Interception loss and rainfall redistribution by three semi-arid growing shrubs in Northeastern Mexico
1
1990
... 降水是陆地生态系统的主要水分来源,尤其在干旱区,水分和养分有效性是植被生长和维持生态系统功能的关键限制因素.大气降水通过植被冠层时再分配为冠层截留、穿透雨和树干茎流三部分,其分配比例受冠层结构[1]、降雨特征[2]和气象条件[3]等因素影响,调控植被与土壤之间的水文和生物地球化学通量[4-5].其中,树干茎流指被树叶、枝条、树干拦截的沿着主干向下运动汇集至根际区的降水.Zhang等[6]基于全球量化研究得出树干茎流占降水比例的中位值为2.7%(四分位距:1.0%~6.3%).尽管占比小,由于降水在植被冠层内对大气沉积物和树木分泌物等的淋洗作用使得树干茎流养分富集[4,7],以及树干茎流在植物地上部分和地下部分运动表现出“双漏斗效应”[8-9],通常认为树干茎流在植被生态系统水分和养分中占有很大份额.尤其是在水分和养分受限的干旱生态系统,树干茎流是植物在贫瘠环境下重要的水分和养分的“点源”输入,对形成灌木根区水分和养分沃岛具有重要贡献[10-12].树干茎流影响着一系列生态水文过程,比如坡面漫流或地表径流产生[13-15]、土壤侵蚀[16-17]、壤中流[18]、地下水补给[19]、土壤水分空间分布格局[20-21]、冠层下方植被和附生植物分配[22]等. ...
Temporal persistence of spatial patterns in throughfall
1
2005
... 降水是陆地生态系统的主要水分来源,尤其在干旱区,水分和养分有效性是植被生长和维持生态系统功能的关键限制因素.大气降水通过植被冠层时再分配为冠层截留、穿透雨和树干茎流三部分,其分配比例受冠层结构[1]、降雨特征[2]和气象条件[3]等因素影响,调控植被与土壤之间的水文和生物地球化学通量[4-5].其中,树干茎流指被树叶、枝条、树干拦截的沿着主干向下运动汇集至根际区的降水.Zhang等[6]基于全球量化研究得出树干茎流占降水比例的中位值为2.7%(四分位距:1.0%~6.3%).尽管占比小,由于降水在植被冠层内对大气沉积物和树木分泌物等的淋洗作用使得树干茎流养分富集[4,7],以及树干茎流在植物地上部分和地下部分运动表现出“双漏斗效应”[8-9],通常认为树干茎流在植被生态系统水分和养分中占有很大份额.尤其是在水分和养分受限的干旱生态系统,树干茎流是植物在贫瘠环境下重要的水分和养分的“点源”输入,对形成灌木根区水分和养分沃岛具有重要贡献[10-12].树干茎流影响着一系列生态水文过程,比如坡面漫流或地表径流产生[13-15]、土壤侵蚀[16-17]、壤中流[18]、地下水补给[19]、土壤水分空间分布格局[20-21]、冠层下方植被和附生植物分配[22]等. ...
Water funnelling by the crown of Flourensia cernua,a Chihuahuan Desert shrub
1
1993
... 降水是陆地生态系统的主要水分来源,尤其在干旱区,水分和养分有效性是植被生长和维持生态系统功能的关键限制因素.大气降水通过植被冠层时再分配为冠层截留、穿透雨和树干茎流三部分,其分配比例受冠层结构[1]、降雨特征[2]和气象条件[3]等因素影响,调控植被与土壤之间的水文和生物地球化学通量[4-5].其中,树干茎流指被树叶、枝条、树干拦截的沿着主干向下运动汇集至根际区的降水.Zhang等[6]基于全球量化研究得出树干茎流占降水比例的中位值为2.7%(四分位距:1.0%~6.3%).尽管占比小,由于降水在植被冠层内对大气沉积物和树木分泌物等的淋洗作用使得树干茎流养分富集[4,7],以及树干茎流在植物地上部分和地下部分运动表现出“双漏斗效应”[8-9],通常认为树干茎流在植被生态系统水分和养分中占有很大份额.尤其是在水分和养分受限的干旱生态系统,树干茎流是植物在贫瘠环境下重要的水分和养分的“点源”输入,对形成灌木根区水分和养分沃岛具有重要贡献[10-12].树干茎流影响着一系列生态水文过程,比如坡面漫流或地表径流产生[13-15]、土壤侵蚀[16-17]、壤中流[18]、地下水补给[19]、土壤水分空间分布格局[20-21]、冠层下方植被和附生植物分配[22]等. ...
A review and evaluation of stemflow literature in the hydrologic and biogeochemical cycles of forested and agricultural ecosystems
2
2003
... 降水是陆地生态系统的主要水分来源,尤其在干旱区,水分和养分有效性是植被生长和维持生态系统功能的关键限制因素.大气降水通过植被冠层时再分配为冠层截留、穿透雨和树干茎流三部分,其分配比例受冠层结构[1]、降雨特征[2]和气象条件[3]等因素影响,调控植被与土壤之间的水文和生物地球化学通量[4-5].其中,树干茎流指被树叶、枝条、树干拦截的沿着主干向下运动汇集至根际区的降水.Zhang等[6]基于全球量化研究得出树干茎流占降水比例的中位值为2.7%(四分位距:1.0%~6.3%).尽管占比小,由于降水在植被冠层内对大气沉积物和树木分泌物等的淋洗作用使得树干茎流养分富集[4,7],以及树干茎流在植物地上部分和地下部分运动表现出“双漏斗效应”[8-9],通常认为树干茎流在植被生态系统水分和养分中占有很大份额.尤其是在水分和养分受限的干旱生态系统,树干茎流是植物在贫瘠环境下重要的水分和养分的“点源”输入,对形成灌木根区水分和养分沃岛具有重要贡献[10-12].树干茎流影响着一系列生态水文过程,比如坡面漫流或地表径流产生[13-15]、土壤侵蚀[16-17]、壤中流[18]、地下水补给[19]、土壤水分空间分布格局[20-21]、冠层下方植被和附生植物分配[22]等. ...
... [4,7],以及树干茎流在植物地上部分和地下部分运动表现出“双漏斗效应”[8-9],通常认为树干茎流在植被生态系统水分和养分中占有很大份额.尤其是在水分和养分受限的干旱生态系统,树干茎流是植物在贫瘠环境下重要的水分和养分的“点源”输入,对形成灌木根区水分和养分沃岛具有重要贡献[10-12].树干茎流影响着一系列生态水文过程,比如坡面漫流或地表径流产生[13-15]、土壤侵蚀[16-17]、壤中流[18]、地下水补给[19]、土壤水分空间分布格局[20-21]、冠层下方植被和附生植物分配[22]等. ...
Rainfall partitioning by vegetation under Mediterranean conditions:a review of studies in Europe
1
2007
... 降水是陆地生态系统的主要水分来源,尤其在干旱区,水分和养分有效性是植被生长和维持生态系统功能的关键限制因素.大气降水通过植被冠层时再分配为冠层截留、穿透雨和树干茎流三部分,其分配比例受冠层结构[1]、降雨特征[2]和气象条件[3]等因素影响,调控植被与土壤之间的水文和生物地球化学通量[4-5].其中,树干茎流指被树叶、枝条、树干拦截的沿着主干向下运动汇集至根际区的降水.Zhang等[6]基于全球量化研究得出树干茎流占降水比例的中位值为2.7%(四分位距:1.0%~6.3%).尽管占比小,由于降水在植被冠层内对大气沉积物和树木分泌物等的淋洗作用使得树干茎流养分富集[4,7],以及树干茎流在植物地上部分和地下部分运动表现出“双漏斗效应”[8-9],通常认为树干茎流在植被生态系统水分和养分中占有很大份额.尤其是在水分和养分受限的干旱生态系统,树干茎流是植物在贫瘠环境下重要的水分和养分的“点源”输入,对形成灌木根区水分和养分沃岛具有重要贡献[10-12].树干茎流影响着一系列生态水文过程,比如坡面漫流或地表径流产生[13-15]、土壤侵蚀[16-17]、壤中流[18]、地下水补给[19]、土壤水分空间分布格局[20-21]、冠层下方植被和附生植物分配[22]等. ...
Global quantitative synthesis of effects of biotic and abiotic factors on stemflow production in woody ecosystems
2
2021
... 降水是陆地生态系统的主要水分来源,尤其在干旱区,水分和养分有效性是植被生长和维持生态系统功能的关键限制因素.大气降水通过植被冠层时再分配为冠层截留、穿透雨和树干茎流三部分,其分配比例受冠层结构[1]、降雨特征[2]和气象条件[3]等因素影响,调控植被与土壤之间的水文和生物地球化学通量[4-5].其中,树干茎流指被树叶、枝条、树干拦截的沿着主干向下运动汇集至根际区的降水.Zhang等[6]基于全球量化研究得出树干茎流占降水比例的中位值为2.7%(四分位距:1.0%~6.3%).尽管占比小,由于降水在植被冠层内对大气沉积物和树木分泌物等的淋洗作用使得树干茎流养分富集[4,7],以及树干茎流在植物地上部分和地下部分运动表现出“双漏斗效应”[8-9],通常认为树干茎流在植被生态系统水分和养分中占有很大份额.尤其是在水分和养分受限的干旱生态系统,树干茎流是植物在贫瘠环境下重要的水分和养分的“点源”输入,对形成灌木根区水分和养分沃岛具有重要贡献[10-12].树干茎流影响着一系列生态水文过程,比如坡面漫流或地表径流产生[13-15]、土壤侵蚀[16-17]、壤中流[18]、地下水补给[19]、土壤水分空间分布格局[20-21]、冠层下方植被和附生植物分配[22]等. ...
... 本研究得出,柠条锦鸡儿产生的树干茎流量占降水量的3.6%.Zhang等[6]研究表明全球灌木树干茎流占降水比例的中位值为7.2%(四分位距:5.2%~11.9%),不同灌木种间树干茎流产量差异较大.比如,厄瓜多尔加拉帕戈斯地区的罗宾逊绢木(Miconia robinsoniana)树干茎流百分比仅为0.6%[42],美国费城地区北美鼠刺(Itea virginica)树干茎流百分比为4.4%[43],陕西神木六道沟流域沙柳(Salix psammophila)树干茎流百分比为6.3%[44],西班牙阿尔梅里亚地区脱皮腺果豆(Adenocarpus decorticans)树干茎流百分比为4.4%[45],毛乌素沙地的沙柳树干茎流百分比9.0%[46],青藏高原东北部地区金路梅(Potentilla fruticosa)树干茎流百分比高达49.3%[47].树干茎流占降水比例较低,但由于灌木表现出的漏斗效应,灌木基部树干茎流的汇流量大.例如,Ck_1树干茎流量达43.2 L(图3),树干茎流可进一步快速通过植物根系和土壤大孔隙形成优先流补给深层土壤,在植物根际区形成水分沃岛,这对旱区植物生存及其遭受水分胁迫时增强其环境适应能力和提高水分利用效率具有重要影响[29]. ...
Throughfall and stemflow in the forest nutrient cycle
1
1983
... 降水是陆地生态系统的主要水分来源,尤其在干旱区,水分和养分有效性是植被生长和维持生态系统功能的关键限制因素.大气降水通过植被冠层时再分配为冠层截留、穿透雨和树干茎流三部分,其分配比例受冠层结构[1]、降雨特征[2]和气象条件[3]等因素影响,调控植被与土壤之间的水文和生物地球化学通量[4-5].其中,树干茎流指被树叶、枝条、树干拦截的沿着主干向下运动汇集至根际区的降水.Zhang等[6]基于全球量化研究得出树干茎流占降水比例的中位值为2.7%(四分位距:1.0%~6.3%).尽管占比小,由于降水在植被冠层内对大气沉积物和树木分泌物等的淋洗作用使得树干茎流养分富集[4,7],以及树干茎流在植物地上部分和地下部分运动表现出“双漏斗效应”[8-9],通常认为树干茎流在植被生态系统水分和养分中占有很大份额.尤其是在水分和养分受限的干旱生态系统,树干茎流是植物在贫瘠环境下重要的水分和养分的“点源”输入,对形成灌木根区水分和养分沃岛具有重要贡献[10-12].树干茎流影响着一系列生态水文过程,比如坡面漫流或地表径流产生[13-15]、土壤侵蚀[16-17]、壤中流[18]、地下水补给[19]、土壤水分空间分布格局[20-21]、冠层下方植被和附生植物分配[22]等. ...
Double-funneling of trees:stemflow and root-induced preferential flow
1
2006
... 降水是陆地生态系统的主要水分来源,尤其在干旱区,水分和养分有效性是植被生长和维持生态系统功能的关键限制因素.大气降水通过植被冠层时再分配为冠层截留、穿透雨和树干茎流三部分,其分配比例受冠层结构[1]、降雨特征[2]和气象条件[3]等因素影响,调控植被与土壤之间的水文和生物地球化学通量[4-5].其中,树干茎流指被树叶、枝条、树干拦截的沿着主干向下运动汇集至根际区的降水.Zhang等[6]基于全球量化研究得出树干茎流占降水比例的中位值为2.7%(四分位距:1.0%~6.3%).尽管占比小,由于降水在植被冠层内对大气沉积物和树木分泌物等的淋洗作用使得树干茎流养分富集[4,7],以及树干茎流在植物地上部分和地下部分运动表现出“双漏斗效应”[8-9],通常认为树干茎流在植被生态系统水分和养分中占有很大份额.尤其是在水分和养分受限的干旱生态系统,树干茎流是植物在贫瘠环境下重要的水分和养分的“点源”输入,对形成灌木根区水分和养分沃岛具有重要贡献[10-12].树干茎流影响着一系列生态水文过程,比如坡面漫流或地表径流产生[13-15]、土壤侵蚀[16-17]、壤中流[18]、地下水补给[19]、土壤水分空间分布格局[20-21]、冠层下方植被和附生植物分配[22]等. ...
Pairing geophysical techniques improves understanding of the near-surface critical zone:visualization of preferential routing of stemflow along coarse roots
1
2020
... 降水是陆地生态系统的主要水分来源,尤其在干旱区,水分和养分有效性是植被生长和维持生态系统功能的关键限制因素.大气降水通过植被冠层时再分配为冠层截留、穿透雨和树干茎流三部分,其分配比例受冠层结构[1]、降雨特征[2]和气象条件[3]等因素影响,调控植被与土壤之间的水文和生物地球化学通量[4-5].其中,树干茎流指被树叶、枝条、树干拦截的沿着主干向下运动汇集至根际区的降水.Zhang等[6]基于全球量化研究得出树干茎流占降水比例的中位值为2.7%(四分位距:1.0%~6.3%).尽管占比小,由于降水在植被冠层内对大气沉积物和树木分泌物等的淋洗作用使得树干茎流养分富集[4,7],以及树干茎流在植物地上部分和地下部分运动表现出“双漏斗效应”[8-9],通常认为树干茎流在植被生态系统水分和养分中占有很大份额.尤其是在水分和养分受限的干旱生态系统,树干茎流是植物在贫瘠环境下重要的水分和养分的“点源”输入,对形成灌木根区水分和养分沃岛具有重要贡献[10-12].树干茎流影响着一系列生态水文过程,比如坡面漫流或地表径流产生[13-15]、土壤侵蚀[16-17]、壤中流[18]、地下水补给[19]、土壤水分空间分布格局[20-21]、冠层下方植被和附生植物分配[22]等. ...
Stemflow in two xerophytic shrubs and its significance to soil water and nutrient enrichment
2
2013
... 降水是陆地生态系统的主要水分来源,尤其在干旱区,水分和养分有效性是植被生长和维持生态系统功能的关键限制因素.大气降水通过植被冠层时再分配为冠层截留、穿透雨和树干茎流三部分,其分配比例受冠层结构[1]、降雨特征[2]和气象条件[3]等因素影响,调控植被与土壤之间的水文和生物地球化学通量[4-5].其中,树干茎流指被树叶、枝条、树干拦截的沿着主干向下运动汇集至根际区的降水.Zhang等[6]基于全球量化研究得出树干茎流占降水比例的中位值为2.7%(四分位距:1.0%~6.3%).尽管占比小,由于降水在植被冠层内对大气沉积物和树木分泌物等的淋洗作用使得树干茎流养分富集[4,7],以及树干茎流在植物地上部分和地下部分运动表现出“双漏斗效应”[8-9],通常认为树干茎流在植被生态系统水分和养分中占有很大份额.尤其是在水分和养分受限的干旱生态系统,树干茎流是植物在贫瘠环境下重要的水分和养分的“点源”输入,对形成灌木根区水分和养分沃岛具有重要贡献[10-12].树干茎流影响着一系列生态水文过程,比如坡面漫流或地表径流产生[13-15]、土壤侵蚀[16-17]、壤中流[18]、地下水补给[19]、土壤水分空间分布格局[20-21]、冠层下方植被和附生植物分配[22]等. ...
... 降水抵达地表之前,植被冠层可视为雨水汇集区,不同物种之间树干茎流产量差异明显.Martinez-Meza等[23]报道,奇瓦瓦沙漠灌木腺牧豆树(Prosopis glandulosa)、三齿团香木(Larrea tridentata)和美洲焦油灌木(Flourensia cernua)树干茎流量分别占降水的5.4%、10%、10.5%;黄土高原多枝柽柳(Tamarix ramosissima)、柠条锦鸡儿(Caragana korshinskii)、红砂(Reaumuria soongorica)的树干茎流量分别占降水的2.2%、3.7%、7.2%[24];科尔沁沙地小叶锦鸡儿(Caragana microphylla)、木岩黄芪(Hedysarum fruticosum)、黄柳(Salix gordejevii)的树干茎流量分别占降水的11%、6.3%、5.3%[25];腾格里沙漠柠条锦鸡儿和油蒿(Artemisia ordosica)的树干茎流量分别占降水的8.8%和2.8%[10].冠层结构参数是导致树干茎流产量存在显著差异的关键因素.Kelly[26]发现树木在无叶期树干茎流产量最高;Levia等[27]发现山毛榉(Fagus sylvatica)幼树的树干茎流产量随叶片数量增加呈上升趋势,但达到一定阈值后趋于稳定;Levia等[27]还发现美国马里兰州的温带森林的树干茎流产量与树干大小显著正相关;Brown等[28]研究表明,相较于树皮易脱落的白橡树,具有紧实树皮的黑橡树树干茎流产量更高;高大的植株往往对应更大冠层投影面积,易产生更多的树干茎流[23,29].然而,对于特定植物种,不同个体之间或同一个体不同分枝之间构型差异如何影响树干茎流产量?不同构型参数重要性次序如何?这些问题目前并不清楚.该方面研究对深入认识干旱区灌木水分蓄集和利用策略具有重要意义. ...
Facilitation and succession under the canopy of a leguminous shrub,Retama sphaerocarpa,in a semi-arid environment in South-east Spain
1996
On the spatial pattern of soil nutrients in desert ecosystems:ecological archives E077-002
1
1996
... 降水是陆地生态系统的主要水分来源,尤其在干旱区,水分和养分有效性是植被生长和维持生态系统功能的关键限制因素.大气降水通过植被冠层时再分配为冠层截留、穿透雨和树干茎流三部分,其分配比例受冠层结构[1]、降雨特征[2]和气象条件[3]等因素影响,调控植被与土壤之间的水文和生物地球化学通量[4-5].其中,树干茎流指被树叶、枝条、树干拦截的沿着主干向下运动汇集至根际区的降水.Zhang等[6]基于全球量化研究得出树干茎流占降水比例的中位值为2.7%(四分位距:1.0%~6.3%).尽管占比小,由于降水在植被冠层内对大气沉积物和树木分泌物等的淋洗作用使得树干茎流养分富集[4,7],以及树干茎流在植物地上部分和地下部分运动表现出“双漏斗效应”[8-9],通常认为树干茎流在植被生态系统水分和养分中占有很大份额.尤其是在水分和养分受限的干旱生态系统,树干茎流是植物在贫瘠环境下重要的水分和养分的“点源”输入,对形成灌木根区水分和养分沃岛具有重要贡献[10-12].树干茎流影响着一系列生态水文过程,比如坡面漫流或地表径流产生[13-15]、土壤侵蚀[16-17]、壤中流[18]、地下水补给[19]、土壤水分空间分布格局[20-21]、冠层下方植被和附生植物分配[22]等. ...
Infiltration‐excess caused by stemflow in a cyclone‐prone tropical rainforest
1
1986
... 降水是陆地生态系统的主要水分来源,尤其在干旱区,水分和养分有效性是植被生长和维持生态系统功能的关键限制因素.大气降水通过植被冠层时再分配为冠层截留、穿透雨和树干茎流三部分,其分配比例受冠层结构[1]、降雨特征[2]和气象条件[3]等因素影响,调控植被与土壤之间的水文和生物地球化学通量[4-5].其中,树干茎流指被树叶、枝条、树干拦截的沿着主干向下运动汇集至根际区的降水.Zhang等[6]基于全球量化研究得出树干茎流占降水比例的中位值为2.7%(四分位距:1.0%~6.3%).尽管占比小,由于降水在植被冠层内对大气沉积物和树木分泌物等的淋洗作用使得树干茎流养分富集[4,7],以及树干茎流在植物地上部分和地下部分运动表现出“双漏斗效应”[8-9],通常认为树干茎流在植被生态系统水分和养分中占有很大份额.尤其是在水分和养分受限的干旱生态系统,树干茎流是植物在贫瘠环境下重要的水分和养分的“点源”输入,对形成灌木根区水分和养分沃岛具有重要贡献[10-12].树干茎流影响着一系列生态水文过程,比如坡面漫流或地表径流产生[13-15]、土壤侵蚀[16-17]、壤中流[18]、地下水补给[19]、土壤水分空间分布格局[20-21]、冠层下方植被和附生植物分配[22]等. ...
Vegetation influences on water yields from grassland and shrubland ecosystems in the Chihuahuan Desert
2002
Modelling runoff at the plot scale taking into account rainfall partitioning by vegetation:application to stemflow of banana (Musa spp.) plant
1
2009
... 降水是陆地生态系统的主要水分来源,尤其在干旱区,水分和养分有效性是植被生长和维持生态系统功能的关键限制因素.大气降水通过植被冠层时再分配为冠层截留、穿透雨和树干茎流三部分,其分配比例受冠层结构[1]、降雨特征[2]和气象条件[3]等因素影响,调控植被与土壤之间的水文和生物地球化学通量[4-5].其中,树干茎流指被树叶、枝条、树干拦截的沿着主干向下运动汇集至根际区的降水.Zhang等[6]基于全球量化研究得出树干茎流占降水比例的中位值为2.7%(四分位距:1.0%~6.3%).尽管占比小,由于降水在植被冠层内对大气沉积物和树木分泌物等的淋洗作用使得树干茎流养分富集[4,7],以及树干茎流在植物地上部分和地下部分运动表现出“双漏斗效应”[8-9],通常认为树干茎流在植被生态系统水分和养分中占有很大份额.尤其是在水分和养分受限的干旱生态系统,树干茎流是植物在贫瘠环境下重要的水分和养分的“点源”输入,对形成灌木根区水分和养分沃岛具有重要贡献[10-12].树干茎流影响着一系列生态水文过程,比如坡面漫流或地表径流产生[13-15]、土壤侵蚀[16-17]、壤中流[18]、地下水补给[19]、土壤水分空间分布格局[20-21]、冠层下方植被和附生植物分配[22]等. ...
Buttresses of tropical rainforest trees influence hillslope processes
1
1988
... 降水是陆地生态系统的主要水分来源,尤其在干旱区,水分和养分有效性是植被生长和维持生态系统功能的关键限制因素.大气降水通过植被冠层时再分配为冠层截留、穿透雨和树干茎流三部分,其分配比例受冠层结构[1]、降雨特征[2]和气象条件[3]等因素影响,调控植被与土壤之间的水文和生物地球化学通量[4-5].其中,树干茎流指被树叶、枝条、树干拦截的沿着主干向下运动汇集至根际区的降水.Zhang等[6]基于全球量化研究得出树干茎流占降水比例的中位值为2.7%(四分位距:1.0%~6.3%).尽管占比小,由于降水在植被冠层内对大气沉积物和树木分泌物等的淋洗作用使得树干茎流养分富集[4,7],以及树干茎流在植物地上部分和地下部分运动表现出“双漏斗效应”[8-9],通常认为树干茎流在植被生态系统水分和养分中占有很大份额.尤其是在水分和养分受限的干旱生态系统,树干茎流是植物在贫瘠环境下重要的水分和养分的“点源”输入,对形成灌木根区水分和养分沃岛具有重要贡献[10-12].树干茎流影响着一系列生态水文过程,比如坡面漫流或地表径流产生[13-15]、土壤侵蚀[16-17]、壤中流[18]、地下水补给[19]、土壤水分空间分布格局[20-21]、冠层下方植被和附生植物分配[22]等. ...
Rainfall energy under creosotebush
1
1999
... 降水是陆地生态系统的主要水分来源,尤其在干旱区,水分和养分有效性是植被生长和维持生态系统功能的关键限制因素.大气降水通过植被冠层时再分配为冠层截留、穿透雨和树干茎流三部分,其分配比例受冠层结构[1]、降雨特征[2]和气象条件[3]等因素影响,调控植被与土壤之间的水文和生物地球化学通量[4-5].其中,树干茎流指被树叶、枝条、树干拦截的沿着主干向下运动汇集至根际区的降水.Zhang等[6]基于全球量化研究得出树干茎流占降水比例的中位值为2.7%(四分位距:1.0%~6.3%).尽管占比小,由于降水在植被冠层内对大气沉积物和树木分泌物等的淋洗作用使得树干茎流养分富集[4,7],以及树干茎流在植物地上部分和地下部分运动表现出“双漏斗效应”[8-9],通常认为树干茎流在植被生态系统水分和养分中占有很大份额.尤其是在水分和养分受限的干旱生态系统,树干茎流是植物在贫瘠环境下重要的水分和养分的“点源”输入,对形成灌木根区水分和养分沃岛具有重要贡献[10-12].树干茎流影响着一系列生态水文过程,比如坡面漫流或地表径流产生[13-15]、土壤侵蚀[16-17]、壤中流[18]、地下水补给[19]、土壤水分空间分布格局[20-21]、冠层下方植被和附生植物分配[22]等. ...
Evidence of double-funneling effect of beech trees by visualization of flow pathways using dye tracer
1
2012
... 降水是陆地生态系统的主要水分来源,尤其在干旱区,水分和养分有效性是植被生长和维持生态系统功能的关键限制因素.大气降水通过植被冠层时再分配为冠层截留、穿透雨和树干茎流三部分,其分配比例受冠层结构[1]、降雨特征[2]和气象条件[3]等因素影响,调控植被与土壤之间的水文和生物地球化学通量[4-5].其中,树干茎流指被树叶、枝条、树干拦截的沿着主干向下运动汇集至根际区的降水.Zhang等[6]基于全球量化研究得出树干茎流占降水比例的中位值为2.7%(四分位距:1.0%~6.3%).尽管占比小,由于降水在植被冠层内对大气沉积物和树木分泌物等的淋洗作用使得树干茎流养分富集[4,7],以及树干茎流在植物地上部分和地下部分运动表现出“双漏斗效应”[8-9],通常认为树干茎流在植被生态系统水分和养分中占有很大份额.尤其是在水分和养分受限的干旱生态系统,树干茎流是植物在贫瘠环境下重要的水分和养分的“点源”输入,对形成灌木根区水分和养分沃岛具有重要贡献[10-12].树干茎流影响着一系列生态水文过程,比如坡面漫流或地表径流产生[13-15]、土壤侵蚀[16-17]、壤中流[18]、地下水补给[19]、土壤水分空间分布格局[20-21]、冠层下方植被和附生植物分配[22]等. ...
Significance of stemflow in groundwater recharge 1:evaluation of the stemflow contribution to recharge using a mass balance approach
1
1996
... 降水是陆地生态系统的主要水分来源,尤其在干旱区,水分和养分有效性是植被生长和维持生态系统功能的关键限制因素.大气降水通过植被冠层时再分配为冠层截留、穿透雨和树干茎流三部分,其分配比例受冠层结构[1]、降雨特征[2]和气象条件[3]等因素影响,调控植被与土壤之间的水文和生物地球化学通量[4-5].其中,树干茎流指被树叶、枝条、树干拦截的沿着主干向下运动汇集至根际区的降水.Zhang等[6]基于全球量化研究得出树干茎流占降水比例的中位值为2.7%(四分位距:1.0%~6.3%).尽管占比小,由于降水在植被冠层内对大气沉积物和树木分泌物等的淋洗作用使得树干茎流养分富集[4,7],以及树干茎流在植物地上部分和地下部分运动表现出“双漏斗效应”[8-9],通常认为树干茎流在植被生态系统水分和养分中占有很大份额.尤其是在水分和养分受限的干旱生态系统,树干茎流是植物在贫瘠环境下重要的水分和养分的“点源”输入,对形成灌木根区水分和养分沃岛具有重要贡献[10-12].树干茎流影响着一系列生态水文过程,比如坡面漫流或地表径流产生[13-15]、土壤侵蚀[16-17]、壤中流[18]、地下水补给[19]、土壤水分空间分布格局[20-21]、冠层下方植被和附生植物分配[22]等. ...
The effect of beech stemflow on spatial patterns of soil solution chemistry and seepage fluxes in a mixed beech/oak stand
1
2000
... 降水是陆地生态系统的主要水分来源,尤其在干旱区,水分和养分有效性是植被生长和维持生态系统功能的关键限制因素.大气降水通过植被冠层时再分配为冠层截留、穿透雨和树干茎流三部分,其分配比例受冠层结构[1]、降雨特征[2]和气象条件[3]等因素影响,调控植被与土壤之间的水文和生物地球化学通量[4-5].其中,树干茎流指被树叶、枝条、树干拦截的沿着主干向下运动汇集至根际区的降水.Zhang等[6]基于全球量化研究得出树干茎流占降水比例的中位值为2.7%(四分位距:1.0%~6.3%).尽管占比小,由于降水在植被冠层内对大气沉积物和树木分泌物等的淋洗作用使得树干茎流养分富集[4,7],以及树干茎流在植物地上部分和地下部分运动表现出“双漏斗效应”[8-9],通常认为树干茎流在植被生态系统水分和养分中占有很大份额.尤其是在水分和养分受限的干旱生态系统,树干茎流是植物在贫瘠环境下重要的水分和养分的“点源”输入,对形成灌木根区水分和养分沃岛具有重要贡献[10-12].树干茎流影响着一系列生态水文过程,比如坡面漫流或地表径流产生[13-15]、土壤侵蚀[16-17]、壤中流[18]、地下水补给[19]、土壤水分空间分布格局[20-21]、冠层下方植被和附生植物分配[22]等. ...
Soil water dynamics around a tree on a hillslope with or without rainwater supplied by stemflow
1
2011
... 降水是陆地生态系统的主要水分来源,尤其在干旱区,水分和养分有效性是植被生长和维持生态系统功能的关键限制因素.大气降水通过植被冠层时再分配为冠层截留、穿透雨和树干茎流三部分,其分配比例受冠层结构[1]、降雨特征[2]和气象条件[3]等因素影响,调控植被与土壤之间的水文和生物地球化学通量[4-5].其中,树干茎流指被树叶、枝条、树干拦截的沿着主干向下运动汇集至根际区的降水.Zhang等[6]基于全球量化研究得出树干茎流占降水比例的中位值为2.7%(四分位距:1.0%~6.3%).尽管占比小,由于降水在植被冠层内对大气沉积物和树木分泌物等的淋洗作用使得树干茎流养分富集[4,7],以及树干茎流在植物地上部分和地下部分运动表现出“双漏斗效应”[8-9],通常认为树干茎流在植被生态系统水分和养分中占有很大份额.尤其是在水分和养分受限的干旱生态系统,树干茎流是植物在贫瘠环境下重要的水分和养分的“点源”输入,对形成灌木根区水分和养分沃岛具有重要贡献[10-12].树干茎流影响着一系列生态水文过程,比如坡面漫流或地表径流产生[13-15]、土壤侵蚀[16-17]、壤中流[18]、地下水补给[19]、土壤水分空间分布格局[20-21]、冠层下方植被和附生植物分配[22]等. ...
Stemflow:a source of nutrients in some naturally growing epiphytic orchids of the Sikkim Himalaya
1
1995
... 降水是陆地生态系统的主要水分来源,尤其在干旱区,水分和养分有效性是植被生长和维持生态系统功能的关键限制因素.大气降水通过植被冠层时再分配为冠层截留、穿透雨和树干茎流三部分,其分配比例受冠层结构[1]、降雨特征[2]和气象条件[3]等因素影响,调控植被与土壤之间的水文和生物地球化学通量[4-5].其中,树干茎流指被树叶、枝条、树干拦截的沿着主干向下运动汇集至根际区的降水.Zhang等[6]基于全球量化研究得出树干茎流占降水比例的中位值为2.7%(四分位距:1.0%~6.3%).尽管占比小,由于降水在植被冠层内对大气沉积物和树木分泌物等的淋洗作用使得树干茎流养分富集[4,7],以及树干茎流在植物地上部分和地下部分运动表现出“双漏斗效应”[8-9],通常认为树干茎流在植被生态系统水分和养分中占有很大份额.尤其是在水分和养分受限的干旱生态系统,树干茎流是植物在贫瘠环境下重要的水分和养分的“点源”输入,对形成灌木根区水分和养分沃岛具有重要贡献[10-12].树干茎流影响着一系列生态水文过程,比如坡面漫流或地表径流产生[13-15]、土壤侵蚀[16-17]、壤中流[18]、地下水补给[19]、土壤水分空间分布格局[20-21]、冠层下方植被和附生植物分配[22]等. ...
Stemflow,throughfall and channelization of stemflow by roots in three Chihuahuan Desert shrubs
3
1996
... 降水抵达地表之前,植被冠层可视为雨水汇集区,不同物种之间树干茎流产量差异明显.Martinez-Meza等[23]报道,奇瓦瓦沙漠灌木腺牧豆树(Prosopis glandulosa)、三齿团香木(Larrea tridentata)和美洲焦油灌木(Flourensia cernua)树干茎流量分别占降水的5.4%、10%、10.5%;黄土高原多枝柽柳(Tamarix ramosissima)、柠条锦鸡儿(Caragana korshinskii)、红砂(Reaumuria soongorica)的树干茎流量分别占降水的2.2%、3.7%、7.2%[24];科尔沁沙地小叶锦鸡儿(Caragana microphylla)、木岩黄芪(Hedysarum fruticosum)、黄柳(Salix gordejevii)的树干茎流量分别占降水的11%、6.3%、5.3%[25];腾格里沙漠柠条锦鸡儿和油蒿(Artemisia ordosica)的树干茎流量分别占降水的8.8%和2.8%[10].冠层结构参数是导致树干茎流产量存在显著差异的关键因素.Kelly[26]发现树木在无叶期树干茎流产量最高;Levia等[27]发现山毛榉(Fagus sylvatica)幼树的树干茎流产量随叶片数量增加呈上升趋势,但达到一定阈值后趋于稳定;Levia等[27]还发现美国马里兰州的温带森林的树干茎流产量与树干大小显著正相关;Brown等[28]研究表明,相较于树皮易脱落的白橡树,具有紧实树皮的黑橡树树干茎流产量更高;高大的植株往往对应更大冠层投影面积,易产生更多的树干茎流[23,29].然而,对于特定植物种,不同个体之间或同一个体不同分枝之间构型差异如何影响树干茎流产量?不同构型参数重要性次序如何?这些问题目前并不清楚.该方面研究对深入认识干旱区灌木水分蓄集和利用策略具有重要意义. ...
... [23,29].然而,对于特定植物种,不同个体之间或同一个体不同分枝之间构型差异如何影响树干茎流产量?不同构型参数重要性次序如何?这些问题目前并不清楚.该方面研究对深入认识干旱区灌木水分蓄集和利用策略具有重要意义. ...
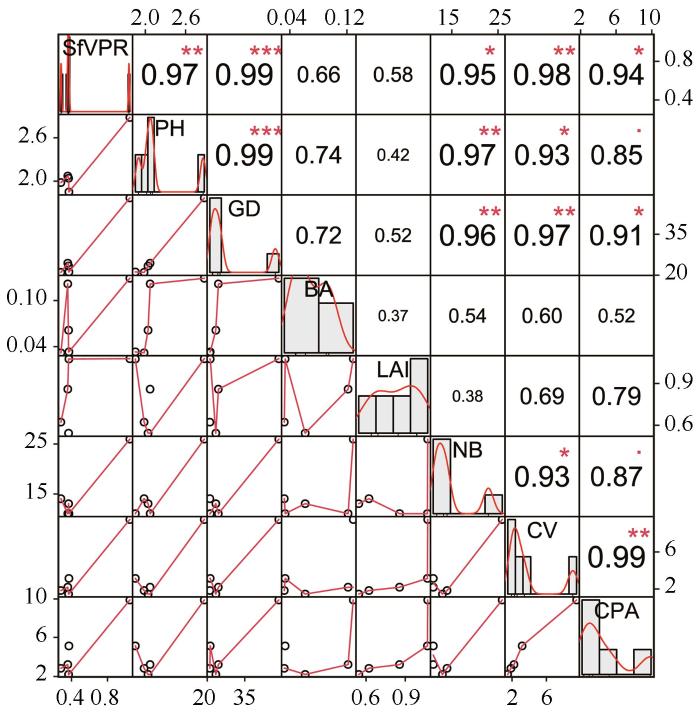
... 现有研究表明,冠层结构参数(株高、基径、基部面积、枝条数、冠层体积及冠层投影面积等)对树干茎流产量具有显著调控作用.本研究结果显示,树干茎流率与株高、基径、枝条数、冠层体积及冠层投影面积之间均呈显著正相关(P<0.05;图4).一般而言,高大植株因其扩展型冠层结构表现出更强的树干茎流生产能力,这种关联性可能与冠层体积与降水截留效率的正反馈机制有关[23,29].值得注意的是,冠层投影面积对树干茎流产量的影响表现出物种特异性,灌木类群中较大的冠层面积结合较高的植株形态更有利于树干茎流形成[48-49].Park等[50]的研究进一步证实,冠层投影面积较大的乔木与小型个体树干茎流产量存在显著差异(P<0.05).同时,基径和枝条数量对树干茎流产量也具有显著影响.例如,沙坡头地区柠条锦鸡儿的树干茎流产量与株高、基径、枝条数、分枝长均呈显著线性正相关(P<0.05)[51-52];黄土高原典型灌木沙棘(Hippophae rhamnoides)的冠层投影面积、分枝长度、枝条数量和冠层体积均与树干茎流产量呈正相关[37].沙拐枣(Calligonum mongolicum)较大的冠层面积和较长的灌木分枝长度增加树干茎流产量[49].此外,树干茎流率与基部面积呈现正相关关系(图4),但相关性不显著性(P=0.663). ...
Stemflow in three shrubs and its effect on soil water enhancement in semiarid loess region of China
1
2008
... 降水抵达地表之前,植被冠层可视为雨水汇集区,不同物种之间树干茎流产量差异明显.Martinez-Meza等[23]报道,奇瓦瓦沙漠灌木腺牧豆树(Prosopis glandulosa)、三齿团香木(Larrea tridentata)和美洲焦油灌木(Flourensia cernua)树干茎流量分别占降水的5.4%、10%、10.5%;黄土高原多枝柽柳(Tamarix ramosissima)、柠条锦鸡儿(Caragana korshinskii)、红砂(Reaumuria soongorica)的树干茎流量分别占降水的2.2%、3.7%、7.2%[24];科尔沁沙地小叶锦鸡儿(Caragana microphylla)、木岩黄芪(Hedysarum fruticosum)、黄柳(Salix gordejevii)的树干茎流量分别占降水的11%、6.3%、5.3%[25];腾格里沙漠柠条锦鸡儿和油蒿(Artemisia ordosica)的树干茎流量分别占降水的8.8%和2.8%[10].冠层结构参数是导致树干茎流产量存在显著差异的关键因素.Kelly[26]发现树木在无叶期树干茎流产量最高;Levia等[27]发现山毛榉(Fagus sylvatica)幼树的树干茎流产量随叶片数量增加呈上升趋势,但达到一定阈值后趋于稳定;Levia等[27]还发现美国马里兰州的温带森林的树干茎流产量与树干大小显著正相关;Brown等[28]研究表明,相较于树皮易脱落的白橡树,具有紧实树皮的黑橡树树干茎流产量更高;高大的植株往往对应更大冠层投影面积,易产生更多的树干茎流[23,29].然而,对于特定植物种,不同个体之间或同一个体不同分枝之间构型差异如何影响树干茎流产量?不同构型参数重要性次序如何?这些问题目前并不清楚.该方面研究对深入认识干旱区灌木水分蓄集和利用策略具有重要意义. ...
Rainfall partitioning characteristics of three typical sand-fixing shrubs in Horqin Sand Land,North-eastern China
1
2017
... 降水抵达地表之前,植被冠层可视为雨水汇集区,不同物种之间树干茎流产量差异明显.Martinez-Meza等[23]报道,奇瓦瓦沙漠灌木腺牧豆树(Prosopis glandulosa)、三齿团香木(Larrea tridentata)和美洲焦油灌木(Flourensia cernua)树干茎流量分别占降水的5.4%、10%、10.5%;黄土高原多枝柽柳(Tamarix ramosissima)、柠条锦鸡儿(Caragana korshinskii)、红砂(Reaumuria soongorica)的树干茎流量分别占降水的2.2%、3.7%、7.2%[24];科尔沁沙地小叶锦鸡儿(Caragana microphylla)、木岩黄芪(Hedysarum fruticosum)、黄柳(Salix gordejevii)的树干茎流量分别占降水的11%、6.3%、5.3%[25];腾格里沙漠柠条锦鸡儿和油蒿(Artemisia ordosica)的树干茎流量分别占降水的8.8%和2.8%[10].冠层结构参数是导致树干茎流产量存在显著差异的关键因素.Kelly[26]发现树木在无叶期树干茎流产量最高;Levia等[27]发现山毛榉(Fagus sylvatica)幼树的树干茎流产量随叶片数量增加呈上升趋势,但达到一定阈值后趋于稳定;Levia等[27]还发现美国马里兰州的温带森林的树干茎流产量与树干大小显著正相关;Brown等[28]研究表明,相较于树皮易脱落的白橡树,具有紧实树皮的黑橡树树干茎流产量更高;高大的植株往往对应更大冠层投影面积,易产生更多的树干茎流[23,29].然而,对于特定植物种,不同个体之间或同一个体不同分枝之间构型差异如何影响树干茎流产量?不同构型参数重要性次序如何?这些问题目前并不清楚.该方面研究对深入认识干旱区灌木水分蓄集和利用策略具有重要意义. ...
Power plant influences on bulk precipitation,throughfall,and stemflow nutrient inputs
1
1984
... 降水抵达地表之前,植被冠层可视为雨水汇集区,不同物种之间树干茎流产量差异明显.Martinez-Meza等[23]报道,奇瓦瓦沙漠灌木腺牧豆树(Prosopis glandulosa)、三齿团香木(Larrea tridentata)和美洲焦油灌木(Flourensia cernua)树干茎流量分别占降水的5.4%、10%、10.5%;黄土高原多枝柽柳(Tamarix ramosissima)、柠条锦鸡儿(Caragana korshinskii)、红砂(Reaumuria soongorica)的树干茎流量分别占降水的2.2%、3.7%、7.2%[24];科尔沁沙地小叶锦鸡儿(Caragana microphylla)、木岩黄芪(Hedysarum fruticosum)、黄柳(Salix gordejevii)的树干茎流量分别占降水的11%、6.3%、5.3%[25];腾格里沙漠柠条锦鸡儿和油蒿(Artemisia ordosica)的树干茎流量分别占降水的8.8%和2.8%[10].冠层结构参数是导致树干茎流产量存在显著差异的关键因素.Kelly[26]发现树木在无叶期树干茎流产量最高;Levia等[27]发现山毛榉(Fagus sylvatica)幼树的树干茎流产量随叶片数量增加呈上升趋势,但达到一定阈值后趋于稳定;Levia等[27]还发现美国马里兰州的温带森林的树干茎流产量与树干大小显著正相关;Brown等[28]研究表明,相较于树皮易脱落的白橡树,具有紧实树皮的黑橡树树干茎流产量更高;高大的植株往往对应更大冠层投影面积,易产生更多的树干茎流[23,29].然而,对于特定植物种,不同个体之间或同一个体不同分枝之间构型差异如何影响树干茎流产量?不同构型参数重要性次序如何?这些问题目前并不清楚.该方面研究对深入认识干旱区灌木水分蓄集和利用策略具有重要意义. ...
Temporal variability of stemflow volume in a beech-yellow poplar forest in relation to tree species and size
2
2010
... 降水抵达地表之前,植被冠层可视为雨水汇集区,不同物种之间树干茎流产量差异明显.Martinez-Meza等[23]报道,奇瓦瓦沙漠灌木腺牧豆树(Prosopis glandulosa)、三齿团香木(Larrea tridentata)和美洲焦油灌木(Flourensia cernua)树干茎流量分别占降水的5.4%、10%、10.5%;黄土高原多枝柽柳(Tamarix ramosissima)、柠条锦鸡儿(Caragana korshinskii)、红砂(Reaumuria soongorica)的树干茎流量分别占降水的2.2%、3.7%、7.2%[24];科尔沁沙地小叶锦鸡儿(Caragana microphylla)、木岩黄芪(Hedysarum fruticosum)、黄柳(Salix gordejevii)的树干茎流量分别占降水的11%、6.3%、5.3%[25];腾格里沙漠柠条锦鸡儿和油蒿(Artemisia ordosica)的树干茎流量分别占降水的8.8%和2.8%[10].冠层结构参数是导致树干茎流产量存在显著差异的关键因素.Kelly[26]发现树木在无叶期树干茎流产量最高;Levia等[27]发现山毛榉(Fagus sylvatica)幼树的树干茎流产量随叶片数量增加呈上升趋势,但达到一定阈值后趋于稳定;Levia等[27]还发现美国马里兰州的温带森林的树干茎流产量与树干大小显著正相关;Brown等[28]研究表明,相较于树皮易脱落的白橡树,具有紧实树皮的黑橡树树干茎流产量更高;高大的植株往往对应更大冠层投影面积,易产生更多的树干茎流[23,29].然而,对于特定植物种,不同个体之间或同一个体不同分枝之间构型差异如何影响树干茎流产量?不同构型参数重要性次序如何?这些问题目前并不清楚.该方面研究对深入认识干旱区灌木水分蓄集和利用策略具有重要意义. ...
... [27]还发现美国马里兰州的温带森林的树干茎流产量与树干大小显著正相关;Brown等[28]研究表明,相较于树皮易脱落的白橡树,具有紧实树皮的黑橡树树干茎流产量更高;高大的植株往往对应更大冠层投影面积,易产生更多的树干茎流[23,29].然而,对于特定植物种,不同个体之间或同一个体不同分枝之间构型差异如何影响树干茎流产量?不同构型参数重要性次序如何?这些问题目前并不清楚.该方面研究对深入认识干旱区灌木水分蓄集和利用策略具有重要意义. ...
An analysis of throughfall and stemflow in mixed oak stands
2
1970
... 降水抵达地表之前,植被冠层可视为雨水汇集区,不同物种之间树干茎流产量差异明显.Martinez-Meza等[23]报道,奇瓦瓦沙漠灌木腺牧豆树(Prosopis glandulosa)、三齿团香木(Larrea tridentata)和美洲焦油灌木(Flourensia cernua)树干茎流量分别占降水的5.4%、10%、10.5%;黄土高原多枝柽柳(Tamarix ramosissima)、柠条锦鸡儿(Caragana korshinskii)、红砂(Reaumuria soongorica)的树干茎流量分别占降水的2.2%、3.7%、7.2%[24];科尔沁沙地小叶锦鸡儿(Caragana microphylla)、木岩黄芪(Hedysarum fruticosum)、黄柳(Salix gordejevii)的树干茎流量分别占降水的11%、6.3%、5.3%[25];腾格里沙漠柠条锦鸡儿和油蒿(Artemisia ordosica)的树干茎流量分别占降水的8.8%和2.8%[10].冠层结构参数是导致树干茎流产量存在显著差异的关键因素.Kelly[26]发现树木在无叶期树干茎流产量最高;Levia等[27]发现山毛榉(Fagus sylvatica)幼树的树干茎流产量随叶片数量增加呈上升趋势,但达到一定阈值后趋于稳定;Levia等[27]还发现美国马里兰州的温带森林的树干茎流产量与树干大小显著正相关;Brown等[28]研究表明,相较于树皮易脱落的白橡树,具有紧实树皮的黑橡树树干茎流产量更高;高大的植株往往对应更大冠层投影面积,易产生更多的树干茎流[23,29].然而,对于特定植物种,不同个体之间或同一个体不同分枝之间构型差异如何影响树干茎流产量?不同构型参数重要性次序如何?这些问题目前并不清楚.该方面研究对深入认识干旱区灌木水分蓄集和利用策略具有重要意义. ...
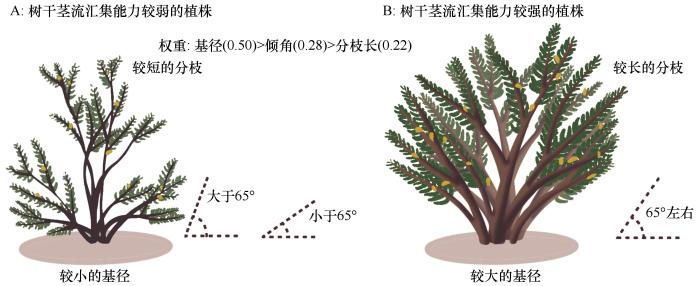
... 树干茎流产生过程并非受单一因素主导,而是多因素综合作用的结果.现有研究多聚焦于不同灌木种间形态参数差异分析[28,50],或分析单一或少数冠层结构参数对树干茎流产量的影响[37,50-51].Honda等[53]和Whitworth-Hulse等[54]研究表明,具有更多分枝且分枝垂直的植物具有更高的树干茎流产流优势;Martinez-Meza[55]发现,当枝条倾角大于45°时,美洲焦油灌木产生的树干茎流量要大于水平角度的枝条,而腺牧豆树在主干角度为70°~75°时具有最大的树干茎流量;泡泡刺(Nitraria sphaerocarpa)树干茎流量随着枝条倾角增加而增加[56].本研究去掉了对树干茎流产量影响较小的分枝形态参数,进而使用熵权法对灌木分枝雨水汇集能力进行了综合评估,依据综合权重值得出对树干茎流产量具有显著作用的关键分枝形态参数为:基径(0.50)>倾角(0.28)>长度(0.22).这说明基径较粗、分枝较长且分枝倾角接近65°的植株具有更高的树干茎流生产效率(图5),这种冠层形态特征可形成更优的雨水截留界面和引流通路.Yuan等[57]在黄土高原开展的研究也表明较粗基径和较长分枝结构可增加沙柳树干茎流产量.本研究量化分析了灌木分枝构型参数对树干茎流产量的影响作用,有助于深入认识干旱区灌木的水分蓄集和利用策略. ...
The effects of canopy structure on stemflow,throughfall and interception loss in a young Sitka spruce plantation
3
1978
... 降水抵达地表之前,植被冠层可视为雨水汇集区,不同物种之间树干茎流产量差异明显.Martinez-Meza等[23]报道,奇瓦瓦沙漠灌木腺牧豆树(Prosopis glandulosa)、三齿团香木(Larrea tridentata)和美洲焦油灌木(Flourensia cernua)树干茎流量分别占降水的5.4%、10%、10.5%;黄土高原多枝柽柳(Tamarix ramosissima)、柠条锦鸡儿(Caragana korshinskii)、红砂(Reaumuria soongorica)的树干茎流量分别占降水的2.2%、3.7%、7.2%[24];科尔沁沙地小叶锦鸡儿(Caragana microphylla)、木岩黄芪(Hedysarum fruticosum)、黄柳(Salix gordejevii)的树干茎流量分别占降水的11%、6.3%、5.3%[25];腾格里沙漠柠条锦鸡儿和油蒿(Artemisia ordosica)的树干茎流量分别占降水的8.8%和2.8%[10].冠层结构参数是导致树干茎流产量存在显著差异的关键因素.Kelly[26]发现树木在无叶期树干茎流产量最高;Levia等[27]发现山毛榉(Fagus sylvatica)幼树的树干茎流产量随叶片数量增加呈上升趋势,但达到一定阈值后趋于稳定;Levia等[27]还发现美国马里兰州的温带森林的树干茎流产量与树干大小显著正相关;Brown等[28]研究表明,相较于树皮易脱落的白橡树,具有紧实树皮的黑橡树树干茎流产量更高;高大的植株往往对应更大冠层投影面积,易产生更多的树干茎流[23,29].然而,对于特定植物种,不同个体之间或同一个体不同分枝之间构型差异如何影响树干茎流产量?不同构型参数重要性次序如何?这些问题目前并不清楚.该方面研究对深入认识干旱区灌木水分蓄集和利用策略具有重要意义. ...
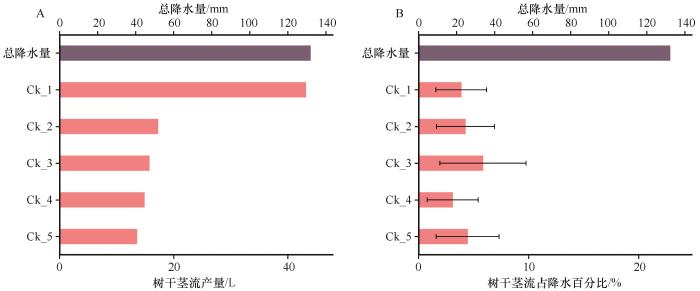
... 本研究得出,柠条锦鸡儿产生的树干茎流量占降水量的3.6%.Zhang等[6]研究表明全球灌木树干茎流占降水比例的中位值为7.2%(四分位距:5.2%~11.9%),不同灌木种间树干茎流产量差异较大.比如,厄瓜多尔加拉帕戈斯地区的罗宾逊绢木(Miconia robinsoniana)树干茎流百分比仅为0.6%[42],美国费城地区北美鼠刺(Itea virginica)树干茎流百分比为4.4%[43],陕西神木六道沟流域沙柳(Salix psammophila)树干茎流百分比为6.3%[44],西班牙阿尔梅里亚地区脱皮腺果豆(Adenocarpus decorticans)树干茎流百分比为4.4%[45],毛乌素沙地的沙柳树干茎流百分比9.0%[46],青藏高原东北部地区金路梅(Potentilla fruticosa)树干茎流百分比高达49.3%[47].树干茎流占降水比例较低,但由于灌木表现出的漏斗效应,灌木基部树干茎流的汇流量大.例如,Ck_1树干茎流量达43.2 L(图3),树干茎流可进一步快速通过植物根系和土壤大孔隙形成优先流补给深层土壤,在植物根际区形成水分沃岛,这对旱区植物生存及其遭受水分胁迫时增强其环境适应能力和提高水分利用效率具有重要影响[29]. ...
... 现有研究表明,冠层结构参数(株高、基径、基部面积、枝条数、冠层体积及冠层投影面积等)对树干茎流产量具有显著调控作用.本研究结果显示,树干茎流率与株高、基径、枝条数、冠层体积及冠层投影面积之间均呈显著正相关(P<0.05;图4).一般而言,高大植株因其扩展型冠层结构表现出更强的树干茎流生产能力,这种关联性可能与冠层体积与降水截留效率的正反馈机制有关[23,29].值得注意的是,冠层投影面积对树干茎流产量的影响表现出物种特异性,灌木类群中较大的冠层面积结合较高的植株形态更有利于树干茎流形成[48-49].Park等[50]的研究进一步证实,冠层投影面积较大的乔木与小型个体树干茎流产量存在显著差异(P<0.05).同时,基径和枝条数量对树干茎流产量也具有显著影响.例如,沙坡头地区柠条锦鸡儿的树干茎流产量与株高、基径、枝条数、分枝长均呈显著线性正相关(P<0.05)[51-52];黄土高原典型灌木沙棘(Hippophae rhamnoides)的冠层投影面积、分枝长度、枝条数量和冠层体积均与树干茎流产量呈正相关[37].沙拐枣(Calligonum mongolicum)较大的冠层面积和较长的灌木分枝长度增加树干茎流产量[49].此外,树干茎流率与基部面积呈现正相关关系(图4),但相关性不显著性(P=0.663). ...
平茬措施对柠条生理特征及土壤水分的影响
2
2012
... 柠条锦鸡儿是具有极强抗逆性的豆科灌木[30],生态适应性广,能在贫瘠沙地、盐碱地和干旱山坡等恶劣环境中生长,是中国北方植被恢复实践中最具代表性的耐旱灌木[31].其在中国主要分布于黄土高原、内蒙古高原及西北荒漠草原区,是干旱区生态系统的关键建群种植物,具有生态适应性强、根系发达、吸水力强等特点[30],在防风固沙、保持水土等方面具有重要生态意义.本研究对腾格里沙漠东南缘沙坡头地区典型固沙灌木柠条锦鸡儿树干茎流进行野外观测,量化灌木枝干结构特征与树干茎流产量的关系,并确定影响树干茎流产量的关键冠层结构指标及其相对重要性.分析灌木分枝构型对树干茎流产流的影响机制,为理解植物形态特征与冠层水文过程的关系提供新的视角,也为评估种内个体形态结构参数对树干茎流产量的影响提供方法参考. ...
... [30],在防风固沙、保持水土等方面具有重要生态意义.本研究对腾格里沙漠东南缘沙坡头地区典型固沙灌木柠条锦鸡儿树干茎流进行野外观测,量化灌木枝干结构特征与树干茎流产量的关系,并确定影响树干茎流产量的关键冠层结构指标及其相对重要性.分析灌木分枝构型对树干茎流产流的影响机制,为理解植物形态特征与冠层水文过程的关系提供新的视角,也为评估种内个体形态结构参数对树干茎流产量的影响提供方法参考. ...
Alteration in isotopic composition of gross rainfall as it is being partitioned into throughfall and stemflow by xerophytic shrub canopies within water-limited arid desert ecosystems
1
2019
... 柠条锦鸡儿是具有极强抗逆性的豆科灌木[30],生态适应性广,能在贫瘠沙地、盐碱地和干旱山坡等恶劣环境中生长,是中国北方植被恢复实践中最具代表性的耐旱灌木[31].其在中国主要分布于黄土高原、内蒙古高原及西北荒漠草原区,是干旱区生态系统的关键建群种植物,具有生态适应性强、根系发达、吸水力强等特点[30],在防风固沙、保持水土等方面具有重要生态意义.本研究对腾格里沙漠东南缘沙坡头地区典型固沙灌木柠条锦鸡儿树干茎流进行野外观测,量化灌木枝干结构特征与树干茎流产量的关系,并确定影响树干茎流产量的关键冠层结构指标及其相对重要性.分析灌木分枝构型对树干茎流产流的影响机制,为理解植物形态特征与冠层水文过程的关系提供新的视角,也为评估种内个体形态结构参数对树干茎流产量的影响提供方法参考. ...
1
2012
... 试验于2024年8—10月在位于宁夏回族自治区中卫市的中国科学院沙坡头沙漠研究试验站水量平衡综合观测场进行(图1).该区地处腾格里沙漠东南缘(37°27′N,105°57′E),海拔1 300 m,属于草原化荒漠,是荒漠与草原的过渡区域.多年平均降水量为188 mm,其中80%的降水发生在7—9月,多年平均潜在蒸发量高达2 500 mm[32].温度变幅较大,年平均气温9.6 ℃,1月平均气温-6.1 ℃,7月平均气温24.7 ℃.盛行西北风,年平均风速2.9~3.5 m·s-1[33-35].土壤类型为沙丘风沙土[35]. ...
Relative contribution of biotic and abiotic factors to stemflow production and funneling efficiency:a long-term field study on a xerophytic shrub species in Tengger Desert of northern China
1
2020
... 试验于2024年8—10月在位于宁夏回族自治区中卫市的中国科学院沙坡头沙漠研究试验站水量平衡综合观测场进行(图1).该区地处腾格里沙漠东南缘(37°27′N,105°57′E),海拔1 300 m,属于草原化荒漠,是荒漠与草原的过渡区域.多年平均降水量为188 mm,其中80%的降水发生在7—9月,多年平均潜在蒸发量高达2 500 mm[32].温度变幅较大,年平均气温9.6 ℃,1月平均气温-6.1 ℃,7月平均气温24.7 ℃.盛行西北风,年平均风速2.9~3.5 m·s-1[33-35].土壤类型为沙丘风沙土[35]. ...
Variation in soil seed banks composition at the desert microhabitats of Caragana korshinskii shrubs
2010
荒漠灌丛降雨再分配对土壤pH值的影响
3
2013
... 试验于2024年8—10月在位于宁夏回族自治区中卫市的中国科学院沙坡头沙漠研究试验站水量平衡综合观测场进行(图1).该区地处腾格里沙漠东南缘(37°27′N,105°57′E),海拔1 300 m,属于草原化荒漠,是荒漠与草原的过渡区域.多年平均降水量为188 mm,其中80%的降水发生在7—9月,多年平均潜在蒸发量高达2 500 mm[32].温度变幅较大,年平均气温9.6 ℃,1月平均气温-6.1 ℃,7月平均气温24.7 ℃.盛行西北风,年平均风速2.9~3.5 m·s-1[33-35].土壤类型为沙丘风沙土[35]. ...
... [35]. ...
... 树干茎流作用不仅与水文过程密切相关,在生态系统养分循环中也扮演着重要角色,是生态系统养分循环的重要驱动因子之一[35,58].大气降水与干沉降过程在冠层交换作用下溶解的气溶胶颗粒物、植物冠层的干沉降物质,存在于植物冠层中间的生产者、消费者和分解者的排泄物与残体,在大气降水的溶解和冲刷作用下,随树干茎流汇集至植物基部,形成灌木沃岛效应[58].本文主要聚焦于柠条锦鸡儿树干茎流的水文过程效应,未来研究将系统探讨养分离子的动态变化特征及其富集规律,以期全面揭示树干茎流在干旱生态系统水分和养分循环中的作用. ...
Have we underestimated stemflow?Lessons from an open tropical rainforest
1
2010
... 柠条锦鸡儿是豆科锦鸡儿属的多年生落叶灌木,多茎干且其表面光滑,冠层呈倒锥形(图1C).叶片为羽状复叶,对生或近对生(长6~10 cm)排列,每片复叶有5~8对卵形小叶(长7~8 mm,宽2~5 mm)[36].本研究选取5株大小不同的柠条锦鸡儿进行树干茎流测量,其形态结构参数如表1所列. ...
Characteristics of Caragana korshinskii and Hippophae rhamnoides stemflow and their significance in soil moisture enhancement in Loess Plateau,China
3
2014
... 树干茎流收集采用铝箔槽导流法[37-38](图1C、1D),具体步骤如下:在柠条锦鸡儿树干基部用铝箔胶纸紧密缠绕两圈确保无空隙;接着,剪取一段铝箔胶纸,沿长度方向对折1/3,并将下1/3的部分沿枝干表面紧紧环绕粘贴,形成一个完全环绕枝干的圆形铝箔水槽;随后,用软管将铝箔水槽与聚乙烯收集容器连接;最后,在铝箔水槽外表面再缠绕至少3层铝箔胶纸加固,形成缠绕枝干呈烟斗状的树干茎流导流水槽.收集测量得到树干茎流产量(L). ...
... 现有研究表明,冠层结构参数(株高、基径、基部面积、枝条数、冠层体积及冠层投影面积等)对树干茎流产量具有显著调控作用.本研究结果显示,树干茎流率与株高、基径、枝条数、冠层体积及冠层投影面积之间均呈显著正相关(P<0.05;图4).一般而言,高大植株因其扩展型冠层结构表现出更强的树干茎流生产能力,这种关联性可能与冠层体积与降水截留效率的正反馈机制有关[23,29].值得注意的是,冠层投影面积对树干茎流产量的影响表现出物种特异性,灌木类群中较大的冠层面积结合较高的植株形态更有利于树干茎流形成[48-49].Park等[50]的研究进一步证实,冠层投影面积较大的乔木与小型个体树干茎流产量存在显著差异(P<0.05).同时,基径和枝条数量对树干茎流产量也具有显著影响.例如,沙坡头地区柠条锦鸡儿的树干茎流产量与株高、基径、枝条数、分枝长均呈显著线性正相关(P<0.05)[51-52];黄土高原典型灌木沙棘(Hippophae rhamnoides)的冠层投影面积、分枝长度、枝条数量和冠层体积均与树干茎流产量呈正相关[37].沙拐枣(Calligonum mongolicum)较大的冠层面积和较长的灌木分枝长度增加树干茎流产量[49].此外,树干茎流率与基部面积呈现正相关关系(图4),但相关性不显著性(P=0.663). ...
... 树干茎流产生过程并非受单一因素主导,而是多因素综合作用的结果.现有研究多聚焦于不同灌木种间形态参数差异分析[28,50],或分析单一或少数冠层结构参数对树干茎流产量的影响[37,50-51].Honda等[53]和Whitworth-Hulse等[54]研究表明,具有更多分枝且分枝垂直的植物具有更高的树干茎流产流优势;Martinez-Meza[55]发现,当枝条倾角大于45°时,美洲焦油灌木产生的树干茎流量要大于水平角度的枝条,而腺牧豆树在主干角度为70°~75°时具有最大的树干茎流量;泡泡刺(Nitraria sphaerocarpa)树干茎流量随着枝条倾角增加而增加[56].本研究去掉了对树干茎流产量影响较小的分枝形态参数,进而使用熵权法对灌木分枝雨水汇集能力进行了综合评估,依据综合权重值得出对树干茎流产量具有显著作用的关键分枝形态参数为:基径(0.50)>倾角(0.28)>长度(0.22).这说明基径较粗、分枝较长且分枝倾角接近65°的植株具有更高的树干茎流生产效率(图5),这种冠层形态特征可形成更优的雨水截留界面和引流通路.Yuan等[57]在黄土高原开展的研究也表明较粗基径和较长分枝结构可增加沙柳树干茎流产量.本研究量化分析了灌木分枝构型参数对树干茎流产量的影响作用,有助于深入认识干旱区灌木的水分蓄集和利用策略. ...
The measurement and modelling of stemflow in an alpine Myricaria squamosa community
1
2015
... 树干茎流收集采用铝箔槽导流法[37-38](图1C、1D),具体步骤如下:在柠条锦鸡儿树干基部用铝箔胶纸紧密缠绕两圈确保无空隙;接着,剪取一段铝箔胶纸,沿长度方向对折1/3,并将下1/3的部分沿枝干表面紧紧环绕粘贴,形成一个完全环绕枝干的圆形铝箔水槽;随后,用软管将铝箔水槽与聚乙烯收集容器连接;最后,在铝箔水槽外表面再缠绕至少3层铝箔胶纸加固,形成缠绕枝干呈烟斗状的树干茎流导流水槽.收集测量得到树干茎流产量(L). ...
Rainfall partitioning into throughfall,stemflow and interception loss by two xerophytic shrubs within a rain-fed re-vegetated desert ecosystem,Northwestern China
2
2015
... 树干茎流量计算方法为[39]: ...
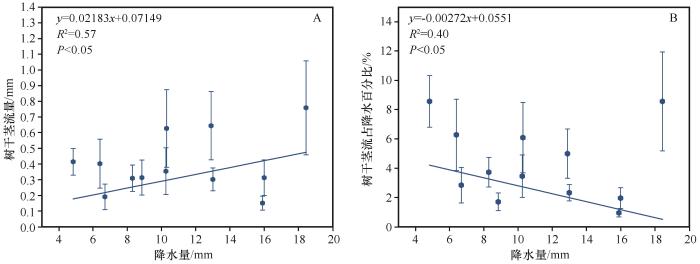
... 为研究不同植株冠层形态差异对树干茎流产量的影响,本研究使用树干茎流率(SfVPR)[39-40],即单位降水产生的树干茎流产量(mL·mm-1),来分析树干茎流产量与冠层参数之间的关系.相较于传统采用绝对体积(mL)或等效水深(mm)的表征方法,该参数具有以下显著优势[40]:首先,该参数通过标准化处理消除了降水量变化对树干茎流产量的影响,有效解决了以往研究中因忽视降水量影响而导致的缺陷;其次,该复合单位(mL·mm-1)具有明确的物理意义,且因其标准化特性易于在水文模型中应用. ...
Stemflow volume per unit rainfall as a good variable to determine the relationship between stemflow amount and morphological metrics of shrubs
4
2017
... 为研究不同植株冠层形态差异对树干茎流产量的影响,本研究使用树干茎流率(SfVPR)[39-40],即单位降水产生的树干茎流产量(mL·mm-1),来分析树干茎流产量与冠层参数之间的关系.相较于传统采用绝对体积(mL)或等效水深(mm)的表征方法,该参数具有以下显著优势[40]:首先,该参数通过标准化处理消除了降水量变化对树干茎流产量的影响,有效解决了以往研究中因忽视降水量影响而导致的缺陷;其次,该复合单位(mL·mm-1)具有明确的物理意义,且因其标准化特性易于在水文模型中应用. ...
... [40]:首先,该参数通过标准化处理消除了降水量变化对树干茎流产量的影响,有效解决了以往研究中因忽视降水量影响而导致的缺陷;其次,该复合单位(mL·mm-1)具有明确的物理意义,且因其标准化特性易于在水文模型中应用. ...
... 树干茎流率计算方法为[40-41]: ...
... 参考Zhang等[40]关于柠条锦鸡儿树干茎流量与树枝倾角呈抛物线关系的研究结果,当倾角为65°时,树干茎流产量达到峰值,本研究以65°倾角为界分析分枝特征与树干茎流产量的关系.树干茎流产量与分枝基径、分枝倾角和分枝长度均呈显著正相关(P<0.05),而其与分枝朝向和枝下高的相关关系不显著(P>0.05,表2). ...
The stemflow of trees in a Bornean lowland tropical forest
1
2004
... 树干茎流率计算方法为[40-41]: ...
Quantification of cloud water interception along the windward slope of Santa Cruz Island,Galapagos (Ecuador)
1
2012
... 本研究得出,柠条锦鸡儿产生的树干茎流量占降水量的3.6%.Zhang等[6]研究表明全球灌木树干茎流占降水比例的中位值为7.2%(四分位距:5.2%~11.9%),不同灌木种间树干茎流产量差异较大.比如,厄瓜多尔加拉帕戈斯地区的罗宾逊绢木(Miconia robinsoniana)树干茎流百分比仅为0.6%[42],美国费城地区北美鼠刺(Itea virginica)树干茎流百分比为4.4%[43],陕西神木六道沟流域沙柳(Salix psammophila)树干茎流百分比为6.3%[44],西班牙阿尔梅里亚地区脱皮腺果豆(Adenocarpus decorticans)树干茎流百分比为4.4%[45],毛乌素沙地的沙柳树干茎流百分比9.0%[46],青藏高原东北部地区金路梅(Potentilla fruticosa)树干茎流百分比高达49.3%[47].树干茎流占降水比例较低,但由于灌木表现出的漏斗效应,灌木基部树干茎流的汇流量大.例如,Ck_1树干茎流量达43.2 L(图3),树干茎流可进一步快速通过植物根系和土壤大孔隙形成优先流补给深层土壤,在植物根际区形成水分沃岛,这对旱区植物生存及其遭受水分胁迫时增强其环境适应能力和提高水分利用效率具有重要影响[29]. ...
Quantification of stemflow in three isolated shrub species in an urban environment
1
2019
... 本研究得出,柠条锦鸡儿产生的树干茎流量占降水量的3.6%.Zhang等[6]研究表明全球灌木树干茎流占降水比例的中位值为7.2%(四分位距:5.2%~11.9%),不同灌木种间树干茎流产量差异较大.比如,厄瓜多尔加拉帕戈斯地区的罗宾逊绢木(Miconia robinsoniana)树干茎流百分比仅为0.6%[42],美国费城地区北美鼠刺(Itea virginica)树干茎流百分比为4.4%[43],陕西神木六道沟流域沙柳(Salix psammophila)树干茎流百分比为6.3%[44],西班牙阿尔梅里亚地区脱皮腺果豆(Adenocarpus decorticans)树干茎流百分比为4.4%[45],毛乌素沙地的沙柳树干茎流百分比9.0%[46],青藏高原东北部地区金路梅(Potentilla fruticosa)树干茎流百分比高达49.3%[47].树干茎流占降水比例较低,但由于灌木表现出的漏斗效应,灌木基部树干茎流的汇流量大.例如,Ck_1树干茎流量达43.2 L(图3),树干茎流可进一步快速通过植物根系和土壤大孔隙形成优先流补给深层土壤,在植物根际区形成水分沃岛,这对旱区植物生存及其遭受水分胁迫时增强其环境适应能力和提高水分利用效率具有重要影响[29]. ...
Stemflow production differ significantly among tree and shrub species on the Chinese Loess Plateau
1
2019
... 本研究得出,柠条锦鸡儿产生的树干茎流量占降水量的3.6%.Zhang等[6]研究表明全球灌木树干茎流占降水比例的中位值为7.2%(四分位距:5.2%~11.9%),不同灌木种间树干茎流产量差异较大.比如,厄瓜多尔加拉帕戈斯地区的罗宾逊绢木(Miconia robinsoniana)树干茎流百分比仅为0.6%[42],美国费城地区北美鼠刺(Itea virginica)树干茎流百分比为4.4%[43],陕西神木六道沟流域沙柳(Salix psammophila)树干茎流百分比为6.3%[44],西班牙阿尔梅里亚地区脱皮腺果豆(Adenocarpus decorticans)树干茎流百分比为4.4%[45],毛乌素沙地的沙柳树干茎流百分比9.0%[46],青藏高原东北部地区金路梅(Potentilla fruticosa)树干茎流百分比高达49.3%[47].树干茎流占降水比例较低,但由于灌木表现出的漏斗效应,灌木基部树干茎流的汇流量大.例如,Ck_1树干茎流量达43.2 L(图3),树干茎流可进一步快速通过植物根系和土壤大孔隙形成优先流补给深层土壤,在植物根际区形成水分沃岛,这对旱区植物生存及其遭受水分胁迫时增强其环境适应能力和提高水分利用效率具有重要影响[29]. ...
Role of vegetation cover in the biogeochemical balances of a small afforested catchment in Southeastern Spain
1
1994
... 本研究得出,柠条锦鸡儿产生的树干茎流量占降水量的3.6%.Zhang等[6]研究表明全球灌木树干茎流占降水比例的中位值为7.2%(四分位距:5.2%~11.9%),不同灌木种间树干茎流产量差异较大.比如,厄瓜多尔加拉帕戈斯地区的罗宾逊绢木(Miconia robinsoniana)树干茎流百分比仅为0.6%[42],美国费城地区北美鼠刺(Itea virginica)树干茎流百分比为4.4%[43],陕西神木六道沟流域沙柳(Salix psammophila)树干茎流百分比为6.3%[44],西班牙阿尔梅里亚地区脱皮腺果豆(Adenocarpus decorticans)树干茎流百分比为4.4%[45],毛乌素沙地的沙柳树干茎流百分比9.0%[46],青藏高原东北部地区金路梅(Potentilla fruticosa)树干茎流百分比高达49.3%[47].树干茎流占降水比例较低,但由于灌木表现出的漏斗效应,灌木基部树干茎流的汇流量大.例如,Ck_1树干茎流量达43.2 L(图3),树干茎流可进一步快速通过植物根系和土壤大孔隙形成优先流补给深层土壤,在植物根际区形成水分沃岛,这对旱区植物生存及其遭受水分胁迫时增强其环境适应能力和提高水分利用效率具有重要影响[29]. ...
毛乌素沙地沙柳灌丛降雨截留与树干茎流特征
1
2008
... 本研究得出,柠条锦鸡儿产生的树干茎流量占降水量的3.6%.Zhang等[6]研究表明全球灌木树干茎流占降水比例的中位值为7.2%(四分位距:5.2%~11.9%),不同灌木种间树干茎流产量差异较大.比如,厄瓜多尔加拉帕戈斯地区的罗宾逊绢木(Miconia robinsoniana)树干茎流百分比仅为0.6%[42],美国费城地区北美鼠刺(Itea virginica)树干茎流百分比为4.4%[43],陕西神木六道沟流域沙柳(Salix psammophila)树干茎流百分比为6.3%[44],西班牙阿尔梅里亚地区脱皮腺果豆(Adenocarpus decorticans)树干茎流百分比为4.4%[45],毛乌素沙地的沙柳树干茎流百分比9.0%[46],青藏高原东北部地区金路梅(Potentilla fruticosa)树干茎流百分比高达49.3%[47].树干茎流占降水比例较低,但由于灌木表现出的漏斗效应,灌木基部树干茎流的汇流量大.例如,Ck_1树干茎流量达43.2 L(图3),树干茎流可进一步快速通过植物根系和土壤大孔隙形成优先流补给深层土壤,在植物根际区形成水分沃岛,这对旱区植物生存及其遭受水分胁迫时增强其环境适应能力和提高水分利用效率具有重要影响[29]. ...
Modelling of rainfall partitioning by a deciduous shrub using a variable parameters Gash model
1
2018
... 本研究得出,柠条锦鸡儿产生的树干茎流量占降水量的3.6%.Zhang等[6]研究表明全球灌木树干茎流占降水比例的中位值为7.2%(四分位距:5.2%~11.9%),不同灌木种间树干茎流产量差异较大.比如,厄瓜多尔加拉帕戈斯地区的罗宾逊绢木(Miconia robinsoniana)树干茎流百分比仅为0.6%[42],美国费城地区北美鼠刺(Itea virginica)树干茎流百分比为4.4%[43],陕西神木六道沟流域沙柳(Salix psammophila)树干茎流百分比为6.3%[44],西班牙阿尔梅里亚地区脱皮腺果豆(Adenocarpus decorticans)树干茎流百分比为4.4%[45],毛乌素沙地的沙柳树干茎流百分比9.0%[46],青藏高原东北部地区金路梅(Potentilla fruticosa)树干茎流百分比高达49.3%[47].树干茎流占降水比例较低,但由于灌木表现出的漏斗效应,灌木基部树干茎流的汇流量大.例如,Ck_1树干茎流量达43.2 L(图3),树干茎流可进一步快速通过植物根系和土壤大孔隙形成优先流补给深层土壤,在植物根际区形成水分沃岛,这对旱区植物生存及其遭受水分胁迫时增强其环境适应能力和提高水分利用效率具有重要影响[29]. ...
干旱地区土壤-植被-水文耦合、响应与适应机制
1
2011
... 现有研究表明,冠层结构参数(株高、基径、基部面积、枝条数、冠层体积及冠层投影面积等)对树干茎流产量具有显著调控作用.本研究结果显示,树干茎流率与株高、基径、枝条数、冠层体积及冠层投影面积之间均呈显著正相关(P<0.05;图4).一般而言,高大植株因其扩展型冠层结构表现出更强的树干茎流生产能力,这种关联性可能与冠层体积与降水截留效率的正反馈机制有关[23,29].值得注意的是,冠层投影面积对树干茎流产量的影响表现出物种特异性,灌木类群中较大的冠层面积结合较高的植株形态更有利于树干茎流形成[48-49].Park等[50]的研究进一步证实,冠层投影面积较大的乔木与小型个体树干茎流产量存在显著差异(P<0.05).同时,基径和枝条数量对树干茎流产量也具有显著影响.例如,沙坡头地区柠条锦鸡儿的树干茎流产量与株高、基径、枝条数、分枝长均呈显著线性正相关(P<0.05)[51-52];黄土高原典型灌木沙棘(Hippophae rhamnoides)的冠层投影面积、分枝长度、枝条数量和冠层体积均与树干茎流产量呈正相关[37].沙拐枣(Calligonum mongolicum)较大的冠层面积和较长的灌木分枝长度增加树干茎流产量[49].此外,树干茎流率与基部面积呈现正相关关系(图4),但相关性不显著性(P=0.663). ...
Connecting ecohydrology and hydropedology in desert shrubs:stemflow as a source of preferential flow in soils
2
2009
... 现有研究表明,冠层结构参数(株高、基径、基部面积、枝条数、冠层体积及冠层投影面积等)对树干茎流产量具有显著调控作用.本研究结果显示,树干茎流率与株高、基径、枝条数、冠层体积及冠层投影面积之间均呈显著正相关(P<0.05;图4).一般而言,高大植株因其扩展型冠层结构表现出更强的树干茎流生产能力,这种关联性可能与冠层体积与降水截留效率的正反馈机制有关[23,29].值得注意的是,冠层投影面积对树干茎流产量的影响表现出物种特异性,灌木类群中较大的冠层面积结合较高的植株形态更有利于树干茎流形成[48-49].Park等[50]的研究进一步证实,冠层投影面积较大的乔木与小型个体树干茎流产量存在显著差异(P<0.05).同时,基径和枝条数量对树干茎流产量也具有显著影响.例如,沙坡头地区柠条锦鸡儿的树干茎流产量与株高、基径、枝条数、分枝长均呈显著线性正相关(P<0.05)[51-52];黄土高原典型灌木沙棘(Hippophae rhamnoides)的冠层投影面积、分枝长度、枝条数量和冠层体积均与树干茎流产量呈正相关[37].沙拐枣(Calligonum mongolicum)较大的冠层面积和较长的灌木分枝长度增加树干茎流产量[49].此外,树干茎流率与基部面积呈现正相关关系(图4),但相关性不显著性(P=0.663). ...
... [49].此外,树干茎流率与基部面积呈现正相关关系(图4),但相关性不显著性(P=0.663). ...
Applicability of stand structural characteristics to stemflow modeling
3
2002
... 现有研究表明,冠层结构参数(株高、基径、基部面积、枝条数、冠层体积及冠层投影面积等)对树干茎流产量具有显著调控作用.本研究结果显示,树干茎流率与株高、基径、枝条数、冠层体积及冠层投影面积之间均呈显著正相关(P<0.05;图4).一般而言,高大植株因其扩展型冠层结构表现出更强的树干茎流生产能力,这种关联性可能与冠层体积与降水截留效率的正反馈机制有关[23,29].值得注意的是,冠层投影面积对树干茎流产量的影响表现出物种特异性,灌木类群中较大的冠层面积结合较高的植株形态更有利于树干茎流形成[48-49].Park等[50]的研究进一步证实,冠层投影面积较大的乔木与小型个体树干茎流产量存在显著差异(P<0.05).同时,基径和枝条数量对树干茎流产量也具有显著影响.例如,沙坡头地区柠条锦鸡儿的树干茎流产量与株高、基径、枝条数、分枝长均呈显著线性正相关(P<0.05)[51-52];黄土高原典型灌木沙棘(Hippophae rhamnoides)的冠层投影面积、分枝长度、枝条数量和冠层体积均与树干茎流产量呈正相关[37].沙拐枣(Calligonum mongolicum)较大的冠层面积和较长的灌木分枝长度增加树干茎流产量[49].此外,树干茎流率与基部面积呈现正相关关系(图4),但相关性不显著性(P=0.663). ...
... 树干茎流产生过程并非受单一因素主导,而是多因素综合作用的结果.现有研究多聚焦于不同灌木种间形态参数差异分析[28,50],或分析单一或少数冠层结构参数对树干茎流产量的影响[37,50-51].Honda等[53]和Whitworth-Hulse等[54]研究表明,具有更多分枝且分枝垂直的植物具有更高的树干茎流产流优势;Martinez-Meza[55]发现,当枝条倾角大于45°时,美洲焦油灌木产生的树干茎流量要大于水平角度的枝条,而腺牧豆树在主干角度为70°~75°时具有最大的树干茎流量;泡泡刺(Nitraria sphaerocarpa)树干茎流量随着枝条倾角增加而增加[56].本研究去掉了对树干茎流产量影响较小的分枝形态参数,进而使用熵权法对灌木分枝雨水汇集能力进行了综合评估,依据综合权重值得出对树干茎流产量具有显著作用的关键分枝形态参数为:基径(0.50)>倾角(0.28)>长度(0.22).这说明基径较粗、分枝较长且分枝倾角接近65°的植株具有更高的树干茎流生产效率(图5),这种冠层形态特征可形成更优的雨水截留界面和引流通路.Yuan等[57]在黄土高原开展的研究也表明较粗基径和较长分枝结构可增加沙柳树干茎流产量.本研究量化分析了灌木分枝构型参数对树干茎流产量的影响作用,有助于深入认识干旱区灌木的水分蓄集和利用策略. ...
... ,50-51].Honda等[53]和Whitworth-Hulse等[54]研究表明,具有更多分枝且分枝垂直的植物具有更高的树干茎流产流优势;Martinez-Meza[55]发现,当枝条倾角大于45°时,美洲焦油灌木产生的树干茎流量要大于水平角度的枝条,而腺牧豆树在主干角度为70°~75°时具有最大的树干茎流量;泡泡刺(Nitraria sphaerocarpa)树干茎流量随着枝条倾角增加而增加[56].本研究去掉了对树干茎流产量影响较小的分枝形态参数,进而使用熵权法对灌木分枝雨水汇集能力进行了综合评估,依据综合权重值得出对树干茎流产量具有显著作用的关键分枝形态参数为:基径(0.50)>倾角(0.28)>长度(0.22).这说明基径较粗、分枝较长且分枝倾角接近65°的植株具有更高的树干茎流生产效率(图5),这种冠层形态特征可形成更优的雨水截留界面和引流通路.Yuan等[57]在黄土高原开展的研究也表明较粗基径和较长分枝结构可增加沙柳树干茎流产量.本研究量化分析了灌木分枝构型参数对树干茎流产量的影响作用,有助于深入认识干旱区灌木的水分蓄集和利用策略. ...
Differential intra-specific stemflow funnelling efficiencies of Caragana korshinskii within arid desert ecosystems
2
2017
... 现有研究表明,冠层结构参数(株高、基径、基部面积、枝条数、冠层体积及冠层投影面积等)对树干茎流产量具有显著调控作用.本研究结果显示,树干茎流率与株高、基径、枝条数、冠层体积及冠层投影面积之间均呈显著正相关(P<0.05;图4).一般而言,高大植株因其扩展型冠层结构表现出更强的树干茎流生产能力,这种关联性可能与冠层体积与降水截留效率的正反馈机制有关[23,29].值得注意的是,冠层投影面积对树干茎流产量的影响表现出物种特异性,灌木类群中较大的冠层面积结合较高的植株形态更有利于树干茎流形成[48-49].Park等[50]的研究进一步证实,冠层投影面积较大的乔木与小型个体树干茎流产量存在显著差异(P<0.05).同时,基径和枝条数量对树干茎流产量也具有显著影响.例如,沙坡头地区柠条锦鸡儿的树干茎流产量与株高、基径、枝条数、分枝长均呈显著线性正相关(P<0.05)[51-52];黄土高原典型灌木沙棘(Hippophae rhamnoides)的冠层投影面积、分枝长度、枝条数量和冠层体积均与树干茎流产量呈正相关[37].沙拐枣(Calligonum mongolicum)较大的冠层面积和较长的灌木分枝长度增加树干茎流产量[49].此外,树干茎流率与基部面积呈现正相关关系(图4),但相关性不显著性(P=0.663). ...
... 树干茎流产生过程并非受单一因素主导,而是多因素综合作用的结果.现有研究多聚焦于不同灌木种间形态参数差异分析[28,50],或分析单一或少数冠层结构参数对树干茎流产量的影响[37,50-51].Honda等[53]和Whitworth-Hulse等[54]研究表明,具有更多分枝且分枝垂直的植物具有更高的树干茎流产流优势;Martinez-Meza[55]发现,当枝条倾角大于45°时,美洲焦油灌木产生的树干茎流量要大于水平角度的枝条,而腺牧豆树在主干角度为70°~75°时具有最大的树干茎流量;泡泡刺(Nitraria sphaerocarpa)树干茎流量随着枝条倾角增加而增加[56].本研究去掉了对树干茎流产量影响较小的分枝形态参数,进而使用熵权法对灌木分枝雨水汇集能力进行了综合评估,依据综合权重值得出对树干茎流产量具有显著作用的关键分枝形态参数为:基径(0.50)>倾角(0.28)>长度(0.22).这说明基径较粗、分枝较长且分枝倾角接近65°的植株具有更高的树干茎流生产效率(图5),这种冠层形态特征可形成更优的雨水截留界面和引流通路.Yuan等[57]在黄土高原开展的研究也表明较粗基径和较长分枝结构可增加沙柳树干茎流产量.本研究量化分析了灌木分枝构型参数对树干茎流产量的影响作用,有助于深入认识干旱区灌木的水分蓄集和利用策略. ...
How do rainfall intensity and raindrop size determine stemflow production?quantitative evaluation from field rainfall simulation experiments
1
2021
... 现有研究表明,冠层结构参数(株高、基径、基部面积、枝条数、冠层体积及冠层投影面积等)对树干茎流产量具有显著调控作用.本研究结果显示,树干茎流率与株高、基径、枝条数、冠层体积及冠层投影面积之间均呈显著正相关(P<0.05;图4).一般而言,高大植株因其扩展型冠层结构表现出更强的树干茎流生产能力,这种关联性可能与冠层体积与降水截留效率的正反馈机制有关[23,29].值得注意的是,冠层投影面积对树干茎流产量的影响表现出物种特异性,灌木类群中较大的冠层面积结合较高的植株形态更有利于树干茎流形成[48-49].Park等[50]的研究进一步证实,冠层投影面积较大的乔木与小型个体树干茎流产量存在显著差异(P<0.05).同时,基径和枝条数量对树干茎流产量也具有显著影响.例如,沙坡头地区柠条锦鸡儿的树干茎流产量与株高、基径、枝条数、分枝长均呈显著线性正相关(P<0.05)[51-52];黄土高原典型灌木沙棘(Hippophae rhamnoides)的冠层投影面积、分枝长度、枝条数量和冠层体积均与树干茎流产量呈正相关[37].沙拐枣(Calligonum mongolicum)较大的冠层面积和较长的灌木分枝长度增加树干茎流产量[49].此外,树干茎流率与基部面积呈现正相关关系(图4),但相关性不显著性(P=0.663). ...
Factors affecting the stemflow of trees in the Brazilian Cerrado
1
2015
... 树干茎流产生过程并非受单一因素主导,而是多因素综合作用的结果.现有研究多聚焦于不同灌木种间形态参数差异分析[28,50],或分析单一或少数冠层结构参数对树干茎流产量的影响[37,50-51].Honda等[53]和Whitworth-Hulse等[54]研究表明,具有更多分枝且分枝垂直的植物具有更高的树干茎流产流优势;Martinez-Meza[55]发现,当枝条倾角大于45°时,美洲焦油灌木产生的树干茎流量要大于水平角度的枝条,而腺牧豆树在主干角度为70°~75°时具有最大的树干茎流量;泡泡刺(Nitraria sphaerocarpa)树干茎流量随着枝条倾角增加而增加[56].本研究去掉了对树干茎流产量影响较小的分枝形态参数,进而使用熵权法对灌木分枝雨水汇集能力进行了综合评估,依据综合权重值得出对树干茎流产量具有显著作用的关键分枝形态参数为:基径(0.50)>倾角(0.28)>长度(0.22).这说明基径较粗、分枝较长且分枝倾角接近65°的植株具有更高的树干茎流生产效率(图5),这种冠层形态特征可形成更优的雨水截留界面和引流通路.Yuan等[57]在黄土高原开展的研究也表明较粗基径和较长分枝结构可增加沙柳树干茎流产量.本研究量化分析了灌木分枝构型参数对树干茎流产量的影响作用,有助于深入认识干旱区灌木的水分蓄集和利用策略. ...
Advantages of rainfall partitioning by the global invader Ligustrum lucidum over the dominant native Lithraea molleoides in a dry forest
1
2020
... 树干茎流产生过程并非受单一因素主导,而是多因素综合作用的结果.现有研究多聚焦于不同灌木种间形态参数差异分析[28,50],或分析单一或少数冠层结构参数对树干茎流产量的影响[37,50-51].Honda等[53]和Whitworth-Hulse等[54]研究表明,具有更多分枝且分枝垂直的植物具有更高的树干茎流产流优势;Martinez-Meza[55]发现,当枝条倾角大于45°时,美洲焦油灌木产生的树干茎流量要大于水平角度的枝条,而腺牧豆树在主干角度为70°~75°时具有最大的树干茎流量;泡泡刺(Nitraria sphaerocarpa)树干茎流量随着枝条倾角增加而增加[56].本研究去掉了对树干茎流产量影响较小的分枝形态参数,进而使用熵权法对灌木分枝雨水汇集能力进行了综合评估,依据综合权重值得出对树干茎流产量具有显著作用的关键分枝形态参数为:基径(0.50)>倾角(0.28)>长度(0.22).这说明基径较粗、分枝较长且分枝倾角接近65°的植株具有更高的树干茎流生产效率(图5),这种冠层形态特征可形成更优的雨水截留界面和引流通路.Yuan等[57]在黄土高原开展的研究也表明较粗基径和较长分枝结构可增加沙柳树干茎流产量.本研究量化分析了灌木分枝构型参数对树干茎流产量的影响作用,有助于深入认识干旱区灌木的水分蓄集和利用策略. ...
Stemflow,Throughfall,and Root Water Channelization by Three Arid Land Shrubs in Southern New Mexico
1
1994
... 树干茎流产生过程并非受单一因素主导,而是多因素综合作用的结果.现有研究多聚焦于不同灌木种间形态参数差异分析[28,50],或分析单一或少数冠层结构参数对树干茎流产量的影响[37,50-51].Honda等[53]和Whitworth-Hulse等[54]研究表明,具有更多分枝且分枝垂直的植物具有更高的树干茎流产流优势;Martinez-Meza[55]发现,当枝条倾角大于45°时,美洲焦油灌木产生的树干茎流量要大于水平角度的枝条,而腺牧豆树在主干角度为70°~75°时具有最大的树干茎流量;泡泡刺(Nitraria sphaerocarpa)树干茎流量随着枝条倾角增加而增加[56].本研究去掉了对树干茎流产量影响较小的分枝形态参数,进而使用熵权法对灌木分枝雨水汇集能力进行了综合评估,依据综合权重值得出对树干茎流产量具有显著作用的关键分枝形态参数为:基径(0.50)>倾角(0.28)>长度(0.22).这说明基径较粗、分枝较长且分枝倾角接近65°的植株具有更高的树干茎流生产效率(图5),这种冠层形态特征可形成更优的雨水截留界面和引流通路.Yuan等[57]在黄土高原开展的研究也表明较粗基径和较长分枝结构可增加沙柳树干茎流产量.本研究量化分析了灌木分枝构型参数对树干茎流产量的影响作用,有助于深入认识干旱区灌木的水分蓄集和利用策略. ...
西北干旱区泡泡刺灌丛的降雨再分配特征及影响因素分析
1
2022
... 树干茎流产生过程并非受单一因素主导,而是多因素综合作用的结果.现有研究多聚焦于不同灌木种间形态参数差异分析[28,50],或分析单一或少数冠层结构参数对树干茎流产量的影响[37,50-51].Honda等[53]和Whitworth-Hulse等[54]研究表明,具有更多分枝且分枝垂直的植物具有更高的树干茎流产流优势;Martinez-Meza[55]发现,当枝条倾角大于45°时,美洲焦油灌木产生的树干茎流量要大于水平角度的枝条,而腺牧豆树在主干角度为70°~75°时具有最大的树干茎流量;泡泡刺(Nitraria sphaerocarpa)树干茎流量随着枝条倾角增加而增加[56].本研究去掉了对树干茎流产量影响较小的分枝形态参数,进而使用熵权法对灌木分枝雨水汇集能力进行了综合评估,依据综合权重值得出对树干茎流产量具有显著作用的关键分枝形态参数为:基径(0.50)>倾角(0.28)>长度(0.22).这说明基径较粗、分枝较长且分枝倾角接近65°的植株具有更高的树干茎流生产效率(图5),这种冠层形态特征可形成更优的雨水截留界面和引流通路.Yuan等[57]在黄土高原开展的研究也表明较粗基径和较长分枝结构可增加沙柳树干茎流产量.本研究量化分析了灌木分枝构型参数对树干茎流产量的影响作用,有助于深入认识干旱区灌木的水分蓄集和利用策略. ...
Comparisons of stemflow and its bio-/abiotic influential factors between two xerophytic shrub species
1
2017
... 树干茎流产生过程并非受单一因素主导,而是多因素综合作用的结果.现有研究多聚焦于不同灌木种间形态参数差异分析[28,50],或分析单一或少数冠层结构参数对树干茎流产量的影响[37,50-51].Honda等[53]和Whitworth-Hulse等[54]研究表明,具有更多分枝且分枝垂直的植物具有更高的树干茎流产流优势;Martinez-Meza[55]发现,当枝条倾角大于45°时,美洲焦油灌木产生的树干茎流量要大于水平角度的枝条,而腺牧豆树在主干角度为70°~75°时具有最大的树干茎流量;泡泡刺(Nitraria sphaerocarpa)树干茎流量随着枝条倾角增加而增加[56].本研究去掉了对树干茎流产量影响较小的分枝形态参数,进而使用熵权法对灌木分枝雨水汇集能力进行了综合评估,依据综合权重值得出对树干茎流产量具有显著作用的关键分枝形态参数为:基径(0.50)>倾角(0.28)>长度(0.22).这说明基径较粗、分枝较长且分枝倾角接近65°的植株具有更高的树干茎流生产效率(图5),这种冠层形态特征可形成更优的雨水截留界面和引流通路.Yuan等[57]在黄土高原开展的研究也表明较粗基径和较长分枝结构可增加沙柳树干茎流产量.本研究量化分析了灌木分枝构型参数对树干茎流产量的影响作用,有助于深入认识干旱区灌木的水分蓄集和利用策略. ...
冠层降雨再分配驱动生态系统养分富集:机制、数量与模式
2
2024
... 树干茎流作用不仅与水文过程密切相关,在生态系统养分循环中也扮演着重要角色,是生态系统养分循环的重要驱动因子之一[35,58].大气降水与干沉降过程在冠层交换作用下溶解的气溶胶颗粒物、植物冠层的干沉降物质,存在于植物冠层中间的生产者、消费者和分解者的排泄物与残体,在大气降水的溶解和冲刷作用下,随树干茎流汇集至植物基部,形成灌木沃岛效应[58].本文主要聚焦于柠条锦鸡儿树干茎流的水文过程效应,未来研究将系统探讨养分离子的动态变化特征及其富集规律,以期全面揭示树干茎流在干旱生态系统水分和养分循环中的作用. ...
... [58].本文主要聚焦于柠条锦鸡儿树干茎流的水文过程效应,未来研究将系统探讨养分离子的动态变化特征及其富集规律,以期全面揭示树干茎流在干旱生态系统水分和养分循环中的作用. ...





 甘公网安备 62010202000688号
甘公网安备 62010202000688号
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}