


0 引言
草本植物作为干旱区植被的重要组成部分,具有较强的指示性和响应性,常被用作环境变化的早期敏感指标[2,7],在维持生态系统初级生产力、养分循环和土壤稳定性等方面发挥着基础性作用[8]。草本植物多样性对地下水埋深具有显著响应[9]。多数情况下,随着地下水埋深增加,土壤水分供应减少,植物面临更严重的水分胁迫,导致物种丰富度和多样性下降[10]。然而,越来越多的研究指出,这一关系并非总是线性,草本植物多样性在一定地下水埋深范围内可能呈现“中间最优”模式,即在特定埋深下达到多样性峰值[11]。这种非线性响应表明地下水调控植物多样性的机制更加复杂,可能受植物生活型、功能性状、水分利用效率以及土壤理化特征等多种因子交互作用的共同影响[9,11]。
土壤作为植物赖以生存的基础介质,其物理和化学性质对植被的结构、功能及分布具有深刻影响[2]。在干旱区,GD作为关键的环境驱动因子,直接调控土壤剖面的水分含量,影响植物可利用水资源,显著改变土壤中的关键养分动态[12]。GD变化能显著影响土壤有机碳、速效氮(尤其是硝态氮)、有效磷和有效钾的浓度。其影响模式可能随埋深范围和具体养分元素而异,并非简单的线性关系[12]。例如,某些养分可能在适中埋深时达到峰值,或在极深埋深条件下因生物地球化学过程(如分解、矿化、淋溶、固定或释放速率)的改变而呈现累积或耗竭[12]。这种由GD驱动的水分和养分有效性的变化,会间接影响植物定居、生长和种间竞争格局[13-14]。此外,地下水埋深波动或长期处于较深水平可能改变土壤盐分平衡,导致盐渍化风险增加,但深层地下水有时也可能因淋溶减弱而在表层土壤中导致某些离子(包括部分营养盐)相对富集[12],进一步筛选能够适应特定盐分和养分条件的物种[15]。
本文选择甘肃敦煌西湖荒漠-湿地生态系统为研究对象,该区域处于绿洲与荒漠交错地带,是典型的地下水维系型生态系统。近年来,区域地下水位波动显著,加之人类活动持续扰动,使得草本植被群落呈现出明显的空间异质性和动态变化特征。然而,尚缺乏系统性实证研究揭示地下水埋深变化如何影响草本植物多样性,及其与土壤理化性质之间的联动机制。因此,本研究旨在探讨以下科学问题:①地下水埋深如何影响敦煌西湖地区荒漠-湿地草本植物的物种多样性与群落结构?②是否存在最适地下水埋深范围以维持较高的植物多样性?③土壤理化特征如何调节地下水对植物多样性的影响?
1 研究区概况
疏勒河地处河西走廊最西端,是甘肃省三大内陆河之一,以冰川融水与山区降水补给为主,经瓜州县流向敦煌市北疏勒河古道;党河由南注入,经敦煌绿洲向西北汇入古疏勒河道,因受上游水库影响,党河和疏勒河主要以定期排放生态补给水沿古疏勒河道流过保护区汇聚于库木塔格沙漠东前沿[16-17]。研究区位于疏勒河流域下游的甘肃敦煌西湖国家级自然保护区(39°45′—40°36′N,92°45′—93°50′E),地处河西走廊最西端,平均海拔960 m,属敦煌盆地的一部分(图1)。该区域为阿尔金山山前洪积倾斜平原,地势南北高、中部低,自东南向西北倾斜[18]。气候类型为典型干旱荒漠气候,多年平均降水量仅39.9 mm,且90%集中于6—9月;多年平均蒸发量高达1 553 mm,干旱显著[19]。
图1
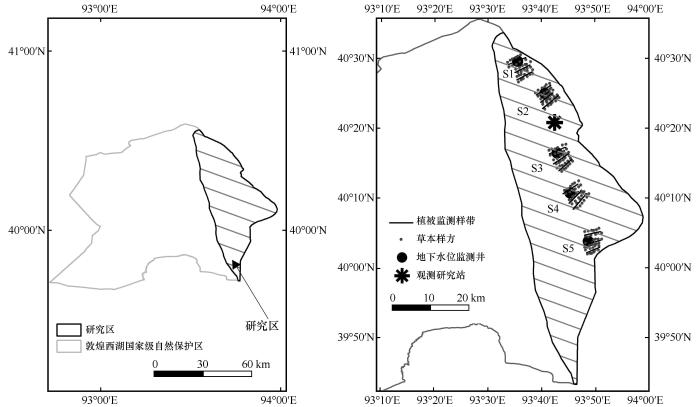
图1
取样点示意图
Fig.1
Diagram of sampling points
2 研究方法
2.1 地下水埋深测定
在甘肃敦煌西湖国家级自然保护区设置5个实验样地(S1~S5),南北横跨约50 km。S1和S2位于马圈湾(沼泽土为主),S3和S4位于后坑(草甸土为主),S5位于火烧湖(棕漠土为主),所有样地土壤表层均无下伏砂石覆盖。每个样地布设地下水埋深监测井[21],监测深度根据区域地下水动态特征调整,确保数据反映植被根系可利用的潜水位范围。其中,S3监测井依托兰州大学野外台站观测平台建立,其余监测井由保护区管护中心建设。监测井数据由敦煌西湖国家级自然保护区提供,采用自动水位记录仪(HOBO U20L,精度±0.1 cm)每小时连续监测地下水埋深(GD)。研究选取2017—2022年生长季(4—10月)数据,以月均值为基础计算各监测点GD多年平均值,用于后续分析(图2)。
图2
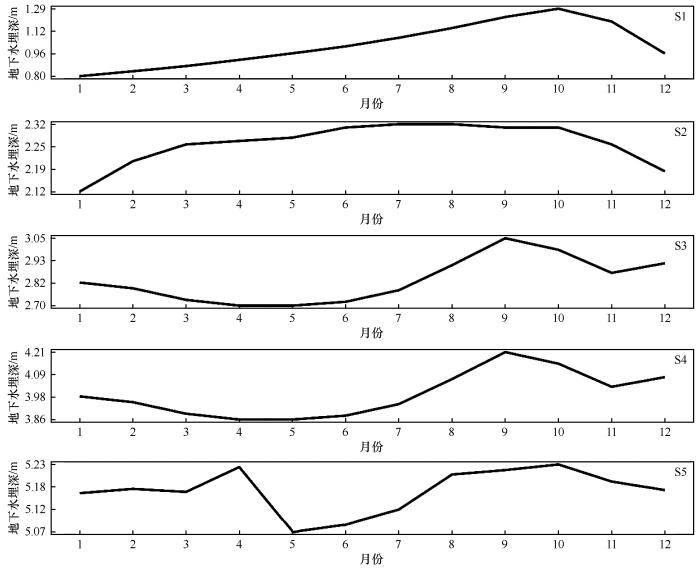
图2
生长季(4—10月)地下水埋深的变化
Fig.2
Changes in depth to groundwater during the growing season (April-October)
2.2 草本植物群落调查
在每个监测样带上设置24个1 m×1 m的样方,用于调查草本的群落参数,5个地点共计180个样方。样方调查于植被生长旺季(7—8月)进行,记录内容包括物种组成(鉴定样方内所有草本植物种名,统计个体数,分株独立计数)、群落盖度(采用目视估测法,按百分比估算样方内植被垂直投影覆盖地表面积)、植株高度(使用精度0.01 m的钢卷尺逐株测量自然伸展高度,取平均值)以及多度。
2.3 土壤样品采集与测定
为评估甘肃敦煌西湖国家级自然保护区草本植物群落土壤养分特征,研究采用五点取样法对每个草本样方(1 m×1 m)进行土壤样品采集。在样方内按对角线及中心点选取5个采样点,清除表层凋落物后,使用土钻采集0~20 cm深度土壤,剔除根系与砾石后混合均匀,形成3份独立混合土样。样品采集后置于阴凉通风处自然风干,研磨过2 mm尼龙筛,保存于密封袋中备用。
土壤总碳(STC)采用重铬酸钾外加热氧化法测定;土壤全氮(STN)通过凯氏定氮法消解后,利用自动定氮仪测定;土壤全磷(STP)采用钼锑抗比色法测定(HClO₄-H₂SO₄消解);总溶解盐(TDS)通过水土比5∶1浸提,电导率法测定浸提液离子总量;土壤含水量(SMC)采用烘干法(105 ℃恒重至质量恒定)计算质量含水率;土壤pH值则通过水土比2.5∶1悬液静置30 min后,采用pH计(精度±0.01)测定[23]。
2.4 数据处理与分析
2.4.1 重要值及多样性指数计算
基于样方调查数据,采用经典多样性指数表征植物群落结构特征。首先,计算每个物种的重要值[24]。
式中:相对频度、相对高度和相对盖度分别为某物种在样方中的频度、高度和盖度与所有物种总频度、总高度和总盖度的比值。
然后计算生物多样性指标[25]:
Margalef丰富度指数(SP) :
Shannon-Wiener多样性指数(H):
Simpson多样性指数(D):
Pielou均匀度指数(J) :
式中:S为样方物种数;N为总个体数;Pi 为第i个物种占所有物种的比例。
2.4.2 数据分析
通过线性回归分析GD与各土壤理化性质(SMC、STC、STN、STP等)关系,评估GD对土壤环境的直接影响。对于物种多样性指数的组间差异比较,采用Kruskal-Wallis单因素方差分析,结合Wilcoxon符号秩检验进行多重比较(显著性阈值α=0.05),以揭示不同GD梯度下群落结构的显著变化。为进一步解析SMC与物种多样性之间的权衡关系,采用分位数回归(90%分位)识别权衡临界点[26],通过R语言quantreg包实现分段回归模型。冗余分析(RDA)用于解析环境因子对物种分布的解释力,结合Hellinger变换的物种数据与标准化环境变量,通过前向选择筛选显著变量(VIF<10),并利用Monte Carlo置换检验(9 999次)验证模型显著性。采用层次分割方法(Hierarchical Partitioning)量化每个环境因子对物种分布的贡献,并进行显著性检验。采用均方根偏差法(RMSD)表征SMC与物种多样性指数的权衡强度[27],RMSD值趋近0时表示协同关系,值增大则反映权衡加剧。所有分析在R4.3.1平台完成,采用vegan包处理群落数据,rdacca.hp和permu.hp包用于层次分割以及显著性检验,采用ggplot2包进行结果可视化。
3 结果与分析
3.1 不同地下水埋深下草本群落组成特征和多样性变化
本研究共记录草本植物12种,隶属7科12属(表1)。其中,禾本科为优势科,包含冰草、芦苇、小獐毛(Aeluropus pungens)和针茅(Stipa capillata)4个物种,占总物种数的33.33%。豆科和莎草科次之,均包含2个物种:豆科有胀果甘草和骆驼刺;莎草科有薹草(Carex parva)和矮藨草(Scirpus pumilus),物种数占比均为16.67%。其余科各记录1个物种,包括海韭菜(Triglochin maritima)、海乳草(Lysimachia maritima)、里海旋覆花(Inula caspica)和罗布麻(Apocynum venetum),分属水麦冬科、报春花科、菊科和夹竹桃科,各占总种数的8.33%。群落主要由多年生植物组成,反映其对干旱环境下水分胁迫的长期适应。
表1 草本群落物种组成及重要值
Table 1
| 物种 | 拉丁名 | 科 | 属 | 重要值 | |||||
|---|---|---|---|---|---|---|---|---|---|
| S1 | S2 | S3 | S4 | S5 | 平均 | ||||
| 矮藨草 | Scirpus pumilus | 莎草科 | 藨草属 | 0.01 | 0.03 | — | — | — | 0.02 |
| 冰草 | Agropyron cristatum | 禾本科 | 冰草属 | 0.24 | 0.16 | 0.11 | — | — | 0.17 |
| 胀果甘草 | Glycyrrhiza inflata | 豆科 | 甘草属 | 0.02 | 0.15 | 0.34 | 0.06 | 0.39 | 0.19 |
| 海韭菜 | Triglochin maritima | 水麦冬科 | 水麦冬属 | 0.06 | 0.05 | — | — | — | 0.06 |
| 海乳草 | Lysimachia maritima | 报春花科 | 珍珠菜属 | — | 0.01 | — | — | — | 0.01 |
| 里海旋覆花 | Inula caspica | 菊科 | 旋覆花属 | — | 0.02 | 0.01 | — | — | 0.02 |
| 芦苇 | Phragmites australis | 禾本科 | 芦苇属 | 0.40 | 0.37 | 0.46 | 0.52 | 0.46 | 0.44 |
| 罗布麻 | Apocynum venetum | 夹竹桃科 | 罗布麻属 | 0.10 | 0.14 | 0.02 | 0.25 | 0.07 | 0.12 |
| 骆驼刺 | Alhagi camelorum | 豆科 | 骆驼刺属 | — | 0.06 | 0.05 | 0.14 | 0.08 | 0.09 |
| 薹草 | Carex parva | 莎草科 | 薹草属 | 0.07 | — | — | — | — | 0.07 |
| 小獐毛 | Aeluropus pungens | 禾本科 | 獐毛属 | 0.10 | — | — | — | — | 0.10 |
| 针茅 | Stipa capillata | 禾本科 | 针茅属 | — | 0.01 | — | — | — | 0.01 |
重要值分析表明,芦苇在5个样地中均占据主导地位,平均重要值达0.44(0.37~0.52),显示其对GD梯度的较强适应性。在浅埋深区(S1~S2,GD为1.07~2.03 m),罗布麻(平均0.12,0.10~0.14)和冰草(平均0.17,0.16~0.24)表现显著,表明其依赖较高SMC。随着地下水埋深增至中等水平(S3~S4,GD为2.78~4.04 m),骆驼刺(平均0.11,0.05~0.17)、罗布麻(平均0.14,0.02~0.25)和胀果甘草(平均0.20,0.06~0.34)重要性上升,反映其较强的耐旱能力。在深埋深区(S5,GD=5.42 m),只有芦苇(0.45)以及胀果甘草(0.39)占据明显优势,群落结构因极端干旱而显著简化。禾本科与豆科植物凭借深根系和耐旱特性,在群落结构与功能中起关键作用。浅埋深区以中生草本为主,深埋深区仅存极端耐旱物种,凸显地下水埋深梯度对物种选择及群落动态的深刻影响。
如图3所示,从S1(GD=1.07 m)到S5(GD=5.42 m),Margalef丰富度指数(SP)呈现连续递减趋势。浅埋深区(S1~S2)物种丰富度较高,以中生草本(如芦苇、冰草)为主;至S3(GD=2.78 m)后,SP值降幅加剧(下降64%),浅根系物种(如海韭菜、矮藨草)逐渐消失;深埋深区(S5)SP值降至最低,仅存极端耐旱种(如骆驼刺)。Shannon-Wiener多样性指数(H)从S1至S5逐级下降,其中S1~S2(GD为1.07~2.03 m)降幅平缓,S3~S4(GD为2.78~4.04 m)显著降低,深埋深区物种组成趋于单一化,仅耐旱种主导。Simpson优势度指数(D)从S1到S5逐渐降低,但趋势呈现两阶段变化:浅埋深区(S1~S2)优势种集中度较低,多物种共存;中及深埋深区(S3~S5)耐旱深根种(如骆驼刺)逐渐占据优势。Pielou均匀度指数(J)较为稳定,且各组之间无差异。物种多样性指数在不同GD梯度下呈现显著分异规律。浅埋深区(GD<3 m)以SP和H值最高、D值最低、J值较高的特征,反映出高物种丰富度、低优势种集中度的中生草本群落特征。中等埋深区(GD为3~4.5 m)多样性指数显著降低,耐旱深根种(如罗布麻和骆驼刺)逐渐占据生态位优势。深埋深区(GD>4.5 m)SP和H值最低、D值最高,仅剩极端耐旱物种存留,群落功能趋于单一化。
图3
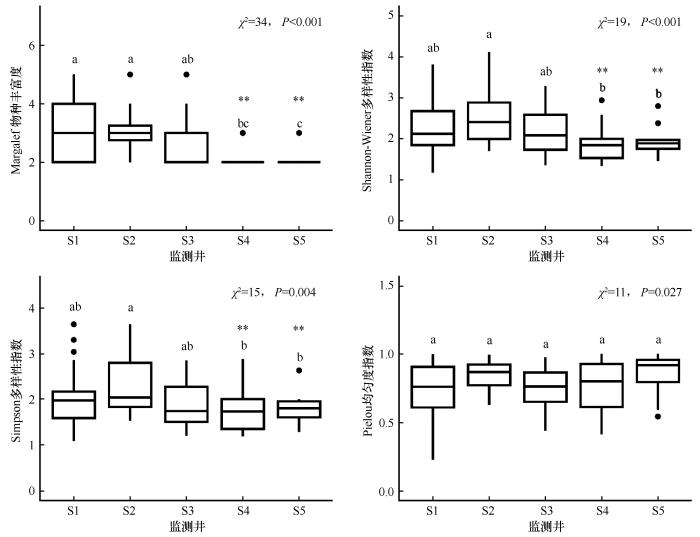
图3
地下水埋深对物种多样性的影响
Fig.3
The influence of groundwater depth on species diversity
3.2 不同地下水埋深下土壤理化性质的变化
本研究基于GD<7 m范围内7个土壤指标的15个观测点数据,分析了其分布特征与变化趋势,并探讨了GD对土壤属性的调控机制。结果显示(图4),STC、STN、SMC和pH随埋深增加呈显著下降趋势(P<0.001),表明GD增大导致水分可利用性降低,抑制了有机质积累与微生物活性。STC/STN和TDS含量则随埋深增加而上升,前者可能与深层有机质分解速率减缓有关,后者或源于蒸发作用下的盐分表聚效应。值得注意的是,STP与埋深无显著关联,暗示其分布受磷矿化或外源输入等非水文因素主导。数据离散性(如STC/STN和TDS)反映了埋深之外其他环境因子的潜在作用。
图4
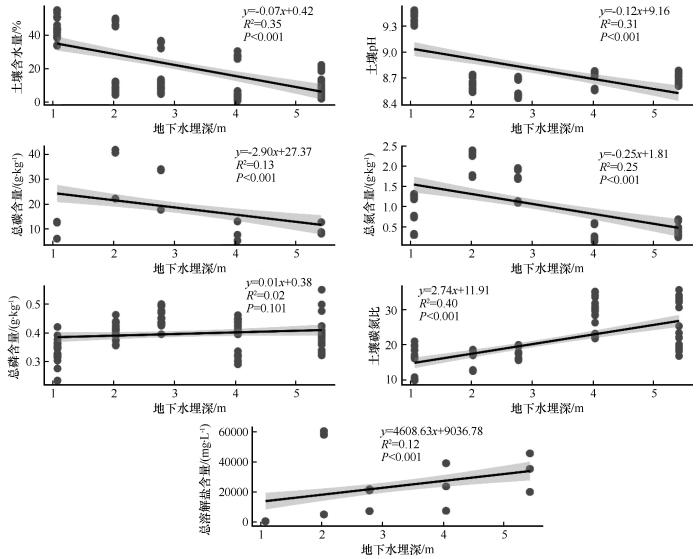
图4
土壤理化性质沿地下水埋深的变化
Fig.4
Changes in soil physical and chemical properties along the depth to groundwater
回归模型进一步量化了GD对土壤理化性质的影响强度。STC和STN随GD的增加逐渐减少,表明水分限制显著削弱了浅层土壤的养分富集能力。SMC与pH下降则直接印证了干旱胁迫加剧与碱性离子淋溶的协同效应。STC/STN随埋深显著上升,表明深层土壤中碳滞留时间延长,而氮矿化受限。TDS虽呈微弱上升趋势,但其高变异性暗示盐分动态可能受局部地下水矿化度或灌溉活动的强烈干扰。STP的独立分布特征进一步支持其受埋深影响有限,主要受控于母质风化或施肥输入。
3.3 环境因子对草本群落物种分布的影响
通过冗余分析(RDA)和层次分割方法分析环境因子对物种分布的影响进行量化分析。RDA结果显示(图5),第一轴(RDA1)解释了45.37%的受限变异,第二轴(RDA2)解释了25.27%,两轴合计解释70.64%的变异。GD箭头最长,夹角最小,表明其与RDA1的相关性最高(R²=0.4287,P=0.001),是群落变化的主要驱动因子。TDS(R²=0.3073,P=0.001)、pH(R²=0.4716,P=0.001)和SMC(R²=0.4916,P=0.001)箭头较长,显示影响显著。STN(R²=0.0300,P=0.179)箭头中等长度。STP(R²=0.0863,P=0.004)和STC(R²=0.0462,P=0.058)箭头较短,影响不显著。S1和S2靠近SMC和pH箭头,表明这两个样地的群落受高SMC和高pH的强正向影响。S5靠近GD和TDS箭头,显示其受高埋深和高盐分驱动。S3和S4位于中间,受到环境因子综合的影响。GD箭头指向正RDA1,胀果甘草沿此方向分布(R²=0.5370,P=0.001),表明其与深层GD正相关。SMC和pH箭头指向负RDA1,矮藨草(R²=0.4117,P=0.001)、里海旋覆花(R²=0.5656,P=0.001)和海乳草(R²=0.4295,P=0.001)分布在此区域,说明高水分和高pH环境有利于这些物种生长。STP箭头影响较弱,芦苇(R²=0.0112,P=0.436)和小獐毛(R²=0.0019,P=0.860)无显著相关性。RDA分析阐明了GD通过调节SMC、pH和TDS,显著驱动草本植物群落格局,其中SMC(49.16%)和pH(47.16%)作用最强。物种分布反映了生态位分化,浅层高水分环境支持矮藨草、海旋覆花和海乳草等湿生植物,深层高盐环境促进胀果甘草的分布。
图5
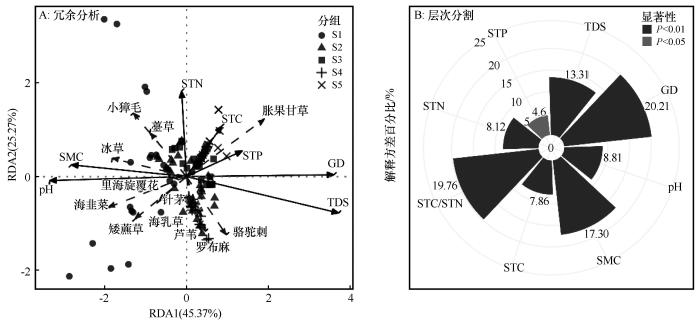
图5
地下水埋深及土壤理化性质对草本群落物种分布的影响
Fig.5
Effects of groundwater depth and soil physicochemical properties on the distribution of herbaceous community species
层次分割结果表明(图5),环境因子对物种分布的总解释力为37.80%。GD是物种分布最主要的驱动因子,其总贡献比例(20.21%)居首位,STC/STN(19.76%)和SMC(17.30%)也有较高的贡献度。TDS、pH、STN、STC和STP的总贡献占比分别为13.31%、8.81%、8.12%、7.86%和4.60%。所有土壤理化因子的置换检验都达到了显著水平,其中STC、STN、SMC、TDS、STC/STN)、pH和GD达到极显著水平(P<0.01)。STP达到显著水平(P=0.041)。
3.4 土壤水分与物种多样性相对收益沿地下水埋深的变化
图6
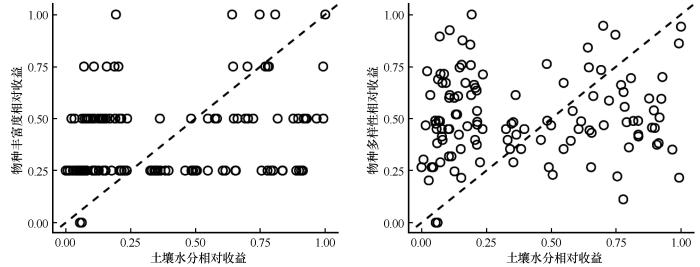
图6
草本群落物种多样性与土壤水分的权衡关系
Fig.6
Trade-offs between species diversity and soil moisture in herbaceous communities
图7
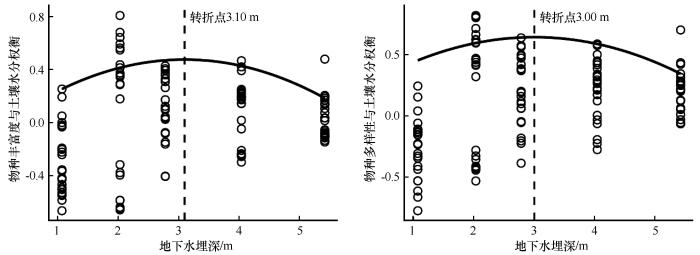
图7
草本群落物种多样性与土壤水分权衡沿地下水埋深的变化
Fig.7
Species diversity and soil water trade-offs in herbaceous communities along groundwater depths
通过分位数回归模型进一步分析发现,物种丰富度、物种多样性和SMC的权衡随GD增加呈现先增后减的抛物线趋势,分别在GD=3.10 m和GD=3.00 m附近达到转折点(图7)。在转折点之前,权衡值逐渐上升,表明SMC对物种多样性的支持作用增强;而在转折点之后,随着GD继续增加(>3.0 m),权衡值明显下降,SMC相对收益减少速度快于物种多样性下降速度。这一现象表明,在GD较深条件下,SMC难以维持现有物种多样性水平,物种多样性维持需要以消耗更多SMC为代价。综合来看,GD在约3.0 m时是物种多样性与SMC权衡的响应阈值。GD持续增加导致SMC显著降低,成为限制荒漠草本群落物种多样性、引发植被退化的主要驱动因素。
4 讨论
4.1 地下水埋深显著影响草本植物群落组成与物种多样性
地下水埋深是调控干旱区植被格局变化的关键水文因子[4,10,14-15]。本研究中,草本植物群落以禾本科和豆科为主,呈现出典型干旱半干旱生态系统物种构成特征。这与干旱半干旱地区研究结果相似[10],这些植物多具备深根性、耐旱性和较高的水分利用效率,可适应不同地下水位的水分胁迫。在本研究区域,芦苇在各个样地中均占优势,显示出对不同地下水位条件的高度适应性。与此同时,在地下水埋深较小的区域(<3 m),如S1与S2样地,胀果甘草和冰草等物种频繁出现,表明其对表层土壤水分依赖较强。随着地下水位下降,群落中逐渐被耐旱性较强的深根植物(如骆驼刺、罗布麻)所替代,反映出植物物种对水分可利用性的生态位分异与选择机制[28-29]。在最大地下水埋深区(>5 m),仅有极少数耐旱物种(如胀果甘草与骆驼刺)能够存活,验证了极端水分胁迫对物种构成的限制作用,这一结果与相似区域的研究结果一致[7,10,28-29]。
本研究进一步发现,物种丰富度与Shannon-Wiener多样性指数随着地下水埋深的增加而显著下降,与已有研究结论相符,即地下水位下降会削弱土壤水分供给,从而降低浅根植物生存能力和群落多样性[[10,25]。此外,Simpson优势度指数在地下水加深过程中逐渐上升,表明深根、耐旱优势种的主导性增强;Pielou均匀度指数未表现出显著差异,提示尽管物种数量减少,但剩余物种的分布趋于均衡。与Zhao等[27]和韩路等[10]提出的浅埋深促进中生植物多样性,而深埋深限制其生长的结论相一致,本研究在不同地下水埋深梯度下,揭示了由中生向旱生植物的群落更替轨迹,并进一步明确了地下水位变化对群落组成的选择性影响。
4.2 地下水埋深通过调节土壤水分与养分特性影响群落构建
然而,地下水埋深对土壤养分的影响并非简单的线性关系,而是呈现出复杂性。研究表明,对于SOC,在浅层模拟地下水条件下(0.4~2.2 m),SOC含量在适中埋深(如1.2 m)可能最高,因为此时水分条件可能最适宜有机质分解与积累的平衡;但在田间深层地下水条件下(2.5~11.0 m),SOC含量随埋深增加而增加,可能因为更深的地下水位导致土壤更干燥,从而抑制了有机碳的分解损失[12]。对于土壤氮素,在浅层地下水条件下,硝态氮含量在适中埋深(0.8~1.2 m)最高,这可能与最佳的水分和好氧条件有利于硝化作用有关;而在深层地下水条件下,硝态氮含量随埋深增加而升高,可能是由于干燥条件抑制了反硝化作用,导致氮素更多地以硝态氮形式累积[12]。本研究结果显示STC/STN随埋深增加而上升,反映了水分限制下氮矿化速率低于碳分解速率,这与深埋深区硝态氮累积但总氮可能降低或有机碳分解更慢的复杂情况相符。对于土壤总磷,本研究表明其与地下水埋深没有显著相关性,推测可能受母质等非水文因素控制[8]。相似地区的研究也表明速效磷(AP)与埋深无显著关系,支持短期内地下水对磷溶解和释放影响不大的观点[12]。然而,在田间深层地下水条件下研究表明速效磷含量在最深处(11.0 m)显著升高,这表明在长期和更极端干旱条件下,磷动态可能发生改变,例如磷淋溶损失减少或难溶性磷缓慢释放累积[12]。这为本研究中STP无显著相关的发现提供了对比,即影响可能取决于埋深范围和时间尺度。
4.3 地下水与其他环境因子共同驱动物种空间分布格局
层次分割分析结果显示,GD对群落组成的解释度最高,其次是STC/STN与SMC,表明地下水深度不仅直接影响群落构建,还通过调控土壤理化性质在更复杂的生态网络中发挥作用。与Zhang等[11]提出干旱生态系统中多因子交互共同塑造植物分布格局的结论相一致。
4.4 土壤水分-物种多样性的权衡关系与生态阈值
分位数回归分析揭示了SMC与物种丰富度和多样性之间存在显著的权衡关系,且这种关系受到GD的调控。在浅埋深区域(GD<3.0 m),SMC与物种多样性呈正相关关系,即水分能够有效促进草本植物的丰富度与均匀度;这一结果与韩路等[10]、Zeng等[9]在塔里木荒漠的研究结果相符。GD超过3.0 m后,正向协同关系出现转折,变为典型的水分-多样性权衡关系:此时维持一定多样性所需的边际水分成本上升,即同等水分输入所带来的物种增益逐渐降低。该现象提示存在一个水分生态阈值(约为3.0 m地下水位),超过该阈值,水分成为限制物种多样性的主导因子。这也与Eamus等[4]和Zhang等[12]关于地下水下降引发生态退化风险的研究结果相似,为区域水资源管理与植被保护提供了理论支撑。
5 结论
本研究系统阐明了GD及关键土壤理化因子对干旱区荒漠-湿地生态系统草本群落结构和多样性的影响调控,揭示了水分-生物多样性协同关系的生态阈值,为区域生态修复提供了重要科学依据。主要研究结论如下:地下水埋深通过水分-养分耦合路径驱动生态过程。GD增加引发SMC下降与盐分富集,通过改变碳氮比化学计量特征重塑土壤养分格局,导致有机质分解受限与氮素有效性降低,形成水文胁迫-养分双重作用机制。植物群落演替呈现显著的水分生态位分化规律。随着GD加深(1.07~5.42 m),中生植物(芦苇等)向旱生物种(骆驼刺等)的替代过程符合环境筛理论,群落结构简化。3.0 m为水分-多样性协同关系的临界阈值。当GD<3.0 m时,SMC与物种多样性呈协同增益;超过该阈值后转为权衡关系。这一发现为确定生态水位红线提供了量化依据。建议将3.0 m作为区域生态水位管控基准,通过水文调控维持土壤水分-养分-盐分平衡,促进中生-旱生草本植物群落协同发展。未来需加强多因子协同作用及植被-土壤反馈机制研究。
参考文献
Global desertification: building a science for dryland development
[J].
Impact of deeper groundwater depth on vegetation and soil in semi-arid region of Eastern China
[J].
Mapping groundwater-dependent ecosystems in arid Central Asia: implications for controlling regional land degradation
[J].
A functional methodology for determining the groundwater regime needed to maintain the health of groundwater-dependent vegetation
[J].
Phreatophytic vegetation and groundwater fluctuations: a review of current research and application of ecosystem response modeling with an emphasis on Great Basin vegetation
[J].
Effects of groundwater depth on vegetation coverage in the Ulan Buh Desert in a recent 20-year period
[J].
Groundwater depth and soil properties are associated with variation in vegetation of a desert riparian ecosystem in an arid area of China
[J].
Effect of groundwater depth on riparian plant diversity along riverside-desert gradients in the Tarim River
[J].
Abiotic stress and human activities reduce plant diversity in desert riparian forests
[J].
Groundwater depth alters soil nutrient concentrations in different environments in an arid desert
[J].
Impact of groundwater depth and soil salinity on riparian plant diversity and distribution in an arid area of China
[J].
Driving factors of community‐level plant functional traits and species distributions in the desert-wetland ecosystem of the Shule River Basin,China
[J].
Desert riparian vegetation and groundwater in the lower reaches of the Tarim River basin
[J].
A review on the driving effect of groundwater depth on the evolution of sandy plant soil systems in arid and semi-arid region
[J].
Effects of different vegetation restoration on soil nutrients,enzyme activities,and microbial communities in degraded karst landscapes in Southwest China
[J].
Effects of groundwater depth on ecological stoichiometric characteristics of assimilated branches and soil of two desert plants
[J].
Vegetation response to groundwater variation in arid environments: visualization of research evolution,synthesis of response types,and estimation of groundwater threshold
[J].

 甘公网安备 62010202000688号
甘公网安备 62010202000688号
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}