解土壤干旱胁迫,防止根系木质部水力失败[16],促进养分运输和提高根际微生物活动等[31].近年来,大量研究显示存在根系向土壤释水现象,如夜间根系向土壤中释放水分以弥补白天植物耗水[16,33-34];在同一土壤剖面内生长在湿润土壤的根系吸水增加量(100%)高于整株植物蒸散量的增加量(40%),即补偿性根系吸水[31,35];在干旱条件下根际土壤水分高于周围土壤水分,在湿润情况时则情况相反[33,36-37].值得一提的是,当土壤极端干旱时,过低的土壤水势阻碍了根系释水过程[38],根毛可能会因为张力过大而断裂[39]. ...
... [16,32].水力再分配有利于土壤水分运输,缓解土壤干旱胁迫,防止根系木质部水力失败[16],促进养分运输和提高根际微生物活动等[31].近年来,大量研究显示存在根系向土壤释水现象,如夜间根系向土壤中释放水分以弥补白天植物耗水[16,33-34];在同一土壤剖面内生长在湿润土壤的根系吸水增加量(100%)高于整株植物蒸散量的增加量(40%),即补偿性根系吸水[31,35];在干旱条件下根际土壤水分高于周围土壤水分,在湿润情况时则情况相反[33,36-37].值得一提的是,当土壤极端干旱时,过低的土壤水势阻碍了根系释水过程[38],根毛可能会因为张力过大而断裂[39]. ...
... [16],促进养分运输和提高根际微生物活动等[31].近年来,大量研究显示存在根系向土壤释水现象,如夜间根系向土壤中释放水分以弥补白天植物耗水[16,33-34];在同一土壤剖面内生长在湿润土壤的根系吸水增加量(100%)高于整株植物蒸散量的增加量(40%),即补偿性根系吸水[31,35];在干旱条件下根际土壤水分高于周围土壤水分,在湿润情况时则情况相反[33,36-37].值得一提的是,当土壤极端干旱时,过低的土壤水势阻碍了根系释水过程[38],根毛可能会因为张力过大而断裂[39]. ...
... [16,33-34];在同一土壤剖面内生长在湿润土壤的根系吸水增加量(100%)高于整株植物蒸散量的增加量(40%),即补偿性根系吸水[31,35];在干旱条件下根际土壤水分高于周围土壤水分,在湿润情况时则情况相反[33,36-37].值得一提的是,当土壤极端干旱时,过低的土壤水势阻碍了根系释水过程[38],根毛可能会因为张力过大而断裂[39]. ...
水分胁迫对土壤-刺槐系统水力特性的影响
1
2021
... 在目前的温度升高和极端降水事件频发的全球气候变化背景下,近10年来中国干旱区平均气温升高0.5 ℃,且降水减少4 mm,呈“暖干化”的变化趋势,气温和降水变化可能会导致干旱区植物死亡[13].量化植物水力导度和水力失败的临界水势值等水力特征有助于预测在未来气候变化背景下植物个体存活、群落组成、物种分布和生态系统功能[13].植物水力传导与植物形态特征息息相关.植物常增加最大根系分布深度和根冠比以增加根系吸水(图1)[14-16],同时降低根系直径,并增加根系表面积和长度以响应干旱胁迫[17].McDowell等[18]基于达西水力推论指出,植物可通过降低高度以适应在气候变化背景下的植物高蒸腾需求;植物茎干通常会减少单位水运输量的木质部横截面投资(地径减小)以应对干旱胁迫[13].植物叶片在干旱胁迫下较厚(高比叶重),往往具有较长的叶肉扩散路径,这增加了叶片木质部外水分移动的阻力,并降低了水力传导性[19],避免植物过多的叶片蒸腾失水导致水力失败[13];植物叶片也可通过落叶和减小叶面积以降低单位边材面积对应的叶面积来保证水平衡.在干旱胁迫下,植物碳投入成本较高的主干和主根通常会舍弃碳投入成本低、水分运输效率低和抗栓塞性低的远端叶片(即落叶)和根系以保证其水力传输(图1),即水力脆弱性分割假说[9].该假说包括水力分割假说和脆弱性分割假说.水力分割假说认为远端器官(根和叶等)水力阻力较大;脆弱性假说认为当面对水势下降时,远端器官(根和叶等)相比于基部主干等器官更脆弱且容易发生空化[7,9,20].Pivovaroff等[9]研究显示半干旱的地中海气候下灌木的叶片水力导度和茎干水力导度负相关,证实了叶片通过增加水力阻力和降低水力导度来保护茎干免受灾难性的水力失败.值得一提的是,水力脆弱性分割现象多发生在受水分限制的干旱、半干旱和季节性干旱地区,湿润地区的植物如常绿阔叶树种由于其叶片碳投入成本昂贵,茎叶的抗栓塞性较强,发生脆弱性分割的可能性较小甚至不存在[21-22]. ...
Darcy's law predicts widespread forest mortality under climate warming
2
2015
... 在目前的温度升高和极端降水事件频发的全球气候变化背景下,近10年来中国干旱区平均气温升高0.5 ℃,且降水减少4 mm,呈“暖干化”的变化趋势,气温和降水变化可能会导致干旱区植物死亡[13].量化植物水力导度和水力失败的临界水势值等水力特征有助于预测在未来气候变化背景下植物个体存活、群落组成、物种分布和生态系统功能[13].植物水力传导与植物形态特征息息相关.植物常增加最大根系分布深度和根冠比以增加根系吸水(图1)[14-16],同时降低根系直径,并增加根系表面积和长度以响应干旱胁迫[17].McDowell等[18]基于达西水力推论指出,植物可通过降低高度以适应在气候变化背景下的植物高蒸腾需求;植物茎干通常会减少单位水运输量的木质部横截面投资(地径减小)以应对干旱胁迫[13].植物叶片在干旱胁迫下较厚(高比叶重),往往具有较长的叶肉扩散路径,这增加了叶片木质部外水分移动的阻力,并降低了水力传导性[19],避免植物过多的叶片蒸腾失水导致水力失败[13];植物叶片也可通过落叶和减小叶面积以降低单位边材面积对应的叶面积来保证水平衡.在干旱胁迫下,植物碳投入成本较高的主干和主根通常会舍弃碳投入成本低、水分运输效率低和抗栓塞性低的远端叶片(即落叶)和根系以保证其水力传输(图1),即水力脆弱性分割假说[9].该假说包括水力分割假说和脆弱性分割假说.水力分割假说认为远端器官(根和叶等)水力阻力较大;脆弱性假说认为当面对水势下降时,远端器官(根和叶等)相比于基部主干等器官更脆弱且容易发生空化[7,9,20].Pivovaroff等[9]研究显示半干旱的地中海气候下灌木的叶片水力导度和茎干水力导度负相关,证实了叶片通过增加水力阻力和降低水力导度来保护茎干免受灾难性的水力失败.值得一提的是,水力脆弱性分割现象多发生在受水分限制的干旱、半干旱和季节性干旱地区,湿润地区的植物如常绿阔叶树种由于其叶片碳投入成本昂贵,茎叶的抗栓塞性较强,发生脆弱性分割的可能性较小甚至不存在[21-22]. ...
... 植物个体水力传导过程包括植物根系吸水、木质部导水和叶片蒸腾失水3个过程,该过程受气象因子、土壤水分有效性和植物水力特性的共同影响.关于SPAC水分传输的模型较多,而基于过程的模型较少.大叶模型将SPAC中植物简化为一片叶片,仅表征土壤水分胁迫与植物叶片蒸腾和光合之间的关系,忽略了植物根系吸水和茎干导水的水力传导过程[57].由于根系的不可见性和难获取性,直接测量根系吸水具有一定困难,建立基于过程的模型准确模拟根系吸水有助于了解土壤水文和植物生长[58-60].根系吸水模拟常采用Feddes模型,其运行需要潜在根系吸水量、水分胁迫函数和根系分布密度函数等,常将其与Richards土壤水分运移模型结合起来使用(即Hydrus模型),用于准确模拟根系吸水量、水分通量和土壤水分等重要水分参数以及植物生长[58].Wang等[59]采用Hydrus模型指出根系吸水量减少和土壤蒸发量增加是阻碍夏玉米在斥水性土壤减产中的主要原因.Tu等[60]采用Hydrus模型模拟农作物在生长季的蒸散量和深层渗漏量,用于指导农业灌溉用水.Sperry模型有效提高了根系吸水、冠层蒸腾和叶片水势间的耦合性,提供了模拟SPAC水分传输的机理性[61-62].由于测量林分尺度上所有植物的单株蒸腾较不现实,因此可通过测量植物株高、冠幅、叶面积指数、地径和占地面积等作为尺度扩展因子将单株蒸腾从个体尺度扩展至林分尺度,目前采用边材面积作为扩展因子推导林分蒸腾较为准确[18,61],边材面积的量化可采用边材面积与胸径之间的经验函数来表示[61]. ...
Coordination of plant hydraulic and photosynthetic traits: confronting optimality theory with field measurements
3
2021
... 在目前的温度升高和极端降水事件频发的全球气候变化背景下,近10年来中国干旱区平均气温升高0.5 ℃,且降水减少4 mm,呈“暖干化”的变化趋势,气温和降水变化可能会导致干旱区植物死亡[13].量化植物水力导度和水力失败的临界水势值等水力特征有助于预测在未来气候变化背景下植物个体存活、群落组成、物种分布和生态系统功能[13].植物水力传导与植物形态特征息息相关.植物常增加最大根系分布深度和根冠比以增加根系吸水(图1)[14-16],同时降低根系直径,并增加根系表面积和长度以响应干旱胁迫[17].McDowell等[18]基于达西水力推论指出,植物可通过降低高度以适应在气候变化背景下的植物高蒸腾需求;植物茎干通常会减少单位水运输量的木质部横截面投资(地径减小)以应对干旱胁迫[13].植物叶片在干旱胁迫下较厚(高比叶重),往往具有较长的叶肉扩散路径,这增加了叶片木质部外水分移动的阻力,并降低了水力传导性[19],避免植物过多的叶片蒸腾失水导致水力失败[13];植物叶片也可通过落叶和减小叶面积以降低单位边材面积对应的叶面积来保证水平衡.在干旱胁迫下,植物碳投入成本较高的主干和主根通常会舍弃碳投入成本低、水分运输效率低和抗栓塞性低的远端叶片(即落叶)和根系以保证其水力传输(图1),即水力脆弱性分割假说[9].该假说包括水力分割假说和脆弱性分割假说.水力分割假说认为远端器官(根和叶等)水力阻力较大;脆弱性假说认为当面对水势下降时,远端器官(根和叶等)相比于基部主干等器官更脆弱且容易发生空化[7,9,20].Pivovaroff等[9]研究显示半干旱的地中海气候下灌木的叶片水力导度和茎干水力导度负相关,证实了叶片通过增加水力阻力和降低水力导度来保护茎干免受灾难性的水力失败.值得一提的是,水力脆弱性分割现象多发生在受水分限制的干旱、半干旱和季节性干旱地区,湿润地区的植物如常绿阔叶树种由于其叶片碳投入成本昂贵,茎叶的抗栓塞性较强,发生脆弱性分割的可能性较小甚至不存在[21-22]. ...
... 植物木质部可通过改变导管结构特征来应对干旱胁迫.木质部常通过减小导管直径,增加导管密度、纹孔膜厚度和木材密度,进而降低木质部导水率并提高水力安全性,且已有大量研究证实(图2)[13,42,44].He等[13]指出相比于湿润环境,在干旱环境下水力失败风险较大且维护成本较高,植物通常会降低导水率以适应环境.Schuldt等[42]发现欧洲山毛榉的木质部导管直径降低7%且纹孔膜厚度增加15%以应对干旱胁迫.导管结构特征影响木质部导水率,Hagen-Poiseuille水力传导方程显示导管导水率与导管直径的4次方成正比,因此导管直径的微小增加会带来导水率的显著提高[41].有研究采用X射线显微断层成像法观测干燥不同时间的赤桉的木质部横截面的导管栓塞情况,结果显示Hagen-Poiseuille水力传导方程模拟的理论导水率高于实际最大导水率[10],原因一是与没有考虑导管长度和导管间的纹孔结构有关[41],导管长度越短、纹孔数量越少和纹孔面积越小,则导管导水率越低[41],纹孔阻力约占整个植株总水力传导阻力的14%~84%[40];二是与导管腔的不规则形状和锥度有关[45];三是与木质部汁液有关,研究显示KCl浓度的增加可导致木质部导水率增加10%[45].枝条形态学基部导管直径通常高于形态学末端[8,40],这一形态结构有利于减小由于植物高度增长带来的水力阻力[8].通常情况下导管直径表现为主干直径>侧枝>枝条交叉点[40].植物体的枝条交叉点、根茎过渡区和节点部位的导管直径较小、导水率较小、胡伯尔值较大和抗栓塞能力较强,这些部位常被称为“水流限制区”和“安全区”[46].胡伯尔值越大,表示维持单位叶面积水分需要的木质部越多,可用于比较不同树种间的水力特征[19,40]. ...
... 植物远端叶片水力阻力较大,占全植株水力阻力的30%,是SPAC水力运输的瓶颈和“安全阀”[9,19,26,47].植物叶片蒸腾约占地球表面蒸腾的70%[26],植物叶片通常通过气孔开闭适应环境.叶片气孔变化主要受两个机制调节:一是由叶表皮细胞和保卫细胞之间的水力连接引发的被动调节机制;二是激素如脱落酸的主动调节机制[4].气孔对水力调节和激素调节的敏感度与植物的等水调节和非等水调节有关[4],前者是指植物叶片气孔在干旱胁迫下迅速关闭以保证植物叶片日最小水势保持不变,在水分充足时复水快,易死于碳饥饿;后者是指植物叶片气孔在干旱胁迫下仍保持打开,降低叶片日最小水势,在水分充足时复水慢,易死于水力失败[6,11-12,48].有趣的是,Kannenberg等[6]认为非等水调节植物在水分胁迫期间并没有增加碳同化量.在自然界中没有严格的等水植物和非等水植物,通常介于两者之间[6].当土壤处于中等干旱条件下,植物通常会趋向于非等水调节;而当植物处于严重干旱条件下,植物趋向于等水调节[6,49].植物的等水和非等水程度的量化常采用水力学面积,即测量边界叶水势和1∶1线围成的多边形面积,水力学面积越小越趋向于等水调节[50];也可采用凌晨和午间水势的斜率来表示等水程度,斜率接近0表示植物偏向于等水植物,接近1或大于1表示植物偏向于非等水植物[51].陈嘉嘉[52]研究结果显示,在腾格里沙漠生态系统中,常见的固沙植物油蒿(Artemisia ordosica)、柠条锦鸡儿(Caragana korshinskii)和花棒(Hedysarum scoparium)的凌晨和午间水势斜率分别是0.33、0.92和1.22,分别属于偏等水植物、偏非等水植物和严格非等水植物.植物叶片除了通过气孔开合应对轻微水分胁迫,也可改变叶片解剖形态特征是对干旱胁迫做出的短期快速响应,如叶片气孔在干旱条件下呈小而密的分布(图2),可通过增加气孔闭合能力来减少蒸腾过程中的水分散失[44-45]. ...
The hydraulic architecture of trees and other woody plants
1
1991
... 在目前的温度升高和极端降水事件频发的全球气候变化背景下,近10年来中国干旱区平均气温升高0.5 ℃,且降水减少4 mm,呈“暖干化”的变化趋势,气温和降水变化可能会导致干旱区植物死亡[13].量化植物水力导度和水力失败的临界水势值等水力特征有助于预测在未来气候变化背景下植物个体存活、群落组成、物种分布和生态系统功能[13].植物水力传导与植物形态特征息息相关.植物常增加最大根系分布深度和根冠比以增加根系吸水(图1)[14-16],同时降低根系直径,并增加根系表面积和长度以响应干旱胁迫[17].McDowell等[18]基于达西水力推论指出,植物可通过降低高度以适应在气候变化背景下的植物高蒸腾需求;植物茎干通常会减少单位水运输量的木质部横截面投资(地径减小)以应对干旱胁迫[13].植物叶片在干旱胁迫下较厚(高比叶重),往往具有较长的叶肉扩散路径,这增加了叶片木质部外水分移动的阻力,并降低了水力传导性[19],避免植物过多的叶片蒸腾失水导致水力失败[13];植物叶片也可通过落叶和减小叶面积以降低单位边材面积对应的叶面积来保证水平衡.在干旱胁迫下,植物碳投入成本较高的主干和主根通常会舍弃碳投入成本低、水分运输效率低和抗栓塞性低的远端叶片(即落叶)和根系以保证其水力传输(图1),即水力脆弱性分割假说[9].该假说包括水力分割假说和脆弱性分割假说.水力分割假说认为远端器官(根和叶等)水力阻力较大;脆弱性假说认为当面对水势下降时,远端器官(根和叶等)相比于基部主干等器官更脆弱且容易发生空化[7,9,20].Pivovaroff等[9]研究显示半干旱的地中海气候下灌木的叶片水力导度和茎干水力导度负相关,证实了叶片通过增加水力阻力和降低水力导度来保护茎干免受灾难性的水力失败.值得一提的是,水力脆弱性分割现象多发生在受水分限制的干旱、半干旱和季节性干旱地区,湿润地区的植物如常绿阔叶树种由于其叶片碳投入成本昂贵,茎叶的抗栓塞性较强,发生脆弱性分割的可能性较小甚至不存在[21-22]. ...
白杨杂交子代栓塞脆弱性分割及与生长的关系
1
2022
... 在目前的温度升高和极端降水事件频发的全球气候变化背景下,近10年来中国干旱区平均气温升高0.5 ℃,且降水减少4 mm,呈“暖干化”的变化趋势,气温和降水变化可能会导致干旱区植物死亡[13].量化植物水力导度和水力失败的临界水势值等水力特征有助于预测在未来气候变化背景下植物个体存活、群落组成、物种分布和生态系统功能[13].植物水力传导与植物形态特征息息相关.植物常增加最大根系分布深度和根冠比以增加根系吸水(图1)[14-16],同时降低根系直径,并增加根系表面积和长度以响应干旱胁迫[17].McDowell等[18]基于达西水力推论指出,植物可通过降低高度以适应在气候变化背景下的植物高蒸腾需求;植物茎干通常会减少单位水运输量的木质部横截面投资(地径减小)以应对干旱胁迫[13].植物叶片在干旱胁迫下较厚(高比叶重),往往具有较长的叶肉扩散路径,这增加了叶片木质部外水分移动的阻力,并降低了水力传导性[19],避免植物过多的叶片蒸腾失水导致水力失败[13];植物叶片也可通过落叶和减小叶面积以降低单位边材面积对应的叶面积来保证水平衡.在干旱胁迫下,植物碳投入成本较高的主干和主根通常会舍弃碳投入成本低、水分运输效率低和抗栓塞性低的远端叶片(即落叶)和根系以保证其水力传输(图1),即水力脆弱性分割假说[9].该假说包括水力分割假说和脆弱性分割假说.水力分割假说认为远端器官(根和叶等)水力阻力较大;脆弱性假说认为当面对水势下降时,远端器官(根和叶等)相比于基部主干等器官更脆弱且容易发生空化[7,9,20].Pivovaroff等[9]研究显示半干旱的地中海气候下灌木的叶片水力导度和茎干水力导度负相关,证实了叶片通过增加水力阻力和降低水力导度来保护茎干免受灾难性的水力失败.值得一提的是,水力脆弱性分割现象多发生在受水分限制的干旱、半干旱和季节性干旱地区,湿润地区的植物如常绿阔叶树种由于其叶片碳投入成本昂贵,茎叶的抗栓塞性较强,发生脆弱性分割的可能性较小甚至不存在[21-22]. ...
Does leaf shedding protect stems from cavitation during seasonal droughts?a test of the hydraulic fuse hypothesis
1
2016
... 在目前的温度升高和极端降水事件频发的全球气候变化背景下,近10年来中国干旱区平均气温升高0.5 ℃,且降水减少4 mm,呈“暖干化”的变化趋势,气温和降水变化可能会导致干旱区植物死亡[13].量化植物水力导度和水力失败的临界水势值等水力特征有助于预测在未来气候变化背景下植物个体存活、群落组成、物种分布和生态系统功能[13].植物水力传导与植物形态特征息息相关.植物常增加最大根系分布深度和根冠比以增加根系吸水(图1)[14-16],同时降低根系直径,并增加根系表面积和长度以响应干旱胁迫[17].McDowell等[18]基于达西水力推论指出,植物可通过降低高度以适应在气候变化背景下的植物高蒸腾需求;植物茎干通常会减少单位水运输量的木质部横截面投资(地径减小)以应对干旱胁迫[13].植物叶片在干旱胁迫下较厚(高比叶重),往往具有较长的叶肉扩散路径,这增加了叶片木质部外水分移动的阻力,并降低了水力传导性[19],避免植物过多的叶片蒸腾失水导致水力失败[13];植物叶片也可通过落叶和减小叶面积以降低单位边材面积对应的叶面积来保证水平衡.在干旱胁迫下,植物碳投入成本较高的主干和主根通常会舍弃碳投入成本低、水分运输效率低和抗栓塞性低的远端叶片(即落叶)和根系以保证其水力传输(图1),即水力脆弱性分割假说[9].该假说包括水力分割假说和脆弱性分割假说.水力分割假说认为远端器官(根和叶等)水力阻力较大;脆弱性假说认为当面对水势下降时,远端器官(根和叶等)相比于基部主干等器官更脆弱且容易发生空化[7,9,20].Pivovaroff等[9]研究显示半干旱的地中海气候下灌木的叶片水力导度和茎干水力导度负相关,证实了叶片通过增加水力阻力和降低水力导度来保护茎干免受灾难性的水力失败.值得一提的是,水力脆弱性分割现象多发生在受水分限制的干旱、半干旱和季节性干旱地区,湿润地区的植物如常绿阔叶树种由于其叶片碳投入成本昂贵,茎叶的抗栓塞性较强,发生脆弱性分割的可能性较小甚至不存在[21-22]. ...
暖温带常见落叶木本植物水分利用策略及干旱死亡机理研究
1
2020
... 不同的干旱强度和时间会影响植物水分运输,在长期的低强度干旱胁迫下,植物水势由于长期处于临界水平,容易死于碳饥饿[23];在短期的严重干旱胁迫下,植物个体易因为水力失败而脱水死亡.目前关于植物水力传导的研究多针对某一过程进行研究,而针对整株植物水力传导过程及不同器官水力性状间的耦合研究较少.植物木质部水力失败是导致植物生理功能失调的主要原因[11].植物水力失败包括3个阶段:一是气孔调节阶段(水力导度降低12%以上对应的水势,>),植物可通过气孔导度调节来减少水分散失[24-25].二是叶片凋落阶段(水力导度降低值12%~88%对应水势),当水势低于时,气孔完全闭合,空气开始从纹孔结构进入导管;当水势低于(水力导度降低50%对应的水势)时,栓塞在木质部中快速扩散影响水分运输[26-27];此后,植株主要通过叶表皮和茎表皮等散失掉活组织、细胞质和细胞间隙中的水分,并通过叶片脱落来减少水分散失[24,28].三是植物死亡阶段(<),造成植物水力系统不可逆转的损伤,并导致植物死亡[24,29].为了保证植物水力系统的完整性,以下将从根系吸水、木质部导水和叶片蒸腾失水3个方面探究植物对干旱胁迫的响应(图2). ...
黄土高原刺槐解剖结构特征及其抗旱生理机制
5
2024
... 不同的干旱强度和时间会影响植物水分运输,在长期的低强度干旱胁迫下,植物水势由于长期处于临界水平,容易死于碳饥饿[23];在短期的严重干旱胁迫下,植物个体易因为水力失败而脱水死亡.目前关于植物水力传导的研究多针对某一过程进行研究,而针对整株植物水力传导过程及不同器官水力性状间的耦合研究较少.植物木质部水力失败是导致植物生理功能失调的主要原因[11].植物水力失败包括3个阶段:一是气孔调节阶段(水力导度降低12%以上对应的水势,>),植物可通过气孔导度调节来减少水分散失[24-25].二是叶片凋落阶段(水力导度降低值12%~88%对应水势),当水势低于时,气孔完全闭合,空气开始从纹孔结构进入导管;当水势低于(水力导度降低50%对应的水势)时,栓塞在木质部中快速扩散影响水分运输[26-27];此后,植株主要通过叶表皮和茎表皮等散失掉活组织、细胞质和细胞间隙中的水分,并通过叶片脱落来减少水分散失[24,28].三是植物死亡阶段(<),造成植物水力系统不可逆转的损伤,并导致植物死亡[24,29].为了保证植物水力系统的完整性,以下将从根系吸水、木质部导水和叶片蒸腾失水3个方面探究植物对干旱胁迫的响应(图2). ...
... [24,28].三是植物死亡阶段(<),造成植物水力系统不可逆转的损伤,并导致植物死亡[24,29].为了保证植物水力系统的完整性,以下将从根系吸水、木质部导水和叶片蒸腾失水3个方面探究植物对干旱胁迫的响应(图2). ...
... [24,29].为了保证植物水力系统的完整性,以下将从根系吸水、木质部导水和叶片蒸腾失水3个方面探究植物对干旱胁迫的响应(图2). ...
... 植物木质部主要包括薄壁细胞、木射线、纤维和导管(管胞)等,薄壁细胞具有储存糖分和水分的作用[24,40],相邻导管间的水分传输通过纹孔结构实现,不相邻导管间的水分传输基于纤维桥假说通过纤维管胞实现[41],纤维具有机械支持作用,导管(管胞)具有水分传输作用.木质部栓塞是植物应对干旱胁迫的被动结果.基于内聚力-张力理论,叶片蒸腾产生的张力大于水分子间的内聚力与水柱和管壁间的附着作用时,木质部导管中水柱断裂并产生气泡,阻碍水分向上输送[7,42-43].气泡的形成可由“气种假说”诠释[43]:①空气泡从纹孔中进入导管;②管道中疏水性区域小气泡膨胀;③木质部中水分流动时,小气泡扩张;④导管壁中小气泡与疏水区脱离.目前关于栓塞的发生主要有两种观点,一种观点指出木质部的空穴和栓塞现象是植物对干旱的响应,在自然界经常发生[43];另一种观点是栓塞只有在极端干旱胁迫下才偶尔发生,且栓塞在水分充足时会被修复[26].木质部栓塞程度随干旱程度增加而增加,常采用栓塞脆弱性曲线来表示[10].木质部抗栓塞性会随植物种类和枝条年龄而异.成年植物个体的木质部有更多的纤维组织来确保机械支撑和水力安全;幼年植物个体的木质部通常会优先形成射线薄壁组织和轴向薄壁组织来进行水分和营养存储[24].Nolf等[10]指出赤桉(Eucalyptus camaldulensis)的当年生枝条相比于老枝更容易受干旱的影响,抗栓塞性较弱;Schuldt等[42]则认为欧洲山毛榉(Fagus sylvatica)生长较慢的老枝比生长较快的新枝的栓塞脆弱性高,可能与老枝空化疲劳有关. ...
... [24].Nolf等[10]指出赤桉(Eucalyptus camaldulensis)的当年生枝条相比于老枝更容易受干旱的影响,抗栓塞性较弱;Schuldt等[42]则认为欧洲山毛榉(Fagus sylvatica)生长较慢的老枝比生长较快的新枝的栓塞脆弱性高,可能与老枝空化疲劳有关. ...
How do stomata respond to water status
1
2019
... 不同的干旱强度和时间会影响植物水分运输,在长期的低强度干旱胁迫下,植物水势由于长期处于临界水平,容易死于碳饥饿[23];在短期的严重干旱胁迫下,植物个体易因为水力失败而脱水死亡.目前关于植物水力传导的研究多针对某一过程进行研究,而针对整株植物水力传导过程及不同器官水力性状间的耦合研究较少.植物木质部水力失败是导致植物生理功能失调的主要原因[11].植物水力失败包括3个阶段:一是气孔调节阶段(水力导度降低12%以上对应的水势,>),植物可通过气孔导度调节来减少水分散失[24-25].二是叶片凋落阶段(水力导度降低值12%~88%对应水势),当水势低于时,气孔完全闭合,空气开始从纹孔结构进入导管;当水势低于(水力导度降低50%对应的水势)时,栓塞在木质部中快速扩散影响水分运输[26-27];此后,植株主要通过叶表皮和茎表皮等散失掉活组织、细胞质和细胞间隙中的水分,并通过叶片脱落来减少水分散失[24,28].三是植物死亡阶段(<),造成植物水力系统不可逆转的损伤,并导致植物死亡[24,29].为了保证植物水力系统的完整性,以下将从根系吸水、木质部导水和叶片蒸腾失水3个方面探究植物对干旱胁迫的响应(图2). ...
干旱胁迫下植物叶水分传导速率与木质部栓塞关系的研究
4
2019
... 不同的干旱强度和时间会影响植物水分运输,在长期的低强度干旱胁迫下,植物水势由于长期处于临界水平,容易死于碳饥饿[23];在短期的严重干旱胁迫下,植物个体易因为水力失败而脱水死亡.目前关于植物水力传导的研究多针对某一过程进行研究,而针对整株植物水力传导过程及不同器官水力性状间的耦合研究较少.植物木质部水力失败是导致植物生理功能失调的主要原因[11].植物水力失败包括3个阶段:一是气孔调节阶段(水力导度降低12%以上对应的水势,>),植物可通过气孔导度调节来减少水分散失[24-25].二是叶片凋落阶段(水力导度降低值12%~88%对应水势),当水势低于时,气孔完全闭合,空气开始从纹孔结构进入导管;当水势低于(水力导度降低50%对应的水势)时,栓塞在木质部中快速扩散影响水分运输[26-27];此后,植株主要通过叶表皮和茎表皮等散失掉活组织、细胞质和细胞间隙中的水分,并通过叶片脱落来减少水分散失[24,28].三是植物死亡阶段(<),造成植物水力系统不可逆转的损伤,并导致植物死亡[24,29].为了保证植物水力系统的完整性,以下将从根系吸水、木质部导水和叶片蒸腾失水3个方面探究植物对干旱胁迫的响应(图2). ...
... 植物木质部主要包括薄壁细胞、木射线、纤维和导管(管胞)等,薄壁细胞具有储存糖分和水分的作用[24,40],相邻导管间的水分传输通过纹孔结构实现,不相邻导管间的水分传输基于纤维桥假说通过纤维管胞实现[41],纤维具有机械支持作用,导管(管胞)具有水分传输作用.木质部栓塞是植物应对干旱胁迫的被动结果.基于内聚力-张力理论,叶片蒸腾产生的张力大于水分子间的内聚力与水柱和管壁间的附着作用时,木质部导管中水柱断裂并产生气泡,阻碍水分向上输送[7,42-43].气泡的形成可由“气种假说”诠释[43]:①空气泡从纹孔中进入导管;②管道中疏水性区域小气泡膨胀;③木质部中水分流动时,小气泡扩张;④导管壁中小气泡与疏水区脱离.目前关于栓塞的发生主要有两种观点,一种观点指出木质部的空穴和栓塞现象是植物对干旱的响应,在自然界经常发生[43];另一种观点是栓塞只有在极端干旱胁迫下才偶尔发生,且栓塞在水分充足时会被修复[26].木质部栓塞程度随干旱程度增加而增加,常采用栓塞脆弱性曲线来表示[10].木质部抗栓塞性会随植物种类和枝条年龄而异.成年植物个体的木质部有更多的纤维组织来确保机械支撑和水力安全;幼年植物个体的木质部通常会优先形成射线薄壁组织和轴向薄壁组织来进行水分和营养存储[24].Nolf等[10]指出赤桉(Eucalyptus camaldulensis)的当年生枝条相比于老枝更容易受干旱的影响,抗栓塞性较弱;Schuldt等[42]则认为欧洲山毛榉(Fagus sylvatica)生长较慢的老枝比生长较快的新枝的栓塞脆弱性高,可能与老枝空化疲劳有关. ...
... 植物远端叶片水力阻力较大,占全植株水力阻力的30%,是SPAC水力运输的瓶颈和“安全阀”[9,19,26,47].植物叶片蒸腾约占地球表面蒸腾的70%[26],植物叶片通常通过气孔开闭适应环境.叶片气孔变化主要受两个机制调节:一是由叶表皮细胞和保卫细胞之间的水力连接引发的被动调节机制;二是激素如脱落酸的主动调节机制[4].气孔对水力调节和激素调节的敏感度与植物的等水调节和非等水调节有关[4],前者是指植物叶片气孔在干旱胁迫下迅速关闭以保证植物叶片日最小水势保持不变,在水分充足时复水快,易死于碳饥饿;后者是指植物叶片气孔在干旱胁迫下仍保持打开,降低叶片日最小水势,在水分充足时复水慢,易死于水力失败[6,11-12,48].有趣的是,Kannenberg等[6]认为非等水调节植物在水分胁迫期间并没有增加碳同化量.在自然界中没有严格的等水植物和非等水植物,通常介于两者之间[6].当土壤处于中等干旱条件下,植物通常会趋向于非等水调节;而当植物处于严重干旱条件下,植物趋向于等水调节[6,49].植物的等水和非等水程度的量化常采用水力学面积,即测量边界叶水势和1∶1线围成的多边形面积,水力学面积越小越趋向于等水调节[50];也可采用凌晨和午间水势的斜率来表示等水程度,斜率接近0表示植物偏向于等水植物,接近1或大于1表示植物偏向于非等水植物[51].陈嘉嘉[52]研究结果显示,在腾格里沙漠生态系统中,常见的固沙植物油蒿(Artemisia ordosica)、柠条锦鸡儿(Caragana korshinskii)和花棒(Hedysarum scoparium)的凌晨和午间水势斜率分别是0.33、0.92和1.22,分别属于偏等水植物、偏非等水植物和严格非等水植物.植物叶片除了通过气孔开合应对轻微水分胁迫,也可改变叶片解剖形态特征是对干旱胁迫做出的短期快速响应,如叶片气孔在干旱条件下呈小而密的分布(图2),可通过增加气孔闭合能力来减少蒸腾过程中的水分散失[44-45]. ...
... [26],植物叶片通常通过气孔开闭适应环境.叶片气孔变化主要受两个机制调节:一是由叶表皮细胞和保卫细胞之间的水力连接引发的被动调节机制;二是激素如脱落酸的主动调节机制[4].气孔对水力调节和激素调节的敏感度与植物的等水调节和非等水调节有关[4],前者是指植物叶片气孔在干旱胁迫下迅速关闭以保证植物叶片日最小水势保持不变,在水分充足时复水快,易死于碳饥饿;后者是指植物叶片气孔在干旱胁迫下仍保持打开,降低叶片日最小水势,在水分充足时复水慢,易死于水力失败[6,11-12,48].有趣的是,Kannenberg等[6]认为非等水调节植物在水分胁迫期间并没有增加碳同化量.在自然界中没有严格的等水植物和非等水植物,通常介于两者之间[6].当土壤处于中等干旱条件下,植物通常会趋向于非等水调节;而当植物处于严重干旱条件下,植物趋向于等水调节[6,49].植物的等水和非等水程度的量化常采用水力学面积,即测量边界叶水势和1∶1线围成的多边形面积,水力学面积越小越趋向于等水调节[50];也可采用凌晨和午间水势的斜率来表示等水程度,斜率接近0表示植物偏向于等水植物,接近1或大于1表示植物偏向于非等水植物[51].陈嘉嘉[52]研究结果显示,在腾格里沙漠生态系统中,常见的固沙植物油蒿(Artemisia ordosica)、柠条锦鸡儿(Caragana korshinskii)和花棒(Hedysarum scoparium)的凌晨和午间水势斜率分别是0.33、0.92和1.22,分别属于偏等水植物、偏非等水植物和严格非等水植物.植物叶片除了通过气孔开合应对轻微水分胁迫,也可改变叶片解剖形态特征是对干旱胁迫做出的短期快速响应,如叶片气孔在干旱条件下呈小而密的分布(图2),可通过增加气孔闭合能力来减少蒸腾过程中的水分散失[44-45]. ...
Xylem embolism threshold for catastrophic hydraulic failure in angiosperm trees
1
2013
... 不同的干旱强度和时间会影响植物水分运输,在长期的低强度干旱胁迫下,植物水势由于长期处于临界水平,容易死于碳饥饿[23];在短期的严重干旱胁迫下,植物个体易因为水力失败而脱水死亡.目前关于植物水力传导的研究多针对某一过程进行研究,而针对整株植物水力传导过程及不同器官水力性状间的耦合研究较少.植物木质部水力失败是导致植物生理功能失调的主要原因[11].植物水力失败包括3个阶段:一是气孔调节阶段(水力导度降低12%以上对应的水势,>),植物可通过气孔导度调节来减少水分散失[24-25].二是叶片凋落阶段(水力导度降低值12%~88%对应水势),当水势低于时,气孔完全闭合,空气开始从纹孔结构进入导管;当水势低于(水力导度降低50%对应的水势)时,栓塞在木质部中快速扩散影响水分运输[26-27];此后,植株主要通过叶表皮和茎表皮等散失掉活组织、细胞质和细胞间隙中的水分,并通过叶片脱落来减少水分散失[24,28].三是植物死亡阶段(<),造成植物水力系统不可逆转的损伤,并导致植物死亡[24,29].为了保证植物水力系统的完整性,以下将从根系吸水、木质部导水和叶片蒸腾失水3个方面探究植物对干旱胁迫的响应(图2). ...
Linking drought-induced xylem embolism resistance to wood anatomical traits in Neotropical trees
1
2021
... 不同的干旱强度和时间会影响植物水分运输,在长期的低强度干旱胁迫下,植物水势由于长期处于临界水平,容易死于碳饥饿[23];在短期的严重干旱胁迫下,植物个体易因为水力失败而脱水死亡.目前关于植物水力传导的研究多针对某一过程进行研究,而针对整株植物水力传导过程及不同器官水力性状间的耦合研究较少.植物木质部水力失败是导致植物生理功能失调的主要原因[11].植物水力失败包括3个阶段:一是气孔调节阶段(水力导度降低12%以上对应的水势,>),植物可通过气孔导度调节来减少水分散失[24-25].二是叶片凋落阶段(水力导度降低值12%~88%对应水势),当水势低于时,气孔完全闭合,空气开始从纹孔结构进入导管;当水势低于(水力导度降低50%对应的水势)时,栓塞在木质部中快速扩散影响水分运输[26-27];此后,植株主要通过叶表皮和茎表皮等散失掉活组织、细胞质和细胞间隙中的水分,并通过叶片脱落来减少水分散失[24,28].三是植物死亡阶段(<),造成植物水力系统不可逆转的损伤,并导致植物死亡[24,29].为了保证植物水力系统的完整性,以下将从根系吸水、木质部导水和叶片蒸腾失水3个方面探究植物对干旱胁迫的响应(图2). ...
Hydraulic failure and tree mortality: from correlation to causation
1
2022
... 不同的干旱强度和时间会影响植物水分运输,在长期的低强度干旱胁迫下,植物水势由于长期处于临界水平,容易死于碳饥饿[23];在短期的严重干旱胁迫下,植物个体易因为水力失败而脱水死亡.目前关于植物水力传导的研究多针对某一过程进行研究,而针对整株植物水力传导过程及不同器官水力性状间的耦合研究较少.植物木质部水力失败是导致植物生理功能失调的主要原因[11].植物水力失败包括3个阶段:一是气孔调节阶段(水力导度降低12%以上对应的水势,>),植物可通过气孔导度调节来减少水分散失[24-25].二是叶片凋落阶段(水力导度降低值12%~88%对应水势),当水势低于时,气孔完全闭合,空气开始从纹孔结构进入导管;当水势低于(水力导度降低50%对应的水势)时,栓塞在木质部中快速扩散影响水分运输[26-27];此后,植株主要通过叶表皮和茎表皮等散失掉活组织、细胞质和细胞间隙中的水分,并通过叶片脱落来减少水分散失[24,28].三是植物死亡阶段(<),造成植物水力系统不可逆转的损伤,并导致植物死亡[24,29].为了保证植物水力系统的完整性,以下将从根系吸水、木质部导水和叶片蒸腾失水3个方面探究植物对干旱胁迫的响应(图2). ...
Hydrological niche segregation defines forest structure and drought tolerance strategies in a seasonal Amazon forest
2
2019
... 水力再分配是植物根系应对干旱的重要策略,能够促进植物利用土壤水分的生态位分化,改变植被分布格局[30].当土壤水势低于植物叶片水势,且根际土壤水分存在明显的梯度时,土壤剖面存在根系吸收湿润区水分运输至干旱区的现象,影响土壤水分环境,即水力再分配[16,31].水力再分配包括水力提升、水力下传和侧向再分配3个过程,分别是指植物根系对水分向上、向下和侧向的运输[16,32].水力再分配有利于土壤水分运输,缓解土壤干旱胁迫,防止根系木质部水力失败[16],促进养分运输和提高根际微生物活动等[31].近年来,大量研究显示存在根系向土壤释水现象,如夜间根系向土壤中释放水分以弥补白天植物耗水[16,33-34];在同一土壤剖面内生长在湿润土壤的根系吸水增加量(100%)高于整株植物蒸散量的增加量(40%),即补偿性根系吸水[31,35];在干旱条件下根际土壤水分高于周围土壤水分,在湿润情况时则情况相反[33,36-37].值得一提的是,当土壤极端干旱时,过低的土壤水势阻碍了根系释水过程[38],根毛可能会因为张力过大而断裂[39]. ...
... 植物进行形态调整,根系水力再分配,改变木质部和叶片的结构特征的同时,也会进行水力效率和水力安全之间的权衡.水力效率常采用水力导度来衡量,它是水力阻力的倒数,根据欧姆定律,水力导度可用液流速率与压力差比值表示[9],常采用、表征植物木质部的抗栓塞性,采用水力导度降低50%和88%时对应的水势(,)表征植物叶片的抗栓塞性,即植物木质部和叶片的水力安全[42],其值越低表示植物越能忍受较低水势,抗栓塞性越强[30].木质部抗栓塞性和干旱度指数呈线性增加趋势,在干旱胁迫发生时木质部能够忍受更低的水势保证水分运输的正常运行[42].“水力权衡假说”认为植物的不同器官间,存在水力传导效率和安全的高低权衡[42-43].Huo等[44]认为在干旱沙区,柠条锦鸡儿通过降低导水率并增加水力安全性来保证植物能够在干旱胁迫下生存;Zhang等[53]发现生长在干旱沙漠生态系统的植物通常具有较高的水力安全性和较低的导水率,植物的存活与植物水力安全密切相关.而目前也有研究显示植物水力效率和安全不存在权衡关系[42-43];低水力效率和低水力安全的植物可在湿润地区生存[43];也有进一步研究指出自然界中该权衡关系并不是非此即彼的,而是根据环境和植物结构特征而变化[54].水力效率和水力安全之间的权衡关系是由导管结构特征和导管间纹孔结构决定的,水力效率高的木质部通常导管直径较大、纹孔结构较多且木材密度较低,相应的水力安全性较低[55].研究显示在中国北方干旱区,灌木物种在相对干旱区根部导管直径较大并采用效率优先的导水策略;而在相对湿润区,灌木物种导管直径较小并采用安全优先的导水策略[56].尤其是在降水稀少且高温的干旱区,水分为植物生存生长的重要限制因子,植物对水力效率和水力安全的权衡关系研究需要进一步探究. ...
Quantification of root water uptake and redistribution using neutron imaging: a review and future directions
3
2022
... 水力再分配是植物根系应对干旱的重要策略,能够促进植物利用土壤水分的生态位分化,改变植被分布格局[30].当土壤水势低于植物叶片水势,且根际土壤水分存在明显的梯度时,土壤剖面存在根系吸收湿润区水分运输至干旱区的现象,影响土壤水分环境,即水力再分配[16,31].水力再分配包括水力提升、水力下传和侧向再分配3个过程,分别是指植物根系对水分向上、向下和侧向的运输[16,32].水力再分配有利于土壤水分运输,缓解土壤干旱胁迫,防止根系木质部水力失败[16],促进养分运输和提高根际微生物活动等[31].近年来,大量研究显示存在根系向土壤释水现象,如夜间根系向土壤中释放水分以弥补白天植物耗水[16,33-34];在同一土壤剖面内生长在湿润土壤的根系吸水增加量(100%)高于整株植物蒸散量的增加量(40%),即补偿性根系吸水[31,35];在干旱条件下根际土壤水分高于周围土壤水分,在湿润情况时则情况相反[33,36-37].值得一提的是,当土壤极端干旱时,过低的土壤水势阻碍了根系释水过程[38],根毛可能会因为张力过大而断裂[39]. ...
... [31].近年来,大量研究显示存在根系向土壤释水现象,如夜间根系向土壤中释放水分以弥补白天植物耗水[16,33-34];在同一土壤剖面内生长在湿润土壤的根系吸水增加量(100%)高于整株植物蒸散量的增加量(40%),即补偿性根系吸水[31,35];在干旱条件下根际土壤水分高于周围土壤水分,在湿润情况时则情况相反[33,36-37].值得一提的是,当土壤极端干旱时,过低的土壤水势阻碍了根系释水过程[38],根毛可能会因为张力过大而断裂[39]. ...
... [31,35];在干旱条件下根际土壤水分高于周围土壤水分,在湿润情况时则情况相反[33,36-37].值得一提的是,当土壤极端干旱时,过低的土壤水势阻碍了根系释水过程[38],根毛可能会因为张力过大而断裂[39]. ...
植物根系水力再分配测定与模拟方法研究进展与展望
1
2015
... 水力再分配是植物根系应对干旱的重要策略,能够促进植物利用土壤水分的生态位分化,改变植被分布格局[30].当土壤水势低于植物叶片水势,且根际土壤水分存在明显的梯度时,土壤剖面存在根系吸收湿润区水分运输至干旱区的现象,影响土壤水分环境,即水力再分配[16,31].水力再分配包括水力提升、水力下传和侧向再分配3个过程,分别是指植物根系对水分向上、向下和侧向的运输[16,32].水力再分配有利于土壤水分运输,缓解土壤干旱胁迫,防止根系木质部水力失败[16],促进养分运输和提高根际微生物活动等[31].近年来,大量研究显示存在根系向土壤释水现象,如夜间根系向土壤中释放水分以弥补白天植物耗水[16,33-34];在同一土壤剖面内生长在湿润土壤的根系吸水增加量(100%)高于整株植物蒸散量的增加量(40%),即补偿性根系吸水[31,35];在干旱条件下根际土壤水分高于周围土壤水分,在湿润情况时则情况相反[33,36-37].值得一提的是,当土壤极端干旱时,过低的土壤水势阻碍了根系释水过程[38],根毛可能会因为张力过大而断裂[39]. ...
Quantification of root water uptake in soil using X-ray computed tomography and image-based modelling
2
2018
... 水力再分配是植物根系应对干旱的重要策略,能够促进植物利用土壤水分的生态位分化,改变植被分布格局[30].当土壤水势低于植物叶片水势,且根际土壤水分存在明显的梯度时,土壤剖面存在根系吸收湿润区水分运输至干旱区的现象,影响土壤水分环境,即水力再分配[16,31].水力再分配包括水力提升、水力下传和侧向再分配3个过程,分别是指植物根系对水分向上、向下和侧向的运输[16,32].水力再分配有利于土壤水分运输,缓解土壤干旱胁迫,防止根系木质部水力失败[16],促进养分运输和提高根际微生物活动等[31].近年来,大量研究显示存在根系向土壤释水现象,如夜间根系向土壤中释放水分以弥补白天植物耗水[16,33-34];在同一土壤剖面内生长在湿润土壤的根系吸水增加量(100%)高于整株植物蒸散量的增加量(40%),即补偿性根系吸水[31,35];在干旱条件下根际土壤水分高于周围土壤水分,在湿润情况时则情况相反[33,36-37].值得一提的是,当土壤极端干旱时,过低的土壤水势阻碍了根系释水过程[38],根毛可能会因为张力过大而断裂[39]. ...
... [33,36-37].值得一提的是,当土壤极端干旱时,过低的土壤水势阻碍了根系释水过程[38],根毛可能会因为张力过大而断裂[39]. ...
Root exudation and rhizosphere biology
1
2003
... 水力再分配是植物根系应对干旱的重要策略,能够促进植物利用土壤水分的生态位分化,改变植被分布格局[30].当土壤水势低于植物叶片水势,且根际土壤水分存在明显的梯度时,土壤剖面存在根系吸收湿润区水分运输至干旱区的现象,影响土壤水分环境,即水力再分配[16,31].水力再分配包括水力提升、水力下传和侧向再分配3个过程,分别是指植物根系对水分向上、向下和侧向的运输[16,32].水力再分配有利于土壤水分运输,缓解土壤干旱胁迫,防止根系木质部水力失败[16],促进养分运输和提高根际微生物活动等[31].近年来,大量研究显示存在根系向土壤释水现象,如夜间根系向土壤中释放水分以弥补白天植物耗水[16,33-34];在同一土壤剖面内生长在湿润土壤的根系吸水增加量(100%)高于整株植物蒸散量的增加量(40%),即补偿性根系吸水[31,35];在干旱条件下根际土壤水分高于周围土壤水分,在湿润情况时则情况相反[33,36-37].值得一提的是,当土壤极端干旱时,过低的土壤水势阻碍了根系释水过程[38],根毛可能会因为张力过大而断裂[39]. ...
Mapping compensating root water uptake in heterogeneous soil conditions via neutron radiography
1
2015
... 水力再分配是植物根系应对干旱的重要策略,能够促进植物利用土壤水分的生态位分化,改变植被分布格局[30].当土壤水势低于植物叶片水势,且根际土壤水分存在明显的梯度时,土壤剖面存在根系吸收湿润区水分运输至干旱区的现象,影响土壤水分环境,即水力再分配[16,31].水力再分配包括水力提升、水力下传和侧向再分配3个过程,分别是指植物根系对水分向上、向下和侧向的运输[16,32].水力再分配有利于土壤水分运输,缓解土壤干旱胁迫,防止根系木质部水力失败[16],促进养分运输和提高根际微生物活动等[31].近年来,大量研究显示存在根系向土壤释水现象,如夜间根系向土壤中释放水分以弥补白天植物耗水[16,33-34];在同一土壤剖面内生长在湿润土壤的根系吸水增加量(100%)高于整株植物蒸散量的增加量(40%),即补偿性根系吸水[31,35];在干旱条件下根际土壤水分高于周围土壤水分,在湿润情况时则情况相反[33,36-37].值得一提的是,当土壤极端干旱时,过低的土壤水势阻碍了根系释水过程[38],根毛可能会因为张力过大而断裂[39]. ...
A Model of root water uptake coupled with rhizosphere dynamics
1
2012
... 水力再分配是植物根系应对干旱的重要策略,能够促进植物利用土壤水分的生态位分化,改变植被分布格局[30].当土壤水势低于植物叶片水势,且根际土壤水分存在明显的梯度时,土壤剖面存在根系吸收湿润区水分运输至干旱区的现象,影响土壤水分环境,即水力再分配[16,31].水力再分配包括水力提升、水力下传和侧向再分配3个过程,分别是指植物根系对水分向上、向下和侧向的运输[16,32].水力再分配有利于土壤水分运输,缓解土壤干旱胁迫,防止根系木质部水力失败[16],促进养分运输和提高根际微生物活动等[31].近年来,大量研究显示存在根系向土壤释水现象,如夜间根系向土壤中释放水分以弥补白天植物耗水[16,33-34];在同一土壤剖面内生长在湿润土壤的根系吸水增加量(100%)高于整株植物蒸散量的增加量(40%),即补偿性根系吸水[31,35];在干旱条件下根际土壤水分高于周围土壤水分,在湿润情况时则情况相反[33,36-37].值得一提的是,当土壤极端干旱时,过低的土壤水势阻碍了根系释水过程[38],根毛可能会因为张力过大而断裂[39]. ...
Three-dimensional visualization and quantification of water content in the rhizosphere
1
2011
... 水力再分配是植物根系应对干旱的重要策略,能够促进植物利用土壤水分的生态位分化,改变植被分布格局[30].当土壤水势低于植物叶片水势,且根际土壤水分存在明显的梯度时,土壤剖面存在根系吸收湿润区水分运输至干旱区的现象,影响土壤水分环境,即水力再分配[16,31].水力再分配包括水力提升、水力下传和侧向再分配3个过程,分别是指植物根系对水分向上、向下和侧向的运输[16,32].水力再分配有利于土壤水分运输,缓解土壤干旱胁迫,防止根系木质部水力失败[16],促进养分运输和提高根际微生物活动等[31].近年来,大量研究显示存在根系向土壤释水现象,如夜间根系向土壤中释放水分以弥补白天植物耗水[16,33-34];在同一土壤剖面内生长在湿润土壤的根系吸水增加量(100%)高于整株植物蒸散量的增加量(40%),即补偿性根系吸水[31,35];在干旱条件下根际土壤水分高于周围土壤水分,在湿润情况时则情况相反[33,36-37].值得一提的是,当土壤极端干旱时,过低的土壤水势阻碍了根系释水过程[38],根毛可能会因为张力过大而断裂[39]. ...
植物根系水力再分配的研究进展
1
2021
... 水力再分配是植物根系应对干旱的重要策略,能够促进植物利用土壤水分的生态位分化,改变植被分布格局[30].当土壤水势低于植物叶片水势,且根际土壤水分存在明显的梯度时,土壤剖面存在根系吸收湿润区水分运输至干旱区的现象,影响土壤水分环境,即水力再分配[16,31].水力再分配包括水力提升、水力下传和侧向再分配3个过程,分别是指植物根系对水分向上、向下和侧向的运输[16,32].水力再分配有利于土壤水分运输,缓解土壤干旱胁迫,防止根系木质部水力失败[16],促进养分运输和提高根际微生物活动等[31].近年来,大量研究显示存在根系向土壤释水现象,如夜间根系向土壤中释放水分以弥补白天植物耗水[16,33-34];在同一土壤剖面内生长在湿润土壤的根系吸水增加量(100%)高于整株植物蒸散量的增加量(40%),即补偿性根系吸水[31,35];在干旱条件下根际土壤水分高于周围土壤水分,在湿润情况时则情况相反[33,36-37].值得一提的是,当土壤极端干旱时,过低的土壤水势阻碍了根系释水过程[38],根毛可能会因为张力过大而断裂[39]. ...
Stomatal closure prevents the drop in soil water potential around roots
1
2020
... 水力再分配是植物根系应对干旱的重要策略,能够促进植物利用土壤水分的生态位分化,改变植被分布格局[30].当土壤水势低于植物叶片水势,且根际土壤水分存在明显的梯度时,土壤剖面存在根系吸收湿润区水分运输至干旱区的现象,影响土壤水分环境,即水力再分配[16,31].水力再分配包括水力提升、水力下传和侧向再分配3个过程,分别是指植物根系对水分向上、向下和侧向的运输[16,32].水力再分配有利于土壤水分运输,缓解土壤干旱胁迫,防止根系木质部水力失败[16],促进养分运输和提高根际微生物活动等[31].近年来,大量研究显示存在根系向土壤释水现象,如夜间根系向土壤中释放水分以弥补白天植物耗水[16,33-34];在同一土壤剖面内生长在湿润土壤的根系吸水增加量(100%)高于整株植物蒸散量的增加量(40%),即补偿性根系吸水[31,35];在干旱条件下根际土壤水分高于周围土壤水分,在湿润情况时则情况相反[33,36-37].值得一提的是,当土壤极端干旱时,过低的土壤水势阻碍了根系释水过程[38],根毛可能会因为张力过大而断裂[39]. ...
植物木质部管道结构和水力特性的关系
5
2017
... 植物木质部主要包括薄壁细胞、木射线、纤维和导管(管胞)等,薄壁细胞具有储存糖分和水分的作用[24,40],相邻导管间的水分传输通过纹孔结构实现,不相邻导管间的水分传输基于纤维桥假说通过纤维管胞实现[41],纤维具有机械支持作用,导管(管胞)具有水分传输作用.木质部栓塞是植物应对干旱胁迫的被动结果.基于内聚力-张力理论,叶片蒸腾产生的张力大于水分子间的内聚力与水柱和管壁间的附着作用时,木质部导管中水柱断裂并产生气泡,阻碍水分向上输送[7,42-43].气泡的形成可由“气种假说”诠释[43]:①空气泡从纹孔中进入导管;②管道中疏水性区域小气泡膨胀;③木质部中水分流动时,小气泡扩张;④导管壁中小气泡与疏水区脱离.目前关于栓塞的发生主要有两种观点,一种观点指出木质部的空穴和栓塞现象是植物对干旱的响应,在自然界经常发生[43];另一种观点是栓塞只有在极端干旱胁迫下才偶尔发生,且栓塞在水分充足时会被修复[26].木质部栓塞程度随干旱程度增加而增加,常采用栓塞脆弱性曲线来表示[10].木质部抗栓塞性会随植物种类和枝条年龄而异.成年植物个体的木质部有更多的纤维组织来确保机械支撑和水力安全;幼年植物个体的木质部通常会优先形成射线薄壁组织和轴向薄壁组织来进行水分和营养存储[24].Nolf等[10]指出赤桉(Eucalyptus camaldulensis)的当年生枝条相比于老枝更容易受干旱的影响,抗栓塞性较弱;Schuldt等[42]则认为欧洲山毛榉(Fagus sylvatica)生长较慢的老枝比生长较快的新枝的栓塞脆弱性高,可能与老枝空化疲劳有关. ...
... 植物木质部可通过改变导管结构特征来应对干旱胁迫.木质部常通过减小导管直径,增加导管密度、纹孔膜厚度和木材密度,进而降低木质部导水率并提高水力安全性,且已有大量研究证实(图2)[13,42,44].He等[13]指出相比于湿润环境,在干旱环境下水力失败风险较大且维护成本较高,植物通常会降低导水率以适应环境.Schuldt等[42]发现欧洲山毛榉的木质部导管直径降低7%且纹孔膜厚度增加15%以应对干旱胁迫.导管结构特征影响木质部导水率,Hagen-Poiseuille水力传导方程显示导管导水率与导管直径的4次方成正比,因此导管直径的微小增加会带来导水率的显著提高[41].有研究采用X射线显微断层成像法观测干燥不同时间的赤桉的木质部横截面的导管栓塞情况,结果显示Hagen-Poiseuille水力传导方程模拟的理论导水率高于实际最大导水率[10],原因一是与没有考虑导管长度和导管间的纹孔结构有关[41],导管长度越短、纹孔数量越少和纹孔面积越小,则导管导水率越低[41],纹孔阻力约占整个植株总水力传导阻力的14%~84%[40];二是与导管腔的不规则形状和锥度有关[45];三是与木质部汁液有关,研究显示KCl浓度的增加可导致木质部导水率增加10%[45].枝条形态学基部导管直径通常高于形态学末端[8,40],这一形态结构有利于减小由于植物高度增长带来的水力阻力[8].通常情况下导管直径表现为主干直径>侧枝>枝条交叉点[40].植物体的枝条交叉点、根茎过渡区和节点部位的导管直径较小、导水率较小、胡伯尔值较大和抗栓塞能力较强,这些部位常被称为“水流限制区”和“安全区”[46].胡伯尔值越大,表示维持单位叶面积水分需要的木质部越多,可用于比较不同树种间的水力特征[19,40]. ...
... ,40],这一形态结构有利于减小由于植物高度增长带来的水力阻力[8].通常情况下导管直径表现为主干直径>侧枝>枝条交叉点[40].植物体的枝条交叉点、根茎过渡区和节点部位的导管直径较小、导水率较小、胡伯尔值较大和抗栓塞能力较强,这些部位常被称为“水流限制区”和“安全区”[46].胡伯尔值越大,表示维持单位叶面积水分需要的木质部越多,可用于比较不同树种间的水力特征[19,40]. ...
... [40].植物体的枝条交叉点、根茎过渡区和节点部位的导管直径较小、导水率较小、胡伯尔值较大和抗栓塞能力较强,这些部位常被称为“水流限制区”和“安全区”[46].胡伯尔值越大,表示维持单位叶面积水分需要的木质部越多,可用于比较不同树种间的水力特征[19,40]. ...
... ,40]. ...
红树植物木质部水力结构与功能研究
4
2021
... 植物木质部主要包括薄壁细胞、木射线、纤维和导管(管胞)等,薄壁细胞具有储存糖分和水分的作用[24,40],相邻导管间的水分传输通过纹孔结构实现,不相邻导管间的水分传输基于纤维桥假说通过纤维管胞实现[41],纤维具有机械支持作用,导管(管胞)具有水分传输作用.木质部栓塞是植物应对干旱胁迫的被动结果.基于内聚力-张力理论,叶片蒸腾产生的张力大于水分子间的内聚力与水柱和管壁间的附着作用时,木质部导管中水柱断裂并产生气泡,阻碍水分向上输送[7,42-43].气泡的形成可由“气种假说”诠释[43]:①空气泡从纹孔中进入导管;②管道中疏水性区域小气泡膨胀;③木质部中水分流动时,小气泡扩张;④导管壁中小气泡与疏水区脱离.目前关于栓塞的发生主要有两种观点,一种观点指出木质部的空穴和栓塞现象是植物对干旱的响应,在自然界经常发生[43];另一种观点是栓塞只有在极端干旱胁迫下才偶尔发生,且栓塞在水分充足时会被修复[26].木质部栓塞程度随干旱程度增加而增加,常采用栓塞脆弱性曲线来表示[10].木质部抗栓塞性会随植物种类和枝条年龄而异.成年植物个体的木质部有更多的纤维组织来确保机械支撑和水力安全;幼年植物个体的木质部通常会优先形成射线薄壁组织和轴向薄壁组织来进行水分和营养存储[24].Nolf等[10]指出赤桉(Eucalyptus camaldulensis)的当年生枝条相比于老枝更容易受干旱的影响,抗栓塞性较弱;Schuldt等[42]则认为欧洲山毛榉(Fagus sylvatica)生长较慢的老枝比生长较快的新枝的栓塞脆弱性高,可能与老枝空化疲劳有关. ...
... 植物木质部可通过改变导管结构特征来应对干旱胁迫.木质部常通过减小导管直径,增加导管密度、纹孔膜厚度和木材密度,进而降低木质部导水率并提高水力安全性,且已有大量研究证实(图2)[13,42,44].He等[13]指出相比于湿润环境,在干旱环境下水力失败风险较大且维护成本较高,植物通常会降低导水率以适应环境.Schuldt等[42]发现欧洲山毛榉的木质部导管直径降低7%且纹孔膜厚度增加15%以应对干旱胁迫.导管结构特征影响木质部导水率,Hagen-Poiseuille水力传导方程显示导管导水率与导管直径的4次方成正比,因此导管直径的微小增加会带来导水率的显著提高[41].有研究采用X射线显微断层成像法观测干燥不同时间的赤桉的木质部横截面的导管栓塞情况,结果显示Hagen-Poiseuille水力传导方程模拟的理论导水率高于实际最大导水率[10],原因一是与没有考虑导管长度和导管间的纹孔结构有关[41],导管长度越短、纹孔数量越少和纹孔面积越小,则导管导水率越低[41],纹孔阻力约占整个植株总水力传导阻力的14%~84%[40];二是与导管腔的不规则形状和锥度有关[45];三是与木质部汁液有关,研究显示KCl浓度的增加可导致木质部导水率增加10%[45].枝条形态学基部导管直径通常高于形态学末端[8,40],这一形态结构有利于减小由于植物高度增长带来的水力阻力[8].通常情况下导管直径表现为主干直径>侧枝>枝条交叉点[40].植物体的枝条交叉点、根茎过渡区和节点部位的导管直径较小、导水率较小、胡伯尔值较大和抗栓塞能力较强,这些部位常被称为“水流限制区”和“安全区”[46].胡伯尔值越大,表示维持单位叶面积水分需要的木质部越多,可用于比较不同树种间的水力特征[19,40]. ...
... [41],导管长度越短、纹孔数量越少和纹孔面积越小,则导管导水率越低[41],纹孔阻力约占整个植株总水力传导阻力的14%~84%[40];二是与导管腔的不规则形状和锥度有关[45];三是与木质部汁液有关,研究显示KCl浓度的增加可导致木质部导水率增加10%[45].枝条形态学基部导管直径通常高于形态学末端[8,40],这一形态结构有利于减小由于植物高度增长带来的水力阻力[8].通常情况下导管直径表现为主干直径>侧枝>枝条交叉点[40].植物体的枝条交叉点、根茎过渡区和节点部位的导管直径较小、导水率较小、胡伯尔值较大和抗栓塞能力较强,这些部位常被称为“水流限制区”和“安全区”[46].胡伯尔值越大,表示维持单位叶面积水分需要的木质部越多,可用于比较不同树种间的水力特征[19,40]. ...
... [41],纹孔阻力约占整个植株总水力传导阻力的14%~84%[40];二是与导管腔的不规则形状和锥度有关[45];三是与木质部汁液有关,研究显示KCl浓度的增加可导致木质部导水率增加10%[45].枝条形态学基部导管直径通常高于形态学末端[8,40],这一形态结构有利于减小由于植物高度增长带来的水力阻力[8].通常情况下导管直径表现为主干直径>侧枝>枝条交叉点[40].植物体的枝条交叉点、根茎过渡区和节点部位的导管直径较小、导水率较小、胡伯尔值较大和抗栓塞能力较强,这些部位常被称为“水流限制区”和“安全区”[46].胡伯尔值越大,表示维持单位叶面积水分需要的木质部越多,可用于比较不同树种间的水力特征[19,40]. ...
How adaptable is the hydraulic system of European beech in the face of climate change-related precipitation reduction
8
2016
... 植物木质部主要包括薄壁细胞、木射线、纤维和导管(管胞)等,薄壁细胞具有储存糖分和水分的作用[24,40],相邻导管间的水分传输通过纹孔结构实现,不相邻导管间的水分传输基于纤维桥假说通过纤维管胞实现[41],纤维具有机械支持作用,导管(管胞)具有水分传输作用.木质部栓塞是植物应对干旱胁迫的被动结果.基于内聚力-张力理论,叶片蒸腾产生的张力大于水分子间的内聚力与水柱和管壁间的附着作用时,木质部导管中水柱断裂并产生气泡,阻碍水分向上输送[7,42-43].气泡的形成可由“气种假说”诠释[43]:①空气泡从纹孔中进入导管;②管道中疏水性区域小气泡膨胀;③木质部中水分流动时,小气泡扩张;④导管壁中小气泡与疏水区脱离.目前关于栓塞的发生主要有两种观点,一种观点指出木质部的空穴和栓塞现象是植物对干旱的响应,在自然界经常发生[43];另一种观点是栓塞只有在极端干旱胁迫下才偶尔发生,且栓塞在水分充足时会被修复[26].木质部栓塞程度随干旱程度增加而增加,常采用栓塞脆弱性曲线来表示[10].木质部抗栓塞性会随植物种类和枝条年龄而异.成年植物个体的木质部有更多的纤维组织来确保机械支撑和水力安全;幼年植物个体的木质部通常会优先形成射线薄壁组织和轴向薄壁组织来进行水分和营养存储[24].Nolf等[10]指出赤桉(Eucalyptus camaldulensis)的当年生枝条相比于老枝更容易受干旱的影响,抗栓塞性较弱;Schuldt等[42]则认为欧洲山毛榉(Fagus sylvatica)生长较慢的老枝比生长较快的新枝的栓塞脆弱性高,可能与老枝空化疲劳有关. ...
... [42]则认为欧洲山毛榉(Fagus sylvatica)生长较慢的老枝比生长较快的新枝的栓塞脆弱性高,可能与老枝空化疲劳有关. ...
... 植物木质部可通过改变导管结构特征来应对干旱胁迫.木质部常通过减小导管直径,增加导管密度、纹孔膜厚度和木材密度,进而降低木质部导水率并提高水力安全性,且已有大量研究证实(图2)[13,42,44].He等[13]指出相比于湿润环境,在干旱环境下水力失败风险较大且维护成本较高,植物通常会降低导水率以适应环境.Schuldt等[42]发现欧洲山毛榉的木质部导管直径降低7%且纹孔膜厚度增加15%以应对干旱胁迫.导管结构特征影响木质部导水率,Hagen-Poiseuille水力传导方程显示导管导水率与导管直径的4次方成正比,因此导管直径的微小增加会带来导水率的显著提高[41].有研究采用X射线显微断层成像法观测干燥不同时间的赤桉的木质部横截面的导管栓塞情况,结果显示Hagen-Poiseuille水力传导方程模拟的理论导水率高于实际最大导水率[10],原因一是与没有考虑导管长度和导管间的纹孔结构有关[41],导管长度越短、纹孔数量越少和纹孔面积越小,则导管导水率越低[41],纹孔阻力约占整个植株总水力传导阻力的14%~84%[40];二是与导管腔的不规则形状和锥度有关[45];三是与木质部汁液有关,研究显示KCl浓度的增加可导致木质部导水率增加10%[45].枝条形态学基部导管直径通常高于形态学末端[8,40],这一形态结构有利于减小由于植物高度增长带来的水力阻力[8].通常情况下导管直径表现为主干直径>侧枝>枝条交叉点[40].植物体的枝条交叉点、根茎过渡区和节点部位的导管直径较小、导水率较小、胡伯尔值较大和抗栓塞能力较强,这些部位常被称为“水流限制区”和“安全区”[46].胡伯尔值越大,表示维持单位叶面积水分需要的木质部越多,可用于比较不同树种间的水力特征[19,40]. ...
... [42]发现欧洲山毛榉的木质部导管直径降低7%且纹孔膜厚度增加15%以应对干旱胁迫.导管结构特征影响木质部导水率,Hagen-Poiseuille水力传导方程显示导管导水率与导管直径的4次方成正比,因此导管直径的微小增加会带来导水率的显著提高[41].有研究采用X射线显微断层成像法观测干燥不同时间的赤桉的木质部横截面的导管栓塞情况,结果显示Hagen-Poiseuille水力传导方程模拟的理论导水率高于实际最大导水率[10],原因一是与没有考虑导管长度和导管间的纹孔结构有关[41],导管长度越短、纹孔数量越少和纹孔面积越小,则导管导水率越低[41],纹孔阻力约占整个植株总水力传导阻力的14%~84%[40];二是与导管腔的不规则形状和锥度有关[45];三是与木质部汁液有关,研究显示KCl浓度的增加可导致木质部导水率增加10%[45].枝条形态学基部导管直径通常高于形态学末端[8,40],这一形态结构有利于减小由于植物高度增长带来的水力阻力[8].通常情况下导管直径表现为主干直径>侧枝>枝条交叉点[40].植物体的枝条交叉点、根茎过渡区和节点部位的导管直径较小、导水率较小、胡伯尔值较大和抗栓塞能力较强,这些部位常被称为“水流限制区”和“安全区”[46].胡伯尔值越大,表示维持单位叶面积水分需要的木质部越多,可用于比较不同树种间的水力特征[19,40]. ...
... 植物进行形态调整,根系水力再分配,改变木质部和叶片的结构特征的同时,也会进行水力效率和水力安全之间的权衡.水力效率常采用水力导度来衡量,它是水力阻力的倒数,根据欧姆定律,水力导度可用液流速率与压力差比值表示[9],常采用、表征植物木质部的抗栓塞性,采用水力导度降低50%和88%时对应的水势(,)表征植物叶片的抗栓塞性,即植物木质部和叶片的水力安全[42],其值越低表示植物越能忍受较低水势,抗栓塞性越强[30].木质部抗栓塞性和干旱度指数呈线性增加趋势,在干旱胁迫发生时木质部能够忍受更低的水势保证水分运输的正常运行[42].“水力权衡假说”认为植物的不同器官间,存在水力传导效率和安全的高低权衡[42-43].Huo等[44]认为在干旱沙区,柠条锦鸡儿通过降低导水率并增加水力安全性来保证植物能够在干旱胁迫下生存;Zhang等[53]发现生长在干旱沙漠生态系统的植物通常具有较高的水力安全性和较低的导水率,植物的存活与植物水力安全密切相关.而目前也有研究显示植物水力效率和安全不存在权衡关系[42-43];低水力效率和低水力安全的植物可在湿润地区生存[43];也有进一步研究指出自然界中该权衡关系并不是非此即彼的,而是根据环境和植物结构特征而变化[54].水力效率和水力安全之间的权衡关系是由导管结构特征和导管间纹孔结构决定的,水力效率高的木质部通常导管直径较大、纹孔结构较多且木材密度较低,相应的水力安全性较低[55].研究显示在中国北方干旱区,灌木物种在相对干旱区根部导管直径较大并采用效率优先的导水策略;而在相对湿润区,灌木物种导管直径较小并采用安全优先的导水策略[56].尤其是在降水稀少且高温的干旱区,水分为植物生存生长的重要限制因子,植物对水力效率和水力安全的权衡关系研究需要进一步探究. ...
... [42].“水力权衡假说”认为植物的不同器官间,存在水力传导效率和安全的高低权衡[42-43].Huo等[44]认为在干旱沙区,柠条锦鸡儿通过降低导水率并增加水力安全性来保证植物能够在干旱胁迫下生存;Zhang等[53]发现生长在干旱沙漠生态系统的植物通常具有较高的水力安全性和较低的导水率,植物的存活与植物水力安全密切相关.而目前也有研究显示植物水力效率和安全不存在权衡关系[42-43];低水力效率和低水力安全的植物可在湿润地区生存[43];也有进一步研究指出自然界中该权衡关系并不是非此即彼的,而是根据环境和植物结构特征而变化[54].水力效率和水力安全之间的权衡关系是由导管结构特征和导管间纹孔结构决定的,水力效率高的木质部通常导管直径较大、纹孔结构较多且木材密度较低,相应的水力安全性较低[55].研究显示在中国北方干旱区,灌木物种在相对干旱区根部导管直径较大并采用效率优先的导水策略;而在相对湿润区,灌木物种导管直径较小并采用安全优先的导水策略[56].尤其是在降水稀少且高温的干旱区,水分为植物生存生长的重要限制因子,植物对水力效率和水力安全的权衡关系研究需要进一步探究. ...
... [42-43].Huo等[44]认为在干旱沙区,柠条锦鸡儿通过降低导水率并增加水力安全性来保证植物能够在干旱胁迫下生存;Zhang等[53]发现生长在干旱沙漠生态系统的植物通常具有较高的水力安全性和较低的导水率,植物的存活与植物水力安全密切相关.而目前也有研究显示植物水力效率和安全不存在权衡关系[42-43];低水力效率和低水力安全的植物可在湿润地区生存[43];也有进一步研究指出自然界中该权衡关系并不是非此即彼的,而是根据环境和植物结构特征而变化[54].水力效率和水力安全之间的权衡关系是由导管结构特征和导管间纹孔结构决定的,水力效率高的木质部通常导管直径较大、纹孔结构较多且木材密度较低,相应的水力安全性较低[55].研究显示在中国北方干旱区,灌木物种在相对干旱区根部导管直径较大并采用效率优先的导水策略;而在相对湿润区,灌木物种导管直径较小并采用安全优先的导水策略[56].尤其是在降水稀少且高温的干旱区,水分为植物生存生长的重要限制因子,植物对水力效率和水力安全的权衡关系研究需要进一步探究. ...
... [42-43];低水力效率和低水力安全的植物可在湿润地区生存[43];也有进一步研究指出自然界中该权衡关系并不是非此即彼的,而是根据环境和植物结构特征而变化[54].水力效率和水力安全之间的权衡关系是由导管结构特征和导管间纹孔结构决定的,水力效率高的木质部通常导管直径较大、纹孔结构较多且木材密度较低,相应的水力安全性较低[55].研究显示在中国北方干旱区,灌木物种在相对干旱区根部导管直径较大并采用效率优先的导水策略;而在相对湿润区,灌木物种导管直径较小并采用安全优先的导水策略[56].尤其是在降水稀少且高温的干旱区,水分为植物生存生长的重要限制因子,植物对水力效率和水力安全的权衡关系研究需要进一步探究. ...
杉木幼苗水力性状特征及其对干旱的响应机制
6
2023
... 植物木质部主要包括薄壁细胞、木射线、纤维和导管(管胞)等,薄壁细胞具有储存糖分和水分的作用[24,40],相邻导管间的水分传输通过纹孔结构实现,不相邻导管间的水分传输基于纤维桥假说通过纤维管胞实现[41],纤维具有机械支持作用,导管(管胞)具有水分传输作用.木质部栓塞是植物应对干旱胁迫的被动结果.基于内聚力-张力理论,叶片蒸腾产生的张力大于水分子间的内聚力与水柱和管壁间的附着作用时,木质部导管中水柱断裂并产生气泡,阻碍水分向上输送[7,42-43].气泡的形成可由“气种假说”诠释[43]:①空气泡从纹孔中进入导管;②管道中疏水性区域小气泡膨胀;③木质部中水分流动时,小气泡扩张;④导管壁中小气泡与疏水区脱离.目前关于栓塞的发生主要有两种观点,一种观点指出木质部的空穴和栓塞现象是植物对干旱的响应,在自然界经常发生[43];另一种观点是栓塞只有在极端干旱胁迫下才偶尔发生,且栓塞在水分充足时会被修复[26].木质部栓塞程度随干旱程度增加而增加,常采用栓塞脆弱性曲线来表示[10].木质部抗栓塞性会随植物种类和枝条年龄而异.成年植物个体的木质部有更多的纤维组织来确保机械支撑和水力安全;幼年植物个体的木质部通常会优先形成射线薄壁组织和轴向薄壁组织来进行水分和营养存储[24].Nolf等[10]指出赤桉(Eucalyptus camaldulensis)的当年生枝条相比于老枝更容易受干旱的影响,抗栓塞性较弱;Schuldt等[42]则认为欧洲山毛榉(Fagus sylvatica)生长较慢的老枝比生长较快的新枝的栓塞脆弱性高,可能与老枝空化疲劳有关. ...
... [43]:①空气泡从纹孔中进入导管;②管道中疏水性区域小气泡膨胀;③木质部中水分流动时,小气泡扩张;④导管壁中小气泡与疏水区脱离.目前关于栓塞的发生主要有两种观点,一种观点指出木质部的空穴和栓塞现象是植物对干旱的响应,在自然界经常发生[43];另一种观点是栓塞只有在极端干旱胁迫下才偶尔发生,且栓塞在水分充足时会被修复[26].木质部栓塞程度随干旱程度增加而增加,常采用栓塞脆弱性曲线来表示[10].木质部抗栓塞性会随植物种类和枝条年龄而异.成年植物个体的木质部有更多的纤维组织来确保机械支撑和水力安全;幼年植物个体的木质部通常会优先形成射线薄壁组织和轴向薄壁组织来进行水分和营养存储[24].Nolf等[10]指出赤桉(Eucalyptus camaldulensis)的当年生枝条相比于老枝更容易受干旱的影响,抗栓塞性较弱;Schuldt等[42]则认为欧洲山毛榉(Fagus sylvatica)生长较慢的老枝比生长较快的新枝的栓塞脆弱性高,可能与老枝空化疲劳有关. ...
... [43];另一种观点是栓塞只有在极端干旱胁迫下才偶尔发生,且栓塞在水分充足时会被修复[26].木质部栓塞程度随干旱程度增加而增加,常采用栓塞脆弱性曲线来表示[10].木质部抗栓塞性会随植物种类和枝条年龄而异.成年植物个体的木质部有更多的纤维组织来确保机械支撑和水力安全;幼年植物个体的木质部通常会优先形成射线薄壁组织和轴向薄壁组织来进行水分和营养存储[24].Nolf等[10]指出赤桉(Eucalyptus camaldulensis)的当年生枝条相比于老枝更容易受干旱的影响,抗栓塞性较弱;Schuldt等[42]则认为欧洲山毛榉(Fagus sylvatica)生长较慢的老枝比生长较快的新枝的栓塞脆弱性高,可能与老枝空化疲劳有关. ...
... 植物进行形态调整,根系水力再分配,改变木质部和叶片的结构特征的同时,也会进行水力效率和水力安全之间的权衡.水力效率常采用水力导度来衡量,它是水力阻力的倒数,根据欧姆定律,水力导度可用液流速率与压力差比值表示[9],常采用、表征植物木质部的抗栓塞性,采用水力导度降低50%和88%时对应的水势(,)表征植物叶片的抗栓塞性,即植物木质部和叶片的水力安全[42],其值越低表示植物越能忍受较低水势,抗栓塞性越强[30].木质部抗栓塞性和干旱度指数呈线性增加趋势,在干旱胁迫发生时木质部能够忍受更低的水势保证水分运输的正常运行[42].“水力权衡假说”认为植物的不同器官间,存在水力传导效率和安全的高低权衡[42-43].Huo等[44]认为在干旱沙区,柠条锦鸡儿通过降低导水率并增加水力安全性来保证植物能够在干旱胁迫下生存;Zhang等[53]发现生长在干旱沙漠生态系统的植物通常具有较高的水力安全性和较低的导水率,植物的存活与植物水力安全密切相关.而目前也有研究显示植物水力效率和安全不存在权衡关系[42-43];低水力效率和低水力安全的植物可在湿润地区生存[43];也有进一步研究指出自然界中该权衡关系并不是非此即彼的,而是根据环境和植物结构特征而变化[54].水力效率和水力安全之间的权衡关系是由导管结构特征和导管间纹孔结构决定的,水力效率高的木质部通常导管直径较大、纹孔结构较多且木材密度较低,相应的水力安全性较低[55].研究显示在中国北方干旱区,灌木物种在相对干旱区根部导管直径较大并采用效率优先的导水策略;而在相对湿润区,灌木物种导管直径较小并采用安全优先的导水策略[56].尤其是在降水稀少且高温的干旱区,水分为植物生存生长的重要限制因子,植物对水力效率和水力安全的权衡关系研究需要进一步探究. ...
... -43];低水力效率和低水力安全的植物可在湿润地区生存[43];也有进一步研究指出自然界中该权衡关系并不是非此即彼的,而是根据环境和植物结构特征而变化[54].水力效率和水力安全之间的权衡关系是由导管结构特征和导管间纹孔结构决定的,水力效率高的木质部通常导管直径较大、纹孔结构较多且木材密度较低,相应的水力安全性较低[55].研究显示在中国北方干旱区,灌木物种在相对干旱区根部导管直径较大并采用效率优先的导水策略;而在相对湿润区,灌木物种导管直径较小并采用安全优先的导水策略[56].尤其是在降水稀少且高温的干旱区,水分为植物生存生长的重要限制因子,植物对水力效率和水力安全的权衡关系研究需要进一步探究. ...
... [43];也有进一步研究指出自然界中该权衡关系并不是非此即彼的,而是根据环境和植物结构特征而变化[54].水力效率和水力安全之间的权衡关系是由导管结构特征和导管间纹孔结构决定的,水力效率高的木质部通常导管直径较大、纹孔结构较多且木材密度较低,相应的水力安全性较低[55].研究显示在中国北方干旱区,灌木物种在相对干旱区根部导管直径较大并采用效率优先的导水策略;而在相对湿润区,灌木物种导管直径较小并采用安全优先的导水策略[56].尤其是在降水稀少且高温的干旱区,水分为植物生存生长的重要限制因子,植物对水力效率和水力安全的权衡关系研究需要进一步探究. ...
Hydraulic mechanism of limiting growth and maintaining survival of desert shrubs in arid habitats
3
2024
... 植物木质部可通过改变导管结构特征来应对干旱胁迫.木质部常通过减小导管直径,增加导管密度、纹孔膜厚度和木材密度,进而降低木质部导水率并提高水力安全性,且已有大量研究证实(图2)[13,42,44].He等[13]指出相比于湿润环境,在干旱环境下水力失败风险较大且维护成本较高,植物通常会降低导水率以适应环境.Schuldt等[42]发现欧洲山毛榉的木质部导管直径降低7%且纹孔膜厚度增加15%以应对干旱胁迫.导管结构特征影响木质部导水率,Hagen-Poiseuille水力传导方程显示导管导水率与导管直径的4次方成正比,因此导管直径的微小增加会带来导水率的显著提高[41].有研究采用X射线显微断层成像法观测干燥不同时间的赤桉的木质部横截面的导管栓塞情况,结果显示Hagen-Poiseuille水力传导方程模拟的理论导水率高于实际最大导水率[10],原因一是与没有考虑导管长度和导管间的纹孔结构有关[41],导管长度越短、纹孔数量越少和纹孔面积越小,则导管导水率越低[41],纹孔阻力约占整个植株总水力传导阻力的14%~84%[40];二是与导管腔的不规则形状和锥度有关[45];三是与木质部汁液有关,研究显示KCl浓度的增加可导致木质部导水率增加10%[45].枝条形态学基部导管直径通常高于形态学末端[8,40],这一形态结构有利于减小由于植物高度增长带来的水力阻力[8].通常情况下导管直径表现为主干直径>侧枝>枝条交叉点[40].植物体的枝条交叉点、根茎过渡区和节点部位的导管直径较小、导水率较小、胡伯尔值较大和抗栓塞能力较强,这些部位常被称为“水流限制区”和“安全区”[46].胡伯尔值越大,表示维持单位叶面积水分需要的木质部越多,可用于比较不同树种间的水力特征[19,40]. ...
... 植物远端叶片水力阻力较大,占全植株水力阻力的30%,是SPAC水力运输的瓶颈和“安全阀”[9,19,26,47].植物叶片蒸腾约占地球表面蒸腾的70%[26],植物叶片通常通过气孔开闭适应环境.叶片气孔变化主要受两个机制调节:一是由叶表皮细胞和保卫细胞之间的水力连接引发的被动调节机制;二是激素如脱落酸的主动调节机制[4].气孔对水力调节和激素调节的敏感度与植物的等水调节和非等水调节有关[4],前者是指植物叶片气孔在干旱胁迫下迅速关闭以保证植物叶片日最小水势保持不变,在水分充足时复水快,易死于碳饥饿;后者是指植物叶片气孔在干旱胁迫下仍保持打开,降低叶片日最小水势,在水分充足时复水慢,易死于水力失败[6,11-12,48].有趣的是,Kannenberg等[6]认为非等水调节植物在水分胁迫期间并没有增加碳同化量.在自然界中没有严格的等水植物和非等水植物,通常介于两者之间[6].当土壤处于中等干旱条件下,植物通常会趋向于非等水调节;而当植物处于严重干旱条件下,植物趋向于等水调节[6,49].植物的等水和非等水程度的量化常采用水力学面积,即测量边界叶水势和1∶1线围成的多边形面积,水力学面积越小越趋向于等水调节[50];也可采用凌晨和午间水势的斜率来表示等水程度,斜率接近0表示植物偏向于等水植物,接近1或大于1表示植物偏向于非等水植物[51].陈嘉嘉[52]研究结果显示,在腾格里沙漠生态系统中,常见的固沙植物油蒿(Artemisia ordosica)、柠条锦鸡儿(Caragana korshinskii)和花棒(Hedysarum scoparium)的凌晨和午间水势斜率分别是0.33、0.92和1.22,分别属于偏等水植物、偏非等水植物和严格非等水植物.植物叶片除了通过气孔开合应对轻微水分胁迫,也可改变叶片解剖形态特征是对干旱胁迫做出的短期快速响应,如叶片气孔在干旱条件下呈小而密的分布(图2),可通过增加气孔闭合能力来减少蒸腾过程中的水分散失[44-45]. ...
... 植物进行形态调整,根系水力再分配,改变木质部和叶片的结构特征的同时,也会进行水力效率和水力安全之间的权衡.水力效率常采用水力导度来衡量,它是水力阻力的倒数,根据欧姆定律,水力导度可用液流速率与压力差比值表示[9],常采用、表征植物木质部的抗栓塞性,采用水力导度降低50%和88%时对应的水势(,)表征植物叶片的抗栓塞性,即植物木质部和叶片的水力安全[42],其值越低表示植物越能忍受较低水势,抗栓塞性越强[30].木质部抗栓塞性和干旱度指数呈线性增加趋势,在干旱胁迫发生时木质部能够忍受更低的水势保证水分运输的正常运行[42].“水力权衡假说”认为植物的不同器官间,存在水力传导效率和安全的高低权衡[42-43].Huo等[44]认为在干旱沙区,柠条锦鸡儿通过降低导水率并增加水力安全性来保证植物能够在干旱胁迫下生存;Zhang等[53]发现生长在干旱沙漠生态系统的植物通常具有较高的水力安全性和较低的导水率,植物的存活与植物水力安全密切相关.而目前也有研究显示植物水力效率和安全不存在权衡关系[42-43];低水力效率和低水力安全的植物可在湿润地区生存[43];也有进一步研究指出自然界中该权衡关系并不是非此即彼的,而是根据环境和植物结构特征而变化[54].水力效率和水力安全之间的权衡关系是由导管结构特征和导管间纹孔结构决定的,水力效率高的木质部通常导管直径较大、纹孔结构较多且木材密度较低,相应的水力安全性较低[55].研究显示在中国北方干旱区,灌木物种在相对干旱区根部导管直径较大并采用效率优先的导水策略;而在相对湿润区,灌木物种导管直径较小并采用安全优先的导水策略[56].尤其是在降水稀少且高温的干旱区,水分为植物生存生长的重要限制因子,植物对水力效率和水力安全的权衡关系研究需要进一步探究. ...
黄土丘陵区典型落叶树种的水力学特性研究
3
2014
... 植物木质部可通过改变导管结构特征来应对干旱胁迫.木质部常通过减小导管直径,增加导管密度、纹孔膜厚度和木材密度,进而降低木质部导水率并提高水力安全性,且已有大量研究证实(图2)[13,42,44].He等[13]指出相比于湿润环境,在干旱环境下水力失败风险较大且维护成本较高,植物通常会降低导水率以适应环境.Schuldt等[42]发现欧洲山毛榉的木质部导管直径降低7%且纹孔膜厚度增加15%以应对干旱胁迫.导管结构特征影响木质部导水率,Hagen-Poiseuille水力传导方程显示导管导水率与导管直径的4次方成正比,因此导管直径的微小增加会带来导水率的显著提高[41].有研究采用X射线显微断层成像法观测干燥不同时间的赤桉的木质部横截面的导管栓塞情况,结果显示Hagen-Poiseuille水力传导方程模拟的理论导水率高于实际最大导水率[10],原因一是与没有考虑导管长度和导管间的纹孔结构有关[41],导管长度越短、纹孔数量越少和纹孔面积越小,则导管导水率越低[41],纹孔阻力约占整个植株总水力传导阻力的14%~84%[40];二是与导管腔的不规则形状和锥度有关[45];三是与木质部汁液有关,研究显示KCl浓度的增加可导致木质部导水率增加10%[45].枝条形态学基部导管直径通常高于形态学末端[8,40],这一形态结构有利于减小由于植物高度增长带来的水力阻力[8].通常情况下导管直径表现为主干直径>侧枝>枝条交叉点[40].植物体的枝条交叉点、根茎过渡区和节点部位的导管直径较小、导水率较小、胡伯尔值较大和抗栓塞能力较强,这些部位常被称为“水流限制区”和“安全区”[46].胡伯尔值越大,表示维持单位叶面积水分需要的木质部越多,可用于比较不同树种间的水力特征[19,40]. ...
... [45].枝条形态学基部导管直径通常高于形态学末端[8,40],这一形态结构有利于减小由于植物高度增长带来的水力阻力[8].通常情况下导管直径表现为主干直径>侧枝>枝条交叉点[40].植物体的枝条交叉点、根茎过渡区和节点部位的导管直径较小、导水率较小、胡伯尔值较大和抗栓塞能力较强,这些部位常被称为“水流限制区”和“安全区”[46].胡伯尔值越大,表示维持单位叶面积水分需要的木质部越多,可用于比较不同树种间的水力特征[19,40]. ...
... 植物远端叶片水力阻力较大,占全植株水力阻力的30%,是SPAC水力运输的瓶颈和“安全阀”[9,19,26,47].植物叶片蒸腾约占地球表面蒸腾的70%[26],植物叶片通常通过气孔开闭适应环境.叶片气孔变化主要受两个机制调节:一是由叶表皮细胞和保卫细胞之间的水力连接引发的被动调节机制;二是激素如脱落酸的主动调节机制[4].气孔对水力调节和激素调节的敏感度与植物的等水调节和非等水调节有关[4],前者是指植物叶片气孔在干旱胁迫下迅速关闭以保证植物叶片日最小水势保持不变,在水分充足时复水快,易死于碳饥饿;后者是指植物叶片气孔在干旱胁迫下仍保持打开,降低叶片日最小水势,在水分充足时复水慢,易死于水力失败[6,11-12,48].有趣的是,Kannenberg等[6]认为非等水调节植物在水分胁迫期间并没有增加碳同化量.在自然界中没有严格的等水植物和非等水植物,通常介于两者之间[6].当土壤处于中等干旱条件下,植物通常会趋向于非等水调节;而当植物处于严重干旱条件下,植物趋向于等水调节[6,49].植物的等水和非等水程度的量化常采用水力学面积,即测量边界叶水势和1∶1线围成的多边形面积,水力学面积越小越趋向于等水调节[50];也可采用凌晨和午间水势的斜率来表示等水程度,斜率接近0表示植物偏向于等水植物,接近1或大于1表示植物偏向于非等水植物[51].陈嘉嘉[52]研究结果显示,在腾格里沙漠生态系统中,常见的固沙植物油蒿(Artemisia ordosica)、柠条锦鸡儿(Caragana korshinskii)和花棒(Hedysarum scoparium)的凌晨和午间水势斜率分别是0.33、0.92和1.22,分别属于偏等水植物、偏非等水植物和严格非等水植物.植物叶片除了通过气孔开合应对轻微水分胁迫,也可改变叶片解剖形态特征是对干旱胁迫做出的短期快速响应,如叶片气孔在干旱条件下呈小而密的分布(图2),可通过增加气孔闭合能力来减少蒸腾过程中的水分散失[44-45]. ...
树木水力结构模型与耐旱机理研究
1
2003
... 植物木质部可通过改变导管结构特征来应对干旱胁迫.木质部常通过减小导管直径,增加导管密度、纹孔膜厚度和木材密度,进而降低木质部导水率并提高水力安全性,且已有大量研究证实(图2)[13,42,44].He等[13]指出相比于湿润环境,在干旱环境下水力失败风险较大且维护成本较高,植物通常会降低导水率以适应环境.Schuldt等[42]发现欧洲山毛榉的木质部导管直径降低7%且纹孔膜厚度增加15%以应对干旱胁迫.导管结构特征影响木质部导水率,Hagen-Poiseuille水力传导方程显示导管导水率与导管直径的4次方成正比,因此导管直径的微小增加会带来导水率的显著提高[41].有研究采用X射线显微断层成像法观测干燥不同时间的赤桉的木质部横截面的导管栓塞情况,结果显示Hagen-Poiseuille水力传导方程模拟的理论导水率高于实际最大导水率[10],原因一是与没有考虑导管长度和导管间的纹孔结构有关[41],导管长度越短、纹孔数量越少和纹孔面积越小,则导管导水率越低[41],纹孔阻力约占整个植株总水力传导阻力的14%~84%[40];二是与导管腔的不规则形状和锥度有关[45];三是与木质部汁液有关,研究显示KCl浓度的增加可导致木质部导水率增加10%[45].枝条形态学基部导管直径通常高于形态学末端[8,40],这一形态结构有利于减小由于植物高度增长带来的水力阻力[8].通常情况下导管直径表现为主干直径>侧枝>枝条交叉点[40].植物体的枝条交叉点、根茎过渡区和节点部位的导管直径较小、导水率较小、胡伯尔值较大和抗栓塞能力较强,这些部位常被称为“水流限制区”和“安全区”[46].胡伯尔值越大,表示维持单位叶面积水分需要的木质部越多,可用于比较不同树种间的水力特征[19,40]. ...
Leaf hydraulics
1
2006
... 植物远端叶片水力阻力较大,占全植株水力阻力的30%,是SPAC水力运输的瓶颈和“安全阀”[9,19,26,47].植物叶片蒸腾约占地球表面蒸腾的70%[26],植物叶片通常通过气孔开闭适应环境.叶片气孔变化主要受两个机制调节:一是由叶表皮细胞和保卫细胞之间的水力连接引发的被动调节机制;二是激素如脱落酸的主动调节机制[4].气孔对水力调节和激素调节的敏感度与植物的等水调节和非等水调节有关[4],前者是指植物叶片气孔在干旱胁迫下迅速关闭以保证植物叶片日最小水势保持不变,在水分充足时复水快,易死于碳饥饿;后者是指植物叶片气孔在干旱胁迫下仍保持打开,降低叶片日最小水势,在水分充足时复水慢,易死于水力失败[6,11-12,48].有趣的是,Kannenberg等[6]认为非等水调节植物在水分胁迫期间并没有增加碳同化量.在自然界中没有严格的等水植物和非等水植物,通常介于两者之间[6].当土壤处于中等干旱条件下,植物通常会趋向于非等水调节;而当植物处于严重干旱条件下,植物趋向于等水调节[6,49].植物的等水和非等水程度的量化常采用水力学面积,即测量边界叶水势和1∶1线围成的多边形面积,水力学面积越小越趋向于等水调节[50];也可采用凌晨和午间水势的斜率来表示等水程度,斜率接近0表示植物偏向于等水植物,接近1或大于1表示植物偏向于非等水植物[51].陈嘉嘉[52]研究结果显示,在腾格里沙漠生态系统中,常见的固沙植物油蒿(Artemisia ordosica)、柠条锦鸡儿(Caragana korshinskii)和花棒(Hedysarum scoparium)的凌晨和午间水势斜率分别是0.33、0.92和1.22,分别属于偏等水植物、偏非等水植物和严格非等水植物.植物叶片除了通过气孔开合应对轻微水分胁迫,也可改变叶片解剖形态特征是对干旱胁迫做出的短期快速响应,如叶片气孔在干旱条件下呈小而密的分布(图2),可通过增加气孔闭合能力来减少蒸腾过程中的水分散失[44-45]. ...
木本植物水力系统对干旱胁迫的响应机制
1
2021
... 植物远端叶片水力阻力较大,占全植株水力阻力的30%,是SPAC水力运输的瓶颈和“安全阀”[9,19,26,47].植物叶片蒸腾约占地球表面蒸腾的70%[26],植物叶片通常通过气孔开闭适应环境.叶片气孔变化主要受两个机制调节:一是由叶表皮细胞和保卫细胞之间的水力连接引发的被动调节机制;二是激素如脱落酸的主动调节机制[4].气孔对水力调节和激素调节的敏感度与植物的等水调节和非等水调节有关[4],前者是指植物叶片气孔在干旱胁迫下迅速关闭以保证植物叶片日最小水势保持不变,在水分充足时复水快,易死于碳饥饿;后者是指植物叶片气孔在干旱胁迫下仍保持打开,降低叶片日最小水势,在水分充足时复水慢,易死于水力失败[6,11-12,48].有趣的是,Kannenberg等[6]认为非等水调节植物在水分胁迫期间并没有增加碳同化量.在自然界中没有严格的等水植物和非等水植物,通常介于两者之间[6].当土壤处于中等干旱条件下,植物通常会趋向于非等水调节;而当植物处于严重干旱条件下,植物趋向于等水调节[6,49].植物的等水和非等水程度的量化常采用水力学面积,即测量边界叶水势和1∶1线围成的多边形面积,水力学面积越小越趋向于等水调节[50];也可采用凌晨和午间水势的斜率来表示等水程度,斜率接近0表示植物偏向于等水植物,接近1或大于1表示植物偏向于非等水植物[51].陈嘉嘉[52]研究结果显示,在腾格里沙漠生态系统中,常见的固沙植物油蒿(Artemisia ordosica)、柠条锦鸡儿(Caragana korshinskii)和花棒(Hedysarum scoparium)的凌晨和午间水势斜率分别是0.33、0.92和1.22,分别属于偏等水植物、偏非等水植物和严格非等水植物.植物叶片除了通过气孔开合应对轻微水分胁迫,也可改变叶片解剖形态特征是对干旱胁迫做出的短期快速响应,如叶片气孔在干旱条件下呈小而密的分布(图2),可通过增加气孔闭合能力来减少蒸腾过程中的水分散失[44-45]. ...
Antecedent soil water content and vapor pressure deficit interactively control water potential in Larrea tridentata
1
2019
... 植物远端叶片水力阻力较大,占全植株水力阻力的30%,是SPAC水力运输的瓶颈和“安全阀”[9,19,26,47].植物叶片蒸腾约占地球表面蒸腾的70%[26],植物叶片通常通过气孔开闭适应环境.叶片气孔变化主要受两个机制调节:一是由叶表皮细胞和保卫细胞之间的水力连接引发的被动调节机制;二是激素如脱落酸的主动调节机制[4].气孔对水力调节和激素调节的敏感度与植物的等水调节和非等水调节有关[4],前者是指植物叶片气孔在干旱胁迫下迅速关闭以保证植物叶片日最小水势保持不变,在水分充足时复水快,易死于碳饥饿;后者是指植物叶片气孔在干旱胁迫下仍保持打开,降低叶片日最小水势,在水分充足时复水慢,易死于水力失败[6,11-12,48].有趣的是,Kannenberg等[6]认为非等水调节植物在水分胁迫期间并没有增加碳同化量.在自然界中没有严格的等水植物和非等水植物,通常介于两者之间[6].当土壤处于中等干旱条件下,植物通常会趋向于非等水调节;而当植物处于严重干旱条件下,植物趋向于等水调节[6,49].植物的等水和非等水程度的量化常采用水力学面积,即测量边界叶水势和1∶1线围成的多边形面积,水力学面积越小越趋向于等水调节[50];也可采用凌晨和午间水势的斜率来表示等水程度,斜率接近0表示植物偏向于等水植物,接近1或大于1表示植物偏向于非等水植物[51].陈嘉嘉[52]研究结果显示,在腾格里沙漠生态系统中,常见的固沙植物油蒿(Artemisia ordosica)、柠条锦鸡儿(Caragana korshinskii)和花棒(Hedysarum scoparium)的凌晨和午间水势斜率分别是0.33、0.92和1.22,分别属于偏等水植物、偏非等水植物和严格非等水植物.植物叶片除了通过气孔开合应对轻微水分胁迫,也可改变叶片解剖形态特征是对干旱胁迫做出的短期快速响应,如叶片气孔在干旱条件下呈小而密的分布(图2),可通过增加气孔闭合能力来减少蒸腾过程中的水分散失[44-45]. ...
不同土壤水分条件下毛白杨不同高度冠层的水分调节特征与机制
1
2020
... 植物远端叶片水力阻力较大,占全植株水力阻力的30%,是SPAC水力运输的瓶颈和“安全阀”[9,19,26,47].植物叶片蒸腾约占地球表面蒸腾的70%[26],植物叶片通常通过气孔开闭适应环境.叶片气孔变化主要受两个机制调节:一是由叶表皮细胞和保卫细胞之间的水力连接引发的被动调节机制;二是激素如脱落酸的主动调节机制[4].气孔对水力调节和激素调节的敏感度与植物的等水调节和非等水调节有关[4],前者是指植物叶片气孔在干旱胁迫下迅速关闭以保证植物叶片日最小水势保持不变,在水分充足时复水快,易死于碳饥饿;后者是指植物叶片气孔在干旱胁迫下仍保持打开,降低叶片日最小水势,在水分充足时复水慢,易死于水力失败[6,11-12,48].有趣的是,Kannenberg等[6]认为非等水调节植物在水分胁迫期间并没有增加碳同化量.在自然界中没有严格的等水植物和非等水植物,通常介于两者之间[6].当土壤处于中等干旱条件下,植物通常会趋向于非等水调节;而当植物处于严重干旱条件下,植物趋向于等水调节[6,49].植物的等水和非等水程度的量化常采用水力学面积,即测量边界叶水势和1∶1线围成的多边形面积,水力学面积越小越趋向于等水调节[50];也可采用凌晨和午间水势的斜率来表示等水程度,斜率接近0表示植物偏向于等水植物,接近1或大于1表示植物偏向于非等水植物[51].陈嘉嘉[52]研究结果显示,在腾格里沙漠生态系统中,常见的固沙植物油蒿(Artemisia ordosica)、柠条锦鸡儿(Caragana korshinskii)和花棒(Hedysarum scoparium)的凌晨和午间水势斜率分别是0.33、0.92和1.22,分别属于偏等水植物、偏非等水植物和严格非等水植物.植物叶片除了通过气孔开合应对轻微水分胁迫,也可改变叶片解剖形态特征是对干旱胁迫做出的短期快速响应,如叶片气孔在干旱条件下呈小而密的分布(图2),可通过增加气孔闭合能力来减少蒸腾过程中的水分散失[44-45]. ...
A new look at water transport regulation in plants
1
2014
... 植物远端叶片水力阻力较大,占全植株水力阻力的30%,是SPAC水力运输的瓶颈和“安全阀”[9,19,26,47].植物叶片蒸腾约占地球表面蒸腾的70%[26],植物叶片通常通过气孔开闭适应环境.叶片气孔变化主要受两个机制调节:一是由叶表皮细胞和保卫细胞之间的水力连接引发的被动调节机制;二是激素如脱落酸的主动调节机制[4].气孔对水力调节和激素调节的敏感度与植物的等水调节和非等水调节有关[4],前者是指植物叶片气孔在干旱胁迫下迅速关闭以保证植物叶片日最小水势保持不变,在水分充足时复水快,易死于碳饥饿;后者是指植物叶片气孔在干旱胁迫下仍保持打开,降低叶片日最小水势,在水分充足时复水慢,易死于水力失败[6,11-12,48].有趣的是,Kannenberg等[6]认为非等水调节植物在水分胁迫期间并没有增加碳同化量.在自然界中没有严格的等水植物和非等水植物,通常介于两者之间[6].当土壤处于中等干旱条件下,植物通常会趋向于非等水调节;而当植物处于严重干旱条件下,植物趋向于等水调节[6,49].植物的等水和非等水程度的量化常采用水力学面积,即测量边界叶水势和1∶1线围成的多边形面积,水力学面积越小越趋向于等水调节[50];也可采用凌晨和午间水势的斜率来表示等水程度,斜率接近0表示植物偏向于等水植物,接近1或大于1表示植物偏向于非等水植物[51].陈嘉嘉[52]研究结果显示,在腾格里沙漠生态系统中,常见的固沙植物油蒿(Artemisia ordosica)、柠条锦鸡儿(Caragana korshinskii)和花棒(Hedysarum scoparium)的凌晨和午间水势斜率分别是0.33、0.92和1.22,分别属于偏等水植物、偏非等水植物和严格非等水植物.植物叶片除了通过气孔开合应对轻微水分胁迫,也可改变叶片解剖形态特征是对干旱胁迫做出的短期快速响应,如叶片气孔在干旱条件下呈小而密的分布(图2),可通过增加气孔闭合能力来减少蒸腾过程中的水分散失[44-45]. ...
干旱和遮荫对典型固沙灌木生存的影响
1
2023
... 植物远端叶片水力阻力较大,占全植株水力阻力的30%,是SPAC水力运输的瓶颈和“安全阀”[9,19,26,47].植物叶片蒸腾约占地球表面蒸腾的70%[26],植物叶片通常通过气孔开闭适应环境.叶片气孔变化主要受两个机制调节:一是由叶表皮细胞和保卫细胞之间的水力连接引发的被动调节机制;二是激素如脱落酸的主动调节机制[4].气孔对水力调节和激素调节的敏感度与植物的等水调节和非等水调节有关[4],前者是指植物叶片气孔在干旱胁迫下迅速关闭以保证植物叶片日最小水势保持不变,在水分充足时复水快,易死于碳饥饿;后者是指植物叶片气孔在干旱胁迫下仍保持打开,降低叶片日最小水势,在水分充足时复水慢,易死于水力失败[6,11-12,48].有趣的是,Kannenberg等[6]认为非等水调节植物在水分胁迫期间并没有增加碳同化量.在自然界中没有严格的等水植物和非等水植物,通常介于两者之间[6].当土壤处于中等干旱条件下,植物通常会趋向于非等水调节;而当植物处于严重干旱条件下,植物趋向于等水调节[6,49].植物的等水和非等水程度的量化常采用水力学面积,即测量边界叶水势和1∶1线围成的多边形面积,水力学面积越小越趋向于等水调节[50];也可采用凌晨和午间水势的斜率来表示等水程度,斜率接近0表示植物偏向于等水植物,接近1或大于1表示植物偏向于非等水植物[51].陈嘉嘉[52]研究结果显示,在腾格里沙漠生态系统中,常见的固沙植物油蒿(Artemisia ordosica)、柠条锦鸡儿(Caragana korshinskii)和花棒(Hedysarum scoparium)的凌晨和午间水势斜率分别是0.33、0.92和1.22,分别属于偏等水植物、偏非等水植物和严格非等水植物.植物叶片除了通过气孔开合应对轻微水分胁迫,也可改变叶片解剖形态特征是对干旱胁迫做出的短期快速响应,如叶片气孔在干旱条件下呈小而密的分布(图2),可通过增加气孔闭合能力来减少蒸腾过程中的水分散失[44-45]. ...
Hydraulic safety and growth rather than climate of origin influence survival in desert shrubs and trees
1
2023
... 植物进行形态调整,根系水力再分配,改变木质部和叶片的结构特征的同时,也会进行水力效率和水力安全之间的权衡.水力效率常采用水力导度来衡量,它是水力阻力的倒数,根据欧姆定律,水力导度可用液流速率与压力差比值表示[9],常采用、表征植物木质部的抗栓塞性,采用水力导度降低50%和88%时对应的水势(,)表征植物叶片的抗栓塞性,即植物木质部和叶片的水力安全[42],其值越低表示植物越能忍受较低水势,抗栓塞性越强[30].木质部抗栓塞性和干旱度指数呈线性增加趋势,在干旱胁迫发生时木质部能够忍受更低的水势保证水分运输的正常运行[42].“水力权衡假说”认为植物的不同器官间,存在水力传导效率和安全的高低权衡[42-43].Huo等[44]认为在干旱沙区,柠条锦鸡儿通过降低导水率并增加水力安全性来保证植物能够在干旱胁迫下生存;Zhang等[53]发现生长在干旱沙漠生态系统的植物通常具有较高的水力安全性和较低的导水率,植物的存活与植物水力安全密切相关.而目前也有研究显示植物水力效率和安全不存在权衡关系[42-43];低水力效率和低水力安全的植物可在湿润地区生存[43];也有进一步研究指出自然界中该权衡关系并不是非此即彼的,而是根据环境和植物结构特征而变化[54].水力效率和水力安全之间的权衡关系是由导管结构特征和导管间纹孔结构决定的,水力效率高的木质部通常导管直径较大、纹孔结构较多且木材密度较低,相应的水力安全性较低[55].研究显示在中国北方干旱区,灌木物种在相对干旱区根部导管直径较大并采用效率优先的导水策略;而在相对湿润区,灌木物种导管直径较小并采用安全优先的导水策略[56].尤其是在降水稀少且高温的干旱区,水分为植物生存生长的重要限制因子,植物对水力效率和水力安全的权衡关系研究需要进一步探究. ...
鸡公山地区不同材性树种枝木质部水力与解剖结构的关系
1
2022
... 植物进行形态调整,根系水力再分配,改变木质部和叶片的结构特征的同时,也会进行水力效率和水力安全之间的权衡.水力效率常采用水力导度来衡量,它是水力阻力的倒数,根据欧姆定律,水力导度可用液流速率与压力差比值表示[9],常采用、表征植物木质部的抗栓塞性,采用水力导度降低50%和88%时对应的水势(,)表征植物叶片的抗栓塞性,即植物木质部和叶片的水力安全[42],其值越低表示植物越能忍受较低水势,抗栓塞性越强[30].木质部抗栓塞性和干旱度指数呈线性增加趋势,在干旱胁迫发生时木质部能够忍受更低的水势保证水分运输的正常运行[42].“水力权衡假说”认为植物的不同器官间,存在水力传导效率和安全的高低权衡[42-43].Huo等[44]认为在干旱沙区,柠条锦鸡儿通过降低导水率并增加水力安全性来保证植物能够在干旱胁迫下生存;Zhang等[53]发现生长在干旱沙漠生态系统的植物通常具有较高的水力安全性和较低的导水率,植物的存活与植物水力安全密切相关.而目前也有研究显示植物水力效率和安全不存在权衡关系[42-43];低水力效率和低水力安全的植物可在湿润地区生存[43];也有进一步研究指出自然界中该权衡关系并不是非此即彼的,而是根据环境和植物结构特征而变化[54].水力效率和水力安全之间的权衡关系是由导管结构特征和导管间纹孔结构决定的,水力效率高的木质部通常导管直径较大、纹孔结构较多且木材密度较低,相应的水力安全性较低[55].研究显示在中国北方干旱区,灌木物种在相对干旱区根部导管直径较大并采用效率优先的导水策略;而在相对湿润区,灌木物种导管直径较小并采用安全优先的导水策略[56].尤其是在降水稀少且高温的干旱区,水分为植物生存生长的重要限制因子,植物对水力效率和水力安全的权衡关系研究需要进一步探究. ...
Hydraulic limits on maximum plant transpiration and the emergence of the safety-efficiency trade-off
1
2013
... 植物进行形态调整,根系水力再分配,改变木质部和叶片的结构特征的同时,也会进行水力效率和水力安全之间的权衡.水力效率常采用水力导度来衡量,它是水力阻力的倒数,根据欧姆定律,水力导度可用液流速率与压力差比值表示[9],常采用、表征植物木质部的抗栓塞性,采用水力导度降低50%和88%时对应的水势(,)表征植物叶片的抗栓塞性,即植物木质部和叶片的水力安全[42],其值越低表示植物越能忍受较低水势,抗栓塞性越强[30].木质部抗栓塞性和干旱度指数呈线性增加趋势,在干旱胁迫发生时木质部能够忍受更低的水势保证水分运输的正常运行[42].“水力权衡假说”认为植物的不同器官间,存在水力传导效率和安全的高低权衡[42-43].Huo等[44]认为在干旱沙区,柠条锦鸡儿通过降低导水率并增加水力安全性来保证植物能够在干旱胁迫下生存;Zhang等[53]发现生长在干旱沙漠生态系统的植物通常具有较高的水力安全性和较低的导水率,植物的存活与植物水力安全密切相关.而目前也有研究显示植物水力效率和安全不存在权衡关系[42-43];低水力效率和低水力安全的植物可在湿润地区生存[43];也有进一步研究指出自然界中该权衡关系并不是非此即彼的,而是根据环境和植物结构特征而变化[54].水力效率和水力安全之间的权衡关系是由导管结构特征和导管间纹孔结构决定的,水力效率高的木质部通常导管直径较大、纹孔结构较多且木材密度较低,相应的水力安全性较低[55].研究显示在中国北方干旱区,灌木物种在相对干旱区根部导管直径较大并采用效率优先的导水策略;而在相对湿润区,灌木物种导管直径较小并采用安全优先的导水策略[56].尤其是在降水稀少且高温的干旱区,水分为植物生存生长的重要限制因子,植物对水力效率和水力安全的权衡关系研究需要进一步探究. ...
中国干旱区草本和灌木根部导管特征及随环境变化
1
2023
... 植物进行形态调整,根系水力再分配,改变木质部和叶片的结构特征的同时,也会进行水力效率和水力安全之间的权衡.水力效率常采用水力导度来衡量,它是水力阻力的倒数,根据欧姆定律,水力导度可用液流速率与压力差比值表示[9],常采用、表征植物木质部的抗栓塞性,采用水力导度降低50%和88%时对应的水势(,)表征植物叶片的抗栓塞性,即植物木质部和叶片的水力安全[42],其值越低表示植物越能忍受较低水势,抗栓塞性越强[30].木质部抗栓塞性和干旱度指数呈线性增加趋势,在干旱胁迫发生时木质部能够忍受更低的水势保证水分运输的正常运行[42].“水力权衡假说”认为植物的不同器官间,存在水力传导效率和安全的高低权衡[42-43].Huo等[44]认为在干旱沙区,柠条锦鸡儿通过降低导水率并增加水力安全性来保证植物能够在干旱胁迫下生存;Zhang等[53]发现生长在干旱沙漠生态系统的植物通常具有较高的水力安全性和较低的导水率,植物的存活与植物水力安全密切相关.而目前也有研究显示植物水力效率和安全不存在权衡关系[42-43];低水力效率和低水力安全的植物可在湿润地区生存[43];也有进一步研究指出自然界中该权衡关系并不是非此即彼的,而是根据环境和植物结构特征而变化[54].水力效率和水力安全之间的权衡关系是由导管结构特征和导管间纹孔结构决定的,水力效率高的木质部通常导管直径较大、纹孔结构较多且木材密度较低,相应的水力安全性较低[55].研究显示在中国北方干旱区,灌木物种在相对干旱区根部导管直径较大并采用效率优先的导水策略;而在相对湿润区,灌木物种导管直径较小并采用安全优先的导水策略[56].尤其是在降水稀少且高温的干旱区,水分为植物生存生长的重要限制因子,植物对水力效率和水力安全的权衡关系研究需要进一步探究. ...
基于植物水力学的中国森林蒸腾模拟研究
1
2022
... 植物个体水力传导过程包括植物根系吸水、木质部导水和叶片蒸腾失水3个过程,该过程受气象因子、土壤水分有效性和植物水力特性的共同影响.关于SPAC水分传输的模型较多,而基于过程的模型较少.大叶模型将SPAC中植物简化为一片叶片,仅表征土壤水分胁迫与植物叶片蒸腾和光合之间的关系,忽略了植物根系吸水和茎干导水的水力传导过程[57].由于根系的不可见性和难获取性,直接测量根系吸水具有一定困难,建立基于过程的模型准确模拟根系吸水有助于了解土壤水文和植物生长[58-60].根系吸水模拟常采用Feddes模型,其运行需要潜在根系吸水量、水分胁迫函数和根系分布密度函数等,常将其与Richards土壤水分运移模型结合起来使用(即Hydrus模型),用于准确模拟根系吸水量、水分通量和土壤水分等重要水分参数以及植物生长[58].Wang等[59]采用Hydrus模型指出根系吸水量减少和土壤蒸发量增加是阻碍夏玉米在斥水性土壤减产中的主要原因.Tu等[60]采用Hydrus模型模拟农作物在生长季的蒸散量和深层渗漏量,用于指导农业灌溉用水.Sperry模型有效提高了根系吸水、冠层蒸腾和叶片水势间的耦合性,提供了模拟SPAC水分传输的机理性[61-62].由于测量林分尺度上所有植物的单株蒸腾较不现实,因此可通过测量植物株高、冠幅、叶面积指数、地径和占地面积等作为尺度扩展因子将单株蒸腾从个体尺度扩展至林分尺度,目前采用边材面积作为扩展因子推导林分蒸腾较为准确[18,61],边材面积的量化可采用边材面积与胸径之间的经验函数来表示[61]. ...
2
1978
... 植物个体水力传导过程包括植物根系吸水、木质部导水和叶片蒸腾失水3个过程,该过程受气象因子、土壤水分有效性和植物水力特性的共同影响.关于SPAC水分传输的模型较多,而基于过程的模型较少.大叶模型将SPAC中植物简化为一片叶片,仅表征土壤水分胁迫与植物叶片蒸腾和光合之间的关系,忽略了植物根系吸水和茎干导水的水力传导过程[57].由于根系的不可见性和难获取性,直接测量根系吸水具有一定困难,建立基于过程的模型准确模拟根系吸水有助于了解土壤水文和植物生长[58-60].根系吸水模拟常采用Feddes模型,其运行需要潜在根系吸水量、水分胁迫函数和根系分布密度函数等,常将其与Richards土壤水分运移模型结合起来使用(即Hydrus模型),用于准确模拟根系吸水量、水分通量和土壤水分等重要水分参数以及植物生长[58].Wang等[59]采用Hydrus模型指出根系吸水量减少和土壤蒸发量增加是阻碍夏玉米在斥水性土壤减产中的主要原因.Tu等[60]采用Hydrus模型模拟农作物在生长季的蒸散量和深层渗漏量,用于指导农业灌溉用水.Sperry模型有效提高了根系吸水、冠层蒸腾和叶片水势间的耦合性,提供了模拟SPAC水分传输的机理性[61-62].由于测量林分尺度上所有植物的单株蒸腾较不现实,因此可通过测量植物株高、冠幅、叶面积指数、地径和占地面积等作为尺度扩展因子将单株蒸腾从个体尺度扩展至林分尺度,目前采用边材面积作为扩展因子推导林分蒸腾较为准确[18,61],边材面积的量化可采用边材面积与胸径之间的经验函数来表示[61]. ...
... [58].Wang等[59]采用Hydrus模型指出根系吸水量减少和土壤蒸发量增加是阻碍夏玉米在斥水性土壤减产中的主要原因.Tu等[60]采用Hydrus模型模拟农作物在生长季的蒸散量和深层渗漏量,用于指导农业灌溉用水.Sperry模型有效提高了根系吸水、冠层蒸腾和叶片水势间的耦合性,提供了模拟SPAC水分传输的机理性[61-62].由于测量林分尺度上所有植物的单株蒸腾较不现实,因此可通过测量植物株高、冠幅、叶面积指数、地径和占地面积等作为尺度扩展因子将单株蒸腾从个体尺度扩展至林分尺度,目前采用边材面积作为扩展因子推导林分蒸腾较为准确[18,61],边材面积的量化可采用边材面积与胸径之间的经验函数来表示[61]. ...
Reduced root water uptake of summer maize grown in water-repellent soils simulated by HYDRUS-1D
1
2021
... 植物个体水力传导过程包括植物根系吸水、木质部导水和叶片蒸腾失水3个过程,该过程受气象因子、土壤水分有效性和植物水力特性的共同影响.关于SPAC水分传输的模型较多,而基于过程的模型较少.大叶模型将SPAC中植物简化为一片叶片,仅表征土壤水分胁迫与植物叶片蒸腾和光合之间的关系,忽略了植物根系吸水和茎干导水的水力传导过程[57].由于根系的不可见性和难获取性,直接测量根系吸水具有一定困难,建立基于过程的模型准确模拟根系吸水有助于了解土壤水文和植物生长[58-60].根系吸水模拟常采用Feddes模型,其运行需要潜在根系吸水量、水分胁迫函数和根系分布密度函数等,常将其与Richards土壤水分运移模型结合起来使用(即Hydrus模型),用于准确模拟根系吸水量、水分通量和土壤水分等重要水分参数以及植物生长[58].Wang等[59]采用Hydrus模型指出根系吸水量减少和土壤蒸发量增加是阻碍夏玉米在斥水性土壤减产中的主要原因.Tu等[60]采用Hydrus模型模拟农作物在生长季的蒸散量和深层渗漏量,用于指导农业灌溉用水.Sperry模型有效提高了根系吸水、冠层蒸腾和叶片水势间的耦合性,提供了模拟SPAC水分传输的机理性[61-62].由于测量林分尺度上所有植物的单株蒸腾较不现实,因此可通过测量植物株高、冠幅、叶面积指数、地径和占地面积等作为尺度扩展因子将单株蒸腾从个体尺度扩展至林分尺度,目前采用边材面积作为扩展因子推导林分蒸腾较为准确[18,61],边材面积的量化可采用边材面积与胸径之间的经验函数来表示[61]. ...
Water budget components estimation for a mature citrus orchard of southern China based on HYDRUS-1D model
2
2021
... 植物个体水力传导过程包括植物根系吸水、木质部导水和叶片蒸腾失水3个过程,该过程受气象因子、土壤水分有效性和植物水力特性的共同影响.关于SPAC水分传输的模型较多,而基于过程的模型较少.大叶模型将SPAC中植物简化为一片叶片,仅表征土壤水分胁迫与植物叶片蒸腾和光合之间的关系,忽略了植物根系吸水和茎干导水的水力传导过程[57].由于根系的不可见性和难获取性,直接测量根系吸水具有一定困难,建立基于过程的模型准确模拟根系吸水有助于了解土壤水文和植物生长[58-60].根系吸水模拟常采用Feddes模型,其运行需要潜在根系吸水量、水分胁迫函数和根系分布密度函数等,常将其与Richards土壤水分运移模型结合起来使用(即Hydrus模型),用于准确模拟根系吸水量、水分通量和土壤水分等重要水分参数以及植物生长[58].Wang等[59]采用Hydrus模型指出根系吸水量减少和土壤蒸发量增加是阻碍夏玉米在斥水性土壤减产中的主要原因.Tu等[60]采用Hydrus模型模拟农作物在生长季的蒸散量和深层渗漏量,用于指导农业灌溉用水.Sperry模型有效提高了根系吸水、冠层蒸腾和叶片水势间的耦合性,提供了模拟SPAC水分传输的机理性[61-62].由于测量林分尺度上所有植物的单株蒸腾较不现实,因此可通过测量植物株高、冠幅、叶面积指数、地径和占地面积等作为尺度扩展因子将单株蒸腾从个体尺度扩展至林分尺度,目前采用边材面积作为扩展因子推导林分蒸腾较为准确[18,61],边材面积的量化可采用边材面积与胸径之间的经验函数来表示[61]. ...
... [60]采用Hydrus模型模拟农作物在生长季的蒸散量和深层渗漏量,用于指导农业灌溉用水.Sperry模型有效提高了根系吸水、冠层蒸腾和叶片水势间的耦合性,提供了模拟SPAC水分传输的机理性[61-62].由于测量林分尺度上所有植物的单株蒸腾较不现实,因此可通过测量植物株高、冠幅、叶面积指数、地径和占地面积等作为尺度扩展因子将单株蒸腾从个体尺度扩展至林分尺度,目前采用边材面积作为扩展因子推导林分蒸腾较为准确[18,61],边材面积的量化可采用边材面积与胸径之间的经验函数来表示[61]. ...
黄土高原刺槐生长及水分传输关键过程对气候变化的响应
3
2024
... 植物个体水力传导过程包括植物根系吸水、木质部导水和叶片蒸腾失水3个过程,该过程受气象因子、土壤水分有效性和植物水力特性的共同影响.关于SPAC水分传输的模型较多,而基于过程的模型较少.大叶模型将SPAC中植物简化为一片叶片,仅表征土壤水分胁迫与植物叶片蒸腾和光合之间的关系,忽略了植物根系吸水和茎干导水的水力传导过程[57].由于根系的不可见性和难获取性,直接测量根系吸水具有一定困难,建立基于过程的模型准确模拟根系吸水有助于了解土壤水文和植物生长[58-60].根系吸水模拟常采用Feddes模型,其运行需要潜在根系吸水量、水分胁迫函数和根系分布密度函数等,常将其与Richards土壤水分运移模型结合起来使用(即Hydrus模型),用于准确模拟根系吸水量、水分通量和土壤水分等重要水分参数以及植物生长[58].Wang等[59]采用Hydrus模型指出根系吸水量减少和土壤蒸发量增加是阻碍夏玉米在斥水性土壤减产中的主要原因.Tu等[60]采用Hydrus模型模拟农作物在生长季的蒸散量和深层渗漏量,用于指导农业灌溉用水.Sperry模型有效提高了根系吸水、冠层蒸腾和叶片水势间的耦合性,提供了模拟SPAC水分传输的机理性[61-62].由于测量林分尺度上所有植物的单株蒸腾较不现实,因此可通过测量植物株高、冠幅、叶面积指数、地径和占地面积等作为尺度扩展因子将单株蒸腾从个体尺度扩展至林分尺度,目前采用边材面积作为扩展因子推导林分蒸腾较为准确[18,61],边材面积的量化可采用边材面积与胸径之间的经验函数来表示[61]. ...
... ,61],边材面积的量化可采用边材面积与胸径之间的经验函数来表示[61]. ...
... [61]. ...
黄土高原降水梯度带刺槐SPAC系统水分传输对干旱环境的适应性
1
2020
... 植物个体水力传导过程包括植物根系吸水、木质部导水和叶片蒸腾失水3个过程,该过程受气象因子、土壤水分有效性和植物水力特性的共同影响.关于SPAC水分传输的模型较多,而基于过程的模型较少.大叶模型将SPAC中植物简化为一片叶片,仅表征土壤水分胁迫与植物叶片蒸腾和光合之间的关系,忽略了植物根系吸水和茎干导水的水力传导过程[57].由于根系的不可见性和难获取性,直接测量根系吸水具有一定困难,建立基于过程的模型准确模拟根系吸水有助于了解土壤水文和植物生长[58-60].根系吸水模拟常采用Feddes模型,其运行需要潜在根系吸水量、水分胁迫函数和根系分布密度函数等,常将其与Richards土壤水分运移模型结合起来使用(即Hydrus模型),用于准确模拟根系吸水量、水分通量和土壤水分等重要水分参数以及植物生长[58].Wang等[59]采用Hydrus模型指出根系吸水量减少和土壤蒸发量增加是阻碍夏玉米在斥水性土壤减产中的主要原因.Tu等[60]采用Hydrus模型模拟农作物在生长季的蒸散量和深层渗漏量,用于指导农业灌溉用水.Sperry模型有效提高了根系吸水、冠层蒸腾和叶片水势间的耦合性,提供了模拟SPAC水分传输的机理性[61-62].由于测量林分尺度上所有植物的单株蒸腾较不现实,因此可通过测量植物株高、冠幅、叶面积指数、地径和占地面积等作为尺度扩展因子将单株蒸腾从个体尺度扩展至林分尺度,目前采用边材面积作为扩展因子推导林分蒸腾较为准确[18,61],边材面积的量化可采用边材面积与胸径之间的经验函数来表示[61]. ...




 甘公网安备 62010202000688号
甘公网安备 62010202000688号
{kind=link}
{kind=link}
{kind=link}
{kind=link}