Drylands in the earth system
1
2010
... 旱区指干旱指数(降水与潜在蒸散量的比值)不高于0.65的地区,大约覆盖全球41%的陆地表面,是地球上最大的陆地生态系统类型,承载着全球38%的人口,其中许多人主要依靠初级生产以维持生计[1-2].10%~20%的旱区已出现不同程度的退化现象[3].预计到21世纪末,旱区面积将进一步扩大11%~23%[4].在全球气候变化的大背景下,旱区退化的风险愈发严峻[5].因此,迫切需要采取措施防止旱区退化、促进旱区恢复与可持续发展,而深入了解旱区生态系统动态是第一步[6]. ...
Drylands extent and environmental issues:a global approach
1
2016
... 旱区指干旱指数(降水与潜在蒸散量的比值)不高于0.65的地区,大约覆盖全球41%的陆地表面,是地球上最大的陆地生态系统类型,承载着全球38%的人口,其中许多人主要依靠初级生产以维持生计[1-2].10%~20%的旱区已出现不同程度的退化现象[3].预计到21世纪末,旱区面积将进一步扩大11%~23%[4].在全球气候变化的大背景下,旱区退化的风险愈发严峻[5].因此,迫切需要采取措施防止旱区退化、促进旱区恢复与可持续发展,而深入了解旱区生态系统动态是第一步[6]. ...
Global drylands:A UN System-wide Response
1
2011
... 旱区指干旱指数(降水与潜在蒸散量的比值)不高于0.65的地区,大约覆盖全球41%的陆地表面,是地球上最大的陆地生态系统类型,承载着全球38%的人口,其中许多人主要依靠初级生产以维持生计[1-2].10%~20%的旱区已出现不同程度的退化现象[3].预计到21世纪末,旱区面积将进一步扩大11%~23%[4].在全球气候变化的大背景下,旱区退化的风险愈发严峻[5].因此,迫切需要采取措施防止旱区退化、促进旱区恢复与可持续发展,而深入了解旱区生态系统动态是第一步[6]. ...
Accelerated dryland expansion under climate change
1
2016
... 旱区指干旱指数(降水与潜在蒸散量的比值)不高于0.65的地区,大约覆盖全球41%的陆地表面,是地球上最大的陆地生态系统类型,承载着全球38%的人口,其中许多人主要依靠初级生产以维持生计[1-2].10%~20%的旱区已出现不同程度的退化现象[3].预计到21世纪末,旱区面积将进一步扩大11%~23%[4].在全球气候变化的大背景下,旱区退化的风险愈发严峻[5].因此,迫切需要采取措施防止旱区退化、促进旱区恢复与可持续发展,而深入了解旱区生态系统动态是第一步[6]. ...
A multiscale,hierarchical model of pulse dynamics in arid-land ecosystems
1
2014
... 旱区指干旱指数(降水与潜在蒸散量的比值)不高于0.65的地区,大约覆盖全球41%的陆地表面,是地球上最大的陆地生态系统类型,承载着全球38%的人口,其中许多人主要依靠初级生产以维持生计[1-2].10%~20%的旱区已出现不同程度的退化现象[3].预计到21世纪末,旱区面积将进一步扩大11%~23%[4].在全球气候变化的大背景下,旱区退化的风险愈发严峻[5].因此,迫切需要采取措施防止旱区退化、促进旱区恢复与可持续发展,而深入了解旱区生态系统动态是第一步[6]. ...
中国西北干旱区水资源与生态环境变化及保护建议
1
2023
... 旱区指干旱指数(降水与潜在蒸散量的比值)不高于0.65的地区,大约覆盖全球41%的陆地表面,是地球上最大的陆地生态系统类型,承载着全球38%的人口,其中许多人主要依靠初级生产以维持生计[1-2].10%~20%的旱区已出现不同程度的退化现象[3].预计到21世纪末,旱区面积将进一步扩大11%~23%[4].在全球气候变化的大背景下,旱区退化的风险愈发严峻[5].因此,迫切需要采取措施防止旱区退化、促进旱区恢复与可持续发展,而深入了解旱区生态系统动态是第一步[6]. ...
1
2016
... 作为旱区特有生物组分[7],生物土壤结皮(由土壤颗粒与不同比例的光合自养生物(蓝藻细菌、藻类、地衣、苔藓植物)和异养生物(细菌、真菌、古细菌)胶结而成的表面土壤复合体[8-9])约覆盖全球旱区面积的30%[10].生物土壤结皮作为土壤与大气间的交互界面[11],在调控生态水文过程(如水分蒸发、径流入渗)和土壤生物地球化学循环(包括水循环、养分循环等)方面发挥着重要的生态系统功能和服务[12-16].生物土壤结皮固氮占全球陆地生物固氮量的40%~85%[10].因此,生物土壤结皮被称为旱区生态系统工程师[17],将其纳入研究框架对理解和管理旱区生态系统十分关键[18]. ...
What is a biocrust?A refined,contemporary definition for a broadening research community
1
2022
... 作为旱区特有生物组分[7],生物土壤结皮(由土壤颗粒与不同比例的光合自养生物(蓝藻细菌、藻类、地衣、苔藓植物)和异养生物(细菌、真菌、古细菌)胶结而成的表面土壤复合体[8-9])约覆盖全球旱区面积的30%[10].生物土壤结皮作为土壤与大气间的交互界面[11],在调控生态水文过程(如水分蒸发、径流入渗)和土壤生物地球化学循环(包括水循环、养分循环等)方面发挥着重要的生态系统功能和服务[12-16].生物土壤结皮固氮占全球陆地生物固氮量的40%~85%[10].因此,生物土壤结皮被称为旱区生态系统工程师[17],将其纳入研究框架对理解和管理旱区生态系统十分关键[18]. ...
1
2003
... 作为旱区特有生物组分[7],生物土壤结皮(由土壤颗粒与不同比例的光合自养生物(蓝藻细菌、藻类、地衣、苔藓植物)和异养生物(细菌、真菌、古细菌)胶结而成的表面土壤复合体[8-9])约覆盖全球旱区面积的30%[10].生物土壤结皮作为土壤与大气间的交互界面[11],在调控生态水文过程(如水分蒸发、径流入渗)和土壤生物地球化学循环(包括水循环、养分循环等)方面发挥着重要的生态系统功能和服务[12-16].生物土壤结皮固氮占全球陆地生物固氮量的40%~85%[10].因此,生物土壤结皮被称为旱区生态系统工程师[17],将其纳入研究框架对理解和管理旱区生态系统十分关键[18]. ...
Dryland photoautotrophic soil surface communities endangered by global change
2
2018
... 作为旱区特有生物组分[7],生物土壤结皮(由土壤颗粒与不同比例的光合自养生物(蓝藻细菌、藻类、地衣、苔藓植物)和异养生物(细菌、真菌、古细菌)胶结而成的表面土壤复合体[8-9])约覆盖全球旱区面积的30%[10].生物土壤结皮作为土壤与大气间的交互界面[11],在调控生态水文过程(如水分蒸发、径流入渗)和土壤生物地球化学循环(包括水循环、养分循环等)方面发挥着重要的生态系统功能和服务[12-16].生物土壤结皮固氮占全球陆地生物固氮量的40%~85%[10].因此,生物土壤结皮被称为旱区生态系统工程师[17],将其纳入研究框架对理解和管理旱区生态系统十分关键[18]. ...
... [10].因此,生物土壤结皮被称为旱区生态系统工程师[17],将其纳入研究框架对理解和管理旱区生态系统十分关键[18]. ...
Carbon flux in deserts depends on soil cover type:a case study in the Gurbantunggute Desert,North China
1
2013
... 作为旱区特有生物组分[7],生物土壤结皮(由土壤颗粒与不同比例的光合自养生物(蓝藻细菌、藻类、地衣、苔藓植物)和异养生物(细菌、真菌、古细菌)胶结而成的表面土壤复合体[8-9])约覆盖全球旱区面积的30%[10].生物土壤结皮作为土壤与大气间的交互界面[11],在调控生态水文过程(如水分蒸发、径流入渗)和土壤生物地球化学循环(包括水循环、养分循环等)方面发挥着重要的生态系统功能和服务[12-16].生物土壤结皮固氮占全球陆地生物固氮量的40%~85%[10].因此,生物土壤结皮被称为旱区生态系统工程师[17],将其纳入研究框架对理解和管理旱区生态系统十分关键[18]. ...
Biological soil crusts as soil stabilizers
1
2016
... 作为旱区特有生物组分[7],生物土壤结皮(由土壤颗粒与不同比例的光合自养生物(蓝藻细菌、藻类、地衣、苔藓植物)和异养生物(细菌、真菌、古细菌)胶结而成的表面土壤复合体[8-9])约覆盖全球旱区面积的30%[10].生物土壤结皮作为土壤与大气间的交互界面[11],在调控生态水文过程(如水分蒸发、径流入渗)和土壤生物地球化学循环(包括水循环、养分循环等)方面发挥着重要的生态系统功能和服务[12-16].生物土壤结皮固氮占全球陆地生物固氮量的40%~85%[10].因此,生物土壤结皮被称为旱区生态系统工程师[17],将其纳入研究框架对理解和管理旱区生态系统十分关键[18]. ...
The role of biocrusts in arid land hydrology
2
2016
... 降水作为旱区生态系统的主要水源,深刻影响生物土壤结皮与维管植物的共存.以较低年降雨量90~150 mm为例,生物土壤结皮相较维管植物往往更具优势[31].主要因为维管植物对水分的需求量大,而生物土壤结皮的需求则小得多.随着降水频率的增加,生物土壤结皮适应的降水条件范围变宽.当日降水频率为0.025时,生物土壤结皮适宜在0~100 mm年降水量范围生长;当日降水频率为1时,生物土壤结皮适宜在0~500 mm年降水量区间生长[33],有力地促进了生物土壤结皮-维管植物共存.干旱事件(降水量显著降低)会降低降雨在土壤中的入渗深度,可能极大降低深根系维管植物的水分利用效率,但并不会对生物土壤结皮产生显著影响[13,46],呈现以生物土壤结皮为主的共存状态.随着降水的增加,生物土壤结皮与维管植物的可利用水分均有所增加,不过两者的竞争优势也随之变化,可能由于过多水分超出生物土壤结皮的可利用范围,增加的降水更多地被维管植物所利用[27].同时,维管植物的盖度过高会极大限制下层光照的有效性,进而导致生物土壤结皮的盖度下降[21].当超过某一临界点时,便会发生从生物土壤结皮主导状态向维管植物主导状态的转变[27].另外,由于生物土壤结皮的存在,降雨模式(降雨量或降雨时间分布)的改变会对灌木斑块和裸露斑块的土壤水分可用性动态产生不同的影响[56].总之,生物土壤结皮和维管植物之间的相互作用对降水的响应是极其复杂的,需考虑研究区域和降水模式的特点[21]. ...
... 在旱区生态系统,植物镶嵌分布在生物土壤结皮斑块中,地表径流是资源转移(如水、营养物质和种子)和再分配的关键途径[28,69].灌木冠层间的生物土壤结皮盖度较高,通常有利于径流的生成,且生物土壤结皮斑块通过向灌木斑块输送径流水源和营养元素,支持现存的灌木斑块[13,49,70-71].就草本而言,例如在尼纳扎的沙丘地中,沙丘间的斜坡上可能无法维持地面芽植物生存,但沙丘间洼地中的地面芽植物却得以生长,这得益于沙丘斜坡上生物土壤结皮产生的径流为洼地提供了水分[44],从而形成了生物土壤结皮-草本共存斑块.沙丘间斜坡上的径流(包括地下流动)是多年生植物密集带形成的主要原因[72].然而,考虑土壤水分水平方向的再分配时,生物土壤结皮也可以和木本植物形成斑块状的共存模式[68,73]. ...
Contribution of cryptogamic covers to the global cycles of carbon and nitrogen
2012
Biological soil crusts as a model system in ecology
2016
Patterns and controls on nitrogen cycling of biological soil crusts
1
2016
... 作为旱区特有生物组分[7],生物土壤结皮(由土壤颗粒与不同比例的光合自养生物(蓝藻细菌、藻类、地衣、苔藓植物)和异养生物(细菌、真菌、古细菌)胶结而成的表面土壤复合体[8-9])约覆盖全球旱区面积的30%[10].生物土壤结皮作为土壤与大气间的交互界面[11],在调控生态水文过程(如水分蒸发、径流入渗)和土壤生物地球化学循环(包括水循环、养分循环等)方面发挥着重要的生态系统功能和服务[12-16].生物土壤结皮固氮占全球陆地生物固氮量的40%~85%[10].因此,生物土壤结皮被称为旱区生态系统工程师[17],将其纳入研究框架对理解和管理旱区生态系统十分关键[18]. ...
Spatial modeling of biological soil crusts to support rangeland assessment and monitoring
1
2006
... 作为旱区特有生物组分[7],生物土壤结皮(由土壤颗粒与不同比例的光合自养生物(蓝藻细菌、藻类、地衣、苔藓植物)和异养生物(细菌、真菌、古细菌)胶结而成的表面土壤复合体[8-9])约覆盖全球旱区面积的30%[10].生物土壤结皮作为土壤与大气间的交互界面[11],在调控生态水文过程(如水分蒸发、径流入渗)和土壤生物地球化学循环(包括水循环、养分循环等)方面发挥着重要的生态系统功能和服务[12-16].生物土壤结皮固氮占全球陆地生物固氮量的40%~85%[10].因此,生物土壤结皮被称为旱区生态系统工程师[17],将其纳入研究框架对理解和管理旱区生态系统十分关键[18]. ...
Biocrust as one of multiple stable states in global drylands
5
2020
... 作为旱区特有生物组分[7],生物土壤结皮(由土壤颗粒与不同比例的光合自养生物(蓝藻细菌、藻类、地衣、苔藓植物)和异养生物(细菌、真菌、古细菌)胶结而成的表面土壤复合体[8-9])约覆盖全球旱区面积的30%[10].生物土壤结皮作为土壤与大气间的交互界面[11],在调控生态水文过程(如水分蒸发、径流入渗)和土壤生物地球化学循环(包括水循环、养分循环等)方面发挥着重要的生态系统功能和服务[12-16].生物土壤结皮固氮占全球陆地生物固氮量的40%~85%[10].因此,生物土壤结皮被称为旱区生态系统工程师[17],将其纳入研究框架对理解和管理旱区生态系统十分关键[18]. ...
... 生物土壤结皮能够与维管植物形成多种共存模式[27].在以色列内盖夫沙漠北部,多年生灌木主导的维管植物斑块中分布着生物土壤结皮,形成了生物土壤结皮-灌木混合态[28].在年降水量低的区域,低降水促进了生物土壤结皮的生长,但限制了维管植物的生长[29],进而促使生物土壤结皮成为主导态.另外,在相同的环境条件下,生物土壤结皮与维管植物的共存可以呈现多种稳定状态,如几乎无植被存在的裸土状态、维管植物主导状态或生物土壤结皮主导状态[18,30-31].Kinast等[31]发现了在不同风速下两种多稳态组合:低风速下的固定结皮沙丘和固定植被沙丘,以及高风速下的固定植被沙丘和移动沙丘.Chen等[18]报道了在低降水量条件下,除裸土状态外,生物土壤结皮也可能形成一种稳定状态,此时生物结皮盖度超过80%且维管植物盖度不超过10%.考虑二者复杂的相互作用,厘清生物土壤结皮与维管植物的共存模式尤为重要. ...
... [18]报道了在低降水量条件下,除裸土状态外,生物土壤结皮也可能形成一种稳定状态,此时生物结皮盖度超过80%且维管植物盖度不超过10%.考虑二者复杂的相互作用,厘清生物土壤结皮与维管植物的共存模式尤为重要. ...
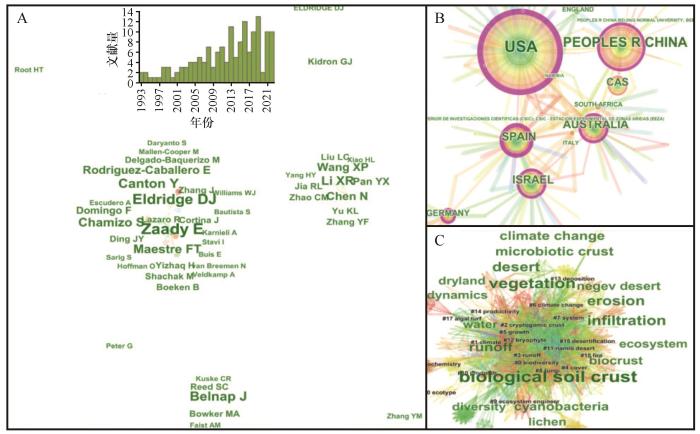
... 旱区生物土壤结皮-维管植物共存的生态水文机制研究,自1993年以来呈逐步上升的趋势(图1).数据库为Web of Science核心合集,检索式为:TS=[(“dryland*” OR “hyper*arid*” OR “arid*” OR “semi*arid*” OR “dry*subhumid*”) AND (“biological soil crust*” OR “biocrust*” OR “biogenic crust*” OR “biological crust*” OR “biotic crust*” OR “microphytic crust*” OR “microbiotic crust*” OR “Cryptogam*” OR “cyanobacteri*” OR “algal*” OR “lichen*” OR “moss*”) AND (“vascular plant” OR “grass” OR “woody” OR “herb*” OR “shrub” OR “bush” OR “turf”) AND (“ecohydrolog*” OR “runoff” OR “infiltration” OR “water flow” OR “ponding” OR “hydrolog*” OR “sorptivity” OR “moisture” OR “erosion” OR “sediment”)],共计检索文献156篇.生态学与水文学交叉融合,为深入探究旱区不同地被类型的存在提供比较全面且系统的阐释[48].目前,生物土壤结皮与维管植物共存的研究主要在生物土壤结皮与维管植被的相互作用[18]、生物土壤结皮与维管植物的生态水文效应[39,49]、生物土壤结皮与维管植物对全球变化的响应等方面[27].然而,对于生物土壤结皮-维管植物共存的生态水文机制研究,目前仍然不足.致力于该领域的研究者以David J.Eldridge、Eli Zaady、Giora J.Kidron、Jayne Belnap、Xinrong Li、Xinping Wang、Ning Chen等人为代表.这些研究主要在旱区分布较广的国家开展,如美国、中国、西班牙、澳大利亚、以色列等. ...
... 在理论方面,将生物土壤结皮与维管植物的共存现象与生态学理论结合,如系统平衡理论(system equilibrium behavior),有助于从理论层面深入理解其生态水文机制,进而更好地理解生物土壤结皮与维管植物的共存.系统平衡理论指出,在相同环境条件下,可存在两种或多种稳定状态,即不同的生物土壤结皮-维管植物共存模式,为探讨不同生态水文过程下旱区生物土壤结皮-维管植物共存提供全新的视角[18,87].因此,从理论层面推动旱区不同地被类型共存问题,是未来值得深入探讨的重要课题. ...
Biological soil crusts in a xeric Florida shrubland:composition,abundance,and spatial heterogeneity of crusts with different disturbance histories
1
2002
... 相较于生物土壤结皮,维管植物一直是旱区研究的核心.生物土壤结皮与维管植物彼此相互作用,又相互影响.二者的相互作用方式多样.生物土壤结皮直接影响维管植物的物种组成、功能性状和丰度,也通过间接作用重新分配降雨、固氮、调节土壤温度等[19-20].这些效应因生物土壤结皮类型、维管植物性状以及局部气候、环境和土壤条件的不同而有所变化[21].在奇瓦瓦沙漠,以蓝藻为主的生物土壤结皮会抑制维管植物幼苗的生长,而以地衣为主的生物土壤结皮则有助于其幼苗生长[22].反之,维管植物也会通过遮阴、产生凋落物以及形成微气候等方式影响生物土壤结皮,而这些影响又与维管植物的物种组成和丰度密切相关[21].在南非西南部卡拉哈里,不同类型的维管植物(如灌木和草本)因其截然不同的微环境改变了土壤表面组成(生物土壤结皮)[23].可见,生物土壤结皮和维管植物盖度之间既有负向作用[24-25],也有正向作用[26]. ...
The potential roles of biological soil crusts in dryland hydrologic cycles
2
2006
... 相较于生物土壤结皮,维管植物一直是旱区研究的核心.生物土壤结皮与维管植物彼此相互作用,又相互影响.二者的相互作用方式多样.生物土壤结皮直接影响维管植物的物种组成、功能性状和丰度,也通过间接作用重新分配降雨、固氮、调节土壤温度等[19-20].这些效应因生物土壤结皮类型、维管植物性状以及局部气候、环境和土壤条件的不同而有所变化[21].在奇瓦瓦沙漠,以蓝藻为主的生物土壤结皮会抑制维管植物幼苗的生长,而以地衣为主的生物土壤结皮则有助于其幼苗生长[22].反之,维管植物也会通过遮阴、产生凋落物以及形成微气候等方式影响生物土壤结皮,而这些影响又与维管植物的物种组成和丰度密切相关[21].在南非西南部卡拉哈里,不同类型的维管植物(如灌木和草本)因其截然不同的微环境改变了土壤表面组成(生物土壤结皮)[23].可见,生物土壤结皮和维管植物盖度之间既有负向作用[24-25],也有正向作用[26]. ...
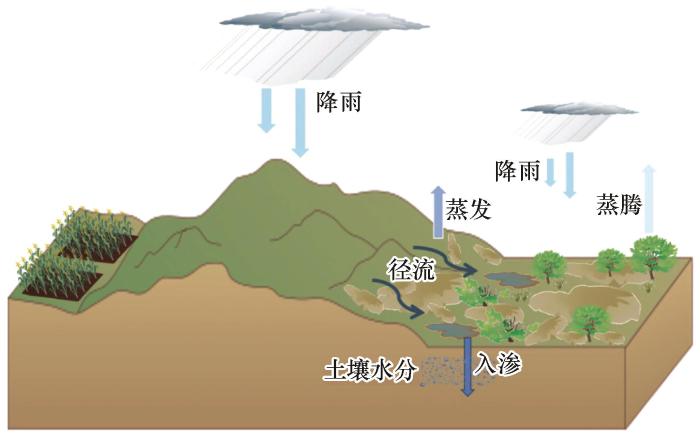
... 水分是旱区生态系统的关键限制因子,生态水文机制在解析生物土壤结皮与维管植物共存模式中可能发挥着核心作用[32].以降水和入渗过程为例,不同种类的生物土壤结皮和维管植物对水分的需求和适应能力不同[27,33],降水频率和强度的变化促使它们在物种组成和数量上做出相应调整,以适应新的水分环境[34-36].水分入渗的速率、深度以及范围等,会改变土壤中的水分分布状况[37],这关系到维管植物与生物土壤结皮在不同土层水分的获取,进而影响二者的生长和竞争关系[38-39].反之,生物土壤结皮与维管植物也并非被动地受水分影响,它们能够通过各自独特的方式影响生态水文过程[40].维管植物凭借自身较为发达的枝叶系统,能够直接截留降水,大大减少降雨对地面的直接冲击,进而减缓地表径流的形成速度[41-42].这不仅改变了水分在土壤表层的分配格局,还对水分的深层渗透和蓄存产生了连锁反应,显著影响了整个区域的水分动态平衡[43].生物土壤结皮可以通过调节径流、入渗和蒸发过程,为维管植物的生长与分布创造适宜的水分条件[44].生物土壤结皮在一定程度上减少了土壤水分的蒸发量,保持了土壤的湿度[20,45].这种对水分的良好调控能力,不仅为维管植物的生长提供了稳定的水分环境,而且通过影响水分的分布状况,进一步影响着不同种类维管植物的分布范围和生长态势[39].此外,生物土壤结皮还能够通过改变土壤水分状况,间接影响灌木的生长[46].土壤水分含量的变化会改变植物的根系生长方向和深度,进而影响植物对养分和水分的获取能力,最终影响灌木的生长状况和分布范围[47]. ...
Interactions of biological soil crusts with vascular plants
4
2016
... 相较于生物土壤结皮,维管植物一直是旱区研究的核心.生物土壤结皮与维管植物彼此相互作用,又相互影响.二者的相互作用方式多样.生物土壤结皮直接影响维管植物的物种组成、功能性状和丰度,也通过间接作用重新分配降雨、固氮、调节土壤温度等[19-20].这些效应因生物土壤结皮类型、维管植物性状以及局部气候、环境和土壤条件的不同而有所变化[21].在奇瓦瓦沙漠,以蓝藻为主的生物土壤结皮会抑制维管植物幼苗的生长,而以地衣为主的生物土壤结皮则有助于其幼苗生长[22].反之,维管植物也会通过遮阴、产生凋落物以及形成微气候等方式影响生物土壤结皮,而这些影响又与维管植物的物种组成和丰度密切相关[21].在南非西南部卡拉哈里,不同类型的维管植物(如灌木和草本)因其截然不同的微环境改变了土壤表面组成(生物土壤结皮)[23].可见,生物土壤结皮和维管植物盖度之间既有负向作用[24-25],也有正向作用[26]. ...
... [21].在南非西南部卡拉哈里,不同类型的维管植物(如灌木和草本)因其截然不同的微环境改变了土壤表面组成(生物土壤结皮)[23].可见,生物土壤结皮和维管植物盖度之间既有负向作用[24-25],也有正向作用[26]. ...
... 降水作为旱区生态系统的主要水源,深刻影响生物土壤结皮与维管植物的共存.以较低年降雨量90~150 mm为例,生物土壤结皮相较维管植物往往更具优势[31].主要因为维管植物对水分的需求量大,而生物土壤结皮的需求则小得多.随着降水频率的增加,生物土壤结皮适应的降水条件范围变宽.当日降水频率为0.025时,生物土壤结皮适宜在0~100 mm年降水量范围生长;当日降水频率为1时,生物土壤结皮适宜在0~500 mm年降水量区间生长[33],有力地促进了生物土壤结皮-维管植物共存.干旱事件(降水量显著降低)会降低降雨在土壤中的入渗深度,可能极大降低深根系维管植物的水分利用效率,但并不会对生物土壤结皮产生显著影响[13,46],呈现以生物土壤结皮为主的共存状态.随着降水的增加,生物土壤结皮与维管植物的可利用水分均有所增加,不过两者的竞争优势也随之变化,可能由于过多水分超出生物土壤结皮的可利用范围,增加的降水更多地被维管植物所利用[27].同时,维管植物的盖度过高会极大限制下层光照的有效性,进而导致生物土壤结皮的盖度下降[21].当超过某一临界点时,便会发生从生物土壤结皮主导状态向维管植物主导状态的转变[27].另外,由于生物土壤结皮的存在,降雨模式(降雨量或降雨时间分布)的改变会对灌木斑块和裸露斑块的土壤水分可用性动态产生不同的影响[56].总之,生物土壤结皮和维管植物之间的相互作用对降水的响应是极其复杂的,需考虑研究区域和降水模式的特点[21]. ...
... [21]. ...
Biocrusts and their disturbance mediate the recruitment of native and exotic grasses from a hot desert ecosystem
1
2018
... 相较于生物土壤结皮,维管植物一直是旱区研究的核心.生物土壤结皮与维管植物彼此相互作用,又相互影响.二者的相互作用方式多样.生物土壤结皮直接影响维管植物的物种组成、功能性状和丰度,也通过间接作用重新分配降雨、固氮、调节土壤温度等[19-20].这些效应因生物土壤结皮类型、维管植物性状以及局部气候、环境和土壤条件的不同而有所变化[21].在奇瓦瓦沙漠,以蓝藻为主的生物土壤结皮会抑制维管植物幼苗的生长,而以地衣为主的生物土壤结皮则有助于其幼苗生长[22].反之,维管植物也会通过遮阴、产生凋落物以及形成微气候等方式影响生物土壤结皮,而这些影响又与维管植物的物种组成和丰度密切相关[21].在南非西南部卡拉哈里,不同类型的维管植物(如灌木和草本)因其截然不同的微环境改变了土壤表面组成(生物土壤结皮)[23].可见,生物土壤结皮和维管植物盖度之间既有负向作用[24-25],也有正向作用[26]. ...
Effects of vegetation on bacterial communities,carbon and nitrogen in dryland soil surfaces:implications for shrub encroachment in the southwest Kalahari
1
2021
... 相较于生物土壤结皮,维管植物一直是旱区研究的核心.生物土壤结皮与维管植物彼此相互作用,又相互影响.二者的相互作用方式多样.生物土壤结皮直接影响维管植物的物种组成、功能性状和丰度,也通过间接作用重新分配降雨、固氮、调节土壤温度等[19-20].这些效应因生物土壤结皮类型、维管植物性状以及局部气候、环境和土壤条件的不同而有所变化[21].在奇瓦瓦沙漠,以蓝藻为主的生物土壤结皮会抑制维管植物幼苗的生长,而以地衣为主的生物土壤结皮则有助于其幼苗生长[22].反之,维管植物也会通过遮阴、产生凋落物以及形成微气候等方式影响生物土壤结皮,而这些影响又与维管植物的物种组成和丰度密切相关[21].在南非西南部卡拉哈里,不同类型的维管植物(如灌木和草本)因其截然不同的微环境改变了土壤表面组成(生物土壤结皮)[23].可见,生物土壤结皮和维管植物盖度之间既有负向作用[24-25],也有正向作用[26]. ...
Structure and function of microphytic soil crusts in wildland ecosystems of arid to semi-arid regions
1
1990
... 相较于生物土壤结皮,维管植物一直是旱区研究的核心.生物土壤结皮与维管植物彼此相互作用,又相互影响.二者的相互作用方式多样.生物土壤结皮直接影响维管植物的物种组成、功能性状和丰度,也通过间接作用重新分配降雨、固氮、调节土壤温度等[19-20].这些效应因生物土壤结皮类型、维管植物性状以及局部气候、环境和土壤条件的不同而有所变化[21].在奇瓦瓦沙漠,以蓝藻为主的生物土壤结皮会抑制维管植物幼苗的生长,而以地衣为主的生物土壤结皮则有助于其幼苗生长[22].反之,维管植物也会通过遮阴、产生凋落物以及形成微气候等方式影响生物土壤结皮,而这些影响又与维管植物的物种组成和丰度密切相关[21].在南非西南部卡拉哈里,不同类型的维管植物(如灌木和草本)因其截然不同的微环境改变了土壤表面组成(生物土壤结皮)[23].可见,生物土壤结皮和维管植物盖度之间既有负向作用[24-25],也有正向作用[26]. ...
Cryptogams,vascular plants,and soil hydrological relations-some preliminary-results from the semiarid woodlands of Eastern Australia
1
1993
... 相较于生物土壤结皮,维管植物一直是旱区研究的核心.生物土壤结皮与维管植物彼此相互作用,又相互影响.二者的相互作用方式多样.生物土壤结皮直接影响维管植物的物种组成、功能性状和丰度,也通过间接作用重新分配降雨、固氮、调节土壤温度等[19-20].这些效应因生物土壤结皮类型、维管植物性状以及局部气候、环境和土壤条件的不同而有所变化[21].在奇瓦瓦沙漠,以蓝藻为主的生物土壤结皮会抑制维管植物幼苗的生长,而以地衣为主的生物土壤结皮则有助于其幼苗生长[22].反之,维管植物也会通过遮阴、产生凋落物以及形成微气候等方式影响生物土壤结皮,而这些影响又与维管植物的物种组成和丰度密切相关[21].在南非西南部卡拉哈里,不同类型的维管植物(如灌木和草本)因其截然不同的微环境改变了土壤表面组成(生物土壤结皮)[23].可见,生物土壤结皮和维管植物盖度之间既有负向作用[24-25],也有正向作用[26]. ...
Non-random distribution of biocrust in a natural arid environment in the Northern Mexican Plateau
1
2018
... 相较于生物土壤结皮,维管植物一直是旱区研究的核心.生物土壤结皮与维管植物彼此相互作用,又相互影响.二者的相互作用方式多样.生物土壤结皮直接影响维管植物的物种组成、功能性状和丰度,也通过间接作用重新分配降雨、固氮、调节土壤温度等[19-20].这些效应因生物土壤结皮类型、维管植物性状以及局部气候、环境和土壤条件的不同而有所变化[21].在奇瓦瓦沙漠,以蓝藻为主的生物土壤结皮会抑制维管植物幼苗的生长,而以地衣为主的生物土壤结皮则有助于其幼苗生长[22].反之,维管植物也会通过遮阴、产生凋落物以及形成微气候等方式影响生物土壤结皮,而这些影响又与维管植物的物种组成和丰度密切相关[21].在南非西南部卡拉哈里,不同类型的维管植物(如灌木和草本)因其截然不同的微环境改变了土壤表面组成(生物土壤结皮)[23].可见,生物土壤结皮和维管植物盖度之间既有负向作用[24-25],也有正向作用[26]. ...
Precipitation and nitrogen deposition alter biocrust-vascular plant coexistence in a desert ecosystem:threshold and mechanisms
8
2022
... 生物土壤结皮能够与维管植物形成多种共存模式[27].在以色列内盖夫沙漠北部,多年生灌木主导的维管植物斑块中分布着生物土壤结皮,形成了生物土壤结皮-灌木混合态[28].在年降水量低的区域,低降水促进了生物土壤结皮的生长,但限制了维管植物的生长[29],进而促使生物土壤结皮成为主导态.另外,在相同的环境条件下,生物土壤结皮与维管植物的共存可以呈现多种稳定状态,如几乎无植被存在的裸土状态、维管植物主导状态或生物土壤结皮主导状态[18,30-31].Kinast等[31]发现了在不同风速下两种多稳态组合:低风速下的固定结皮沙丘和固定植被沙丘,以及高风速下的固定植被沙丘和移动沙丘.Chen等[18]报道了在低降水量条件下,除裸土状态外,生物土壤结皮也可能形成一种稳定状态,此时生物结皮盖度超过80%且维管植物盖度不超过10%.考虑二者复杂的相互作用,厘清生物土壤结皮与维管植物的共存模式尤为重要. ...
... 水分是旱区生态系统的关键限制因子,生态水文机制在解析生物土壤结皮与维管植物共存模式中可能发挥着核心作用[32].以降水和入渗过程为例,不同种类的生物土壤结皮和维管植物对水分的需求和适应能力不同[27,33],降水频率和强度的变化促使它们在物种组成和数量上做出相应调整,以适应新的水分环境[34-36].水分入渗的速率、深度以及范围等,会改变土壤中的水分分布状况[37],这关系到维管植物与生物土壤结皮在不同土层水分的获取,进而影响二者的生长和竞争关系[38-39].反之,生物土壤结皮与维管植物也并非被动地受水分影响,它们能够通过各自独特的方式影响生态水文过程[40].维管植物凭借自身较为发达的枝叶系统,能够直接截留降水,大大减少降雨对地面的直接冲击,进而减缓地表径流的形成速度[41-42].这不仅改变了水分在土壤表层的分配格局,还对水分的深层渗透和蓄存产生了连锁反应,显著影响了整个区域的水分动态平衡[43].生物土壤结皮可以通过调节径流、入渗和蒸发过程,为维管植物的生长与分布创造适宜的水分条件[44].生物土壤结皮在一定程度上减少了土壤水分的蒸发量,保持了土壤的湿度[20,45].这种对水分的良好调控能力,不仅为维管植物的生长提供了稳定的水分环境,而且通过影响水分的分布状况,进一步影响着不同种类维管植物的分布范围和生长态势[39].此外,生物土壤结皮还能够通过改变土壤水分状况,间接影响灌木的生长[46].土壤水分含量的变化会改变植物的根系生长方向和深度,进而影响植物对养分和水分的获取能力,最终影响灌木的生长状况和分布范围[47]. ...
... 旱区生物土壤结皮-维管植物共存的生态水文机制研究,自1993年以来呈逐步上升的趋势(图1).数据库为Web of Science核心合集,检索式为:TS=[(“dryland*” OR “hyper*arid*” OR “arid*” OR “semi*arid*” OR “dry*subhumid*”) AND (“biological soil crust*” OR “biocrust*” OR “biogenic crust*” OR “biological crust*” OR “biotic crust*” OR “microphytic crust*” OR “microbiotic crust*” OR “Cryptogam*” OR “cyanobacteri*” OR “algal*” OR “lichen*” OR “moss*”) AND (“vascular plant” OR “grass” OR “woody” OR “herb*” OR “shrub” OR “bush” OR “turf”) AND (“ecohydrolog*” OR “runoff” OR “infiltration” OR “water flow” OR “ponding” OR “hydrolog*” OR “sorptivity” OR “moisture” OR “erosion” OR “sediment”)],共计检索文献156篇.生态学与水文学交叉融合,为深入探究旱区不同地被类型的存在提供比较全面且系统的阐释[48].目前,生物土壤结皮与维管植物共存的研究主要在生物土壤结皮与维管植被的相互作用[18]、生物土壤结皮与维管植物的生态水文效应[39,49]、生物土壤结皮与维管植物对全球变化的响应等方面[27].然而,对于生物土壤结皮-维管植物共存的生态水文机制研究,目前仍然不足.致力于该领域的研究者以David J.Eldridge、Eli Zaady、Giora J.Kidron、Jayne Belnap、Xinrong Li、Xinping Wang、Ning Chen等人为代表.这些研究主要在旱区分布较广的国家开展,如美国、中国、西班牙、澳大利亚、以色列等. ...
... 1993年,于澳大利亚东部半干旱林地的控制实验表明,维管植物盖度与隐花植物盖度存在极为紧密的关联.随后,于2002年在以色列内盖夫沙漠北部发现以多年生灌木为主导的景观斑块被生物土壤结皮所隔开[28].2008年,Li等[50]发现在腾格里沙漠的灌木生态系统中,维管植物和生物土壤结皮斑块之间存在源-汇关系.借助长期监测数据和野外实验,通过改变土壤水分及水循环状况,确定腾格里沙漠生物土壤结皮的出现与发育是否会导致固沙植被的变化[51-52].2013年,生物土壤结皮的动态被纳入了模型,构建了一个维管植被和生物土壤结皮耦合模型,一定程度助力了该领域研究的发展[31].研究者在美国加州半干旱草原,利用2010—2014年的实验,探讨在春季仅浇水、施肥+浇水两种试验处理分别对藓类生物土壤结皮和维管植物的影响,试图验证藓类生物土壤结皮和矮维管植物的盖度和多样性会因降雨和养分(两类同时添加)的增加而减少,从而导致高维管植物盖度的增加这一假设[53].Xiao等[46]于2017年在黄土高原的一个半干旱流域布设18个采样点,以验证生物土壤结皮是否会降低土壤湿度,进而导致人工种植灌木退化,经分析得出人工灌木林土壤湿度变化的主导因素是高盖度藓类为主的生物土壤结皮.2018年,于以色列内盖夫沙漠的尼扎纳研究点发现,生物土壤结皮通过增加蒸发导致沙丘斜坡上大多数多年生植物死亡,但同时通过产生地表径流有利于沙丘间洼地浅根系多年生植物的生存[44].同年及次年,Chen等[37,54]构建了一个明确考虑三层土壤(生物土壤结皮层、浅层和深层)的生态水文模型,以此研究不同类型生物土壤结皮对旱区恢复动态的影响.2019年,Jia等[33]开发了一个生态水文模型,并将生物土壤结皮作为系统状态变量,探讨生物土壤结皮、草本及灌木对降雨变化的响应,这有力推动了对全球气候变化下旱区生态系统动态的理解,并有助于预防荒漠化.2022年,She等[27]于毛乌素沙地开展为期5年的降雨和氮添加试验,研究了全球变化如何影响生物土壤结皮-维管植物共存及其机制. ...
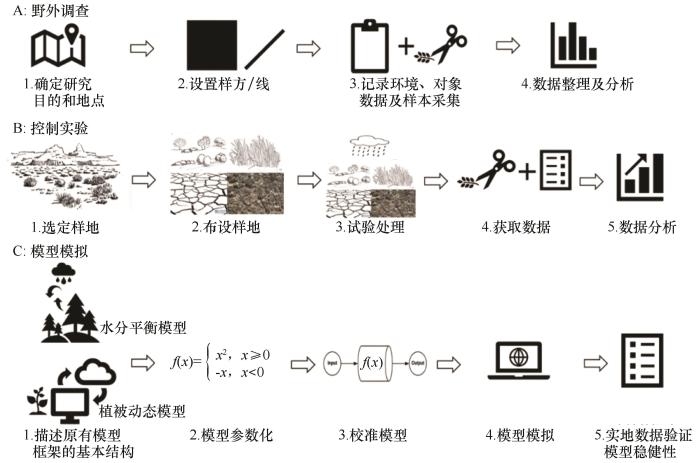
... 围绕生物土壤结皮-维管植物共存的生态水文机制,通过Web of Science数据库核心合集检索,共筛选出156篇文献.这些研究在方法运用上,主要采用了野外调查和控制实验两大类方法[27,49].其中,野外调查因其能直接获取自然状态下生物土壤结皮与维管植物共存的实际状况和相关数据,所以在研究中占据主导地位,约有90%的研究侧重于此方法.对不同环境下的生物土壤结皮和维管植物进行直接观测、采样与记录,以全面了解它们之间的相互关系.虽然少部分研究使用控制实验,但这种方法能够在人为设定的条件下,对特定因素进行精准控制和研究,有助于深入剖析某些关键因素对生物土壤结皮-维管植物共存生态水文机制的影响(图2). ...
... 降水作为旱区生态系统的主要水源,深刻影响生物土壤结皮与维管植物的共存.以较低年降雨量90~150 mm为例,生物土壤结皮相较维管植物往往更具优势[31].主要因为维管植物对水分的需求量大,而生物土壤结皮的需求则小得多.随着降水频率的增加,生物土壤结皮适应的降水条件范围变宽.当日降水频率为0.025时,生物土壤结皮适宜在0~100 mm年降水量范围生长;当日降水频率为1时,生物土壤结皮适宜在0~500 mm年降水量区间生长[33],有力地促进了生物土壤结皮-维管植物共存.干旱事件(降水量显著降低)会降低降雨在土壤中的入渗深度,可能极大降低深根系维管植物的水分利用效率,但并不会对生物土壤结皮产生显著影响[13,46],呈现以生物土壤结皮为主的共存状态.随着降水的增加,生物土壤结皮与维管植物的可利用水分均有所增加,不过两者的竞争优势也随之变化,可能由于过多水分超出生物土壤结皮的可利用范围,增加的降水更多地被维管植物所利用[27].同时,维管植物的盖度过高会极大限制下层光照的有效性,进而导致生物土壤结皮的盖度下降[21].当超过某一临界点时,便会发生从生物土壤结皮主导状态向维管植物主导状态的转变[27].另外,由于生物土壤结皮的存在,降雨模式(降雨量或降雨时间分布)的改变会对灌木斑块和裸露斑块的土壤水分可用性动态产生不同的影响[56].总之,生物土壤结皮和维管植物之间的相互作用对降水的响应是极其复杂的,需考虑研究区域和降水模式的特点[21]. ...
... [27].另外,由于生物土壤结皮的存在,降雨模式(降雨量或降雨时间分布)的改变会对灌木斑块和裸露斑块的土壤水分可用性动态产生不同的影响[56].总之,生物土壤结皮和维管植物之间的相互作用对降水的响应是极其复杂的,需考虑研究区域和降水模式的特点[21]. ...
... 生物土壤结皮-维管植物的共存现象在旱区普遍存在[27,49],这种共存关系对植被恢复及旱区生态系统功能和服务维持有着不可忽视的作用.自21世纪初,研究者们已开始关注两者共存的潜在机制,然而,从生态水文视角探讨其共存模式的研究仍显匮乏.现有研究多关注单一生态水文过程,而涉及多个生态水文过程的综合研究尚不多见.因此,未来研究应综合考虑多个生态水文过程,以全面解析旱区植被共存的潜在机制. ...
Microphytic crusts,shrub patches and water harvesting in the Negev Desert:the Shikim system
3
2002
... 生物土壤结皮能够与维管植物形成多种共存模式[27].在以色列内盖夫沙漠北部,多年生灌木主导的维管植物斑块中分布着生物土壤结皮,形成了生物土壤结皮-灌木混合态[28].在年降水量低的区域,低降水促进了生物土壤结皮的生长,但限制了维管植物的生长[29],进而促使生物土壤结皮成为主导态.另外,在相同的环境条件下,生物土壤结皮与维管植物的共存可以呈现多种稳定状态,如几乎无植被存在的裸土状态、维管植物主导状态或生物土壤结皮主导状态[18,30-31].Kinast等[31]发现了在不同风速下两种多稳态组合:低风速下的固定结皮沙丘和固定植被沙丘,以及高风速下的固定植被沙丘和移动沙丘.Chen等[18]报道了在低降水量条件下,除裸土状态外,生物土壤结皮也可能形成一种稳定状态,此时生物结皮盖度超过80%且维管植物盖度不超过10%.考虑二者复杂的相互作用,厘清生物土壤结皮与维管植物的共存模式尤为重要. ...
... 1993年,于澳大利亚东部半干旱林地的控制实验表明,维管植物盖度与隐花植物盖度存在极为紧密的关联.随后,于2002年在以色列内盖夫沙漠北部发现以多年生灌木为主导的景观斑块被生物土壤结皮所隔开[28].2008年,Li等[50]发现在腾格里沙漠的灌木生态系统中,维管植物和生物土壤结皮斑块之间存在源-汇关系.借助长期监测数据和野外实验,通过改变土壤水分及水循环状况,确定腾格里沙漠生物土壤结皮的出现与发育是否会导致固沙植被的变化[51-52].2013年,生物土壤结皮的动态被纳入了模型,构建了一个维管植被和生物土壤结皮耦合模型,一定程度助力了该领域研究的发展[31].研究者在美国加州半干旱草原,利用2010—2014年的实验,探讨在春季仅浇水、施肥+浇水两种试验处理分别对藓类生物土壤结皮和维管植物的影响,试图验证藓类生物土壤结皮和矮维管植物的盖度和多样性会因降雨和养分(两类同时添加)的增加而减少,从而导致高维管植物盖度的增加这一假设[53].Xiao等[46]于2017年在黄土高原的一个半干旱流域布设18个采样点,以验证生物土壤结皮是否会降低土壤湿度,进而导致人工种植灌木退化,经分析得出人工灌木林土壤湿度变化的主导因素是高盖度藓类为主的生物土壤结皮.2018年,于以色列内盖夫沙漠的尼扎纳研究点发现,生物土壤结皮通过增加蒸发导致沙丘斜坡上大多数多年生植物死亡,但同时通过产生地表径流有利于沙丘间洼地浅根系多年生植物的生存[44].同年及次年,Chen等[37,54]构建了一个明确考虑三层土壤(生物土壤结皮层、浅层和深层)的生态水文模型,以此研究不同类型生物土壤结皮对旱区恢复动态的影响.2019年,Jia等[33]开发了一个生态水文模型,并将生物土壤结皮作为系统状态变量,探讨生物土壤结皮、草本及灌木对降雨变化的响应,这有力推动了对全球气候变化下旱区生态系统动态的理解,并有助于预防荒漠化.2022年,She等[27]于毛乌素沙地开展为期5年的降雨和氮添加试验,研究了全球变化如何影响生物土壤结皮-维管植物共存及其机制. ...
... 在旱区生态系统,植物镶嵌分布在生物土壤结皮斑块中,地表径流是资源转移(如水、营养物质和种子)和再分配的关键途径[28,69].灌木冠层间的生物土壤结皮盖度较高,通常有利于径流的生成,且生物土壤结皮斑块通过向灌木斑块输送径流水源和营养元素,支持现存的灌木斑块[13,49,70-71].就草本而言,例如在尼纳扎的沙丘地中,沙丘间的斜坡上可能无法维持地面芽植物生存,但沙丘间洼地中的地面芽植物却得以生长,这得益于沙丘斜坡上生物土壤结皮产生的径流为洼地提供了水分[44],从而形成了生物土壤结皮-草本共存斑块.沙丘间斜坡上的径流(包括地下流动)是多年生植物密集带形成的主要原因[72].然而,考虑土壤水分水平方向的再分配时,生物土壤结皮也可以和木本植物形成斑块状的共存模式[68,73]. ...
Effects of biocrust on soil erosion and organic carbon losses under natural rainfall
1
2017
... 生物土壤结皮能够与维管植物形成多种共存模式[27].在以色列内盖夫沙漠北部,多年生灌木主导的维管植物斑块中分布着生物土壤结皮,形成了生物土壤结皮-灌木混合态[28].在年降水量低的区域,低降水促进了生物土壤结皮的生长,但限制了维管植物的生长[29],进而促使生物土壤结皮成为主导态.另外,在相同的环境条件下,生物土壤结皮与维管植物的共存可以呈现多种稳定状态,如几乎无植被存在的裸土状态、维管植物主导状态或生物土壤结皮主导状态[18,30-31].Kinast等[31]发现了在不同风速下两种多稳态组合:低风速下的固定结皮沙丘和固定植被沙丘,以及高风速下的固定植被沙丘和移动沙丘.Chen等[18]报道了在低降水量条件下,除裸土状态外,生物土壤结皮也可能形成一种稳定状态,此时生物结皮盖度超过80%且维管植物盖度不超过10%.考虑二者复杂的相互作用,厘清生物土壤结皮与维管植物的共存模式尤为重要. ...
Hydrologic variability in dryland regions:impacts on ecosystem dynamics and food security
1
2012
... 生物土壤结皮能够与维管植物形成多种共存模式[27].在以色列内盖夫沙漠北部,多年生灌木主导的维管植物斑块中分布着生物土壤结皮,形成了生物土壤结皮-灌木混合态[28].在年降水量低的区域,低降水促进了生物土壤结皮的生长,但限制了维管植物的生长[29],进而促使生物土壤结皮成为主导态.另外,在相同的环境条件下,生物土壤结皮与维管植物的共存可以呈现多种稳定状态,如几乎无植被存在的裸土状态、维管植物主导状态或生物土壤结皮主导状态[18,30-31].Kinast等[31]发现了在不同风速下两种多稳态组合:低风速下的固定结皮沙丘和固定植被沙丘,以及高风速下的固定植被沙丘和移动沙丘.Chen等[18]报道了在低降水量条件下,除裸土状态外,生物土壤结皮也可能形成一种稳定状态,此时生物结皮盖度超过80%且维管植物盖度不超过10%.考虑二者复杂的相互作用,厘清生物土壤结皮与维管植物的共存模式尤为重要. ...
Biogenic crust dynamics on sand dunes
4
2013
... 生物土壤结皮能够与维管植物形成多种共存模式[27].在以色列内盖夫沙漠北部,多年生灌木主导的维管植物斑块中分布着生物土壤结皮,形成了生物土壤结皮-灌木混合态[28].在年降水量低的区域,低降水促进了生物土壤结皮的生长,但限制了维管植物的生长[29],进而促使生物土壤结皮成为主导态.另外,在相同的环境条件下,生物土壤结皮与维管植物的共存可以呈现多种稳定状态,如几乎无植被存在的裸土状态、维管植物主导状态或生物土壤结皮主导状态[18,30-31].Kinast等[31]发现了在不同风速下两种多稳态组合:低风速下的固定结皮沙丘和固定植被沙丘,以及高风速下的固定植被沙丘和移动沙丘.Chen等[18]报道了在低降水量条件下,除裸土状态外,生物土壤结皮也可能形成一种稳定状态,此时生物结皮盖度超过80%且维管植物盖度不超过10%.考虑二者复杂的相互作用,厘清生物土壤结皮与维管植物的共存模式尤为重要. ...
... [31]发现了在不同风速下两种多稳态组合:低风速下的固定结皮沙丘和固定植被沙丘,以及高风速下的固定植被沙丘和移动沙丘.Chen等[18]报道了在低降水量条件下,除裸土状态外,生物土壤结皮也可能形成一种稳定状态,此时生物结皮盖度超过80%且维管植物盖度不超过10%.考虑二者复杂的相互作用,厘清生物土壤结皮与维管植物的共存模式尤为重要. ...
... 1993年,于澳大利亚东部半干旱林地的控制实验表明,维管植物盖度与隐花植物盖度存在极为紧密的关联.随后,于2002年在以色列内盖夫沙漠北部发现以多年生灌木为主导的景观斑块被生物土壤结皮所隔开[28].2008年,Li等[50]发现在腾格里沙漠的灌木生态系统中,维管植物和生物土壤结皮斑块之间存在源-汇关系.借助长期监测数据和野外实验,通过改变土壤水分及水循环状况,确定腾格里沙漠生物土壤结皮的出现与发育是否会导致固沙植被的变化[51-52].2013年,生物土壤结皮的动态被纳入了模型,构建了一个维管植被和生物土壤结皮耦合模型,一定程度助力了该领域研究的发展[31].研究者在美国加州半干旱草原,利用2010—2014年的实验,探讨在春季仅浇水、施肥+浇水两种试验处理分别对藓类生物土壤结皮和维管植物的影响,试图验证藓类生物土壤结皮和矮维管植物的盖度和多样性会因降雨和养分(两类同时添加)的增加而减少,从而导致高维管植物盖度的增加这一假设[53].Xiao等[46]于2017年在黄土高原的一个半干旱流域布设18个采样点,以验证生物土壤结皮是否会降低土壤湿度,进而导致人工种植灌木退化,经分析得出人工灌木林土壤湿度变化的主导因素是高盖度藓类为主的生物土壤结皮.2018年,于以色列内盖夫沙漠的尼扎纳研究点发现,生物土壤结皮通过增加蒸发导致沙丘斜坡上大多数多年生植物死亡,但同时通过产生地表径流有利于沙丘间洼地浅根系多年生植物的生存[44].同年及次年,Chen等[37,54]构建了一个明确考虑三层土壤(生物土壤结皮层、浅层和深层)的生态水文模型,以此研究不同类型生物土壤结皮对旱区恢复动态的影响.2019年,Jia等[33]开发了一个生态水文模型,并将生物土壤结皮作为系统状态变量,探讨生物土壤结皮、草本及灌木对降雨变化的响应,这有力推动了对全球气候变化下旱区生态系统动态的理解,并有助于预防荒漠化.2022年,She等[27]于毛乌素沙地开展为期5年的降雨和氮添加试验,研究了全球变化如何影响生物土壤结皮-维管植物共存及其机制. ...
... 降水作为旱区生态系统的主要水源,深刻影响生物土壤结皮与维管植物的共存.以较低年降雨量90~150 mm为例,生物土壤结皮相较维管植物往往更具优势[31].主要因为维管植物对水分的需求量大,而生物土壤结皮的需求则小得多.随着降水频率的增加,生物土壤结皮适应的降水条件范围变宽.当日降水频率为0.025时,生物土壤结皮适宜在0~100 mm年降水量范围生长;当日降水频率为1时,生物土壤结皮适宜在0~500 mm年降水量区间生长[33],有力地促进了生物土壤结皮-维管植物共存.干旱事件(降水量显著降低)会降低降雨在土壤中的入渗深度,可能极大降低深根系维管植物的水分利用效率,但并不会对生物土壤结皮产生显著影响[13,46],呈现以生物土壤结皮为主的共存状态.随着降水的增加,生物土壤结皮与维管植物的可利用水分均有所增加,不过两者的竞争优势也随之变化,可能由于过多水分超出生物土壤结皮的可利用范围,增加的降水更多地被维管植物所利用[27].同时,维管植物的盖度过高会极大限制下层光照的有效性,进而导致生物土壤结皮的盖度下降[21].当超过某一临界点时,便会发生从生物土壤结皮主导状态向维管植物主导状态的转变[27].另外,由于生物土壤结皮的存在,降雨模式(降雨量或降雨时间分布)的改变会对灌木斑块和裸露斑块的土壤水分可用性动态产生不同的影响[56].总之,生物土壤结皮和维管植物之间的相互作用对降水的响应是极其复杂的,需考虑研究区域和降水模式的特点[21]. ...
Biocrust-induced partitioning of soil water between grass and shrub in a desert steppe of Northwest China
2
2023
... 水分是旱区生态系统的关键限制因子,生态水文机制在解析生物土壤结皮与维管植物共存模式中可能发挥着核心作用[32].以降水和入渗过程为例,不同种类的生物土壤结皮和维管植物对水分的需求和适应能力不同[27,33],降水频率和强度的变化促使它们在物种组成和数量上做出相应调整,以适应新的水分环境[34-36].水分入渗的速率、深度以及范围等,会改变土壤中的水分分布状况[37],这关系到维管植物与生物土壤结皮在不同土层水分的获取,进而影响二者的生长和竞争关系[38-39].反之,生物土壤结皮与维管植物也并非被动地受水分影响,它们能够通过各自独特的方式影响生态水文过程[40].维管植物凭借自身较为发达的枝叶系统,能够直接截留降水,大大减少降雨对地面的直接冲击,进而减缓地表径流的形成速度[41-42].这不仅改变了水分在土壤表层的分配格局,还对水分的深层渗透和蓄存产生了连锁反应,显著影响了整个区域的水分动态平衡[43].生物土壤结皮可以通过调节径流、入渗和蒸发过程,为维管植物的生长与分布创造适宜的水分条件[44].生物土壤结皮在一定程度上减少了土壤水分的蒸发量,保持了土壤的湿度[20,45].这种对水分的良好调控能力,不仅为维管植物的生长提供了稳定的水分环境,而且通过影响水分的分布状况,进一步影响着不同种类维管植物的分布范围和生长态势[39].此外,生物土壤结皮还能够通过改变土壤水分状况,间接影响灌木的生长[46].土壤水分含量的变化会改变植物的根系生长方向和深度,进而影响植物对养分和水分的获取能力,最终影响灌木的生长状况和分布范围[47]. ...
... 在研究对象方面,现有旱区植被共存模式的研究相对单一,主要在生物土壤结皮-草本共存或生物土壤结皮-木本植物(灌木)的共存上,而较少涉及生物土壤结皮-草本-灌木植物共存,全生物要素(乔木-灌木-草本-生物土壤结皮)共存问题的研究更少.Yang等[32]发现生物土壤结皮-灌木-草本共存模式在浅层土壤水分上存在竞争关系,打破了传统上旱区草本-灌木共存的土壤水分利用分层现象.因此,从全生物要素角度探讨共存问题是未来研究的重要方向.而首要任务是明确生物土壤结皮-维管植物的共存模式,进而系统性地探究水文过程如何影响并塑造这些共存模式. ...
High rainfall frequency promotes the dominance of biocrust under low annual rainfall
4
2019
... 水分是旱区生态系统的关键限制因子,生态水文机制在解析生物土壤结皮与维管植物共存模式中可能发挥着核心作用[32].以降水和入渗过程为例,不同种类的生物土壤结皮和维管植物对水分的需求和适应能力不同[27,33],降水频率和强度的变化促使它们在物种组成和数量上做出相应调整,以适应新的水分环境[34-36].水分入渗的速率、深度以及范围等,会改变土壤中的水分分布状况[37],这关系到维管植物与生物土壤结皮在不同土层水分的获取,进而影响二者的生长和竞争关系[38-39].反之,生物土壤结皮与维管植物也并非被动地受水分影响,它们能够通过各自独特的方式影响生态水文过程[40].维管植物凭借自身较为发达的枝叶系统,能够直接截留降水,大大减少降雨对地面的直接冲击,进而减缓地表径流的形成速度[41-42].这不仅改变了水分在土壤表层的分配格局,还对水分的深层渗透和蓄存产生了连锁反应,显著影响了整个区域的水分动态平衡[43].生物土壤结皮可以通过调节径流、入渗和蒸发过程,为维管植物的生长与分布创造适宜的水分条件[44].生物土壤结皮在一定程度上减少了土壤水分的蒸发量,保持了土壤的湿度[20,45].这种对水分的良好调控能力,不仅为维管植物的生长提供了稳定的水分环境,而且通过影响水分的分布状况,进一步影响着不同种类维管植物的分布范围和生长态势[39].此外,生物土壤结皮还能够通过改变土壤水分状况,间接影响灌木的生长[46].土壤水分含量的变化会改变植物的根系生长方向和深度,进而影响植物对养分和水分的获取能力,最终影响灌木的生长状况和分布范围[47]. ...
... 1993年,于澳大利亚东部半干旱林地的控制实验表明,维管植物盖度与隐花植物盖度存在极为紧密的关联.随后,于2002年在以色列内盖夫沙漠北部发现以多年生灌木为主导的景观斑块被生物土壤结皮所隔开[28].2008年,Li等[50]发现在腾格里沙漠的灌木生态系统中,维管植物和生物土壤结皮斑块之间存在源-汇关系.借助长期监测数据和野外实验,通过改变土壤水分及水循环状况,确定腾格里沙漠生物土壤结皮的出现与发育是否会导致固沙植被的变化[51-52].2013年,生物土壤结皮的动态被纳入了模型,构建了一个维管植被和生物土壤结皮耦合模型,一定程度助力了该领域研究的发展[31].研究者在美国加州半干旱草原,利用2010—2014年的实验,探讨在春季仅浇水、施肥+浇水两种试验处理分别对藓类生物土壤结皮和维管植物的影响,试图验证藓类生物土壤结皮和矮维管植物的盖度和多样性会因降雨和养分(两类同时添加)的增加而减少,从而导致高维管植物盖度的增加这一假设[53].Xiao等[46]于2017年在黄土高原的一个半干旱流域布设18个采样点,以验证生物土壤结皮是否会降低土壤湿度,进而导致人工种植灌木退化,经分析得出人工灌木林土壤湿度变化的主导因素是高盖度藓类为主的生物土壤结皮.2018年,于以色列内盖夫沙漠的尼扎纳研究点发现,生物土壤结皮通过增加蒸发导致沙丘斜坡上大多数多年生植物死亡,但同时通过产生地表径流有利于沙丘间洼地浅根系多年生植物的生存[44].同年及次年,Chen等[37,54]构建了一个明确考虑三层土壤(生物土壤结皮层、浅层和深层)的生态水文模型,以此研究不同类型生物土壤结皮对旱区恢复动态的影响.2019年,Jia等[33]开发了一个生态水文模型,并将生物土壤结皮作为系统状态变量,探讨生物土壤结皮、草本及灌木对降雨变化的响应,这有力推动了对全球气候变化下旱区生态系统动态的理解,并有助于预防荒漠化.2022年,She等[27]于毛乌素沙地开展为期5年的降雨和氮添加试验,研究了全球变化如何影响生物土壤结皮-维管植物共存及其机制. ...
... 近年来,随着科技的飞速发展和数据的日益丰富,模型模拟凭借其独特的优势,在该领域发挥着越来越重要的作用.模型模拟具有灵活性,能够根据不同的研究需求和假设进行灵活调整和优化;具备可操控性,可以人为设定和改变各种输入参数,以探究不同条件下生态水文机制的变化;易获取性高,研究人员可以利用已有的数据和软件工具快速构建模型.特别是生态水文模型,它能够综合考虑多种生态水文过程,模拟生物土壤结皮与维管植物在不同条件下的生长、水分运移等过程,有望在未来研究中占据主导地位[33,37,55].构建模型涉及到对生态水文系统的结构、功能和各个组成部分之间关系的抽象和数学描述;接着进行参数化与模型校准,即根据实际数据和研究成果确定模型中各个参数的值,调整模型,使输出结果与实际观测数据尽可能吻合;最后按照设定的条件和参数运行模型并对模拟结果进行验证,通过与更多实际观测数据对比,评估模型的准确性和可靠性. ...
... 降水作为旱区生态系统的主要水源,深刻影响生物土壤结皮与维管植物的共存.以较低年降雨量90~150 mm为例,生物土壤结皮相较维管植物往往更具优势[31].主要因为维管植物对水分的需求量大,而生物土壤结皮的需求则小得多.随着降水频率的增加,生物土壤结皮适应的降水条件范围变宽.当日降水频率为0.025时,生物土壤结皮适宜在0~100 mm年降水量范围生长;当日降水频率为1时,生物土壤结皮适宜在0~500 mm年降水量区间生长[33],有力地促进了生物土壤结皮-维管植物共存.干旱事件(降水量显著降低)会降低降雨在土壤中的入渗深度,可能极大降低深根系维管植物的水分利用效率,但并不会对生物土壤结皮产生显著影响[13,46],呈现以生物土壤结皮为主的共存状态.随着降水的增加,生物土壤结皮与维管植物的可利用水分均有所增加,不过两者的竞争优势也随之变化,可能由于过多水分超出生物土壤结皮的可利用范围,增加的降水更多地被维管植物所利用[27].同时,维管植物的盖度过高会极大限制下层光照的有效性,进而导致生物土壤结皮的盖度下降[21].当超过某一临界点时,便会发生从生物土壤结皮主导状态向维管植物主导状态的转变[27].另外,由于生物土壤结皮的存在,降雨模式(降雨量或降雨时间分布)的改变会对灌木斑块和裸露斑块的土壤水分可用性动态产生不同的影响[56].总之,生物土壤结皮和维管植物之间的相互作用对降水的响应是极其复杂的,需考虑研究区域和降水模式的特点[21]. ...
Biological soil crusts (biocrusts) as a model system in community,landscape and ecosystem ecology
1
2014
... 水分是旱区生态系统的关键限制因子,生态水文机制在解析生物土壤结皮与维管植物共存模式中可能发挥着核心作用[32].以降水和入渗过程为例,不同种类的生物土壤结皮和维管植物对水分的需求和适应能力不同[27,33],降水频率和强度的变化促使它们在物种组成和数量上做出相应调整,以适应新的水分环境[34-36].水分入渗的速率、深度以及范围等,会改变土壤中的水分分布状况[37],这关系到维管植物与生物土壤结皮在不同土层水分的获取,进而影响二者的生长和竞争关系[38-39].反之,生物土壤结皮与维管植物也并非被动地受水分影响,它们能够通过各自独特的方式影响生态水文过程[40].维管植物凭借自身较为发达的枝叶系统,能够直接截留降水,大大减少降雨对地面的直接冲击,进而减缓地表径流的形成速度[41-42].这不仅改变了水分在土壤表层的分配格局,还对水分的深层渗透和蓄存产生了连锁反应,显著影响了整个区域的水分动态平衡[43].生物土壤结皮可以通过调节径流、入渗和蒸发过程,为维管植物的生长与分布创造适宜的水分条件[44].生物土壤结皮在一定程度上减少了土壤水分的蒸发量,保持了土壤的湿度[20,45].这种对水分的良好调控能力,不仅为维管植物的生长提供了稳定的水分环境,而且通过影响水分的分布状况,进一步影响着不同种类维管植物的分布范围和生长态势[39].此外,生物土壤结皮还能够通过改变土壤水分状况,间接影响灌木的生长[46].土壤水分含量的变化会改变植物的根系生长方向和深度,进而影响植物对养分和水分的获取能力,最终影响灌木的生长状况和分布范围[47]. ...
Primary production of the central grassland region of the united-states
1988
Changes to dryland rainfall result in rapid moss mortality and altered soil fertility
1
2012
... 水分是旱区生态系统的关键限制因子,生态水文机制在解析生物土壤结皮与维管植物共存模式中可能发挥着核心作用[32].以降水和入渗过程为例,不同种类的生物土壤结皮和维管植物对水分的需求和适应能力不同[27,33],降水频率和强度的变化促使它们在物种组成和数量上做出相应调整,以适应新的水分环境[34-36].水分入渗的速率、深度以及范围等,会改变土壤中的水分分布状况[37],这关系到维管植物与生物土壤结皮在不同土层水分的获取,进而影响二者的生长和竞争关系[38-39].反之,生物土壤结皮与维管植物也并非被动地受水分影响,它们能够通过各自独特的方式影响生态水文过程[40].维管植物凭借自身较为发达的枝叶系统,能够直接截留降水,大大减少降雨对地面的直接冲击,进而减缓地表径流的形成速度[41-42].这不仅改变了水分在土壤表层的分配格局,还对水分的深层渗透和蓄存产生了连锁反应,显著影响了整个区域的水分动态平衡[43].生物土壤结皮可以通过调节径流、入渗和蒸发过程,为维管植物的生长与分布创造适宜的水分条件[44].生物土壤结皮在一定程度上减少了土壤水分的蒸发量,保持了土壤的湿度[20,45].这种对水分的良好调控能力,不仅为维管植物的生长提供了稳定的水分环境,而且通过影响水分的分布状况,进一步影响着不同种类维管植物的分布范围和生长态势[39].此外,生物土壤结皮还能够通过改变土壤水分状况,间接影响灌木的生长[46].土壤水分含量的变化会改变植物的根系生长方向和深度,进而影响植物对养分和水分的获取能力,最终影响灌木的生长状况和分布范围[47]. ...
Ecohydrological effects of biological soil crust on the vegetation dynamics of restoration in a dryland ecosystem
5
2018
... 水分是旱区生态系统的关键限制因子,生态水文机制在解析生物土壤结皮与维管植物共存模式中可能发挥着核心作用[32].以降水和入渗过程为例,不同种类的生物土壤结皮和维管植物对水分的需求和适应能力不同[27,33],降水频率和强度的变化促使它们在物种组成和数量上做出相应调整,以适应新的水分环境[34-36].水分入渗的速率、深度以及范围等,会改变土壤中的水分分布状况[37],这关系到维管植物与生物土壤结皮在不同土层水分的获取,进而影响二者的生长和竞争关系[38-39].反之,生物土壤结皮与维管植物也并非被动地受水分影响,它们能够通过各自独特的方式影响生态水文过程[40].维管植物凭借自身较为发达的枝叶系统,能够直接截留降水,大大减少降雨对地面的直接冲击,进而减缓地表径流的形成速度[41-42].这不仅改变了水分在土壤表层的分配格局,还对水分的深层渗透和蓄存产生了连锁反应,显著影响了整个区域的水分动态平衡[43].生物土壤结皮可以通过调节径流、入渗和蒸发过程,为维管植物的生长与分布创造适宜的水分条件[44].生物土壤结皮在一定程度上减少了土壤水分的蒸发量,保持了土壤的湿度[20,45].这种对水分的良好调控能力,不仅为维管植物的生长提供了稳定的水分环境,而且通过影响水分的分布状况,进一步影响着不同种类维管植物的分布范围和生长态势[39].此外,生物土壤结皮还能够通过改变土壤水分状况,间接影响灌木的生长[46].土壤水分含量的变化会改变植物的根系生长方向和深度,进而影响植物对养分和水分的获取能力,最终影响灌木的生长状况和分布范围[47]. ...
... 1993年,于澳大利亚东部半干旱林地的控制实验表明,维管植物盖度与隐花植物盖度存在极为紧密的关联.随后,于2002年在以色列内盖夫沙漠北部发现以多年生灌木为主导的景观斑块被生物土壤结皮所隔开[28].2008年,Li等[50]发现在腾格里沙漠的灌木生态系统中,维管植物和生物土壤结皮斑块之间存在源-汇关系.借助长期监测数据和野外实验,通过改变土壤水分及水循环状况,确定腾格里沙漠生物土壤结皮的出现与发育是否会导致固沙植被的变化[51-52].2013年,生物土壤结皮的动态被纳入了模型,构建了一个维管植被和生物土壤结皮耦合模型,一定程度助力了该领域研究的发展[31].研究者在美国加州半干旱草原,利用2010—2014年的实验,探讨在春季仅浇水、施肥+浇水两种试验处理分别对藓类生物土壤结皮和维管植物的影响,试图验证藓类生物土壤结皮和矮维管植物的盖度和多样性会因降雨和养分(两类同时添加)的增加而减少,从而导致高维管植物盖度的增加这一假设[53].Xiao等[46]于2017年在黄土高原的一个半干旱流域布设18个采样点,以验证生物土壤结皮是否会降低土壤湿度,进而导致人工种植灌木退化,经分析得出人工灌木林土壤湿度变化的主导因素是高盖度藓类为主的生物土壤结皮.2018年,于以色列内盖夫沙漠的尼扎纳研究点发现,生物土壤结皮通过增加蒸发导致沙丘斜坡上大多数多年生植物死亡,但同时通过产生地表径流有利于沙丘间洼地浅根系多年生植物的生存[44].同年及次年,Chen等[37,54]构建了一个明确考虑三层土壤(生物土壤结皮层、浅层和深层)的生态水文模型,以此研究不同类型生物土壤结皮对旱区恢复动态的影响.2019年,Jia等[33]开发了一个生态水文模型,并将生物土壤结皮作为系统状态变量,探讨生物土壤结皮、草本及灌木对降雨变化的响应,这有力推动了对全球气候变化下旱区生态系统动态的理解,并有助于预防荒漠化.2022年,She等[27]于毛乌素沙地开展为期5年的降雨和氮添加试验,研究了全球变化如何影响生物土壤结皮-维管植物共存及其机制. ...
... 近年来,随着科技的飞速发展和数据的日益丰富,模型模拟凭借其独特的优势,在该领域发挥着越来越重要的作用.模型模拟具有灵活性,能够根据不同的研究需求和假设进行灵活调整和优化;具备可操控性,可以人为设定和改变各种输入参数,以探究不同条件下生态水文机制的变化;易获取性高,研究人员可以利用已有的数据和软件工具快速构建模型.特别是生态水文模型,它能够综合考虑多种生态水文过程,模拟生物土壤结皮与维管植物在不同条件下的生长、水分运移等过程,有望在未来研究中占据主导地位[33,37,55].构建模型涉及到对生态水文系统的结构、功能和各个组成部分之间关系的抽象和数学描述;接着进行参数化与模型校准,即根据实际数据和研究成果确定模型中各个参数的值,调整模型,使输出结果与实际观测数据尽可能吻合;最后按照设定的条件和参数运行模型并对模拟结果进行验证,通过与更多实际观测数据对比,评估模型的准确性和可靠性. ...
... 降雨后的土壤入渗是一个重要的生态水文过程,是土壤水分补给的主要途径[57].藓类生物土壤结皮的出现限制水分向深层土壤的入渗,会降低深根系植物的可利用水分,最终致使植物群落由深根系旱生灌木向浅根系的草本植物群落转变[52,58-62].在风沙严重的旱区,风沙沉积作用致使水分渗透减少,进而导致深层土壤干燥,生物土壤结皮广泛取代旱生灌木[63].此外,不同类型的生物土壤结皮对降雨土壤入渗的影响存在明显差异,因此,生物土壤结皮的演替(从藻类到地衣和藓类)会使浅层土壤降水入渗比例增加,从而提高草本植物的盖度,使旱区生态系统从裸土状态转变为草本植物主导的荒漠草原状态,实现恢复[37]. ...
... 蒸散作用(蒸发与蒸腾)对于土壤-植物-大气连续体(Soil-Plant-Atmosphere Continuum,SPAC)至关重要[74].地球表面约60%的降水通过蒸散作用返回大气,而在旱区这一比例可高达90%[75].蒸发又作为土壤与大气之间水分和能量平衡的核心要素,在旱区生态水文过程发挥重要的作用[76].然而,降雨强度对蒸发过程具有显著影响.在低强度降雨时,生物土壤结皮通过抑制蒸发保持土壤水分,而在高强度降雨时,则转变为促进蒸发,可能导致土壤水分的损失[59,77].以腾格里沙漠东南缘沙坡头地区为例,约4/5的降雨事件低于5 mm,生物土壤结皮通过抑制或延缓蒸发而保存土壤水分,这为微生物和草本植物的生长提供了有利条件,进而推动植被演替[78],最终形成草本植物主导状态.在极端干旱年份(年降雨量约30 mm),生物土壤结皮较大的蒸发量是一年生植物萌发与生长的主要限制,而未覆盖生物土壤结皮的沙地受此影响较小[79].不同的是,草本植物下方的生物土壤结皮降低了深层土壤水分含量,可能通过减少草根可获得的水分来限制草本植物的蒸腾作用,即生物土壤结皮通过增加土壤蒸发和减少草本植物蒸腾作用加剧了草地生态系统蒸散[80].当植物蒸腾作用所利用水分超过土壤蒸发作用时,存在生物土壤结皮的生态系统最终转变为草本植物主导状态[37],呈现出高盖度草本植物-低盖度生物土壤结皮共存模式.此外,其他生态水文过程的综合影响亦不容忽视.蒸发增加导致沙丘间多数多年生植物死亡,而生物土壤结皮通过其在径流产生中的作用,有助于沙丘间洼地浅根系多年生植物的生长和存活[44],从而促进生物土壤结皮与多年生植物的共存.蒸发与入渗的共同作用对土壤水分的改变也会影响维管植物的生长[62],进而对生物土壤结皮-维管植物的共存产生影响. ...
Refining the stress-gradient hypothesis for competition and facilitation in plant communities
1
2009
... 水分是旱区生态系统的关键限制因子,生态水文机制在解析生物土壤结皮与维管植物共存模式中可能发挥着核心作用[32].以降水和入渗过程为例,不同种类的生物土壤结皮和维管植物对水分的需求和适应能力不同[27,33],降水频率和强度的变化促使它们在物种组成和数量上做出相应调整,以适应新的水分环境[34-36].水分入渗的速率、深度以及范围等,会改变土壤中的水分分布状况[37],这关系到维管植物与生物土壤结皮在不同土层水分的获取,进而影响二者的生长和竞争关系[38-39].反之,生物土壤结皮与维管植物也并非被动地受水分影响,它们能够通过各自独特的方式影响生态水文过程[40].维管植物凭借自身较为发达的枝叶系统,能够直接截留降水,大大减少降雨对地面的直接冲击,进而减缓地表径流的形成速度[41-42].这不仅改变了水分在土壤表层的分配格局,还对水分的深层渗透和蓄存产生了连锁反应,显著影响了整个区域的水分动态平衡[43].生物土壤结皮可以通过调节径流、入渗和蒸发过程,为维管植物的生长与分布创造适宜的水分条件[44].生物土壤结皮在一定程度上减少了土壤水分的蒸发量,保持了土壤的湿度[20,45].这种对水分的良好调控能力,不仅为维管植物的生长提供了稳定的水分环境,而且通过影响水分的分布状况,进一步影响着不同种类维管植物的分布范围和生长态势[39].此外,生物土壤结皮还能够通过改变土壤水分状况,间接影响灌木的生长[46].土壤水分含量的变化会改变植物的根系生长方向和深度,进而影响植物对养分和水分的获取能力,最终影响灌木的生长状况和分布范围[47]. ...
The pervasive and multifaceted influence of biocrusts on water in the world's drylands
3
2020
... 水分是旱区生态系统的关键限制因子,生态水文机制在解析生物土壤结皮与维管植物共存模式中可能发挥着核心作用[32].以降水和入渗过程为例,不同种类的生物土壤结皮和维管植物对水分的需求和适应能力不同[27,33],降水频率和强度的变化促使它们在物种组成和数量上做出相应调整,以适应新的水分环境[34-36].水分入渗的速率、深度以及范围等,会改变土壤中的水分分布状况[37],这关系到维管植物与生物土壤结皮在不同土层水分的获取,进而影响二者的生长和竞争关系[38-39].反之,生物土壤结皮与维管植物也并非被动地受水分影响,它们能够通过各自独特的方式影响生态水文过程[40].维管植物凭借自身较为发达的枝叶系统,能够直接截留降水,大大减少降雨对地面的直接冲击,进而减缓地表径流的形成速度[41-42].这不仅改变了水分在土壤表层的分配格局,还对水分的深层渗透和蓄存产生了连锁反应,显著影响了整个区域的水分动态平衡[43].生物土壤结皮可以通过调节径流、入渗和蒸发过程,为维管植物的生长与分布创造适宜的水分条件[44].生物土壤结皮在一定程度上减少了土壤水分的蒸发量,保持了土壤的湿度[20,45].这种对水分的良好调控能力,不仅为维管植物的生长提供了稳定的水分环境,而且通过影响水分的分布状况,进一步影响着不同种类维管植物的分布范围和生长态势[39].此外,生物土壤结皮还能够通过改变土壤水分状况,间接影响灌木的生长[46].土壤水分含量的变化会改变植物的根系生长方向和深度,进而影响植物对养分和水分的获取能力,最终影响灌木的生长状况和分布范围[47]. ...
... [39].此外,生物土壤结皮还能够通过改变土壤水分状况,间接影响灌木的生长[46].土壤水分含量的变化会改变植物的根系生长方向和深度,进而影响植物对养分和水分的获取能力,最终影响灌木的生长状况和分布范围[47]. ...
... 旱区生物土壤结皮-维管植物共存的生态水文机制研究,自1993年以来呈逐步上升的趋势(图1).数据库为Web of Science核心合集,检索式为:TS=[(“dryland*” OR “hyper*arid*” OR “arid*” OR “semi*arid*” OR “dry*subhumid*”) AND (“biological soil crust*” OR “biocrust*” OR “biogenic crust*” OR “biological crust*” OR “biotic crust*” OR “microphytic crust*” OR “microbiotic crust*” OR “Cryptogam*” OR “cyanobacteri*” OR “algal*” OR “lichen*” OR “moss*”) AND (“vascular plant” OR “grass” OR “woody” OR “herb*” OR “shrub” OR “bush” OR “turf”) AND (“ecohydrolog*” OR “runoff” OR “infiltration” OR “water flow” OR “ponding” OR “hydrolog*” OR “sorptivity” OR “moisture” OR “erosion” OR “sediment”)],共计检索文献156篇.生态学与水文学交叉融合,为深入探究旱区不同地被类型的存在提供比较全面且系统的阐释[48].目前,生物土壤结皮与维管植物共存的研究主要在生物土壤结皮与维管植被的相互作用[18]、生物土壤结皮与维管植物的生态水文效应[39,49]、生物土壤结皮与维管植物对全球变化的响应等方面[27].然而,对于生物土壤结皮-维管植物共存的生态水文机制研究,目前仍然不足.致力于该领域的研究者以David J.Eldridge、Eli Zaady、Giora J.Kidron、Jayne Belnap、Xinrong Li、Xinping Wang、Ning Chen等人为代表.这些研究主要在旱区分布较广的国家开展,如美国、中国、西班牙、澳大利亚、以色列等. ...
Biocrusts positively affect the soil water balance in semiarid ecosystems
2
2016
... 水分是旱区生态系统的关键限制因子,生态水文机制在解析生物土壤结皮与维管植物共存模式中可能发挥着核心作用[32].以降水和入渗过程为例,不同种类的生物土壤结皮和维管植物对水分的需求和适应能力不同[27,33],降水频率和强度的变化促使它们在物种组成和数量上做出相应调整,以适应新的水分环境[34-36].水分入渗的速率、深度以及范围等,会改变土壤中的水分分布状况[37],这关系到维管植物与生物土壤结皮在不同土层水分的获取,进而影响二者的生长和竞争关系[38-39].反之,生物土壤结皮与维管植物也并非被动地受水分影响,它们能够通过各自独特的方式影响生态水文过程[40].维管植物凭借自身较为发达的枝叶系统,能够直接截留降水,大大减少降雨对地面的直接冲击,进而减缓地表径流的形成速度[41-42].这不仅改变了水分在土壤表层的分配格局,还对水分的深层渗透和蓄存产生了连锁反应,显著影响了整个区域的水分动态平衡[43].生物土壤结皮可以通过调节径流、入渗和蒸发过程,为维管植物的生长与分布创造适宜的水分条件[44].生物土壤结皮在一定程度上减少了土壤水分的蒸发量,保持了土壤的湿度[20,45].这种对水分的良好调控能力,不仅为维管植物的生长提供了稳定的水分环境,而且通过影响水分的分布状况,进一步影响着不同种类维管植物的分布范围和生长态势[39].此外,生物土壤结皮还能够通过改变土壤水分状况,间接影响灌木的生长[46].土壤水分含量的变化会改变植物的根系生长方向和深度,进而影响植物对养分和水分的获取能力,最终影响灌木的生长状况和分布范围[47]. ...
... 土壤水分是影响旱区生态系统动态的另一关键参数[64],对生态系统的年际变化和演变起着决定性作用[65].半干旱时,土壤水分的变化主要受生物土壤结皮盖度的影响,可能导致深层土壤水分供应不足,进而使得该区域的维管植物受到水分胁迫,如人工种植的油蒿(Artemisia ordosica)因水分限制而发生退化甚至死亡[46,62].从另一角度看,生物土壤结皮具有截留降雨的能力,有助于延长表层土壤水分的保持时间并增加浅层土壤水分含量[40,55],从而促进浅根系草本植物的生长和发育[66],进而有利于草本与生物土壤结皮共存,却对灌木与生物土壤结皮共存不利.生物土壤结皮存在的土壤表面更加稳定,有助于提高维管植物盖度;然而,由于土壤水分有效性的降低,生物土壤结皮可能对维管植物产生负面影响[67],最终导致生物土壤结皮占主导地位.另外,由于土壤水分水平的再分配,生物土壤结皮和木本植物也会形成斑块状的植被模式[68]. ...
Climatology and ecohydrology of precipitation pulses in arid and semiarid ecosystems of the western USA
1
2004
... 水分是旱区生态系统的关键限制因子,生态水文机制在解析生物土壤结皮与维管植物共存模式中可能发挥着核心作用[32].以降水和入渗过程为例,不同种类的生物土壤结皮和维管植物对水分的需求和适应能力不同[27,33],降水频率和强度的变化促使它们在物种组成和数量上做出相应调整,以适应新的水分环境[34-36].水分入渗的速率、深度以及范围等,会改变土壤中的水分分布状况[37],这关系到维管植物与生物土壤结皮在不同土层水分的获取,进而影响二者的生长和竞争关系[38-39].反之,生物土壤结皮与维管植物也并非被动地受水分影响,它们能够通过各自独特的方式影响生态水文过程[40].维管植物凭借自身较为发达的枝叶系统,能够直接截留降水,大大减少降雨对地面的直接冲击,进而减缓地表径流的形成速度[41-42].这不仅改变了水分在土壤表层的分配格局,还对水分的深层渗透和蓄存产生了连锁反应,显著影响了整个区域的水分动态平衡[43].生物土壤结皮可以通过调节径流、入渗和蒸发过程,为维管植物的生长与分布创造适宜的水分条件[44].生物土壤结皮在一定程度上减少了土壤水分的蒸发量,保持了土壤的湿度[20,45].这种对水分的良好调控能力,不仅为维管植物的生长提供了稳定的水分环境,而且通过影响水分的分布状况,进一步影响着不同种类维管植物的分布范围和生长态势[39].此外,生物土壤结皮还能够通过改变土壤水分状况,间接影响灌木的生长[46].土壤水分含量的变化会改变植物的根系生长方向和深度,进而影响植物对养分和水分的获取能力,最终影响灌木的生长状况和分布范围[47]. ...
Vegetation patches and runoff-erosion as interacting ecohydrological processes in semiarid landscapes
1
2005
... 水分是旱区生态系统的关键限制因子,生态水文机制在解析生物土壤结皮与维管植物共存模式中可能发挥着核心作用[32].以降水和入渗过程为例,不同种类的生物土壤结皮和维管植物对水分的需求和适应能力不同[27,33],降水频率和强度的变化促使它们在物种组成和数量上做出相应调整,以适应新的水分环境[34-36].水分入渗的速率、深度以及范围等,会改变土壤中的水分分布状况[37],这关系到维管植物与生物土壤结皮在不同土层水分的获取,进而影响二者的生长和竞争关系[38-39].反之,生物土壤结皮与维管植物也并非被动地受水分影响,它们能够通过各自独特的方式影响生态水文过程[40].维管植物凭借自身较为发达的枝叶系统,能够直接截留降水,大大减少降雨对地面的直接冲击,进而减缓地表径流的形成速度[41-42].这不仅改变了水分在土壤表层的分配格局,还对水分的深层渗透和蓄存产生了连锁反应,显著影响了整个区域的水分动态平衡[43].生物土壤结皮可以通过调节径流、入渗和蒸发过程,为维管植物的生长与分布创造适宜的水分条件[44].生物土壤结皮在一定程度上减少了土壤水分的蒸发量,保持了土壤的湿度[20,45].这种对水分的良好调控能力,不仅为维管植物的生长提供了稳定的水分环境,而且通过影响水分的分布状况,进一步影响着不同种类维管植物的分布范围和生长态势[39].此外,生物土壤结皮还能够通过改变土壤水分状况,间接影响灌木的生长[46].土壤水分含量的变化会改变植物的根系生长方向和深度,进而影响植物对养分和水分的获取能力,最终影响灌木的生长状况和分布范围[47]. ...
Ecohydrology of water-limited environments:a scientific vision
1
2006
... 水分是旱区生态系统的关键限制因子,生态水文机制在解析生物土壤结皮与维管植物共存模式中可能发挥着核心作用[32].以降水和入渗过程为例,不同种类的生物土壤结皮和维管植物对水分的需求和适应能力不同[27,33],降水频率和强度的变化促使它们在物种组成和数量上做出相应调整,以适应新的水分环境[34-36].水分入渗的速率、深度以及范围等,会改变土壤中的水分分布状况[37],这关系到维管植物与生物土壤结皮在不同土层水分的获取,进而影响二者的生长和竞争关系[38-39].反之,生物土壤结皮与维管植物也并非被动地受水分影响,它们能够通过各自独特的方式影响生态水文过程[40].维管植物凭借自身较为发达的枝叶系统,能够直接截留降水,大大减少降雨对地面的直接冲击,进而减缓地表径流的形成速度[41-42].这不仅改变了水分在土壤表层的分配格局,还对水分的深层渗透和蓄存产生了连锁反应,显著影响了整个区域的水分动态平衡[43].生物土壤结皮可以通过调节径流、入渗和蒸发过程,为维管植物的生长与分布创造适宜的水分条件[44].生物土壤结皮在一定程度上减少了土壤水分的蒸发量,保持了土壤的湿度[20,45].这种对水分的良好调控能力,不仅为维管植物的生长提供了稳定的水分环境,而且通过影响水分的分布状况,进一步影响着不同种类维管植物的分布范围和生长态势[39].此外,生物土壤结皮还能够通过改变土壤水分状况,间接影响灌木的生长[46].土壤水分含量的变化会改变植物的根系生长方向和深度,进而影响植物对养分和水分的获取能力,最终影响灌木的生长状况和分布范围[47]. ...
The contrasting effect of biocrusts on shallow-rooted perennial plants (hemicryptophytes):increasing mortality (through evaporation) or survival (through runoff)
4
2018
... 水分是旱区生态系统的关键限制因子,生态水文机制在解析生物土壤结皮与维管植物共存模式中可能发挥着核心作用[32].以降水和入渗过程为例,不同种类的生物土壤结皮和维管植物对水分的需求和适应能力不同[27,33],降水频率和强度的变化促使它们在物种组成和数量上做出相应调整,以适应新的水分环境[34-36].水分入渗的速率、深度以及范围等,会改变土壤中的水分分布状况[37],这关系到维管植物与生物土壤结皮在不同土层水分的获取,进而影响二者的生长和竞争关系[38-39].反之,生物土壤结皮与维管植物也并非被动地受水分影响,它们能够通过各自独特的方式影响生态水文过程[40].维管植物凭借自身较为发达的枝叶系统,能够直接截留降水,大大减少降雨对地面的直接冲击,进而减缓地表径流的形成速度[41-42].这不仅改变了水分在土壤表层的分配格局,还对水分的深层渗透和蓄存产生了连锁反应,显著影响了整个区域的水分动态平衡[43].生物土壤结皮可以通过调节径流、入渗和蒸发过程,为维管植物的生长与分布创造适宜的水分条件[44].生物土壤结皮在一定程度上减少了土壤水分的蒸发量,保持了土壤的湿度[20,45].这种对水分的良好调控能力,不仅为维管植物的生长提供了稳定的水分环境,而且通过影响水分的分布状况,进一步影响着不同种类维管植物的分布范围和生长态势[39].此外,生物土壤结皮还能够通过改变土壤水分状况,间接影响灌木的生长[46].土壤水分含量的变化会改变植物的根系生长方向和深度,进而影响植物对养分和水分的获取能力,最终影响灌木的生长状况和分布范围[47]. ...
... 1993年,于澳大利亚东部半干旱林地的控制实验表明,维管植物盖度与隐花植物盖度存在极为紧密的关联.随后,于2002年在以色列内盖夫沙漠北部发现以多年生灌木为主导的景观斑块被生物土壤结皮所隔开[28].2008年,Li等[50]发现在腾格里沙漠的灌木生态系统中,维管植物和生物土壤结皮斑块之间存在源-汇关系.借助长期监测数据和野外实验,通过改变土壤水分及水循环状况,确定腾格里沙漠生物土壤结皮的出现与发育是否会导致固沙植被的变化[51-52].2013年,生物土壤结皮的动态被纳入了模型,构建了一个维管植被和生物土壤结皮耦合模型,一定程度助力了该领域研究的发展[31].研究者在美国加州半干旱草原,利用2010—2014年的实验,探讨在春季仅浇水、施肥+浇水两种试验处理分别对藓类生物土壤结皮和维管植物的影响,试图验证藓类生物土壤结皮和矮维管植物的盖度和多样性会因降雨和养分(两类同时添加)的增加而减少,从而导致高维管植物盖度的增加这一假设[53].Xiao等[46]于2017年在黄土高原的一个半干旱流域布设18个采样点,以验证生物土壤结皮是否会降低土壤湿度,进而导致人工种植灌木退化,经分析得出人工灌木林土壤湿度变化的主导因素是高盖度藓类为主的生物土壤结皮.2018年,于以色列内盖夫沙漠的尼扎纳研究点发现,生物土壤结皮通过增加蒸发导致沙丘斜坡上大多数多年生植物死亡,但同时通过产生地表径流有利于沙丘间洼地浅根系多年生植物的生存[44].同年及次年,Chen等[37,54]构建了一个明确考虑三层土壤(生物土壤结皮层、浅层和深层)的生态水文模型,以此研究不同类型生物土壤结皮对旱区恢复动态的影响.2019年,Jia等[33]开发了一个生态水文模型,并将生物土壤结皮作为系统状态变量,探讨生物土壤结皮、草本及灌木对降雨变化的响应,这有力推动了对全球气候变化下旱区生态系统动态的理解,并有助于预防荒漠化.2022年,She等[27]于毛乌素沙地开展为期5年的降雨和氮添加试验,研究了全球变化如何影响生物土壤结皮-维管植物共存及其机制. ...
... 在旱区生态系统,植物镶嵌分布在生物土壤结皮斑块中,地表径流是资源转移(如水、营养物质和种子)和再分配的关键途径[28,69].灌木冠层间的生物土壤结皮盖度较高,通常有利于径流的生成,且生物土壤结皮斑块通过向灌木斑块输送径流水源和营养元素,支持现存的灌木斑块[13,49,70-71].就草本而言,例如在尼纳扎的沙丘地中,沙丘间的斜坡上可能无法维持地面芽植物生存,但沙丘间洼地中的地面芽植物却得以生长,这得益于沙丘斜坡上生物土壤结皮产生的径流为洼地提供了水分[44],从而形成了生物土壤结皮-草本共存斑块.沙丘间斜坡上的径流(包括地下流动)是多年生植物密集带形成的主要原因[72].然而,考虑土壤水分水平方向的再分配时,生物土壤结皮也可以和木本植物形成斑块状的共存模式[68,73]. ...
... 蒸散作用(蒸发与蒸腾)对于土壤-植物-大气连续体(Soil-Plant-Atmosphere Continuum,SPAC)至关重要[74].地球表面约60%的降水通过蒸散作用返回大气,而在旱区这一比例可高达90%[75].蒸发又作为土壤与大气之间水分和能量平衡的核心要素,在旱区生态水文过程发挥重要的作用[76].然而,降雨强度对蒸发过程具有显著影响.在低强度降雨时,生物土壤结皮通过抑制蒸发保持土壤水分,而在高强度降雨时,则转变为促进蒸发,可能导致土壤水分的损失[59,77].以腾格里沙漠东南缘沙坡头地区为例,约4/5的降雨事件低于5 mm,生物土壤结皮通过抑制或延缓蒸发而保存土壤水分,这为微生物和草本植物的生长提供了有利条件,进而推动植被演替[78],最终形成草本植物主导状态.在极端干旱年份(年降雨量约30 mm),生物土壤结皮较大的蒸发量是一年生植物萌发与生长的主要限制,而未覆盖生物土壤结皮的沙地受此影响较小[79].不同的是,草本植物下方的生物土壤结皮降低了深层土壤水分含量,可能通过减少草根可获得的水分来限制草本植物的蒸腾作用,即生物土壤结皮通过增加土壤蒸发和减少草本植物蒸腾作用加剧了草地生态系统蒸散[80].当植物蒸腾作用所利用水分超过土壤蒸发作用时,存在生物土壤结皮的生态系统最终转变为草本植物主导状态[37],呈现出高盖度草本植物-低盖度生物土壤结皮共存模式.此外,其他生态水文过程的综合影响亦不容忽视.蒸发增加导致沙丘间多数多年生植物死亡,而生物土壤结皮通过其在径流产生中的作用,有助于沙丘间洼地浅根系多年生植物的生长和存活[44],从而促进生物土壤结皮与多年生植物的共存.蒸发与入渗的共同作用对土壤水分的改变也会影响维管植物的生长[62],进而对生物土壤结皮-维管植物的共存产生影响. ...
Effects of surface characteristics on infiltration patterns in an and shrub desert
1
2007
... 水分是旱区生态系统的关键限制因子,生态水文机制在解析生物土壤结皮与维管植物共存模式中可能发挥着核心作用[32].以降水和入渗过程为例,不同种类的生物土壤结皮和维管植物对水分的需求和适应能力不同[27,33],降水频率和强度的变化促使它们在物种组成和数量上做出相应调整,以适应新的水分环境[34-36].水分入渗的速率、深度以及范围等,会改变土壤中的水分分布状况[37],这关系到维管植物与生物土壤结皮在不同土层水分的获取,进而影响二者的生长和竞争关系[38-39].反之,生物土壤结皮与维管植物也并非被动地受水分影响,它们能够通过各自独特的方式影响生态水文过程[40].维管植物凭借自身较为发达的枝叶系统,能够直接截留降水,大大减少降雨对地面的直接冲击,进而减缓地表径流的形成速度[41-42].这不仅改变了水分在土壤表层的分配格局,还对水分的深层渗透和蓄存产生了连锁反应,显著影响了整个区域的水分动态平衡[43].生物土壤结皮可以通过调节径流、入渗和蒸发过程,为维管植物的生长与分布创造适宜的水分条件[44].生物土壤结皮在一定程度上减少了土壤水分的蒸发量,保持了土壤的湿度[20,45].这种对水分的良好调控能力,不仅为维管植物的生长提供了稳定的水分环境,而且通过影响水分的分布状况,进一步影响着不同种类维管植物的分布范围和生长态势[39].此外,生物土壤结皮还能够通过改变土壤水分状况,间接影响灌木的生长[46].土壤水分含量的变化会改变植物的根系生长方向和深度,进而影响植物对养分和水分的获取能力,最终影响灌木的生长状况和分布范围[47]. ...
Moss-dominated biocrusts decrease soil moisture and result in the degradation of artificially planted shrubs under semiarid climate
4
2017
... 水分是旱区生态系统的关键限制因子,生态水文机制在解析生物土壤结皮与维管植物共存模式中可能发挥着核心作用[32].以降水和入渗过程为例,不同种类的生物土壤结皮和维管植物对水分的需求和适应能力不同[27,33],降水频率和强度的变化促使它们在物种组成和数量上做出相应调整,以适应新的水分环境[34-36].水分入渗的速率、深度以及范围等,会改变土壤中的水分分布状况[37],这关系到维管植物与生物土壤结皮在不同土层水分的获取,进而影响二者的生长和竞争关系[38-39].反之,生物土壤结皮与维管植物也并非被动地受水分影响,它们能够通过各自独特的方式影响生态水文过程[40].维管植物凭借自身较为发达的枝叶系统,能够直接截留降水,大大减少降雨对地面的直接冲击,进而减缓地表径流的形成速度[41-42].这不仅改变了水分在土壤表层的分配格局,还对水分的深层渗透和蓄存产生了连锁反应,显著影响了整个区域的水分动态平衡[43].生物土壤结皮可以通过调节径流、入渗和蒸发过程,为维管植物的生长与分布创造适宜的水分条件[44].生物土壤结皮在一定程度上减少了土壤水分的蒸发量,保持了土壤的湿度[20,45].这种对水分的良好调控能力,不仅为维管植物的生长提供了稳定的水分环境,而且通过影响水分的分布状况,进一步影响着不同种类维管植物的分布范围和生长态势[39].此外,生物土壤结皮还能够通过改变土壤水分状况,间接影响灌木的生长[46].土壤水分含量的变化会改变植物的根系生长方向和深度,进而影响植物对养分和水分的获取能力,最终影响灌木的生长状况和分布范围[47]. ...
... 1993年,于澳大利亚东部半干旱林地的控制实验表明,维管植物盖度与隐花植物盖度存在极为紧密的关联.随后,于2002年在以色列内盖夫沙漠北部发现以多年生灌木为主导的景观斑块被生物土壤结皮所隔开[28].2008年,Li等[50]发现在腾格里沙漠的灌木生态系统中,维管植物和生物土壤结皮斑块之间存在源-汇关系.借助长期监测数据和野外实验,通过改变土壤水分及水循环状况,确定腾格里沙漠生物土壤结皮的出现与发育是否会导致固沙植被的变化[51-52].2013年,生物土壤结皮的动态被纳入了模型,构建了一个维管植被和生物土壤结皮耦合模型,一定程度助力了该领域研究的发展[31].研究者在美国加州半干旱草原,利用2010—2014年的实验,探讨在春季仅浇水、施肥+浇水两种试验处理分别对藓类生物土壤结皮和维管植物的影响,试图验证藓类生物土壤结皮和矮维管植物的盖度和多样性会因降雨和养分(两类同时添加)的增加而减少,从而导致高维管植物盖度的增加这一假设[53].Xiao等[46]于2017年在黄土高原的一个半干旱流域布设18个采样点,以验证生物土壤结皮是否会降低土壤湿度,进而导致人工种植灌木退化,经分析得出人工灌木林土壤湿度变化的主导因素是高盖度藓类为主的生物土壤结皮.2018年,于以色列内盖夫沙漠的尼扎纳研究点发现,生物土壤结皮通过增加蒸发导致沙丘斜坡上大多数多年生植物死亡,但同时通过产生地表径流有利于沙丘间洼地浅根系多年生植物的生存[44].同年及次年,Chen等[37,54]构建了一个明确考虑三层土壤(生物土壤结皮层、浅层和深层)的生态水文模型,以此研究不同类型生物土壤结皮对旱区恢复动态的影响.2019年,Jia等[33]开发了一个生态水文模型,并将生物土壤结皮作为系统状态变量,探讨生物土壤结皮、草本及灌木对降雨变化的响应,这有力推动了对全球气候变化下旱区生态系统动态的理解,并有助于预防荒漠化.2022年,She等[27]于毛乌素沙地开展为期5年的降雨和氮添加试验,研究了全球变化如何影响生物土壤结皮-维管植物共存及其机制. ...
... 降水作为旱区生态系统的主要水源,深刻影响生物土壤结皮与维管植物的共存.以较低年降雨量90~150 mm为例,生物土壤结皮相较维管植物往往更具优势[31].主要因为维管植物对水分的需求量大,而生物土壤结皮的需求则小得多.随着降水频率的增加,生物土壤结皮适应的降水条件范围变宽.当日降水频率为0.025时,生物土壤结皮适宜在0~100 mm年降水量范围生长;当日降水频率为1时,生物土壤结皮适宜在0~500 mm年降水量区间生长[33],有力地促进了生物土壤结皮-维管植物共存.干旱事件(降水量显著降低)会降低降雨在土壤中的入渗深度,可能极大降低深根系维管植物的水分利用效率,但并不会对生物土壤结皮产生显著影响[13,46],呈现以生物土壤结皮为主的共存状态.随着降水的增加,生物土壤结皮与维管植物的可利用水分均有所增加,不过两者的竞争优势也随之变化,可能由于过多水分超出生物土壤结皮的可利用范围,增加的降水更多地被维管植物所利用[27].同时,维管植物的盖度过高会极大限制下层光照的有效性,进而导致生物土壤结皮的盖度下降[21].当超过某一临界点时,便会发生从生物土壤结皮主导状态向维管植物主导状态的转变[27].另外,由于生物土壤结皮的存在,降雨模式(降雨量或降雨时间分布)的改变会对灌木斑块和裸露斑块的土壤水分可用性动态产生不同的影响[56].总之,生物土壤结皮和维管植物之间的相互作用对降水的响应是极其复杂的,需考虑研究区域和降水模式的特点[21]. ...
... 土壤水分是影响旱区生态系统动态的另一关键参数[64],对生态系统的年际变化和演变起着决定性作用[65].半干旱时,土壤水分的变化主要受生物土壤结皮盖度的影响,可能导致深层土壤水分供应不足,进而使得该区域的维管植物受到水分胁迫,如人工种植的油蒿(Artemisia ordosica)因水分限制而发生退化甚至死亡[46,62].从另一角度看,生物土壤结皮具有截留降雨的能力,有助于延长表层土壤水分的保持时间并增加浅层土壤水分含量[40,55],从而促进浅根系草本植物的生长和发育[66],进而有利于草本与生物土壤结皮共存,却对灌木与生物土壤结皮共存不利.生物土壤结皮存在的土壤表面更加稳定,有助于提高维管植物盖度;然而,由于土壤水分有效性的降低,生物土壤结皮可能对维管植物产生负面影响[67],最终导致生物土壤结皮占主导地位.另外,由于土壤水分水平的再分配,生物土壤结皮和木本植物也会形成斑块状的植被模式[68]. ...
西北地区地下水依赖型植被生态水文过程研究进展与展望
1
2025
... 水分是旱区生态系统的关键限制因子,生态水文机制在解析生物土壤结皮与维管植物共存模式中可能发挥着核心作用[32].以降水和入渗过程为例,不同种类的生物土壤结皮和维管植物对水分的需求和适应能力不同[27,33],降水频率和强度的变化促使它们在物种组成和数量上做出相应调整,以适应新的水分环境[34-36].水分入渗的速率、深度以及范围等,会改变土壤中的水分分布状况[37],这关系到维管植物与生物土壤结皮在不同土层水分的获取,进而影响二者的生长和竞争关系[38-39].反之,生物土壤结皮与维管植物也并非被动地受水分影响,它们能够通过各自独特的方式影响生态水文过程[40].维管植物凭借自身较为发达的枝叶系统,能够直接截留降水,大大减少降雨对地面的直接冲击,进而减缓地表径流的形成速度[41-42].这不仅改变了水分在土壤表层的分配格局,还对水分的深层渗透和蓄存产生了连锁反应,显著影响了整个区域的水分动态平衡[43].生物土壤结皮可以通过调节径流、入渗和蒸发过程,为维管植物的生长与分布创造适宜的水分条件[44].生物土壤结皮在一定程度上减少了土壤水分的蒸发量,保持了土壤的湿度[20,45].这种对水分的良好调控能力,不仅为维管植物的生长提供了稳定的水分环境,而且通过影响水分的分布状况,进一步影响着不同种类维管植物的分布范围和生长态势[39].此外,生物土壤结皮还能够通过改变土壤水分状况,间接影响灌木的生长[46].土壤水分含量的变化会改变植物的根系生长方向和深度,进而影响植物对养分和水分的获取能力,最终影响灌木的生长状况和分布范围[47]. ...
Dryland ecohydrology and climate change:critical issues and technical advances
1
2012
... 旱区生物土壤结皮-维管植物共存的生态水文机制研究,自1993年以来呈逐步上升的趋势(图1).数据库为Web of Science核心合集,检索式为:TS=[(“dryland*” OR “hyper*arid*” OR “arid*” OR “semi*arid*” OR “dry*subhumid*”) AND (“biological soil crust*” OR “biocrust*” OR “biogenic crust*” OR “biological crust*” OR “biotic crust*” OR “microphytic crust*” OR “microbiotic crust*” OR “Cryptogam*” OR “cyanobacteri*” OR “algal*” OR “lichen*” OR “moss*”) AND (“vascular plant” OR “grass” OR “woody” OR “herb*” OR “shrub” OR “bush” OR “turf”) AND (“ecohydrolog*” OR “runoff” OR “infiltration” OR “water flow” OR “ponding” OR “hydrolog*” OR “sorptivity” OR “moisture” OR “erosion” OR “sediment”)],共计检索文献156篇.生态学与水文学交叉融合,为深入探究旱区不同地被类型的存在提供比较全面且系统的阐释[48].目前,生物土壤结皮与维管植物共存的研究主要在生物土壤结皮与维管植被的相互作用[18]、生物土壤结皮与维管植物的生态水文效应[39,49]、生物土壤结皮与维管植物对全球变化的响应等方面[27].然而,对于生物土壤结皮-维管植物共存的生态水文机制研究,目前仍然不足.致力于该领域的研究者以David J.Eldridge、Eli Zaady、Giora J.Kidron、Jayne Belnap、Xinrong Li、Xinping Wang、Ning Chen等人为代表.这些研究主要在旱区分布较广的国家开展,如美国、中国、西班牙、澳大利亚、以色列等. ...
The dynamic of the eco-hydrological interrelations between shrubs and biocrusts in the Negev shrublands:empiric assessments and perspectives for shrubland rehabilitation
4
2022
... 旱区生物土壤结皮-维管植物共存的生态水文机制研究,自1993年以来呈逐步上升的趋势(图1).数据库为Web of Science核心合集,检索式为:TS=[(“dryland*” OR “hyper*arid*” OR “arid*” OR “semi*arid*” OR “dry*subhumid*”) AND (“biological soil crust*” OR “biocrust*” OR “biogenic crust*” OR “biological crust*” OR “biotic crust*” OR “microphytic crust*” OR “microbiotic crust*” OR “Cryptogam*” OR “cyanobacteri*” OR “algal*” OR “lichen*” OR “moss*”) AND (“vascular plant” OR “grass” OR “woody” OR “herb*” OR “shrub” OR “bush” OR “turf”) AND (“ecohydrolog*” OR “runoff” OR “infiltration” OR “water flow” OR “ponding” OR “hydrolog*” OR “sorptivity” OR “moisture” OR “erosion” OR “sediment”)],共计检索文献156篇.生态学与水文学交叉融合,为深入探究旱区不同地被类型的存在提供比较全面且系统的阐释[48].目前,生物土壤结皮与维管植物共存的研究主要在生物土壤结皮与维管植被的相互作用[18]、生物土壤结皮与维管植物的生态水文效应[39,49]、生物土壤结皮与维管植物对全球变化的响应等方面[27].然而,对于生物土壤结皮-维管植物共存的生态水文机制研究,目前仍然不足.致力于该领域的研究者以David J.Eldridge、Eli Zaady、Giora J.Kidron、Jayne Belnap、Xinrong Li、Xinping Wang、Ning Chen等人为代表.这些研究主要在旱区分布较广的国家开展,如美国、中国、西班牙、澳大利亚、以色列等. ...
... 围绕生物土壤结皮-维管植物共存的生态水文机制,通过Web of Science数据库核心合集检索,共筛选出156篇文献.这些研究在方法运用上,主要采用了野外调查和控制实验两大类方法[27,49].其中,野外调查因其能直接获取自然状态下生物土壤结皮与维管植物共存的实际状况和相关数据,所以在研究中占据主导地位,约有90%的研究侧重于此方法.对不同环境下的生物土壤结皮和维管植物进行直接观测、采样与记录,以全面了解它们之间的相互关系.虽然少部分研究使用控制实验,但这种方法能够在人为设定的条件下,对特定因素进行精准控制和研究,有助于深入剖析某些关键因素对生物土壤结皮-维管植物共存生态水文机制的影响(图2). ...
... 在旱区生态系统,植物镶嵌分布在生物土壤结皮斑块中,地表径流是资源转移(如水、营养物质和种子)和再分配的关键途径[28,69].灌木冠层间的生物土壤结皮盖度较高,通常有利于径流的生成,且生物土壤结皮斑块通过向灌木斑块输送径流水源和营养元素,支持现存的灌木斑块[13,49,70-71].就草本而言,例如在尼纳扎的沙丘地中,沙丘间的斜坡上可能无法维持地面芽植物生存,但沙丘间洼地中的地面芽植物却得以生长,这得益于沙丘斜坡上生物土壤结皮产生的径流为洼地提供了水分[44],从而形成了生物土壤结皮-草本共存斑块.沙丘间斜坡上的径流(包括地下流动)是多年生植物密集带形成的主要原因[72].然而,考虑土壤水分水平方向的再分配时,生物土壤结皮也可以和木本植物形成斑块状的共存模式[68,73]. ...
... 生物土壤结皮-维管植物的共存现象在旱区普遍存在[27,49],这种共存关系对植被恢复及旱区生态系统功能和服务维持有着不可忽视的作用.自21世纪初,研究者们已开始关注两者共存的潜在机制,然而,从生态水文视角探讨其共存模式的研究仍显匮乏.现有研究多关注单一生态水文过程,而涉及多个生态水文过程的综合研究尚不多见.因此,未来研究应综合考虑多个生态水文过程,以全面解析旱区植被共存的潜在机制. ...
Effects of crust and shrub patches on runoff,sedimentation,and related nutrient (C,N) redistribution in the desertified steppe zone of the Tengger Desert,Northern China
1
2008
... 1993年,于澳大利亚东部半干旱林地的控制实验表明,维管植物盖度与隐花植物盖度存在极为紧密的关联.随后,于2002年在以色列内盖夫沙漠北部发现以多年生灌木为主导的景观斑块被生物土壤结皮所隔开[28].2008年,Li等[50]发现在腾格里沙漠的灌木生态系统中,维管植物和生物土壤结皮斑块之间存在源-汇关系.借助长期监测数据和野外实验,通过改变土壤水分及水循环状况,确定腾格里沙漠生物土壤结皮的出现与发育是否会导致固沙植被的变化[51-52].2013年,生物土壤结皮的动态被纳入了模型,构建了一个维管植被和生物土壤结皮耦合模型,一定程度助力了该领域研究的发展[31].研究者在美国加州半干旱草原,利用2010—2014年的实验,探讨在春季仅浇水、施肥+浇水两种试验处理分别对藓类生物土壤结皮和维管植物的影响,试图验证藓类生物土壤结皮和矮维管植物的盖度和多样性会因降雨和养分(两类同时添加)的增加而减少,从而导致高维管植物盖度的增加这一假设[53].Xiao等[46]于2017年在黄土高原的一个半干旱流域布设18个采样点,以验证生物土壤结皮是否会降低土壤湿度,进而导致人工种植灌木退化,经分析得出人工灌木林土壤湿度变化的主导因素是高盖度藓类为主的生物土壤结皮.2018年,于以色列内盖夫沙漠的尼扎纳研究点发现,生物土壤结皮通过增加蒸发导致沙丘斜坡上大多数多年生植物死亡,但同时通过产生地表径流有利于沙丘间洼地浅根系多年生植物的生存[44].同年及次年,Chen等[37,54]构建了一个明确考虑三层土壤(生物土壤结皮层、浅层和深层)的生态水文模型,以此研究不同类型生物土壤结皮对旱区恢复动态的影响.2019年,Jia等[33]开发了一个生态水文模型,并将生物土壤结皮作为系统状态变量,探讨生物土壤结皮、草本及灌木对降雨变化的响应,这有力推动了对全球气候变化下旱区生态系统动态的理解,并有助于预防荒漠化.2022年,She等[27]于毛乌素沙地开展为期5年的降雨和氮添加试验,研究了全球变化如何影响生物土壤结皮-维管植物共存及其机制. ...
Biological soil crust distribution is related to patterns of fragmentation and landuse in a dryland agricultural landscape of southern Australia
1
2008
... 1993年,于澳大利亚东部半干旱林地的控制实验表明,维管植物盖度与隐花植物盖度存在极为紧密的关联.随后,于2002年在以色列内盖夫沙漠北部发现以多年生灌木为主导的景观斑块被生物土壤结皮所隔开[28].2008年,Li等[50]发现在腾格里沙漠的灌木生态系统中,维管植物和生物土壤结皮斑块之间存在源-汇关系.借助长期监测数据和野外实验,通过改变土壤水分及水循环状况,确定腾格里沙漠生物土壤结皮的出现与发育是否会导致固沙植被的变化[51-52].2013年,生物土壤结皮的动态被纳入了模型,构建了一个维管植被和生物土壤结皮耦合模型,一定程度助力了该领域研究的发展[31].研究者在美国加州半干旱草原,利用2010—2014年的实验,探讨在春季仅浇水、施肥+浇水两种试验处理分别对藓类生物土壤结皮和维管植物的影响,试图验证藓类生物土壤结皮和矮维管植物的盖度和多样性会因降雨和养分(两类同时添加)的增加而减少,从而导致高维管植物盖度的增加这一假设[53].Xiao等[46]于2017年在黄土高原的一个半干旱流域布设18个采样点,以验证生物土壤结皮是否会降低土壤湿度,进而导致人工种植灌木退化,经分析得出人工灌木林土壤湿度变化的主导因素是高盖度藓类为主的生物土壤结皮.2018年,于以色列内盖夫沙漠的尼扎纳研究点发现,生物土壤结皮通过增加蒸发导致沙丘斜坡上大多数多年生植物死亡,但同时通过产生地表径流有利于沙丘间洼地浅根系多年生植物的生存[44].同年及次年,Chen等[37,54]构建了一个明确考虑三层土壤(生物土壤结皮层、浅层和深层)的生态水文模型,以此研究不同类型生物土壤结皮对旱区恢复动态的影响.2019年,Jia等[33]开发了一个生态水文模型,并将生物土壤结皮作为系统状态变量,探讨生物土壤结皮、草本及灌木对降雨变化的响应,这有力推动了对全球气候变化下旱区生态系统动态的理解,并有助于预防荒漠化.2022年,She等[27]于毛乌素沙地开展为期5年的降雨和氮添加试验,研究了全球变化如何影响生物土壤结皮-维管植物共存及其机制. ...
Do biological soil crusts determine vegetation changes in sandy deserts?Implications for managing artificial vegetation
3
2010
... 1993年,于澳大利亚东部半干旱林地的控制实验表明,维管植物盖度与隐花植物盖度存在极为紧密的关联.随后,于2002年在以色列内盖夫沙漠北部发现以多年生灌木为主导的景观斑块被生物土壤结皮所隔开[28].2008年,Li等[50]发现在腾格里沙漠的灌木生态系统中,维管植物和生物土壤结皮斑块之间存在源-汇关系.借助长期监测数据和野外实验,通过改变土壤水分及水循环状况,确定腾格里沙漠生物土壤结皮的出现与发育是否会导致固沙植被的变化[51-52].2013年,生物土壤结皮的动态被纳入了模型,构建了一个维管植被和生物土壤结皮耦合模型,一定程度助力了该领域研究的发展[31].研究者在美国加州半干旱草原,利用2010—2014年的实验,探讨在春季仅浇水、施肥+浇水两种试验处理分别对藓类生物土壤结皮和维管植物的影响,试图验证藓类生物土壤结皮和矮维管植物的盖度和多样性会因降雨和养分(两类同时添加)的增加而减少,从而导致高维管植物盖度的增加这一假设[53].Xiao等[46]于2017年在黄土高原的一个半干旱流域布设18个采样点,以验证生物土壤结皮是否会降低土壤湿度,进而导致人工种植灌木退化,经分析得出人工灌木林土壤湿度变化的主导因素是高盖度藓类为主的生物土壤结皮.2018年,于以色列内盖夫沙漠的尼扎纳研究点发现,生物土壤结皮通过增加蒸发导致沙丘斜坡上大多数多年生植物死亡,但同时通过产生地表径流有利于沙丘间洼地浅根系多年生植物的生存[44].同年及次年,Chen等[37,54]构建了一个明确考虑三层土壤(生物土壤结皮层、浅层和深层)的生态水文模型,以此研究不同类型生物土壤结皮对旱区恢复动态的影响.2019年,Jia等[33]开发了一个生态水文模型,并将生物土壤结皮作为系统状态变量,探讨生物土壤结皮、草本及灌木对降雨变化的响应,这有力推动了对全球气候变化下旱区生态系统动态的理解,并有助于预防荒漠化.2022年,She等[27]于毛乌素沙地开展为期5年的降雨和氮添加试验,研究了全球变化如何影响生物土壤结皮-维管植物共存及其机制. ...
... 随着研究工作的持续推进和深入挖掘,长期和短期监测逐渐成为该领域研究的热点话题,受到广泛关注[52].这一转变主要因为生态水文过程的动态变化,时间尺度的差异会导致不同的生态水文现象和机制显现.通过长期的连续监测,可以捕捉到生态水文过程的长期趋势和周期性变化;短期监测则能够敏锐地捕捉到一些快速变化的现象和短期内的响应机制,二者相互补充,为全面理解生物土壤结皮-维管植物共存的生态水文机制提供更丰富的数据和信息. ...
... 降雨后的土壤入渗是一个重要的生态水文过程,是土壤水分补给的主要途径[57].藓类生物土壤结皮的出现限制水分向深层土壤的入渗,会降低深根系植物的可利用水分,最终致使植物群落由深根系旱生灌木向浅根系的草本植物群落转变[52,58-62].在风沙严重的旱区,风沙沉积作用致使水分渗透减少,进而导致深层土壤干燥,生物土壤结皮广泛取代旱生灌木[63].此外,不同类型的生物土壤结皮对降雨土壤入渗的影响存在明显差异,因此,生物土壤结皮的演替(从藻类到地衣和藓类)会使浅层土壤降水入渗比例增加,从而提高草本植物的盖度,使旱区生态系统从裸土状态转变为草本植物主导的荒漠草原状态,实现恢复[37]. ...
Comparing the responses of bryophytes and short-statured vascular plants to climate shifts and eutrophication
1
2017
... 1993年,于澳大利亚东部半干旱林地的控制实验表明,维管植物盖度与隐花植物盖度存在极为紧密的关联.随后,于2002年在以色列内盖夫沙漠北部发现以多年生灌木为主导的景观斑块被生物土壤结皮所隔开[28].2008年,Li等[50]发现在腾格里沙漠的灌木生态系统中,维管植物和生物土壤结皮斑块之间存在源-汇关系.借助长期监测数据和野外实验,通过改变土壤水分及水循环状况,确定腾格里沙漠生物土壤结皮的出现与发育是否会导致固沙植被的变化[51-52].2013年,生物土壤结皮的动态被纳入了模型,构建了一个维管植被和生物土壤结皮耦合模型,一定程度助力了该领域研究的发展[31].研究者在美国加州半干旱草原,利用2010—2014年的实验,探讨在春季仅浇水、施肥+浇水两种试验处理分别对藓类生物土壤结皮和维管植物的影响,试图验证藓类生物土壤结皮和矮维管植物的盖度和多样性会因降雨和养分(两类同时添加)的增加而减少,从而导致高维管植物盖度的增加这一假设[53].Xiao等[46]于2017年在黄土高原的一个半干旱流域布设18个采样点,以验证生物土壤结皮是否会降低土壤湿度,进而导致人工种植灌木退化,经分析得出人工灌木林土壤湿度变化的主导因素是高盖度藓类为主的生物土壤结皮.2018年,于以色列内盖夫沙漠的尼扎纳研究点发现,生物土壤结皮通过增加蒸发导致沙丘斜坡上大多数多年生植物死亡,但同时通过产生地表径流有利于沙丘间洼地浅根系多年生植物的生存[44].同年及次年,Chen等[37,54]构建了一个明确考虑三层土壤(生物土壤结皮层、浅层和深层)的生态水文模型,以此研究不同类型生物土壤结皮对旱区恢复动态的影响.2019年,Jia等[33]开发了一个生态水文模型,并将生物土壤结皮作为系统状态变量,探讨生物土壤结皮、草本及灌木对降雨变化的响应,这有力推动了对全球气候变化下旱区生态系统动态的理解,并有助于预防荒漠化.2022年,She等[27]于毛乌素沙地开展为期5年的降雨和氮添加试验,研究了全球变化如何影响生物土壤结皮-维管植物共存及其机制. ...
Ecohydrological effects of biocrust type on restoration dynamics in drylands
1
2019
... 1993年,于澳大利亚东部半干旱林地的控制实验表明,维管植物盖度与隐花植物盖度存在极为紧密的关联.随后,于2002年在以色列内盖夫沙漠北部发现以多年生灌木为主导的景观斑块被生物土壤结皮所隔开[28].2008年,Li等[50]发现在腾格里沙漠的灌木生态系统中,维管植物和生物土壤结皮斑块之间存在源-汇关系.借助长期监测数据和野外实验,通过改变土壤水分及水循环状况,确定腾格里沙漠生物土壤结皮的出现与发育是否会导致固沙植被的变化[51-52].2013年,生物土壤结皮的动态被纳入了模型,构建了一个维管植被和生物土壤结皮耦合模型,一定程度助力了该领域研究的发展[31].研究者在美国加州半干旱草原,利用2010—2014年的实验,探讨在春季仅浇水、施肥+浇水两种试验处理分别对藓类生物土壤结皮和维管植物的影响,试图验证藓类生物土壤结皮和矮维管植物的盖度和多样性会因降雨和养分(两类同时添加)的增加而减少,从而导致高维管植物盖度的增加这一假设[53].Xiao等[46]于2017年在黄土高原的一个半干旱流域布设18个采样点,以验证生物土壤结皮是否会降低土壤湿度,进而导致人工种植灌木退化,经分析得出人工灌木林土壤湿度变化的主导因素是高盖度藓类为主的生物土壤结皮.2018年,于以色列内盖夫沙漠的尼扎纳研究点发现,生物土壤结皮通过增加蒸发导致沙丘斜坡上大多数多年生植物死亡,但同时通过产生地表径流有利于沙丘间洼地浅根系多年生植物的生存[44].同年及次年,Chen等[37,54]构建了一个明确考虑三层土壤(生物土壤结皮层、浅层和深层)的生态水文模型,以此研究不同类型生物土壤结皮对旱区恢复动态的影响.2019年,Jia等[33]开发了一个生态水文模型,并将生物土壤结皮作为系统状态变量,探讨生物土壤结皮、草本及灌木对降雨变化的响应,这有力推动了对全球气候变化下旱区生态系统动态的理解,并有助于预防荒漠化.2022年,She等[27]于毛乌素沙地开展为期5年的降雨和氮添加试验,研究了全球变化如何影响生物土壤结皮-维管植物共存及其机制. ...
Ecohydrological role of biological soil crusts across a gradient in levels of development
2
2017
... 近年来,随着科技的飞速发展和数据的日益丰富,模型模拟凭借其独特的优势,在该领域发挥着越来越重要的作用.模型模拟具有灵活性,能够根据不同的研究需求和假设进行灵活调整和优化;具备可操控性,可以人为设定和改变各种输入参数,以探究不同条件下生态水文机制的变化;易获取性高,研究人员可以利用已有的数据和软件工具快速构建模型.特别是生态水文模型,它能够综合考虑多种生态水文过程,模拟生物土壤结皮与维管植物在不同条件下的生长、水分运移等过程,有望在未来研究中占据主导地位[33,37,55].构建模型涉及到对生态水文系统的结构、功能和各个组成部分之间关系的抽象和数学描述;接着进行参数化与模型校准,即根据实际数据和研究成果确定模型中各个参数的值,调整模型,使输出结果与实际观测数据尽可能吻合;最后按照设定的条件和参数运行模型并对模拟结果进行验证,通过与更多实际观测数据对比,评估模型的准确性和可靠性. ...
... 土壤水分是影响旱区生态系统动态的另一关键参数[64],对生态系统的年际变化和演变起着决定性作用[65].半干旱时,土壤水分的变化主要受生物土壤结皮盖度的影响,可能导致深层土壤水分供应不足,进而使得该区域的维管植物受到水分胁迫,如人工种植的油蒿(Artemisia ordosica)因水分限制而发生退化甚至死亡[46,62].从另一角度看,生物土壤结皮具有截留降雨的能力,有助于延长表层土壤水分的保持时间并增加浅层土壤水分含量[40,55],从而促进浅根系草本植物的生长和发育[66],进而有利于草本与生物土壤结皮共存,却对灌木与生物土壤结皮共存不利.生物土壤结皮存在的土壤表面更加稳定,有助于提高维管植物盖度;然而,由于土壤水分有效性的降低,生物土壤结皮可能对维管植物产生负面影响[67],最终导致生物土壤结皮占主导地位.另外,由于土壤水分水平的再分配,生物土壤结皮和木本植物也会形成斑块状的植被模式[68]. ...
Divergent effects of biological soil crusts on soil respiration between bare patches and shrub patches under simulated rainfall in a desert ecosystem in Northwest China
1
2021
... 降水作为旱区生态系统的主要水源,深刻影响生物土壤结皮与维管植物的共存.以较低年降雨量90~150 mm为例,生物土壤结皮相较维管植物往往更具优势[31].主要因为维管植物对水分的需求量大,而生物土壤结皮的需求则小得多.随着降水频率的增加,生物土壤结皮适应的降水条件范围变宽.当日降水频率为0.025时,生物土壤结皮适宜在0~100 mm年降水量范围生长;当日降水频率为1时,生物土壤结皮适宜在0~500 mm年降水量区间生长[33],有力地促进了生物土壤结皮-维管植物共存.干旱事件(降水量显著降低)会降低降雨在土壤中的入渗深度,可能极大降低深根系维管植物的水分利用效率,但并不会对生物土壤结皮产生显著影响[13,46],呈现以生物土壤结皮为主的共存状态.随着降水的增加,生物土壤结皮与维管植物的可利用水分均有所增加,不过两者的竞争优势也随之变化,可能由于过多水分超出生物土壤结皮的可利用范围,增加的降水更多地被维管植物所利用[27].同时,维管植物的盖度过高会极大限制下层光照的有效性,进而导致生物土壤结皮的盖度下降[21].当超过某一临界点时,便会发生从生物土壤结皮主导状态向维管植物主导状态的转变[27].另外,由于生物土壤结皮的存在,降雨模式(降雨量或降雨时间分布)的改变会对灌木斑块和裸露斑块的土壤水分可用性动态产生不同的影响[56].总之,生物土壤结皮和维管植物之间的相互作用对降水的响应是极其复杂的,需考虑研究区域和降水模式的特点[21]. ...
Seasonal and land use dependent variability of soil hydraulic and soil hydrological properties of two Northern German soils
1
2008
... 降雨后的土壤入渗是一个重要的生态水文过程,是土壤水分补给的主要途径[57].藓类生物土壤结皮的出现限制水分向深层土壤的入渗,会降低深根系植物的可利用水分,最终致使植物群落由深根系旱生灌木向浅根系的草本植物群落转变[52,58-62].在风沙严重的旱区,风沙沉积作用致使水分渗透减少,进而导致深层土壤干燥,生物土壤结皮广泛取代旱生灌木[63].此外,不同类型的生物土壤结皮对降雨土壤入渗的影响存在明显差异,因此,生物土壤结皮的演替(从藻类到地衣和藓类)会使浅层土壤降水入渗比例增加,从而提高草本植物的盖度,使旱区生态系统从裸土状态转变为草本植物主导的荒漠草原状态,实现恢复[37]. ...
固定沙丘结皮层藓类植物多样性及固沙作用研究
1
2003
... 降雨后的土壤入渗是一个重要的生态水文过程,是土壤水分补给的主要途径[57].藓类生物土壤结皮的出现限制水分向深层土壤的入渗,会降低深根系植物的可利用水分,最终致使植物群落由深根系旱生灌木向浅根系的草本植物群落转变[52,58-62].在风沙严重的旱区,风沙沉积作用致使水分渗透减少,进而导致深层土壤干燥,生物土壤结皮广泛取代旱生灌木[63].此外,不同类型的生物土壤结皮对降雨土壤入渗的影响存在明显差异,因此,生物土壤结皮的演替(从藻类到地衣和藓类)会使浅层土壤降水入渗比例增加,从而提高草本植物的盖度,使旱区生态系统从裸土状态转变为草本植物主导的荒漠草原状态,实现恢复[37]. ...
Long-term effects of revegetation on soil water content of sand dunes in arid region of Northern China
1
2004
... 蒸散作用(蒸发与蒸腾)对于土壤-植物-大气连续体(Soil-Plant-Atmosphere Continuum,SPAC)至关重要[74].地球表面约60%的降水通过蒸散作用返回大气,而在旱区这一比例可高达90%[75].蒸发又作为土壤与大气之间水分和能量平衡的核心要素,在旱区生态水文过程发挥重要的作用[76].然而,降雨强度对蒸发过程具有显著影响.在低强度降雨时,生物土壤结皮通过抑制蒸发保持土壤水分,而在高强度降雨时,则转变为促进蒸发,可能导致土壤水分的损失[59,77].以腾格里沙漠东南缘沙坡头地区为例,约4/5的降雨事件低于5 mm,生物土壤结皮通过抑制或延缓蒸发而保存土壤水分,这为微生物和草本植物的生长提供了有利条件,进而推动植被演替[78],最终形成草本植物主导状态.在极端干旱年份(年降雨量约30 mm),生物土壤结皮较大的蒸发量是一年生植物萌发与生长的主要限制,而未覆盖生物土壤结皮的沙地受此影响较小[79].不同的是,草本植物下方的生物土壤结皮降低了深层土壤水分含量,可能通过减少草根可获得的水分来限制草本植物的蒸腾作用,即生物土壤结皮通过增加土壤蒸发和减少草本植物蒸腾作用加剧了草地生态系统蒸散[80].当植物蒸腾作用所利用水分超过土壤蒸发作用时,存在生物土壤结皮的生态系统最终转变为草本植物主导状态[37],呈现出高盖度草本植物-低盖度生物土壤结皮共存模式.此外,其他生态水文过程的综合影响亦不容忽视.蒸发增加导致沙丘间多数多年生植物死亡,而生物土壤结皮通过其在径流产生中的作用,有助于沙丘间洼地浅根系多年生植物的生长和存活[44],从而促进生物土壤结皮与多年生植物的共存.蒸发与入渗的共同作用对土壤水分的改变也会影响维管植物的生长[62],进而对生物土壤结皮-维管植物的共存产生影响. ...
Negative and positive effects of topsoil biological crusts on water availability along a rainfall gradient in a sandy and area
2007
Differential hydrological response of biological topsoil crusts along a rainfall gradient in a sandy arid area:Northern Negev Desert,Israel
2011
Does biocrust successional stage determine the degradation of vascular vegetation via alterations in its hydrological roles in semi-arid ecosystem?
3
2019
... 降雨后的土壤入渗是一个重要的生态水文过程,是土壤水分补给的主要途径[57].藓类生物土壤结皮的出现限制水分向深层土壤的入渗,会降低深根系植物的可利用水分,最终致使植物群落由深根系旱生灌木向浅根系的草本植物群落转变[52,58-62].在风沙严重的旱区,风沙沉积作用致使水分渗透减少,进而导致深层土壤干燥,生物土壤结皮广泛取代旱生灌木[63].此外,不同类型的生物土壤结皮对降雨土壤入渗的影响存在明显差异,因此,生物土壤结皮的演替(从藻类到地衣和藓类)会使浅层土壤降水入渗比例增加,从而提高草本植物的盖度,使旱区生态系统从裸土状态转变为草本植物主导的荒漠草原状态,实现恢复[37]. ...
... 土壤水分是影响旱区生态系统动态的另一关键参数[64],对生态系统的年际变化和演变起着决定性作用[65].半干旱时,土壤水分的变化主要受生物土壤结皮盖度的影响,可能导致深层土壤水分供应不足,进而使得该区域的维管植物受到水分胁迫,如人工种植的油蒿(Artemisia ordosica)因水分限制而发生退化甚至死亡[46,62].从另一角度看,生物土壤结皮具有截留降雨的能力,有助于延长表层土壤水分的保持时间并增加浅层土壤水分含量[40,55],从而促进浅根系草本植物的生长和发育[66],进而有利于草本与生物土壤结皮共存,却对灌木与生物土壤结皮共存不利.生物土壤结皮存在的土壤表面更加稳定,有助于提高维管植物盖度;然而,由于土壤水分有效性的降低,生物土壤结皮可能对维管植物产生负面影响[67],最终导致生物土壤结皮占主导地位.另外,由于土壤水分水平的再分配,生物土壤结皮和木本植物也会形成斑块状的植被模式[68]. ...
... 蒸散作用(蒸发与蒸腾)对于土壤-植物-大气连续体(Soil-Plant-Atmosphere Continuum,SPAC)至关重要[74].地球表面约60%的降水通过蒸散作用返回大气,而在旱区这一比例可高达90%[75].蒸发又作为土壤与大气之间水分和能量平衡的核心要素,在旱区生态水文过程发挥重要的作用[76].然而,降雨强度对蒸发过程具有显著影响.在低强度降雨时,生物土壤结皮通过抑制蒸发保持土壤水分,而在高强度降雨时,则转变为促进蒸发,可能导致土壤水分的损失[59,77].以腾格里沙漠东南缘沙坡头地区为例,约4/5的降雨事件低于5 mm,生物土壤结皮通过抑制或延缓蒸发而保存土壤水分,这为微生物和草本植物的生长提供了有利条件,进而推动植被演替[78],最终形成草本植物主导状态.在极端干旱年份(年降雨量约30 mm),生物土壤结皮较大的蒸发量是一年生植物萌发与生长的主要限制,而未覆盖生物土壤结皮的沙地受此影响较小[79].不同的是,草本植物下方的生物土壤结皮降低了深层土壤水分含量,可能通过减少草根可获得的水分来限制草本植物的蒸腾作用,即生物土壤结皮通过增加土壤蒸发和减少草本植物蒸腾作用加剧了草地生态系统蒸散[80].当植物蒸腾作用所利用水分超过土壤蒸发作用时,存在生物土壤结皮的生态系统最终转变为草本植物主导状态[37],呈现出高盖度草本植物-低盖度生物土壤结皮共存模式.此外,其他生态水文过程的综合影响亦不容忽视.蒸发增加导致沙丘间多数多年生植物死亡,而生物土壤结皮通过其在径流产生中的作用,有助于沙丘间洼地浅根系多年生植物的生长和存活[44],从而促进生物土壤结皮与多年生植物的共存.蒸发与入渗的共同作用对土壤水分的改变也会影响维管植物的生长[62],进而对生物土壤结皮-维管植物的共存产生影响. ...
Aeolian deposition and its effect on soil and vegetation changes on stabilised desert dunes in northern China
1
1998
... 降雨后的土壤入渗是一个重要的生态水文过程,是土壤水分补给的主要途径[57].藓类生物土壤结皮的出现限制水分向深层土壤的入渗,会降低深根系植物的可利用水分,最终致使植物群落由深根系旱生灌木向浅根系的草本植物群落转变[52,58-62].在风沙严重的旱区,风沙沉积作用致使水分渗透减少,进而导致深层土壤干燥,生物土壤结皮广泛取代旱生灌木[63].此外,不同类型的生物土壤结皮对降雨土壤入渗的影响存在明显差异,因此,生物土壤结皮的演替(从藻类到地衣和藓类)会使浅层土壤降水入渗比例增加,从而提高草本植物的盖度,使旱区生态系统从裸土状态转变为草本植物主导的荒漠草原状态,实现恢复[37]. ...
Dynamics of deep soil moisture in response to vegetational restoration on the Loess Plateau of China
1
2014
... 土壤水分是影响旱区生态系统动态的另一关键参数[64],对生态系统的年际变化和演变起着决定性作用[65].半干旱时,土壤水分的变化主要受生物土壤结皮盖度的影响,可能导致深层土壤水分供应不足,进而使得该区域的维管植物受到水分胁迫,如人工种植的油蒿(Artemisia ordosica)因水分限制而发生退化甚至死亡[46,62].从另一角度看,生物土壤结皮具有截留降雨的能力,有助于延长表层土壤水分的保持时间并增加浅层土壤水分含量[40,55],从而促进浅根系草本植物的生长和发育[66],进而有利于草本与生物土壤结皮共存,却对灌木与生物土壤结皮共存不利.生物土壤结皮存在的土壤表面更加稳定,有助于提高维管植物盖度;然而,由于土壤水分有效性的降低,生物土壤结皮可能对维管植物产生负面影响[67],最终导致生物土壤结皮占主导地位.另外,由于土壤水分水平的再分配,生物土壤结皮和木本植物也会形成斑块状的植被模式[68]. ...
Ecohydrological change mechanism of a rainfed revegetation ecosystem at southeastern edge of Tengger Desert,Northwest China
1
2004
... 土壤水分是影响旱区生态系统动态的另一关键参数[64],对生态系统的年际变化和演变起着决定性作用[65].半干旱时,土壤水分的变化主要受生物土壤结皮盖度的影响,可能导致深层土壤水分供应不足,进而使得该区域的维管植物受到水分胁迫,如人工种植的油蒿(Artemisia ordosica)因水分限制而发生退化甚至死亡[46,62].从另一角度看,生物土壤结皮具有截留降雨的能力,有助于延长表层土壤水分的保持时间并增加浅层土壤水分含量[40,55],从而促进浅根系草本植物的生长和发育[66],进而有利于草本与生物土壤结皮共存,却对灌木与生物土壤结皮共存不利.生物土壤结皮存在的土壤表面更加稳定,有助于提高维管植物盖度;然而,由于土壤水分有效性的降低,生物土壤结皮可能对维管植物产生负面影响[67],最终导致生物土壤结皮占主导地位.另外,由于土壤水分水平的再分配,生物土壤结皮和木本植物也会形成斑块状的植被模式[68]. ...
Ecological restoration and recovery in the wind-blown sand hazard areas of northern China:relationship between soil water and carrying capacity for vegetation in the Tengger Desert
1
2014
... 土壤水分是影响旱区生态系统动态的另一关键参数[64],对生态系统的年际变化和演变起着决定性作用[65].半干旱时,土壤水分的变化主要受生物土壤结皮盖度的影响,可能导致深层土壤水分供应不足,进而使得该区域的维管植物受到水分胁迫,如人工种植的油蒿(Artemisia ordosica)因水分限制而发生退化甚至死亡[46,62].从另一角度看,生物土壤结皮具有截留降雨的能力,有助于延长表层土壤水分的保持时间并增加浅层土壤水分含量[40,55],从而促进浅根系草本植物的生长和发育[66],进而有利于草本与生物土壤结皮共存,却对灌木与生物土壤结皮共存不利.生物土壤结皮存在的土壤表面更加稳定,有助于提高维管植物盖度;然而,由于土壤水分有效性的降低,生物土壤结皮可能对维管植物产生负面影响[67],最终导致生物土壤结皮占主导地位.另外,由于土壤水分水平的再分配,生物土壤结皮和木本植物也会形成斑块状的植被模式[68]. ...
The dual effect of sand-covered biocrusts on annual plants:increasing cover but reducing individual plant biomass and fecundity
1
2019
... 土壤水分是影响旱区生态系统动态的另一关键参数[64],对生态系统的年际变化和演变起着决定性作用[65].半干旱时,土壤水分的变化主要受生物土壤结皮盖度的影响,可能导致深层土壤水分供应不足,进而使得该区域的维管植物受到水分胁迫,如人工种植的油蒿(Artemisia ordosica)因水分限制而发生退化甚至死亡[46,62].从另一角度看,生物土壤结皮具有截留降雨的能力,有助于延长表层土壤水分的保持时间并增加浅层土壤水分含量[40,55],从而促进浅根系草本植物的生长和发育[66],进而有利于草本与生物土壤结皮共存,却对灌木与生物土壤结皮共存不利.生物土壤结皮存在的土壤表面更加稳定,有助于提高维管植物盖度;然而,由于土壤水分有效性的降低,生物土壤结皮可能对维管植物产生负面影响[67],最终导致生物土壤结皮占主导地位.另外,由于土壤水分水平的再分配,生物土壤结皮和木本植物也会形成斑块状的植被模式[68]. ...
A coupled vegetation-crust model for patchy landscapes
2
2016
... 土壤水分是影响旱区生态系统动态的另一关键参数[64],对生态系统的年际变化和演变起着决定性作用[65].半干旱时,土壤水分的变化主要受生物土壤结皮盖度的影响,可能导致深层土壤水分供应不足,进而使得该区域的维管植物受到水分胁迫,如人工种植的油蒿(Artemisia ordosica)因水分限制而发生退化甚至死亡[46,62].从另一角度看,生物土壤结皮具有截留降雨的能力,有助于延长表层土壤水分的保持时间并增加浅层土壤水分含量[40,55],从而促进浅根系草本植物的生长和发育[66],进而有利于草本与生物土壤结皮共存,却对灌木与生物土壤结皮共存不利.生物土壤结皮存在的土壤表面更加稳定,有助于提高维管植物盖度;然而,由于土壤水分有效性的降低,生物土壤结皮可能对维管植物产生负面影响[67],最终导致生物土壤结皮占主导地位.另外,由于土壤水分水平的再分配,生物土壤结皮和木本植物也会形成斑块状的植被模式[68]. ...
... 在旱区生态系统,植物镶嵌分布在生物土壤结皮斑块中,地表径流是资源转移(如水、营养物质和种子)和再分配的关键途径[28,69].灌木冠层间的生物土壤结皮盖度较高,通常有利于径流的生成,且生物土壤结皮斑块通过向灌木斑块输送径流水源和营养元素,支持现存的灌木斑块[13,49,70-71].就草本而言,例如在尼纳扎的沙丘地中,沙丘间的斜坡上可能无法维持地面芽植物生存,但沙丘间洼地中的地面芽植物却得以生长,这得益于沙丘斜坡上生物土壤结皮产生的径流为洼地提供了水分[44],从而形成了生物土壤结皮-草本共存斑块.沙丘间斜坡上的径流(包括地下流动)是多年生植物密集带形成的主要原因[72].然而,考虑土壤水分水平方向的再分配时,生物土壤结皮也可以和木本植物形成斑块状的共存模式[68,73]. ...
Linkages between microbial and hydrologic processes in arid and semiarid watersheds
1
2005
... 在旱区生态系统,植物镶嵌分布在生物土壤结皮斑块中,地表径流是资源转移(如水、营养物质和种子)和再分配的关键途径[28,69].灌木冠层间的生物土壤结皮盖度较高,通常有利于径流的生成,且生物土壤结皮斑块通过向灌木斑块输送径流水源和营养元素,支持现存的灌木斑块[13,49,70-71].就草本而言,例如在尼纳扎的沙丘地中,沙丘间的斜坡上可能无法维持地面芽植物生存,但沙丘间洼地中的地面芽植物却得以生长,这得益于沙丘斜坡上生物土壤结皮产生的径流为洼地提供了水分[44],从而形成了生物土壤结皮-草本共存斑块.沙丘间斜坡上的径流(包括地下流动)是多年生植物密集带形成的主要原因[72].然而,考虑土壤水分水平方向的再分配时,生物土壤结皮也可以和木本植物形成斑块状的共存模式[68,73]. ...
Hydrological function of rapidly induced biocrusts
1
2019
... 在旱区生态系统,植物镶嵌分布在生物土壤结皮斑块中,地表径流是资源转移(如水、营养物质和种子)和再分配的关键途径[28,69].灌木冠层间的生物土壤结皮盖度较高,通常有利于径流的生成,且生物土壤结皮斑块通过向灌木斑块输送径流水源和营养元素,支持现存的灌木斑块[13,49,70-71].就草本而言,例如在尼纳扎的沙丘地中,沙丘间的斜坡上可能无法维持地面芽植物生存,但沙丘间洼地中的地面芽植物却得以生长,这得益于沙丘斜坡上生物土壤结皮产生的径流为洼地提供了水分[44],从而形成了生物土壤结皮-草本共存斑块.沙丘间斜坡上的径流(包括地下流动)是多年生植物密集带形成的主要原因[72].然而,考虑土壤水分水平方向的再分配时,生物土壤结皮也可以和木本植物形成斑块状的共存模式[68,73]. ...
Linkages between biocrust development and water erosion and implications for erosion model implementation
1
2020
... 在旱区生态系统,植物镶嵌分布在生物土壤结皮斑块中,地表径流是资源转移(如水、营养物质和种子)和再分配的关键途径[28,69].灌木冠层间的生物土壤结皮盖度较高,通常有利于径流的生成,且生物土壤结皮斑块通过向灌木斑块输送径流水源和营养元素,支持现存的灌木斑块[13,49,70-71].就草本而言,例如在尼纳扎的沙丘地中,沙丘间的斜坡上可能无法维持地面芽植物生存,但沙丘间洼地中的地面芽植物却得以生长,这得益于沙丘斜坡上生物土壤结皮产生的径流为洼地提供了水分[44],从而形成了生物土壤结皮-草本共存斑块.沙丘间斜坡上的径流(包括地下流动)是多年生植物密集带形成的主要原因[72].然而,考虑土壤水分水平方向的再分配时,生物土壤结皮也可以和木本植物形成斑块状的共存模式[68,73]. ...
Differential water distribution over dune slopes as affected by slope position and microbiotic crust,Negev Desert,Israel
1
1999
... 在旱区生态系统,植物镶嵌分布在生物土壤结皮斑块中,地表径流是资源转移(如水、营养物质和种子)和再分配的关键途径[28,69].灌木冠层间的生物土壤结皮盖度较高,通常有利于径流的生成,且生物土壤结皮斑块通过向灌木斑块输送径流水源和营养元素,支持现存的灌木斑块[13,49,70-71].就草本而言,例如在尼纳扎的沙丘地中,沙丘间的斜坡上可能无法维持地面芽植物生存,但沙丘间洼地中的地面芽植物却得以生长,这得益于沙丘斜坡上生物土壤结皮产生的径流为洼地提供了水分[44],从而形成了生物土壤结皮-草本共存斑块.沙丘间斜坡上的径流(包括地下流动)是多年生植物密集带形成的主要原因[72].然而,考虑土壤水分水平方向的再分配时,生物土壤结皮也可以和木本植物形成斑块状的共存模式[68,73]. ...
Self-organization of vegetation in arid ecosystems
1
2002
... 在旱区生态系统,植物镶嵌分布在生物土壤结皮斑块中,地表径流是资源转移(如水、营养物质和种子)和再分配的关键途径[28,69].灌木冠层间的生物土壤结皮盖度较高,通常有利于径流的生成,且生物土壤结皮斑块通过向灌木斑块输送径流水源和营养元素,支持现存的灌木斑块[13,49,70-71].就草本而言,例如在尼纳扎的沙丘地中,沙丘间的斜坡上可能无法维持地面芽植物生存,但沙丘间洼地中的地面芽植物却得以生长,这得益于沙丘斜坡上生物土壤结皮产生的径流为洼地提供了水分[44],从而形成了生物土壤结皮-草本共存斑块.沙丘间斜坡上的径流(包括地下流动)是多年生植物密集带形成的主要原因[72].然而,考虑土壤水分水平方向的再分配时,生物土壤结皮也可以和木本植物形成斑块状的共存模式[68,73]. ...
The future of evapotranspiration:global requirements for ecosystem functioning,carbon and climate feedbacks,agricultural management,and water resources
1
2017
... 蒸散作用(蒸发与蒸腾)对于土壤-植物-大气连续体(Soil-Plant-Atmosphere Continuum,SPAC)至关重要[74].地球表面约60%的降水通过蒸散作用返回大气,而在旱区这一比例可高达90%[75].蒸发又作为土壤与大气之间水分和能量平衡的核心要素,在旱区生态水文过程发挥重要的作用[76].然而,降雨强度对蒸发过程具有显著影响.在低强度降雨时,生物土壤结皮通过抑制蒸发保持土壤水分,而在高强度降雨时,则转变为促进蒸发,可能导致土壤水分的损失[59,77].以腾格里沙漠东南缘沙坡头地区为例,约4/5的降雨事件低于5 mm,生物土壤结皮通过抑制或延缓蒸发而保存土壤水分,这为微生物和草本植物的生长提供了有利条件,进而推动植被演替[78],最终形成草本植物主导状态.在极端干旱年份(年降雨量约30 mm),生物土壤结皮较大的蒸发量是一年生植物萌发与生长的主要限制,而未覆盖生物土壤结皮的沙地受此影响较小[79].不同的是,草本植物下方的生物土壤结皮降低了深层土壤水分含量,可能通过减少草根可获得的水分来限制草本植物的蒸腾作用,即生物土壤结皮通过增加土壤蒸发和减少草本植物蒸腾作用加剧了草地生态系统蒸散[80].当植物蒸腾作用所利用水分超过土壤蒸发作用时,存在生物土壤结皮的生态系统最终转变为草本植物主导状态[37],呈现出高盖度草本植物-低盖度生物土壤结皮共存模式.此外,其他生态水文过程的综合影响亦不容忽视.蒸发增加导致沙丘间多数多年生植物死亡,而生物土壤结皮通过其在径流产生中的作用,有助于沙丘间洼地浅根系多年生植物的生长和存活[44],从而促进生物土壤结皮与多年生植物的共存.蒸发与入渗的共同作用对土壤水分的改变也会影响维管植物的生长[62],进而对生物土壤结皮-维管植物的共存产生影响. ...
1
2012
... 蒸散作用(蒸发与蒸腾)对于土壤-植物-大气连续体(Soil-Plant-Atmosphere Continuum,SPAC)至关重要[74].地球表面约60%的降水通过蒸散作用返回大气,而在旱区这一比例可高达90%[75].蒸发又作为土壤与大气之间水分和能量平衡的核心要素,在旱区生态水文过程发挥重要的作用[76].然而,降雨强度对蒸发过程具有显著影响.在低强度降雨时,生物土壤结皮通过抑制蒸发保持土壤水分,而在高强度降雨时,则转变为促进蒸发,可能导致土壤水分的损失[59,77].以腾格里沙漠东南缘沙坡头地区为例,约4/5的降雨事件低于5 mm,生物土壤结皮通过抑制或延缓蒸发而保存土壤水分,这为微生物和草本植物的生长提供了有利条件,进而推动植被演替[78],最终形成草本植物主导状态.在极端干旱年份(年降雨量约30 mm),生物土壤结皮较大的蒸发量是一年生植物萌发与生长的主要限制,而未覆盖生物土壤结皮的沙地受此影响较小[79].不同的是,草本植物下方的生物土壤结皮降低了深层土壤水分含量,可能通过减少草根可获得的水分来限制草本植物的蒸腾作用,即生物土壤结皮通过增加土壤蒸发和减少草本植物蒸腾作用加剧了草地生态系统蒸散[80].当植物蒸腾作用所利用水分超过土壤蒸发作用时,存在生物土壤结皮的生态系统最终转变为草本植物主导状态[37],呈现出高盖度草本植物-低盖度生物土壤结皮共存模式.此外,其他生态水文过程的综合影响亦不容忽视.蒸发增加导致沙丘间多数多年生植物死亡,而生物土壤结皮通过其在径流产生中的作用,有助于沙丘间洼地浅根系多年生植物的生长和存活[44],从而促进生物土壤结皮与多年生植物的共存.蒸发与入渗的共同作用对土壤水分的改变也会影响维管植物的生长[62],进而对生物土壤结皮-维管植物的共存产生影响. ...
Increasing effect of biocrusts on evaporation is evidenced by simulating evaporation and diffusion experiments and water stable isotope analysis
1
2024
... 蒸散作用(蒸发与蒸腾)对于土壤-植物-大气连续体(Soil-Plant-Atmosphere Continuum,SPAC)至关重要[74].地球表面约60%的降水通过蒸散作用返回大气,而在旱区这一比例可高达90%[75].蒸发又作为土壤与大气之间水分和能量平衡的核心要素,在旱区生态水文过程发挥重要的作用[76].然而,降雨强度对蒸发过程具有显著影响.在低强度降雨时,生物土壤结皮通过抑制蒸发保持土壤水分,而在高强度降雨时,则转变为促进蒸发,可能导致土壤水分的损失[59,77].以腾格里沙漠东南缘沙坡头地区为例,约4/5的降雨事件低于5 mm,生物土壤结皮通过抑制或延缓蒸发而保存土壤水分,这为微生物和草本植物的生长提供了有利条件,进而推动植被演替[78],最终形成草本植物主导状态.在极端干旱年份(年降雨量约30 mm),生物土壤结皮较大的蒸发量是一年生植物萌发与生长的主要限制,而未覆盖生物土壤结皮的沙地受此影响较小[79].不同的是,草本植物下方的生物土壤结皮降低了深层土壤水分含量,可能通过减少草根可获得的水分来限制草本植物的蒸腾作用,即生物土壤结皮通过增加土壤蒸发和减少草本植物蒸腾作用加剧了草地生态系统蒸散[80].当植物蒸腾作用所利用水分超过土壤蒸发作用时,存在生物土壤结皮的生态系统最终转变为草本植物主导状态[37],呈现出高盖度草本植物-低盖度生物土壤结皮共存模式.此外,其他生态水文过程的综合影响亦不容忽视.蒸发增加导致沙丘间多数多年生植物死亡,而生物土壤结皮通过其在径流产生中的作用,有助于沙丘间洼地浅根系多年生植物的生长和存活[44],从而促进生物土壤结皮与多年生植物的共存.蒸发与入渗的共同作用对土壤水分的改变也会影响维管植物的生长[62],进而对生物土壤结皮-维管植物的共存产生影响. ...
Soil water and temperature patterns in an arid desert dune sand
1
1996
... 蒸散作用(蒸发与蒸腾)对于土壤-植物-大气连续体(Soil-Plant-Atmosphere Continuum,SPAC)至关重要[74].地球表面约60%的降水通过蒸散作用返回大气,而在旱区这一比例可高达90%[75].蒸发又作为土壤与大气之间水分和能量平衡的核心要素,在旱区生态水文过程发挥重要的作用[76].然而,降雨强度对蒸发过程具有显著影响.在低强度降雨时,生物土壤结皮通过抑制蒸发保持土壤水分,而在高强度降雨时,则转变为促进蒸发,可能导致土壤水分的损失[59,77].以腾格里沙漠东南缘沙坡头地区为例,约4/5的降雨事件低于5 mm,生物土壤结皮通过抑制或延缓蒸发而保存土壤水分,这为微生物和草本植物的生长提供了有利条件,进而推动植被演替[78],最终形成草本植物主导状态.在极端干旱年份(年降雨量约30 mm),生物土壤结皮较大的蒸发量是一年生植物萌发与生长的主要限制,而未覆盖生物土壤结皮的沙地受此影响较小[79].不同的是,草本植物下方的生物土壤结皮降低了深层土壤水分含量,可能通过减少草根可获得的水分来限制草本植物的蒸腾作用,即生物土壤结皮通过增加土壤蒸发和减少草本植物蒸腾作用加剧了草地生态系统蒸散[80].当植物蒸腾作用所利用水分超过土壤蒸发作用时,存在生物土壤结皮的生态系统最终转变为草本植物主导状态[37],呈现出高盖度草本植物-低盖度生物土壤结皮共存模式.此外,其他生态水文过程的综合影响亦不容忽视.蒸发增加导致沙丘间多数多年生植物死亡,而生物土壤结皮通过其在径流产生中的作用,有助于沙丘间洼地浅根系多年生植物的生长和存活[44],从而促进生物土壤结皮与多年生植物的共存.蒸发与入渗的共同作用对土壤水分的改变也会影响维管植物的生长[62],进而对生物土壤结皮-维管植物的共存产生影响. ...
Evaporation properties of a revegetated area of the Tengger Desert,North China
1
2008
... 蒸散作用(蒸发与蒸腾)对于土壤-植物-大气连续体(Soil-Plant-Atmosphere Continuum,SPAC)至关重要[74].地球表面约60%的降水通过蒸散作用返回大气,而在旱区这一比例可高达90%[75].蒸发又作为土壤与大气之间水分和能量平衡的核心要素,在旱区生态水文过程发挥重要的作用[76].然而,降雨强度对蒸发过程具有显著影响.在低强度降雨时,生物土壤结皮通过抑制蒸发保持土壤水分,而在高强度降雨时,则转变为促进蒸发,可能导致土壤水分的损失[59,77].以腾格里沙漠东南缘沙坡头地区为例,约4/5的降雨事件低于5 mm,生物土壤结皮通过抑制或延缓蒸发而保存土壤水分,这为微生物和草本植物的生长提供了有利条件,进而推动植被演替[78],最终形成草本植物主导状态.在极端干旱年份(年降雨量约30 mm),生物土壤结皮较大的蒸发量是一年生植物萌发与生长的主要限制,而未覆盖生物土壤结皮的沙地受此影响较小[79].不同的是,草本植物下方的生物土壤结皮降低了深层土壤水分含量,可能通过减少草根可获得的水分来限制草本植物的蒸腾作用,即生物土壤结皮通过增加土壤蒸发和减少草本植物蒸腾作用加剧了草地生态系统蒸散[80].当植物蒸腾作用所利用水分超过土壤蒸发作用时,存在生物土壤结皮的生态系统最终转变为草本植物主导状态[37],呈现出高盖度草本植物-低盖度生物土壤结皮共存模式.此外,其他生态水文过程的综合影响亦不容忽视.蒸发增加导致沙丘间多数多年生植物死亡,而生物土壤结皮通过其在径流产生中的作用,有助于沙丘间洼地浅根系多年生植物的生长和存活[44],从而促进生物土壤结皮与多年生植物的共存.蒸发与入渗的共同作用对土壤水分的改变也会影响维管植物的生长[62],进而对生物土壤结皮-维管植物的共存产生影响. ...
The negative effect of biocrusts upon annual-plant growth on sand dunes during extreme droughts
1
2014
... 蒸散作用(蒸发与蒸腾)对于土壤-植物-大气连续体(Soil-Plant-Atmosphere Continuum,SPAC)至关重要[74].地球表面约60%的降水通过蒸散作用返回大气,而在旱区这一比例可高达90%[75].蒸发又作为土壤与大气之间水分和能量平衡的核心要素,在旱区生态水文过程发挥重要的作用[76].然而,降雨强度对蒸发过程具有显著影响.在低强度降雨时,生物土壤结皮通过抑制蒸发保持土壤水分,而在高强度降雨时,则转变为促进蒸发,可能导致土壤水分的损失[59,77].以腾格里沙漠东南缘沙坡头地区为例,约4/5的降雨事件低于5 mm,生物土壤结皮通过抑制或延缓蒸发而保存土壤水分,这为微生物和草本植物的生长提供了有利条件,进而推动植被演替[78],最终形成草本植物主导状态.在极端干旱年份(年降雨量约30 mm),生物土壤结皮较大的蒸发量是一年生植物萌发与生长的主要限制,而未覆盖生物土壤结皮的沙地受此影响较小[79].不同的是,草本植物下方的生物土壤结皮降低了深层土壤水分含量,可能通过减少草根可获得的水分来限制草本植物的蒸腾作用,即生物土壤结皮通过增加土壤蒸发和减少草本植物蒸腾作用加剧了草地生态系统蒸散[80].当植物蒸腾作用所利用水分超过土壤蒸发作用时,存在生物土壤结皮的生态系统最终转变为草本植物主导状态[37],呈现出高盖度草本植物-低盖度生物土壤结皮共存模式.此外,其他生态水文过程的综合影响亦不容忽视.蒸发增加导致沙丘间多数多年生植物死亡,而生物土壤结皮通过其在径流产生中的作用,有助于沙丘间洼地浅根系多年生植物的生长和存活[44],从而促进生物土壤结皮与多年生植物的共存.蒸发与入渗的共同作用对土壤水分的改变也会影响维管植物的生长[62],进而对生物土壤结皮-维管植物的共存产生影响. ...
Biocrusts intensify grassland evapotranspiration through increasing evaporation and reducing transpiration in a semi-arid ecosystem
1
2024
... 蒸散作用(蒸发与蒸腾)对于土壤-植物-大气连续体(Soil-Plant-Atmosphere Continuum,SPAC)至关重要[74].地球表面约60%的降水通过蒸散作用返回大气,而在旱区这一比例可高达90%[75].蒸发又作为土壤与大气之间水分和能量平衡的核心要素,在旱区生态水文过程发挥重要的作用[76].然而,降雨强度对蒸发过程具有显著影响.在低强度降雨时,生物土壤结皮通过抑制蒸发保持土壤水分,而在高强度降雨时,则转变为促进蒸发,可能导致土壤水分的损失[59,77].以腾格里沙漠东南缘沙坡头地区为例,约4/5的降雨事件低于5 mm,生物土壤结皮通过抑制或延缓蒸发而保存土壤水分,这为微生物和草本植物的生长提供了有利条件,进而推动植被演替[78],最终形成草本植物主导状态.在极端干旱年份(年降雨量约30 mm),生物土壤结皮较大的蒸发量是一年生植物萌发与生长的主要限制,而未覆盖生物土壤结皮的沙地受此影响较小[79].不同的是,草本植物下方的生物土壤结皮降低了深层土壤水分含量,可能通过减少草根可获得的水分来限制草本植物的蒸腾作用,即生物土壤结皮通过增加土壤蒸发和减少草本植物蒸腾作用加剧了草地生态系统蒸散[80].当植物蒸腾作用所利用水分超过土壤蒸发作用时,存在生物土壤结皮的生态系统最终转变为草本植物主导状态[37],呈现出高盖度草本植物-低盖度生物土壤结皮共存模式.此外,其他生态水文过程的综合影响亦不容忽视.蒸发增加导致沙丘间多数多年生植物死亡,而生物土壤结皮通过其在径流产生中的作用,有助于沙丘间洼地浅根系多年生植物的生长和存活[44],从而促进生物土壤结皮与多年生植物的共存.蒸发与入渗的共同作用对土壤水分的改变也会影响维管植物的生长[62],进而对生物土壤结皮-维管植物的共存产生影响. ...
Dew formation characteristics at annual and daily scale in xerophyte shrub plantations at southeast margin of Tengger Desert,Northern China
1
2018
... 露水、雾和大气水汽,是除降雨外的重要水分输入,对旱区生物土壤结皮和维管植物共存至为关键[81-82].通常,露水弥补了由于土壤蒸发和叶片蒸腾导致的水分损耗,为旱生灌木提供关键的水源[83],促使灌木主导生态系统状态.此外,在黎明时分,相较于裸露的沙土,藓类和地衣生物土壤结皮凝结的露水量显著升高,也可为维管植物生存提供一定的水分[84-85],从而为生物土壤结皮-维管植物的共存创造有利条件.因此,露水量的增加有利于生物土壤结皮-维管植物共存状态的形成.在植被形成的间隙空间内,较高的露水量驱动微生物及孢子的生长和发育,进一步加速了生物土壤结皮的形成,反过来,生物土壤结皮的存在促进了露水的形成[83],进而形成了露水形成-共存状态的正反馈. ...
Dew and fog as possible evolutionary drivers?The expansion of crustose and fruticose lichens in the Negev is respectively mainly dictated by dew and fog
1
2022
... 露水、雾和大气水汽,是除降雨外的重要水分输入,对旱区生物土壤结皮和维管植物共存至为关键[81-82].通常,露水弥补了由于土壤蒸发和叶片蒸腾导致的水分损耗,为旱生灌木提供关键的水源[83],促使灌木主导生态系统状态.此外,在黎明时分,相较于裸露的沙土,藓类和地衣生物土壤结皮凝结的露水量显著升高,也可为维管植物生存提供一定的水分[84-85],从而为生物土壤结皮-维管植物的共存创造有利条件.因此,露水量的增加有利于生物土壤结皮-维管植物共存状态的形成.在植被形成的间隙空间内,较高的露水量驱动微生物及孢子的生长和发育,进一步加速了生物土壤结皮的形成,反过来,生物土壤结皮的存在促进了露水的形成[83],进而形成了露水形成-共存状态的正反馈. ...
Dew formation characteristics in a revegetation-stabilized desert ecosystem in Shapotou area,Northern China
2
2010
... 露水、雾和大气水汽,是除降雨外的重要水分输入,对旱区生物土壤结皮和维管植物共存至为关键[81-82].通常,露水弥补了由于土壤蒸发和叶片蒸腾导致的水分损耗,为旱生灌木提供关键的水源[83],促使灌木主导生态系统状态.此外,在黎明时分,相较于裸露的沙土,藓类和地衣生物土壤结皮凝结的露水量显著升高,也可为维管植物生存提供一定的水分[84-85],从而为生物土壤结皮-维管植物的共存创造有利条件.因此,露水量的增加有利于生物土壤结皮-维管植物共存状态的形成.在植被形成的间隙空间内,较高的露水量驱动微生物及孢子的生长和发育,进一步加速了生物土壤结皮的形成,反过来,生物土壤结皮的存在促进了露水的形成[83],进而形成了露水形成-共存状态的正反馈. ...
... [83],进而形成了露水形成-共存状态的正反馈. ...
The influence of biological soil crusts on dew deposition in Gurbantunggut Desert,Northwestern China
1
2009
... 露水、雾和大气水汽,是除降雨外的重要水分输入,对旱区生物土壤结皮和维管植物共存至为关键[81-82].通常,露水弥补了由于土壤蒸发和叶片蒸腾导致的水分损耗,为旱生灌木提供关键的水源[83],促使灌木主导生态系统状态.此外,在黎明时分,相较于裸露的沙土,藓类和地衣生物土壤结皮凝结的露水量显著升高,也可为维管植物生存提供一定的水分[84-85],从而为生物土壤结皮-维管植物的共存创造有利条件.因此,露水量的增加有利于生物土壤结皮-维管植物共存状态的形成.在植被形成的间隙空间内,较高的露水量驱动微生物及孢子的生长和发育,进一步加速了生物土壤结皮的形成,反过来,生物土壤结皮的存在促进了露水的形成[83],进而形成了露水形成-共存状态的正反馈. ...
Species composition,distribution patterns andecological functions of biological soil crusts in the Gurbantunggut Desert
1
2010
... 露水、雾和大气水汽,是除降雨外的重要水分输入,对旱区生物土壤结皮和维管植物共存至为关键[81-82].通常,露水弥补了由于土壤蒸发和叶片蒸腾导致的水分损耗,为旱生灌木提供关键的水源[83],促使灌木主导生态系统状态.此外,在黎明时分,相较于裸露的沙土,藓类和地衣生物土壤结皮凝结的露水量显著升高,也可为维管植物生存提供一定的水分[84-85],从而为生物土壤结皮-维管植物的共存创造有利条件.因此,露水量的增加有利于生物土壤结皮-维管植物共存状态的形成.在植被形成的间隙空间内,较高的露水量驱动微生物及孢子的生长和发育,进一步加速了生物土壤结皮的形成,反过来,生物土壤结皮的存在促进了露水的形成[83],进而形成了露水形成-共存状态的正反馈. ...
Advancing studies on global biocrust distribution
1
2024
... 在研究尺度方面,目前主要在小尺度(局地区域),跨越两个或更多地点的研究案例极少,这极大地约束了研究问题的普适性,无法有力地支撑旱区生态系统恢复与功能维持.未来研究应致力于扩大研究的空间尺度,通过大尺度调查、遥感产品、联网观测和实验平台,收集更为完备的数据资源,从而支撑更全面客观的研究.另外,很多现有的大型数据库(如Splot、Grassplot、VegBank、BLM_AIM等)并非专门针对生物土壤结皮开展调查,仍有大量关于生物土壤结皮的数据,可有力地支持区域甚至全球尺度研究,例如,BLM_AIM数据库报道了超过5 200个地点生物土壤结皮盖度数据.更多关于如何借助现有数据库扩展大尺度生物土壤结皮的方法可参考文献[86]. ...
Catastrophic shifts in ecosystems
1
2001
... 在理论方面,将生物土壤结皮与维管植物的共存现象与生态学理论结合,如系统平衡理论(system equilibrium behavior),有助于从理论层面深入理解其生态水文机制,进而更好地理解生物土壤结皮与维管植物的共存.系统平衡理论指出,在相同环境条件下,可存在两种或多种稳定状态,即不同的生物土壤结皮-维管植物共存模式,为探讨不同生态水文过程下旱区生物土壤结皮-维管植物共存提供全新的视角[18,87].因此,从理论层面推动旱区不同地被类型共存问题,是未来值得深入探讨的重要课题. ...




 甘公网安备 62010202000688号
甘公网安备 62010202000688号
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}