巴丹吉林沙漠边缘地带植物区系及其资源利用
1
2016
... 荒漠植物是荒漠生态系统的核心,维持着荒漠生态系统营养物质循环和能量流动[1],增加地表不可蚀面积,保护地表免遭风蚀,同时增大地表粗糙度,分解风力,降低风速,改变风沙流结构,减小输沙率,截留沙物质,减缓绿洲流沙入侵及沙漠扩张[2-6].沙丘与丘间地交错分布是沙漠主要景观表现[7],荒漠植物主要分布在生境条件较好的丘间地[8].沿主导风向移动的沙丘会将位于背风坡坡脚的荒漠植物埋压在其底部,切断植物与外界的水、热、气交换,破坏植物生境,导致植物死亡,直至其移动过境后植物枯落物会在迎风坡坡脚暴露于空气和光照环境. ...
1
1980
... 荒漠植物是荒漠生态系统的核心,维持着荒漠生态系统营养物质循环和能量流动[1],增加地表不可蚀面积,保护地表免遭风蚀,同时增大地表粗糙度,分解风力,降低风速,改变风沙流结构,减小输沙率,截留沙物质,减缓绿洲流沙入侵及沙漠扩张[2-6].沙丘与丘间地交错分布是沙漠主要景观表现[7],荒漠植物主要分布在生境条件较好的丘间地[8].沿主导风向移动的沙丘会将位于背风坡坡脚的荒漠植物埋压在其底部,切断植物与外界的水、热、气交换,破坏植物生境,导致植物死亡,直至其移动过境后植物枯落物会在迎风坡坡脚暴露于空气和光照环境. ...
The morphology and origin of Nabkhas,region of Mopti,Mali,West Africa
1994
Influence of vegetation cover on sand transport by wind: field studies at Owens Lake,California
2015
The effect of single vegetation elements on wind speed and sediment transport in the Sahelian zone of Burkina Faso
2007
The protective effects of nebkhas on an oasis
1
2016
... 荒漠植物是荒漠生态系统的核心,维持着荒漠生态系统营养物质循环和能量流动[1],增加地表不可蚀面积,保护地表免遭风蚀,同时增大地表粗糙度,分解风力,降低风速,改变风沙流结构,减小输沙率,截留沙物质,减缓绿洲流沙入侵及沙漠扩张[2-6].沙丘与丘间地交错分布是沙漠主要景观表现[7],荒漠植物主要分布在生境条件较好的丘间地[8].沿主导风向移动的沙丘会将位于背风坡坡脚的荒漠植物埋压在其底部,切断植物与外界的水、热、气交换,破坏植物生境,导致植物死亡,直至其移动过境后植物枯落物会在迎风坡坡脚暴露于空气和光照环境. ...
World distribution of arid and semi-arid homoclimates
1
1953
... 荒漠植物是荒漠生态系统的核心,维持着荒漠生态系统营养物质循环和能量流动[1],增加地表不可蚀面积,保护地表免遭风蚀,同时增大地表粗糙度,分解风力,降低风速,改变风沙流结构,减小输沙率,截留沙物质,减缓绿洲流沙入侵及沙漠扩张[2-6].沙丘与丘间地交错分布是沙漠主要景观表现[7],荒漠植物主要分布在生境条件较好的丘间地[8].沿主导风向移动的沙丘会将位于背风坡坡脚的荒漠植物埋压在其底部,切断植物与外界的水、热、气交换,破坏植物生境,导致植物死亡,直至其移动过境后植物枯落物会在迎风坡坡脚暴露于空气和光照环境. ...
1
1996
... 荒漠植物是荒漠生态系统的核心,维持着荒漠生态系统营养物质循环和能量流动[1],增加地表不可蚀面积,保护地表免遭风蚀,同时增大地表粗糙度,分解风力,降低风速,改变风沙流结构,减小输沙率,截留沙物质,减缓绿洲流沙入侵及沙漠扩张[2-6].沙丘与丘间地交错分布是沙漠主要景观表现[7],荒漠植物主要分布在生境条件较好的丘间地[8].沿主导风向移动的沙丘会将位于背风坡坡脚的荒漠植物埋压在其底部,切断植物与外界的水、热、气交换,破坏植物生境,导致植物死亡,直至其移动过境后植物枯落物会在迎风坡坡脚暴露于空气和光照环境. ...
森林凋落物研究开展
3
2017
... 植物枯落物分解能从植物中释放各种元素至周围环境,在干旱地区维持生态系统功能、物质循环和能量流动等方面具有重要作用[9].以往关于枯落物分解的研究大多在森林[9-13]、草地[14-18]和沙地[19-22],深入探讨植物枯落物的分解速率及其影响因素[9-24].环境条件恶劣,荒漠植被群落结构简单,微生物活性低,干旱区枯落物分解易受光降解影响[25-26],目前对沙丘埋压后植物枯落物在无光照和水热交换条件下的分解情况知之甚少.鉴于此,本文系统研究了沙丘对荒漠植物的埋压时长与沙丘形态的关系,分析了随沙丘埋压时长植物枝条枯落物在分解过程中质量损失、C和N元素等其他养分含量的动态变化,探讨干旱风沙环境对植物枯落物分解的影响,对准确预测荒漠植物碳氮循环具有重要意义. ...
... [9-13]、草地[14-18]和沙地[19-22],深入探讨植物枯落物的分解速率及其影响因素[9-24].环境条件恶劣,荒漠植被群落结构简单,微生物活性低,干旱区枯落物分解易受光降解影响[25-26],目前对沙丘埋压后植物枯落物在无光照和水热交换条件下的分解情况知之甚少.鉴于此,本文系统研究了沙丘对荒漠植物的埋压时长与沙丘形态的关系,分析了随沙丘埋压时长植物枝条枯落物在分解过程中质量损失、C和N元素等其他养分含量的动态变化,探讨干旱风沙环境对植物枯落物分解的影响,对准确预测荒漠植物碳氮循环具有重要意义. ...
... [9-24].环境条件恶劣,荒漠植被群落结构简单,微生物活性低,干旱区枯落物分解易受光降解影响[25-26],目前对沙丘埋压后植物枯落物在无光照和水热交换条件下的分解情况知之甚少.鉴于此,本文系统研究了沙丘对荒漠植物的埋压时长与沙丘形态的关系,分析了随沙丘埋压时长植物枝条枯落物在分解过程中质量损失、C和N元素等其他养分含量的动态变化,探讨干旱风沙环境对植物枯落物分解的影响,对准确预测荒漠植物碳氮循环具有重要意义. ...
土壤微生物群落对麻栎-刺槐混交林凋落物分解的影响
2021
呼伦贝尔沙地樟子松天然林结构多样性和竞争对树木生长的影响
2024
Inorganic fertilizer enrichment of soil: effect on decomposition of plant litter under subhumid tropical conditions
1999
川西亚高山森林凋落物不同分解阶段基质质量特征
2
2017
... 植物枯落物分解能从植物中释放各种元素至周围环境,在干旱地区维持生态系统功能、物质循环和能量流动等方面具有重要作用[9].以往关于枯落物分解的研究大多在森林[9-13]、草地[14-18]和沙地[19-22],深入探讨植物枯落物的分解速率及其影响因素[9-24].环境条件恶劣,荒漠植被群落结构简单,微生物活性低,干旱区枯落物分解易受光降解影响[25-26],目前对沙丘埋压后植物枯落物在无光照和水热交换条件下的分解情况知之甚少.鉴于此,本文系统研究了沙丘对荒漠植物的埋压时长与沙丘形态的关系,分析了随沙丘埋压时长植物枝条枯落物在分解过程中质量损失、C和N元素等其他养分含量的动态变化,探讨干旱风沙环境对植物枯落物分解的影响,对准确预测荒漠植物碳氮循环具有重要意义. ...
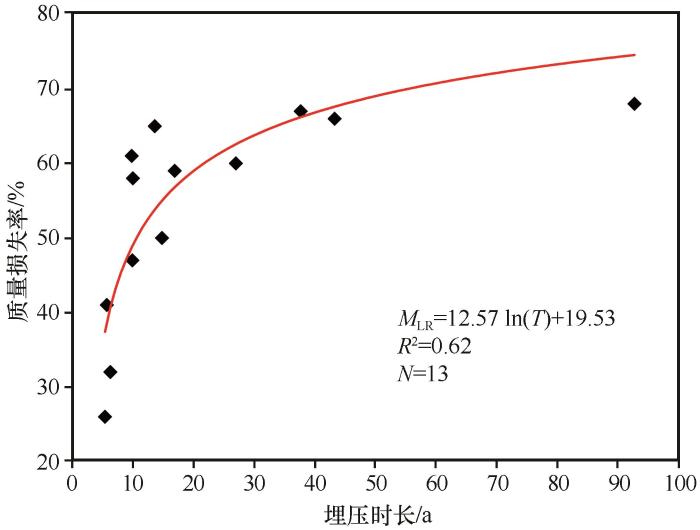
... 枯落物的分解是缓慢的过程,分解初期速率较快,后期逐渐平缓[25-26].本研究发现白刺枝条在沙丘埋压情况下,其枯落物质量损失在被埋压的前25年较大,之后趋于平缓,这与王嘉年等[25]和范琳杰等[26]对干旱区荒漠植物分解速率随时间变化规律的研究结果一致,但时间转折点明显晚于其研究结果.这是由于枯落物分解前期主要与非生物因素(如物理破碎等)有关,枯落物中水溶性化合物先分解,在分解过程中像木质素等难以分解的物质累积,导致枯落物分解速率逐渐减缓,此时,枯落物的分解主要由生物因素主导[13,25,36-37],而分解速率快慢转折点出现的时间较晚的原因可能一方面是由于本研究分析的是荒漠植物枝条的分解速率,而其他研究关注的是荒漠植物的叶片,组成物质的差异造成枝条分解速率原本要远低于叶片,另一方面是被沙丘埋压的条件下植物的分解速率要低于自然条件. ...
草地枯落物分解研究进展及展望
1
2010
... 植物枯落物分解能从植物中释放各种元素至周围环境,在干旱地区维持生态系统功能、物质循环和能量流动等方面具有重要作用[9].以往关于枯落物分解的研究大多在森林[9-13]、草地[14-18]和沙地[19-22],深入探讨植物枯落物的分解速率及其影响因素[9-24].环境条件恶劣,荒漠植被群落结构简单,微生物活性低,干旱区枯落物分解易受光降解影响[25-26],目前对沙丘埋压后植物枯落物在无光照和水热交换条件下的分解情况知之甚少.鉴于此,本文系统研究了沙丘对荒漠植物的埋压时长与沙丘形态的关系,分析了随沙丘埋压时长植物枝条枯落物在分解过程中质量损失、C和N元素等其他养分含量的动态变化,探讨干旱风沙环境对植物枯落物分解的影响,对准确预测荒漠植物碳氮循环具有重要意义. ...
青藏高原不同气候区高寒沙地两种优势植物及其根际土壤的养分特征
2018
Litter decomposition and nutrient release as affected by soil nitrogen availability and litter quality in a semiarid grassland ecosystem
2010
宁夏荒漠草原单种与混合植物枯落物分解过程及土壤动物的贡献
2019
松嫩草地不同放牧强度下植物物种枯落物分解的变化规律研究
1
2013
... 植物枯落物分解能从植物中释放各种元素至周围环境,在干旱地区维持生态系统功能、物质循环和能量流动等方面具有重要作用[9].以往关于枯落物分解的研究大多在森林[9-13]、草地[14-18]和沙地[19-22],深入探讨植物枯落物的分解速率及其影响因素[9-24].环境条件恶劣,荒漠植被群落结构简单,微生物活性低,干旱区枯落物分解易受光降解影响[25-26],目前对沙丘埋压后植物枯落物在无光照和水热交换条件下的分解情况知之甚少.鉴于此,本文系统研究了沙丘对荒漠植物的埋压时长与沙丘形态的关系,分析了随沙丘埋压时长植物枝条枯落物在分解过程中质量损失、C和N元素等其他养分含量的动态变化,探讨干旱风沙环境对植物枯落物分解的影响,对准确预测荒漠植物碳氮循环具有重要意义. ...
凋落物多样性及组成对凋落物分解和土壤微生物群落的影响
1
2019
... 植物枯落物分解能从植物中释放各种元素至周围环境,在干旱地区维持生态系统功能、物质循环和能量流动等方面具有重要作用[9].以往关于枯落物分解的研究大多在森林[9-13]、草地[14-18]和沙地[19-22],深入探讨植物枯落物的分解速率及其影响因素[9-24].环境条件恶劣,荒漠植被群落结构简单,微生物活性低,干旱区枯落物分解易受光降解影响[25-26],目前对沙丘埋压后植物枯落物在无光照和水热交换条件下的分解情况知之甚少.鉴于此,本文系统研究了沙丘对荒漠植物的埋压时长与沙丘形态的关系,分析了随沙丘埋压时长植物枝条枯落物在分解过程中质量损失、C和N元素等其他养分含量的动态变化,探讨干旱风沙环境对植物枯落物分解的影响,对准确预测荒漠植物碳氮循环具有重要意义. ...
环境因素对干旱半干旱区凋落物分解的影响研究进展
2013
降水格局对科尔沁沙地三种草本植物凋落物分解的影响
2018
Resolving the dryland decomposition conundrum:some new perspectives on potential drivers
1
2009
... 植物枯落物分解能从植物中释放各种元素至周围环境,在干旱地区维持生态系统功能、物质循环和能量流动等方面具有重要作用[9].以往关于枯落物分解的研究大多在森林[9-13]、草地[14-18]和沙地[19-22],深入探讨植物枯落物的分解速率及其影响因素[9-24].环境条件恶劣,荒漠植被群落结构简单,微生物活性低,干旱区枯落物分解易受光降解影响[25-26],目前对沙丘埋压后植物枯落物在无光照和水热交换条件下的分解情况知之甚少.鉴于此,本文系统研究了沙丘对荒漠植物的埋压时长与沙丘形态的关系,分析了随沙丘埋压时长植物枝条枯落物在分解过程中质量损失、C和N元素等其他养分含量的动态变化,探讨干旱风沙环境对植物枯落物分解的影响,对准确预测荒漠植物碳氮循环具有重要意义. ...
Litter complexity and composition are determinants of the diversity and species composition of oribatid mites in litter bags
1
1998
... 植物枯落物分解能从植物中释放各种元素至周围环境,在干旱地区维持生态系统功能、物质循环和能量流动等方面具有重要作用[9].以往关于枯落物分解的研究大多在森林[9-13]、草地[14-18]和沙地[19-22],深入探讨植物枯落物的分解速率及其影响因素[9-24].环境条件恶劣,荒漠植被群落结构简单,微生物活性低,干旱区枯落物分解易受光降解影响[25-26],目前对沙丘埋压后植物枯落物在无光照和水热交换条件下的分解情况知之甚少.鉴于此,本文系统研究了沙丘对荒漠植物的埋压时长与沙丘形态的关系,分析了随沙丘埋压时长植物枝条枯落物在分解过程中质量损失、C和N元素等其他养分含量的动态变化,探讨干旱风沙环境对植物枯落物分解的影响,对准确预测荒漠植物碳氮循环具有重要意义. ...
自然光照和荫蔽条件下两种荒漠植物叶片凋落物分解特征研究
6
2023
... 植物枯落物分解能从植物中释放各种元素至周围环境,在干旱地区维持生态系统功能、物质循环和能量流动等方面具有重要作用[9].以往关于枯落物分解的研究大多在森林[9-13]、草地[14-18]和沙地[19-22],深入探讨植物枯落物的分解速率及其影响因素[9-24].环境条件恶劣,荒漠植被群落结构简单,微生物活性低,干旱区枯落物分解易受光降解影响[25-26],目前对沙丘埋压后植物枯落物在无光照和水热交换条件下的分解情况知之甚少.鉴于此,本文系统研究了沙丘对荒漠植物的埋压时长与沙丘形态的关系,分析了随沙丘埋压时长植物枝条枯落物在分解过程中质量损失、C和N元素等其他养分含量的动态变化,探讨干旱风沙环境对植物枯落物分解的影响,对准确预测荒漠植物碳氮循环具有重要意义. ...
... 被沙丘埋压的白刺枝条枯落物质量损失率与埋压时长间呈对数正相关,说明在被沙丘埋压的极端环境下,荒漠植物枝条枯落物的分解依然进行;但分解速率与时间之间的关系模型与在草原和干旱区正常环境中植物叶片分解速率与时间之间的单指数模型、双指数模型和渐近模型有差异[25,33],说明被沙丘埋压后枝条的分解速率要明显低于草原,也低于干旱区自然光照和荫蔽条件下枯落物分解速率.这可能是由于植物分解过程与环境因子密切相关[34],被沙丘埋压后隔绝了植物枝条与外界环境的水热交换和光照等条件,致使植物枯落物在正常环境下分解过程的某些环节减弱或者不能正常进行;同时,本研究探讨的是植物枝条枯落物的分解速率而其他研究分析的是植物的叶片,由于不同植物器官元素组成的较大差异[35],枝条本身比叶片更难分解,分解速率要更慢. ...
... 枯落物的分解是缓慢的过程,分解初期速率较快,后期逐渐平缓[25-26].本研究发现白刺枝条在沙丘埋压情况下,其枯落物质量损失在被埋压的前25年较大,之后趋于平缓,这与王嘉年等[25]和范琳杰等[26]对干旱区荒漠植物分解速率随时间变化规律的研究结果一致,但时间转折点明显晚于其研究结果.这是由于枯落物分解前期主要与非生物因素(如物理破碎等)有关,枯落物中水溶性化合物先分解,在分解过程中像木质素等难以分解的物质累积,导致枯落物分解速率逐渐减缓,此时,枯落物的分解主要由生物因素主导[13,25,36-37],而分解速率快慢转折点出现的时间较晚的原因可能一方面是由于本研究分析的是荒漠植物枝条的分解速率,而其他研究关注的是荒漠植物的叶片,组成物质的差异造成枝条分解速率原本要远低于叶片,另一方面是被沙丘埋压的条件下植物的分解速率要低于自然条件. ...
... [25]和范琳杰等[26]对干旱区荒漠植物分解速率随时间变化规律的研究结果一致,但时间转折点明显晚于其研究结果.这是由于枯落物分解前期主要与非生物因素(如物理破碎等)有关,枯落物中水溶性化合物先分解,在分解过程中像木质素等难以分解的物质累积,导致枯落物分解速率逐渐减缓,此时,枯落物的分解主要由生物因素主导[13,25,36-37],而分解速率快慢转折点出现的时间较晚的原因可能一方面是由于本研究分析的是荒漠植物枝条的分解速率,而其他研究关注的是荒漠植物的叶片,组成物质的差异造成枝条分解速率原本要远低于叶片,另一方面是被沙丘埋压的条件下植物的分解速率要低于自然条件. ...
... ,25,36-37],而分解速率快慢转折点出现的时间较晚的原因可能一方面是由于本研究分析的是荒漠植物枝条的分解速率,而其他研究关注的是荒漠植物的叶片,组成物质的差异造成枝条分解速率原本要远低于叶片,另一方面是被沙丘埋压的条件下植物的分解速率要低于自然条件. ...
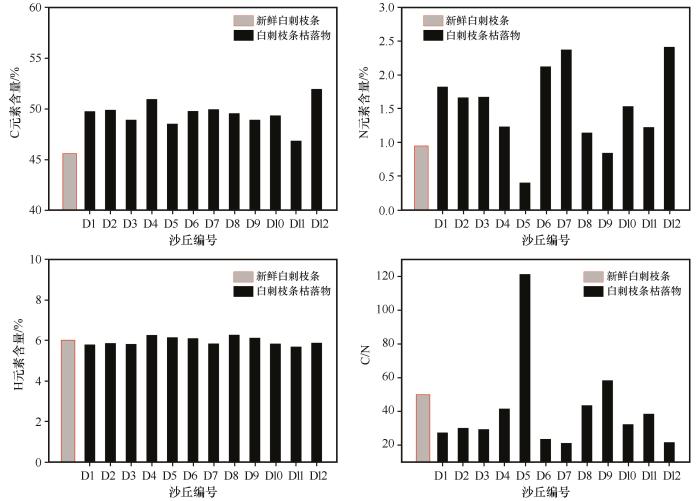
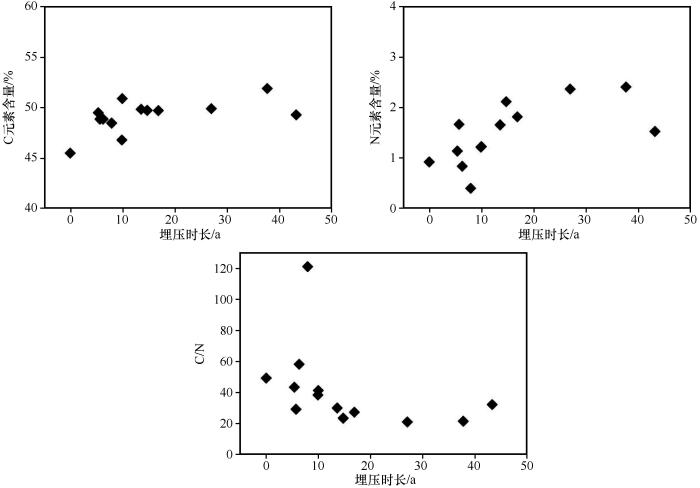
... 碳是构成植物枯落物有机质的主要组成元素,在其分解过程中,只有20%的C元素留存在土壤有机质中[45],枯落物分解过程中C元素的释放速度影响全球生态系统碳平衡[46],而C元素在枯落物分解过程中并不总被释放出来,其与枯落物自身特性、所在环境以及分解时间有关[47].本研究发现,被沙丘埋压后,植物枝条枯落物C和N的含量升高,C/N比在降低,这与孙本华等[48]在荒漠条件下对大豆秆的分解过程中C和N及C/N的变化规律的研究结果一致.主要是由于大规模的沙埋对植物有一定的压实作用,可能会改变植物内部结构,影响植物分解速率;另外植物在被沙丘埋压后与空气隔绝,仅依靠自身含氧发生厌氧细菌的分解作用,从而开始脱水、去羧基,减少腐殖酸等物质,使植物中C元素含量相对增高,而氧含量相对减少,存在碳化过程.而含N元素物质基本未参与或者参与较少分解反应,在分解过程中其相对含量增加比C多,故C/N比降低.随着埋压时间增加,氧气不充足条件下植物的分解反应继续进行,C和N相对含量越来越高,而C/N比越来越低.因此,被沙丘埋压后植物枯落物分解是C和N的富集过程,这与王嘉年等[25]在干旱区对枯落物的分解做遮光处理后C和N含量的变化结果一致,间接地反映出在干旱生态系统中光降解在C周转中的重要作用.但由于本研究只在沙丘附近非丘间地区域分别采集了1株生长旺盛的白刺和1株小叶锦鸡儿植株,对于C和N元素的富集度的精度有一定影响,在以后研究中需加大对鲜活植株的采样和测量.同时,被沙丘埋压不同阶段枯落物的物质组成以及导致C、N元素富集的具体原因亟待进一步探究. ...
极端干旱区花花柴(Karelinia caspia)和胡杨(Populus euphratica)叶凋落物分解和养分释放特征
3
2021
... 植物枯落物分解能从植物中释放各种元素至周围环境,在干旱地区维持生态系统功能、物质循环和能量流动等方面具有重要作用[9].以往关于枯落物分解的研究大多在森林[9-13]、草地[14-18]和沙地[19-22],深入探讨植物枯落物的分解速率及其影响因素[9-24].环境条件恶劣,荒漠植被群落结构简单,微生物活性低,干旱区枯落物分解易受光降解影响[25-26],目前对沙丘埋压后植物枯落物在无光照和水热交换条件下的分解情况知之甚少.鉴于此,本文系统研究了沙丘对荒漠植物的埋压时长与沙丘形态的关系,分析了随沙丘埋压时长植物枝条枯落物在分解过程中质量损失、C和N元素等其他养分含量的动态变化,探讨干旱风沙环境对植物枯落物分解的影响,对准确预测荒漠植物碳氮循环具有重要意义. ...
... 枯落物的分解是缓慢的过程,分解初期速率较快,后期逐渐平缓[25-26].本研究发现白刺枝条在沙丘埋压情况下,其枯落物质量损失在被埋压的前25年较大,之后趋于平缓,这与王嘉年等[25]和范琳杰等[26]对干旱区荒漠植物分解速率随时间变化规律的研究结果一致,但时间转折点明显晚于其研究结果.这是由于枯落物分解前期主要与非生物因素(如物理破碎等)有关,枯落物中水溶性化合物先分解,在分解过程中像木质素等难以分解的物质累积,导致枯落物分解速率逐渐减缓,此时,枯落物的分解主要由生物因素主导[13,25,36-37],而分解速率快慢转折点出现的时间较晚的原因可能一方面是由于本研究分析的是荒漠植物枝条的分解速率,而其他研究关注的是荒漠植物的叶片,组成物质的差异造成枝条分解速率原本要远低于叶片,另一方面是被沙丘埋压的条件下植物的分解速率要低于自然条件. ...
... [26]对干旱区荒漠植物分解速率随时间变化规律的研究结果一致,但时间转折点明显晚于其研究结果.这是由于枯落物分解前期主要与非生物因素(如物理破碎等)有关,枯落物中水溶性化合物先分解,在分解过程中像木质素等难以分解的物质累积,导致枯落物分解速率逐渐减缓,此时,枯落物的分解主要由生物因素主导[13,25,36-37],而分解速率快慢转折点出现的时间较晚的原因可能一方面是由于本研究分析的是荒漠植物枝条的分解速率,而其他研究关注的是荒漠植物的叶片,组成物质的差异造成枝条分解速率原本要远低于叶片,另一方面是被沙丘埋压的条件下植物的分解速率要低于自然条件. ...
基于遥感的巴丹吉林沙漠范围与面积分析
1
2010
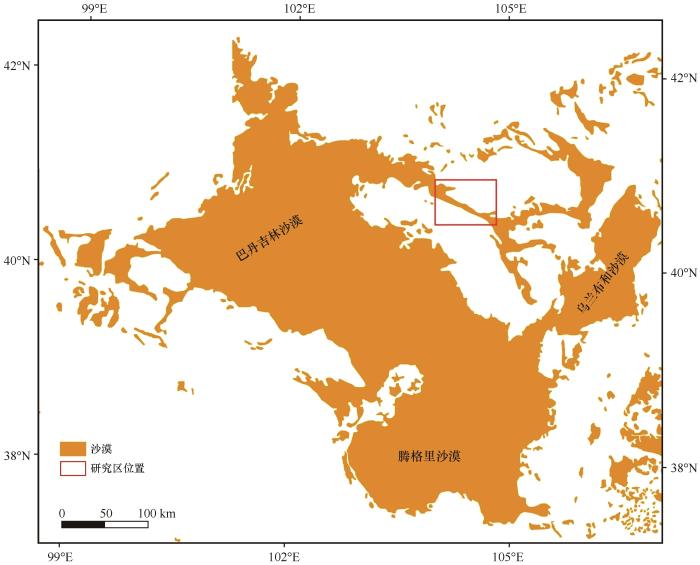
... 巴丹吉林沙漠位于阿拉善高原的中心,是中国第二大流动沙漠[27].巴丹吉林沙漠-乌兰布和沙漠输沙带(39°30′—41°00′N,104°00′—106°00′E)[28]从巴丹吉林沙漠东南部马拉盖山南侧开始延伸至乌兰布和沙漠北部,长约250 km,宽5~45 km[28-29](图1).年平均气温为10 ℃,多年平均降水量为150 mm,全年降水集中在6—9月(占全年降水量的70%)[29],年平均大风日数大于50 d,年平均风速为3.7 m·s-1,主导风向为W、WNW和NW[29-30].多年平均潜在蒸散量为1 447 mm,干燥指数为0.07,属于典型干旱荒漠区[31].植被组成以旱生超旱生和盐生灌木、小灌木和小半灌木为主,主要有霸王(Zygophyllum xanthoxylon)、白刺(Nitraria tangutorum)、小叶锦鸡儿(Caragana microphylla)、珍珠(Salsola passerina)、红砂(Reaumuria soongorica)和猫头刺(Oxytropis aciphylla)等[32].输沙带沙丘以新月形沙丘和沙丘链为主.新月形沙丘的高度为3~20 m,沙丘链长约200~5 000 km,高度为5~15 m[28].新月形沙丘和沙丘链的丘间地存在大量白刺灌丛沙堆和锦鸡儿灌丛沙堆[28].地带性土壤为灰漠土,砾石、粗砂含量高,丘间地有薄层覆沙[29]. ...
阿拉善高原风沙地貌过程研究
4
2015
... 巴丹吉林沙漠位于阿拉善高原的中心,是中国第二大流动沙漠[27].巴丹吉林沙漠-乌兰布和沙漠输沙带(39°30′—41°00′N,104°00′—106°00′E)[28]从巴丹吉林沙漠东南部马拉盖山南侧开始延伸至乌兰布和沙漠北部,长约250 km,宽5~45 km[28-29](图1).年平均气温为10 ℃,多年平均降水量为150 mm,全年降水集中在6—9月(占全年降水量的70%)[29],年平均大风日数大于50 d,年平均风速为3.7 m·s-1,主导风向为W、WNW和NW[29-30].多年平均潜在蒸散量为1 447 mm,干燥指数为0.07,属于典型干旱荒漠区[31].植被组成以旱生超旱生和盐生灌木、小灌木和小半灌木为主,主要有霸王(Zygophyllum xanthoxylon)、白刺(Nitraria tangutorum)、小叶锦鸡儿(Caragana microphylla)、珍珠(Salsola passerina)、红砂(Reaumuria soongorica)和猫头刺(Oxytropis aciphylla)等[32].输沙带沙丘以新月形沙丘和沙丘链为主.新月形沙丘的高度为3~20 m,沙丘链长约200~5 000 km,高度为5~15 m[28].新月形沙丘和沙丘链的丘间地存在大量白刺灌丛沙堆和锦鸡儿灌丛沙堆[28].地带性土壤为灰漠土,砾石、粗砂含量高,丘间地有薄层覆沙[29]. ...
... [28-29](图1).年平均气温为10 ℃,多年平均降水量为150 mm,全年降水集中在6—9月(占全年降水量的70%)[29],年平均大风日数大于50 d,年平均风速为3.7 m·s-1,主导风向为W、WNW和NW[29-30].多年平均潜在蒸散量为1 447 mm,干燥指数为0.07,属于典型干旱荒漠区[31].植被组成以旱生超旱生和盐生灌木、小灌木和小半灌木为主,主要有霸王(Zygophyllum xanthoxylon)、白刺(Nitraria tangutorum)、小叶锦鸡儿(Caragana microphylla)、珍珠(Salsola passerina)、红砂(Reaumuria soongorica)和猫头刺(Oxytropis aciphylla)等[32].输沙带沙丘以新月形沙丘和沙丘链为主.新月形沙丘的高度为3~20 m,沙丘链长约200~5 000 km,高度为5~15 m[28].新月形沙丘和沙丘链的丘间地存在大量白刺灌丛沙堆和锦鸡儿灌丛沙堆[28].地带性土壤为灰漠土,砾石、粗砂含量高,丘间地有薄层覆沙[29]. ...
... [28].新月形沙丘和沙丘链的丘间地存在大量白刺灌丛沙堆和锦鸡儿灌丛沙堆[28].地带性土壤为灰漠土,砾石、粗砂含量高,丘间地有薄层覆沙[29]. ...
... [28].地带性土壤为灰漠土,砾石、粗砂含量高,丘间地有薄层覆沙[29]. ...
巴丹吉林-乌兰布和沙漠输沙带新月形沙丘动态
5
2024
... 巴丹吉林沙漠位于阿拉善高原的中心,是中国第二大流动沙漠[27].巴丹吉林沙漠-乌兰布和沙漠输沙带(39°30′—41°00′N,104°00′—106°00′E)[28]从巴丹吉林沙漠东南部马拉盖山南侧开始延伸至乌兰布和沙漠北部,长约250 km,宽5~45 km[28-29](图1).年平均气温为10 ℃,多年平均降水量为150 mm,全年降水集中在6—9月(占全年降水量的70%)[29],年平均大风日数大于50 d,年平均风速为3.7 m·s-1,主导风向为W、WNW和NW[29-30].多年平均潜在蒸散量为1 447 mm,干燥指数为0.07,属于典型干旱荒漠区[31].植被组成以旱生超旱生和盐生灌木、小灌木和小半灌木为主,主要有霸王(Zygophyllum xanthoxylon)、白刺(Nitraria tangutorum)、小叶锦鸡儿(Caragana microphylla)、珍珠(Salsola passerina)、红砂(Reaumuria soongorica)和猫头刺(Oxytropis aciphylla)等[32].输沙带沙丘以新月形沙丘和沙丘链为主.新月形沙丘的高度为3~20 m,沙丘链长约200~5 000 km,高度为5~15 m[28].新月形沙丘和沙丘链的丘间地存在大量白刺灌丛沙堆和锦鸡儿灌丛沙堆[28].地带性土壤为灰漠土,砾石、粗砂含量高,丘间地有薄层覆沙[29]. ...
... [29],年平均大风日数大于50 d,年平均风速为3.7 m·s-1,主导风向为W、WNW和NW[29-30].多年平均潜在蒸散量为1 447 mm,干燥指数为0.07,属于典型干旱荒漠区[31].植被组成以旱生超旱生和盐生灌木、小灌木和小半灌木为主,主要有霸王(Zygophyllum xanthoxylon)、白刺(Nitraria tangutorum)、小叶锦鸡儿(Caragana microphylla)、珍珠(Salsola passerina)、红砂(Reaumuria soongorica)和猫头刺(Oxytropis aciphylla)等[32].输沙带沙丘以新月形沙丘和沙丘链为主.新月形沙丘的高度为3~20 m,沙丘链长约200~5 000 km,高度为5~15 m[28].新月形沙丘和沙丘链的丘间地存在大量白刺灌丛沙堆和锦鸡儿灌丛沙堆[28].地带性土壤为灰漠土,砾石、粗砂含量高,丘间地有薄层覆沙[29]. ...
... [29-30].多年平均潜在蒸散量为1 447 mm,干燥指数为0.07,属于典型干旱荒漠区[31].植被组成以旱生超旱生和盐生灌木、小灌木和小半灌木为主,主要有霸王(Zygophyllum xanthoxylon)、白刺(Nitraria tangutorum)、小叶锦鸡儿(Caragana microphylla)、珍珠(Salsola passerina)、红砂(Reaumuria soongorica)和猫头刺(Oxytropis aciphylla)等[32].输沙带沙丘以新月形沙丘和沙丘链为主.新月形沙丘的高度为3~20 m,沙丘链长约200~5 000 km,高度为5~15 m[28].新月形沙丘和沙丘链的丘间地存在大量白刺灌丛沙堆和锦鸡儿灌丛沙堆[28].地带性土壤为灰漠土,砾石、粗砂含量高,丘间地有薄层覆沙[29]. ...
... [29]. ...
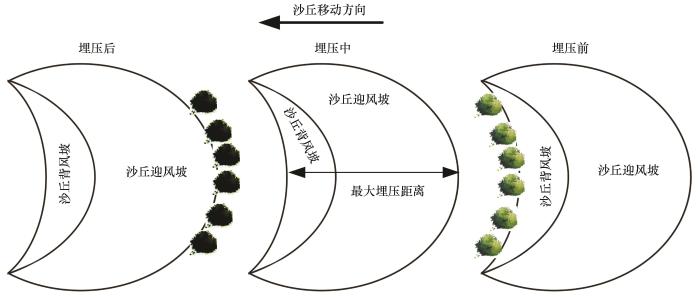
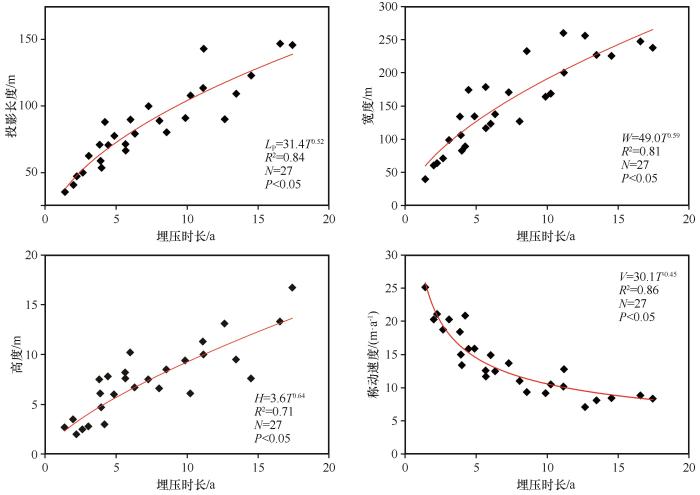
... 根据形态特征,在研究区选择27个新月形沙丘(不包括采集样品的所有沙丘),利用QuickBird遥感影像,测量沙丘宽度(W)、背风坡长度、迎风坡和背风坡在水平面投影的最大长度(Ld)和两期影像间沙丘移动距离,并计算对应沙丘高度(H)和移动速度(V,单位为m·a-1)[29-30].同一沙丘不同部位迎风坡和背风坡在地面投影长度不同,对植物埋压时长亦不相同,由于植物枝条枯落物均采集自沙丘中轴线附近的迎风坡坡脚,故本研究中沙丘对荒漠植物的埋压时长(T,单位为a)为沿沙丘移动方向沙丘迎风坡和背风坡在水平面投影的最大长度除以沙丘移动速度,即为沙丘对植物的最大埋压时长,具体表达式为: ...
Shaping effects of sand flow channels on aeolian geomorphology:a case study of the Badain Jaran,Tengger,and Ulan Buh Deserts,northern China
2
2022
... 巴丹吉林沙漠位于阿拉善高原的中心,是中国第二大流动沙漠[27].巴丹吉林沙漠-乌兰布和沙漠输沙带(39°30′—41°00′N,104°00′—106°00′E)[28]从巴丹吉林沙漠东南部马拉盖山南侧开始延伸至乌兰布和沙漠北部,长约250 km,宽5~45 km[28-29](图1).年平均气温为10 ℃,多年平均降水量为150 mm,全年降水集中在6—9月(占全年降水量的70%)[29],年平均大风日数大于50 d,年平均风速为3.7 m·s-1,主导风向为W、WNW和NW[29-30].多年平均潜在蒸散量为1 447 mm,干燥指数为0.07,属于典型干旱荒漠区[31].植被组成以旱生超旱生和盐生灌木、小灌木和小半灌木为主,主要有霸王(Zygophyllum xanthoxylon)、白刺(Nitraria tangutorum)、小叶锦鸡儿(Caragana microphylla)、珍珠(Salsola passerina)、红砂(Reaumuria soongorica)和猫头刺(Oxytropis aciphylla)等[32].输沙带沙丘以新月形沙丘和沙丘链为主.新月形沙丘的高度为3~20 m,沙丘链长约200~5 000 km,高度为5~15 m[28].新月形沙丘和沙丘链的丘间地存在大量白刺灌丛沙堆和锦鸡儿灌丛沙堆[28].地带性土壤为灰漠土,砾石、粗砂含量高,丘间地有薄层覆沙[29]. ...
... 根据形态特征,在研究区选择27个新月形沙丘(不包括采集样品的所有沙丘),利用QuickBird遥感影像,测量沙丘宽度(W)、背风坡长度、迎风坡和背风坡在水平面投影的最大长度(Ld)和两期影像间沙丘移动距离,并计算对应沙丘高度(H)和移动速度(V,单位为m·a-1)[29-30].同一沙丘不同部位迎风坡和背风坡在地面投影长度不同,对植物埋压时长亦不相同,由于植物枝条枯落物均采集自沙丘中轴线附近的迎风坡坡脚,故本研究中沙丘对荒漠植物的埋压时长(T,单位为a)为沿沙丘移动方向沙丘迎风坡和背风坡在水平面投影的最大长度除以沙丘移动速度,即为沙丘对植物的最大埋压时长,具体表达式为: ...
巴丹吉林沙漠与亚玛雷克沙漠之间连接带的动态研究
1
2021
... 巴丹吉林沙漠位于阿拉善高原的中心,是中国第二大流动沙漠[27].巴丹吉林沙漠-乌兰布和沙漠输沙带(39°30′—41°00′N,104°00′—106°00′E)[28]从巴丹吉林沙漠东南部马拉盖山南侧开始延伸至乌兰布和沙漠北部,长约250 km,宽5~45 km[28-29](图1).年平均气温为10 ℃,多年平均降水量为150 mm,全年降水集中在6—9月(占全年降水量的70%)[29],年平均大风日数大于50 d,年平均风速为3.7 m·s-1,主导风向为W、WNW和NW[29-30].多年平均潜在蒸散量为1 447 mm,干燥指数为0.07,属于典型干旱荒漠区[31].植被组成以旱生超旱生和盐生灌木、小灌木和小半灌木为主,主要有霸王(Zygophyllum xanthoxylon)、白刺(Nitraria tangutorum)、小叶锦鸡儿(Caragana microphylla)、珍珠(Salsola passerina)、红砂(Reaumuria soongorica)和猫头刺(Oxytropis aciphylla)等[32].输沙带沙丘以新月形沙丘和沙丘链为主.新月形沙丘的高度为3~20 m,沙丘链长约200~5 000 km,高度为5~15 m[28].新月形沙丘和沙丘链的丘间地存在大量白刺灌丛沙堆和锦鸡儿灌丛沙堆[28].地带性土壤为灰漠土,砾石、粗砂含量高,丘间地有薄层覆沙[29]. ...
巴丹吉林沙漠东缘主要植物种群空间分布关系
1
2021
... 巴丹吉林沙漠位于阿拉善高原的中心,是中国第二大流动沙漠[27].巴丹吉林沙漠-乌兰布和沙漠输沙带(39°30′—41°00′N,104°00′—106°00′E)[28]从巴丹吉林沙漠东南部马拉盖山南侧开始延伸至乌兰布和沙漠北部,长约250 km,宽5~45 km[28-29](图1).年平均气温为10 ℃,多年平均降水量为150 mm,全年降水集中在6—9月(占全年降水量的70%)[29],年平均大风日数大于50 d,年平均风速为3.7 m·s-1,主导风向为W、WNW和NW[29-30].多年平均潜在蒸散量为1 447 mm,干燥指数为0.07,属于典型干旱荒漠区[31].植被组成以旱生超旱生和盐生灌木、小灌木和小半灌木为主,主要有霸王(Zygophyllum xanthoxylon)、白刺(Nitraria tangutorum)、小叶锦鸡儿(Caragana microphylla)、珍珠(Salsola passerina)、红砂(Reaumuria soongorica)和猫头刺(Oxytropis aciphylla)等[32].输沙带沙丘以新月形沙丘和沙丘链为主.新月形沙丘的高度为3~20 m,沙丘链长约200~5 000 km,高度为5~15 m[28].新月形沙丘和沙丘链的丘间地存在大量白刺灌丛沙堆和锦鸡儿灌丛沙堆[28].地带性土壤为灰漠土,砾石、粗砂含量高,丘间地有薄层覆沙[29]. ...
克氏针茅草原植物凋落物分解特性及其对环境变化的响应
1
2006
... 被沙丘埋压的白刺枝条枯落物质量损失率与埋压时长间呈对数正相关,说明在被沙丘埋压的极端环境下,荒漠植物枝条枯落物的分解依然进行;但分解速率与时间之间的关系模型与在草原和干旱区正常环境中植物叶片分解速率与时间之间的单指数模型、双指数模型和渐近模型有差异[25,33],说明被沙丘埋压后枝条的分解速率要明显低于草原,也低于干旱区自然光照和荫蔽条件下枯落物分解速率.这可能是由于植物分解过程与环境因子密切相关[34],被沙丘埋压后隔绝了植物枝条与外界环境的水热交换和光照等条件,致使植物枯落物在正常环境下分解过程的某些环节减弱或者不能正常进行;同时,本研究探讨的是植物枝条枯落物的分解速率而其他研究分析的是植物的叶片,由于不同植物器官元素组成的较大差异[35],枝条本身比叶片更难分解,分解速率要更慢. ...
陆地生态系统凋落物分解研究进展
1
2010
... 被沙丘埋压的白刺枝条枯落物质量损失率与埋压时长间呈对数正相关,说明在被沙丘埋压的极端环境下,荒漠植物枝条枯落物的分解依然进行;但分解速率与时间之间的关系模型与在草原和干旱区正常环境中植物叶片分解速率与时间之间的单指数模型、双指数模型和渐近模型有差异[25,33],说明被沙丘埋压后枝条的分解速率要明显低于草原,也低于干旱区自然光照和荫蔽条件下枯落物分解速率.这可能是由于植物分解过程与环境因子密切相关[34],被沙丘埋压后隔绝了植物枝条与外界环境的水热交换和光照等条件,致使植物枯落物在正常环境下分解过程的某些环节减弱或者不能正常进行;同时,本研究探讨的是植物枝条枯落物的分解速率而其他研究分析的是植物的叶片,由于不同植物器官元素组成的较大差异[35],枝条本身比叶片更难分解,分解速率要更慢. ...
河西走廊荒漠区不同功能类群植物叶片建成成本的比较
1
2023
... 被沙丘埋压的白刺枝条枯落物质量损失率与埋压时长间呈对数正相关,说明在被沙丘埋压的极端环境下,荒漠植物枝条枯落物的分解依然进行;但分解速率与时间之间的关系模型与在草原和干旱区正常环境中植物叶片分解速率与时间之间的单指数模型、双指数模型和渐近模型有差异[25,33],说明被沙丘埋压后枝条的分解速率要明显低于草原,也低于干旱区自然光照和荫蔽条件下枯落物分解速率.这可能是由于植物分解过程与环境因子密切相关[34],被沙丘埋压后隔绝了植物枝条与外界环境的水热交换和光照等条件,致使植物枯落物在正常环境下分解过程的某些环节减弱或者不能正常进行;同时,本研究探讨的是植物枝条枯落物的分解速率而其他研究分析的是植物的叶片,由于不同植物器官元素组成的较大差异[35],枝条本身比叶片更难分解,分解速率要更慢. ...
森林凋落物分解研究进展
1
2006
... 枯落物的分解是缓慢的过程,分解初期速率较快,后期逐渐平缓[25-26].本研究发现白刺枝条在沙丘埋压情况下,其枯落物质量损失在被埋压的前25年较大,之后趋于平缓,这与王嘉年等[25]和范琳杰等[26]对干旱区荒漠植物分解速率随时间变化规律的研究结果一致,但时间转折点明显晚于其研究结果.这是由于枯落物分解前期主要与非生物因素(如物理破碎等)有关,枯落物中水溶性化合物先分解,在分解过程中像木质素等难以分解的物质累积,导致枯落物分解速率逐渐减缓,此时,枯落物的分解主要由生物因素主导[13,25,36-37],而分解速率快慢转折点出现的时间较晚的原因可能一方面是由于本研究分析的是荒漠植物枝条的分解速率,而其他研究关注的是荒漠植物的叶片,组成物质的差异造成枝条分解速率原本要远低于叶片,另一方面是被沙丘埋压的条件下植物的分解速率要低于自然条件. ...
Impacts of global warming on litter decomposition
1
2014
... 枯落物的分解是缓慢的过程,分解初期速率较快,后期逐渐平缓[25-26].本研究发现白刺枝条在沙丘埋压情况下,其枯落物质量损失在被埋压的前25年较大,之后趋于平缓,这与王嘉年等[25]和范琳杰等[26]对干旱区荒漠植物分解速率随时间变化规律的研究结果一致,但时间转折点明显晚于其研究结果.这是由于枯落物分解前期主要与非生物因素(如物理破碎等)有关,枯落物中水溶性化合物先分解,在分解过程中像木质素等难以分解的物质累积,导致枯落物分解速率逐渐减缓,此时,枯落物的分解主要由生物因素主导[13,25,36-37],而分解速率快慢转折点出现的时间较晚的原因可能一方面是由于本研究分析的是荒漠植物枝条的分解速率,而其他研究关注的是荒漠植物的叶片,组成物质的差异造成枝条分解速率原本要远低于叶片,另一方面是被沙丘埋压的条件下植物的分解速率要低于自然条件. ...
Earthworm effects on carbon and nitrogen dynamics of surface litter in corn agroecosystems
1
1997
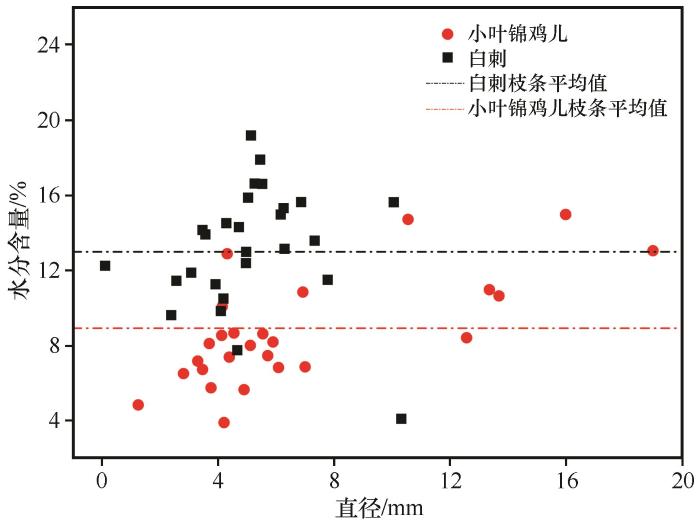
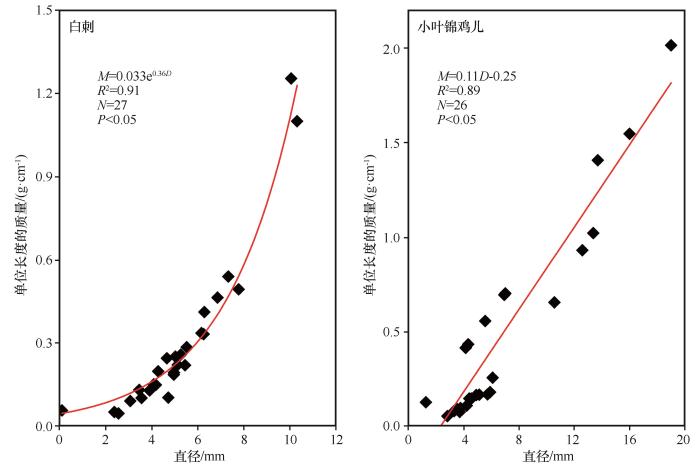
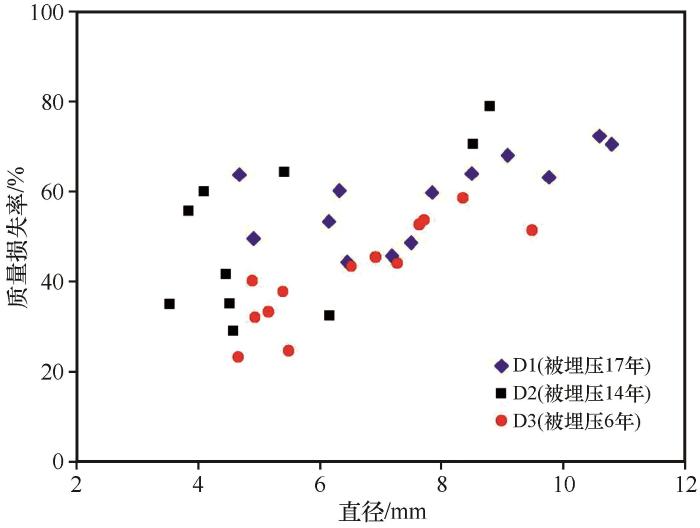
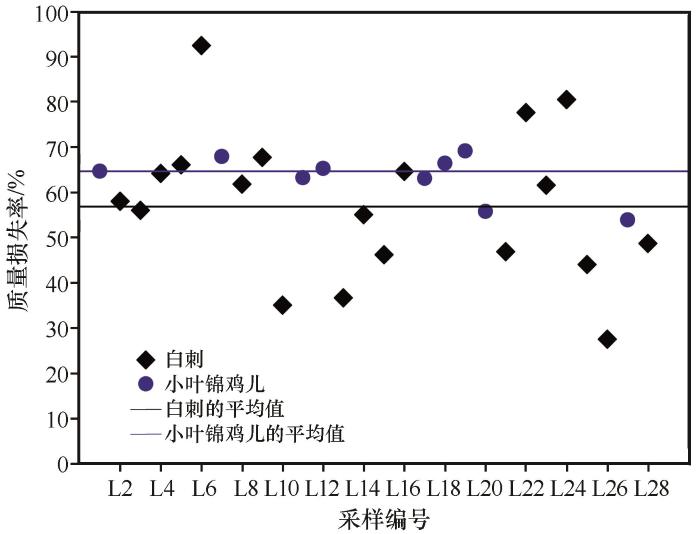
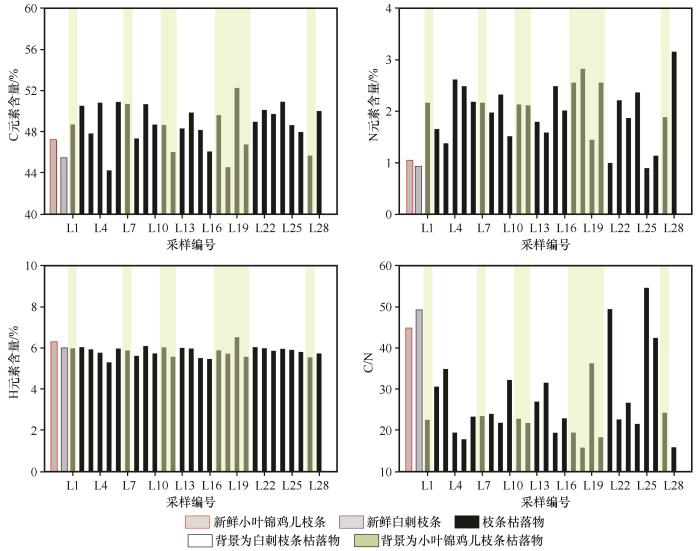
... 枯落物的初始化学组成可以用来很好地预测枯落物的分解,一般来说,枯落物中N含量越低、C/N越高,枯落物分解速率越慢[38-40];相反,枯落物中N含量高,可以促进微生物活性,从而加快凋落物的分解[41-44].在本研究中相同被沙丘埋压状况下白刺枝条枯落物的质量损失率低于小叶锦鸡儿,说明在相同埋压环境下,白刺枝条的分解速率小于小叶锦鸡儿,这是由于白刺枝条纤维素含量与C/N比均高于小叶锦鸡儿,因此其分解速率较低.此外,沙丘不同部位移动速度相同,但从中轴线向两翼角方向沙丘迎风坡坡长和背风坡坡长在地面的投影长度在变小,故同一沙丘从中轴线向两翼角方向沙丘对植物的埋压时长是变小的.研究所选的沙丘D13对迎风坡坡脚中轴线附近植物的埋压时长约比对离中轴线最远采样位置植物埋压时长多20年.但由于植物枝条枯落物采样位置相距很近,采样时无法通过普通GPS进行精准定位,无法估算沙丘D13紧邻迎风坡坡脚不同位置的植物枯落物的具体C、N埋压时长;另外由于被沙丘埋压93年,采样时沙丘中轴线附近的植物枯落物只剩粗枝条,造成不同位置枯落物质量损失率较大幅度变化,且出现了从沙丘近翼角位置向沙丘中轴线方向,随埋压时长的增加植物枯落物的分解速率及C和N元素含量有减小趋势的反规律变化.这说明沙丘对植物埋压过程中在刚开始被埋压阶段直径较粗枝条的分解速率大于较细的枝条,但随埋压时长增加较细枝条枯落物首先会分解殆尽,不同直径的枝条枯落物存在分解殆尽的埋压时长阈值,未来需加强不同规模沙丘的不同部位的埋压对植物枯落物分解的影响研究. ...
Litter decomposition rates in Canadian forests
1999
Temporal dynamics of microbial communities on decomposing leaf litter of 10 plant species in relation to decomposition rate
1
2012
... 枯落物的初始化学组成可以用来很好地预测枯落物的分解,一般来说,枯落物中N含量越低、C/N越高,枯落物分解速率越慢[38-40];相反,枯落物中N含量高,可以促进微生物活性,从而加快凋落物的分解[41-44].在本研究中相同被沙丘埋压状况下白刺枝条枯落物的质量损失率低于小叶锦鸡儿,说明在相同埋压环境下,白刺枝条的分解速率小于小叶锦鸡儿,这是由于白刺枝条纤维素含量与C/N比均高于小叶锦鸡儿,因此其分解速率较低.此外,沙丘不同部位移动速度相同,但从中轴线向两翼角方向沙丘迎风坡坡长和背风坡坡长在地面的投影长度在变小,故同一沙丘从中轴线向两翼角方向沙丘对植物的埋压时长是变小的.研究所选的沙丘D13对迎风坡坡脚中轴线附近植物的埋压时长约比对离中轴线最远采样位置植物埋压时长多20年.但由于植物枝条枯落物采样位置相距很近,采样时无法通过普通GPS进行精准定位,无法估算沙丘D13紧邻迎风坡坡脚不同位置的植物枯落物的具体C、N埋压时长;另外由于被沙丘埋压93年,采样时沙丘中轴线附近的植物枯落物只剩粗枝条,造成不同位置枯落物质量损失率较大幅度变化,且出现了从沙丘近翼角位置向沙丘中轴线方向,随埋压时长的增加植物枯落物的分解速率及C和N元素含量有减小趋势的反规律变化.这说明沙丘对植物埋压过程中在刚开始被埋压阶段直径较粗枝条的分解速率大于较细的枝条,但随埋压时长增加较细枝条枯落物首先会分解殆尽,不同直径的枝条枯落物存在分解殆尽的埋压时长阈值,未来需加强不同规模沙丘的不同部位的埋压对植物枯落物分解的影响研究. ...
不同调控措施对杉木枯落物分解的影响
1
2006
... 枯落物的初始化学组成可以用来很好地预测枯落物的分解,一般来说,枯落物中N含量越低、C/N越高,枯落物分解速率越慢[38-40];相反,枯落物中N含量高,可以促进微生物活性,从而加快凋落物的分解[41-44].在本研究中相同被沙丘埋压状况下白刺枝条枯落物的质量损失率低于小叶锦鸡儿,说明在相同埋压环境下,白刺枝条的分解速率小于小叶锦鸡儿,这是由于白刺枝条纤维素含量与C/N比均高于小叶锦鸡儿,因此其分解速率较低.此外,沙丘不同部位移动速度相同,但从中轴线向两翼角方向沙丘迎风坡坡长和背风坡坡长在地面的投影长度在变小,故同一沙丘从中轴线向两翼角方向沙丘对植物的埋压时长是变小的.研究所选的沙丘D13对迎风坡坡脚中轴线附近植物的埋压时长约比对离中轴线最远采样位置植物埋压时长多20年.但由于植物枝条枯落物采样位置相距很近,采样时无法通过普通GPS进行精准定位,无法估算沙丘D13紧邻迎风坡坡脚不同位置的植物枯落物的具体C、N埋压时长;另外由于被沙丘埋压93年,采样时沙丘中轴线附近的植物枯落物只剩粗枝条,造成不同位置枯落物质量损失率较大幅度变化,且出现了从沙丘近翼角位置向沙丘中轴线方向,随埋压时长的增加植物枯落物的分解速率及C和N元素含量有减小趋势的反规律变化.这说明沙丘对植物埋压过程中在刚开始被埋压阶段直径较粗枝条的分解速率大于较细的枝条,但随埋压时长增加较细枝条枯落物首先会分解殆尽,不同直径的枝条枯落物存在分解殆尽的埋压时长阈值,未来需加强不同规模沙丘的不同部位的埋压对植物枯落物分解的影响研究. ...
Litter composition rather than plant presence affects decomposition of tropical litter mixtures
2011
Decomposition of dominant plant species litter in a semi-arid grassland
2003
草原凋落物的分解及影响元素的分解与累积
1
2012
... 枯落物的初始化学组成可以用来很好地预测枯落物的分解,一般来说,枯落物中N含量越低、C/N越高,枯落物分解速率越慢[38-40];相反,枯落物中N含量高,可以促进微生物活性,从而加快凋落物的分解[41-44].在本研究中相同被沙丘埋压状况下白刺枝条枯落物的质量损失率低于小叶锦鸡儿,说明在相同埋压环境下,白刺枝条的分解速率小于小叶锦鸡儿,这是由于白刺枝条纤维素含量与C/N比均高于小叶锦鸡儿,因此其分解速率较低.此外,沙丘不同部位移动速度相同,但从中轴线向两翼角方向沙丘迎风坡坡长和背风坡坡长在地面的投影长度在变小,故同一沙丘从中轴线向两翼角方向沙丘对植物的埋压时长是变小的.研究所选的沙丘D13对迎风坡坡脚中轴线附近植物的埋压时长约比对离中轴线最远采样位置植物埋压时长多20年.但由于植物枝条枯落物采样位置相距很近,采样时无法通过普通GPS进行精准定位,无法估算沙丘D13紧邻迎风坡坡脚不同位置的植物枯落物的具体C、N埋压时长;另外由于被沙丘埋压93年,采样时沙丘中轴线附近的植物枯落物只剩粗枝条,造成不同位置枯落物质量损失率较大幅度变化,且出现了从沙丘近翼角位置向沙丘中轴线方向,随埋压时长的增加植物枯落物的分解速率及C和N元素含量有减小趋势的反规律变化.这说明沙丘对植物埋压过程中在刚开始被埋压阶段直径较粗枝条的分解速率大于较细的枝条,但随埋压时长增加较细枝条枯落物首先会分解殆尽,不同直径的枝条枯落物存在分解殆尽的埋压时长阈值,未来需加强不同规模沙丘的不同部位的埋压对植物枯落物分解的影响研究. ...
Estimating the contribution of leaf litter decomposition to soil CO2 efflux in a beech forest using 13C-depleted litter
1
2005
... 碳是构成植物枯落物有机质的主要组成元素,在其分解过程中,只有20%的C元素留存在土壤有机质中[45],枯落物分解过程中C元素的释放速度影响全球生态系统碳平衡[46],而C元素在枯落物分解过程中并不总被释放出来,其与枯落物自身特性、所在环境以及分解时间有关[47].本研究发现,被沙丘埋压后,植物枝条枯落物C和N的含量升高,C/N比在降低,这与孙本华等[48]在荒漠条件下对大豆秆的分解过程中C和N及C/N的变化规律的研究结果一致.主要是由于大规模的沙埋对植物有一定的压实作用,可能会改变植物内部结构,影响植物分解速率;另外植物在被沙丘埋压后与空气隔绝,仅依靠自身含氧发生厌氧细菌的分解作用,从而开始脱水、去羧基,减少腐殖酸等物质,使植物中C元素含量相对增高,而氧含量相对减少,存在碳化过程.而含N元素物质基本未参与或者参与较少分解反应,在分解过程中其相对含量增加比C多,故C/N比降低.随着埋压时间增加,氧气不充足条件下植物的分解反应继续进行,C和N相对含量越来越高,而C/N比越来越低.因此,被沙丘埋压后植物枯落物分解是C和N的富集过程,这与王嘉年等[25]在干旱区对枯落物的分解做遮光处理后C和N含量的变化结果一致,间接地反映出在干旱生态系统中光降解在C周转中的重要作用.但由于本研究只在沙丘附近非丘间地区域分别采集了1株生长旺盛的白刺和1株小叶锦鸡儿植株,对于C和N元素的富集度的精度有一定影响,在以后研究中需加大对鲜活植株的采样和测量.同时,被沙丘埋压不同阶段枯落物的物质组成以及导致C、N元素富集的具体原因亟待进一步探究. ...
The global carbon dioxide flux in soil respiration and its relationship to vegetation and climate
1
1992
... 碳是构成植物枯落物有机质的主要组成元素,在其分解过程中,只有20%的C元素留存在土壤有机质中[45],枯落物分解过程中C元素的释放速度影响全球生态系统碳平衡[46],而C元素在枯落物分解过程中并不总被释放出来,其与枯落物自身特性、所在环境以及分解时间有关[47].本研究发现,被沙丘埋压后,植物枝条枯落物C和N的含量升高,C/N比在降低,这与孙本华等[48]在荒漠条件下对大豆秆的分解过程中C和N及C/N的变化规律的研究结果一致.主要是由于大规模的沙埋对植物有一定的压实作用,可能会改变植物内部结构,影响植物分解速率;另外植物在被沙丘埋压后与空气隔绝,仅依靠自身含氧发生厌氧细菌的分解作用,从而开始脱水、去羧基,减少腐殖酸等物质,使植物中C元素含量相对增高,而氧含量相对减少,存在碳化过程.而含N元素物质基本未参与或者参与较少分解反应,在分解过程中其相对含量增加比C多,故C/N比降低.随着埋压时间增加,氧气不充足条件下植物的分解反应继续进行,C和N相对含量越来越高,而C/N比越来越低.因此,被沙丘埋压后植物枯落物分解是C和N的富集过程,这与王嘉年等[25]在干旱区对枯落物的分解做遮光处理后C和N含量的变化结果一致,间接地反映出在干旱生态系统中光降解在C周转中的重要作用.但由于本研究只在沙丘附近非丘间地区域分别采集了1株生长旺盛的白刺和1株小叶锦鸡儿植株,对于C和N元素的富集度的精度有一定影响,在以后研究中需加大对鲜活植株的采样和测量.同时,被沙丘埋压不同阶段枯落物的物质组成以及导致C、N元素富集的具体原因亟待进一步探究. ...
Leaching,accumulation and release of nitrogen in decomposing forest litter
1
1981
... 碳是构成植物枯落物有机质的主要组成元素,在其分解过程中,只有20%的C元素留存在土壤有机质中[45],枯落物分解过程中C元素的释放速度影响全球生态系统碳平衡[46],而C元素在枯落物分解过程中并不总被释放出来,其与枯落物自身特性、所在环境以及分解时间有关[47].本研究发现,被沙丘埋压后,植物枝条枯落物C和N的含量升高,C/N比在降低,这与孙本华等[48]在荒漠条件下对大豆秆的分解过程中C和N及C/N的变化规律的研究结果一致.主要是由于大规模的沙埋对植物有一定的压实作用,可能会改变植物内部结构,影响植物分解速率;另外植物在被沙丘埋压后与空气隔绝,仅依靠自身含氧发生厌氧细菌的分解作用,从而开始脱水、去羧基,减少腐殖酸等物质,使植物中C元素含量相对增高,而氧含量相对减少,存在碳化过程.而含N元素物质基本未参与或者参与较少分解反应,在分解过程中其相对含量增加比C多,故C/N比降低.随着埋压时间增加,氧气不充足条件下植物的分解反应继续进行,C和N相对含量越来越高,而C/N比越来越低.因此,被沙丘埋压后植物枯落物分解是C和N的富集过程,这与王嘉年等[25]在干旱区对枯落物的分解做遮光处理后C和N含量的变化结果一致,间接地反映出在干旱生态系统中光降解在C周转中的重要作用.但由于本研究只在沙丘附近非丘间地区域分别采集了1株生长旺盛的白刺和1株小叶锦鸡儿植株,对于C和N元素的富集度的精度有一定影响,在以后研究中需加大对鲜活植株的采样和测量.同时,被沙丘埋压不同阶段枯落物的物质组成以及导致C、N元素富集的具体原因亟待进一步探究. ...
荒漠生态条件下植物残体分解和养分释放的研究
1
2005
... 碳是构成植物枯落物有机质的主要组成元素,在其分解过程中,只有20%的C元素留存在土壤有机质中[45],枯落物分解过程中C元素的释放速度影响全球生态系统碳平衡[46],而C元素在枯落物分解过程中并不总被释放出来,其与枯落物自身特性、所在环境以及分解时间有关[47].本研究发现,被沙丘埋压后,植物枝条枯落物C和N的含量升高,C/N比在降低,这与孙本华等[48]在荒漠条件下对大豆秆的分解过程中C和N及C/N的变化规律的研究结果一致.主要是由于大规模的沙埋对植物有一定的压实作用,可能会改变植物内部结构,影响植物分解速率;另外植物在被沙丘埋压后与空气隔绝,仅依靠自身含氧发生厌氧细菌的分解作用,从而开始脱水、去羧基,减少腐殖酸等物质,使植物中C元素含量相对增高,而氧含量相对减少,存在碳化过程.而含N元素物质基本未参与或者参与较少分解反应,在分解过程中其相对含量增加比C多,故C/N比降低.随着埋压时间增加,氧气不充足条件下植物的分解反应继续进行,C和N相对含量越来越高,而C/N比越来越低.因此,被沙丘埋压后植物枯落物分解是C和N的富集过程,这与王嘉年等[25]在干旱区对枯落物的分解做遮光处理后C和N含量的变化结果一致,间接地反映出在干旱生态系统中光降解在C周转中的重要作用.但由于本研究只在沙丘附近非丘间地区域分别采集了1株生长旺盛的白刺和1株小叶锦鸡儿植株,对于C和N元素的富集度的精度有一定影响,在以后研究中需加大对鲜活植株的采样和测量.同时,被沙丘埋压不同阶段枯落物的物质组成以及导致C、N元素富集的具体原因亟待进一步探究. ...






 甘公网安备 62010202000688号
甘公网安备 62010202000688号
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}