Soil carbon sequestration impacts on global climate change and food security
1
2004
... 土壤有机碳(Soil Organic Carbon,SOC)是陆地碳库的核心组成部分,其储量分别约为大气和植被碳库储量的2倍和3倍[1],在调控陆地环境稳定和全球气候方面至关重要.尽管旱区SOC含量较低,但仍占全球土壤碳储量的27.9%[2].其中,植物光合作用、动植物残骸以及土壤自养微生物的光合固碳,共同构成了旱地SOC的潜在来源[3-4].生物土壤结皮作为旱地生态系统中最具代表性的生物成分,是主要由藻类、地衣和藓类等光合自养群落与异养细菌、真菌和古菌等生物类群与土壤颗粒紧密结合形成的土壤复合体[5].生物土壤结皮约占全球旱区面积的30%[6],约占中国旱区面积的13.9%,主要分布在中国西北荒漠区,其中藻结皮约占中国旱地面积的10.7%[7].藻结皮具有极强的环境适应性和生存能力,能够在极端干旱、贫瘠的环境中迅速定居和繁殖,为地衣和藓类结皮的形成提供条件.同时由于藻结皮具备一定的碳固定能力[8],使其成为干旱半干旱地区土壤的重要碳源,对于维持该区域生物多样性和生态系统功能具有不可替代的作用.为此,研究中国西北荒漠区藻结皮对SOC的影响,对深入理解土壤有机碳固存和碳循环具有重要的意义. ...
Environmental drivers of soil carbon and nitrogen accumulation in global drylands
1
2024
... 土壤有机碳(Soil Organic Carbon,SOC)是陆地碳库的核心组成部分,其储量分别约为大气和植被碳库储量的2倍和3倍[1],在调控陆地环境稳定和全球气候方面至关重要.尽管旱区SOC含量较低,但仍占全球土壤碳储量的27.9%[2].其中,植物光合作用、动植物残骸以及土壤自养微生物的光合固碳,共同构成了旱地SOC的潜在来源[3-4].生物土壤结皮作为旱地生态系统中最具代表性的生物成分,是主要由藻类、地衣和藓类等光合自养群落与异养细菌、真菌和古菌等生物类群与土壤颗粒紧密结合形成的土壤复合体[5].生物土壤结皮约占全球旱区面积的30%[6],约占中国旱区面积的13.9%,主要分布在中国西北荒漠区,其中藻结皮约占中国旱地面积的10.7%[7].藻结皮具有极强的环境适应性和生存能力,能够在极端干旱、贫瘠的环境中迅速定居和繁殖,为地衣和藓类结皮的形成提供条件.同时由于藻结皮具备一定的碳固定能力[8],使其成为干旱半干旱地区土壤的重要碳源,对于维持该区域生物多样性和生态系统功能具有不可替代的作用.为此,研究中国西北荒漠区藻结皮对SOC的影响,对深入理解土壤有机碳固存和碳循环具有重要的意义. ...
Carbon sequestration in dryland ecosystems
1
2004
... 土壤有机碳(Soil Organic Carbon,SOC)是陆地碳库的核心组成部分,其储量分别约为大气和植被碳库储量的2倍和3倍[1],在调控陆地环境稳定和全球气候方面至关重要.尽管旱区SOC含量较低,但仍占全球土壤碳储量的27.9%[2].其中,植物光合作用、动植物残骸以及土壤自养微生物的光合固碳,共同构成了旱地SOC的潜在来源[3-4].生物土壤结皮作为旱地生态系统中最具代表性的生物成分,是主要由藻类、地衣和藓类等光合自养群落与异养细菌、真菌和古菌等生物类群与土壤颗粒紧密结合形成的土壤复合体[5].生物土壤结皮约占全球旱区面积的30%[6],约占中国旱区面积的13.9%,主要分布在中国西北荒漠区,其中藻结皮约占中国旱地面积的10.7%[7].藻结皮具有极强的环境适应性和生存能力,能够在极端干旱、贫瘠的环境中迅速定居和繁殖,为地衣和藓类结皮的形成提供条件.同时由于藻结皮具备一定的碳固定能力[8],使其成为干旱半干旱地区土壤的重要碳源,对于维持该区域生物多样性和生态系统功能具有不可替代的作用.为此,研究中国西北荒漠区藻结皮对SOC的影响,对深入理解土壤有机碳固存和碳循环具有重要的意义. ...
The contentious nature of soil organic matter
1
2015
... 土壤有机碳(Soil Organic Carbon,SOC)是陆地碳库的核心组成部分,其储量分别约为大气和植被碳库储量的2倍和3倍[1],在调控陆地环境稳定和全球气候方面至关重要.尽管旱区SOC含量较低,但仍占全球土壤碳储量的27.9%[2].其中,植物光合作用、动植物残骸以及土壤自养微生物的光合固碳,共同构成了旱地SOC的潜在来源[3-4].生物土壤结皮作为旱地生态系统中最具代表性的生物成分,是主要由藻类、地衣和藓类等光合自养群落与异养细菌、真菌和古菌等生物类群与土壤颗粒紧密结合形成的土壤复合体[5].生物土壤结皮约占全球旱区面积的30%[6],约占中国旱区面积的13.9%,主要分布在中国西北荒漠区,其中藻结皮约占中国旱地面积的10.7%[7].藻结皮具有极强的环境适应性和生存能力,能够在极端干旱、贫瘠的环境中迅速定居和繁殖,为地衣和藓类结皮的形成提供条件.同时由于藻结皮具备一定的碳固定能力[8],使其成为干旱半干旱地区土壤的重要碳源,对于维持该区域生物多样性和生态系统功能具有不可替代的作用.为此,研究中国西北荒漠区藻结皮对SOC的影响,对深入理解土壤有机碳固存和碳循环具有重要的意义. ...
The potential roles of biological soil crusts in dryland hydrologic cycles
1
2006
... 土壤有机碳(Soil Organic Carbon,SOC)是陆地碳库的核心组成部分,其储量分别约为大气和植被碳库储量的2倍和3倍[1],在调控陆地环境稳定和全球气候方面至关重要.尽管旱区SOC含量较低,但仍占全球土壤碳储量的27.9%[2].其中,植物光合作用、动植物残骸以及土壤自养微生物的光合固碳,共同构成了旱地SOC的潜在来源[3-4].生物土壤结皮作为旱地生态系统中最具代表性的生物成分,是主要由藻类、地衣和藓类等光合自养群落与异养细菌、真菌和古菌等生物类群与土壤颗粒紧密结合形成的土壤复合体[5].生物土壤结皮约占全球旱区面积的30%[6],约占中国旱区面积的13.9%,主要分布在中国西北荒漠区,其中藻结皮约占中国旱地面积的10.7%[7].藻结皮具有极强的环境适应性和生存能力,能够在极端干旱、贫瘠的环境中迅速定居和繁殖,为地衣和藓类结皮的形成提供条件.同时由于藻结皮具备一定的碳固定能力[8],使其成为干旱半干旱地区土壤的重要碳源,对于维持该区域生物多样性和生态系统功能具有不可替代的作用.为此,研究中国西北荒漠区藻结皮对SOC的影响,对深入理解土壤有机碳固存和碳循环具有重要的意义. ...
Biocrust as one of multiple stable states in global drylands
1
2020
... 土壤有机碳(Soil Organic Carbon,SOC)是陆地碳库的核心组成部分,其储量分别约为大气和植被碳库储量的2倍和3倍[1],在调控陆地环境稳定和全球气候方面至关重要.尽管旱区SOC含量较低,但仍占全球土壤碳储量的27.9%[2].其中,植物光合作用、动植物残骸以及土壤自养微生物的光合固碳,共同构成了旱地SOC的潜在来源[3-4].生物土壤结皮作为旱地生态系统中最具代表性的生物成分,是主要由藻类、地衣和藓类等光合自养群落与异养细菌、真菌和古菌等生物类群与土壤颗粒紧密结合形成的土壤复合体[5].生物土壤结皮约占全球旱区面积的30%[6],约占中国旱区面积的13.9%,主要分布在中国西北荒漠区,其中藻结皮约占中国旱地面积的10.7%[7].藻结皮具有极强的环境适应性和生存能力,能够在极端干旱、贫瘠的环境中迅速定居和繁殖,为地衣和藓类结皮的形成提供条件.同时由于藻结皮具备一定的碳固定能力[8],使其成为干旱半干旱地区土壤的重要碳源,对于维持该区域生物多样性和生态系统功能具有不可替代的作用.为此,研究中国西北荒漠区藻结皮对SOC的影响,对深入理解土壤有机碳固存和碳循环具有重要的意义. ...
Mapping biocrust distribution in China’s drylands under changing climate
1
2023
... 土壤有机碳(Soil Organic Carbon,SOC)是陆地碳库的核心组成部分,其储量分别约为大气和植被碳库储量的2倍和3倍[1],在调控陆地环境稳定和全球气候方面至关重要.尽管旱区SOC含量较低,但仍占全球土壤碳储量的27.9%[2].其中,植物光合作用、动植物残骸以及土壤自养微生物的光合固碳,共同构成了旱地SOC的潜在来源[3-4].生物土壤结皮作为旱地生态系统中最具代表性的生物成分,是主要由藻类、地衣和藓类等光合自养群落与异养细菌、真菌和古菌等生物类群与土壤颗粒紧密结合形成的土壤复合体[5].生物土壤结皮约占全球旱区面积的30%[6],约占中国旱区面积的13.9%,主要分布在中国西北荒漠区,其中藻结皮约占中国旱地面积的10.7%[7].藻结皮具有极强的环境适应性和生存能力,能够在极端干旱、贫瘠的环境中迅速定居和繁殖,为地衣和藓类结皮的形成提供条件.同时由于藻结皮具备一定的碳固定能力[8],使其成为干旱半干旱地区土壤的重要碳源,对于维持该区域生物多样性和生态系统功能具有不可替代的作用.为此,研究中国西北荒漠区藻结皮对SOC的影响,对深入理解土壤有机碳固存和碳循环具有重要的意义. ...
Effects of biological soil crusts on soil labile organic carbon of patchy alpine meadows in the Source Zone of the Yellow River,West China
2
2023
... 土壤有机碳(Soil Organic Carbon,SOC)是陆地碳库的核心组成部分,其储量分别约为大气和植被碳库储量的2倍和3倍[1],在调控陆地环境稳定和全球气候方面至关重要.尽管旱区SOC含量较低,但仍占全球土壤碳储量的27.9%[2].其中,植物光合作用、动植物残骸以及土壤自养微生物的光合固碳,共同构成了旱地SOC的潜在来源[3-4].生物土壤结皮作为旱地生态系统中最具代表性的生物成分,是主要由藻类、地衣和藓类等光合自养群落与异养细菌、真菌和古菌等生物类群与土壤颗粒紧密结合形成的土壤复合体[5].生物土壤结皮约占全球旱区面积的30%[6],约占中国旱区面积的13.9%,主要分布在中国西北荒漠区,其中藻结皮约占中国旱地面积的10.7%[7].藻结皮具有极强的环境适应性和生存能力,能够在极端干旱、贫瘠的环境中迅速定居和繁殖,为地衣和藓类结皮的形成提供条件.同时由于藻结皮具备一定的碳固定能力[8],使其成为干旱半干旱地区土壤的重要碳源,对于维持该区域生物多样性和生态系统功能具有不可替代的作用.为此,研究中国西北荒漠区藻结皮对SOC的影响,对深入理解土壤有机碳固存和碳循环具有重要的意义. ...
... 土壤理化特征变化会影响土壤微生物的活性以及植物凋落物的输入,这些变化会对SOC含量产生影响[33].本研究结果表明,土壤养分(TN、NH-N和AP等)含量与裸沙和藻结皮POC和MAOC含量呈正相关,这一结果与Sun等[8]对生物土壤结皮斑块碳组分的研究结果相一致.氮作为干旱区的主要限制因素之一,氮富集会增加植物源碳的输入同时为微生物提供氮源,进而减少微生物对有机质的分解,使得POC含量增加[15,29,34].同时,速效养分(NH-N和NO-N)以及磷元素通过共同影响微生物活性(其中磷主要通过与氮的化学计量关系来调节微生物多样性与活性),调节微生物残体和植物凋落物的分解过程,进而促进POC和MAOC的形成[35-36].本研究发现裸沙POC和MAOC含量与pH值负相关,与Zhou等[30]的研究结果一致.pH对微生物影响较大,研究区pH呈碱性,微生物活动受到高pH值抑制后活性降低,进而影响有机质的分解和转化过程,减少MAOC的积累[30];另外,高pH会抑制植物生产力,减少碳输入,影响POC和MAOC含量[37].因此,荒漠区裸沙与藻结皮SOC组分含量分布特征受到多方面因素的综合作用,本研究结果为干旱区土壤碳动态评估提供了理论指导. ...
Dynamic interactions at the mineral-organic matter interface
1
2021
... SOC的固存与土壤矿物间相互作用密切相关,通过形成物理和化学屏障调节SOC周转[9].因此,通常将SOC划分为颗粒态有机碳(Particulate Organic Carbon,POC)和矿物结合态有机碳(Mineral-Associated Organic Carbon,MAOC),分别研究SOC的形成机制和稳定性[10].其中,POC主要源自植物残体的分解过程,易被微生物利用,平均停留时间较短[11],代表不稳定易利用的土壤碳;MAOC的形成涉及微生物残体、分解产物和可溶性植物残留物吸附到土壤矿物表面[12-14],可指示相对稳定且周转周期较长的碳,因此POC/MAOC值一定程度上反映了SOC的稳定性[15].由于POC和MAOC在形成和稳定性方面存在差异,其分布受不同因素驱动.研究表明气候是POC和MAOC储存的重要影响因素[16],气候变暖和降水量增加促使POC含量增加,而MAOC则呈下降或稳定趋势[17].此外,土壤理化性质对土壤微生物的活跃程度以及有机质分解速率具有显著影响,同时土壤中氮磷元素可通过影响植物生长和微生物代谢调控SOC组分的输入量与分解速率[18].因此,解析SOC的稳定性,需要综合考虑气候和土壤特征等影响因子. ...
Conceptualizing soil organic matter into particulate and mineral‐associated forms to address global change in the 21st century
3
2020
... SOC的固存与土壤矿物间相互作用密切相关,通过形成物理和化学屏障调节SOC周转[9].因此,通常将SOC划分为颗粒态有机碳(Particulate Organic Carbon,POC)和矿物结合态有机碳(Mineral-Associated Organic Carbon,MAOC),分别研究SOC的形成机制和稳定性[10].其中,POC主要源自植物残体的分解过程,易被微生物利用,平均停留时间较短[11],代表不稳定易利用的土壤碳;MAOC的形成涉及微生物残体、分解产物和可溶性植物残留物吸附到土壤矿物表面[12-14],可指示相对稳定且周转周期较长的碳,因此POC/MAOC值一定程度上反映了SOC的稳定性[15].由于POC和MAOC在形成和稳定性方面存在差异,其分布受不同因素驱动.研究表明气候是POC和MAOC储存的重要影响因素[16],气候变暖和降水量增加促使POC含量增加,而MAOC则呈下降或稳定趋势[17].此外,土壤理化性质对土壤微生物的活跃程度以及有机质分解速率具有显著影响,同时土壤中氮磷元素可通过影响植物生长和微生物代谢调控SOC组分的输入量与分解速率[18].因此,解析SOC的稳定性,需要综合考虑气候和土壤特征等影响因子. ...
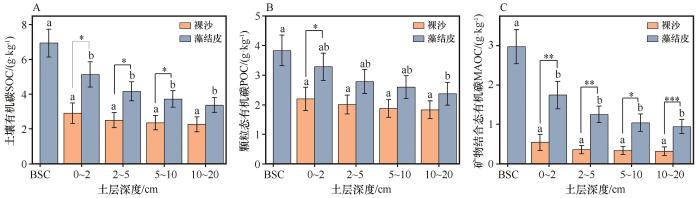
... 随着土层深度的增加,藻结皮层和0~5 cm土层SOC及其组分含量显著高于下层土壤,表明藻结皮层存在养分聚集效应.藻结皮层具有丰富的藻类,可通过光合作用合成有机物,加之藻结皮的覆盖,可以在一定程度上减少土壤表层养分流失,进而使得表层SOC组分含量高于下层[27].同时,表层SOC增加又为土壤微生物生长提供适宜的条件,导致土壤微生物数量随土层深度增加呈现下降趋势.微生物是SOC形成与周转的主要参与者和调控者,在其增殖、生长直至死亡的生命周期中,会产生大量的细胞残留物及其分解过程中释放的小分子代谢产物,部分微生物残体与土壤矿物质发生紧密结合,促进MAOC的积累[10].这一系列过程共同作用使得藻结皮在固定与储存SOC方面展现出显著的能力,成为干旱区中重要的碳汇. ...
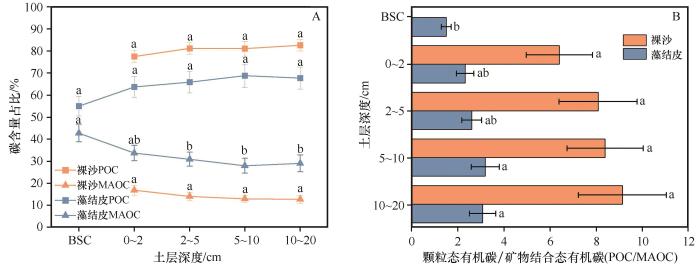
... 裸沙与藻结皮中MAOC占SOC含量的比例呈现出随深度增加而下降的趋势.具体而言,从表层开始,裸沙MAOC含量占SOC的比例由16.85%降至12.69%,而藻结皮中这一比例则由42.70%降至28.95%.这些观测值显著低于先前研究中报道的比例,即SOC的60%以上被储存在MAOC库中[10].西北荒漠区MAOC整体含量较低,一方面,荒漠区降水稀少使得土壤SWC处于较低水平,进而微生物活动受到限制,导致其对植物残体的分解速度相对较低,土壤有机质周转速度减慢,从而有利于POC含量的积累,导致MAOC占比降低.另一方面,荒漠区土壤中粗砂颗粒含量占据优势,其细砂颗粒含量相对较少,有机物质难与矿物表面充分接触和结合,不利于MAOC的形成[28].由于POC易于被微生物分解的特性以及对环境变化的高度敏感性,通过POC/MAOC的值可以反映出SOC稳定性的相对大小[15].本研究中,藻结皮SOC稳定性高于裸沙,主要原因是藻结皮能够增加土壤水分保持能力,为微生物生长和活动提供更适宜的条件,更有利于MAOC含量积累,进而提高了SOC的稳定性[29]. ...
SOM fractionation methods:relevance to functional pools and to stabilization mechanisms
1
2007
... SOC的固存与土壤矿物间相互作用密切相关,通过形成物理和化学屏障调节SOC周转[9].因此,通常将SOC划分为颗粒态有机碳(Particulate Organic Carbon,POC)和矿物结合态有机碳(Mineral-Associated Organic Carbon,MAOC),分别研究SOC的形成机制和稳定性[10].其中,POC主要源自植物残体的分解过程,易被微生物利用,平均停留时间较短[11],代表不稳定易利用的土壤碳;MAOC的形成涉及微生物残体、分解产物和可溶性植物残留物吸附到土壤矿物表面[12-14],可指示相对稳定且周转周期较长的碳,因此POC/MAOC值一定程度上反映了SOC的稳定性[15].由于POC和MAOC在形成和稳定性方面存在差异,其分布受不同因素驱动.研究表明气候是POC和MAOC储存的重要影响因素[16],气候变暖和降水量增加促使POC含量增加,而MAOC则呈下降或稳定趋势[17].此外,土壤理化性质对土壤微生物的活跃程度以及有机质分解速率具有显著影响,同时土壤中氮磷元素可通过影响植物生长和微生物代谢调控SOC组分的输入量与分解速率[18].因此,解析SOC的稳定性,需要综合考虑气候和土壤特征等影响因子. ...
Quantitative assessment of microbial necromass contribution to soil organic matter
1
2019
... SOC的固存与土壤矿物间相互作用密切相关,通过形成物理和化学屏障调节SOC周转[9].因此,通常将SOC划分为颗粒态有机碳(Particulate Organic Carbon,POC)和矿物结合态有机碳(Mineral-Associated Organic Carbon,MAOC),分别研究SOC的形成机制和稳定性[10].其中,POC主要源自植物残体的分解过程,易被微生物利用,平均停留时间较短[11],代表不稳定易利用的土壤碳;MAOC的形成涉及微生物残体、分解产物和可溶性植物残留物吸附到土壤矿物表面[12-14],可指示相对稳定且周转周期较长的碳,因此POC/MAOC值一定程度上反映了SOC的稳定性[15].由于POC和MAOC在形成和稳定性方面存在差异,其分布受不同因素驱动.研究表明气候是POC和MAOC储存的重要影响因素[16],气候变暖和降水量增加促使POC含量增加,而MAOC则呈下降或稳定趋势[17].此外,土壤理化性质对土壤微生物的活跃程度以及有机质分解速率具有显著影响,同时土壤中氮磷元素可通过影响植物生长和微生物代谢调控SOC组分的输入量与分解速率[18].因此,解析SOC的稳定性,需要综合考虑气候和土壤特征等影响因子. ...
Dual,differential isotope labeling shows the preferential movement of labile plant constituents into mineral-bonded soil organic matter
0
2016
Direct evidence for microbial-derived soil organic matter formation and its ecophysiological controls
1
2016
... SOC的固存与土壤矿物间相互作用密切相关,通过形成物理和化学屏障调节SOC周转[9].因此,通常将SOC划分为颗粒态有机碳(Particulate Organic Carbon,POC)和矿物结合态有机碳(Mineral-Associated Organic Carbon,MAOC),分别研究SOC的形成机制和稳定性[10].其中,POC主要源自植物残体的分解过程,易被微生物利用,平均停留时间较短[11],代表不稳定易利用的土壤碳;MAOC的形成涉及微生物残体、分解产物和可溶性植物残留物吸附到土壤矿物表面[12-14],可指示相对稳定且周转周期较长的碳,因此POC/MAOC值一定程度上反映了SOC的稳定性[15].由于POC和MAOC在形成和稳定性方面存在差异,其分布受不同因素驱动.研究表明气候是POC和MAOC储存的重要影响因素[16],气候变暖和降水量增加促使POC含量增加,而MAOC则呈下降或稳定趋势[17].此外,土壤理化性质对土壤微生物的活跃程度以及有机质分解速率具有显著影响,同时土壤中氮磷元素可通过影响植物生长和微生物代谢调控SOC组分的输入量与分解速率[18].因此,解析SOC的稳定性,需要综合考虑气候和土壤特征等影响因子. ...
Soil organic carbon fractions in China: spatial distribution,drivers,and future changes
4
2024
... SOC的固存与土壤矿物间相互作用密切相关,通过形成物理和化学屏障调节SOC周转[9].因此,通常将SOC划分为颗粒态有机碳(Particulate Organic Carbon,POC)和矿物结合态有机碳(Mineral-Associated Organic Carbon,MAOC),分别研究SOC的形成机制和稳定性[10].其中,POC主要源自植物残体的分解过程,易被微生物利用,平均停留时间较短[11],代表不稳定易利用的土壤碳;MAOC的形成涉及微生物残体、分解产物和可溶性植物残留物吸附到土壤矿物表面[12-14],可指示相对稳定且周转周期较长的碳,因此POC/MAOC值一定程度上反映了SOC的稳定性[15].由于POC和MAOC在形成和稳定性方面存在差异,其分布受不同因素驱动.研究表明气候是POC和MAOC储存的重要影响因素[16],气候变暖和降水量增加促使POC含量增加,而MAOC则呈下降或稳定趋势[17].此外,土壤理化性质对土壤微生物的活跃程度以及有机质分解速率具有显著影响,同时土壤中氮磷元素可通过影响植物生长和微生物代谢调控SOC组分的输入量与分解速率[18].因此,解析SOC的稳定性,需要综合考虑气候和土壤特征等影响因子. ...
... 裸沙与藻结皮中MAOC占SOC含量的比例呈现出随深度增加而下降的趋势.具体而言,从表层开始,裸沙MAOC含量占SOC的比例由16.85%降至12.69%,而藻结皮中这一比例则由42.70%降至28.95%.这些观测值显著低于先前研究中报道的比例,即SOC的60%以上被储存在MAOC库中[10].西北荒漠区MAOC整体含量较低,一方面,荒漠区降水稀少使得土壤SWC处于较低水平,进而微生物活动受到限制,导致其对植物残体的分解速度相对较低,土壤有机质周转速度减慢,从而有利于POC含量的积累,导致MAOC占比降低.另一方面,荒漠区土壤中粗砂颗粒含量占据优势,其细砂颗粒含量相对较少,有机物质难与矿物表面充分接触和结合,不利于MAOC的形成[28].由于POC易于被微生物分解的特性以及对环境变化的高度敏感性,通过POC/MAOC的值可以反映出SOC稳定性的相对大小[15].本研究中,藻结皮SOC稳定性高于裸沙,主要原因是藻结皮能够增加土壤水分保持能力,为微生物生长和活动提供更适宜的条件,更有利于MAOC含量积累,进而提高了SOC的稳定性[29]. ...
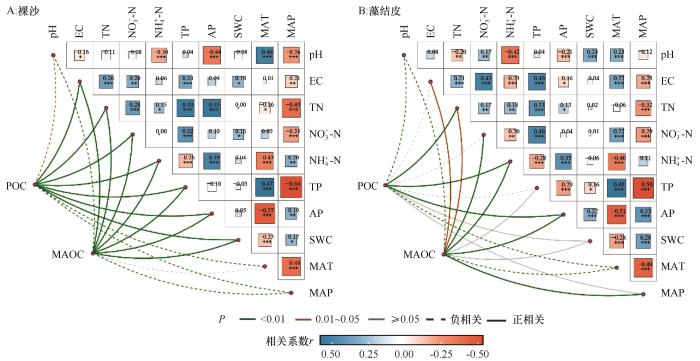
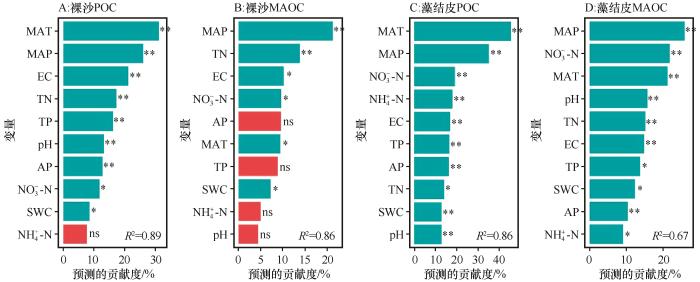
... 在区域尺度下,SOC的变化主要受气候、土壤和植被条件的影响[16].本研究也表明,裸沙和藻结皮SOC组分变化的重要预测变量为MAP和MAT.多数研究认为,生物土壤结皮只有在外界温度和水分条件达到一定程度才能正常生长,其生长发育所需要的水分来自近地表土壤水分,引起微生物的快速响应[24,30],进而影响其SOC含量.在干旱区,降水对SOC的影响主要通过土壤微生物活性而非植被[31].温度升高可能会增加微生物生长和周转,提高微生物代谢活性,从而会加速POC分解和损失,使得SOC含量减少.另外,变暖会引起土壤真菌相对丰度增加,促进MAOC分解,从而导致MAOC储量减少[15].因此,二者与MAT呈负相关关系.小尺度上,一般来说,适宜的SWC有利于SOC积累,但是干旱区SWC比较低,SWC增加使得藻结皮增加植物碳输入,从而减轻微生物抑制,使得POC和MAOC含量增加[32].因此,SWC与POC和MAOC含量呈正相关. ...
... 土壤理化特征变化会影响土壤微生物的活性以及植物凋落物的输入,这些变化会对SOC含量产生影响[33].本研究结果表明,土壤养分(TN、NH-N和AP等)含量与裸沙和藻结皮POC和MAOC含量呈正相关,这一结果与Sun等[8]对生物土壤结皮斑块碳组分的研究结果相一致.氮作为干旱区的主要限制因素之一,氮富集会增加植物源碳的输入同时为微生物提供氮源,进而减少微生物对有机质的分解,使得POC含量增加[15,29,34].同时,速效养分(NH-N和NO-N)以及磷元素通过共同影响微生物活性(其中磷主要通过与氮的化学计量关系来调节微生物多样性与活性),调节微生物残体和植物凋落物的分解过程,进而促进POC和MAOC的形成[35-36].本研究发现裸沙POC和MAOC含量与pH值负相关,与Zhou等[30]的研究结果一致.pH对微生物影响较大,研究区pH呈碱性,微生物活动受到高pH值抑制后活性降低,进而影响有机质的分解和转化过程,减少MAOC的积累[30];另外,高pH会抑制植物生产力,减少碳输入,影响POC和MAOC含量[37].因此,荒漠区裸沙与藻结皮SOC组分含量分布特征受到多方面因素的综合作用,本研究结果为干旱区土壤碳动态评估提供了理论指导. ...
Distinct,direct and climate-mediated environmental controls on global particulate and mineral-associated organic carbon storage
2
2024
... SOC的固存与土壤矿物间相互作用密切相关,通过形成物理和化学屏障调节SOC周转[9].因此,通常将SOC划分为颗粒态有机碳(Particulate Organic Carbon,POC)和矿物结合态有机碳(Mineral-Associated Organic Carbon,MAOC),分别研究SOC的形成机制和稳定性[10].其中,POC主要源自植物残体的分解过程,易被微生物利用,平均停留时间较短[11],代表不稳定易利用的土壤碳;MAOC的形成涉及微生物残体、分解产物和可溶性植物残留物吸附到土壤矿物表面[12-14],可指示相对稳定且周转周期较长的碳,因此POC/MAOC值一定程度上反映了SOC的稳定性[15].由于POC和MAOC在形成和稳定性方面存在差异,其分布受不同因素驱动.研究表明气候是POC和MAOC储存的重要影响因素[16],气候变暖和降水量增加促使POC含量增加,而MAOC则呈下降或稳定趋势[17].此外,土壤理化性质对土壤微生物的活跃程度以及有机质分解速率具有显著影响,同时土壤中氮磷元素可通过影响植物生长和微生物代谢调控SOC组分的输入量与分解速率[18].因此,解析SOC的稳定性,需要综合考虑气候和土壤特征等影响因子. ...
... 在区域尺度下,SOC的变化主要受气候、土壤和植被条件的影响[16].本研究也表明,裸沙和藻结皮SOC组分变化的重要预测变量为MAP和MAT.多数研究认为,生物土壤结皮只有在外界温度和水分条件达到一定程度才能正常生长,其生长发育所需要的水分来自近地表土壤水分,引起微生物的快速响应[24,30],进而影响其SOC含量.在干旱区,降水对SOC的影响主要通过土壤微生物活性而非植被[31].温度升高可能会增加微生物生长和周转,提高微生物代谢活性,从而会加速POC分解和损失,使得SOC含量减少.另外,变暖会引起土壤真菌相对丰度增加,促进MAOC分解,从而导致MAOC储量减少[15].因此,二者与MAT呈负相关关系.小尺度上,一般来说,适宜的SWC有利于SOC积累,但是干旱区SWC比较低,SWC增加使得藻结皮增加植物碳输入,从而减轻微生物抑制,使得POC和MAOC含量增加[32].因此,SWC与POC和MAOC含量呈正相关. ...
Soil organic carbon response to global environmental change depends on its distribution between mineral-associated and particulate organic matter:a meta-analysis
1
2021
... SOC的固存与土壤矿物间相互作用密切相关,通过形成物理和化学屏障调节SOC周转[9].因此,通常将SOC划分为颗粒态有机碳(Particulate Organic Carbon,POC)和矿物结合态有机碳(Mineral-Associated Organic Carbon,MAOC),分别研究SOC的形成机制和稳定性[10].其中,POC主要源自植物残体的分解过程,易被微生物利用,平均停留时间较短[11],代表不稳定易利用的土壤碳;MAOC的形成涉及微生物残体、分解产物和可溶性植物残留物吸附到土壤矿物表面[12-14],可指示相对稳定且周转周期较长的碳,因此POC/MAOC值一定程度上反映了SOC的稳定性[15].由于POC和MAOC在形成和稳定性方面存在差异,其分布受不同因素驱动.研究表明气候是POC和MAOC储存的重要影响因素[16],气候变暖和降水量增加促使POC含量增加,而MAOC则呈下降或稳定趋势[17].此外,土壤理化性质对土壤微生物的活跃程度以及有机质分解速率具有显著影响,同时土壤中氮磷元素可通过影响植物生长和微生物代谢调控SOC组分的输入量与分解速率[18].因此,解析SOC的稳定性,需要综合考虑气候和土壤特征等影响因子. ...
Soil microbial carbon and nitrogen limitation constraints soil organic carbon stability in arid and semi-arid grasslands
1
2025
... SOC的固存与土壤矿物间相互作用密切相关,通过形成物理和化学屏障调节SOC周转[9].因此,通常将SOC划分为颗粒态有机碳(Particulate Organic Carbon,POC)和矿物结合态有机碳(Mineral-Associated Organic Carbon,MAOC),分别研究SOC的形成机制和稳定性[10].其中,POC主要源自植物残体的分解过程,易被微生物利用,平均停留时间较短[11],代表不稳定易利用的土壤碳;MAOC的形成涉及微生物残体、分解产物和可溶性植物残留物吸附到土壤矿物表面[12-14],可指示相对稳定且周转周期较长的碳,因此POC/MAOC值一定程度上反映了SOC的稳定性[15].由于POC和MAOC在形成和稳定性方面存在差异,其分布受不同因素驱动.研究表明气候是POC和MAOC储存的重要影响因素[16],气候变暖和降水量增加促使POC含量增加,而MAOC则呈下降或稳定趋势[17].此外,土壤理化性质对土壤微生物的活跃程度以及有机质分解速率具有显著影响,同时土壤中氮磷元素可通过影响植物生长和微生物代谢调控SOC组分的输入量与分解速率[18].因此,解析SOC的稳定性,需要综合考虑气候和土壤特征等影响因子. ...
Soil nitrogen and climate drive the positive effect of biological soil crusts on soil organic carbon sequestration in drylands:a meta-analysis
1
2022
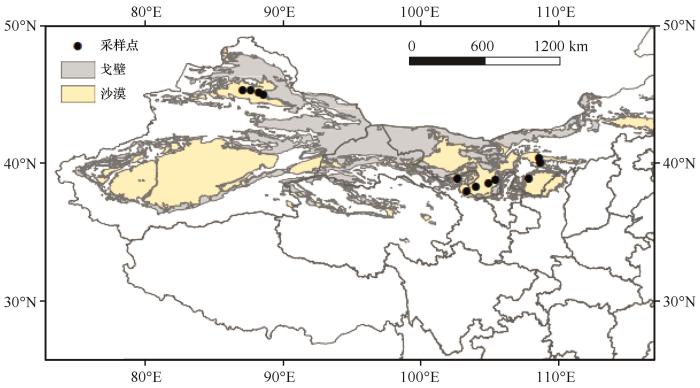
... 国内外关于生物土壤结皮SOC周转方面已取得诸多进展.研究表明生物土壤结皮对荒漠SOC的积极影响强于草原[19];此外,在黄土高原北部小流域的研究表明,生物土壤结皮显著提高了SOC含量,并增加了表层土壤碳储量[20].目前相关研究大多局限于特定的点位,对于区域生物土壤结皮覆盖对SOC组分含量及其驱动因子的研究较少.中国西北荒漠区具有特殊的光-温-湿的气候梯度分布格局,这些因子如何影响藻结皮覆盖下SOC含量及其垂直分布特征仍缺乏深入的理解.本研究调查了西北荒漠区具有生物土壤结皮分布的12个样点,并结合气候数据和土壤理化特征,分析了藻结皮覆盖下0~20 cm不同土层中SOC组分含量的分布特征及其影响因素,旨在揭示藻结皮在该区域SOC动态变化中的重要作用,为区域碳汇评估与管理提供科学依据. ...
Contributions of three types of biocrusts to soil carbon stock and annual efflux in a small watershed of Northern Chinese Loess Plateau
1
2022
... 国内外关于生物土壤结皮SOC周转方面已取得诸多进展.研究表明生物土壤结皮对荒漠SOC的积极影响强于草原[19];此外,在黄土高原北部小流域的研究表明,生物土壤结皮显著提高了SOC含量,并增加了表层土壤碳储量[20].目前相关研究大多局限于特定的点位,对于区域生物土壤结皮覆盖对SOC组分含量及其驱动因子的研究较少.中国西北荒漠区具有特殊的光-温-湿的气候梯度分布格局,这些因子如何影响藻结皮覆盖下SOC含量及其垂直分布特征仍缺乏深入的理解.本研究调查了西北荒漠区具有生物土壤结皮分布的12个样点,并结合气候数据和土壤理化特征,分析了藻结皮覆盖下0~20 cm不同土层中SOC组分含量的分布特征及其影响因素,旨在揭示藻结皮在该区域SOC动态变化中的重要作用,为区域碳汇评估与管理提供科学依据. ...
西北荒漠区土壤氮素组分沿干旱梯度的空间变化
1
2024
... 中国西北荒漠区地处中纬度地带的欧亚大陆腹地,以山地和荒漠为主体,自西北向东南经过的主要沙漠为古尔班通古特沙漠、巴丹吉林沙漠、腾格里沙漠以及毛乌素沙地等.该地区降水稀少,日照强烈,气温变化强烈,其植被覆盖主要是以盐生旱生和超旱生以及耐盐碱的草本、灌木及小乔木为建群种构成的稀疏植被,同时生物土壤结皮广泛分布,藻结皮是西北荒漠区主要的结皮类型[21]. ...
Particulate Soil organic-matter changes across a grassland cultivation sequence
1
1992
... POC和MAOC含量采用六偏磷酸钠分散法分离测定[22].取10 g风干土壤样品放入100 mL塑料瓶中,加入60 mL 0.5%六偏磷酸钠溶液,置于摇床(90 r·min-1)振摇18 h分散样品,以破坏聚集体.将土壤悬液过53 µm筛,反复用蒸馏水漂洗,直至水流清澈.筛下的细质部分(˂53 µm)为矿物结合态有机质(MAOM),留在筛上的粗质部分(>53 µm)为颗粒态有机质(POM),将筛上下组分在60 ℃烘箱中烘48 h至恒重,称重后收集,将烘干土壤研磨,使用碳氮分析仪测定其碳含量. ...
黄土高原生物土壤结皮发育过程中颗粒态和矿物结合态有机碳变化特征
1
2023
... 随着土层深度的增加,裸沙和藻结皮的SOC、POC和MAOC含量均呈下降趋势,且同土层SOC组分含量表现为藻结皮高于裸沙.这与已有研究结果一致,在黄土高原,生物土壤结皮POC和MAOC含量随土层增加而降低[23].对毛乌素沙地藻结皮研究发现,其覆盖下SOC含量为裸沙的2.1倍[24].藻结皮发育在干旱区土壤发育和稳定中具有重要的生态功能,一方面,藻结皮作为生物土壤结皮初级演替阶段,其固碳作用主要通过原核生物和光合生物固碳途径完成[25],为有机质的累积提供了基础.另一方面,干旱区土壤普遍缺乏氮素,藻结皮中的固氮蓝藻在生物固氮中发挥重要作用,可为其他植物及土壤微生物供给充足的氮素资源[26],不仅促进了植物生长和土壤微生物活性,还增强了土壤中有机碳输入量,逐步增加生物土壤结皮的稳定性. ...
毛乌素沙地区域尺度生物结皮有机碳空间分布特征及其影响因素
2
2022
... 随着土层深度的增加,裸沙和藻结皮的SOC、POC和MAOC含量均呈下降趋势,且同土层SOC组分含量表现为藻结皮高于裸沙.这与已有研究结果一致,在黄土高原,生物土壤结皮POC和MAOC含量随土层增加而降低[23].对毛乌素沙地藻结皮研究发现,其覆盖下SOC含量为裸沙的2.1倍[24].藻结皮发育在干旱区土壤发育和稳定中具有重要的生态功能,一方面,藻结皮作为生物土壤结皮初级演替阶段,其固碳作用主要通过原核生物和光合生物固碳途径完成[25],为有机质的累积提供了基础.另一方面,干旱区土壤普遍缺乏氮素,藻结皮中的固氮蓝藻在生物固氮中发挥重要作用,可为其他植物及土壤微生物供给充足的氮素资源[26],不仅促进了植物生长和土壤微生物活性,还增强了土壤中有机碳输入量,逐步增加生物土壤结皮的稳定性. ...
... 在区域尺度下,SOC的变化主要受气候、土壤和植被条件的影响[16].本研究也表明,裸沙和藻结皮SOC组分变化的重要预测变量为MAP和MAT.多数研究认为,生物土壤结皮只有在外界温度和水分条件达到一定程度才能正常生长,其生长发育所需要的水分来自近地表土壤水分,引起微生物的快速响应[24,30],进而影响其SOC含量.在干旱区,降水对SOC的影响主要通过土壤微生物活性而非植被[31].温度升高可能会增加微生物生长和周转,提高微生物代谢活性,从而会加速POC分解和损失,使得SOC含量减少.另外,变暖会引起土壤真菌相对丰度增加,促进MAOC分解,从而导致MAOC储量减少[15].因此,二者与MAT呈负相关关系.小尺度上,一般来说,适宜的SWC有利于SOC积累,但是干旱区SWC比较低,SWC增加使得藻结皮增加植物碳输入,从而减轻微生物抑制,使得POC和MAOC含量增加[32].因此,SWC与POC和MAOC含量呈正相关. ...
古尔班通古特沙漠不同区域藻类结皮微生物结构和潜在功能
1
2024
... 随着土层深度的增加,裸沙和藻结皮的SOC、POC和MAOC含量均呈下降趋势,且同土层SOC组分含量表现为藻结皮高于裸沙.这与已有研究结果一致,在黄土高原,生物土壤结皮POC和MAOC含量随土层增加而降低[23].对毛乌素沙地藻结皮研究发现,其覆盖下SOC含量为裸沙的2.1倍[24].藻结皮发育在干旱区土壤发育和稳定中具有重要的生态功能,一方面,藻结皮作为生物土壤结皮初级演替阶段,其固碳作用主要通过原核生物和光合生物固碳途径完成[25],为有机质的累积提供了基础.另一方面,干旱区土壤普遍缺乏氮素,藻结皮中的固氮蓝藻在生物固氮中发挥重要作用,可为其他植物及土壤微生物供给充足的氮素资源[26],不仅促进了植物生长和土壤微生物活性,还增强了土壤中有机碳输入量,逐步增加生物土壤结皮的稳定性. ...
黄土高原生物结皮对土壤养分的表层聚集与吸附固持效应
1
2021
... 随着土层深度的增加,裸沙和藻结皮的SOC、POC和MAOC含量均呈下降趋势,且同土层SOC组分含量表现为藻结皮高于裸沙.这与已有研究结果一致,在黄土高原,生物土壤结皮POC和MAOC含量随土层增加而降低[23].对毛乌素沙地藻结皮研究发现,其覆盖下SOC含量为裸沙的2.1倍[24].藻结皮发育在干旱区土壤发育和稳定中具有重要的生态功能,一方面,藻结皮作为生物土壤结皮初级演替阶段,其固碳作用主要通过原核生物和光合生物固碳途径完成[25],为有机质的累积提供了基础.另一方面,干旱区土壤普遍缺乏氮素,藻结皮中的固氮蓝藻在生物固氮中发挥重要作用,可为其他植物及土壤微生物供给充足的氮素资源[26],不仅促进了植物生长和土壤微生物活性,还增强了土壤中有机碳输入量,逐步增加生物土壤结皮的稳定性. ...
古尔班通古特沙漠生物土壤结皮下土壤有机碳垂直分布特征及影响因素
1
2024
... 随着土层深度的增加,藻结皮层和0~5 cm土层SOC及其组分含量显著高于下层土壤,表明藻结皮层存在养分聚集效应.藻结皮层具有丰富的藻类,可通过光合作用合成有机物,加之藻结皮的覆盖,可以在一定程度上减少土壤表层养分流失,进而使得表层SOC组分含量高于下层[27].同时,表层SOC增加又为土壤微生物生长提供适宜的条件,导致土壤微生物数量随土层深度增加呈现下降趋势.微生物是SOC形成与周转的主要参与者和调控者,在其增殖、生长直至死亡的生命周期中,会产生大量的细胞残留物及其分解过程中释放的小分子代谢产物,部分微生物残体与土壤矿物质发生紧密结合,促进MAOC的积累[10].这一系列过程共同作用使得藻结皮在固定与储存SOC方面展现出显著的能力,成为干旱区中重要的碳汇. ...
腾格里沙漠南缘植被恢复对土壤有机碳组成及稳定性的影响
1
2024
... 裸沙与藻结皮中MAOC占SOC含量的比例呈现出随深度增加而下降的趋势.具体而言,从表层开始,裸沙MAOC含量占SOC的比例由16.85%降至12.69%,而藻结皮中这一比例则由42.70%降至28.95%.这些观测值显著低于先前研究中报道的比例,即SOC的60%以上被储存在MAOC库中[10].西北荒漠区MAOC整体含量较低,一方面,荒漠区降水稀少使得土壤SWC处于较低水平,进而微生物活动受到限制,导致其对植物残体的分解速度相对较低,土壤有机质周转速度减慢,从而有利于POC含量的积累,导致MAOC占比降低.另一方面,荒漠区土壤中粗砂颗粒含量占据优势,其细砂颗粒含量相对较少,有机物质难与矿物表面充分接触和结合,不利于MAOC的形成[28].由于POC易于被微生物分解的特性以及对环境变化的高度敏感性,通过POC/MAOC的值可以反映出SOC稳定性的相对大小[15].本研究中,藻结皮SOC稳定性高于裸沙,主要原因是藻结皮能够增加土壤水分保持能力,为微生物生长和活动提供更适宜的条件,更有利于MAOC含量积累,进而提高了SOC的稳定性[29]. ...
Biocrusts enhance soil organic carbon stability and regulate the fate of new-input carbon in semiarid desert ecosystems
2
2024
... 裸沙与藻结皮中MAOC占SOC含量的比例呈现出随深度增加而下降的趋势.具体而言,从表层开始,裸沙MAOC含量占SOC的比例由16.85%降至12.69%,而藻结皮中这一比例则由42.70%降至28.95%.这些观测值显著低于先前研究中报道的比例,即SOC的60%以上被储存在MAOC库中[10].西北荒漠区MAOC整体含量较低,一方面,荒漠区降水稀少使得土壤SWC处于较低水平,进而微生物活动受到限制,导致其对植物残体的分解速度相对较低,土壤有机质周转速度减慢,从而有利于POC含量的积累,导致MAOC占比降低.另一方面,荒漠区土壤中粗砂颗粒含量占据优势,其细砂颗粒含量相对较少,有机物质难与矿物表面充分接触和结合,不利于MAOC的形成[28].由于POC易于被微生物分解的特性以及对环境变化的高度敏感性,通过POC/MAOC的值可以反映出SOC稳定性的相对大小[15].本研究中,藻结皮SOC稳定性高于裸沙,主要原因是藻结皮能够增加土壤水分保持能力,为微生物生长和活动提供更适宜的条件,更有利于MAOC含量积累,进而提高了SOC的稳定性[29]. ...
... 土壤理化特征变化会影响土壤微生物的活性以及植物凋落物的输入,这些变化会对SOC含量产生影响[33].本研究结果表明,土壤养分(TN、NH-N和AP等)含量与裸沙和藻结皮POC和MAOC含量呈正相关,这一结果与Sun等[8]对生物土壤结皮斑块碳组分的研究结果相一致.氮作为干旱区的主要限制因素之一,氮富集会增加植物源碳的输入同时为微生物提供氮源,进而减少微生物对有机质的分解,使得POC含量增加[15,29,34].同时,速效养分(NH-N和NO-N)以及磷元素通过共同影响微生物活性(其中磷主要通过与氮的化学计量关系来调节微生物多样性与活性),调节微生物残体和植物凋落物的分解过程,进而促进POC和MAOC的形成[35-36].本研究发现裸沙POC和MAOC含量与pH值负相关,与Zhou等[30]的研究结果一致.pH对微生物影响较大,研究区pH呈碱性,微生物活动受到高pH值抑制后活性降低,进而影响有机质的分解和转化过程,减少MAOC的积累[30];另外,高pH会抑制植物生产力,减少碳输入,影响POC和MAOC含量[37].因此,荒漠区裸沙与藻结皮SOC组分含量分布特征受到多方面因素的综合作用,本研究结果为干旱区土壤碳动态评估提供了理论指导. ...
Global turnover of soil mineral-associated and particulate organic carbon
3
2024
... 在区域尺度下,SOC的变化主要受气候、土壤和植被条件的影响[16].本研究也表明,裸沙和藻结皮SOC组分变化的重要预测变量为MAP和MAT.多数研究认为,生物土壤结皮只有在外界温度和水分条件达到一定程度才能正常生长,其生长发育所需要的水分来自近地表土壤水分,引起微生物的快速响应[24,30],进而影响其SOC含量.在干旱区,降水对SOC的影响主要通过土壤微生物活性而非植被[31].温度升高可能会增加微生物生长和周转,提高微生物代谢活性,从而会加速POC分解和损失,使得SOC含量减少.另外,变暖会引起土壤真菌相对丰度增加,促进MAOC分解,从而导致MAOC储量减少[15].因此,二者与MAT呈负相关关系.小尺度上,一般来说,适宜的SWC有利于SOC积累,但是干旱区SWC比较低,SWC增加使得藻结皮增加植物碳输入,从而减轻微生物抑制,使得POC和MAOC含量增加[32].因此,SWC与POC和MAOC含量呈正相关. ...
... 土壤理化特征变化会影响土壤微生物的活性以及植物凋落物的输入,这些变化会对SOC含量产生影响[33].本研究结果表明,土壤养分(TN、NH-N和AP等)含量与裸沙和藻结皮POC和MAOC含量呈正相关,这一结果与Sun等[8]对生物土壤结皮斑块碳组分的研究结果相一致.氮作为干旱区的主要限制因素之一,氮富集会增加植物源碳的输入同时为微生物提供氮源,进而减少微生物对有机质的分解,使得POC含量增加[15,29,34].同时,速效养分(NH-N和NO-N)以及磷元素通过共同影响微生物活性(其中磷主要通过与氮的化学计量关系来调节微生物多样性与活性),调节微生物残体和植物凋落物的分解过程,进而促进POC和MAOC的形成[35-36].本研究发现裸沙POC和MAOC含量与pH值负相关,与Zhou等[30]的研究结果一致.pH对微生物影响较大,研究区pH呈碱性,微生物活动受到高pH值抑制后活性降低,进而影响有机质的分解和转化过程,减少MAOC的积累[30];另外,高pH会抑制植物生产力,减少碳输入,影响POC和MAOC含量[37].因此,荒漠区裸沙与藻结皮SOC组分含量分布特征受到多方面因素的综合作用,本研究结果为干旱区土壤碳动态评估提供了理论指导. ...
... [30];另外,高pH会抑制植物生产力,减少碳输入,影响POC和MAOC含量[37].因此,荒漠区裸沙与藻结皮SOC组分含量分布特征受到多方面因素的综合作用,本研究结果为干旱区土壤碳动态评估提供了理论指导. ...
Spatiotemporal dynamics of soil organic carbon in desert region and its response to climate change:a case study of Heihe river
1
2025
... 在区域尺度下,SOC的变化主要受气候、土壤和植被条件的影响[16].本研究也表明,裸沙和藻结皮SOC组分变化的重要预测变量为MAP和MAT.多数研究认为,生物土壤结皮只有在外界温度和水分条件达到一定程度才能正常生长,其生长发育所需要的水分来自近地表土壤水分,引起微生物的快速响应[24,30],进而影响其SOC含量.在干旱区,降水对SOC的影响主要通过土壤微生物活性而非植被[31].温度升高可能会增加微生物生长和周转,提高微生物代谢活性,从而会加速POC分解和损失,使得SOC含量减少.另外,变暖会引起土壤真菌相对丰度增加,促进MAOC分解,从而导致MAOC储量减少[15].因此,二者与MAT呈负相关关系.小尺度上,一般来说,适宜的SWC有利于SOC积累,但是干旱区SWC比较低,SWC增加使得藻结皮增加植物碳输入,从而减轻微生物抑制,使得POC和MAOC含量增加[32].因此,SWC与POC和MAOC含量呈正相关. ...
Impacts of altered precipitation regimes on soil communities and biogeochemistry in arid and semi‐arid ecosystems
1
2015
... 在区域尺度下,SOC的变化主要受气候、土壤和植被条件的影响[16].本研究也表明,裸沙和藻结皮SOC组分变化的重要预测变量为MAP和MAT.多数研究认为,生物土壤结皮只有在外界温度和水分条件达到一定程度才能正常生长,其生长发育所需要的水分来自近地表土壤水分,引起微生物的快速响应[24,30],进而影响其SOC含量.在干旱区,降水对SOC的影响主要通过土壤微生物活性而非植被[31].温度升高可能会增加微生物生长和周转,提高微生物代谢活性,从而会加速POC分解和损失,使得SOC含量减少.另外,变暖会引起土壤真菌相对丰度增加,促进MAOC分解,从而导致MAOC储量减少[15].因此,二者与MAT呈负相关关系.小尺度上,一般来说,适宜的SWC有利于SOC积累,但是干旱区SWC比较低,SWC增加使得藻结皮增加植物碳输入,从而减轻微生物抑制,使得POC和MAOC含量增加[32].因此,SWC与POC和MAOC含量呈正相关. ...
生物结皮对土壤养分及碳循环影响的研究进展
1
2023
... 土壤理化特征变化会影响土壤微生物的活性以及植物凋落物的输入,这些变化会对SOC含量产生影响[33].本研究结果表明,土壤养分(TN、NH-N和AP等)含量与裸沙和藻结皮POC和MAOC含量呈正相关,这一结果与Sun等[8]对生物土壤结皮斑块碳组分的研究结果相一致.氮作为干旱区的主要限制因素之一,氮富集会增加植物源碳的输入同时为微生物提供氮源,进而减少微生物对有机质的分解,使得POC含量增加[15,29,34].同时,速效养分(NH-N和NO-N)以及磷元素通过共同影响微生物活性(其中磷主要通过与氮的化学计量关系来调节微生物多样性与活性),调节微生物残体和植物凋落物的分解过程,进而促进POC和MAOC的形成[35-36].本研究发现裸沙POC和MAOC含量与pH值负相关,与Zhou等[30]的研究结果一致.pH对微生物影响较大,研究区pH呈碱性,微生物活动受到高pH值抑制后活性降低,进而影响有机质的分解和转化过程,减少MAOC的积累[30];另外,高pH会抑制植物生产力,减少碳输入,影响POC和MAOC含量[37].因此,荒漠区裸沙与藻结皮SOC组分含量分布特征受到多方面因素的综合作用,本研究结果为干旱区土壤碳动态评估提供了理论指导. ...
Nitrogen input alleviates the priming effects of biochar addition on soil organic carbon decomposition
1
2025
... 土壤理化特征变化会影响土壤微生物的活性以及植物凋落物的输入,这些变化会对SOC含量产生影响[33].本研究结果表明,土壤养分(TN、NH-N和AP等)含量与裸沙和藻结皮POC和MAOC含量呈正相关,这一结果与Sun等[8]对生物土壤结皮斑块碳组分的研究结果相一致.氮作为干旱区的主要限制因素之一,氮富集会增加植物源碳的输入同时为微生物提供氮源,进而减少微生物对有机质的分解,使得POC含量增加[15,29,34].同时,速效养分(NH-N和NO-N)以及磷元素通过共同影响微生物活性(其中磷主要通过与氮的化学计量关系来调节微生物多样性与活性),调节微生物残体和植物凋落物的分解过程,进而促进POC和MAOC的形成[35-36].本研究发现裸沙POC和MAOC含量与pH值负相关,与Zhou等[30]的研究结果一致.pH对微生物影响较大,研究区pH呈碱性,微生物活动受到高pH值抑制后活性降低,进而影响有机质的分解和转化过程,减少MAOC的积累[30];另外,高pH会抑制植物生产力,减少碳输入,影响POC和MAOC含量[37].因此,荒漠区裸沙与藻结皮SOC组分含量分布特征受到多方面因素的综合作用,本研究结果为干旱区土壤碳动态评估提供了理论指导. ...
Latitudinal patterns and drivers of plant lignin and microbial necromass accumulation in forest soils:disentangling microbial and abiotic controls
1
2024
... 土壤理化特征变化会影响土壤微生物的活性以及植物凋落物的输入,这些变化会对SOC含量产生影响[33].本研究结果表明,土壤养分(TN、NH-N和AP等)含量与裸沙和藻结皮POC和MAOC含量呈正相关,这一结果与Sun等[8]对生物土壤结皮斑块碳组分的研究结果相一致.氮作为干旱区的主要限制因素之一,氮富集会增加植物源碳的输入同时为微生物提供氮源,进而减少微生物对有机质的分解,使得POC含量增加[15,29,34].同时,速效养分(NH-N和NO-N)以及磷元素通过共同影响微生物活性(其中磷主要通过与氮的化学计量关系来调节微生物多样性与活性),调节微生物残体和植物凋落物的分解过程,进而促进POC和MAOC的形成[35-36].本研究发现裸沙POC和MAOC含量与pH值负相关,与Zhou等[30]的研究结果一致.pH对微生物影响较大,研究区pH呈碱性,微生物活动受到高pH值抑制后活性降低,进而影响有机质的分解和转化过程,减少MAOC的积累[30];另外,高pH会抑制植物生产力,减少碳输入,影响POC和MAOC含量[37].因此,荒漠区裸沙与藻结皮SOC组分含量分布特征受到多方面因素的综合作用,本研究结果为干旱区土壤碳动态评估提供了理论指导. ...
The amounts and ratio of nitrogen and phosphorus addition drive the rate of litter decomposition in a subtropical forest
1
2022
... 土壤理化特征变化会影响土壤微生物的活性以及植物凋落物的输入,这些变化会对SOC含量产生影响[33].本研究结果表明,土壤养分(TN、NH-N和AP等)含量与裸沙和藻结皮POC和MAOC含量呈正相关,这一结果与Sun等[8]对生物土壤结皮斑块碳组分的研究结果相一致.氮作为干旱区的主要限制因素之一,氮富集会增加植物源碳的输入同时为微生物提供氮源,进而减少微生物对有机质的分解,使得POC含量增加[15,29,34].同时,速效养分(NH-N和NO-N)以及磷元素通过共同影响微生物活性(其中磷主要通过与氮的化学计量关系来调节微生物多样性与活性),调节微生物残体和植物凋落物的分解过程,进而促进POC和MAOC的形成[35-36].本研究发现裸沙POC和MAOC含量与pH值负相关,与Zhou等[30]的研究结果一致.pH对微生物影响较大,研究区pH呈碱性,微生物活动受到高pH值抑制后活性降低,进而影响有机质的分解和转化过程,减少MAOC的积累[30];另外,高pH会抑制植物生产力,减少碳输入,影响POC和MAOC含量[37].因此,荒漠区裸沙与藻结皮SOC组分含量分布特征受到多方面因素的综合作用,本研究结果为干旱区土壤碳动态评估提供了理论指导. ...
Soil organic matter priming:the pH effects
1
2024
... 土壤理化特征变化会影响土壤微生物的活性以及植物凋落物的输入,这些变化会对SOC含量产生影响[33].本研究结果表明,土壤养分(TN、NH-N和AP等)含量与裸沙和藻结皮POC和MAOC含量呈正相关,这一结果与Sun等[8]对生物土壤结皮斑块碳组分的研究结果相一致.氮作为干旱区的主要限制因素之一,氮富集会增加植物源碳的输入同时为微生物提供氮源,进而减少微生物对有机质的分解,使得POC含量增加[15,29,34].同时,速效养分(NH-N和NO-N)以及磷元素通过共同影响微生物活性(其中磷主要通过与氮的化学计量关系来调节微生物多样性与活性),调节微生物残体和植物凋落物的分解过程,进而促进POC和MAOC的形成[35-36].本研究发现裸沙POC和MAOC含量与pH值负相关,与Zhou等[30]的研究结果一致.pH对微生物影响较大,研究区pH呈碱性,微生物活动受到高pH值抑制后活性降低,进而影响有机质的分解和转化过程,减少MAOC的积累[30];另外,高pH会抑制植物生产力,减少碳输入,影响POC和MAOC含量[37].因此,荒漠区裸沙与藻结皮SOC组分含量分布特征受到多方面因素的综合作用,本研究结果为干旱区土壤碳动态评估提供了理论指导. ...




 甘公网安备 62010202000688号
甘公网安备 62010202000688号
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}