我国沙区人工植被系统生态-水文过程和互馈机理研究评述
5
2013
... 在干旱荒漠人工植被区,水分是一切生态系统格局和过程的驱动力[1],植被建设与有限的水资源互为矛盾,亦存在互依互调关系.植物的水分利用策略决定着生态系统格局和动态,而生态系统演变又不断改变土壤-植被的水文过程[2-3].阐明植被生态系统水分利用特征及其稳定性维持机理,是实现干旱沙区生态恢复和植被系统可持续发展的先决条件.在干旱生态系统中,植物的降水利用对策复杂多样,许多研究者提出了各种模型描述和解释.最具有代表性的为土壤水分资源双层分布利用模型基于土壤水分在垂直方向上的异质性,将土壤分为两层,认为不同植物功能类型通过利用不同深度土壤水分资源实现共存[4].通过稳定同位素技术分析证实不同生活型植物对水分利用策略存在显著差异.乔木优先利用地下水,灌木呈现多元水源利用特征,而草本则根据根系深浅形成资源获取的垂直生态位互补[5],这种互补性不仅促进物种共存,还能通过浅根系植物固持表层水分、中深根系植物维持深层水资源协同作用,提升群落整体水分利用效率[6].干旱区诸多生态建设也验证了这一假说[7].之后又有学者提出脉动-存储理论[8],即在干旱区生态系统中,降水事件作为脉动触发一系列生态过程,而植物和微生物等生物体则通过储备来应对脉动带来的资源变化.脉动-存储理论的核心为干旱区生态系统受到间歇性降水事件的影响,降水事件为植物和微生物提供短暂的资源脉冲,植物和微生物利用脉冲资源进行生长和繁殖,并在资源丰富时积累储备,以维持干旱期间生命活动.这种储备包括碳、氮等营养物质,还包括水分和其他关键资源[9].Reynolds等[10]对脉动-存储理论进行了改进,指出干旱半干旱环境降水并不直接影响生态系统生产力状况,而是通过影响土壤水分间接控制其响应过程,植物响应程度与物种丰富度、前期土壤水分条件和季节因素关系密切.而针对干旱区植被斑块状的分布特征,Ludwig等[11]提出了诱发-迁移-储备-脉动(TTRP)模型.该模型基于干旱半干旱生态系统资源有限特点,强调降水事件作为触发器,以及植被斑块在资源存储和利用中的核心作用.资源(主要是水分和养分)通过地表径流等方式从植被稀疏区域迁移到植被斑块区域.植被斑块作为储备中心将资源存储起来,在后续的生长和繁殖中以脉动的形式利用这些资源.如果考虑不同植物功能性状及其对降水利用的时滞性,Ogle等[12]通过整合植物功能类型、脉动、阈值和时滞等提出阈值-时滞模型,深入探讨了植物在干旱环境中水分利用和生长策略.只有当降水量超过这一阈值时,植物才会启动生长和繁殖活动.植物对脉动事件大小和持续时间具有严格的响应阈值,如浅根植物与深根植物所依赖的水源有所差异,大降水事件通常能导致降水入渗较深,而小降水事件降水入渗较浅,导致不同植物生长响应的降水阈值不同[13].Schwinning等[14]提出等级响应模型,由于降水特征在植物生长过程、种群动态和生态系统功能等方面的重要作用.这些模型在不同程度上揭示干旱生态系统中植物对降水的利用机制,为深入理解植被生态系统在干旱环境中的适应性及稳定性提供了理论基础.同时,也为生态恢复和水资源管理提供重要的指导,有助于制定合理的生态恢复策略和水资源分配方案. ...
... 树干径流是干旱半干旱区植被在降水再分配过程中不可忽视的组成部分[54-55].其在形成过程中能够改变降水的空间分布格局,影响水分和养分循环的速度和方向,亦对植物生存和生长起着至关重要作用[56].目前,国内外学者对干旱荒漠地区灌木的自然集水特征及其对植物根部土壤水分和营养元素补给进行研究[57-58],通过实验观测和模型方法确定不同灌木树干径流产生条件、影响因素、树干径流量与雨水特征关系等[55].这些研究表明荒漠灌木能够通过树干径流收集5%~10%(有时可达到40%)的降水并直接运送到植物根部,之后通过根道系统和土壤大孔隙下渗,将水分贮存在较深的土层中供植物在干旱缺水时利用[59],同时缓解了根际区降水入渗的浅层化问题[60-61].据估算通过灌木树干径流的聚集作用,根部土壤收集到的水分是当地降雨量的11~221倍[62],土壤湿润锋增加1.2~4.5倍,土壤水分含量提高10%~140%[63].不同植被覆盖类型下热带、温带以及干旱半干旱区生态系统中平均树干径流量占同期降雨量的3.5%、11.3%和19.0%[64].这一过程可能是荒漠耐旱灌木适应干旱瘠薄立地条件重要生态水文机理[1].在沙坡头人工固沙植被区,柠条和油蒿的树干径流占降雨量的8.99%和2.82%[65],但也有研究认为这有可能过高估计树干径流对灌丛根际区的补给作用[66].主要原因有:一是沙坡头地区以小降雨事件为主,加之较高的潜在蒸散量,不利于树干径流形成;二是由于植被区灌木和半灌木茎干直径较小,且柠条枝条一般为多枝结构,油蒿叶表面覆盖一层不透水蜡质,这种特殊植物冠层结构不利于树干径流形成[18].虽然这一比例较小,但在干旱地区,即使是少量的水分补充对植物的生长也具有重要意义. ...
... 叶片吸水是指植物不通过根系,而直接通过叶片来吸取水分的能力,同时也是植物利用隐匿降水(如雾或凝结水)的重要方式[67],特别是在小降水事件占主导的干旱荒漠人工植被区[1].关于植物叶片吸水的研究可以追溯到20世纪50年代,Stone[68]在北美西部地区首次发现黄松(Pinus ponderosa)植物通过叶片摄取水分,但是由于局限于特定环境的特定对象,此现象并未得到足够重视[69].直到近些年来越来越多的研究发现叶片吸水是植物最大限度地利用有限的水资源的重要途径[70].迄今为止,国内外发现有70余种植物通过叶片吸收水分[71].主要分布在北方阔叶针叶林[72-73]、热带雨林[74]、稀疏草原[75]、温带森林[76]和温带草原[77]等.国际上对云雾林的研究也证实了这一现象,雾水可直接通过叶片角质层扩散至根茎,占叶片总含水量的42%[71-72].对生活在地中海地区的常绿灌木迷迭香(Rosmarinus officinalis)和西班牙薰衣草(Lavandula stoechas)的研究结果表明:干旱时期有一半时间出现凝结水吸收使叶片相对含水量提高了72%,水势提高了0.5 MPa,水分亏缺状况得到缓解[78].国内关于植物叶片吸水的研究主要在西双版纳雨林生态系统的附生植物与非附生植物叶片[79],及干旱荒漠地区如准噶尔盆地[80]、民勤[81]、毛乌素沙地[82-83]和腾格里沙漠人工植被区[26]等植物均具有叶片吸水现象,且荒漠植物地上(叶片)和地下(根系)部分耗水及吸水策略存在权衡[80]. ...
... 在荒漠人工植被生态系统中,水分是限制植物生长的主要环境因子,不同植物类群对水分适应的多样性是维持生态系统平衡和可持续性的关键因素[1].对于固沙灌木和半灌木,不同生物学特性决定其维持生存的生态水文学机理不同.如柠条叶与嫩茎上丰富的软毛,为叶片吸收凝结水提供了有利条件;而油蒿根系主要分布在浅层土壤(0~20 cm),对树干径流的利用比例可能高于柠条[55,65].同时,植物水分利用对策也可能随着降水梯度的变化而改变,如东非萨瓦纳生境中特别干旱时段,水力提升现象和树干径流基本消失,合欢(Acacia tortilis)主要通过叶片吸水来适应干旱胁迫[87];当降水量为1.5~5.8 mm,许多植物如三齿团香木(Larrea tridentate)、美洲焦油菊(Flourensia cernua)和腺牧豆树(Prosopis glandulosa)产生树干径流,通过根系优先流来补充土壤水分[88];干季末期大降雨事件(50 mm)之后,萨瓦纳8种主要植物的根系传导水分状况表明:水分在水势梯度驱动下,先从浅层土壤进入浅层根,之后进入主根,最后再由主根进入深层较干燥土壤中,完成水力再分配[89].这说明植物水分利用方式或者对策会随着降雨量的变化而变化. ...
... 固沙灌木对降水的利用受多种因素制约,其中降雨量是关键[1,85].根据沙坡头站实验观测数据,建立不同降水梯度下固沙植物对降水利用的生态水文调节阈值模型(图1),确定固沙植物根系水力再分配、树干径流和叶片吸水这3种水分适应对策状态转移的拐点.当降水量为0~1 mm,不存在树干径流,植物适应干旱的主要对策为叶片吸水;而当降水量增大至P3~P4时,如在1~5 mm时,叶片吸水作用可以被忽略,树干径流起主要作用;当降水量大于5 mm时,降水可通过穿透雨、入渗等对土壤水进行大量补充,根土界面的水力再分配是植物适应干旱的主要对策.降水量逐渐变化过程中,根土界面水力再分配、树干径流和叶片吸水对植物生存都有影响,因此在模型中定义了混合状态,即代表这3种对策同时存在,只是在某一降水区间,某一种对策可能对植物生存起主要作用.这说明在小降雨事件中,植物主要通过叶片吸水和树干径流等方式快速利用有限水分资源,以满足短期的生理需求;而在较大降雨事件中,根系水力再分配则发挥主导作用,将多余的水分储存于土壤中,供植物在干旱时期使用. ...
流沙固定过程中土壤-植被系统演变
2
2003
... 在干旱荒漠人工植被区,水分是一切生态系统格局和过程的驱动力[1],植被建设与有限的水资源互为矛盾,亦存在互依互调关系.植物的水分利用策略决定着生态系统格局和动态,而生态系统演变又不断改变土壤-植被的水文过程[2-3].阐明植被生态系统水分利用特征及其稳定性维持机理,是实现干旱沙区生态恢复和植被系统可持续发展的先决条件.在干旱生态系统中,植物的降水利用对策复杂多样,许多研究者提出了各种模型描述和解释.最具有代表性的为土壤水分资源双层分布利用模型基于土壤水分在垂直方向上的异质性,将土壤分为两层,认为不同植物功能类型通过利用不同深度土壤水分资源实现共存[4].通过稳定同位素技术分析证实不同生活型植物对水分利用策略存在显著差异.乔木优先利用地下水,灌木呈现多元水源利用特征,而草本则根据根系深浅形成资源获取的垂直生态位互补[5],这种互补性不仅促进物种共存,还能通过浅根系植物固持表层水分、中深根系植物维持深层水资源协同作用,提升群落整体水分利用效率[6].干旱区诸多生态建设也验证了这一假说[7].之后又有学者提出脉动-存储理论[8],即在干旱区生态系统中,降水事件作为脉动触发一系列生态过程,而植物和微生物等生物体则通过储备来应对脉动带来的资源变化.脉动-存储理论的核心为干旱区生态系统受到间歇性降水事件的影响,降水事件为植物和微生物提供短暂的资源脉冲,植物和微生物利用脉冲资源进行生长和繁殖,并在资源丰富时积累储备,以维持干旱期间生命活动.这种储备包括碳、氮等营养物质,还包括水分和其他关键资源[9].Reynolds等[10]对脉动-存储理论进行了改进,指出干旱半干旱环境降水并不直接影响生态系统生产力状况,而是通过影响土壤水分间接控制其响应过程,植物响应程度与物种丰富度、前期土壤水分条件和季节因素关系密切.而针对干旱区植被斑块状的分布特征,Ludwig等[11]提出了诱发-迁移-储备-脉动(TTRP)模型.该模型基于干旱半干旱生态系统资源有限特点,强调降水事件作为触发器,以及植被斑块在资源存储和利用中的核心作用.资源(主要是水分和养分)通过地表径流等方式从植被稀疏区域迁移到植被斑块区域.植被斑块作为储备中心将资源存储起来,在后续的生长和繁殖中以脉动的形式利用这些资源.如果考虑不同植物功能性状及其对降水利用的时滞性,Ogle等[12]通过整合植物功能类型、脉动、阈值和时滞等提出阈值-时滞模型,深入探讨了植物在干旱环境中水分利用和生长策略.只有当降水量超过这一阈值时,植物才会启动生长和繁殖活动.植物对脉动事件大小和持续时间具有严格的响应阈值,如浅根植物与深根植物所依赖的水源有所差异,大降水事件通常能导致降水入渗较深,而小降水事件降水入渗较浅,导致不同植物生长响应的降水阈值不同[13].Schwinning等[14]提出等级响应模型,由于降水特征在植物生长过程、种群动态和生态系统功能等方面的重要作用.这些模型在不同程度上揭示干旱生态系统中植物对降水的利用机制,为深入理解植被生态系统在干旱环境中的适应性及稳定性提供了理论基础.同时,也为生态恢复和水资源管理提供重要的指导,有助于制定合理的生态恢复策略和水资源分配方案. ...
... 植被和土壤之间协同演替关系是沙漠生态系统稳定性的基础[2].腾格里沙漠随着固沙植被建立,植被在从人工系统向自然系统的演变中经历了覆盖度从增加到减少的自疏过程、从灌木到半灌木再到草本的变化,土壤从流动风沙土向钙积旱成土发育[130].固沙灌木与土壤之间存在着复杂的互馈关系,这种关系在维持生态系统稳定性方面发挥着关键作用.一方面,固沙植物通过根系生长、凋落物分解等过程影响土壤的物理和化学性质[19,131].例如植物根系在生长过程中分泌有机物质,增加土壤有机碳含量,改善土壤结构和保水能力[132];同时,根系周转和凋落物分解也为土壤提供丰富养分,促进土壤肥力提升.另一方面,土壤水分和养分状况又反作用于植物的生长和发育.适宜的土壤水分条件能够促进植物根系扩展和吸收功能,提高植物水分利用效率和抗旱能力;而土壤养分则直接影响植物生长速度、生物量积累以及繁殖能力[133].不同演替阶段柠条的水分利用特征表明:演替早期,土壤水分含量决定柠条的水分利用来源,演替中后期,其决定因子分别为植物的基径和土壤黏粉粒含量[134]. ...
我国北方风沙危害区生态重建与恢复:腾格里沙漠土壤水分与植被承载力的探讨
2
2014
... 在干旱荒漠人工植被区,水分是一切生态系统格局和过程的驱动力[1],植被建设与有限的水资源互为矛盾,亦存在互依互调关系.植物的水分利用策略决定着生态系统格局和动态,而生态系统演变又不断改变土壤-植被的水文过程[2-3].阐明植被生态系统水分利用特征及其稳定性维持机理,是实现干旱沙区生态恢复和植被系统可持续发展的先决条件.在干旱生态系统中,植物的降水利用对策复杂多样,许多研究者提出了各种模型描述和解释.最具有代表性的为土壤水分资源双层分布利用模型基于土壤水分在垂直方向上的异质性,将土壤分为两层,认为不同植物功能类型通过利用不同深度土壤水分资源实现共存[4].通过稳定同位素技术分析证实不同生活型植物对水分利用策略存在显著差异.乔木优先利用地下水,灌木呈现多元水源利用特征,而草本则根据根系深浅形成资源获取的垂直生态位互补[5],这种互补性不仅促进物种共存,还能通过浅根系植物固持表层水分、中深根系植物维持深层水资源协同作用,提升群落整体水分利用效率[6].干旱区诸多生态建设也验证了这一假说[7].之后又有学者提出脉动-存储理论[8],即在干旱区生态系统中,降水事件作为脉动触发一系列生态过程,而植物和微生物等生物体则通过储备来应对脉动带来的资源变化.脉动-存储理论的核心为干旱区生态系统受到间歇性降水事件的影响,降水事件为植物和微生物提供短暂的资源脉冲,植物和微生物利用脉冲资源进行生长和繁殖,并在资源丰富时积累储备,以维持干旱期间生命活动.这种储备包括碳、氮等营养物质,还包括水分和其他关键资源[9].Reynolds等[10]对脉动-存储理论进行了改进,指出干旱半干旱环境降水并不直接影响生态系统生产力状况,而是通过影响土壤水分间接控制其响应过程,植物响应程度与物种丰富度、前期土壤水分条件和季节因素关系密切.而针对干旱区植被斑块状的分布特征,Ludwig等[11]提出了诱发-迁移-储备-脉动(TTRP)模型.该模型基于干旱半干旱生态系统资源有限特点,强调降水事件作为触发器,以及植被斑块在资源存储和利用中的核心作用.资源(主要是水分和养分)通过地表径流等方式从植被稀疏区域迁移到植被斑块区域.植被斑块作为储备中心将资源存储起来,在后续的生长和繁殖中以脉动的形式利用这些资源.如果考虑不同植物功能性状及其对降水利用的时滞性,Ogle等[12]通过整合植物功能类型、脉动、阈值和时滞等提出阈值-时滞模型,深入探讨了植物在干旱环境中水分利用和生长策略.只有当降水量超过这一阈值时,植物才会启动生长和繁殖活动.植物对脉动事件大小和持续时间具有严格的响应阈值,如浅根植物与深根植物所依赖的水源有所差异,大降水事件通常能导致降水入渗较深,而小降水事件降水入渗较浅,导致不同植物生长响应的降水阈值不同[13].Schwinning等[14]提出等级响应模型,由于降水特征在植物生长过程、种群动态和生态系统功能等方面的重要作用.这些模型在不同程度上揭示干旱生态系统中植物对降水的利用机制,为深入理解植被生态系统在干旱环境中的适应性及稳定性提供了理论基础.同时,也为生态恢复和水资源管理提供重要的指导,有助于制定合理的生态恢复策略和水资源分配方案. ...
... 腾格里沙漠东南缘人工植被区是中国植物固沙领域的先例.自1956年起科研人员在此开展系统固沙植被建设,历经近70年不懈努力,成功探索出有效的治沙模式,为中国乃至全球荒漠化治理提供实践经验.在治理过程中,首先采用草方格固沙技术,通过增加地表粗糙度有效降低风速,成功固定流沙,为植被的生长创造了有利条件.在此基础上,筛选并种植柠条(Caragana korshinskii)、油蒿(Artemisia ordosica)、花棒(Hedysarum scoparium)等耐旱抗风沙植物,逐步构建起稳定的植被群落.经过多年的持续治理,包兰铁路两侧形成了一条长55 km、宽500~1 000 m的绿色长廊,确保包兰铁路畅通无阻,极大地改善了区域生态环境[28].但近年来由于全球气候变化,人工植被生态系统抵御自然风险能力降低,植被稳定性问题引起了国内外学者广泛关注[29].一方面随着流动沙丘固定和生境改变,生物类群剧增,人工植被逐渐向多功能复合系统演变[7].另一方面,由于严酷的生境条件,特别是有限的水分,驱使物种之间竞争加剧,使早期栽植的固沙灌木逐渐退化,植被盖度大幅下降.其主要的原因是土壤水分降低和水分循环特征改变[3].植被演变直接关系到防风固沙效益,反过来又影响整个固沙防护体系的稳定性[30].因此,深入了解腾格里沙漠典型固沙灌木生存的生态水文学机理成为目前植被建设中关键的科学问题.为此,本文分析和综述了腾格里沙漠典型固沙灌木多途径水分利用策略及其稳定性维持机制,为干旱沙区植被恢复与重建后的植被稳定性及其变化趋势评价提供理论依据. ...
Walter's two-layer hypothesis revisited:back to the roots!
1
2013
... 在干旱荒漠人工植被区,水分是一切生态系统格局和过程的驱动力[1],植被建设与有限的水资源互为矛盾,亦存在互依互调关系.植物的水分利用策略决定着生态系统格局和动态,而生态系统演变又不断改变土壤-植被的水文过程[2-3].阐明植被生态系统水分利用特征及其稳定性维持机理,是实现干旱沙区生态恢复和植被系统可持续发展的先决条件.在干旱生态系统中,植物的降水利用对策复杂多样,许多研究者提出了各种模型描述和解释.最具有代表性的为土壤水分资源双层分布利用模型基于土壤水分在垂直方向上的异质性,将土壤分为两层,认为不同植物功能类型通过利用不同深度土壤水分资源实现共存[4].通过稳定同位素技术分析证实不同生活型植物对水分利用策略存在显著差异.乔木优先利用地下水,灌木呈现多元水源利用特征,而草本则根据根系深浅形成资源获取的垂直生态位互补[5],这种互补性不仅促进物种共存,还能通过浅根系植物固持表层水分、中深根系植物维持深层水资源协同作用,提升群落整体水分利用效率[6].干旱区诸多生态建设也验证了这一假说[7].之后又有学者提出脉动-存储理论[8],即在干旱区生态系统中,降水事件作为脉动触发一系列生态过程,而植物和微生物等生物体则通过储备来应对脉动带来的资源变化.脉动-存储理论的核心为干旱区生态系统受到间歇性降水事件的影响,降水事件为植物和微生物提供短暂的资源脉冲,植物和微生物利用脉冲资源进行生长和繁殖,并在资源丰富时积累储备,以维持干旱期间生命活动.这种储备包括碳、氮等营养物质,还包括水分和其他关键资源[9].Reynolds等[10]对脉动-存储理论进行了改进,指出干旱半干旱环境降水并不直接影响生态系统生产力状况,而是通过影响土壤水分间接控制其响应过程,植物响应程度与物种丰富度、前期土壤水分条件和季节因素关系密切.而针对干旱区植被斑块状的分布特征,Ludwig等[11]提出了诱发-迁移-储备-脉动(TTRP)模型.该模型基于干旱半干旱生态系统资源有限特点,强调降水事件作为触发器,以及植被斑块在资源存储和利用中的核心作用.资源(主要是水分和养分)通过地表径流等方式从植被稀疏区域迁移到植被斑块区域.植被斑块作为储备中心将资源存储起来,在后续的生长和繁殖中以脉动的形式利用这些资源.如果考虑不同植物功能性状及其对降水利用的时滞性,Ogle等[12]通过整合植物功能类型、脉动、阈值和时滞等提出阈值-时滞模型,深入探讨了植物在干旱环境中水分利用和生长策略.只有当降水量超过这一阈值时,植物才会启动生长和繁殖活动.植物对脉动事件大小和持续时间具有严格的响应阈值,如浅根植物与深根植物所依赖的水源有所差异,大降水事件通常能导致降水入渗较深,而小降水事件降水入渗较浅,导致不同植物生长响应的降水阈值不同[13].Schwinning等[14]提出等级响应模型,由于降水特征在植物生长过程、种群动态和生态系统功能等方面的重要作用.这些模型在不同程度上揭示干旱生态系统中植物对降水的利用机制,为深入理解植被生态系统在干旱环境中的适应性及稳定性提供了理论基础.同时,也为生态恢复和水资源管理提供重要的指导,有助于制定合理的生态恢复策略和水资源分配方案. ...
干旱区荒漠植物体内潜在水源差异及利用策略分析
1
2019
... 在干旱荒漠人工植被区,水分是一切生态系统格局和过程的驱动力[1],植被建设与有限的水资源互为矛盾,亦存在互依互调关系.植物的水分利用策略决定着生态系统格局和动态,而生态系统演变又不断改变土壤-植被的水文过程[2-3].阐明植被生态系统水分利用特征及其稳定性维持机理,是实现干旱沙区生态恢复和植被系统可持续发展的先决条件.在干旱生态系统中,植物的降水利用对策复杂多样,许多研究者提出了各种模型描述和解释.最具有代表性的为土壤水分资源双层分布利用模型基于土壤水分在垂直方向上的异质性,将土壤分为两层,认为不同植物功能类型通过利用不同深度土壤水分资源实现共存[4].通过稳定同位素技术分析证实不同生活型植物对水分利用策略存在显著差异.乔木优先利用地下水,灌木呈现多元水源利用特征,而草本则根据根系深浅形成资源获取的垂直生态位互补[5],这种互补性不仅促进物种共存,还能通过浅根系植物固持表层水分、中深根系植物维持深层水资源协同作用,提升群落整体水分利用效率[6].干旱区诸多生态建设也验证了这一假说[7].之后又有学者提出脉动-存储理论[8],即在干旱区生态系统中,降水事件作为脉动触发一系列生态过程,而植物和微生物等生物体则通过储备来应对脉动带来的资源变化.脉动-存储理论的核心为干旱区生态系统受到间歇性降水事件的影响,降水事件为植物和微生物提供短暂的资源脉冲,植物和微生物利用脉冲资源进行生长和繁殖,并在资源丰富时积累储备,以维持干旱期间生命活动.这种储备包括碳、氮等营养物质,还包括水分和其他关键资源[9].Reynolds等[10]对脉动-存储理论进行了改进,指出干旱半干旱环境降水并不直接影响生态系统生产力状况,而是通过影响土壤水分间接控制其响应过程,植物响应程度与物种丰富度、前期土壤水分条件和季节因素关系密切.而针对干旱区植被斑块状的分布特征,Ludwig等[11]提出了诱发-迁移-储备-脉动(TTRP)模型.该模型基于干旱半干旱生态系统资源有限特点,强调降水事件作为触发器,以及植被斑块在资源存储和利用中的核心作用.资源(主要是水分和养分)通过地表径流等方式从植被稀疏区域迁移到植被斑块区域.植被斑块作为储备中心将资源存储起来,在后续的生长和繁殖中以脉动的形式利用这些资源.如果考虑不同植物功能性状及其对降水利用的时滞性,Ogle等[12]通过整合植物功能类型、脉动、阈值和时滞等提出阈值-时滞模型,深入探讨了植物在干旱环境中水分利用和生长策略.只有当降水量超过这一阈值时,植物才会启动生长和繁殖活动.植物对脉动事件大小和持续时间具有严格的响应阈值,如浅根植物与深根植物所依赖的水源有所差异,大降水事件通常能导致降水入渗较深,而小降水事件降水入渗较浅,导致不同植物生长响应的降水阈值不同[13].Schwinning等[14]提出等级响应模型,由于降水特征在植物生长过程、种群动态和生态系统功能等方面的重要作用.这些模型在不同程度上揭示干旱生态系统中植物对降水的利用机制,为深入理解植被生态系统在干旱环境中的适应性及稳定性提供了理论基础.同时,也为生态恢复和水资源管理提供重要的指导,有助于制定合理的生态恢复策略和水资源分配方案. ...
内蒙古荒漠草原植物水分利用特征差异及对环境因子的响应
1
2023
... 在干旱荒漠人工植被区,水分是一切生态系统格局和过程的驱动力[1],植被建设与有限的水资源互为矛盾,亦存在互依互调关系.植物的水分利用策略决定着生态系统格局和动态,而生态系统演变又不断改变土壤-植被的水文过程[2-3].阐明植被生态系统水分利用特征及其稳定性维持机理,是实现干旱沙区生态恢复和植被系统可持续发展的先决条件.在干旱生态系统中,植物的降水利用对策复杂多样,许多研究者提出了各种模型描述和解释.最具有代表性的为土壤水分资源双层分布利用模型基于土壤水分在垂直方向上的异质性,将土壤分为两层,认为不同植物功能类型通过利用不同深度土壤水分资源实现共存[4].通过稳定同位素技术分析证实不同生活型植物对水分利用策略存在显著差异.乔木优先利用地下水,灌木呈现多元水源利用特征,而草本则根据根系深浅形成资源获取的垂直生态位互补[5],这种互补性不仅促进物种共存,还能通过浅根系植物固持表层水分、中深根系植物维持深层水资源协同作用,提升群落整体水分利用效率[6].干旱区诸多生态建设也验证了这一假说[7].之后又有学者提出脉动-存储理论[8],即在干旱区生态系统中,降水事件作为脉动触发一系列生态过程,而植物和微生物等生物体则通过储备来应对脉动带来的资源变化.脉动-存储理论的核心为干旱区生态系统受到间歇性降水事件的影响,降水事件为植物和微生物提供短暂的资源脉冲,植物和微生物利用脉冲资源进行生长和繁殖,并在资源丰富时积累储备,以维持干旱期间生命活动.这种储备包括碳、氮等营养物质,还包括水分和其他关键资源[9].Reynolds等[10]对脉动-存储理论进行了改进,指出干旱半干旱环境降水并不直接影响生态系统生产力状况,而是通过影响土壤水分间接控制其响应过程,植物响应程度与物种丰富度、前期土壤水分条件和季节因素关系密切.而针对干旱区植被斑块状的分布特征,Ludwig等[11]提出了诱发-迁移-储备-脉动(TTRP)模型.该模型基于干旱半干旱生态系统资源有限特点,强调降水事件作为触发器,以及植被斑块在资源存储和利用中的核心作用.资源(主要是水分和养分)通过地表径流等方式从植被稀疏区域迁移到植被斑块区域.植被斑块作为储备中心将资源存储起来,在后续的生长和繁殖中以脉动的形式利用这些资源.如果考虑不同植物功能性状及其对降水利用的时滞性,Ogle等[12]通过整合植物功能类型、脉动、阈值和时滞等提出阈值-时滞模型,深入探讨了植物在干旱环境中水分利用和生长策略.只有当降水量超过这一阈值时,植物才会启动生长和繁殖活动.植物对脉动事件大小和持续时间具有严格的响应阈值,如浅根植物与深根植物所依赖的水源有所差异,大降水事件通常能导致降水入渗较深,而小降水事件降水入渗较浅,导致不同植物生长响应的降水阈值不同[13].Schwinning等[14]提出等级响应模型,由于降水特征在植物生长过程、种群动态和生态系统功能等方面的重要作用.这些模型在不同程度上揭示干旱生态系统中植物对降水的利用机制,为深入理解植被生态系统在干旱环境中的适应性及稳定性提供了理论基础.同时,也为生态恢复和水资源管理提供重要的指导,有助于制定合理的生态恢复策略和水资源分配方案. ...
干旱沙区土壤空间异质性变化对植被恢复的影响
2
2005
... 在干旱荒漠人工植被区,水分是一切生态系统格局和过程的驱动力[1],植被建设与有限的水资源互为矛盾,亦存在互依互调关系.植物的水分利用策略决定着生态系统格局和动态,而生态系统演变又不断改变土壤-植被的水文过程[2-3].阐明植被生态系统水分利用特征及其稳定性维持机理,是实现干旱沙区生态恢复和植被系统可持续发展的先决条件.在干旱生态系统中,植物的降水利用对策复杂多样,许多研究者提出了各种模型描述和解释.最具有代表性的为土壤水分资源双层分布利用模型基于土壤水分在垂直方向上的异质性,将土壤分为两层,认为不同植物功能类型通过利用不同深度土壤水分资源实现共存[4].通过稳定同位素技术分析证实不同生活型植物对水分利用策略存在显著差异.乔木优先利用地下水,灌木呈现多元水源利用特征,而草本则根据根系深浅形成资源获取的垂直生态位互补[5],这种互补性不仅促进物种共存,还能通过浅根系植物固持表层水分、中深根系植物维持深层水资源协同作用,提升群落整体水分利用效率[6].干旱区诸多生态建设也验证了这一假说[7].之后又有学者提出脉动-存储理论[8],即在干旱区生态系统中,降水事件作为脉动触发一系列生态过程,而植物和微生物等生物体则通过储备来应对脉动带来的资源变化.脉动-存储理论的核心为干旱区生态系统受到间歇性降水事件的影响,降水事件为植物和微生物提供短暂的资源脉冲,植物和微生物利用脉冲资源进行生长和繁殖,并在资源丰富时积累储备,以维持干旱期间生命活动.这种储备包括碳、氮等营养物质,还包括水分和其他关键资源[9].Reynolds等[10]对脉动-存储理论进行了改进,指出干旱半干旱环境降水并不直接影响生态系统生产力状况,而是通过影响土壤水分间接控制其响应过程,植物响应程度与物种丰富度、前期土壤水分条件和季节因素关系密切.而针对干旱区植被斑块状的分布特征,Ludwig等[11]提出了诱发-迁移-储备-脉动(TTRP)模型.该模型基于干旱半干旱生态系统资源有限特点,强调降水事件作为触发器,以及植被斑块在资源存储和利用中的核心作用.资源(主要是水分和养分)通过地表径流等方式从植被稀疏区域迁移到植被斑块区域.植被斑块作为储备中心将资源存储起来,在后续的生长和繁殖中以脉动的形式利用这些资源.如果考虑不同植物功能性状及其对降水利用的时滞性,Ogle等[12]通过整合植物功能类型、脉动、阈值和时滞等提出阈值-时滞模型,深入探讨了植物在干旱环境中水分利用和生长策略.只有当降水量超过这一阈值时,植物才会启动生长和繁殖活动.植物对脉动事件大小和持续时间具有严格的响应阈值,如浅根植物与深根植物所依赖的水源有所差异,大降水事件通常能导致降水入渗较深,而小降水事件降水入渗较浅,导致不同植物生长响应的降水阈值不同[13].Schwinning等[14]提出等级响应模型,由于降水特征在植物生长过程、种群动态和生态系统功能等方面的重要作用.这些模型在不同程度上揭示干旱生态系统中植物对降水的利用机制,为深入理解植被生态系统在干旱环境中的适应性及稳定性提供了理论基础.同时,也为生态恢复和水资源管理提供重要的指导,有助于制定合理的生态恢复策略和水资源分配方案. ...
... 腾格里沙漠东南缘人工植被区是中国植物固沙领域的先例.自1956年起科研人员在此开展系统固沙植被建设,历经近70年不懈努力,成功探索出有效的治沙模式,为中国乃至全球荒漠化治理提供实践经验.在治理过程中,首先采用草方格固沙技术,通过增加地表粗糙度有效降低风速,成功固定流沙,为植被的生长创造了有利条件.在此基础上,筛选并种植柠条(Caragana korshinskii)、油蒿(Artemisia ordosica)、花棒(Hedysarum scoparium)等耐旱抗风沙植物,逐步构建起稳定的植被群落.经过多年的持续治理,包兰铁路两侧形成了一条长55 km、宽500~1 000 m的绿色长廊,确保包兰铁路畅通无阻,极大地改善了区域生态环境[28].但近年来由于全球气候变化,人工植被生态系统抵御自然风险能力降低,植被稳定性问题引起了国内外学者广泛关注[29].一方面随着流动沙丘固定和生境改变,生物类群剧增,人工植被逐渐向多功能复合系统演变[7].另一方面,由于严酷的生境条件,特别是有限的水分,驱使物种之间竞争加剧,使早期栽植的固沙灌木逐渐退化,植被盖度大幅下降.其主要的原因是土壤水分降低和水分循环特征改变[3].植被演变直接关系到防风固沙效益,反过来又影响整个固沙防护体系的稳定性[30].因此,深入了解腾格里沙漠典型固沙灌木生存的生态水文学机理成为目前植被建设中关键的科学问题.为此,本文分析和综述了腾格里沙漠典型固沙灌木多途径水分利用策略及其稳定性维持机制,为干旱沙区植被恢复与重建后的植被稳定性及其变化趋势评价提供理论依据. ...
Desert ecosystems:environment and producers
1
1973
... 在干旱荒漠人工植被区,水分是一切生态系统格局和过程的驱动力[1],植被建设与有限的水资源互为矛盾,亦存在互依互调关系.植物的水分利用策略决定着生态系统格局和动态,而生态系统演变又不断改变土壤-植被的水文过程[2-3].阐明植被生态系统水分利用特征及其稳定性维持机理,是实现干旱沙区生态恢复和植被系统可持续发展的先决条件.在干旱生态系统中,植物的降水利用对策复杂多样,许多研究者提出了各种模型描述和解释.最具有代表性的为土壤水分资源双层分布利用模型基于土壤水分在垂直方向上的异质性,将土壤分为两层,认为不同植物功能类型通过利用不同深度土壤水分资源实现共存[4].通过稳定同位素技术分析证实不同生活型植物对水分利用策略存在显著差异.乔木优先利用地下水,灌木呈现多元水源利用特征,而草本则根据根系深浅形成资源获取的垂直生态位互补[5],这种互补性不仅促进物种共存,还能通过浅根系植物固持表层水分、中深根系植物维持深层水资源协同作用,提升群落整体水分利用效率[6].干旱区诸多生态建设也验证了这一假说[7].之后又有学者提出脉动-存储理论[8],即在干旱区生态系统中,降水事件作为脉动触发一系列生态过程,而植物和微生物等生物体则通过储备来应对脉动带来的资源变化.脉动-存储理论的核心为干旱区生态系统受到间歇性降水事件的影响,降水事件为植物和微生物提供短暂的资源脉冲,植物和微生物利用脉冲资源进行生长和繁殖,并在资源丰富时积累储备,以维持干旱期间生命活动.这种储备包括碳、氮等营养物质,还包括水分和其他关键资源[9].Reynolds等[10]对脉动-存储理论进行了改进,指出干旱半干旱环境降水并不直接影响生态系统生产力状况,而是通过影响土壤水分间接控制其响应过程,植物响应程度与物种丰富度、前期土壤水分条件和季节因素关系密切.而针对干旱区植被斑块状的分布特征,Ludwig等[11]提出了诱发-迁移-储备-脉动(TTRP)模型.该模型基于干旱半干旱生态系统资源有限特点,强调降水事件作为触发器,以及植被斑块在资源存储和利用中的核心作用.资源(主要是水分和养分)通过地表径流等方式从植被稀疏区域迁移到植被斑块区域.植被斑块作为储备中心将资源存储起来,在后续的生长和繁殖中以脉动的形式利用这些资源.如果考虑不同植物功能性状及其对降水利用的时滞性,Ogle等[12]通过整合植物功能类型、脉动、阈值和时滞等提出阈值-时滞模型,深入探讨了植物在干旱环境中水分利用和生长策略.只有当降水量超过这一阈值时,植物才会启动生长和繁殖活动.植物对脉动事件大小和持续时间具有严格的响应阈值,如浅根植物与深根植物所依赖的水源有所差异,大降水事件通常能导致降水入渗较深,而小降水事件降水入渗较浅,导致不同植物生长响应的降水阈值不同[13].Schwinning等[14]提出等级响应模型,由于降水特征在植物生长过程、种群动态和生态系统功能等方面的重要作用.这些模型在不同程度上揭示干旱生态系统中植物对降水的利用机制,为深入理解植被生态系统在干旱环境中的适应性及稳定性提供了理论基础.同时,也为生态恢复和水资源管理提供重要的指导,有助于制定合理的生态恢复策略和水资源分配方案. ...
干旱、半干旱环境降水脉动对生态系统的影响
1
2011
... 在干旱荒漠人工植被区,水分是一切生态系统格局和过程的驱动力[1],植被建设与有限的水资源互为矛盾,亦存在互依互调关系.植物的水分利用策略决定着生态系统格局和动态,而生态系统演变又不断改变土壤-植被的水文过程[2-3].阐明植被生态系统水分利用特征及其稳定性维持机理,是实现干旱沙区生态恢复和植被系统可持续发展的先决条件.在干旱生态系统中,植物的降水利用对策复杂多样,许多研究者提出了各种模型描述和解释.最具有代表性的为土壤水分资源双层分布利用模型基于土壤水分在垂直方向上的异质性,将土壤分为两层,认为不同植物功能类型通过利用不同深度土壤水分资源实现共存[4].通过稳定同位素技术分析证实不同生活型植物对水分利用策略存在显著差异.乔木优先利用地下水,灌木呈现多元水源利用特征,而草本则根据根系深浅形成资源获取的垂直生态位互补[5],这种互补性不仅促进物种共存,还能通过浅根系植物固持表层水分、中深根系植物维持深层水资源协同作用,提升群落整体水分利用效率[6].干旱区诸多生态建设也验证了这一假说[7].之后又有学者提出脉动-存储理论[8],即在干旱区生态系统中,降水事件作为脉动触发一系列生态过程,而植物和微生物等生物体则通过储备来应对脉动带来的资源变化.脉动-存储理论的核心为干旱区生态系统受到间歇性降水事件的影响,降水事件为植物和微生物提供短暂的资源脉冲,植物和微生物利用脉冲资源进行生长和繁殖,并在资源丰富时积累储备,以维持干旱期间生命活动.这种储备包括碳、氮等营养物质,还包括水分和其他关键资源[9].Reynolds等[10]对脉动-存储理论进行了改进,指出干旱半干旱环境降水并不直接影响生态系统生产力状况,而是通过影响土壤水分间接控制其响应过程,植物响应程度与物种丰富度、前期土壤水分条件和季节因素关系密切.而针对干旱区植被斑块状的分布特征,Ludwig等[11]提出了诱发-迁移-储备-脉动(TTRP)模型.该模型基于干旱半干旱生态系统资源有限特点,强调降水事件作为触发器,以及植被斑块在资源存储和利用中的核心作用.资源(主要是水分和养分)通过地表径流等方式从植被稀疏区域迁移到植被斑块区域.植被斑块作为储备中心将资源存储起来,在后续的生长和繁殖中以脉动的形式利用这些资源.如果考虑不同植物功能性状及其对降水利用的时滞性,Ogle等[12]通过整合植物功能类型、脉动、阈值和时滞等提出阈值-时滞模型,深入探讨了植物在干旱环境中水分利用和生长策略.只有当降水量超过这一阈值时,植物才会启动生长和繁殖活动.植物对脉动事件大小和持续时间具有严格的响应阈值,如浅根植物与深根植物所依赖的水源有所差异,大降水事件通常能导致降水入渗较深,而小降水事件降水入渗较浅,导致不同植物生长响应的降水阈值不同[13].Schwinning等[14]提出等级响应模型,由于降水特征在植物生长过程、种群动态和生态系统功能等方面的重要作用.这些模型在不同程度上揭示干旱生态系统中植物对降水的利用机制,为深入理解植被生态系统在干旱环境中的适应性及稳定性提供了理论基础.同时,也为生态恢复和水资源管理提供重要的指导,有助于制定合理的生态恢复策略和水资源分配方案. ...
Modifying the 'pulse-reserve' paradigm for deserts of North America:precipitation pulses,soil water,and plant responses
1
2004
... 在干旱荒漠人工植被区,水分是一切生态系统格局和过程的驱动力[1],植被建设与有限的水资源互为矛盾,亦存在互依互调关系.植物的水分利用策略决定着生态系统格局和动态,而生态系统演变又不断改变土壤-植被的水文过程[2-3].阐明植被生态系统水分利用特征及其稳定性维持机理,是实现干旱沙区生态恢复和植被系统可持续发展的先决条件.在干旱生态系统中,植物的降水利用对策复杂多样,许多研究者提出了各种模型描述和解释.最具有代表性的为土壤水分资源双层分布利用模型基于土壤水分在垂直方向上的异质性,将土壤分为两层,认为不同植物功能类型通过利用不同深度土壤水分资源实现共存[4].通过稳定同位素技术分析证实不同生活型植物对水分利用策略存在显著差异.乔木优先利用地下水,灌木呈现多元水源利用特征,而草本则根据根系深浅形成资源获取的垂直生态位互补[5],这种互补性不仅促进物种共存,还能通过浅根系植物固持表层水分、中深根系植物维持深层水资源协同作用,提升群落整体水分利用效率[6].干旱区诸多生态建设也验证了这一假说[7].之后又有学者提出脉动-存储理论[8],即在干旱区生态系统中,降水事件作为脉动触发一系列生态过程,而植物和微生物等生物体则通过储备来应对脉动带来的资源变化.脉动-存储理论的核心为干旱区生态系统受到间歇性降水事件的影响,降水事件为植物和微生物提供短暂的资源脉冲,植物和微生物利用脉冲资源进行生长和繁殖,并在资源丰富时积累储备,以维持干旱期间生命活动.这种储备包括碳、氮等营养物质,还包括水分和其他关键资源[9].Reynolds等[10]对脉动-存储理论进行了改进,指出干旱半干旱环境降水并不直接影响生态系统生产力状况,而是通过影响土壤水分间接控制其响应过程,植物响应程度与物种丰富度、前期土壤水分条件和季节因素关系密切.而针对干旱区植被斑块状的分布特征,Ludwig等[11]提出了诱发-迁移-储备-脉动(TTRP)模型.该模型基于干旱半干旱生态系统资源有限特点,强调降水事件作为触发器,以及植被斑块在资源存储和利用中的核心作用.资源(主要是水分和养分)通过地表径流等方式从植被稀疏区域迁移到植被斑块区域.植被斑块作为储备中心将资源存储起来,在后续的生长和繁殖中以脉动的形式利用这些资源.如果考虑不同植物功能性状及其对降水利用的时滞性,Ogle等[12]通过整合植物功能类型、脉动、阈值和时滞等提出阈值-时滞模型,深入探讨了植物在干旱环境中水分利用和生长策略.只有当降水量超过这一阈值时,植物才会启动生长和繁殖活动.植物对脉动事件大小和持续时间具有严格的响应阈值,如浅根植物与深根植物所依赖的水源有所差异,大降水事件通常能导致降水入渗较深,而小降水事件降水入渗较浅,导致不同植物生长响应的降水阈值不同[13].Schwinning等[14]提出等级响应模型,由于降水特征在植物生长过程、种群动态和生态系统功能等方面的重要作用.这些模型在不同程度上揭示干旱生态系统中植物对降水的利用机制,为深入理解植被生态系统在干旱环境中的适应性及稳定性提供了理论基础.同时,也为生态恢复和水资源管理提供重要的指导,有助于制定合理的生态恢复策略和水资源分配方案. ...
A scaling rule for landscape patches and how it applies to conserving soil resources in Savannas
1
2000
... 在干旱荒漠人工植被区,水分是一切生态系统格局和过程的驱动力[1],植被建设与有限的水资源互为矛盾,亦存在互依互调关系.植物的水分利用策略决定着生态系统格局和动态,而生态系统演变又不断改变土壤-植被的水文过程[2-3].阐明植被生态系统水分利用特征及其稳定性维持机理,是实现干旱沙区生态恢复和植被系统可持续发展的先决条件.在干旱生态系统中,植物的降水利用对策复杂多样,许多研究者提出了各种模型描述和解释.最具有代表性的为土壤水分资源双层分布利用模型基于土壤水分在垂直方向上的异质性,将土壤分为两层,认为不同植物功能类型通过利用不同深度土壤水分资源实现共存[4].通过稳定同位素技术分析证实不同生活型植物对水分利用策略存在显著差异.乔木优先利用地下水,灌木呈现多元水源利用特征,而草本则根据根系深浅形成资源获取的垂直生态位互补[5],这种互补性不仅促进物种共存,还能通过浅根系植物固持表层水分、中深根系植物维持深层水资源协同作用,提升群落整体水分利用效率[6].干旱区诸多生态建设也验证了这一假说[7].之后又有学者提出脉动-存储理论[8],即在干旱区生态系统中,降水事件作为脉动触发一系列生态过程,而植物和微生物等生物体则通过储备来应对脉动带来的资源变化.脉动-存储理论的核心为干旱区生态系统受到间歇性降水事件的影响,降水事件为植物和微生物提供短暂的资源脉冲,植物和微生物利用脉冲资源进行生长和繁殖,并在资源丰富时积累储备,以维持干旱期间生命活动.这种储备包括碳、氮等营养物质,还包括水分和其他关键资源[9].Reynolds等[10]对脉动-存储理论进行了改进,指出干旱半干旱环境降水并不直接影响生态系统生产力状况,而是通过影响土壤水分间接控制其响应过程,植物响应程度与物种丰富度、前期土壤水分条件和季节因素关系密切.而针对干旱区植被斑块状的分布特征,Ludwig等[11]提出了诱发-迁移-储备-脉动(TTRP)模型.该模型基于干旱半干旱生态系统资源有限特点,强调降水事件作为触发器,以及植被斑块在资源存储和利用中的核心作用.资源(主要是水分和养分)通过地表径流等方式从植被稀疏区域迁移到植被斑块区域.植被斑块作为储备中心将资源存储起来,在后续的生长和繁殖中以脉动的形式利用这些资源.如果考虑不同植物功能性状及其对降水利用的时滞性,Ogle等[12]通过整合植物功能类型、脉动、阈值和时滞等提出阈值-时滞模型,深入探讨了植物在干旱环境中水分利用和生长策略.只有当降水量超过这一阈值时,植物才会启动生长和繁殖活动.植物对脉动事件大小和持续时间具有严格的响应阈值,如浅根植物与深根植物所依赖的水源有所差异,大降水事件通常能导致降水入渗较深,而小降水事件降水入渗较浅,导致不同植物生长响应的降水阈值不同[13].Schwinning等[14]提出等级响应模型,由于降水特征在植物生长过程、种群动态和生态系统功能等方面的重要作用.这些模型在不同程度上揭示干旱生态系统中植物对降水的利用机制,为深入理解植被生态系统在干旱环境中的适应性及稳定性提供了理论基础.同时,也为生态恢复和水资源管理提供重要的指导,有助于制定合理的生态恢复策略和水资源分配方案. ...
Plant responses to precipitation in desert ecosystems:integrating functional types,pulses,thresholds,and delays
1
2004
... 在干旱荒漠人工植被区,水分是一切生态系统格局和过程的驱动力[1],植被建设与有限的水资源互为矛盾,亦存在互依互调关系.植物的水分利用策略决定着生态系统格局和动态,而生态系统演变又不断改变土壤-植被的水文过程[2-3].阐明植被生态系统水分利用特征及其稳定性维持机理,是实现干旱沙区生态恢复和植被系统可持续发展的先决条件.在干旱生态系统中,植物的降水利用对策复杂多样,许多研究者提出了各种模型描述和解释.最具有代表性的为土壤水分资源双层分布利用模型基于土壤水分在垂直方向上的异质性,将土壤分为两层,认为不同植物功能类型通过利用不同深度土壤水分资源实现共存[4].通过稳定同位素技术分析证实不同生活型植物对水分利用策略存在显著差异.乔木优先利用地下水,灌木呈现多元水源利用特征,而草本则根据根系深浅形成资源获取的垂直生态位互补[5],这种互补性不仅促进物种共存,还能通过浅根系植物固持表层水分、中深根系植物维持深层水资源协同作用,提升群落整体水分利用效率[6].干旱区诸多生态建设也验证了这一假说[7].之后又有学者提出脉动-存储理论[8],即在干旱区生态系统中,降水事件作为脉动触发一系列生态过程,而植物和微生物等生物体则通过储备来应对脉动带来的资源变化.脉动-存储理论的核心为干旱区生态系统受到间歇性降水事件的影响,降水事件为植物和微生物提供短暂的资源脉冲,植物和微生物利用脉冲资源进行生长和繁殖,并在资源丰富时积累储备,以维持干旱期间生命活动.这种储备包括碳、氮等营养物质,还包括水分和其他关键资源[9].Reynolds等[10]对脉动-存储理论进行了改进,指出干旱半干旱环境降水并不直接影响生态系统生产力状况,而是通过影响土壤水分间接控制其响应过程,植物响应程度与物种丰富度、前期土壤水分条件和季节因素关系密切.而针对干旱区植被斑块状的分布特征,Ludwig等[11]提出了诱发-迁移-储备-脉动(TTRP)模型.该模型基于干旱半干旱生态系统资源有限特点,强调降水事件作为触发器,以及植被斑块在资源存储和利用中的核心作用.资源(主要是水分和养分)通过地表径流等方式从植被稀疏区域迁移到植被斑块区域.植被斑块作为储备中心将资源存储起来,在后续的生长和繁殖中以脉动的形式利用这些资源.如果考虑不同植物功能性状及其对降水利用的时滞性,Ogle等[12]通过整合植物功能类型、脉动、阈值和时滞等提出阈值-时滞模型,深入探讨了植物在干旱环境中水分利用和生长策略.只有当降水量超过这一阈值时,植物才会启动生长和繁殖活动.植物对脉动事件大小和持续时间具有严格的响应阈值,如浅根植物与深根植物所依赖的水源有所差异,大降水事件通常能导致降水入渗较深,而小降水事件降水入渗较浅,导致不同植物生长响应的降水阈值不同[13].Schwinning等[14]提出等级响应模型,由于降水特征在植物生长过程、种群动态和生态系统功能等方面的重要作用.这些模型在不同程度上揭示干旱生态系统中植物对降水的利用机制,为深入理解植被生态系统在干旱环境中的适应性及稳定性提供了理论基础.同时,也为生态恢复和水资源管理提供重要的指导,有助于制定合理的生态恢复策略和水资源分配方案. ...
Phenological events and their environmental triggers in Mojave Desert ecosystems
1
1974
... 在干旱荒漠人工植被区,水分是一切生态系统格局和过程的驱动力[1],植被建设与有限的水资源互为矛盾,亦存在互依互调关系.植物的水分利用策略决定着生态系统格局和动态,而生态系统演变又不断改变土壤-植被的水文过程[2-3].阐明植被生态系统水分利用特征及其稳定性维持机理,是实现干旱沙区生态恢复和植被系统可持续发展的先决条件.在干旱生态系统中,植物的降水利用对策复杂多样,许多研究者提出了各种模型描述和解释.最具有代表性的为土壤水分资源双层分布利用模型基于土壤水分在垂直方向上的异质性,将土壤分为两层,认为不同植物功能类型通过利用不同深度土壤水分资源实现共存[4].通过稳定同位素技术分析证实不同生活型植物对水分利用策略存在显著差异.乔木优先利用地下水,灌木呈现多元水源利用特征,而草本则根据根系深浅形成资源获取的垂直生态位互补[5],这种互补性不仅促进物种共存,还能通过浅根系植物固持表层水分、中深根系植物维持深层水资源协同作用,提升群落整体水分利用效率[6].干旱区诸多生态建设也验证了这一假说[7].之后又有学者提出脉动-存储理论[8],即在干旱区生态系统中,降水事件作为脉动触发一系列生态过程,而植物和微生物等生物体则通过储备来应对脉动带来的资源变化.脉动-存储理论的核心为干旱区生态系统受到间歇性降水事件的影响,降水事件为植物和微生物提供短暂的资源脉冲,植物和微生物利用脉冲资源进行生长和繁殖,并在资源丰富时积累储备,以维持干旱期间生命活动.这种储备包括碳、氮等营养物质,还包括水分和其他关键资源[9].Reynolds等[10]对脉动-存储理论进行了改进,指出干旱半干旱环境降水并不直接影响生态系统生产力状况,而是通过影响土壤水分间接控制其响应过程,植物响应程度与物种丰富度、前期土壤水分条件和季节因素关系密切.而针对干旱区植被斑块状的分布特征,Ludwig等[11]提出了诱发-迁移-储备-脉动(TTRP)模型.该模型基于干旱半干旱生态系统资源有限特点,强调降水事件作为触发器,以及植被斑块在资源存储和利用中的核心作用.资源(主要是水分和养分)通过地表径流等方式从植被稀疏区域迁移到植被斑块区域.植被斑块作为储备中心将资源存储起来,在后续的生长和繁殖中以脉动的形式利用这些资源.如果考虑不同植物功能性状及其对降水利用的时滞性,Ogle等[12]通过整合植物功能类型、脉动、阈值和时滞等提出阈值-时滞模型,深入探讨了植物在干旱环境中水分利用和生长策略.只有当降水量超过这一阈值时,植物才会启动生长和繁殖活动.植物对脉动事件大小和持续时间具有严格的响应阈值,如浅根植物与深根植物所依赖的水源有所差异,大降水事件通常能导致降水入渗较深,而小降水事件降水入渗较浅,导致不同植物生长响应的降水阈值不同[13].Schwinning等[14]提出等级响应模型,由于降水特征在植物生长过程、种群动态和生态系统功能等方面的重要作用.这些模型在不同程度上揭示干旱生态系统中植物对降水的利用机制,为深入理解植被生态系统在干旱环境中的适应性及稳定性提供了理论基础.同时,也为生态恢复和水资源管理提供重要的指导,有助于制定合理的生态恢复策略和水资源分配方案. ...
Thresholds,memory,and seasonality:understanding pulse dynamics in arid/semi-arid ecosystems
1
2004
... 在干旱荒漠人工植被区,水分是一切生态系统格局和过程的驱动力[1],植被建设与有限的水资源互为矛盾,亦存在互依互调关系.植物的水分利用策略决定着生态系统格局和动态,而生态系统演变又不断改变土壤-植被的水文过程[2-3].阐明植被生态系统水分利用特征及其稳定性维持机理,是实现干旱沙区生态恢复和植被系统可持续发展的先决条件.在干旱生态系统中,植物的降水利用对策复杂多样,许多研究者提出了各种模型描述和解释.最具有代表性的为土壤水分资源双层分布利用模型基于土壤水分在垂直方向上的异质性,将土壤分为两层,认为不同植物功能类型通过利用不同深度土壤水分资源实现共存[4].通过稳定同位素技术分析证实不同生活型植物对水分利用策略存在显著差异.乔木优先利用地下水,灌木呈现多元水源利用特征,而草本则根据根系深浅形成资源获取的垂直生态位互补[5],这种互补性不仅促进物种共存,还能通过浅根系植物固持表层水分、中深根系植物维持深层水资源协同作用,提升群落整体水分利用效率[6].干旱区诸多生态建设也验证了这一假说[7].之后又有学者提出脉动-存储理论[8],即在干旱区生态系统中,降水事件作为脉动触发一系列生态过程,而植物和微生物等生物体则通过储备来应对脉动带来的资源变化.脉动-存储理论的核心为干旱区生态系统受到间歇性降水事件的影响,降水事件为植物和微生物提供短暂的资源脉冲,植物和微生物利用脉冲资源进行生长和繁殖,并在资源丰富时积累储备,以维持干旱期间生命活动.这种储备包括碳、氮等营养物质,还包括水分和其他关键资源[9].Reynolds等[10]对脉动-存储理论进行了改进,指出干旱半干旱环境降水并不直接影响生态系统生产力状况,而是通过影响土壤水分间接控制其响应过程,植物响应程度与物种丰富度、前期土壤水分条件和季节因素关系密切.而针对干旱区植被斑块状的分布特征,Ludwig等[11]提出了诱发-迁移-储备-脉动(TTRP)模型.该模型基于干旱半干旱生态系统资源有限特点,强调降水事件作为触发器,以及植被斑块在资源存储和利用中的核心作用.资源(主要是水分和养分)通过地表径流等方式从植被稀疏区域迁移到植被斑块区域.植被斑块作为储备中心将资源存储起来,在后续的生长和繁殖中以脉动的形式利用这些资源.如果考虑不同植物功能性状及其对降水利用的时滞性,Ogle等[12]通过整合植物功能类型、脉动、阈值和时滞等提出阈值-时滞模型,深入探讨了植物在干旱环境中水分利用和生长策略.只有当降水量超过这一阈值时,植物才会启动生长和繁殖活动.植物对脉动事件大小和持续时间具有严格的响应阈值,如浅根植物与深根植物所依赖的水源有所差异,大降水事件通常能导致降水入渗较深,而小降水事件降水入渗较浅,导致不同植物生长响应的降水阈值不同[13].Schwinning等[14]提出等级响应模型,由于降水特征在植物生长过程、种群动态和生态系统功能等方面的重要作用.这些模型在不同程度上揭示干旱生态系统中植物对降水的利用机制,为深入理解植被生态系统在干旱环境中的适应性及稳定性提供了理论基础.同时,也为生态恢复和水资源管理提供重要的指导,有助于制定合理的生态恢复策略和水资源分配方案. ...
Anatomical,morphological and metabolic acclimation in the resurrection plant Reaumuria soongorica during dehydration and rehydration
1
2007
... 干旱区植物生态适应性是其水分利用策略的重要组成部分.在长期的进化过程中,植物形成了多种适应干旱环境的机制,如特定的细胞损伤修复机理[15]、异化的叶片或同化枝[16]、发达的根系[17-19]、特殊的植物体渗透调节能力[20-21]、独特的解剖结构和水分生理代谢方式[22]等.在水文过程中主要体现为大孔隙优先流入根道及土壤动物活动产生的孔隙[23]、根系水力再分配[24]、树干径流[25]和叶片吸水[26]等特殊的干旱区植物生存和水分利用策略.Li等[27]从生态系统角度分析,发现干旱区固沙植被在演替中,土壤表面轻度干扰,如土壤动物(蚂蚁、蜥蜴等)洞穴等促进降水向深层土壤入渗. ...
中国干旱半干旱区荒漠植物叶片(或同化枝)表皮微形态特征
2
2016
... 干旱区植物生态适应性是其水分利用策略的重要组成部分.在长期的进化过程中,植物形成了多种适应干旱环境的机制,如特定的细胞损伤修复机理[15]、异化的叶片或同化枝[16]、发达的根系[17-19]、特殊的植物体渗透调节能力[20-21]、独特的解剖结构和水分生理代谢方式[22]等.在水文过程中主要体现为大孔隙优先流入根道及土壤动物活动产生的孔隙[23]、根系水力再分配[24]、树干径流[25]和叶片吸水[26]等特殊的干旱区植物生存和水分利用策略.Li等[27]从生态系统角度分析,发现干旱区固沙植被在演替中,土壤表面轻度干扰,如土壤动物(蚂蚁、蜥蜴等)洞穴等促进降水向深层土壤入渗. ...
... 植物水分利用策略多样性是荒漠生态系统维持功能稳定核心机制,其通过根系水力再分配、树干径流和叶片吸水3种关键生态水文适应机制的协同作用,塑造了独特的资源分配格局与抗逆能力.根系水力再分配作为深根系植物(如胡杨(Populus euphratica)、柽柳)核心策略,通过夜间液流将深层(60~240 cm)地下水提升至浅层土壤(0~150 cm),缓解自身干旱胁迫,为浅根系草本提供水分支持,促进多物种共存[42].胡杨根系水力提升可使0~150 cm土层含水量提升16%~38%,同时通过根际优先流增强养分循环效率[42-43].树干径流则通过茎干截留降水并定向输导至根区,形成局部沃岛效应,富集有机质(提升15%~30%)和养分(如N、P含量增加20%~45%)[57-59].叶片吸水通过形态(肉质化、角质层增厚)与生理(CAM途径、气孔夜间开放)双重适应,将蒸腾速率降低至日间最大值的20%~38%,同时利用渗透调节物质(如脯氨酸、可溶性糖)维持细胞水势稳定(ΔΨ≥ -1.5 MPa)[16].这3类策略时空耦合形成多层次水分利用网络.在时间维度上水力再分配主导夜间水分再分配,树干径流响应降水事件,而叶片吸水则在晨间高湿度时段激活;空间维度上调控地下水源网络、地表径流再分布与大气水分捕获.这共同构建土壤-植物-大气连续体动态平衡,通过增强水分时空异质性、维持关键种生理功能及促进微生境形成,最终保障荒漠生态系统的结构稳定与功能可持续性. ...
Distribution and seasonal dynamics of roots in a revegetated stand of Artemisia ordosica Kracsh.in the Tengger Desert (North China)
2
2008
... 干旱区植物生态适应性是其水分利用策略的重要组成部分.在长期的进化过程中,植物形成了多种适应干旱环境的机制,如特定的细胞损伤修复机理[15]、异化的叶片或同化枝[16]、发达的根系[17-19]、特殊的植物体渗透调节能力[20-21]、独特的解剖结构和水分生理代谢方式[22]等.在水文过程中主要体现为大孔隙优先流入根道及土壤动物活动产生的孔隙[23]、根系水力再分配[24]、树干径流[25]和叶片吸水[26]等特殊的干旱区植物生存和水分利用策略.Li等[27]从生态系统角度分析,发现干旱区固沙植被在演替中,土壤表面轻度干扰,如土壤动物(蚂蚁、蜥蜴等)洞穴等促进降水向深层土壤入渗. ...
... 荒漠植物生理生态特征对降水响应亦具有尺度依赖性.植物自身生理特点,如植物光合和蒸腾作用对脉冲降水响应,主要表现在短时间尺度上[99].在日尺度上,小降水事件增加植物蒸腾,而大降水事件降低植物蒸腾,主要原因是小降水事件降低植物叶片水势,而后在很短时间内迅速恢复.但在降水量较大时,植物叶片水势增大,太阳辐射、空气温度以及饱和水汽压差均在之后很长一段时间内保持较低水平,因此植物蒸腾量下降.另外树干液流与蒸腾之间存在时间滞后[100].在长时间尺度上,植物对脉冲降水响应主要体现在种群结构、植被盖度或植物优势种的变化等方面.柠条和油蒿的叶面积指数,NBL和蒸腾均与当地年降水量显著线性正相关.间歇性降水,特别是0~5 mm和0~5 d内降水量,对水分胁迫下植被生存起着积极作用.很少水量可以激活干旱沙漠地区诸多生物过程,如种子发芽和繁殖等.降水集中于某一个时间段,则有可能造成土壤水分流失影响植被的可持续性和生存[101].间歇性降水导致土壤水分在局部和短时间内升高,植物能进行充分利用,同时一部分降水在土壤表层快速蒸发后通过入渗进入深层土壤,干旱期间供植物使用[102].通常情况下,这种类型的脉冲降水更有利于油蒿生存,油蒿根系主要分布在土壤表层(0~40 cm),柠条根部主要分布在1 m的土层之下,其根系利用渗透水的机会较少[17-18].另外小降水事件能促进土壤养分吸收.研究表明较小强度降水事件(降水强度2.8 mm·h-1)中,37.1%侵蚀产物、30.3%有机质、48.3%氮和30.0%溶解养分被重新分配至灌丛斑块;而在较大强度降水事件中(降水强度5.7 mm·h-1),31.1%侵蚀产物、9.84%有机质、19.0%氮和10.0%溶解养分被重新分配至灌丛斑块,其中大部分被径流所带走[103]. ...
Distribution,biomass,and dynamics of roots in a revegetated stand of Caragana korshinskii in the Tengger Desert,Northwestern China
2
2009
... 树干径流是干旱半干旱区植被在降水再分配过程中不可忽视的组成部分[54-55].其在形成过程中能够改变降水的空间分布格局,影响水分和养分循环的速度和方向,亦对植物生存和生长起着至关重要作用[56].目前,国内外学者对干旱荒漠地区灌木的自然集水特征及其对植物根部土壤水分和营养元素补给进行研究[57-58],通过实验观测和模型方法确定不同灌木树干径流产生条件、影响因素、树干径流量与雨水特征关系等[55].这些研究表明荒漠灌木能够通过树干径流收集5%~10%(有时可达到40%)的降水并直接运送到植物根部,之后通过根道系统和土壤大孔隙下渗,将水分贮存在较深的土层中供植物在干旱缺水时利用[59],同时缓解了根际区降水入渗的浅层化问题[60-61].据估算通过灌木树干径流的聚集作用,根部土壤收集到的水分是当地降雨量的11~221倍[62],土壤湿润锋增加1.2~4.5倍,土壤水分含量提高10%~140%[63].不同植被覆盖类型下热带、温带以及干旱半干旱区生态系统中平均树干径流量占同期降雨量的3.5%、11.3%和19.0%[64].这一过程可能是荒漠耐旱灌木适应干旱瘠薄立地条件重要生态水文机理[1].在沙坡头人工固沙植被区,柠条和油蒿的树干径流占降雨量的8.99%和2.82%[65],但也有研究认为这有可能过高估计树干径流对灌丛根际区的补给作用[66].主要原因有:一是沙坡头地区以小降雨事件为主,加之较高的潜在蒸散量,不利于树干径流形成;二是由于植被区灌木和半灌木茎干直径较小,且柠条枝条一般为多枝结构,油蒿叶表面覆盖一层不透水蜡质,这种特殊植物冠层结构不利于树干径流形成[18].虽然这一比例较小,但在干旱地区,即使是少量的水分补充对植物的生长也具有重要意义. ...
... 荒漠植物生理生态特征对降水响应亦具有尺度依赖性.植物自身生理特点,如植物光合和蒸腾作用对脉冲降水响应,主要表现在短时间尺度上[99].在日尺度上,小降水事件增加植物蒸腾,而大降水事件降低植物蒸腾,主要原因是小降水事件降低植物叶片水势,而后在很短时间内迅速恢复.但在降水量较大时,植物叶片水势增大,太阳辐射、空气温度以及饱和水汽压差均在之后很长一段时间内保持较低水平,因此植物蒸腾量下降.另外树干液流与蒸腾之间存在时间滞后[100].在长时间尺度上,植物对脉冲降水响应主要体现在种群结构、植被盖度或植物优势种的变化等方面.柠条和油蒿的叶面积指数,NBL和蒸腾均与当地年降水量显著线性正相关.间歇性降水,特别是0~5 mm和0~5 d内降水量,对水分胁迫下植被生存起着积极作用.很少水量可以激活干旱沙漠地区诸多生物过程,如种子发芽和繁殖等.降水集中于某一个时间段,则有可能造成土壤水分流失影响植被的可持续性和生存[101].间歇性降水导致土壤水分在局部和短时间内升高,植物能进行充分利用,同时一部分降水在土壤表层快速蒸发后通过入渗进入深层土壤,干旱期间供植物使用[102].通常情况下,这种类型的脉冲降水更有利于油蒿生存,油蒿根系主要分布在土壤表层(0~40 cm),柠条根部主要分布在1 m的土层之下,其根系利用渗透水的机会较少[17-18].另外小降水事件能促进土壤养分吸收.研究表明较小强度降水事件(降水强度2.8 mm·h-1)中,37.1%侵蚀产物、30.3%有机质、48.3%氮和30.0%溶解养分被重新分配至灌丛斑块;而在较大强度降水事件中(降水强度5.7 mm·h-1),31.1%侵蚀产物、9.84%有机质、19.0%氮和10.0%溶解养分被重新分配至灌丛斑块,其中大部分被径流所带走[103]. ...
腾格里沙漠东南缘不同年代固沙区根系分布
2
2013
... 干旱区植物生态适应性是其水分利用策略的重要组成部分.在长期的进化过程中,植物形成了多种适应干旱环境的机制,如特定的细胞损伤修复机理[15]、异化的叶片或同化枝[16]、发达的根系[17-19]、特殊的植物体渗透调节能力[20-21]、独特的解剖结构和水分生理代谢方式[22]等.在水文过程中主要体现为大孔隙优先流入根道及土壤动物活动产生的孔隙[23]、根系水力再分配[24]、树干径流[25]和叶片吸水[26]等特殊的干旱区植物生存和水分利用策略.Li等[27]从生态系统角度分析,发现干旱区固沙植被在演替中,土壤表面轻度干扰,如土壤动物(蚂蚁、蜥蜴等)洞穴等促进降水向深层土壤入渗. ...
... 植被和土壤之间协同演替关系是沙漠生态系统稳定性的基础[2].腾格里沙漠随着固沙植被建立,植被在从人工系统向自然系统的演变中经历了覆盖度从增加到减少的自疏过程、从灌木到半灌木再到草本的变化,土壤从流动风沙土向钙积旱成土发育[130].固沙灌木与土壤之间存在着复杂的互馈关系,这种关系在维持生态系统稳定性方面发挥着关键作用.一方面,固沙植物通过根系生长、凋落物分解等过程影响土壤的物理和化学性质[19,131].例如植物根系在生长过程中分泌有机物质,增加土壤有机碳含量,改善土壤结构和保水能力[132];同时,根系周转和凋落物分解也为土壤提供丰富养分,促进土壤肥力提升.另一方面,土壤水分和养分状况又反作用于植物的生长和发育.适宜的土壤水分条件能够促进植物根系扩展和吸收功能,提高植物水分利用效率和抗旱能力;而土壤养分则直接影响植物生长速度、生物量积累以及繁殖能力[133].不同演替阶段柠条的水分利用特征表明:演替早期,土壤水分含量决定柠条的水分利用来源,演替中后期,其决定因子分别为植物的基径和土壤黏粉粒含量[134]. ...
油蒿不同部位水分关系研究
1
2007
... 干旱区植物生态适应性是其水分利用策略的重要组成部分.在长期的进化过程中,植物形成了多种适应干旱环境的机制,如特定的细胞损伤修复机理[15]、异化的叶片或同化枝[16]、发达的根系[17-19]、特殊的植物体渗透调节能力[20-21]、独特的解剖结构和水分生理代谢方式[22]等.在水文过程中主要体现为大孔隙优先流入根道及土壤动物活动产生的孔隙[23]、根系水力再分配[24]、树干径流[25]和叶片吸水[26]等特殊的干旱区植物生存和水分利用策略.Li等[27]从生态系统角度分析,发现干旱区固沙植被在演替中,土壤表面轻度干扰,如土壤动物(蚂蚁、蜥蜴等)洞穴等促进降水向深层土壤入渗. ...
More sensitive to drought of young tissues with weak water potential adjustment capacity in two desert shrubs
1
2021
... 干旱区植物生态适应性是其水分利用策略的重要组成部分.在长期的进化过程中,植物形成了多种适应干旱环境的机制,如特定的细胞损伤修复机理[15]、异化的叶片或同化枝[16]、发达的根系[17-19]、特殊的植物体渗透调节能力[20-21]、独特的解剖结构和水分生理代谢方式[22]等.在水文过程中主要体现为大孔隙优先流入根道及土壤动物活动产生的孔隙[23]、根系水力再分配[24]、树干径流[25]和叶片吸水[26]等特殊的干旱区植物生存和水分利用策略.Li等[27]从生态系统角度分析,发现干旱区固沙植被在演替中,土壤表面轻度干扰,如土壤动物(蚂蚁、蜥蜴等)洞穴等促进降水向深层土壤入渗. ...
油蒿(Artemisia ordosica)和柠条(Caragana korshinskii)生理生态特性的昼夜变化特征与调节机制
1
2015
... 干旱区植物生态适应性是其水分利用策略的重要组成部分.在长期的进化过程中,植物形成了多种适应干旱环境的机制,如特定的细胞损伤修复机理[15]、异化的叶片或同化枝[16]、发达的根系[17-19]、特殊的植物体渗透调节能力[20-21]、独特的解剖结构和水分生理代谢方式[22]等.在水文过程中主要体现为大孔隙优先流入根道及土壤动物活动产生的孔隙[23]、根系水力再分配[24]、树干径流[25]和叶片吸水[26]等特殊的干旱区植物生存和水分利用策略.Li等[27]从生态系统角度分析,发现干旱区固沙植被在演替中,土壤表面轻度干扰,如土壤动物(蚂蚁、蜥蜴等)洞穴等促进降水向深层土壤入渗. ...
Desert shrub stemflow and its significance in soil moisture replenishment
1
2011
... 干旱区植物生态适应性是其水分利用策略的重要组成部分.在长期的进化过程中,植物形成了多种适应干旱环境的机制,如特定的细胞损伤修复机理[15]、异化的叶片或同化枝[16]、发达的根系[17-19]、特殊的植物体渗透调节能力[20-21]、独特的解剖结构和水分生理代谢方式[22]等.在水文过程中主要体现为大孔隙优先流入根道及土壤动物活动产生的孔隙[23]、根系水力再分配[24]、树干径流[25]和叶片吸水[26]等特殊的干旱区植物生存和水分利用策略.Li等[27]从生态系统角度分析,发现干旱区固沙植被在演替中,土壤表面轻度干扰,如土壤动物(蚂蚁、蜥蜴等)洞穴等促进降水向深层土壤入渗. ...
Magnitude and determinants of plant root hydraulic redistribution:a global synthesis analysis
2
2022
... 干旱区植物生态适应性是其水分利用策略的重要组成部分.在长期的进化过程中,植物形成了多种适应干旱环境的机制,如特定的细胞损伤修复机理[15]、异化的叶片或同化枝[16]、发达的根系[17-19]、特殊的植物体渗透调节能力[20-21]、独特的解剖结构和水分生理代谢方式[22]等.在水文过程中主要体现为大孔隙优先流入根道及土壤动物活动产生的孔隙[23]、根系水力再分配[24]、树干径流[25]和叶片吸水[26]等特殊的干旱区植物生存和水分利用策略.Li等[27]从生态系统角度分析,发现干旱区固沙植被在演替中,土壤表面轻度干扰,如土壤动物(蚂蚁、蜥蜴等)洞穴等促进降水向深层土壤入渗. ...
... 在干旱沙区,降水进入根际土壤后,根土界面的水力再分配(HR)被认为是植物适应干旱逆境的重要生态水文机制[31-32],主要包括水力提升、逆向和侧向水力提升3种模式[33].自Richards等[34]首次明确灌木三齿篙(Artemisia tridentata)在野外条件下存在水力提升现象以来,植物根-土界面的水力再分配已成为国际生态学界研究的热点,并取得了重要进展[35-39].水力再分配为植物提供了在土壤中暂时的储水机制,作为邻近植物的重要水源,能够极大地促进植物生长,对净初级生产力、生态系统水分平衡和养分循环有着重要影响[40].国内学者也对此进行了大量的报道和研究[41-49],在不同时空尺度上检验和评价HR过程机理及其生态学意义.迄今为止已经证明有近120种植物存在水力再分配现象[32,50],主要分布于干旱半干旱地区[51]或有明显季节性土壤干旱的湿润和半湿润地区[52].在全球尺度上,HR的平均大小为0.249 mm·d-1(95% Cl,0.113~0.384),其对植物蒸腾作用的贡献平均为27.4%(3%~79%),温带森林的HR最大(0.502 mm·d-1),而针叶林和阔叶林的HR最小(0.100 mm·d-1),被子植物的HR显著高于裸子植物(P<0.05),干燥指数和土壤质地显著影响HR[24].在沙坡头固沙植被区,研究结果表明柠条和油蒿等典型固沙灌木根系能够在土壤不同层次之间进行水分重新分配[53].例如,在遭受干旱胁迫时,柠条和油蒿根系会从深层向表层输送水分,维持表层土壤的湿润程度,促进灌丛下草本植物的生长.监测显示柠条根系水力再分配量为0.01~0.182 mm·d-1,5月达到最大值,10月则降至最小,平均值为0.079 mm·d-1;油蒿根系水力再分配量相对较低为0.01~0.05 mm·d-1,平均值为0.03 mm·d-1.这表明柠条在水力再分配方面能力较强,可能与其根系形态和分布特征有关. ...
Rainfall partitioning by vegetation in China:a quantitative synthesis
1
2022
... 干旱区植物生态适应性是其水分利用策略的重要组成部分.在长期的进化过程中,植物形成了多种适应干旱环境的机制,如特定的细胞损伤修复机理[15]、异化的叶片或同化枝[16]、发达的根系[17-19]、特殊的植物体渗透调节能力[20-21]、独特的解剖结构和水分生理代谢方式[22]等.在水文过程中主要体现为大孔隙优先流入根道及土壤动物活动产生的孔隙[23]、根系水力再分配[24]、树干径流[25]和叶片吸水[26]等特殊的干旱区植物生存和水分利用策略.Li等[27]从生态系统角度分析,发现干旱区固沙植被在演替中,土壤表面轻度干扰,如土壤动物(蚂蚁、蜥蜴等)洞穴等促进降水向深层土壤入渗. ...
柠条(Caragana korshinskii)、油蒿(Artemisia ordosica)、花棒(Hedysarum scoparium)叶片吸水潜力及影响因素
3
2020
... 干旱区植物生态适应性是其水分利用策略的重要组成部分.在长期的进化过程中,植物形成了多种适应干旱环境的机制,如特定的细胞损伤修复机理[15]、异化的叶片或同化枝[16]、发达的根系[17-19]、特殊的植物体渗透调节能力[20-21]、独特的解剖结构和水分生理代谢方式[22]等.在水文过程中主要体现为大孔隙优先流入根道及土壤动物活动产生的孔隙[23]、根系水力再分配[24]、树干径流[25]和叶片吸水[26]等特殊的干旱区植物生存和水分利用策略.Li等[27]从生态系统角度分析,发现干旱区固沙植被在演替中,土壤表面轻度干扰,如土壤动物(蚂蚁、蜥蜴等)洞穴等促进降水向深层土壤入渗. ...
... 叶片吸水是指植物不通过根系,而直接通过叶片来吸取水分的能力,同时也是植物利用隐匿降水(如雾或凝结水)的重要方式[67],特别是在小降水事件占主导的干旱荒漠人工植被区[1].关于植物叶片吸水的研究可以追溯到20世纪50年代,Stone[68]在北美西部地区首次发现黄松(Pinus ponderosa)植物通过叶片摄取水分,但是由于局限于特定环境的特定对象,此现象并未得到足够重视[69].直到近些年来越来越多的研究发现叶片吸水是植物最大限度地利用有限的水资源的重要途径[70].迄今为止,国内外发现有70余种植物通过叶片吸收水分[71].主要分布在北方阔叶针叶林[72-73]、热带雨林[74]、稀疏草原[75]、温带森林[76]和温带草原[77]等.国际上对云雾林的研究也证实了这一现象,雾水可直接通过叶片角质层扩散至根茎,占叶片总含水量的42%[71-72].对生活在地中海地区的常绿灌木迷迭香(Rosmarinus officinalis)和西班牙薰衣草(Lavandula stoechas)的研究结果表明:干旱时期有一半时间出现凝结水吸收使叶片相对含水量提高了72%,水势提高了0.5 MPa,水分亏缺状况得到缓解[78].国内关于植物叶片吸水的研究主要在西双版纳雨林生态系统的附生植物与非附生植物叶片[79],及干旱荒漠地区如准噶尔盆地[80]、民勤[81]、毛乌素沙地[82-83]和腾格里沙漠人工植被区[26]等植物均具有叶片吸水现象,且荒漠植物地上(叶片)和地下(根系)部分耗水及吸水策略存在权衡[80]. ...
... 叶片吸水对干旱沙区植物具有重要生理和生态作用.如光合性能改善,植物组织含水量增加,甚至延长叶片寿命、增加结实量、促进植物生长等[75,84].在沙坡头人工固沙植被区,小于5 mm的降水占到总降水频次的63.9%[85],凝结水量也占全年降水量的10%~20%[86],通过同位素示踪技术,在沙坡头人工植被区证明了典型固沙灌木也存在叶片吸水的现象,量化了柠条和油蒿单位面积吸水量分别为3.03~11.02、5.86~24.76 g,单位面积植物群落水分吸收能力分别为300~600、600~800 g,相当于0.3~0.8 mm的同期降雨量.[26].这表明叶片吸水在一定程度上能够补充植物体内的水分亏缺,尤其在干旱时期对维持植物的生理活性具有重要作用. ...
Ants mediate soil water in arid desert ecosystems:mitigating rainfall interception induced by biological soil crusts?
1
2014
... 干旱区植物生态适应性是其水分利用策略的重要组成部分.在长期的进化过程中,植物形成了多种适应干旱环境的机制,如特定的细胞损伤修复机理[15]、异化的叶片或同化枝[16]、发达的根系[17-19]、特殊的植物体渗透调节能力[20-21]、独特的解剖结构和水分生理代谢方式[22]等.在水文过程中主要体现为大孔隙优先流入根道及土壤动物活动产生的孔隙[23]、根系水力再分配[24]、树干径流[25]和叶片吸水[26]等特殊的干旱区植物生存和水分利用策略.Li等[27]从生态系统角度分析,发现干旱区固沙植被在演替中,土壤表面轻度干扰,如土壤动物(蚂蚁、蜥蜴等)洞穴等促进降水向深层土壤入渗. ...
长期生态学研究引领中国沙区的生态重建与恢复
1
2017
... 腾格里沙漠东南缘人工植被区是中国植物固沙领域的先例.自1956年起科研人员在此开展系统固沙植被建设,历经近70年不懈努力,成功探索出有效的治沙模式,为中国乃至全球荒漠化治理提供实践经验.在治理过程中,首先采用草方格固沙技术,通过增加地表粗糙度有效降低风速,成功固定流沙,为植被的生长创造了有利条件.在此基础上,筛选并种植柠条(Caragana korshinskii)、油蒿(Artemisia ordosica)、花棒(Hedysarum scoparium)等耐旱抗风沙植物,逐步构建起稳定的植被群落.经过多年的持续治理,包兰铁路两侧形成了一条长55 km、宽500~1 000 m的绿色长廊,确保包兰铁路畅通无阻,极大地改善了区域生态环境[28].但近年来由于全球气候变化,人工植被生态系统抵御自然风险能力降低,植被稳定性问题引起了国内外学者广泛关注[29].一方面随着流动沙丘固定和生境改变,生物类群剧增,人工植被逐渐向多功能复合系统演变[7].另一方面,由于严酷的生境条件,特别是有限的水分,驱使物种之间竞争加剧,使早期栽植的固沙灌木逐渐退化,植被盖度大幅下降.其主要的原因是土壤水分降低和水分循环特征改变[3].植被演变直接关系到防风固沙效益,反过来又影响整个固沙防护体系的稳定性[30].因此,深入了解腾格里沙漠典型固沙灌木生存的生态水文学机理成为目前植被建设中关键的科学问题.为此,本文分析和综述了腾格里沙漠典型固沙灌木多途径水分利用策略及其稳定性维持机制,为干旱沙区植被恢复与重建后的植被稳定性及其变化趋势评价提供理论依据. ...
The stability of revegetated ecosystems in sandy areas:an assessment and prediction index
1
2015
... 腾格里沙漠东南缘人工植被区是中国植物固沙领域的先例.自1956年起科研人员在此开展系统固沙植被建设,历经近70年不懈努力,成功探索出有效的治沙模式,为中国乃至全球荒漠化治理提供实践经验.在治理过程中,首先采用草方格固沙技术,通过增加地表粗糙度有效降低风速,成功固定流沙,为植被的生长创造了有利条件.在此基础上,筛选并种植柠条(Caragana korshinskii)、油蒿(Artemisia ordosica)、花棒(Hedysarum scoparium)等耐旱抗风沙植物,逐步构建起稳定的植被群落.经过多年的持续治理,包兰铁路两侧形成了一条长55 km、宽500~1 000 m的绿色长廊,确保包兰铁路畅通无阻,极大地改善了区域生态环境[28].但近年来由于全球气候变化,人工植被生态系统抵御自然风险能力降低,植被稳定性问题引起了国内外学者广泛关注[29].一方面随着流动沙丘固定和生境改变,生物类群剧增,人工植被逐渐向多功能复合系统演变[7].另一方面,由于严酷的生境条件,特别是有限的水分,驱使物种之间竞争加剧,使早期栽植的固沙灌木逐渐退化,植被盖度大幅下降.其主要的原因是土壤水分降低和水分循环特征改变[3].植被演变直接关系到防风固沙效益,反过来又影响整个固沙防护体系的稳定性[30].因此,深入了解腾格里沙漠典型固沙灌木生存的生态水文学机理成为目前植被建设中关键的科学问题.为此,本文分析和综述了腾格里沙漠典型固沙灌木多途径水分利用策略及其稳定性维持机制,为干旱沙区植被恢复与重建后的植被稳定性及其变化趋势评价提供理论依据. ...
中国干旱区恢复生态学研究进展及趋势评述
1
2014
... 腾格里沙漠东南缘人工植被区是中国植物固沙领域的先例.自1956年起科研人员在此开展系统固沙植被建设,历经近70年不懈努力,成功探索出有效的治沙模式,为中国乃至全球荒漠化治理提供实践经验.在治理过程中,首先采用草方格固沙技术,通过增加地表粗糙度有效降低风速,成功固定流沙,为植被的生长创造了有利条件.在此基础上,筛选并种植柠条(Caragana korshinskii)、油蒿(Artemisia ordosica)、花棒(Hedysarum scoparium)等耐旱抗风沙植物,逐步构建起稳定的植被群落.经过多年的持续治理,包兰铁路两侧形成了一条长55 km、宽500~1 000 m的绿色长廊,确保包兰铁路畅通无阻,极大地改善了区域生态环境[28].但近年来由于全球气候变化,人工植被生态系统抵御自然风险能力降低,植被稳定性问题引起了国内外学者广泛关注[29].一方面随着流动沙丘固定和生境改变,生物类群剧增,人工植被逐渐向多功能复合系统演变[7].另一方面,由于严酷的生境条件,特别是有限的水分,驱使物种之间竞争加剧,使早期栽植的固沙灌木逐渐退化,植被盖度大幅下降.其主要的原因是土壤水分降低和水分循环特征改变[3].植被演变直接关系到防风固沙效益,反过来又影响整个固沙防护体系的稳定性[30].因此,深入了解腾格里沙漠典型固沙灌木生存的生态水文学机理成为目前植被建设中关键的科学问题.为此,本文分析和综述了腾格里沙漠典型固沙灌木多途径水分利用策略及其稳定性维持机制,为干旱沙区植被恢复与重建后的植被稳定性及其变化趋势评价提供理论依据. ...
干旱区根土界面水分再分配及其生态水文效应研究进展与展望
1
2008
... 在干旱沙区,降水进入根际土壤后,根土界面的水力再分配(HR)被认为是植物适应干旱逆境的重要生态水文机制[31-32],主要包括水力提升、逆向和侧向水力提升3种模式[33].自Richards等[34]首次明确灌木三齿篙(Artemisia tridentata)在野外条件下存在水力提升现象以来,植物根-土界面的水力再分配已成为国际生态学界研究的热点,并取得了重要进展[35-39].水力再分配为植物提供了在土壤中暂时的储水机制,作为邻近植物的重要水源,能够极大地促进植物生长,对净初级生产力、生态系统水分平衡和养分循环有着重要影响[40].国内学者也对此进行了大量的报道和研究[41-49],在不同时空尺度上检验和评价HR过程机理及其生态学意义.迄今为止已经证明有近120种植物存在水力再分配现象[32,50],主要分布于干旱半干旱地区[51]或有明显季节性土壤干旱的湿润和半湿润地区[52].在全球尺度上,HR的平均大小为0.249 mm·d-1(95% Cl,0.113~0.384),其对植物蒸腾作用的贡献平均为27.4%(3%~79%),温带森林的HR最大(0.502 mm·d-1),而针叶林和阔叶林的HR最小(0.100 mm·d-1),被子植物的HR显著高于裸子植物(P<0.05),干燥指数和土壤质地显著影响HR[24].在沙坡头固沙植被区,研究结果表明柠条和油蒿等典型固沙灌木根系能够在土壤不同层次之间进行水分重新分配[53].例如,在遭受干旱胁迫时,柠条和油蒿根系会从深层向表层输送水分,维持表层土壤的湿润程度,促进灌丛下草本植物的生长.监测显示柠条根系水力再分配量为0.01~0.182 mm·d-1,5月达到最大值,10月则降至最小,平均值为0.079 mm·d-1;油蒿根系水力再分配量相对较低为0.01~0.05 mm·d-1,平均值为0.03 mm·d-1.这表明柠条在水力再分配方面能力较强,可能与其根系形态和分布特征有关. ...
The magnitude of hydraulic redistribution by plant roots:a review and synthesis of empirical and modeling studies
2
2012
... 在干旱沙区,降水进入根际土壤后,根土界面的水力再分配(HR)被认为是植物适应干旱逆境的重要生态水文机制[31-32],主要包括水力提升、逆向和侧向水力提升3种模式[33].自Richards等[34]首次明确灌木三齿篙(Artemisia tridentata)在野外条件下存在水力提升现象以来,植物根-土界面的水力再分配已成为国际生态学界研究的热点,并取得了重要进展[35-39].水力再分配为植物提供了在土壤中暂时的储水机制,作为邻近植物的重要水源,能够极大地促进植物生长,对净初级生产力、生态系统水分平衡和养分循环有着重要影响[40].国内学者也对此进行了大量的报道和研究[41-49],在不同时空尺度上检验和评价HR过程机理及其生态学意义.迄今为止已经证明有近120种植物存在水力再分配现象[32,50],主要分布于干旱半干旱地区[51]或有明显季节性土壤干旱的湿润和半湿润地区[52].在全球尺度上,HR的平均大小为0.249 mm·d-1(95% Cl,0.113~0.384),其对植物蒸腾作用的贡献平均为27.4%(3%~79%),温带森林的HR最大(0.502 mm·d-1),而针叶林和阔叶林的HR最小(0.100 mm·d-1),被子植物的HR显著高于裸子植物(P<0.05),干燥指数和土壤质地显著影响HR[24].在沙坡头固沙植被区,研究结果表明柠条和油蒿等典型固沙灌木根系能够在土壤不同层次之间进行水分重新分配[53].例如,在遭受干旱胁迫时,柠条和油蒿根系会从深层向表层输送水分,维持表层土壤的湿润程度,促进灌丛下草本植物的生长.监测显示柠条根系水力再分配量为0.01~0.182 mm·d-1,5月达到最大值,10月则降至最小,平均值为0.079 mm·d-1;油蒿根系水力再分配量相对较低为0.01~0.05 mm·d-1,平均值为0.03 mm·d-1.这表明柠条在水力再分配方面能力较强,可能与其根系形态和分布特征有关. ...
... [32,50],主要分布于干旱半干旱地区[51]或有明显季节性土壤干旱的湿润和半湿润地区[52].在全球尺度上,HR的平均大小为0.249 mm·d-1(95% Cl,0.113~0.384),其对植物蒸腾作用的贡献平均为27.4%(3%~79%),温带森林的HR最大(0.502 mm·d-1),而针叶林和阔叶林的HR最小(0.100 mm·d-1),被子植物的HR显著高于裸子植物(P<0.05),干燥指数和土壤质地显著影响HR[24].在沙坡头固沙植被区,研究结果表明柠条和油蒿等典型固沙灌木根系能够在土壤不同层次之间进行水分重新分配[53].例如,在遭受干旱胁迫时,柠条和油蒿根系会从深层向表层输送水分,维持表层土壤的湿润程度,促进灌丛下草本植物的生长.监测显示柠条根系水力再分配量为0.01~0.182 mm·d-1,5月达到最大值,10月则降至最小,平均值为0.079 mm·d-1;油蒿根系水力再分配量相对较低为0.01~0.05 mm·d-1,平均值为0.03 mm·d-1.这表明柠条在水力再分配方面能力较强,可能与其根系形态和分布特征有关. ...
The redistribution of soil water by tree root systems
1
1998
... 在干旱沙区,降水进入根际土壤后,根土界面的水力再分配(HR)被认为是植物适应干旱逆境的重要生态水文机制[31-32],主要包括水力提升、逆向和侧向水力提升3种模式[33].自Richards等[34]首次明确灌木三齿篙(Artemisia tridentata)在野外条件下存在水力提升现象以来,植物根-土界面的水力再分配已成为国际生态学界研究的热点,并取得了重要进展[35-39].水力再分配为植物提供了在土壤中暂时的储水机制,作为邻近植物的重要水源,能够极大地促进植物生长,对净初级生产力、生态系统水分平衡和养分循环有着重要影响[40].国内学者也对此进行了大量的报道和研究[41-49],在不同时空尺度上检验和评价HR过程机理及其生态学意义.迄今为止已经证明有近120种植物存在水力再分配现象[32,50],主要分布于干旱半干旱地区[51]或有明显季节性土壤干旱的湿润和半湿润地区[52].在全球尺度上,HR的平均大小为0.249 mm·d-1(95% Cl,0.113~0.384),其对植物蒸腾作用的贡献平均为27.4%(3%~79%),温带森林的HR最大(0.502 mm·d-1),而针叶林和阔叶林的HR最小(0.100 mm·d-1),被子植物的HR显著高于裸子植物(P<0.05),干燥指数和土壤质地显著影响HR[24].在沙坡头固沙植被区,研究结果表明柠条和油蒿等典型固沙灌木根系能够在土壤不同层次之间进行水分重新分配[53].例如,在遭受干旱胁迫时,柠条和油蒿根系会从深层向表层输送水分,维持表层土壤的湿润程度,促进灌丛下草本植物的生长.监测显示柠条根系水力再分配量为0.01~0.182 mm·d-1,5月达到最大值,10月则降至最小,平均值为0.079 mm·d-1;油蒿根系水力再分配量相对较低为0.01~0.05 mm·d-1,平均值为0.03 mm·d-1.这表明柠条在水力再分配方面能力较强,可能与其根系形态和分布特征有关. ...
Hydraulic lift:substantial nocturnal water transport between soil layers by Artemisia tridentata roots
1
1987
... 在干旱沙区,降水进入根际土壤后,根土界面的水力再分配(HR)被认为是植物适应干旱逆境的重要生态水文机制[31-32],主要包括水力提升、逆向和侧向水力提升3种模式[33].自Richards等[34]首次明确灌木三齿篙(Artemisia tridentata)在野外条件下存在水力提升现象以来,植物根-土界面的水力再分配已成为国际生态学界研究的热点,并取得了重要进展[35-39].水力再分配为植物提供了在土壤中暂时的储水机制,作为邻近植物的重要水源,能够极大地促进植物生长,对净初级生产力、生态系统水分平衡和养分循环有着重要影响[40].国内学者也对此进行了大量的报道和研究[41-49],在不同时空尺度上检验和评价HR过程机理及其生态学意义.迄今为止已经证明有近120种植物存在水力再分配现象[32,50],主要分布于干旱半干旱地区[51]或有明显季节性土壤干旱的湿润和半湿润地区[52].在全球尺度上,HR的平均大小为0.249 mm·d-1(95% Cl,0.113~0.384),其对植物蒸腾作用的贡献平均为27.4%(3%~79%),温带森林的HR最大(0.502 mm·d-1),而针叶林和阔叶林的HR最小(0.100 mm·d-1),被子植物的HR显著高于裸子植物(P<0.05),干燥指数和土壤质地显著影响HR[24].在沙坡头固沙植被区,研究结果表明柠条和油蒿等典型固沙灌木根系能够在土壤不同层次之间进行水分重新分配[53].例如,在遭受干旱胁迫时,柠条和油蒿根系会从深层向表层输送水分,维持表层土壤的湿润程度,促进灌丛下草本植物的生长.监测显示柠条根系水力再分配量为0.01~0.182 mm·d-1,5月达到最大值,10月则降至最小,平均值为0.079 mm·d-1;油蒿根系水力再分配量相对较低为0.01~0.05 mm·d-1,平均值为0.03 mm·d-1.这表明柠条在水力再分配方面能力较强,可能与其根系形态和分布特征有关. ...
Hydraulic redistribution through the root systems of senesced plants
1
2005
... 在干旱沙区,降水进入根际土壤后,根土界面的水力再分配(HR)被认为是植物适应干旱逆境的重要生态水文机制[31-32],主要包括水力提升、逆向和侧向水力提升3种模式[33].自Richards等[34]首次明确灌木三齿篙(Artemisia tridentata)在野外条件下存在水力提升现象以来,植物根-土界面的水力再分配已成为国际生态学界研究的热点,并取得了重要进展[35-39].水力再分配为植物提供了在土壤中暂时的储水机制,作为邻近植物的重要水源,能够极大地促进植物生长,对净初级生产力、生态系统水分平衡和养分循环有着重要影响[40].国内学者也对此进行了大量的报道和研究[41-49],在不同时空尺度上检验和评价HR过程机理及其生态学意义.迄今为止已经证明有近120种植物存在水力再分配现象[32,50],主要分布于干旱半干旱地区[51]或有明显季节性土壤干旱的湿润和半湿润地区[52].在全球尺度上,HR的平均大小为0.249 mm·d-1(95% Cl,0.113~0.384),其对植物蒸腾作用的贡献平均为27.4%(3%~79%),温带森林的HR最大(0.502 mm·d-1),而针叶林和阔叶林的HR最小(0.100 mm·d-1),被子植物的HR显著高于裸子植物(P<0.05),干燥指数和土壤质地显著影响HR[24].在沙坡头固沙植被区,研究结果表明柠条和油蒿等典型固沙灌木根系能够在土壤不同层次之间进行水分重新分配[53].例如,在遭受干旱胁迫时,柠条和油蒿根系会从深层向表层输送水分,维持表层土壤的湿润程度,促进灌丛下草本植物的生长.监测显示柠条根系水力再分配量为0.01~0.182 mm·d-1,5月达到最大值,10月则降至最小,平均值为0.079 mm·d-1;油蒿根系水力再分配量相对较低为0.01~0.05 mm·d-1,平均值为0.03 mm·d-1.这表明柠条在水力再分配方面能力较强,可能与其根系形态和分布特征有关. ...
The ecohydrologic significance of hydraulic redistribution in a semiarid Savanna
2008
Impact of hydraulic redistribution on multispecies vegetation water use in a semiarid savanna ecosystem:an experimental and modeling synthesis
2018
Hydraulic lift:processes,methods,and practical implications for society
2021
Tree water uptake patterns across the globe
1
2024
... 在干旱沙区,降水进入根际土壤后,根土界面的水力再分配(HR)被认为是植物适应干旱逆境的重要生态水文机制[31-32],主要包括水力提升、逆向和侧向水力提升3种模式[33].自Richards等[34]首次明确灌木三齿篙(Artemisia tridentata)在野外条件下存在水力提升现象以来,植物根-土界面的水力再分配已成为国际生态学界研究的热点,并取得了重要进展[35-39].水力再分配为植物提供了在土壤中暂时的储水机制,作为邻近植物的重要水源,能够极大地促进植物生长,对净初级生产力、生态系统水分平衡和养分循环有着重要影响[40].国内学者也对此进行了大量的报道和研究[41-49],在不同时空尺度上检验和评价HR过程机理及其生态学意义.迄今为止已经证明有近120种植物存在水力再分配现象[32,50],主要分布于干旱半干旱地区[51]或有明显季节性土壤干旱的湿润和半湿润地区[52].在全球尺度上,HR的平均大小为0.249 mm·d-1(95% Cl,0.113~0.384),其对植物蒸腾作用的贡献平均为27.4%(3%~79%),温带森林的HR最大(0.502 mm·d-1),而针叶林和阔叶林的HR最小(0.100 mm·d-1),被子植物的HR显著高于裸子植物(P<0.05),干燥指数和土壤质地显著影响HR[24].在沙坡头固沙植被区,研究结果表明柠条和油蒿等典型固沙灌木根系能够在土壤不同层次之间进行水分重新分配[53].例如,在遭受干旱胁迫时,柠条和油蒿根系会从深层向表层输送水分,维持表层土壤的湿润程度,促进灌丛下草本植物的生长.监测显示柠条根系水力再分配量为0.01~0.182 mm·d-1,5月达到最大值,10月则降至最小,平均值为0.079 mm·d-1;油蒿根系水力再分配量相对较低为0.01~0.05 mm·d-1,平均值为0.03 mm·d-1.这表明柠条在水力再分配方面能力较强,可能与其根系形态和分布特征有关. ...
Hydraulic redistribution by deeply rooted grasses and its ecohydrologic implications in the southern Great Plains of North America
1
2021
... 在干旱沙区,降水进入根际土壤后,根土界面的水力再分配(HR)被认为是植物适应干旱逆境的重要生态水文机制[31-32],主要包括水力提升、逆向和侧向水力提升3种模式[33].自Richards等[34]首次明确灌木三齿篙(Artemisia tridentata)在野外条件下存在水力提升现象以来,植物根-土界面的水力再分配已成为国际生态学界研究的热点,并取得了重要进展[35-39].水力再分配为植物提供了在土壤中暂时的储水机制,作为邻近植物的重要水源,能够极大地促进植物生长,对净初级生产力、生态系统水分平衡和养分循环有着重要影响[40].国内学者也对此进行了大量的报道和研究[41-49],在不同时空尺度上检验和评价HR过程机理及其生态学意义.迄今为止已经证明有近120种植物存在水力再分配现象[32,50],主要分布于干旱半干旱地区[51]或有明显季节性土壤干旱的湿润和半湿润地区[52].在全球尺度上,HR的平均大小为0.249 mm·d-1(95% Cl,0.113~0.384),其对植物蒸腾作用的贡献平均为27.4%(3%~79%),温带森林的HR最大(0.502 mm·d-1),而针叶林和阔叶林的HR最小(0.100 mm·d-1),被子植物的HR显著高于裸子植物(P<0.05),干燥指数和土壤质地显著影响HR[24].在沙坡头固沙植被区,研究结果表明柠条和油蒿等典型固沙灌木根系能够在土壤不同层次之间进行水分重新分配[53].例如,在遭受干旱胁迫时,柠条和油蒿根系会从深层向表层输送水分,维持表层土壤的湿润程度,促进灌丛下草本植物的生长.监测显示柠条根系水力再分配量为0.01~0.182 mm·d-1,5月达到最大值,10月则降至最小,平均值为0.079 mm·d-1;油蒿根系水力再分配量相对较低为0.01~0.05 mm·d-1,平均值为0.03 mm·d-1.这表明柠条在水力再分配方面能力较强,可能与其根系形态和分布特征有关. ...
土壤-根系统水分再分配:土壤-植物-大气连续体中的一个小通路
1
2007
... 在干旱沙区,降水进入根际土壤后,根土界面的水力再分配(HR)被认为是植物适应干旱逆境的重要生态水文机制[31-32],主要包括水力提升、逆向和侧向水力提升3种模式[33].自Richards等[34]首次明确灌木三齿篙(Artemisia tridentata)在野外条件下存在水力提升现象以来,植物根-土界面的水力再分配已成为国际生态学界研究的热点,并取得了重要进展[35-39].水力再分配为植物提供了在土壤中暂时的储水机制,作为邻近植物的重要水源,能够极大地促进植物生长,对净初级生产力、生态系统水分平衡和养分循环有着重要影响[40].国内学者也对此进行了大量的报道和研究[41-49],在不同时空尺度上检验和评价HR过程机理及其生态学意义.迄今为止已经证明有近120种植物存在水力再分配现象[32,50],主要分布于干旱半干旱地区[51]或有明显季节性土壤干旱的湿润和半湿润地区[52].在全球尺度上,HR的平均大小为0.249 mm·d-1(95% Cl,0.113~0.384),其对植物蒸腾作用的贡献平均为27.4%(3%~79%),温带森林的HR最大(0.502 mm·d-1),而针叶林和阔叶林的HR最小(0.100 mm·d-1),被子植物的HR显著高于裸子植物(P<0.05),干燥指数和土壤质地显著影响HR[24].在沙坡头固沙植被区,研究结果表明柠条和油蒿等典型固沙灌木根系能够在土壤不同层次之间进行水分重新分配[53].例如,在遭受干旱胁迫时,柠条和油蒿根系会从深层向表层输送水分,维持表层土壤的湿润程度,促进灌丛下草本植物的生长.监测显示柠条根系水力再分配量为0.01~0.182 mm·d-1,5月达到最大值,10月则降至最小,平均值为0.079 mm·d-1;油蒿根系水力再分配量相对较低为0.01~0.05 mm·d-1,平均值为0.03 mm·d-1.这表明柠条在水力再分配方面能力较强,可能与其根系形态和分布特征有关. ...
胡杨根系水力提升作用的证据及其生态学意义
2
2009
... 植物水分利用策略多样性是荒漠生态系统维持功能稳定核心机制,其通过根系水力再分配、树干径流和叶片吸水3种关键生态水文适应机制的协同作用,塑造了独特的资源分配格局与抗逆能力.根系水力再分配作为深根系植物(如胡杨(Populus euphratica)、柽柳)核心策略,通过夜间液流将深层(60~240 cm)地下水提升至浅层土壤(0~150 cm),缓解自身干旱胁迫,为浅根系草本提供水分支持,促进多物种共存[42].胡杨根系水力提升可使0~150 cm土层含水量提升16%~38%,同时通过根际优先流增强养分循环效率[42-43].树干径流则通过茎干截留降水并定向输导至根区,形成局部沃岛效应,富集有机质(提升15%~30%)和养分(如N、P含量增加20%~45%)[57-59].叶片吸水通过形态(肉质化、角质层增厚)与生理(CAM途径、气孔夜间开放)双重适应,将蒸腾速率降低至日间最大值的20%~38%,同时利用渗透调节物质(如脯氨酸、可溶性糖)维持细胞水势稳定(ΔΨ≥ -1.5 MPa)[16].这3类策略时空耦合形成多层次水分利用网络.在时间维度上水力再分配主导夜间水分再分配,树干径流响应降水事件,而叶片吸水则在晨间高湿度时段激活;空间维度上调控地下水源网络、地表径流再分布与大气水分捕获.这共同构建土壤-植物-大气连续体动态平衡,通过增强水分时空异质性、维持关键种生理功能及促进微生境形成,最终保障荒漠生态系统的结构稳定与功能可持续性. ...
... [42-43].树干径流则通过茎干截留降水并定向输导至根区,形成局部沃岛效应,富集有机质(提升15%~30%)和养分(如N、P含量增加20%~45%)[57-59].叶片吸水通过形态(肉质化、角质层增厚)与生理(CAM途径、气孔夜间开放)双重适应,将蒸腾速率降低至日间最大值的20%~38%,同时利用渗透调节物质(如脯氨酸、可溶性糖)维持细胞水势稳定(ΔΨ≥ -1.5 MPa)[16].这3类策略时空耦合形成多层次水分利用网络.在时间维度上水力再分配主导夜间水分再分配,树干径流响应降水事件,而叶片吸水则在晨间高湿度时段激活;空间维度上调控地下水源网络、地表径流再分布与大气水分捕获.这共同构建土壤-植物-大气连续体动态平衡,通过增强水分时空异质性、维持关键种生理功能及促进微生境形成,最终保障荒漠生态系统的结构稳定与功能可持续性. ...
胡杨根系水力再分配的模式、大小及其影响因子
1
2014
... 植物水分利用策略多样性是荒漠生态系统维持功能稳定核心机制,其通过根系水力再分配、树干径流和叶片吸水3种关键生态水文适应机制的协同作用,塑造了独特的资源分配格局与抗逆能力.根系水力再分配作为深根系植物(如胡杨(Populus euphratica)、柽柳)核心策略,通过夜间液流将深层(60~240 cm)地下水提升至浅层土壤(0~150 cm),缓解自身干旱胁迫,为浅根系草本提供水分支持,促进多物种共存[42].胡杨根系水力提升可使0~150 cm土层含水量提升16%~38%,同时通过根际优先流增强养分循环效率[42-43].树干径流则通过茎干截留降水并定向输导至根区,形成局部沃岛效应,富集有机质(提升15%~30%)和养分(如N、P含量增加20%~45%)[57-59].叶片吸水通过形态(肉质化、角质层增厚)与生理(CAM途径、气孔夜间开放)双重适应,将蒸腾速率降低至日间最大值的20%~38%,同时利用渗透调节物质(如脯氨酸、可溶性糖)维持细胞水势稳定(ΔΨ≥ -1.5 MPa)[16].这3类策略时空耦合形成多层次水分利用网络.在时间维度上水力再分配主导夜间水分再分配,树干径流响应降水事件,而叶片吸水则在晨间高湿度时段激活;空间维度上调控地下水源网络、地表径流再分布与大气水分捕获.这共同构建土壤-植物-大气连续体动态平衡,通过增强水分时空异质性、维持关键种生理功能及促进微生境形成,最终保障荒漠生态系统的结构稳定与功能可持续性. ...
根系水力再分配对陆地碳水循环的影响:以亚马孙流域为例
2019
Evidence of hydraulic redistribution and its water sharing mechanism in a karst field of Southwest China
2022
Soil water hydraulic redistribution in a subtropical monsoon evergreen forest
2022
Populus euphratica counteracts drought stress through the dew coupling and root hydraulic redistribution processes
2023
毛白杨根系液流与水力再分配特征
1
2023
... 在干旱沙区,降水进入根际土壤后,根土界面的水力再分配(HR)被认为是植物适应干旱逆境的重要生态水文机制[31-32],主要包括水力提升、逆向和侧向水力提升3种模式[33].自Richards等[34]首次明确灌木三齿篙(Artemisia tridentata)在野外条件下存在水力提升现象以来,植物根-土界面的水力再分配已成为国际生态学界研究的热点,并取得了重要进展[35-39].水力再分配为植物提供了在土壤中暂时的储水机制,作为邻近植物的重要水源,能够极大地促进植物生长,对净初级生产力、生态系统水分平衡和养分循环有着重要影响[40].国内学者也对此进行了大量的报道和研究[41-49],在不同时空尺度上检验和评价HR过程机理及其生态学意义.迄今为止已经证明有近120种植物存在水力再分配现象[32,50],主要分布于干旱半干旱地区[51]或有明显季节性土壤干旱的湿润和半湿润地区[52].在全球尺度上,HR的平均大小为0.249 mm·d-1(95% Cl,0.113~0.384),其对植物蒸腾作用的贡献平均为27.4%(3%~79%),温带森林的HR最大(0.502 mm·d-1),而针叶林和阔叶林的HR最小(0.100 mm·d-1),被子植物的HR显著高于裸子植物(P<0.05),干燥指数和土壤质地显著影响HR[24].在沙坡头固沙植被区,研究结果表明柠条和油蒿等典型固沙灌木根系能够在土壤不同层次之间进行水分重新分配[53].例如,在遭受干旱胁迫时,柠条和油蒿根系会从深层向表层输送水分,维持表层土壤的湿润程度,促进灌丛下草本植物的生长.监测显示柠条根系水力再分配量为0.01~0.182 mm·d-1,5月达到最大值,10月则降至最小,平均值为0.079 mm·d-1;油蒿根系水力再分配量相对较低为0.01~0.05 mm·d-1,平均值为0.03 mm·d-1.这表明柠条在水力再分配方面能力较强,可能与其根系形态和分布特征有关. ...
植物根系水力再分配量及影响因素分析
1
2021
... 在干旱沙区,降水进入根际土壤后,根土界面的水力再分配(HR)被认为是植物适应干旱逆境的重要生态水文机制[31-32],主要包括水力提升、逆向和侧向水力提升3种模式[33].自Richards等[34]首次明确灌木三齿篙(Artemisia tridentata)在野外条件下存在水力提升现象以来,植物根-土界面的水力再分配已成为国际生态学界研究的热点,并取得了重要进展[35-39].水力再分配为植物提供了在土壤中暂时的储水机制,作为邻近植物的重要水源,能够极大地促进植物生长,对净初级生产力、生态系统水分平衡和养分循环有着重要影响[40].国内学者也对此进行了大量的报道和研究[41-49],在不同时空尺度上检验和评价HR过程机理及其生态学意义.迄今为止已经证明有近120种植物存在水力再分配现象[32,50],主要分布于干旱半干旱地区[51]或有明显季节性土壤干旱的湿润和半湿润地区[52].在全球尺度上,HR的平均大小为0.249 mm·d-1(95% Cl,0.113~0.384),其对植物蒸腾作用的贡献平均为27.4%(3%~79%),温带森林的HR最大(0.502 mm·d-1),而针叶林和阔叶林的HR最小(0.100 mm·d-1),被子植物的HR显著高于裸子植物(P<0.05),干燥指数和土壤质地显著影响HR[24].在沙坡头固沙植被区,研究结果表明柠条和油蒿等典型固沙灌木根系能够在土壤不同层次之间进行水分重新分配[53].例如,在遭受干旱胁迫时,柠条和油蒿根系会从深层向表层输送水分,维持表层土壤的湿润程度,促进灌丛下草本植物的生长.监测显示柠条根系水力再分配量为0.01~0.182 mm·d-1,5月达到最大值,10月则降至最小,平均值为0.079 mm·d-1;油蒿根系水力再分配量相对较低为0.01~0.05 mm·d-1,平均值为0.03 mm·d-1.这表明柠条在水力再分配方面能力较强,可能与其根系形态和分布特征有关. ...
Seasonal and diurnal patterns of soil water potential in the rhizosphere of blue oaks:evidence for hydraulic lift
1
2000
... 在干旱沙区,降水进入根际土壤后,根土界面的水力再分配(HR)被认为是植物适应干旱逆境的重要生态水文机制[31-32],主要包括水力提升、逆向和侧向水力提升3种模式[33].自Richards等[34]首次明确灌木三齿篙(Artemisia tridentata)在野外条件下存在水力提升现象以来,植物根-土界面的水力再分配已成为国际生态学界研究的热点,并取得了重要进展[35-39].水力再分配为植物提供了在土壤中暂时的储水机制,作为邻近植物的重要水源,能够极大地促进植物生长,对净初级生产力、生态系统水分平衡和养分循环有着重要影响[40].国内学者也对此进行了大量的报道和研究[41-49],在不同时空尺度上检验和评价HR过程机理及其生态学意义.迄今为止已经证明有近120种植物存在水力再分配现象[32,50],主要分布于干旱半干旱地区[51]或有明显季节性土壤干旱的湿润和半湿润地区[52].在全球尺度上,HR的平均大小为0.249 mm·d-1(95% Cl,0.113~0.384),其对植物蒸腾作用的贡献平均为27.4%(3%~79%),温带森林的HR最大(0.502 mm·d-1),而针叶林和阔叶林的HR最小(0.100 mm·d-1),被子植物的HR显著高于裸子植物(P<0.05),干燥指数和土壤质地显著影响HR[24].在沙坡头固沙植被区,研究结果表明柠条和油蒿等典型固沙灌木根系能够在土壤不同层次之间进行水分重新分配[53].例如,在遭受干旱胁迫时,柠条和油蒿根系会从深层向表层输送水分,维持表层土壤的湿润程度,促进灌丛下草本植物的生长.监测显示柠条根系水力再分配量为0.01~0.182 mm·d-1,5月达到最大值,10月则降至最小,平均值为0.079 mm·d-1;油蒿根系水力再分配量相对较低为0.01~0.05 mm·d-1,平均值为0.03 mm·d-1.这表明柠条在水力再分配方面能力较强,可能与其根系形态和分布特征有关. ...
Hydraulic redistribution of soil water by neotropical Savanna trees
1
2002
... 在干旱沙区,降水进入根际土壤后,根土界面的水力再分配(HR)被认为是植物适应干旱逆境的重要生态水文机制[31-32],主要包括水力提升、逆向和侧向水力提升3种模式[33].自Richards等[34]首次明确灌木三齿篙(Artemisia tridentata)在野外条件下存在水力提升现象以来,植物根-土界面的水力再分配已成为国际生态学界研究的热点,并取得了重要进展[35-39].水力再分配为植物提供了在土壤中暂时的储水机制,作为邻近植物的重要水源,能够极大地促进植物生长,对净初级生产力、生态系统水分平衡和养分循环有着重要影响[40].国内学者也对此进行了大量的报道和研究[41-49],在不同时空尺度上检验和评价HR过程机理及其生态学意义.迄今为止已经证明有近120种植物存在水力再分配现象[32,50],主要分布于干旱半干旱地区[51]或有明显季节性土壤干旱的湿润和半湿润地区[52].在全球尺度上,HR的平均大小为0.249 mm·d-1(95% Cl,0.113~0.384),其对植物蒸腾作用的贡献平均为27.4%(3%~79%),温带森林的HR最大(0.502 mm·d-1),而针叶林和阔叶林的HR最小(0.100 mm·d-1),被子植物的HR显著高于裸子植物(P<0.05),干燥指数和土壤质地显著影响HR[24].在沙坡头固沙植被区,研究结果表明柠条和油蒿等典型固沙灌木根系能够在土壤不同层次之间进行水分重新分配[53].例如,在遭受干旱胁迫时,柠条和油蒿根系会从深层向表层输送水分,维持表层土壤的湿润程度,促进灌丛下草本植物的生长.监测显示柠条根系水力再分配量为0.01~0.182 mm·d-1,5月达到最大值,10月则降至最小,平均值为0.079 mm·d-1;油蒿根系水力再分配量相对较低为0.01~0.05 mm·d-1,平均值为0.03 mm·d-1.这表明柠条在水力再分配方面能力较强,可能与其根系形态和分布特征有关. ...
Stable isotopic analysis on water utilization sources of two xerophytic shrubs in a revegetated desert area:Tengger Desert,China
2
2015
... 在干旱沙区,降水进入根际土壤后,根土界面的水力再分配(HR)被认为是植物适应干旱逆境的重要生态水文机制[31-32],主要包括水力提升、逆向和侧向水力提升3种模式[33].自Richards等[34]首次明确灌木三齿篙(Artemisia tridentata)在野外条件下存在水力提升现象以来,植物根-土界面的水力再分配已成为国际生态学界研究的热点,并取得了重要进展[35-39].水力再分配为植物提供了在土壤中暂时的储水机制,作为邻近植物的重要水源,能够极大地促进植物生长,对净初级生产力、生态系统水分平衡和养分循环有着重要影响[40].国内学者也对此进行了大量的报道和研究[41-49],在不同时空尺度上检验和评价HR过程机理及其生态学意义.迄今为止已经证明有近120种植物存在水力再分配现象[32,50],主要分布于干旱半干旱地区[51]或有明显季节性土壤干旱的湿润和半湿润地区[52].在全球尺度上,HR的平均大小为0.249 mm·d-1(95% Cl,0.113~0.384),其对植物蒸腾作用的贡献平均为27.4%(3%~79%),温带森林的HR最大(0.502 mm·d-1),而针叶林和阔叶林的HR最小(0.100 mm·d-1),被子植物的HR显著高于裸子植物(P<0.05),干燥指数和土壤质地显著影响HR[24].在沙坡头固沙植被区,研究结果表明柠条和油蒿等典型固沙灌木根系能够在土壤不同层次之间进行水分重新分配[53].例如,在遭受干旱胁迫时,柠条和油蒿根系会从深层向表层输送水分,维持表层土壤的湿润程度,促进灌丛下草本植物的生长.监测显示柠条根系水力再分配量为0.01~0.182 mm·d-1,5月达到最大值,10月则降至最小,平均值为0.079 mm·d-1;油蒿根系水力再分配量相对较低为0.01~0.05 mm·d-1,平均值为0.03 mm·d-1.这表明柠条在水力再分配方面能力较强,可能与其根系形态和分布特征有关. ...
... 固沙灌木地上和地下部分在生长发育过程中存在密切协调关系,是植物适应干旱环境、维持自身稳定的关键[122].干旱区植物根系通常具有深根性和广布性.例如白梭梭和梭梭的垂直根深逾5 m,可至地下水层吸收水分[123].柽柳主根和侧根均极为发达,主根可伸至地下水层,最深10 m以上[124].此外,一些植物如沙拐枣根系多为水平分布,但在地下水较深的地区,其根系也能垂直向下发展以获取水分[125].以上根系的形态和分布使得植物能够有效地利用土壤水分资源.长期干旱时植物通过落叶(如梭梭)、茎干休眠等方式减少地上部分的水分消耗,同时维持根系活力(如沙拐枣根系持续生长寻找水源).植物地上和地下部分在生理功能上也表现为协调关系[126].植物通过冠层截留降水,减少土壤水分蒸发.同时,植物根系能够将截留水分吸收并输送到地上部分,支持植物蒸腾作用和光合作用.固沙灌木冠层截留和树干径流在水分再分配中起着重要作用,能够提高根际区水分供应[55].植物在资源分配上也表现出对干旱环境的适应性,根据环境条件调整地上和地下部分的生物量分配,以优化对光、水和养分等资源的利用[127].干旱条件下植物可能会将更多的资源分配到根系,以增强水分获取能力.同时,植物叶片和细根在碳、氮、磷等元素的分配中也存在协同关系.西北干旱区荒漠植物叶片和细根在碳、氮、磷含量及其计量比上表现出显著的正相关关系[128],这表明植物在完成正常的生命活动时,地上与地下部分的养分分配存在协同作用.深根系植物(如梭梭)主根深入地下水源,地上部分冠层稀疏(减少蒸腾面积),形成高而疏的形态,减少水分竞争.浅根系植物(如沙柳)侧根广泛分布于表层,冠层密集,依赖小降水后对表层水分的快速利用.植物地上和地下部分之间存在信息流和信号传递的协调机制[129].植物可通过木质部和韧皮部传递激素、压力波和电信号等,以协调地上和地下部分的生长和发育.信号传递能够帮助植物快速响应环境变化,如干旱胁迫下气孔调节和根系水分吸收调整.柠条和油蒿细根生长量和死亡量分别受土壤水分和温度共同影响,且不同年代植物对土壤水分利用深度和比例存在差异[53].柠条主要利用60、100 cm的土壤水分,二者占全部水源的75%左右;而油蒿主要利用10、20 cm的土壤水分,二者占全部水源的80%左右.随着固沙植被年限增加,植物对土壤表层水分利用比例逐渐减少,这是植物为了适应深层土壤水分相对稳定环境而做出的生理调整,也反映了植物地上和地下部分在资源获取和分配上的协调性.干旱区植物通过地上与地下部分的形态互补、资源分配动态调整及生理信号联动,实现了水分获取、运输与消耗的高效协调[85].这种根冠协同机制是其适应极端干旱的核心策略,也为人工植被恢复中的物种选择和生态管理提供了理论依据. ...
Connecting ecohydrology and hydropedology in desert shrubs:stem flow as a source of preferential flow in soils
1
2009
... 树干径流是干旱半干旱区植被在降水再分配过程中不可忽视的组成部分[54-55].其在形成过程中能够改变降水的空间分布格局,影响水分和养分循环的速度和方向,亦对植物生存和生长起着至关重要作用[56].目前,国内外学者对干旱荒漠地区灌木的自然集水特征及其对植物根部土壤水分和营养元素补给进行研究[57-58],通过实验观测和模型方法确定不同灌木树干径流产生条件、影响因素、树干径流量与雨水特征关系等[55].这些研究表明荒漠灌木能够通过树干径流收集5%~10%(有时可达到40%)的降水并直接运送到植物根部,之后通过根道系统和土壤大孔隙下渗,将水分贮存在较深的土层中供植物在干旱缺水时利用[59],同时缓解了根际区降水入渗的浅层化问题[60-61].据估算通过灌木树干径流的聚集作用,根部土壤收集到的水分是当地降雨量的11~221倍[62],土壤湿润锋增加1.2~4.5倍,土壤水分含量提高10%~140%[63].不同植被覆盖类型下热带、温带以及干旱半干旱区生态系统中平均树干径流量占同期降雨量的3.5%、11.3%和19.0%[64].这一过程可能是荒漠耐旱灌木适应干旱瘠薄立地条件重要生态水文机理[1].在沙坡头人工固沙植被区,柠条和油蒿的树干径流占降雨量的8.99%和2.82%[65],但也有研究认为这有可能过高估计树干径流对灌丛根际区的补给作用[66].主要原因有:一是沙坡头地区以小降雨事件为主,加之较高的潜在蒸散量,不利于树干径流形成;二是由于植被区灌木和半灌木茎干直径较小,且柠条枝条一般为多枝结构,油蒿叶表面覆盖一层不透水蜡质,这种特殊植物冠层结构不利于树干径流形成[18].虽然这一比例较小,但在干旱地区,即使是少量的水分补充对植物的生长也具有重要意义. ...
Global quantitative synthesis of effects of biotic and abiotic factors on stemflow production in woody ecosystems
5
2021
... 树干径流是干旱半干旱区植被在降水再分配过程中不可忽视的组成部分[54-55].其在形成过程中能够改变降水的空间分布格局,影响水分和养分循环的速度和方向,亦对植物生存和生长起着至关重要作用[56].目前,国内外学者对干旱荒漠地区灌木的自然集水特征及其对植物根部土壤水分和营养元素补给进行研究[57-58],通过实验观测和模型方法确定不同灌木树干径流产生条件、影响因素、树干径流量与雨水特征关系等[55].这些研究表明荒漠灌木能够通过树干径流收集5%~10%(有时可达到40%)的降水并直接运送到植物根部,之后通过根道系统和土壤大孔隙下渗,将水分贮存在较深的土层中供植物在干旱缺水时利用[59],同时缓解了根际区降水入渗的浅层化问题[60-61].据估算通过灌木树干径流的聚集作用,根部土壤收集到的水分是当地降雨量的11~221倍[62],土壤湿润锋增加1.2~4.5倍,土壤水分含量提高10%~140%[63].不同植被覆盖类型下热带、温带以及干旱半干旱区生态系统中平均树干径流量占同期降雨量的3.5%、11.3%和19.0%[64].这一过程可能是荒漠耐旱灌木适应干旱瘠薄立地条件重要生态水文机理[1].在沙坡头人工固沙植被区,柠条和油蒿的树干径流占降雨量的8.99%和2.82%[65],但也有研究认为这有可能过高估计树干径流对灌丛根际区的补给作用[66].主要原因有:一是沙坡头地区以小降雨事件为主,加之较高的潜在蒸散量,不利于树干径流形成;二是由于植被区灌木和半灌木茎干直径较小,且柠条枝条一般为多枝结构,油蒿叶表面覆盖一层不透水蜡质,这种特殊植物冠层结构不利于树干径流形成[18].虽然这一比例较小,但在干旱地区,即使是少量的水分补充对植物的生长也具有重要意义. ...
... [55].这些研究表明荒漠灌木能够通过树干径流收集5%~10%(有时可达到40%)的降水并直接运送到植物根部,之后通过根道系统和土壤大孔隙下渗,将水分贮存在较深的土层中供植物在干旱缺水时利用[59],同时缓解了根际区降水入渗的浅层化问题[60-61].据估算通过灌木树干径流的聚集作用,根部土壤收集到的水分是当地降雨量的11~221倍[62],土壤湿润锋增加1.2~4.5倍,土壤水分含量提高10%~140%[63].不同植被覆盖类型下热带、温带以及干旱半干旱区生态系统中平均树干径流量占同期降雨量的3.5%、11.3%和19.0%[64].这一过程可能是荒漠耐旱灌木适应干旱瘠薄立地条件重要生态水文机理[1].在沙坡头人工固沙植被区,柠条和油蒿的树干径流占降雨量的8.99%和2.82%[65],但也有研究认为这有可能过高估计树干径流对灌丛根际区的补给作用[66].主要原因有:一是沙坡头地区以小降雨事件为主,加之较高的潜在蒸散量,不利于树干径流形成;二是由于植被区灌木和半灌木茎干直径较小,且柠条枝条一般为多枝结构,油蒿叶表面覆盖一层不透水蜡质,这种特殊植物冠层结构不利于树干径流形成[18].虽然这一比例较小,但在干旱地区,即使是少量的水分补充对植物的生长也具有重要意义. ...
... 在荒漠人工植被生态系统中,水分是限制植物生长的主要环境因子,不同植物类群对水分适应的多样性是维持生态系统平衡和可持续性的关键因素[1].对于固沙灌木和半灌木,不同生物学特性决定其维持生存的生态水文学机理不同.如柠条叶与嫩茎上丰富的软毛,为叶片吸收凝结水提供了有利条件;而油蒿根系主要分布在浅层土壤(0~20 cm),对树干径流的利用比例可能高于柠条[55,65].同时,植物水分利用对策也可能随着降水梯度的变化而改变,如东非萨瓦纳生境中特别干旱时段,水力提升现象和树干径流基本消失,合欢(Acacia tortilis)主要通过叶片吸水来适应干旱胁迫[87];当降水量为1.5~5.8 mm,许多植物如三齿团香木(Larrea tridentate)、美洲焦油菊(Flourensia cernua)和腺牧豆树(Prosopis glandulosa)产生树干径流,通过根系优先流来补充土壤水分[88];干季末期大降雨事件(50 mm)之后,萨瓦纳8种主要植物的根系传导水分状况表明:水分在水势梯度驱动下,先从浅层土壤进入浅层根,之后进入主根,最后再由主根进入深层较干燥土壤中,完成水力再分配[89].这说明植物水分利用方式或者对策会随着降雨量的变化而变化. ...
... 通过相关资料计算得出柠条和油蒿水量平衡各分量的分配比例[90](图2),结果显示在沙坡头多年平均降雨量(186 mm)与凝结水(25 mm)情况下,柠条冠层截留量较高(21.85 mm,占输入量的10.36%),穿透雨为163.5 mm,叶片吸水量可达5.2 mm,蒸腾(82.87 mm)与蒸发(121.16 mm)为主要耗水途径,树干径流为4.7 mm,土壤储水量波动较大(±23.92 mm).油蒿冠层截留量较柠条降至14.9 mm(占比6.74%),穿透雨增加至173.7 mm,叶片吸水量升至9.7 mm,蒸腾(83.9 mm)与蒸发(122.2 mm)略有增强,但树干径流大幅减少至1.3 mm,土壤储水量波动较小(±14.56 mm).柠条冠层截留能力更强,而油蒿冠层透水性更高,穿透雨比例增加,水分分配稳定性提升.两物种的水文差异可能源于冠层结构特性(如叶片密度、茎干形态)对降水截留、再分配及土壤水分动态调控作用[55,57]. ...
... 固沙灌木地上和地下部分在生长发育过程中存在密切协调关系,是植物适应干旱环境、维持自身稳定的关键[122].干旱区植物根系通常具有深根性和广布性.例如白梭梭和梭梭的垂直根深逾5 m,可至地下水层吸收水分[123].柽柳主根和侧根均极为发达,主根可伸至地下水层,最深10 m以上[124].此外,一些植物如沙拐枣根系多为水平分布,但在地下水较深的地区,其根系也能垂直向下发展以获取水分[125].以上根系的形态和分布使得植物能够有效地利用土壤水分资源.长期干旱时植物通过落叶(如梭梭)、茎干休眠等方式减少地上部分的水分消耗,同时维持根系活力(如沙拐枣根系持续生长寻找水源).植物地上和地下部分在生理功能上也表现为协调关系[126].植物通过冠层截留降水,减少土壤水分蒸发.同时,植物根系能够将截留水分吸收并输送到地上部分,支持植物蒸腾作用和光合作用.固沙灌木冠层截留和树干径流在水分再分配中起着重要作用,能够提高根际区水分供应[55].植物在资源分配上也表现出对干旱环境的适应性,根据环境条件调整地上和地下部分的生物量分配,以优化对光、水和养分等资源的利用[127].干旱条件下植物可能会将更多的资源分配到根系,以增强水分获取能力.同时,植物叶片和细根在碳、氮、磷等元素的分配中也存在协同关系.西北干旱区荒漠植物叶片和细根在碳、氮、磷含量及其计量比上表现出显著的正相关关系[128],这表明植物在完成正常的生命活动时,地上与地下部分的养分分配存在协同作用.深根系植物(如梭梭)主根深入地下水源,地上部分冠层稀疏(减少蒸腾面积),形成高而疏的形态,减少水分竞争.浅根系植物(如沙柳)侧根广泛分布于表层,冠层密集,依赖小降水后对表层水分的快速利用.植物地上和地下部分之间存在信息流和信号传递的协调机制[129].植物可通过木质部和韧皮部传递激素、压力波和电信号等,以协调地上和地下部分的生长和发育.信号传递能够帮助植物快速响应环境变化,如干旱胁迫下气孔调节和根系水分吸收调整.柠条和油蒿细根生长量和死亡量分别受土壤水分和温度共同影响,且不同年代植物对土壤水分利用深度和比例存在差异[53].柠条主要利用60、100 cm的土壤水分,二者占全部水源的75%左右;而油蒿主要利用10、20 cm的土壤水分,二者占全部水源的80%左右.随着固沙植被年限增加,植物对土壤表层水分利用比例逐渐减少,这是植物为了适应深层土壤水分相对稳定环境而做出的生理调整,也反映了植物地上和地下部分在资源获取和分配上的协调性.干旱区植物通过地上与地下部分的形态互补、资源分配动态调整及生理信号联动,实现了水分获取、运输与消耗的高效协调[85].这种根冠协同机制是其适应极端干旱的核心策略,也为人工植被恢复中的物种选择和生态管理提供了理论依据. ...
On the importance of stemflow to the woody plants in drylands:individual vs.ecosystem scales
1
2021
... 树干径流是干旱半干旱区植被在降水再分配过程中不可忽视的组成部分[54-55].其在形成过程中能够改变降水的空间分布格局,影响水分和养分循环的速度和方向,亦对植物生存和生长起着至关重要作用[56].目前,国内外学者对干旱荒漠地区灌木的自然集水特征及其对植物根部土壤水分和营养元素补给进行研究[57-58],通过实验观测和模型方法确定不同灌木树干径流产生条件、影响因素、树干径流量与雨水特征关系等[55].这些研究表明荒漠灌木能够通过树干径流收集5%~10%(有时可达到40%)的降水并直接运送到植物根部,之后通过根道系统和土壤大孔隙下渗,将水分贮存在较深的土层中供植物在干旱缺水时利用[59],同时缓解了根际区降水入渗的浅层化问题[60-61].据估算通过灌木树干径流的聚集作用,根部土壤收集到的水分是当地降雨量的11~221倍[62],土壤湿润锋增加1.2~4.5倍,土壤水分含量提高10%~140%[63].不同植被覆盖类型下热带、温带以及干旱半干旱区生态系统中平均树干径流量占同期降雨量的3.5%、11.3%和19.0%[64].这一过程可能是荒漠耐旱灌木适应干旱瘠薄立地条件重要生态水文机理[1].在沙坡头人工固沙植被区,柠条和油蒿的树干径流占降雨量的8.99%和2.82%[65],但也有研究认为这有可能过高估计树干径流对灌丛根际区的补给作用[66].主要原因有:一是沙坡头地区以小降雨事件为主,加之较高的潜在蒸散量,不利于树干径流形成;二是由于植被区灌木和半灌木茎干直径较小,且柠条枝条一般为多枝结构,油蒿叶表面覆盖一层不透水蜡质,这种特殊植物冠层结构不利于树干径流形成[18].虽然这一比例较小,但在干旱地区,即使是少量的水分补充对植物的生长也具有重要意义. ...
冠层降水再分配驱动生态系统养分富集:机制、数量与模式
3
2024
... 树干径流是干旱半干旱区植被在降水再分配过程中不可忽视的组成部分[54-55].其在形成过程中能够改变降水的空间分布格局,影响水分和养分循环的速度和方向,亦对植物生存和生长起着至关重要作用[56].目前,国内外学者对干旱荒漠地区灌木的自然集水特征及其对植物根部土壤水分和营养元素补给进行研究[57-58],通过实验观测和模型方法确定不同灌木树干径流产生条件、影响因素、树干径流量与雨水特征关系等[55].这些研究表明荒漠灌木能够通过树干径流收集5%~10%(有时可达到40%)的降水并直接运送到植物根部,之后通过根道系统和土壤大孔隙下渗,将水分贮存在较深的土层中供植物在干旱缺水时利用[59],同时缓解了根际区降水入渗的浅层化问题[60-61].据估算通过灌木树干径流的聚集作用,根部土壤收集到的水分是当地降雨量的11~221倍[62],土壤湿润锋增加1.2~4.5倍,土壤水分含量提高10%~140%[63].不同植被覆盖类型下热带、温带以及干旱半干旱区生态系统中平均树干径流量占同期降雨量的3.5%、11.3%和19.0%[64].这一过程可能是荒漠耐旱灌木适应干旱瘠薄立地条件重要生态水文机理[1].在沙坡头人工固沙植被区,柠条和油蒿的树干径流占降雨量的8.99%和2.82%[65],但也有研究认为这有可能过高估计树干径流对灌丛根际区的补给作用[66].主要原因有:一是沙坡头地区以小降雨事件为主,加之较高的潜在蒸散量,不利于树干径流形成;二是由于植被区灌木和半灌木茎干直径较小,且柠条枝条一般为多枝结构,油蒿叶表面覆盖一层不透水蜡质,这种特殊植物冠层结构不利于树干径流形成[18].虽然这一比例较小,但在干旱地区,即使是少量的水分补充对植物的生长也具有重要意义. ...
... 通过相关资料计算得出柠条和油蒿水量平衡各分量的分配比例[90](图2),结果显示在沙坡头多年平均降雨量(186 mm)与凝结水(25 mm)情况下,柠条冠层截留量较高(21.85 mm,占输入量的10.36%),穿透雨为163.5 mm,叶片吸水量可达5.2 mm,蒸腾(82.87 mm)与蒸发(121.16 mm)为主要耗水途径,树干径流为4.7 mm,土壤储水量波动较大(±23.92 mm).油蒿冠层截留量较柠条降至14.9 mm(占比6.74%),穿透雨增加至173.7 mm,叶片吸水量升至9.7 mm,蒸腾(83.9 mm)与蒸发(122.2 mm)略有增强,但树干径流大幅减少至1.3 mm,土壤储水量波动较小(±14.56 mm).柠条冠层截留能力更强,而油蒿冠层透水性更高,穿透雨比例增加,水分分配稳定性提升.两物种的水文差异可能源于冠层结构特性(如叶片密度、茎干形态)对降水截留、再分配及土壤水分动态调控作用[55,57]. ...
... 植物水分利用策略多样性是荒漠生态系统维持功能稳定核心机制,其通过根系水力再分配、树干径流和叶片吸水3种关键生态水文适应机制的协同作用,塑造了独特的资源分配格局与抗逆能力.根系水力再分配作为深根系植物(如胡杨(Populus euphratica)、柽柳)核心策略,通过夜间液流将深层(60~240 cm)地下水提升至浅层土壤(0~150 cm),缓解自身干旱胁迫,为浅根系草本提供水分支持,促进多物种共存[42].胡杨根系水力提升可使0~150 cm土层含水量提升16%~38%,同时通过根际优先流增强养分循环效率[42-43].树干径流则通过茎干截留降水并定向输导至根区,形成局部沃岛效应,富集有机质(提升15%~30%)和养分(如N、P含量增加20%~45%)[57-59].叶片吸水通过形态(肉质化、角质层增厚)与生理(CAM途径、气孔夜间开放)双重适应,将蒸腾速率降低至日间最大值的20%~38%,同时利用渗透调节物质(如脯氨酸、可溶性糖)维持细胞水势稳定(ΔΨ≥ -1.5 MPa)[16].这3类策略时空耦合形成多层次水分利用网络.在时间维度上水力再分配主导夜间水分再分配,树干径流响应降水事件,而叶片吸水则在晨间高湿度时段激活;空间维度上调控地下水源网络、地表径流再分布与大气水分捕获.这共同构建土壤-植物-大气连续体动态平衡,通过增强水分时空异质性、维持关键种生理功能及促进微生境形成,最终保障荒漠生态系统的结构稳定与功能可持续性. ...
Deciphering global patterns of forest canopy rainfall interception (FCRI):a synthesis of geographical,forest species,and methodological influences
1
2024
... 树干径流是干旱半干旱区植被在降水再分配过程中不可忽视的组成部分[54-55].其在形成过程中能够改变降水的空间分布格局,影响水分和养分循环的速度和方向,亦对植物生存和生长起着至关重要作用[56].目前,国内外学者对干旱荒漠地区灌木的自然集水特征及其对植物根部土壤水分和营养元素补给进行研究[57-58],通过实验观测和模型方法确定不同灌木树干径流产生条件、影响因素、树干径流量与雨水特征关系等[55].这些研究表明荒漠灌木能够通过树干径流收集5%~10%(有时可达到40%)的降水并直接运送到植物根部,之后通过根道系统和土壤大孔隙下渗,将水分贮存在较深的土层中供植物在干旱缺水时利用[59],同时缓解了根际区降水入渗的浅层化问题[60-61].据估算通过灌木树干径流的聚集作用,根部土壤收集到的水分是当地降雨量的11~221倍[62],土壤湿润锋增加1.2~4.5倍,土壤水分含量提高10%~140%[63].不同植被覆盖类型下热带、温带以及干旱半干旱区生态系统中平均树干径流量占同期降雨量的3.5%、11.3%和19.0%[64].这一过程可能是荒漠耐旱灌木适应干旱瘠薄立地条件重要生态水文机理[1].在沙坡头人工固沙植被区,柠条和油蒿的树干径流占降雨量的8.99%和2.82%[65],但也有研究认为这有可能过高估计树干径流对灌丛根际区的补给作用[66].主要原因有:一是沙坡头地区以小降雨事件为主,加之较高的潜在蒸散量,不利于树干径流形成;二是由于植被区灌木和半灌木茎干直径较小,且柠条枝条一般为多枝结构,油蒿叶表面覆盖一层不透水蜡质,这种特殊植物冠层结构不利于树干径流形成[18].虽然这一比例较小,但在干旱地区,即使是少量的水分补充对植物的生长也具有重要意义. ...
荒漠灌木树干茎流及其生态水文效应研究进展
2
2010
... 树干径流是干旱半干旱区植被在降水再分配过程中不可忽视的组成部分[54-55].其在形成过程中能够改变降水的空间分布格局,影响水分和养分循环的速度和方向,亦对植物生存和生长起着至关重要作用[56].目前,国内外学者对干旱荒漠地区灌木的自然集水特征及其对植物根部土壤水分和营养元素补给进行研究[57-58],通过实验观测和模型方法确定不同灌木树干径流产生条件、影响因素、树干径流量与雨水特征关系等[55].这些研究表明荒漠灌木能够通过树干径流收集5%~10%(有时可达到40%)的降水并直接运送到植物根部,之后通过根道系统和土壤大孔隙下渗,将水分贮存在较深的土层中供植物在干旱缺水时利用[59],同时缓解了根际区降水入渗的浅层化问题[60-61].据估算通过灌木树干径流的聚集作用,根部土壤收集到的水分是当地降雨量的11~221倍[62],土壤湿润锋增加1.2~4.5倍,土壤水分含量提高10%~140%[63].不同植被覆盖类型下热带、温带以及干旱半干旱区生态系统中平均树干径流量占同期降雨量的3.5%、11.3%和19.0%[64].这一过程可能是荒漠耐旱灌木适应干旱瘠薄立地条件重要生态水文机理[1].在沙坡头人工固沙植被区,柠条和油蒿的树干径流占降雨量的8.99%和2.82%[65],但也有研究认为这有可能过高估计树干径流对灌丛根际区的补给作用[66].主要原因有:一是沙坡头地区以小降雨事件为主,加之较高的潜在蒸散量,不利于树干径流形成;二是由于植被区灌木和半灌木茎干直径较小,且柠条枝条一般为多枝结构,油蒿叶表面覆盖一层不透水蜡质,这种特殊植物冠层结构不利于树干径流形成[18].虽然这一比例较小,但在干旱地区,即使是少量的水分补充对植物的生长也具有重要意义. ...
... 植物水分利用策略多样性是荒漠生态系统维持功能稳定核心机制,其通过根系水力再分配、树干径流和叶片吸水3种关键生态水文适应机制的协同作用,塑造了独特的资源分配格局与抗逆能力.根系水力再分配作为深根系植物(如胡杨(Populus euphratica)、柽柳)核心策略,通过夜间液流将深层(60~240 cm)地下水提升至浅层土壤(0~150 cm),缓解自身干旱胁迫,为浅根系草本提供水分支持,促进多物种共存[42].胡杨根系水力提升可使0~150 cm土层含水量提升16%~38%,同时通过根际优先流增强养分循环效率[42-43].树干径流则通过茎干截留降水并定向输导至根区,形成局部沃岛效应,富集有机质(提升15%~30%)和养分(如N、P含量增加20%~45%)[57-59].叶片吸水通过形态(肉质化、角质层增厚)与生理(CAM途径、气孔夜间开放)双重适应,将蒸腾速率降低至日间最大值的20%~38%,同时利用渗透调节物质(如脯氨酸、可溶性糖)维持细胞水势稳定(ΔΨ≥ -1.5 MPa)[16].这3类策略时空耦合形成多层次水分利用网络.在时间维度上水力再分配主导夜间水分再分配,树干径流响应降水事件,而叶片吸水则在晨间高湿度时段激活;空间维度上调控地下水源网络、地表径流再分布与大气水分捕获.这共同构建土壤-植物-大气连续体动态平衡,通过增强水分时空异质性、维持关键种生理功能及促进微生境形成,最终保障荒漠生态系统的结构稳定与功能可持续性. ...
黄土高原水蚀风蚀交错带降水及灌木林冠截留特性研究
1
2005
... 树干径流是干旱半干旱区植被在降水再分配过程中不可忽视的组成部分[54-55].其在形成过程中能够改变降水的空间分布格局,影响水分和养分循环的速度和方向,亦对植物生存和生长起着至关重要作用[56].目前,国内外学者对干旱荒漠地区灌木的自然集水特征及其对植物根部土壤水分和营养元素补给进行研究[57-58],通过实验观测和模型方法确定不同灌木树干径流产生条件、影响因素、树干径流量与雨水特征关系等[55].这些研究表明荒漠灌木能够通过树干径流收集5%~10%(有时可达到40%)的降水并直接运送到植物根部,之后通过根道系统和土壤大孔隙下渗,将水分贮存在较深的土层中供植物在干旱缺水时利用[59],同时缓解了根际区降水入渗的浅层化问题[60-61].据估算通过灌木树干径流的聚集作用,根部土壤收集到的水分是当地降雨量的11~221倍[62],土壤湿润锋增加1.2~4.5倍,土壤水分含量提高10%~140%[63].不同植被覆盖类型下热带、温带以及干旱半干旱区生态系统中平均树干径流量占同期降雨量的3.5%、11.3%和19.0%[64].这一过程可能是荒漠耐旱灌木适应干旱瘠薄立地条件重要生态水文机理[1].在沙坡头人工固沙植被区,柠条和油蒿的树干径流占降雨量的8.99%和2.82%[65],但也有研究认为这有可能过高估计树干径流对灌丛根际区的补给作用[66].主要原因有:一是沙坡头地区以小降雨事件为主,加之较高的潜在蒸散量,不利于树干径流形成;二是由于植被区灌木和半灌木茎干直径较小,且柠条枝条一般为多枝结构,油蒿叶表面覆盖一层不透水蜡质,这种特殊植物冠层结构不利于树干径流形成[18].虽然这一比例较小,但在干旱地区,即使是少量的水分补充对植物的生长也具有重要意义. ...
古尔班通古特沙漠梭梭和白梭梭树干茎流特征
1
2013
... 树干径流是干旱半干旱区植被在降水再分配过程中不可忽视的组成部分[54-55].其在形成过程中能够改变降水的空间分布格局,影响水分和养分循环的速度和方向,亦对植物生存和生长起着至关重要作用[56].目前,国内外学者对干旱荒漠地区灌木的自然集水特征及其对植物根部土壤水分和营养元素补给进行研究[57-58],通过实验观测和模型方法确定不同灌木树干径流产生条件、影响因素、树干径流量与雨水特征关系等[55].这些研究表明荒漠灌木能够通过树干径流收集5%~10%(有时可达到40%)的降水并直接运送到植物根部,之后通过根道系统和土壤大孔隙下渗,将水分贮存在较深的土层中供植物在干旱缺水时利用[59],同时缓解了根际区降水入渗的浅层化问题[60-61].据估算通过灌木树干径流的聚集作用,根部土壤收集到的水分是当地降雨量的11~221倍[62],土壤湿润锋增加1.2~4.5倍,土壤水分含量提高10%~140%[63].不同植被覆盖类型下热带、温带以及干旱半干旱区生态系统中平均树干径流量占同期降雨量的3.5%、11.3%和19.0%[64].这一过程可能是荒漠耐旱灌木适应干旱瘠薄立地条件重要生态水文机理[1].在沙坡头人工固沙植被区,柠条和油蒿的树干径流占降雨量的8.99%和2.82%[65],但也有研究认为这有可能过高估计树干径流对灌丛根际区的补给作用[66].主要原因有:一是沙坡头地区以小降雨事件为主,加之较高的潜在蒸散量,不利于树干径流形成;二是由于植被区灌木和半灌木茎干直径较小,且柠条枝条一般为多枝结构,油蒿叶表面覆盖一层不透水蜡质,这种特殊植物冠层结构不利于树干径流形成[18].虽然这一比例较小,但在干旱地区,即使是少量的水分补充对植物的生长也具有重要意义. ...
干旱地区土壤-植被-水文耦合、响应与适应机制
1
2011
... 树干径流是干旱半干旱区植被在降水再分配过程中不可忽视的组成部分[54-55].其在形成过程中能够改变降水的空间分布格局,影响水分和养分循环的速度和方向,亦对植物生存和生长起着至关重要作用[56].目前,国内外学者对干旱荒漠地区灌木的自然集水特征及其对植物根部土壤水分和营养元素补给进行研究[57-58],通过实验观测和模型方法确定不同灌木树干径流产生条件、影响因素、树干径流量与雨水特征关系等[55].这些研究表明荒漠灌木能够通过树干径流收集5%~10%(有时可达到40%)的降水并直接运送到植物根部,之后通过根道系统和土壤大孔隙下渗,将水分贮存在较深的土层中供植物在干旱缺水时利用[59],同时缓解了根际区降水入渗的浅层化问题[60-61].据估算通过灌木树干径流的聚集作用,根部土壤收集到的水分是当地降雨量的11~221倍[62],土壤湿润锋增加1.2~4.5倍,土壤水分含量提高10%~140%[63].不同植被覆盖类型下热带、温带以及干旱半干旱区生态系统中平均树干径流量占同期降雨量的3.5%、11.3%和19.0%[64].这一过程可能是荒漠耐旱灌木适应干旱瘠薄立地条件重要生态水文机理[1].在沙坡头人工固沙植被区,柠条和油蒿的树干径流占降雨量的8.99%和2.82%[65],但也有研究认为这有可能过高估计树干径流对灌丛根际区的补给作用[66].主要原因有:一是沙坡头地区以小降雨事件为主,加之较高的潜在蒸散量,不利于树干径流形成;二是由于植被区灌木和半灌木茎干直径较小,且柠条枝条一般为多枝结构,油蒿叶表面覆盖一层不透水蜡质,这种特殊植物冠层结构不利于树干径流形成[18].虽然这一比例较小,但在干旱地区,即使是少量的水分补充对植物的生长也具有重要意义. ...
Stem flow in three shrubs and its effect on soil water enhancement in semiarid loess region of China
1
2008
... 树干径流是干旱半干旱区植被在降水再分配过程中不可忽视的组成部分[54-55].其在形成过程中能够改变降水的空间分布格局,影响水分和养分循环的速度和方向,亦对植物生存和生长起着至关重要作用[56].目前,国内外学者对干旱荒漠地区灌木的自然集水特征及其对植物根部土壤水分和营养元素补给进行研究[57-58],通过实验观测和模型方法确定不同灌木树干径流产生条件、影响因素、树干径流量与雨水特征关系等[55].这些研究表明荒漠灌木能够通过树干径流收集5%~10%(有时可达到40%)的降水并直接运送到植物根部,之后通过根道系统和土壤大孔隙下渗,将水分贮存在较深的土层中供植物在干旱缺水时利用[59],同时缓解了根际区降水入渗的浅层化问题[60-61].据估算通过灌木树干径流的聚集作用,根部土壤收集到的水分是当地降雨量的11~221倍[62],土壤湿润锋增加1.2~4.5倍,土壤水分含量提高10%~140%[63].不同植被覆盖类型下热带、温带以及干旱半干旱区生态系统中平均树干径流量占同期降雨量的3.5%、11.3%和19.0%[64].这一过程可能是荒漠耐旱灌木适应干旱瘠薄立地条件重要生态水文机理[1].在沙坡头人工固沙植被区,柠条和油蒿的树干径流占降雨量的8.99%和2.82%[65],但也有研究认为这有可能过高估计树干径流对灌丛根际区的补给作用[66].主要原因有:一是沙坡头地区以小降雨事件为主,加之较高的潜在蒸散量,不利于树干径流形成;二是由于植被区灌木和半灌木茎干直径较小,且柠条枝条一般为多枝结构,油蒿叶表面覆盖一层不透水蜡质,这种特殊植物冠层结构不利于树干径流形成[18].虽然这一比例较小,但在干旱地区,即使是少量的水分补充对植物的生长也具有重要意义. ...
Rainfall partitioning by vegetation under Mediterranean conditions:a review of studies in Europe
1
2007
... 树干径流是干旱半干旱区植被在降水再分配过程中不可忽视的组成部分[54-55].其在形成过程中能够改变降水的空间分布格局,影响水分和养分循环的速度和方向,亦对植物生存和生长起着至关重要作用[56].目前,国内外学者对干旱荒漠地区灌木的自然集水特征及其对植物根部土壤水分和营养元素补给进行研究[57-58],通过实验观测和模型方法确定不同灌木树干径流产生条件、影响因素、树干径流量与雨水特征关系等[55].这些研究表明荒漠灌木能够通过树干径流收集5%~10%(有时可达到40%)的降水并直接运送到植物根部,之后通过根道系统和土壤大孔隙下渗,将水分贮存在较深的土层中供植物在干旱缺水时利用[59],同时缓解了根际区降水入渗的浅层化问题[60-61].据估算通过灌木树干径流的聚集作用,根部土壤收集到的水分是当地降雨量的11~221倍[62],土壤湿润锋增加1.2~4.5倍,土壤水分含量提高10%~140%[63].不同植被覆盖类型下热带、温带以及干旱半干旱区生态系统中平均树干径流量占同期降雨量的3.5%、11.3%和19.0%[64].这一过程可能是荒漠耐旱灌木适应干旱瘠薄立地条件重要生态水文机理[1].在沙坡头人工固沙植被区,柠条和油蒿的树干径流占降雨量的8.99%和2.82%[65],但也有研究认为这有可能过高估计树干径流对灌丛根际区的补给作用[66].主要原因有:一是沙坡头地区以小降雨事件为主,加之较高的潜在蒸散量,不利于树干径流形成;二是由于植被区灌木和半灌木茎干直径较小,且柠条枝条一般为多枝结构,油蒿叶表面覆盖一层不透水蜡质,这种特殊植物冠层结构不利于树干径流形成[18].虽然这一比例较小,但在干旱地区,即使是少量的水分补充对植物的生长也具有重要意义. ...
荒漠灌丛树干茎流及其入渗、再分配特征
2
2010
... 树干径流是干旱半干旱区植被在降水再分配过程中不可忽视的组成部分[54-55].其在形成过程中能够改变降水的空间分布格局,影响水分和养分循环的速度和方向,亦对植物生存和生长起着至关重要作用[56].目前,国内外学者对干旱荒漠地区灌木的自然集水特征及其对植物根部土壤水分和营养元素补给进行研究[57-58],通过实验观测和模型方法确定不同灌木树干径流产生条件、影响因素、树干径流量与雨水特征关系等[55].这些研究表明荒漠灌木能够通过树干径流收集5%~10%(有时可达到40%)的降水并直接运送到植物根部,之后通过根道系统和土壤大孔隙下渗,将水分贮存在较深的土层中供植物在干旱缺水时利用[59],同时缓解了根际区降水入渗的浅层化问题[60-61].据估算通过灌木树干径流的聚集作用,根部土壤收集到的水分是当地降雨量的11~221倍[62],土壤湿润锋增加1.2~4.5倍,土壤水分含量提高10%~140%[63].不同植被覆盖类型下热带、温带以及干旱半干旱区生态系统中平均树干径流量占同期降雨量的3.5%、11.3%和19.0%[64].这一过程可能是荒漠耐旱灌木适应干旱瘠薄立地条件重要生态水文机理[1].在沙坡头人工固沙植被区,柠条和油蒿的树干径流占降雨量的8.99%和2.82%[65],但也有研究认为这有可能过高估计树干径流对灌丛根际区的补给作用[66].主要原因有:一是沙坡头地区以小降雨事件为主,加之较高的潜在蒸散量,不利于树干径流形成;二是由于植被区灌木和半灌木茎干直径较小,且柠条枝条一般为多枝结构,油蒿叶表面覆盖一层不透水蜡质,这种特殊植物冠层结构不利于树干径流形成[18].虽然这一比例较小,但在干旱地区,即使是少量的水分补充对植物的生长也具有重要意义. ...
... 在荒漠人工植被生态系统中,水分是限制植物生长的主要环境因子,不同植物类群对水分适应的多样性是维持生态系统平衡和可持续性的关键因素[1].对于固沙灌木和半灌木,不同生物学特性决定其维持生存的生态水文学机理不同.如柠条叶与嫩茎上丰富的软毛,为叶片吸收凝结水提供了有利条件;而油蒿根系主要分布在浅层土壤(0~20 cm),对树干径流的利用比例可能高于柠条[55,65].同时,植物水分利用对策也可能随着降水梯度的变化而改变,如东非萨瓦纳生境中特别干旱时段,水力提升现象和树干径流基本消失,合欢(Acacia tortilis)主要通过叶片吸水来适应干旱胁迫[87];当降水量为1.5~5.8 mm,许多植物如三齿团香木(Larrea tridentate)、美洲焦油菊(Flourensia cernua)和腺牧豆树(Prosopis glandulosa)产生树干径流,通过根系优先流来补充土壤水分[88];干季末期大降雨事件(50 mm)之后,萨瓦纳8种主要植物的根系传导水分状况表明:水分在水势梯度驱动下,先从浅层土壤进入浅层根,之后进入主根,最后再由主根进入深层较干燥土壤中,完成水力再分配[89].这说明植物水分利用方式或者对策会随着降雨量的变化而变化. ...
沙漠人工植被区的蒸发蒸腾
2
2005
... 树干径流是干旱半干旱区植被在降水再分配过程中不可忽视的组成部分[54-55].其在形成过程中能够改变降水的空间分布格局,影响水分和养分循环的速度和方向,亦对植物生存和生长起着至关重要作用[56].目前,国内外学者对干旱荒漠地区灌木的自然集水特征及其对植物根部土壤水分和营养元素补给进行研究[57-58],通过实验观测和模型方法确定不同灌木树干径流产生条件、影响因素、树干径流量与雨水特征关系等[55].这些研究表明荒漠灌木能够通过树干径流收集5%~10%(有时可达到40%)的降水并直接运送到植物根部,之后通过根道系统和土壤大孔隙下渗,将水分贮存在较深的土层中供植物在干旱缺水时利用[59],同时缓解了根际区降水入渗的浅层化问题[60-61].据估算通过灌木树干径流的聚集作用,根部土壤收集到的水分是当地降雨量的11~221倍[62],土壤湿润锋增加1.2~4.5倍,土壤水分含量提高10%~140%[63].不同植被覆盖类型下热带、温带以及干旱半干旱区生态系统中平均树干径流量占同期降雨量的3.5%、11.3%和19.0%[64].这一过程可能是荒漠耐旱灌木适应干旱瘠薄立地条件重要生态水文机理[1].在沙坡头人工固沙植被区,柠条和油蒿的树干径流占降雨量的8.99%和2.82%[65],但也有研究认为这有可能过高估计树干径流对灌丛根际区的补给作用[66].主要原因有:一是沙坡头地区以小降雨事件为主,加之较高的潜在蒸散量,不利于树干径流形成;二是由于植被区灌木和半灌木茎干直径较小,且柠条枝条一般为多枝结构,油蒿叶表面覆盖一层不透水蜡质,这种特殊植物冠层结构不利于树干径流形成[18].虽然这一比例较小,但在干旱地区,即使是少量的水分补充对植物的生长也具有重要意义. ...
... 水量平衡动态变化直接关系到植被生长与存活,是植被-土壤系统稳定和健康的核心[90].水量平衡各组分构成在时空尺度上的变异直接影响水量平衡.前期对固沙植被水量平衡研究发现,植物蒸腾量和土壤蒸发量均随着年降水量减少而减少;但是随着降水减少,蒸腾量占蒸散量比例依次减少,而蒸发量占蒸散量比例则依次增大[66].这说明干旱胁迫不仅降低了植物的蒸腾量和蒸散量,而且增加降水的蒸发损失,可供植物利用的水分大幅减少,使得水量平衡各组分发生变化.固沙区生物土壤结皮的形成同样对降水再分配产生影响.生物土壤结皮能够拦截大部分降水[91],不利于高等植物吸收利用.生物土壤结皮对蒸发的影响具有降水依赖性,在大降水条件下(>5 mm)促进土壤蒸发,而在小降水后则抑制蒸发[92].水量平衡各组分对水分来源(主要是降水)依赖性的差异影响到其本身.对水量平衡中各组分的区分及量化是了解其时空异质性的关键所在. ...
Foliar water uptake:processes,pathways,and integration into plant water budgets
1
2019
... 叶片吸水是指植物不通过根系,而直接通过叶片来吸取水分的能力,同时也是植物利用隐匿降水(如雾或凝结水)的重要方式[67],特别是在小降水事件占主导的干旱荒漠人工植被区[1].关于植物叶片吸水的研究可以追溯到20世纪50年代,Stone[68]在北美西部地区首次发现黄松(Pinus ponderosa)植物通过叶片摄取水分,但是由于局限于特定环境的特定对象,此现象并未得到足够重视[69].直到近些年来越来越多的研究发现叶片吸水是植物最大限度地利用有限的水资源的重要途径[70].迄今为止,国内外发现有70余种植物通过叶片吸收水分[71].主要分布在北方阔叶针叶林[72-73]、热带雨林[74]、稀疏草原[75]、温带森林[76]和温带草原[77]等.国际上对云雾林的研究也证实了这一现象,雾水可直接通过叶片角质层扩散至根茎,占叶片总含水量的42%[71-72].对生活在地中海地区的常绿灌木迷迭香(Rosmarinus officinalis)和西班牙薰衣草(Lavandula stoechas)的研究结果表明:干旱时期有一半时间出现凝结水吸收使叶片相对含水量提高了72%,水势提高了0.5 MPa,水分亏缺状况得到缓解[78].国内关于植物叶片吸水的研究主要在西双版纳雨林生态系统的附生植物与非附生植物叶片[79],及干旱荒漠地区如准噶尔盆地[80]、民勤[81]、毛乌素沙地[82-83]和腾格里沙漠人工植被区[26]等植物均具有叶片吸水现象,且荒漠植物地上(叶片)和地下(根系)部分耗水及吸水策略存在权衡[80]. ...
Dew as an ecological factor:II.the effect of artificial dew on the survival of Pinus ponderosa and associated species
1
1957
... 叶片吸水是指植物不通过根系,而直接通过叶片来吸取水分的能力,同时也是植物利用隐匿降水(如雾或凝结水)的重要方式[67],特别是在小降水事件占主导的干旱荒漠人工植被区[1].关于植物叶片吸水的研究可以追溯到20世纪50年代,Stone[68]在北美西部地区首次发现黄松(Pinus ponderosa)植物通过叶片摄取水分,但是由于局限于特定环境的特定对象,此现象并未得到足够重视[69].直到近些年来越来越多的研究发现叶片吸水是植物最大限度地利用有限的水资源的重要途径[70].迄今为止,国内外发现有70余种植物通过叶片吸收水分[71].主要分布在北方阔叶针叶林[72-73]、热带雨林[74]、稀疏草原[75]、温带森林[76]和温带草原[77]等.国际上对云雾林的研究也证实了这一现象,雾水可直接通过叶片角质层扩散至根茎,占叶片总含水量的42%[71-72].对生活在地中海地区的常绿灌木迷迭香(Rosmarinus officinalis)和西班牙薰衣草(Lavandula stoechas)的研究结果表明:干旱时期有一半时间出现凝结水吸收使叶片相对含水量提高了72%,水势提高了0.5 MPa,水分亏缺状况得到缓解[78].国内关于植物叶片吸水的研究主要在西双版纳雨林生态系统的附生植物与非附生植物叶片[79],及干旱荒漠地区如准噶尔盆地[80]、民勤[81]、毛乌素沙地[82-83]和腾格里沙漠人工植被区[26]等植物均具有叶片吸水现象,且荒漠植物地上(叶片)和地下(根系)部分耗水及吸水策略存在权衡[80]. ...
Polystichum munitum (Dryopteridaceae) varies geographically in its capacity to absorb fog water by foliar uptake within the redwood forest ecosystem
1
2010
... 叶片吸水是指植物不通过根系,而直接通过叶片来吸取水分的能力,同时也是植物利用隐匿降水(如雾或凝结水)的重要方式[67],特别是在小降水事件占主导的干旱荒漠人工植被区[1].关于植物叶片吸水的研究可以追溯到20世纪50年代,Stone[68]在北美西部地区首次发现黄松(Pinus ponderosa)植物通过叶片摄取水分,但是由于局限于特定环境的特定对象,此现象并未得到足够重视[69].直到近些年来越来越多的研究发现叶片吸水是植物最大限度地利用有限的水资源的重要途径[70].迄今为止,国内外发现有70余种植物通过叶片吸收水分[71].主要分布在北方阔叶针叶林[72-73]、热带雨林[74]、稀疏草原[75]、温带森林[76]和温带草原[77]等.国际上对云雾林的研究也证实了这一现象,雾水可直接通过叶片角质层扩散至根茎,占叶片总含水量的42%[71-72].对生活在地中海地区的常绿灌木迷迭香(Rosmarinus officinalis)和西班牙薰衣草(Lavandula stoechas)的研究结果表明:干旱时期有一半时间出现凝结水吸收使叶片相对含水量提高了72%,水势提高了0.5 MPa,水分亏缺状况得到缓解[78].国内关于植物叶片吸水的研究主要在西双版纳雨林生态系统的附生植物与非附生植物叶片[79],及干旱荒漠地区如准噶尔盆地[80]、民勤[81]、毛乌素沙地[82-83]和腾格里沙漠人工植被区[26]等植物均具有叶片吸水现象,且荒漠植物地上(叶片)和地下(根系)部分耗水及吸水策略存在权衡[80]. ...
The contribution of fog to the water relations of Sequoia sempervirens (D.Don):foliar uptake and prevention of dehydration
1
2004
... 叶片吸水是指植物不通过根系,而直接通过叶片来吸取水分的能力,同时也是植物利用隐匿降水(如雾或凝结水)的重要方式[67],特别是在小降水事件占主导的干旱荒漠人工植被区[1].关于植物叶片吸水的研究可以追溯到20世纪50年代,Stone[68]在北美西部地区首次发现黄松(Pinus ponderosa)植物通过叶片摄取水分,但是由于局限于特定环境的特定对象,此现象并未得到足够重视[69].直到近些年来越来越多的研究发现叶片吸水是植物最大限度地利用有限的水资源的重要途径[70].迄今为止,国内外发现有70余种植物通过叶片吸收水分[71].主要分布在北方阔叶针叶林[72-73]、热带雨林[74]、稀疏草原[75]、温带森林[76]和温带草原[77]等.国际上对云雾林的研究也证实了这一现象,雾水可直接通过叶片角质层扩散至根茎,占叶片总含水量的42%[71-72].对生活在地中海地区的常绿灌木迷迭香(Rosmarinus officinalis)和西班牙薰衣草(Lavandula stoechas)的研究结果表明:干旱时期有一半时间出现凝结水吸收使叶片相对含水量提高了72%,水势提高了0.5 MPa,水分亏缺状况得到缓解[78].国内关于植物叶片吸水的研究主要在西双版纳雨林生态系统的附生植物与非附生植物叶片[79],及干旱荒漠地区如准噶尔盆地[80]、民勤[81]、毛乌素沙地[82-83]和腾格里沙漠人工植被区[26]等植物均具有叶片吸水现象,且荒漠植物地上(叶片)和地下(根系)部分耗水及吸水策略存在权衡[80]. ...
The incidence and implications of clouds for cloud forest plant water relations
2
2013
... 叶片吸水是指植物不通过根系,而直接通过叶片来吸取水分的能力,同时也是植物利用隐匿降水(如雾或凝结水)的重要方式[67],特别是在小降水事件占主导的干旱荒漠人工植被区[1].关于植物叶片吸水的研究可以追溯到20世纪50年代,Stone[68]在北美西部地区首次发现黄松(Pinus ponderosa)植物通过叶片摄取水分,但是由于局限于特定环境的特定对象,此现象并未得到足够重视[69].直到近些年来越来越多的研究发现叶片吸水是植物最大限度地利用有限的水资源的重要途径[70].迄今为止,国内外发现有70余种植物通过叶片吸收水分[71].主要分布在北方阔叶针叶林[72-73]、热带雨林[74]、稀疏草原[75]、温带森林[76]和温带草原[77]等.国际上对云雾林的研究也证实了这一现象,雾水可直接通过叶片角质层扩散至根茎,占叶片总含水量的42%[71-72].对生活在地中海地区的常绿灌木迷迭香(Rosmarinus officinalis)和西班牙薰衣草(Lavandula stoechas)的研究结果表明:干旱时期有一半时间出现凝结水吸收使叶片相对含水量提高了72%,水势提高了0.5 MPa,水分亏缺状况得到缓解[78].国内关于植物叶片吸水的研究主要在西双版纳雨林生态系统的附生植物与非附生植物叶片[79],及干旱荒漠地区如准噶尔盆地[80]、民勤[81]、毛乌素沙地[82-83]和腾格里沙漠人工植被区[26]等植物均具有叶片吸水现象,且荒漠植物地上(叶片)和地下(根系)部分耗水及吸水策略存在权衡[80]. ...
... [71-72].对生活在地中海地区的常绿灌木迷迭香(Rosmarinus officinalis)和西班牙薰衣草(Lavandula stoechas)的研究结果表明:干旱时期有一半时间出现凝结水吸收使叶片相对含水量提高了72%,水势提高了0.5 MPa,水分亏缺状况得到缓解[78].国内关于植物叶片吸水的研究主要在西双版纳雨林生态系统的附生植物与非附生植物叶片[79],及干旱荒漠地区如准噶尔盆地[80]、民勤[81]、毛乌素沙地[82-83]和腾格里沙漠人工植被区[26]等植物均具有叶片吸水现象,且荒漠植物地上(叶片)和地下(根系)部分耗水及吸水策略存在权衡[80]. ...
Cloud forest trees with higher foliar water uptake capacity and anisohydric behavior are more vulnerable to drought and climate change
2
2016
... 叶片吸水是指植物不通过根系,而直接通过叶片来吸取水分的能力,同时也是植物利用隐匿降水(如雾或凝结水)的重要方式[67],特别是在小降水事件占主导的干旱荒漠人工植被区[1].关于植物叶片吸水的研究可以追溯到20世纪50年代,Stone[68]在北美西部地区首次发现黄松(Pinus ponderosa)植物通过叶片摄取水分,但是由于局限于特定环境的特定对象,此现象并未得到足够重视[69].直到近些年来越来越多的研究发现叶片吸水是植物最大限度地利用有限的水资源的重要途径[70].迄今为止,国内外发现有70余种植物通过叶片吸收水分[71].主要分布在北方阔叶针叶林[72-73]、热带雨林[74]、稀疏草原[75]、温带森林[76]和温带草原[77]等.国际上对云雾林的研究也证实了这一现象,雾水可直接通过叶片角质层扩散至根茎,占叶片总含水量的42%[71-72].对生活在地中海地区的常绿灌木迷迭香(Rosmarinus officinalis)和西班牙薰衣草(Lavandula stoechas)的研究结果表明:干旱时期有一半时间出现凝结水吸收使叶片相对含水量提高了72%,水势提高了0.5 MPa,水分亏缺状况得到缓解[78].国内关于植物叶片吸水的研究主要在西双版纳雨林生态系统的附生植物与非附生植物叶片[79],及干旱荒漠地区如准噶尔盆地[80]、民勤[81]、毛乌素沙地[82-83]和腾格里沙漠人工植被区[26]等植物均具有叶片吸水现象,且荒漠植物地上(叶片)和地下(根系)部分耗水及吸水策略存在权衡[80]. ...
... -72].对生活在地中海地区的常绿灌木迷迭香(Rosmarinus officinalis)和西班牙薰衣草(Lavandula stoechas)的研究结果表明:干旱时期有一半时间出现凝结水吸收使叶片相对含水量提高了72%,水势提高了0.5 MPa,水分亏缺状况得到缓解[78].国内关于植物叶片吸水的研究主要在西双版纳雨林生态系统的附生植物与非附生植物叶片[79],及干旱荒漠地区如准噶尔盆地[80]、民勤[81]、毛乌素沙地[82-83]和腾格里沙漠人工植被区[26]等植物均具有叶片吸水现象,且荒漠植物地上(叶片)和地下(根系)部分耗水及吸水策略存在权衡[80]. ...
Evidence of foliar water uptake in a conifer species
1
2021
... 叶片吸水是指植物不通过根系,而直接通过叶片来吸取水分的能力,同时也是植物利用隐匿降水(如雾或凝结水)的重要方式[67],特别是在小降水事件占主导的干旱荒漠人工植被区[1].关于植物叶片吸水的研究可以追溯到20世纪50年代,Stone[68]在北美西部地区首次发现黄松(Pinus ponderosa)植物通过叶片摄取水分,但是由于局限于特定环境的特定对象,此现象并未得到足够重视[69].直到近些年来越来越多的研究发现叶片吸水是植物最大限度地利用有限的水资源的重要途径[70].迄今为止,国内外发现有70余种植物通过叶片吸收水分[71].主要分布在北方阔叶针叶林[72-73]、热带雨林[74]、稀疏草原[75]、温带森林[76]和温带草原[77]等.国际上对云雾林的研究也证实了这一现象,雾水可直接通过叶片角质层扩散至根茎,占叶片总含水量的42%[71-72].对生活在地中海地区的常绿灌木迷迭香(Rosmarinus officinalis)和西班牙薰衣草(Lavandula stoechas)的研究结果表明:干旱时期有一半时间出现凝结水吸收使叶片相对含水量提高了72%,水势提高了0.5 MPa,水分亏缺状况得到缓解[78].国内关于植物叶片吸水的研究主要在西双版纳雨林生态系统的附生植物与非附生植物叶片[79],及干旱荒漠地区如准噶尔盆地[80]、民勤[81]、毛乌素沙地[82-83]和腾格里沙漠人工植被区[26]等植物均具有叶片吸水现象,且荒漠植物地上(叶片)和地下(根系)部分耗水及吸水策略存在权衡[80]. ...
Foliar water uptake in Amazonian trees:evidence and consequences
1
2019
... 叶片吸水是指植物不通过根系,而直接通过叶片来吸取水分的能力,同时也是植物利用隐匿降水(如雾或凝结水)的重要方式[67],特别是在小降水事件占主导的干旱荒漠人工植被区[1].关于植物叶片吸水的研究可以追溯到20世纪50年代,Stone[68]在北美西部地区首次发现黄松(Pinus ponderosa)植物通过叶片摄取水分,但是由于局限于特定环境的特定对象,此现象并未得到足够重视[69].直到近些年来越来越多的研究发现叶片吸水是植物最大限度地利用有限的水资源的重要途径[70].迄今为止,国内外发现有70余种植物通过叶片吸收水分[71].主要分布在北方阔叶针叶林[72-73]、热带雨林[74]、稀疏草原[75]、温带森林[76]和温带草原[77]等.国际上对云雾林的研究也证实了这一现象,雾水可直接通过叶片角质层扩散至根茎,占叶片总含水量的42%[71-72].对生活在地中海地区的常绿灌木迷迭香(Rosmarinus officinalis)和西班牙薰衣草(Lavandula stoechas)的研究结果表明:干旱时期有一半时间出现凝结水吸收使叶片相对含水量提高了72%,水势提高了0.5 MPa,水分亏缺状况得到缓解[78].国内关于植物叶片吸水的研究主要在西双版纳雨林生态系统的附生植物与非附生植物叶片[79],及干旱荒漠地区如准噶尔盆地[80]、民勤[81]、毛乌素沙地[82-83]和腾格里沙漠人工植被区[26]等植物均具有叶片吸水现象,且荒漠植物地上(叶片)和地下(根系)部分耗水及吸水策略存在权衡[80]. ...
Foliar water uptake in arid ecosystems:seasonal variability and ecophysiological consequences
2
2020
... 叶片吸水是指植物不通过根系,而直接通过叶片来吸取水分的能力,同时也是植物利用隐匿降水(如雾或凝结水)的重要方式[67],特别是在小降水事件占主导的干旱荒漠人工植被区[1].关于植物叶片吸水的研究可以追溯到20世纪50年代,Stone[68]在北美西部地区首次发现黄松(Pinus ponderosa)植物通过叶片摄取水分,但是由于局限于特定环境的特定对象,此现象并未得到足够重视[69].直到近些年来越来越多的研究发现叶片吸水是植物最大限度地利用有限的水资源的重要途径[70].迄今为止,国内外发现有70余种植物通过叶片吸收水分[71].主要分布在北方阔叶针叶林[72-73]、热带雨林[74]、稀疏草原[75]、温带森林[76]和温带草原[77]等.国际上对云雾林的研究也证实了这一现象,雾水可直接通过叶片角质层扩散至根茎,占叶片总含水量的42%[71-72].对生活在地中海地区的常绿灌木迷迭香(Rosmarinus officinalis)和西班牙薰衣草(Lavandula stoechas)的研究结果表明:干旱时期有一半时间出现凝结水吸收使叶片相对含水量提高了72%,水势提高了0.5 MPa,水分亏缺状况得到缓解[78].国内关于植物叶片吸水的研究主要在西双版纳雨林生态系统的附生植物与非附生植物叶片[79],及干旱荒漠地区如准噶尔盆地[80]、民勤[81]、毛乌素沙地[82-83]和腾格里沙漠人工植被区[26]等植物均具有叶片吸水现象,且荒漠植物地上(叶片)和地下(根系)部分耗水及吸水策略存在权衡[80]. ...
... 叶片吸水对干旱沙区植物具有重要生理和生态作用.如光合性能改善,植物组织含水量增加,甚至延长叶片寿命、增加结实量、促进植物生长等[75,84].在沙坡头人工固沙植被区,小于5 mm的降水占到总降水频次的63.9%[85],凝结水量也占全年降水量的10%~20%[86],通过同位素示踪技术,在沙坡头人工植被区证明了典型固沙灌木也存在叶片吸水的现象,量化了柠条和油蒿单位面积吸水量分别为3.03~11.02、5.86~24.76 g,单位面积植物群落水分吸收能力分别为300~600、600~800 g,相当于0.3~0.8 mm的同期降雨量.[26].这表明叶片吸水在一定程度上能够补充植物体内的水分亏缺,尤其在干旱时期对维持植物的生理活性具有重要作用. ...
Influence of drought on foliar water uptake capacity of temperate tree species
1
2019
... 叶片吸水是指植物不通过根系,而直接通过叶片来吸取水分的能力,同时也是植物利用隐匿降水(如雾或凝结水)的重要方式[67],特别是在小降水事件占主导的干旱荒漠人工植被区[1].关于植物叶片吸水的研究可以追溯到20世纪50年代,Stone[68]在北美西部地区首次发现黄松(Pinus ponderosa)植物通过叶片摄取水分,但是由于局限于特定环境的特定对象,此现象并未得到足够重视[69].直到近些年来越来越多的研究发现叶片吸水是植物最大限度地利用有限的水资源的重要途径[70].迄今为止,国内外发现有70余种植物通过叶片吸收水分[71].主要分布在北方阔叶针叶林[72-73]、热带雨林[74]、稀疏草原[75]、温带森林[76]和温带草原[77]等.国际上对云雾林的研究也证实了这一现象,雾水可直接通过叶片角质层扩散至根茎,占叶片总含水量的42%[71-72].对生活在地中海地区的常绿灌木迷迭香(Rosmarinus officinalis)和西班牙薰衣草(Lavandula stoechas)的研究结果表明:干旱时期有一半时间出现凝结水吸收使叶片相对含水量提高了72%,水势提高了0.5 MPa,水分亏缺状况得到缓解[78].国内关于植物叶片吸水的研究主要在西双版纳雨林生态系统的附生植物与非附生植物叶片[79],及干旱荒漠地区如准噶尔盆地[80]、民勤[81]、毛乌素沙地[82-83]和腾格里沙漠人工植被区[26]等植物均具有叶片吸水现象,且荒漠植物地上(叶片)和地下(根系)部分耗水及吸水策略存在权衡[80]. ...
Foliar uptake of dew in the sandy ecosystem of the Mongolia Plateau:a life-sustaining and carbon accumulation strategy shared differently by C3 and C4 grasses
1
2020
... 叶片吸水是指植物不通过根系,而直接通过叶片来吸取水分的能力,同时也是植物利用隐匿降水(如雾或凝结水)的重要方式[67],特别是在小降水事件占主导的干旱荒漠人工植被区[1].关于植物叶片吸水的研究可以追溯到20世纪50年代,Stone[68]在北美西部地区首次发现黄松(Pinus ponderosa)植物通过叶片摄取水分,但是由于局限于特定环境的特定对象,此现象并未得到足够重视[69].直到近些年来越来越多的研究发现叶片吸水是植物最大限度地利用有限的水资源的重要途径[70].迄今为止,国内外发现有70余种植物通过叶片吸收水分[71].主要分布在北方阔叶针叶林[72-73]、热带雨林[74]、稀疏草原[75]、温带森林[76]和温带草原[77]等.国际上对云雾林的研究也证实了这一现象,雾水可直接通过叶片角质层扩散至根茎,占叶片总含水量的42%[71-72].对生活在地中海地区的常绿灌木迷迭香(Rosmarinus officinalis)和西班牙薰衣草(Lavandula stoechas)的研究结果表明:干旱时期有一半时间出现凝结水吸收使叶片相对含水量提高了72%,水势提高了0.5 MPa,水分亏缺状况得到缓解[78].国内关于植物叶片吸水的研究主要在西双版纳雨林生态系统的附生植物与非附生植物叶片[79],及干旱荒漠地区如准噶尔盆地[80]、民勤[81]、毛乌素沙地[82-83]和腾格里沙漠人工植被区[26]等植物均具有叶片吸水现象,且荒漠植物地上(叶片)和地下(根系)部分耗水及吸水策略存在权衡[80]. ...
Diurnal variations of photosynthesis and dew absorption by leaves in two evergreen shrubs growing in Mediterranean field conditions
1
1999
... 叶片吸水是指植物不通过根系,而直接通过叶片来吸取水分的能力,同时也是植物利用隐匿降水(如雾或凝结水)的重要方式[67],特别是在小降水事件占主导的干旱荒漠人工植被区[1].关于植物叶片吸水的研究可以追溯到20世纪50年代,Stone[68]在北美西部地区首次发现黄松(Pinus ponderosa)植物通过叶片摄取水分,但是由于局限于特定环境的特定对象,此现象并未得到足够重视[69].直到近些年来越来越多的研究发现叶片吸水是植物最大限度地利用有限的水资源的重要途径[70].迄今为止,国内外发现有70余种植物通过叶片吸收水分[71].主要分布在北方阔叶针叶林[72-73]、热带雨林[74]、稀疏草原[75]、温带森林[76]和温带草原[77]等.国际上对云雾林的研究也证实了这一现象,雾水可直接通过叶片角质层扩散至根茎,占叶片总含水量的42%[71-72].对生活在地中海地区的常绿灌木迷迭香(Rosmarinus officinalis)和西班牙薰衣草(Lavandula stoechas)的研究结果表明:干旱时期有一半时间出现凝结水吸收使叶片相对含水量提高了72%,水势提高了0.5 MPa,水分亏缺状况得到缓解[78].国内关于植物叶片吸水的研究主要在西双版纳雨林生态系统的附生植物与非附生植物叶片[79],及干旱荒漠地区如准噶尔盆地[80]、民勤[81]、毛乌素沙地[82-83]和腾格里沙漠人工植被区[26]等植物均具有叶片吸水现象,且荒漠植物地上(叶片)和地下(根系)部分耗水及吸水策略存在权衡[80]. ...
西双版纳地区附生与非附生植物叶片对雾水的吸收
1
2006
... 叶片吸水是指植物不通过根系,而直接通过叶片来吸取水分的能力,同时也是植物利用隐匿降水(如雾或凝结水)的重要方式[67],特别是在小降水事件占主导的干旱荒漠人工植被区[1].关于植物叶片吸水的研究可以追溯到20世纪50年代,Stone[68]在北美西部地区首次发现黄松(Pinus ponderosa)植物通过叶片摄取水分,但是由于局限于特定环境的特定对象,此现象并未得到足够重视[69].直到近些年来越来越多的研究发现叶片吸水是植物最大限度地利用有限的水资源的重要途径[70].迄今为止,国内外发现有70余种植物通过叶片吸收水分[71].主要分布在北方阔叶针叶林[72-73]、热带雨林[74]、稀疏草原[75]、温带森林[76]和温带草原[77]等.国际上对云雾林的研究也证实了这一现象,雾水可直接通过叶片角质层扩散至根茎,占叶片总含水量的42%[71-72].对生活在地中海地区的常绿灌木迷迭香(Rosmarinus officinalis)和西班牙薰衣草(Lavandula stoechas)的研究结果表明:干旱时期有一半时间出现凝结水吸收使叶片相对含水量提高了72%,水势提高了0.5 MPa,水分亏缺状况得到缓解[78].国内关于植物叶片吸水的研究主要在西双版纳雨林生态系统的附生植物与非附生植物叶片[79],及干旱荒漠地区如准噶尔盆地[80]、民勤[81]、毛乌素沙地[82-83]和腾格里沙漠人工植被区[26]等植物均具有叶片吸水现象,且荒漠植物地上(叶片)和地下(根系)部分耗水及吸水策略存在权衡[80]. ...
准噶尔盆地荒漠植物的叶片水分吸收策略
2
2011
... 叶片吸水是指植物不通过根系,而直接通过叶片来吸取水分的能力,同时也是植物利用隐匿降水(如雾或凝结水)的重要方式[67],特别是在小降水事件占主导的干旱荒漠人工植被区[1].关于植物叶片吸水的研究可以追溯到20世纪50年代,Stone[68]在北美西部地区首次发现黄松(Pinus ponderosa)植物通过叶片摄取水分,但是由于局限于特定环境的特定对象,此现象并未得到足够重视[69].直到近些年来越来越多的研究发现叶片吸水是植物最大限度地利用有限的水资源的重要途径[70].迄今为止,国内外发现有70余种植物通过叶片吸收水分[71].主要分布在北方阔叶针叶林[72-73]、热带雨林[74]、稀疏草原[75]、温带森林[76]和温带草原[77]等.国际上对云雾林的研究也证实了这一现象,雾水可直接通过叶片角质层扩散至根茎,占叶片总含水量的42%[71-72].对生活在地中海地区的常绿灌木迷迭香(Rosmarinus officinalis)和西班牙薰衣草(Lavandula stoechas)的研究结果表明:干旱时期有一半时间出现凝结水吸收使叶片相对含水量提高了72%,水势提高了0.5 MPa,水分亏缺状况得到缓解[78].国内关于植物叶片吸水的研究主要在西双版纳雨林生态系统的附生植物与非附生植物叶片[79],及干旱荒漠地区如准噶尔盆地[80]、民勤[81]、毛乌素沙地[82-83]和腾格里沙漠人工植被区[26]等植物均具有叶片吸水现象,且荒漠植物地上(叶片)和地下(根系)部分耗水及吸水策略存在权衡[80]. ...
... [80]. ...
民勤荒漠植物叶片水分吸收性状研究
1
2020
... 叶片吸水是指植物不通过根系,而直接通过叶片来吸取水分的能力,同时也是植物利用隐匿降水(如雾或凝结水)的重要方式[67],特别是在小降水事件占主导的干旱荒漠人工植被区[1].关于植物叶片吸水的研究可以追溯到20世纪50年代,Stone[68]在北美西部地区首次发现黄松(Pinus ponderosa)植物通过叶片摄取水分,但是由于局限于特定环境的特定对象,此现象并未得到足够重视[69].直到近些年来越来越多的研究发现叶片吸水是植物最大限度地利用有限的水资源的重要途径[70].迄今为止,国内外发现有70余种植物通过叶片吸收水分[71].主要分布在北方阔叶针叶林[72-73]、热带雨林[74]、稀疏草原[75]、温带森林[76]和温带草原[77]等.国际上对云雾林的研究也证实了这一现象,雾水可直接通过叶片角质层扩散至根茎,占叶片总含水量的42%[71-72].对生活在地中海地区的常绿灌木迷迭香(Rosmarinus officinalis)和西班牙薰衣草(Lavandula stoechas)的研究结果表明:干旱时期有一半时间出现凝结水吸收使叶片相对含水量提高了72%,水势提高了0.5 MPa,水分亏缺状况得到缓解[78].国内关于植物叶片吸水的研究主要在西双版纳雨林生态系统的附生植物与非附生植物叶片[79],及干旱荒漠地区如准噶尔盆地[80]、民勤[81]、毛乌素沙地[82-83]和腾格里沙漠人工植被区[26]等植物均具有叶片吸水现象,且荒漠植物地上(叶片)和地下(根系)部分耗水及吸水策略存在权衡[80]. ...
毛乌素沙地两种典型灌木叶片凝结水吸收能力及吸水途径
1
2021
... 叶片吸水是指植物不通过根系,而直接通过叶片来吸取水分的能力,同时也是植物利用隐匿降水(如雾或凝结水)的重要方式[67],特别是在小降水事件占主导的干旱荒漠人工植被区[1].关于植物叶片吸水的研究可以追溯到20世纪50年代,Stone[68]在北美西部地区首次发现黄松(Pinus ponderosa)植物通过叶片摄取水分,但是由于局限于特定环境的特定对象,此现象并未得到足够重视[69].直到近些年来越来越多的研究发现叶片吸水是植物最大限度地利用有限的水资源的重要途径[70].迄今为止,国内外发现有70余种植物通过叶片吸收水分[71].主要分布在北方阔叶针叶林[72-73]、热带雨林[74]、稀疏草原[75]、温带森林[76]和温带草原[77]等.国际上对云雾林的研究也证实了这一现象,雾水可直接通过叶片角质层扩散至根茎,占叶片总含水量的42%[71-72].对生活在地中海地区的常绿灌木迷迭香(Rosmarinus officinalis)和西班牙薰衣草(Lavandula stoechas)的研究结果表明:干旱时期有一半时间出现凝结水吸收使叶片相对含水量提高了72%,水势提高了0.5 MPa,水分亏缺状况得到缓解[78].国内关于植物叶片吸水的研究主要在西双版纳雨林生态系统的附生植物与非附生植物叶片[79],及干旱荒漠地区如准噶尔盆地[80]、民勤[81]、毛乌素沙地[82-83]和腾格里沙漠人工植被区[26]等植物均具有叶片吸水现象,且荒漠植物地上(叶片)和地下(根系)部分耗水及吸水策略存在权衡[80]. ...
Foliar water uptake of four shrub species in a semi-arid desert
1
2021
... 叶片吸水是指植物不通过根系,而直接通过叶片来吸取水分的能力,同时也是植物利用隐匿降水(如雾或凝结水)的重要方式[67],特别是在小降水事件占主导的干旱荒漠人工植被区[1].关于植物叶片吸水的研究可以追溯到20世纪50年代,Stone[68]在北美西部地区首次发现黄松(Pinus ponderosa)植物通过叶片摄取水分,但是由于局限于特定环境的特定对象,此现象并未得到足够重视[69].直到近些年来越来越多的研究发现叶片吸水是植物最大限度地利用有限的水资源的重要途径[70].迄今为止,国内外发现有70余种植物通过叶片吸收水分[71].主要分布在北方阔叶针叶林[72-73]、热带雨林[74]、稀疏草原[75]、温带森林[76]和温带草原[77]等.国际上对云雾林的研究也证实了这一现象,雾水可直接通过叶片角质层扩散至根茎,占叶片总含水量的42%[71-72].对生活在地中海地区的常绿灌木迷迭香(Rosmarinus officinalis)和西班牙薰衣草(Lavandula stoechas)的研究结果表明:干旱时期有一半时间出现凝结水吸收使叶片相对含水量提高了72%,水势提高了0.5 MPa,水分亏缺状况得到缓解[78].国内关于植物叶片吸水的研究主要在西双版纳雨林生态系统的附生植物与非附生植物叶片[79],及干旱荒漠地区如准噶尔盆地[80]、民勤[81]、毛乌素沙地[82-83]和腾格里沙漠人工植被区[26]等植物均具有叶片吸水现象,且荒漠植物地上(叶片)和地下(根系)部分耗水及吸水策略存在权衡[80]. ...
Foliar water uptake:a common water acquisition strategy for plants of the redwood forest
1
2009
... 叶片吸水对干旱沙区植物具有重要生理和生态作用.如光合性能改善,植物组织含水量增加,甚至延长叶片寿命、增加结实量、促进植物生长等[75,84].在沙坡头人工固沙植被区,小于5 mm的降水占到总降水频次的63.9%[85],凝结水量也占全年降水量的10%~20%[86],通过同位素示踪技术,在沙坡头人工植被区证明了典型固沙灌木也存在叶片吸水的现象,量化了柠条和油蒿单位面积吸水量分别为3.03~11.02、5.86~24.76 g,单位面积植物群落水分吸收能力分别为300~600、600~800 g,相当于0.3~0.8 mm的同期降雨量.[26].这表明叶片吸水在一定程度上能够补充植物体内的水分亏缺,尤其在干旱时期对维持植物的生理活性具有重要作用. ...
吸湿凝结水对荒漠地区生物土壤结皮生态功能的影响综述
4
2022
... 叶片吸水对干旱沙区植物具有重要生理和生态作用.如光合性能改善,植物组织含水量增加,甚至延长叶片寿命、增加结实量、促进植物生长等[75,84].在沙坡头人工固沙植被区,小于5 mm的降水占到总降水频次的63.9%[85],凝结水量也占全年降水量的10%~20%[86],通过同位素示踪技术,在沙坡头人工植被区证明了典型固沙灌木也存在叶片吸水的现象,量化了柠条和油蒿单位面积吸水量分别为3.03~11.02、5.86~24.76 g,单位面积植物群落水分吸收能力分别为300~600、600~800 g,相当于0.3~0.8 mm的同期降雨量.[26].这表明叶片吸水在一定程度上能够补充植物体内的水分亏缺,尤其在干旱时期对维持植物的生理活性具有重要作用. ...
... 固沙灌木对降水的利用受多种因素制约,其中降雨量是关键[1,85].根据沙坡头站实验观测数据,建立不同降水梯度下固沙植物对降水利用的生态水文调节阈值模型(图1),确定固沙植物根系水力再分配、树干径流和叶片吸水这3种水分适应对策状态转移的拐点.当降水量为0~1 mm,不存在树干径流,植物适应干旱的主要对策为叶片吸水;而当降水量增大至P3~P4时,如在1~5 mm时,叶片吸水作用可以被忽略,树干径流起主要作用;当降水量大于5 mm时,降水可通过穿透雨、入渗等对土壤水进行大量补充,根土界面的水力再分配是植物适应干旱的主要对策.降水量逐渐变化过程中,根土界面水力再分配、树干径流和叶片吸水对植物生存都有影响,因此在模型中定义了混合状态,即代表这3种对策同时存在,只是在某一降水区间,某一种对策可能对植物生存起主要作用.这说明在小降雨事件中,植物主要通过叶片吸水和树干径流等方式快速利用有限水分资源,以满足短期的生理需求;而在较大降雨事件中,根系水力再分配则发挥主导作用,将多余的水分储存于土壤中,供植物在干旱时期使用. ...
... 作为干旱区重要水分补给源降水,具有显著的时空异质性和随机性.在荒漠人工植被区,脉冲式降水引起土壤水分和植被格局的动态变化[93].降水为随机的脉冲现象,考虑其对植被的影响,探讨其生态水文过程,已成为国际生态水文学界关注的热点[94-95].腾格里沙漠降水具有明显的脉冲特征,0~5 mm小降水事件占全年降水事件的77.8%,且降水间隔期以0~10 d为主.这种脉冲降水模式对固沙灌木生长和生理过程产生重要影响.固沙灌木对不同强度和频率的降水脉冲响应机制不同[85].0~5 d降水间隔对于油蒿叶面积指数(LAI)、新枝长和蒸腾耗水影响较大(50%左右),其次是5~10 d降水间隔.当降水间隔大于10 d时,其对油蒿影响较为稳定.0~5 d降水间隔对油蒿生理生态特征影响起决定作用的为0~5 mm的小降水事件.柠条LAI和蒸腾耗水对0~5 d降水间隔较为敏感,而对新枝长影响最大则为5~10 d降水间隔.与油蒿相比,0~5 d降水间隔对柠条生理生态特征影响较小,而当降水间隔大于5 d时,其对柠条影响作用要大于油蒿.这说明在干旱人工植被区,间歇性小降水事件更有利于油蒿生长发育. ...
... 固沙灌木地上和地下部分在生长发育过程中存在密切协调关系,是植物适应干旱环境、维持自身稳定的关键[122].干旱区植物根系通常具有深根性和广布性.例如白梭梭和梭梭的垂直根深逾5 m,可至地下水层吸收水分[123].柽柳主根和侧根均极为发达,主根可伸至地下水层,最深10 m以上[124].此外,一些植物如沙拐枣根系多为水平分布,但在地下水较深的地区,其根系也能垂直向下发展以获取水分[125].以上根系的形态和分布使得植物能够有效地利用土壤水分资源.长期干旱时植物通过落叶(如梭梭)、茎干休眠等方式减少地上部分的水分消耗,同时维持根系活力(如沙拐枣根系持续生长寻找水源).植物地上和地下部分在生理功能上也表现为协调关系[126].植物通过冠层截留降水,减少土壤水分蒸发.同时,植物根系能够将截留水分吸收并输送到地上部分,支持植物蒸腾作用和光合作用.固沙灌木冠层截留和树干径流在水分再分配中起着重要作用,能够提高根际区水分供应[55].植物在资源分配上也表现出对干旱环境的适应性,根据环境条件调整地上和地下部分的生物量分配,以优化对光、水和养分等资源的利用[127].干旱条件下植物可能会将更多的资源分配到根系,以增强水分获取能力.同时,植物叶片和细根在碳、氮、磷等元素的分配中也存在协同关系.西北干旱区荒漠植物叶片和细根在碳、氮、磷含量及其计量比上表现出显著的正相关关系[128],这表明植物在完成正常的生命活动时,地上与地下部分的养分分配存在协同作用.深根系植物(如梭梭)主根深入地下水源,地上部分冠层稀疏(减少蒸腾面积),形成高而疏的形态,减少水分竞争.浅根系植物(如沙柳)侧根广泛分布于表层,冠层密集,依赖小降水后对表层水分的快速利用.植物地上和地下部分之间存在信息流和信号传递的协调机制[129].植物可通过木质部和韧皮部传递激素、压力波和电信号等,以协调地上和地下部分的生长和发育.信号传递能够帮助植物快速响应环境变化,如干旱胁迫下气孔调节和根系水分吸收调整.柠条和油蒿细根生长量和死亡量分别受土壤水分和温度共同影响,且不同年代植物对土壤水分利用深度和比例存在差异[53].柠条主要利用60、100 cm的土壤水分,二者占全部水源的75%左右;而油蒿主要利用10、20 cm的土壤水分,二者占全部水源的80%左右.随着固沙植被年限增加,植物对土壤表层水分利用比例逐渐减少,这是植物为了适应深层土壤水分相对稳定环境而做出的生理调整,也反映了植物地上和地下部分在资源获取和分配上的协调性.干旱区植物通过地上与地下部分的形态互补、资源分配动态调整及生理信号联动,实现了水分获取、运输与消耗的高效协调[85].这种根冠协同机制是其适应极端干旱的核心策略,也为人工植被恢复中的物种选择和生态管理提供了理论依据. ...
Effect of rainfall pulses on plant growth and transpiration of two xerophytic shrubs in a revegetated desert area:Tengger Desert,China
1
2016
... 叶片吸水对干旱沙区植物具有重要生理和生态作用.如光合性能改善,植物组织含水量增加,甚至延长叶片寿命、增加结实量、促进植物生长等[75,84].在沙坡头人工固沙植被区,小于5 mm的降水占到总降水频次的63.9%[85],凝结水量也占全年降水量的10%~20%[86],通过同位素示踪技术,在沙坡头人工植被区证明了典型固沙灌木也存在叶片吸水的现象,量化了柠条和油蒿单位面积吸水量分别为3.03~11.02、5.86~24.76 g,单位面积植物群落水分吸收能力分别为300~600、600~800 g,相当于0.3~0.8 mm的同期降雨量.[26].这表明叶片吸水在一定程度上能够补充植物体内的水分亏缺,尤其在干旱时期对维持植物的生理活性具有重要作用. ...
Hydraulic lift in Acacia tortilis trees on an east African Savannah
1
2003
... 在荒漠人工植被生态系统中,水分是限制植物生长的主要环境因子,不同植物类群对水分适应的多样性是维持生态系统平衡和可持续性的关键因素[1].对于固沙灌木和半灌木,不同生物学特性决定其维持生存的生态水文学机理不同.如柠条叶与嫩茎上丰富的软毛,为叶片吸收凝结水提供了有利条件;而油蒿根系主要分布在浅层土壤(0~20 cm),对树干径流的利用比例可能高于柠条[55,65].同时,植物水分利用对策也可能随着降水梯度的变化而改变,如东非萨瓦纳生境中特别干旱时段,水力提升现象和树干径流基本消失,合欢(Acacia tortilis)主要通过叶片吸水来适应干旱胁迫[87];当降水量为1.5~5.8 mm,许多植物如三齿团香木(Larrea tridentate)、美洲焦油菊(Flourensia cernua)和腺牧豆树(Prosopis glandulosa)产生树干径流,通过根系优先流来补充土壤水分[88];干季末期大降雨事件(50 mm)之后,萨瓦纳8种主要植物的根系传导水分状况表明:水分在水势梯度驱动下,先从浅层土壤进入浅层根,之后进入主根,最后再由主根进入深层较干燥土壤中,完成水力再分配[89].这说明植物水分利用方式或者对策会随着降雨量的变化而变化. ...
Stemflow in two xerophytic shrubs and its significance to soil water and nutrient enrichment
1
2013
... 在荒漠人工植被生态系统中,水分是限制植物生长的主要环境因子,不同植物类群对水分适应的多样性是维持生态系统平衡和可持续性的关键因素[1].对于固沙灌木和半灌木,不同生物学特性决定其维持生存的生态水文学机理不同.如柠条叶与嫩茎上丰富的软毛,为叶片吸收凝结水提供了有利条件;而油蒿根系主要分布在浅层土壤(0~20 cm),对树干径流的利用比例可能高于柠条[55,65].同时,植物水分利用对策也可能随着降水梯度的变化而改变,如东非萨瓦纳生境中特别干旱时段,水力提升现象和树干径流基本消失,合欢(Acacia tortilis)主要通过叶片吸水来适应干旱胁迫[87];当降水量为1.5~5.8 mm,许多植物如三齿团香木(Larrea tridentate)、美洲焦油菊(Flourensia cernua)和腺牧豆树(Prosopis glandulosa)产生树干径流,通过根系优先流来补充土壤水分[88];干季末期大降雨事件(50 mm)之后,萨瓦纳8种主要植物的根系传导水分状况表明:水分在水势梯度驱动下,先从浅层土壤进入浅层根,之后进入主根,最后再由主根进入深层较干燥土壤中,完成水力再分配[89].这说明植物水分利用方式或者对策会随着降雨量的变化而变化. ...
Hydraulic redistribution of soil water by Neotropical Savanna trees
1
2002
... 在荒漠人工植被生态系统中,水分是限制植物生长的主要环境因子,不同植物类群对水分适应的多样性是维持生态系统平衡和可持续性的关键因素[1].对于固沙灌木和半灌木,不同生物学特性决定其维持生存的生态水文学机理不同.如柠条叶与嫩茎上丰富的软毛,为叶片吸收凝结水提供了有利条件;而油蒿根系主要分布在浅层土壤(0~20 cm),对树干径流的利用比例可能高于柠条[55,65].同时,植物水分利用对策也可能随着降水梯度的变化而改变,如东非萨瓦纳生境中特别干旱时段,水力提升现象和树干径流基本消失,合欢(Acacia tortilis)主要通过叶片吸水来适应干旱胁迫[87];当降水量为1.5~5.8 mm,许多植物如三齿团香木(Larrea tridentate)、美洲焦油菊(Flourensia cernua)和腺牧豆树(Prosopis glandulosa)产生树干径流,通过根系优先流来补充土壤水分[88];干季末期大降雨事件(50 mm)之后,萨瓦纳8种主要植物的根系传导水分状况表明:水分在水势梯度驱动下,先从浅层土壤进入浅层根,之后进入主根,最后再由主根进入深层较干燥土壤中,完成水力再分配[89].这说明植物水分利用方式或者对策会随着降雨量的变化而变化. ...
2
2016
... 水量平衡动态变化直接关系到植被生长与存活,是植被-土壤系统稳定和健康的核心[90].水量平衡各组分构成在时空尺度上的变异直接影响水量平衡.前期对固沙植被水量平衡研究发现,植物蒸腾量和土壤蒸发量均随着年降水量减少而减少;但是随着降水减少,蒸腾量占蒸散量比例依次减少,而蒸发量占蒸散量比例则依次增大[66].这说明干旱胁迫不仅降低了植物的蒸腾量和蒸散量,而且增加降水的蒸发损失,可供植物利用的水分大幅减少,使得水量平衡各组分发生变化.固沙区生物土壤结皮的形成同样对降水再分配产生影响.生物土壤结皮能够拦截大部分降水[91],不利于高等植物吸收利用.生物土壤结皮对蒸发的影响具有降水依赖性,在大降水条件下(>5 mm)促进土壤蒸发,而在小降水后则抑制蒸发[92].水量平衡各组分对水分来源(主要是降水)依赖性的差异影响到其本身.对水量平衡中各组分的区分及量化是了解其时空异质性的关键所在. ...
... 通过相关资料计算得出柠条和油蒿水量平衡各分量的分配比例[90](图2),结果显示在沙坡头多年平均降雨量(186 mm)与凝结水(25 mm)情况下,柠条冠层截留量较高(21.85 mm,占输入量的10.36%),穿透雨为163.5 mm,叶片吸水量可达5.2 mm,蒸腾(82.87 mm)与蒸发(121.16 mm)为主要耗水途径,树干径流为4.7 mm,土壤储水量波动较大(±23.92 mm).油蒿冠层截留量较柠条降至14.9 mm(占比6.74%),穿透雨增加至173.7 mm,叶片吸水量升至9.7 mm,蒸腾(83.9 mm)与蒸发(122.2 mm)略有增强,但树干径流大幅减少至1.3 mm,土壤储水量波动较小(±14.56 mm).柠条冠层截留能力更强,而油蒿冠层透水性更高,穿透雨比例增加,水分分配稳定性提升.两物种的水文差异可能源于冠层结构特性(如叶片密度、茎干形态)对降水截留、再分配及土壤水分动态调控作用[55,57]. ...
Long-term ecosystem effects of sand-binding vegetation in the Tengger Desert,Northern China
1
2004
... 水量平衡动态变化直接关系到植被生长与存活,是植被-土壤系统稳定和健康的核心[90].水量平衡各组分构成在时空尺度上的变异直接影响水量平衡.前期对固沙植被水量平衡研究发现,植物蒸腾量和土壤蒸发量均随着年降水量减少而减少;但是随着降水减少,蒸腾量占蒸散量比例依次减少,而蒸发量占蒸散量比例则依次增大[66].这说明干旱胁迫不仅降低了植物的蒸腾量和蒸散量,而且增加降水的蒸发损失,可供植物利用的水分大幅减少,使得水量平衡各组分发生变化.固沙区生物土壤结皮的形成同样对降水再分配产生影响.生物土壤结皮能够拦截大部分降水[91],不利于高等植物吸收利用.生物土壤结皮对蒸发的影响具有降水依赖性,在大降水条件下(>5 mm)促进土壤蒸发,而在小降水后则抑制蒸发[92].水量平衡各组分对水分来源(主要是降水)依赖性的差异影响到其本身.对水量平衡中各组分的区分及量化是了解其时空异质性的关键所在. ...
Evaporation properties of a revegetated area of the Tengger Desert,North China
1
2008
... 水量平衡动态变化直接关系到植被生长与存活,是植被-土壤系统稳定和健康的核心[90].水量平衡各组分构成在时空尺度上的变异直接影响水量平衡.前期对固沙植被水量平衡研究发现,植物蒸腾量和土壤蒸发量均随着年降水量减少而减少;但是随着降水减少,蒸腾量占蒸散量比例依次减少,而蒸发量占蒸散量比例则依次增大[66].这说明干旱胁迫不仅降低了植物的蒸腾量和蒸散量,而且增加降水的蒸发损失,可供植物利用的水分大幅减少,使得水量平衡各组分发生变化.固沙区生物土壤结皮的形成同样对降水再分配产生影响.生物土壤结皮能够拦截大部分降水[91],不利于高等植物吸收利用.生物土壤结皮对蒸发的影响具有降水依赖性,在大降水条件下(>5 mm)促进土壤蒸发,而在小降水后则抑制蒸发[92].水量平衡各组分对水分来源(主要是降水)依赖性的差异影响到其本身.对水量平衡中各组分的区分及量化是了解其时空异质性的关键所在. ...
Hierarchy of responses to resource pulses in arid and semi-arid ecosystems
1
2004
... 作为干旱区重要水分补给源降水,具有显著的时空异质性和随机性.在荒漠人工植被区,脉冲式降水引起土壤水分和植被格局的动态变化[93].降水为随机的脉冲现象,考虑其对植被的影响,探讨其生态水文过程,已成为国际生态水文学界关注的热点[94-95].腾格里沙漠降水具有明显的脉冲特征,0~5 mm小降水事件占全年降水事件的77.8%,且降水间隔期以0~10 d为主.这种脉冲降水模式对固沙灌木生长和生理过程产生重要影响.固沙灌木对不同强度和频率的降水脉冲响应机制不同[85].0~5 d降水间隔对于油蒿叶面积指数(LAI)、新枝长和蒸腾耗水影响较大(50%左右),其次是5~10 d降水间隔.当降水间隔大于10 d时,其对油蒿影响较为稳定.0~5 d降水间隔对油蒿生理生态特征影响起决定作用的为0~5 mm的小降水事件.柠条LAI和蒸腾耗水对0~5 d降水间隔较为敏感,而对新枝长影响最大则为5~10 d降水间隔.与油蒿相比,0~5 d降水间隔对柠条生理生态特征影响较小,而当降水间隔大于5 d时,其对柠条影响作用要大于油蒿.这说明在干旱人工植被区,间歇性小降水事件更有利于油蒿生长发育. ...
Rainfall pulse response of carbon exchange to the timing of natural intra-annual rainfall in a temperate grass ecosystem
1
2020
... 作为干旱区重要水分补给源降水,具有显著的时空异质性和随机性.在荒漠人工植被区,脉冲式降水引起土壤水分和植被格局的动态变化[93].降水为随机的脉冲现象,考虑其对植被的影响,探讨其生态水文过程,已成为国际生态水文学界关注的热点[94-95].腾格里沙漠降水具有明显的脉冲特征,0~5 mm小降水事件占全年降水事件的77.8%,且降水间隔期以0~10 d为主.这种脉冲降水模式对固沙灌木生长和生理过程产生重要影响.固沙灌木对不同强度和频率的降水脉冲响应机制不同[85].0~5 d降水间隔对于油蒿叶面积指数(LAI)、新枝长和蒸腾耗水影响较大(50%左右),其次是5~10 d降水间隔.当降水间隔大于10 d时,其对油蒿影响较为稳定.0~5 d降水间隔对油蒿生理生态特征影响起决定作用的为0~5 mm的小降水事件.柠条LAI和蒸腾耗水对0~5 d降水间隔较为敏感,而对新枝长影响最大则为5~10 d降水间隔.与油蒿相比,0~5 d降水间隔对柠条生理生态特征影响较小,而当降水间隔大于5 d时,其对柠条影响作用要大于油蒿.这说明在干旱人工植被区,间歇性小降水事件更有利于油蒿生长发育. ...
Plant responses to changing rainfall frequency and intensity
1
2024
... 作为干旱区重要水分补给源降水,具有显著的时空异质性和随机性.在荒漠人工植被区,脉冲式降水引起土壤水分和植被格局的动态变化[93].降水为随机的脉冲现象,考虑其对植被的影响,探讨其生态水文过程,已成为国际生态水文学界关注的热点[94-95].腾格里沙漠降水具有明显的脉冲特征,0~5 mm小降水事件占全年降水事件的77.8%,且降水间隔期以0~10 d为主.这种脉冲降水模式对固沙灌木生长和生理过程产生重要影响.固沙灌木对不同强度和频率的降水脉冲响应机制不同[85].0~5 d降水间隔对于油蒿叶面积指数(LAI)、新枝长和蒸腾耗水影响较大(50%左右),其次是5~10 d降水间隔.当降水间隔大于10 d时,其对油蒿影响较为稳定.0~5 d降水间隔对油蒿生理生态特征影响起决定作用的为0~5 mm的小降水事件.柠条LAI和蒸腾耗水对0~5 d降水间隔较为敏感,而对新枝长影响最大则为5~10 d降水间隔.与油蒿相比,0~5 d降水间隔对柠条生理生态特征影响较小,而当降水间隔大于5 d时,其对柠条影响作用要大于油蒿.这说明在干旱人工植被区,间歇性小降水事件更有利于油蒿生长发育. ...
Water-use strategy of three central Asian desert shrubs and their responses to rain pulse events
1
2006
... 小降水可以增加浅层土壤水,更有利于草本植物生长,而深根植物如灌木对小降水则无显著响应,而更倾向于利用大降水[96].对古尔班通古特沙漠南缘降水模式和不同生活型植物对小降水生理响应研究表明所有草本植物(11种)对2~5 mm降水都有不同程度响应,只有1种灌木即驼绒藜(Ceratoides latens)对4 mm降水有响应.这12种植物在降水后第1天,叶片相对含水量显著增加,水分状况得到显著改善,表明这些植物对小降水(≤5 mm)有显著响应,而半灌木沙漠绢蒿(Seriphidium santolinum)、灌木梭梭(Haloxylon ammodendron)和柽柳(Tamarix ramosissima)、小乔木白梭梭(Haloxylon persicum)对5 mm以下降水均无显著响应,小降水主要被1年生草本植物所利用[97].通过统计各降水梯度上响应植物种数发现,在降水量达到3 mm时开始响应植物种类最多.1 mm降水后,多数草本植物净光合速率(Pn)、气孔导度(gs)上升,对干旱植物组织起到一定缓解作用[98].这可能是维持干旱沙区植被覆盖度及自组织格局的重要原因. ...
古尔班通古特沙漠不同生活型植物对小雨量降雨的响应
1
2009
... 小降水可以增加浅层土壤水,更有利于草本植物生长,而深根植物如灌木对小降水则无显著响应,而更倾向于利用大降水[96].对古尔班通古特沙漠南缘降水模式和不同生活型植物对小降水生理响应研究表明所有草本植物(11种)对2~5 mm降水都有不同程度响应,只有1种灌木即驼绒藜(Ceratoides latens)对4 mm降水有响应.这12种植物在降水后第1天,叶片相对含水量显著增加,水分状况得到显著改善,表明这些植物对小降水(≤5 mm)有显著响应,而半灌木沙漠绢蒿(Seriphidium santolinum)、灌木梭梭(Haloxylon ammodendron)和柽柳(Tamarix ramosissima)、小乔木白梭梭(Haloxylon persicum)对5 mm以下降水均无显著响应,小降水主要被1年生草本植物所利用[97].通过统计各降水梯度上响应植物种数发现,在降水量达到3 mm时开始响应植物种类最多.1 mm降水后,多数草本植物净光合速率(Pn)、气孔导度(gs)上升,对干旱植物组织起到一定缓解作用[98].这可能是维持干旱沙区植被覆盖度及自组织格局的重要原因. ...
荒漠草本植物在不同降雨模式下的光合响应和生物量分配策略
1
2013
... 小降水可以增加浅层土壤水,更有利于草本植物生长,而深根植物如灌木对小降水则无显著响应,而更倾向于利用大降水[96].对古尔班通古特沙漠南缘降水模式和不同生活型植物对小降水生理响应研究表明所有草本植物(11种)对2~5 mm降水都有不同程度响应,只有1种灌木即驼绒藜(Ceratoides latens)对4 mm降水有响应.这12种植物在降水后第1天,叶片相对含水量显著增加,水分状况得到显著改善,表明这些植物对小降水(≤5 mm)有显著响应,而半灌木沙漠绢蒿(Seriphidium santolinum)、灌木梭梭(Haloxylon ammodendron)和柽柳(Tamarix ramosissima)、小乔木白梭梭(Haloxylon persicum)对5 mm以下降水均无显著响应,小降水主要被1年生草本植物所利用[97].通过统计各降水梯度上响应植物种数发现,在降水量达到3 mm时开始响应植物种类最多.1 mm降水后,多数草本植物净光合速率(Pn)、气孔导度(gs)上升,对干旱植物组织起到一定缓解作用[98].这可能是维持干旱沙区植被覆盖度及自组织格局的重要原因. ...
Sap flow of Artemisia ordosica and the influence of environmental factors in a revegetated desert area:Tengger Desert,China
1
2010
... 荒漠植物生理生态特征对降水响应亦具有尺度依赖性.植物自身生理特点,如植物光合和蒸腾作用对脉冲降水响应,主要表现在短时间尺度上[99].在日尺度上,小降水事件增加植物蒸腾,而大降水事件降低植物蒸腾,主要原因是小降水事件降低植物叶片水势,而后在很短时间内迅速恢复.但在降水量较大时,植物叶片水势增大,太阳辐射、空气温度以及饱和水汽压差均在之后很长一段时间内保持较低水平,因此植物蒸腾量下降.另外树干液流与蒸腾之间存在时间滞后[100].在长时间尺度上,植物对脉冲降水响应主要体现在种群结构、植被盖度或植物优势种的变化等方面.柠条和油蒿的叶面积指数,NBL和蒸腾均与当地年降水量显著线性正相关.间歇性降水,特别是0~5 mm和0~5 d内降水量,对水分胁迫下植被生存起着积极作用.很少水量可以激活干旱沙漠地区诸多生物过程,如种子发芽和繁殖等.降水集中于某一个时间段,则有可能造成土壤水分流失影响植被的可持续性和生存[101].间歇性降水导致土壤水分在局部和短时间内升高,植物能进行充分利用,同时一部分降水在土壤表层快速蒸发后通过入渗进入深层土壤,干旱期间供植物使用[102].通常情况下,这种类型的脉冲降水更有利于油蒿生存,油蒿根系主要分布在土壤表层(0~40 cm),柠条根部主要分布在1 m的土层之下,其根系利用渗透水的机会较少[17-18].另外小降水事件能促进土壤养分吸收.研究表明较小强度降水事件(降水强度2.8 mm·h-1)中,37.1%侵蚀产物、30.3%有机质、48.3%氮和30.0%溶解养分被重新分配至灌丛斑块;而在较大强度降水事件中(降水强度5.7 mm·h-1),31.1%侵蚀产物、9.84%有机质、19.0%氮和10.0%溶解养分被重新分配至灌丛斑块,其中大部分被径流所带走[103]. ...
Less than 20-min time lags between transpiration and stem sap flow in emergent trees in a Bornean tropical rainforest
1
2008
... 荒漠植物生理生态特征对降水响应亦具有尺度依赖性.植物自身生理特点,如植物光合和蒸腾作用对脉冲降水响应,主要表现在短时间尺度上[99].在日尺度上,小降水事件增加植物蒸腾,而大降水事件降低植物蒸腾,主要原因是小降水事件降低植物叶片水势,而后在很短时间内迅速恢复.但在降水量较大时,植物叶片水势增大,太阳辐射、空气温度以及饱和水汽压差均在之后很长一段时间内保持较低水平,因此植物蒸腾量下降.另外树干液流与蒸腾之间存在时间滞后[100].在长时间尺度上,植物对脉冲降水响应主要体现在种群结构、植被盖度或植物优势种的变化等方面.柠条和油蒿的叶面积指数,NBL和蒸腾均与当地年降水量显著线性正相关.间歇性降水,特别是0~5 mm和0~5 d内降水量,对水分胁迫下植被生存起着积极作用.很少水量可以激活干旱沙漠地区诸多生物过程,如种子发芽和繁殖等.降水集中于某一个时间段,则有可能造成土壤水分流失影响植被的可持续性和生存[101].间歇性降水导致土壤水分在局部和短时间内升高,植物能进行充分利用,同时一部分降水在土壤表层快速蒸发后通过入渗进入深层土壤,干旱期间供植物使用[102].通常情况下,这种类型的脉冲降水更有利于油蒿生存,油蒿根系主要分布在土壤表层(0~40 cm),柠条根部主要分布在1 m的土层之下,其根系利用渗透水的机会较少[17-18].另外小降水事件能促进土壤养分吸收.研究表明较小强度降水事件(降水强度2.8 mm·h-1)中,37.1%侵蚀产物、30.3%有机质、48.3%氮和30.0%溶解养分被重新分配至灌丛斑块;而在较大强度降水事件中(降水强度5.7 mm·h-1),31.1%侵蚀产物、9.84%有机质、19.0%氮和10.0%溶解养分被重新分配至灌丛斑块,其中大部分被径流所带走[103]. ...
Vegetation response to rainfall intermittency in drylands:results from a simple ecohydrological box model
1
2007
... 荒漠植物生理生态特征对降水响应亦具有尺度依赖性.植物自身生理特点,如植物光合和蒸腾作用对脉冲降水响应,主要表现在短时间尺度上[99].在日尺度上,小降水事件增加植物蒸腾,而大降水事件降低植物蒸腾,主要原因是小降水事件降低植物叶片水势,而后在很短时间内迅速恢复.但在降水量较大时,植物叶片水势增大,太阳辐射、空气温度以及饱和水汽压差均在之后很长一段时间内保持较低水平,因此植物蒸腾量下降.另外树干液流与蒸腾之间存在时间滞后[100].在长时间尺度上,植物对脉冲降水响应主要体现在种群结构、植被盖度或植物优势种的变化等方面.柠条和油蒿的叶面积指数,NBL和蒸腾均与当地年降水量显著线性正相关.间歇性降水,特别是0~5 mm和0~5 d内降水量,对水分胁迫下植被生存起着积极作用.很少水量可以激活干旱沙漠地区诸多生物过程,如种子发芽和繁殖等.降水集中于某一个时间段,则有可能造成土壤水分流失影响植被的可持续性和生存[101].间歇性降水导致土壤水分在局部和短时间内升高,植物能进行充分利用,同时一部分降水在土壤表层快速蒸发后通过入渗进入深层土壤,干旱期间供植物使用[102].通常情况下,这种类型的脉冲降水更有利于油蒿生存,油蒿根系主要分布在土壤表层(0~40 cm),柠条根部主要分布在1 m的土层之下,其根系利用渗透水的机会较少[17-18].另外小降水事件能促进土壤养分吸收.研究表明较小强度降水事件(降水强度2.8 mm·h-1)中,37.1%侵蚀产物、30.3%有机质、48.3%氮和30.0%溶解养分被重新分配至灌丛斑块;而在较大强度降水事件中(降水强度5.7 mm·h-1),31.1%侵蚀产物、9.84%有机质、19.0%氮和10.0%溶解养分被重新分配至灌丛斑块,其中大部分被径流所带走[103]. ...
Patterned vegetation and rainfall intermittency
1
2009
... 荒漠植物生理生态特征对降水响应亦具有尺度依赖性.植物自身生理特点,如植物光合和蒸腾作用对脉冲降水响应,主要表现在短时间尺度上[99].在日尺度上,小降水事件增加植物蒸腾,而大降水事件降低植物蒸腾,主要原因是小降水事件降低植物叶片水势,而后在很短时间内迅速恢复.但在降水量较大时,植物叶片水势增大,太阳辐射、空气温度以及饱和水汽压差均在之后很长一段时间内保持较低水平,因此植物蒸腾量下降.另外树干液流与蒸腾之间存在时间滞后[100].在长时间尺度上,植物对脉冲降水响应主要体现在种群结构、植被盖度或植物优势种的变化等方面.柠条和油蒿的叶面积指数,NBL和蒸腾均与当地年降水量显著线性正相关.间歇性降水,特别是0~5 mm和0~5 d内降水量,对水分胁迫下植被生存起着积极作用.很少水量可以激活干旱沙漠地区诸多生物过程,如种子发芽和繁殖等.降水集中于某一个时间段,则有可能造成土壤水分流失影响植被的可持续性和生存[101].间歇性降水导致土壤水分在局部和短时间内升高,植物能进行充分利用,同时一部分降水在土壤表层快速蒸发后通过入渗进入深层土壤,干旱期间供植物使用[102].通常情况下,这种类型的脉冲降水更有利于油蒿生存,油蒿根系主要分布在土壤表层(0~40 cm),柠条根部主要分布在1 m的土层之下,其根系利用渗透水的机会较少[17-18].另外小降水事件能促进土壤养分吸收.研究表明较小强度降水事件(降水强度2.8 mm·h-1)中,37.1%侵蚀产物、30.3%有机质、48.3%氮和30.0%溶解养分被重新分配至灌丛斑块;而在较大强度降水事件中(降水强度5.7 mm·h-1),31.1%侵蚀产物、9.84%有机质、19.0%氮和10.0%溶解养分被重新分配至灌丛斑块,其中大部分被径流所带走[103]. ...
Effects of crust and shrub patches on runoff,sedimentation,and related nutrient (C,N) redistribution in the desertified steppe zone of the Tengger Desert,Northern China
1
2008
... 荒漠植物生理生态特征对降水响应亦具有尺度依赖性.植物自身生理特点,如植物光合和蒸腾作用对脉冲降水响应,主要表现在短时间尺度上[99].在日尺度上,小降水事件增加植物蒸腾,而大降水事件降低植物蒸腾,主要原因是小降水事件降低植物叶片水势,而后在很短时间内迅速恢复.但在降水量较大时,植物叶片水势增大,太阳辐射、空气温度以及饱和水汽压差均在之后很长一段时间内保持较低水平,因此植物蒸腾量下降.另外树干液流与蒸腾之间存在时间滞后[100].在长时间尺度上,植物对脉冲降水响应主要体现在种群结构、植被盖度或植物优势种的变化等方面.柠条和油蒿的叶面积指数,NBL和蒸腾均与当地年降水量显著线性正相关.间歇性降水,特别是0~5 mm和0~5 d内降水量,对水分胁迫下植被生存起着积极作用.很少水量可以激活干旱沙漠地区诸多生物过程,如种子发芽和繁殖等.降水集中于某一个时间段,则有可能造成土壤水分流失影响植被的可持续性和生存[101].间歇性降水导致土壤水分在局部和短时间内升高,植物能进行充分利用,同时一部分降水在土壤表层快速蒸发后通过入渗进入深层土壤,干旱期间供植物使用[102].通常情况下,这种类型的脉冲降水更有利于油蒿生存,油蒿根系主要分布在土壤表层(0~40 cm),柠条根部主要分布在1 m的土层之下,其根系利用渗透水的机会较少[17-18].另外小降水事件能促进土壤养分吸收.研究表明较小强度降水事件(降水强度2.8 mm·h-1)中,37.1%侵蚀产物、30.3%有机质、48.3%氮和30.0%溶解养分被重新分配至灌丛斑块;而在较大强度降水事件中(降水强度5.7 mm·h-1),31.1%侵蚀产物、9.84%有机质、19.0%氮和10.0%溶解养分被重新分配至灌丛斑块,其中大部分被径流所带走[103]. ...
Plant hydraulics as a central hub integrating plant and ecosystem function:meeting report for 'Emerging Frontiers in Plant Hydraulics' (Washington,DC,May 2015)
1
2016
... 为适应环境水分变化而形成植物水力性状是植物水分传输和利用的核心,也是整合植物和生态系统功能的中心枢纽[104-105],能较好地描述植物对干旱胁迫的响应[106].植物在生长过程中通过构建特定叶片水力系统来保持植株在生长过程中水分运输效率[107].植物为适应陆地环境进化出木质部维管系统,通过水力学机制向光合器官长距离运输水分,同时通过木质部水力结构调整进行栓塞修复等[108]. ...
木本植物水力系统对干旱胁迫的响应机制
1
2021
... 为适应环境水分变化而形成植物水力性状是植物水分传输和利用的核心,也是整合植物和生态系统功能的中心枢纽[104-105],能较好地描述植物对干旱胁迫的响应[106].植物在生长过程中通过构建特定叶片水力系统来保持植株在生长过程中水分运输效率[107].植物为适应陆地环境进化出木质部维管系统,通过水力学机制向光合器官长距离运输水分,同时通过木质部水力结构调整进行栓塞修复等[108]. ...
基于植物功能性状的生态学研究进展:从个体水平到全球尺度
2
2019
... 为适应环境水分变化而形成植物水力性状是植物水分传输和利用的核心,也是整合植物和生态系统功能的中心枢纽[104-105],能较好地描述植物对干旱胁迫的响应[106].植物在生长过程中通过构建特定叶片水力系统来保持植株在生长过程中水分运输效率[107].植物为适应陆地环境进化出木质部维管系统,通过水力学机制向光合器官长距离运输水分,同时通过木质部水力结构调整进行栓塞修复等[108]. ...
... 水力功能性状决定植物水分利用特征,植物对不同环境水分条件的长期适应形成多种水分利用策略.个体尺度上,植物通过增加根系的吸水能力,如一些典型荒漠植物梭梭和琵琶柴(Reaumuria soongarica)可根据水分条件调节根系和冠层生长,从而维持正常光合作用[109],也可以通过叶片吸水、木质部光合作用或者根系水力提升等措施来提高植物水分利用效率[110].植物在遭受轻度水分胁迫时,水势调节和气孔运动联动避免水力障碍,中度水分胁迫时,木质部水力特性权衡与协调保证植物体内水分正常运输,极端干旱时,植物通过水力分割等保证植物存活[106].另一方面,水力结构影响物种分布、抗逆能力等,植物水力性状决定极端干旱引起的植物枯死率[111].木质部水力安全是喀斯特木本植物适应干旱的特征,也是其生态位分化和群落构建的潜在机制[112].水力结构特征决定了热带喀斯特及沟谷雨林木本植物种类组成[113],水力结构特征影响物种多样性和空间分布[114],如水分运输能力较高的植物表现出较高的碳同化能力和生长速率,偏好生长于水分可利用性较高生境[115].群落不同层片植物水力结构策略的权衡是维持不同垂直层次物种共存的重要机制[116].区域尺度上,沿降水梯度分布的锦鸡儿属植物叶片光合及水力供给特征分化明显[117].不同海拔梯度树木的功能性状也存在显著差异,如海拔分布上限较低的树种具有较大的导管直径和水力直径,水分传输效率和生长速率较高;而海拔分布上限较高的树种具有更大的边材密度和导管密度,抵抗冰冻胁迫造成气穴化栓塞的能力更高[107]. ...
The hydraulic architecture of an arborescent monocot:ontogeny-related adjustments in vessel size and leaf area compensate for increased resistance
2
2021
... 为适应环境水分变化而形成植物水力性状是植物水分传输和利用的核心,也是整合植物和生态系统功能的中心枢纽[104-105],能较好地描述植物对干旱胁迫的响应[106].植物在生长过程中通过构建特定叶片水力系统来保持植株在生长过程中水分运输效率[107].植物为适应陆地环境进化出木质部维管系统,通过水力学机制向光合器官长距离运输水分,同时通过木质部水力结构调整进行栓塞修复等[108]. ...
... 水力功能性状决定植物水分利用特征,植物对不同环境水分条件的长期适应形成多种水分利用策略.个体尺度上,植物通过增加根系的吸水能力,如一些典型荒漠植物梭梭和琵琶柴(Reaumuria soongarica)可根据水分条件调节根系和冠层生长,从而维持正常光合作用[109],也可以通过叶片吸水、木质部光合作用或者根系水力提升等措施来提高植物水分利用效率[110].植物在遭受轻度水分胁迫时,水势调节和气孔运动联动避免水力障碍,中度水分胁迫时,木质部水力特性权衡与协调保证植物体内水分正常运输,极端干旱时,植物通过水力分割等保证植物存活[106].另一方面,水力结构影响物种分布、抗逆能力等,植物水力性状决定极端干旱引起的植物枯死率[111].木质部水力安全是喀斯特木本植物适应干旱的特征,也是其生态位分化和群落构建的潜在机制[112].水力结构特征决定了热带喀斯特及沟谷雨林木本植物种类组成[113],水力结构特征影响物种多样性和空间分布[114],如水分运输能力较高的植物表现出较高的碳同化能力和生长速率,偏好生长于水分可利用性较高生境[115].群落不同层片植物水力结构策略的权衡是维持不同垂直层次物种共存的重要机制[116].区域尺度上,沿降水梯度分布的锦鸡儿属植物叶片光合及水力供给特征分化明显[117].不同海拔梯度树木的功能性状也存在显著差异,如海拔分布上限较低的树种具有较大的导管直径和水力直径,水分传输效率和生长速率较高;而海拔分布上限较高的树种具有更大的边材密度和导管密度,抵抗冰冻胁迫造成气穴化栓塞的能力更高[107]. ...
植物木质部水力学研究进展
1
2020
... 为适应环境水分变化而形成植物水力性状是植物水分传输和利用的核心,也是整合植物和生态系统功能的中心枢纽[104-105],能较好地描述植物对干旱胁迫的响应[106].植物在生长过程中通过构建特定叶片水力系统来保持植株在生长过程中水分运输效率[107].植物为适应陆地环境进化出木质部维管系统,通过水力学机制向光合器官长距离运输水分,同时通过木质部水力结构调整进行栓塞修复等[108]. ...
3种荒漠灌木的用水策略及相关的叶片生理表现
1
2005
... 水力功能性状决定植物水分利用特征,植物对不同环境水分条件的长期适应形成多种水分利用策略.个体尺度上,植物通过增加根系的吸水能力,如一些典型荒漠植物梭梭和琵琶柴(Reaumuria soongarica)可根据水分条件调节根系和冠层生长,从而维持正常光合作用[109],也可以通过叶片吸水、木质部光合作用或者根系水力提升等措施来提高植物水分利用效率[110].植物在遭受轻度水分胁迫时,水势调节和气孔运动联动避免水力障碍,中度水分胁迫时,木质部水力特性权衡与协调保证植物体内水分正常运输,极端干旱时,植物通过水力分割等保证植物存活[106].另一方面,水力结构影响物种分布、抗逆能力等,植物水力性状决定极端干旱引起的植物枯死率[111].木质部水力安全是喀斯特木本植物适应干旱的特征,也是其生态位分化和群落构建的潜在机制[112].水力结构特征决定了热带喀斯特及沟谷雨林木本植物种类组成[113],水力结构特征影响物种多样性和空间分布[114],如水分运输能力较高的植物表现出较高的碳同化能力和生长速率,偏好生长于水分可利用性较高生境[115].群落不同层片植物水力结构策略的权衡是维持不同垂直层次物种共存的重要机制[116].区域尺度上,沿降水梯度分布的锦鸡儿属植物叶片光合及水力供给特征分化明显[117].不同海拔梯度树木的功能性状也存在显著差异,如海拔分布上限较低的树种具有较大的导管直径和水力直径,水分传输效率和生长速率较高;而海拔分布上限较高的树种具有更大的边材密度和导管密度,抵抗冰冻胁迫造成气穴化栓塞的能力更高[107]. ...
中国干旱沙区的生态重建与恢复:沙坡头站60年重要研究进展综述
1
2016
... 水力功能性状决定植物水分利用特征,植物对不同环境水分条件的长期适应形成多种水分利用策略.个体尺度上,植物通过增加根系的吸水能力,如一些典型荒漠植物梭梭和琵琶柴(Reaumuria soongarica)可根据水分条件调节根系和冠层生长,从而维持正常光合作用[109],也可以通过叶片吸水、木质部光合作用或者根系水力提升等措施来提高植物水分利用效率[110].植物在遭受轻度水分胁迫时,水势调节和气孔运动联动避免水力障碍,中度水分胁迫时,木质部水力特性权衡与协调保证植物体内水分正常运输,极端干旱时,植物通过水力分割等保证植物存活[106].另一方面,水力结构影响物种分布、抗逆能力等,植物水力性状决定极端干旱引起的植物枯死率[111].木质部水力安全是喀斯特木本植物适应干旱的特征,也是其生态位分化和群落构建的潜在机制[112].水力结构特征决定了热带喀斯特及沟谷雨林木本植物种类组成[113],水力结构特征影响物种多样性和空间分布[114],如水分运输能力较高的植物表现出较高的碳同化能力和生长速率,偏好生长于水分可利用性较高生境[115].群落不同层片植物水力结构策略的权衡是维持不同垂直层次物种共存的重要机制[116].区域尺度上,沿降水梯度分布的锦鸡儿属植物叶片光合及水力供给特征分化明显[117].不同海拔梯度树木的功能性状也存在显著差异,如海拔分布上限较低的树种具有较大的导管直径和水力直径,水分传输效率和生长速率较高;而海拔分布上限较高的树种具有更大的边材密度和导管密度,抵抗冰冻胁迫造成气穴化栓塞的能力更高[107]. ...
Hydraulic prediction of drought-induced plant dieback and top-kill depends on leaf habit and growth form
1
2021
... 水力功能性状决定植物水分利用特征,植物对不同环境水分条件的长期适应形成多种水分利用策略.个体尺度上,植物通过增加根系的吸水能力,如一些典型荒漠植物梭梭和琵琶柴(Reaumuria soongarica)可根据水分条件调节根系和冠层生长,从而维持正常光合作用[109],也可以通过叶片吸水、木质部光合作用或者根系水力提升等措施来提高植物水分利用效率[110].植物在遭受轻度水分胁迫时,水势调节和气孔运动联动避免水力障碍,中度水分胁迫时,木质部水力特性权衡与协调保证植物体内水分正常运输,极端干旱时,植物通过水力分割等保证植物存活[106].另一方面,水力结构影响物种分布、抗逆能力等,植物水力性状决定极端干旱引起的植物枯死率[111].木质部水力安全是喀斯特木本植物适应干旱的特征,也是其生态位分化和群落构建的潜在机制[112].水力结构特征决定了热带喀斯特及沟谷雨林木本植物种类组成[113],水力结构特征影响物种多样性和空间分布[114],如水分运输能力较高的植物表现出较高的碳同化能力和生长速率,偏好生长于水分可利用性较高生境[115].群落不同层片植物水力结构策略的权衡是维持不同垂直层次物种共存的重要机制[116].区域尺度上,沿降水梯度分布的锦鸡儿属植物叶片光合及水力供给特征分化明显[117].不同海拔梯度树木的功能性状也存在显著差异,如海拔分布上限较低的树种具有较大的导管直径和水力直径,水分传输效率和生长速率较高;而海拔分布上限较高的树种具有更大的边材密度和导管密度,抵抗冰冻胁迫造成气穴化栓塞的能力更高[107]. ...
热带亚热带喀斯特森林树种的水分生理研究进展
1
2023
... 水力功能性状决定植物水分利用特征,植物对不同环境水分条件的长期适应形成多种水分利用策略.个体尺度上,植物通过增加根系的吸水能力,如一些典型荒漠植物梭梭和琵琶柴(Reaumuria soongarica)可根据水分条件调节根系和冠层生长,从而维持正常光合作用[109],也可以通过叶片吸水、木质部光合作用或者根系水力提升等措施来提高植物水分利用效率[110].植物在遭受轻度水分胁迫时,水势调节和气孔运动联动避免水力障碍,中度水分胁迫时,木质部水力特性权衡与协调保证植物体内水分正常运输,极端干旱时,植物通过水力分割等保证植物存活[106].另一方面,水力结构影响物种分布、抗逆能力等,植物水力性状决定极端干旱引起的植物枯死率[111].木质部水力安全是喀斯特木本植物适应干旱的特征,也是其生态位分化和群落构建的潜在机制[112].水力结构特征决定了热带喀斯特及沟谷雨林木本植物种类组成[113],水力结构特征影响物种多样性和空间分布[114],如水分运输能力较高的植物表现出较高的碳同化能力和生长速率,偏好生长于水分可利用性较高生境[115].群落不同层片植物水力结构策略的权衡是维持不同垂直层次物种共存的重要机制[116].区域尺度上,沿降水梯度分布的锦鸡儿属植物叶片光合及水力供给特征分化明显[117].不同海拔梯度树木的功能性状也存在显著差异,如海拔分布上限较低的树种具有较大的导管直径和水力直径,水分传输效率和生长速率较高;而海拔分布上限较高的树种具有更大的边材密度和导管密度,抵抗冰冻胁迫造成气穴化栓塞的能力更高[107]. ...
Different hydraulic traits of woody plants from tropical forests with contrasting soil water availability
1
2017
... 水力功能性状决定植物水分利用特征,植物对不同环境水分条件的长期适应形成多种水分利用策略.个体尺度上,植物通过增加根系的吸水能力,如一些典型荒漠植物梭梭和琵琶柴(Reaumuria soongarica)可根据水分条件调节根系和冠层生长,从而维持正常光合作用[109],也可以通过叶片吸水、木质部光合作用或者根系水力提升等措施来提高植物水分利用效率[110].植物在遭受轻度水分胁迫时,水势调节和气孔运动联动避免水力障碍,中度水分胁迫时,木质部水力特性权衡与协调保证植物体内水分正常运输,极端干旱时,植物通过水力分割等保证植物存活[106].另一方面,水力结构影响物种分布、抗逆能力等,植物水力性状决定极端干旱引起的植物枯死率[111].木质部水力安全是喀斯特木本植物适应干旱的特征,也是其生态位分化和群落构建的潜在机制[112].水力结构特征决定了热带喀斯特及沟谷雨林木本植物种类组成[113],水力结构特征影响物种多样性和空间分布[114],如水分运输能力较高的植物表现出较高的碳同化能力和生长速率,偏好生长于水分可利用性较高生境[115].群落不同层片植物水力结构策略的权衡是维持不同垂直层次物种共存的重要机制[116].区域尺度上,沿降水梯度分布的锦鸡儿属植物叶片光合及水力供给特征分化明显[117].不同海拔梯度树木的功能性状也存在显著差异,如海拔分布上限较低的树种具有较大的导管直径和水力直径,水分传输效率和生长速率较高;而海拔分布上限较高的树种具有更大的边材密度和导管密度,抵抗冰冻胁迫造成气穴化栓塞的能力更高[107]. ...
广西弄岗喀斯特森林9种主要树种水力结构功能特征及其相关性分析
1
2023
... 水力功能性状决定植物水分利用特征,植物对不同环境水分条件的长期适应形成多种水分利用策略.个体尺度上,植物通过增加根系的吸水能力,如一些典型荒漠植物梭梭和琵琶柴(Reaumuria soongarica)可根据水分条件调节根系和冠层生长,从而维持正常光合作用[109],也可以通过叶片吸水、木质部光合作用或者根系水力提升等措施来提高植物水分利用效率[110].植物在遭受轻度水分胁迫时,水势调节和气孔运动联动避免水力障碍,中度水分胁迫时,木质部水力特性权衡与协调保证植物体内水分正常运输,极端干旱时,植物通过水力分割等保证植物存活[106].另一方面,水力结构影响物种分布、抗逆能力等,植物水力性状决定极端干旱引起的植物枯死率[111].木质部水力安全是喀斯特木本植物适应干旱的特征,也是其生态位分化和群落构建的潜在机制[112].水力结构特征决定了热带喀斯特及沟谷雨林木本植物种类组成[113],水力结构特征影响物种多样性和空间分布[114],如水分运输能力较高的植物表现出较高的碳同化能力和生长速率,偏好生长于水分可利用性较高生境[115].群落不同层片植物水力结构策略的权衡是维持不同垂直层次物种共存的重要机制[116].区域尺度上,沿降水梯度分布的锦鸡儿属植物叶片光合及水力供给特征分化明显[117].不同海拔梯度树木的功能性状也存在显著差异,如海拔分布上限较低的树种具有较大的导管直径和水力直径,水分传输效率和生长速率较高;而海拔分布上限较高的树种具有更大的边材密度和导管密度,抵抗冰冻胁迫造成气穴化栓塞的能力更高[107]. ...
Divergences in hydraulic architecture form an important basis for niche differentiation between diploid and polyploid Betula species in NE China
1
2017
... 水力功能性状决定植物水分利用特征,植物对不同环境水分条件的长期适应形成多种水分利用策略.个体尺度上,植物通过增加根系的吸水能力,如一些典型荒漠植物梭梭和琵琶柴(Reaumuria soongarica)可根据水分条件调节根系和冠层生长,从而维持正常光合作用[109],也可以通过叶片吸水、木质部光合作用或者根系水力提升等措施来提高植物水分利用效率[110].植物在遭受轻度水分胁迫时,水势调节和气孔运动联动避免水力障碍,中度水分胁迫时,木质部水力特性权衡与协调保证植物体内水分正常运输,极端干旱时,植物通过水力分割等保证植物存活[106].另一方面,水力结构影响物种分布、抗逆能力等,植物水力性状决定极端干旱引起的植物枯死率[111].木质部水力安全是喀斯特木本植物适应干旱的特征,也是其生态位分化和群落构建的潜在机制[112].水力结构特征决定了热带喀斯特及沟谷雨林木本植物种类组成[113],水力结构特征影响物种多样性和空间分布[114],如水分运输能力较高的植物表现出较高的碳同化能力和生长速率,偏好生长于水分可利用性较高生境[115].群落不同层片植物水力结构策略的权衡是维持不同垂直层次物种共存的重要机制[116].区域尺度上,沿降水梯度分布的锦鸡儿属植物叶片光合及水力供给特征分化明显[117].不同海拔梯度树木的功能性状也存在显著差异,如海拔分布上限较低的树种具有较大的导管直径和水力直径,水分传输效率和生长速率较高;而海拔分布上限较高的树种具有更大的边材密度和导管密度,抵抗冰冻胁迫造成气穴化栓塞的能力更高[107]. ...
浙江天童木本植物水力结构与树高的关联性
1
2020
... 水力功能性状决定植物水分利用特征,植物对不同环境水分条件的长期适应形成多种水分利用策略.个体尺度上,植物通过增加根系的吸水能力,如一些典型荒漠植物梭梭和琵琶柴(Reaumuria soongarica)可根据水分条件调节根系和冠层生长,从而维持正常光合作用[109],也可以通过叶片吸水、木质部光合作用或者根系水力提升等措施来提高植物水分利用效率[110].植物在遭受轻度水分胁迫时,水势调节和气孔运动联动避免水力障碍,中度水分胁迫时,木质部水力特性权衡与协调保证植物体内水分正常运输,极端干旱时,植物通过水力分割等保证植物存活[106].另一方面,水力结构影响物种分布、抗逆能力等,植物水力性状决定极端干旱引起的植物枯死率[111].木质部水力安全是喀斯特木本植物适应干旱的特征,也是其生态位分化和群落构建的潜在机制[112].水力结构特征决定了热带喀斯特及沟谷雨林木本植物种类组成[113],水力结构特征影响物种多样性和空间分布[114],如水分运输能力较高的植物表现出较高的碳同化能力和生长速率,偏好生长于水分可利用性较高生境[115].群落不同层片植物水力结构策略的权衡是维持不同垂直层次物种共存的重要机制[116].区域尺度上,沿降水梯度分布的锦鸡儿属植物叶片光合及水力供给特征分化明显[117].不同海拔梯度树木的功能性状也存在显著差异,如海拔分布上限较低的树种具有较大的导管直径和水力直径,水分传输效率和生长速率较高;而海拔分布上限较高的树种具有更大的边材密度和导管密度,抵抗冰冻胁迫造成气穴化栓塞的能力更高[107]. ...
Combined high leaf hydraulic safety and efficiency provides drought tolerance in Caragana species adapted to low mean annual precipitation
1
2021
... 水力功能性状决定植物水分利用特征,植物对不同环境水分条件的长期适应形成多种水分利用策略.个体尺度上,植物通过增加根系的吸水能力,如一些典型荒漠植物梭梭和琵琶柴(Reaumuria soongarica)可根据水分条件调节根系和冠层生长,从而维持正常光合作用[109],也可以通过叶片吸水、木质部光合作用或者根系水力提升等措施来提高植物水分利用效率[110].植物在遭受轻度水分胁迫时,水势调节和气孔运动联动避免水力障碍,中度水分胁迫时,木质部水力特性权衡与协调保证植物体内水分正常运输,极端干旱时,植物通过水力分割等保证植物存活[106].另一方面,水力结构影响物种分布、抗逆能力等,植物水力性状决定极端干旱引起的植物枯死率[111].木质部水力安全是喀斯特木本植物适应干旱的特征,也是其生态位分化和群落构建的潜在机制[112].水力结构特征决定了热带喀斯特及沟谷雨林木本植物种类组成[113],水力结构特征影响物种多样性和空间分布[114],如水分运输能力较高的植物表现出较高的碳同化能力和生长速率,偏好生长于水分可利用性较高生境[115].群落不同层片植物水力结构策略的权衡是维持不同垂直层次物种共存的重要机制[116].区域尺度上,沿降水梯度分布的锦鸡儿属植物叶片光合及水力供给特征分化明显[117].不同海拔梯度树木的功能性状也存在显著差异,如海拔分布上限较低的树种具有较大的导管直径和水力直径,水分传输效率和生长速率较高;而海拔分布上限较高的树种具有更大的边材密度和导管密度,抵抗冰冻胁迫造成气穴化栓塞的能力更高[107]. ...
木本植物应对干旱胁迫的响应机制:基于水力学性状视角
1
2024
... 随着灌木年龄增加,水力传输效率显著降低,但黎明前(φpd)、正午(φmd)及日变化(∆φmax)水势在不同年龄之间无显著变化,表明植物在长期生长过程中通过调整自身水力学特性来适应干旱环境[118].固沙灌木的木质部结构和功能对其水分传导效率至关重要.固沙灌木的木质部导水率受土壤水分含量影响.在水分充足条件下,木质部导水率较高,能够快速输送水分至植物各部分;干旱条件下木质部导水率降低,以减少气穴化栓塞的风险.例如,柠条和中间锦鸡儿(Caragana liouana)木质部导水率在不同季节不同土壤水分条件下表现出显著差异[119].腾格里沙漠东南缘沙坡头人工固沙植被区1956、1964、1981、1987年不同演替阶段柠条、油蒿和花棒水力参数研究表明:随着建植年限增加,3种固沙灌木水势和导水率呈现出逐渐降低趋势,其中柠条导水率下降最快,油蒿导水率下降最慢,木质部导水率损失百分数(PLC)则呈现出相反的变化趋势,3种固沙灌木PLC随着建植年限增加而上升,其中柠条和花棒的PLC增加速率高于油蒿[120].对沙坡头地区35种木本植物生长、水力传输效率及水力传输安全特征的研究表明,植物水力安全和生长速率对植物生存有直接影响,水力安全性越高且生长速率越慢的植物,对环境适应性及抗逆性越强.灌木保持低水分传输效率及低生长速率以维持生存,采取保守型生存策略,而乔木则具有较高水分传输效率且优先生长,采取索取型生存策略[121].固沙灌木通过根系分异、木质部抗栓塞、叶片形态调整及高效水分利用等水力学属性,适应干旱环境并发挥固沙功能. ...
固沙灌木柠条锦鸡儿和中间锦鸡儿木质部导水与叶片光合能力对土壤水分的响应
1
2023
... 随着灌木年龄增加,水力传输效率显著降低,但黎明前(φpd)、正午(φmd)及日变化(∆φmax)水势在不同年龄之间无显著变化,表明植物在长期生长过程中通过调整自身水力学特性来适应干旱环境[118].固沙灌木的木质部结构和功能对其水分传导效率至关重要.固沙灌木的木质部导水率受土壤水分含量影响.在水分充足条件下,木质部导水率较高,能够快速输送水分至植物各部分;干旱条件下木质部导水率降低,以减少气穴化栓塞的风险.例如,柠条和中间锦鸡儿(Caragana liouana)木质部导水率在不同季节不同土壤水分条件下表现出显著差异[119].腾格里沙漠东南缘沙坡头人工固沙植被区1956、1964、1981、1987年不同演替阶段柠条、油蒿和花棒水力参数研究表明:随着建植年限增加,3种固沙灌木水势和导水率呈现出逐渐降低趋势,其中柠条导水率下降最快,油蒿导水率下降最慢,木质部导水率损失百分数(PLC)则呈现出相反的变化趋势,3种固沙灌木PLC随着建植年限增加而上升,其中柠条和花棒的PLC增加速率高于油蒿[120].对沙坡头地区35种木本植物生长、水力传输效率及水力传输安全特征的研究表明,植物水力安全和生长速率对植物生存有直接影响,水力安全性越高且生长速率越慢的植物,对环境适应性及抗逆性越强.灌木保持低水分传输效率及低生长速率以维持生存,采取保守型生存策略,而乔木则具有较高水分传输效率且优先生长,采取索取型生存策略[121].固沙灌木通过根系分异、木质部抗栓塞、叶片形态调整及高效水分利用等水力学属性,适应干旱环境并发挥固沙功能. ...
干旱和遮荫对典型固沙灌木生存的影响
1
2023
... 随着灌木年龄增加,水力传输效率显著降低,但黎明前(φpd)、正午(φmd)及日变化(∆φmax)水势在不同年龄之间无显著变化,表明植物在长期生长过程中通过调整自身水力学特性来适应干旱环境[118].固沙灌木的木质部结构和功能对其水分传导效率至关重要.固沙灌木的木质部导水率受土壤水分含量影响.在水分充足条件下,木质部导水率较高,能够快速输送水分至植物各部分;干旱条件下木质部导水率降低,以减少气穴化栓塞的风险.例如,柠条和中间锦鸡儿(Caragana liouana)木质部导水率在不同季节不同土壤水分条件下表现出显著差异[119].腾格里沙漠东南缘沙坡头人工固沙植被区1956、1964、1981、1987年不同演替阶段柠条、油蒿和花棒水力参数研究表明:随着建植年限增加,3种固沙灌木水势和导水率呈现出逐渐降低趋势,其中柠条导水率下降最快,油蒿导水率下降最慢,木质部导水率损失百分数(PLC)则呈现出相反的变化趋势,3种固沙灌木PLC随着建植年限增加而上升,其中柠条和花棒的PLC增加速率高于油蒿[120].对沙坡头地区35种木本植物生长、水力传输效率及水力传输安全特征的研究表明,植物水力安全和生长速率对植物生存有直接影响,水力安全性越高且生长速率越慢的植物,对环境适应性及抗逆性越强.灌木保持低水分传输效率及低生长速率以维持生存,采取保守型生存策略,而乔木则具有较高水分传输效率且优先生长,采取索取型生存策略[121].固沙灌木通过根系分异、木质部抗栓塞、叶片形态调整及高效水分利用等水力学属性,适应干旱环境并发挥固沙功能. ...
Hydraulic safety and growth rather than climate of origin influence survival in desert shrubs and trees
1
2023
... 随着灌木年龄增加,水力传输效率显著降低,但黎明前(φpd)、正午(φmd)及日变化(∆φmax)水势在不同年龄之间无显著变化,表明植物在长期生长过程中通过调整自身水力学特性来适应干旱环境[118].固沙灌木的木质部结构和功能对其水分传导效率至关重要.固沙灌木的木质部导水率受土壤水分含量影响.在水分充足条件下,木质部导水率较高,能够快速输送水分至植物各部分;干旱条件下木质部导水率降低,以减少气穴化栓塞的风险.例如,柠条和中间锦鸡儿(Caragana liouana)木质部导水率在不同季节不同土壤水分条件下表现出显著差异[119].腾格里沙漠东南缘沙坡头人工固沙植被区1956、1964、1981、1987年不同演替阶段柠条、油蒿和花棒水力参数研究表明:随着建植年限增加,3种固沙灌木水势和导水率呈现出逐渐降低趋势,其中柠条导水率下降最快,油蒿导水率下降最慢,木质部导水率损失百分数(PLC)则呈现出相反的变化趋势,3种固沙灌木PLC随着建植年限增加而上升,其中柠条和花棒的PLC增加速率高于油蒿[120].对沙坡头地区35种木本植物生长、水力传输效率及水力传输安全特征的研究表明,植物水力安全和生长速率对植物生存有直接影响,水力安全性越高且生长速率越慢的植物,对环境适应性及抗逆性越强.灌木保持低水分传输效率及低生长速率以维持生存,采取保守型生存策略,而乔木则具有较高水分传输效率且优先生长,采取索取型生存策略[121].固沙灌木通过根系分异、木质部抗栓塞、叶片形态调整及高效水分利用等水力学属性,适应干旱环境并发挥固沙功能. ...
“植物-土壤”相互反馈的关键生态学问题:格局、过程与机制
1
2020
... 固沙灌木地上和地下部分在生长发育过程中存在密切协调关系,是植物适应干旱环境、维持自身稳定的关键[122].干旱区植物根系通常具有深根性和广布性.例如白梭梭和梭梭的垂直根深逾5 m,可至地下水层吸收水分[123].柽柳主根和侧根均极为发达,主根可伸至地下水层,最深10 m以上[124].此外,一些植物如沙拐枣根系多为水平分布,但在地下水较深的地区,其根系也能垂直向下发展以获取水分[125].以上根系的形态和分布使得植物能够有效地利用土壤水分资源.长期干旱时植物通过落叶(如梭梭)、茎干休眠等方式减少地上部分的水分消耗,同时维持根系活力(如沙拐枣根系持续生长寻找水源).植物地上和地下部分在生理功能上也表现为协调关系[126].植物通过冠层截留降水,减少土壤水分蒸发.同时,植物根系能够将截留水分吸收并输送到地上部分,支持植物蒸腾作用和光合作用.固沙灌木冠层截留和树干径流在水分再分配中起着重要作用,能够提高根际区水分供应[55].植物在资源分配上也表现出对干旱环境的适应性,根据环境条件调整地上和地下部分的生物量分配,以优化对光、水和养分等资源的利用[127].干旱条件下植物可能会将更多的资源分配到根系,以增强水分获取能力.同时,植物叶片和细根在碳、氮、磷等元素的分配中也存在协同关系.西北干旱区荒漠植物叶片和细根在碳、氮、磷含量及其计量比上表现出显著的正相关关系[128],这表明植物在完成正常的生命活动时,地上与地下部分的养分分配存在协同作用.深根系植物(如梭梭)主根深入地下水源,地上部分冠层稀疏(减少蒸腾面积),形成高而疏的形态,减少水分竞争.浅根系植物(如沙柳)侧根广泛分布于表层,冠层密集,依赖小降水后对表层水分的快速利用.植物地上和地下部分之间存在信息流和信号传递的协调机制[129].植物可通过木质部和韧皮部传递激素、压力波和电信号等,以协调地上和地下部分的生长和发育.信号传递能够帮助植物快速响应环境变化,如干旱胁迫下气孔调节和根系水分吸收调整.柠条和油蒿细根生长量和死亡量分别受土壤水分和温度共同影响,且不同年代植物对土壤水分利用深度和比例存在差异[53].柠条主要利用60、100 cm的土壤水分,二者占全部水源的75%左右;而油蒿主要利用10、20 cm的土壤水分,二者占全部水源的80%左右.随着固沙植被年限增加,植物对土壤表层水分利用比例逐渐减少,这是植物为了适应深层土壤水分相对稳定环境而做出的生理调整,也反映了植物地上和地下部分在资源获取和分配上的协调性.干旱区植物通过地上与地下部分的形态互补、资源分配动态调整及生理信号联动,实现了水分获取、运输与消耗的高效协调[85].这种根冠协同机制是其适应极端干旱的核心策略,也为人工植被恢复中的物种选择和生态管理提供了理论依据. ...
根深决定不同个体大小梭梭对夏季干旱生理响应的差异
1
2021
... 固沙灌木地上和地下部分在生长发育过程中存在密切协调关系,是植物适应干旱环境、维持自身稳定的关键[122].干旱区植物根系通常具有深根性和广布性.例如白梭梭和梭梭的垂直根深逾5 m,可至地下水层吸收水分[123].柽柳主根和侧根均极为发达,主根可伸至地下水层,最深10 m以上[124].此外,一些植物如沙拐枣根系多为水平分布,但在地下水较深的地区,其根系也能垂直向下发展以获取水分[125].以上根系的形态和分布使得植物能够有效地利用土壤水分资源.长期干旱时植物通过落叶(如梭梭)、茎干休眠等方式减少地上部分的水分消耗,同时维持根系活力(如沙拐枣根系持续生长寻找水源).植物地上和地下部分在生理功能上也表现为协调关系[126].植物通过冠层截留降水,减少土壤水分蒸发.同时,植物根系能够将截留水分吸收并输送到地上部分,支持植物蒸腾作用和光合作用.固沙灌木冠层截留和树干径流在水分再分配中起着重要作用,能够提高根际区水分供应[55].植物在资源分配上也表现出对干旱环境的适应性,根据环境条件调整地上和地下部分的生物量分配,以优化对光、水和养分等资源的利用[127].干旱条件下植物可能会将更多的资源分配到根系,以增强水分获取能力.同时,植物叶片和细根在碳、氮、磷等元素的分配中也存在协同关系.西北干旱区荒漠植物叶片和细根在碳、氮、磷含量及其计量比上表现出显著的正相关关系[128],这表明植物在完成正常的生命活动时,地上与地下部分的养分分配存在协同作用.深根系植物(如梭梭)主根深入地下水源,地上部分冠层稀疏(减少蒸腾面积),形成高而疏的形态,减少水分竞争.浅根系植物(如沙柳)侧根广泛分布于表层,冠层密集,依赖小降水后对表层水分的快速利用.植物地上和地下部分之间存在信息流和信号传递的协调机制[129].植物可通过木质部和韧皮部传递激素、压力波和电信号等,以协调地上和地下部分的生长和发育.信号传递能够帮助植物快速响应环境变化,如干旱胁迫下气孔调节和根系水分吸收调整.柠条和油蒿细根生长量和死亡量分别受土壤水分和温度共同影响,且不同年代植物对土壤水分利用深度和比例存在差异[53].柠条主要利用60、100 cm的土壤水分,二者占全部水源的75%左右;而油蒿主要利用10、20 cm的土壤水分,二者占全部水源的80%左右.随着固沙植被年限增加,植物对土壤表层水分利用比例逐渐减少,这是植物为了适应深层土壤水分相对稳定环境而做出的生理调整,也反映了植物地上和地下部分在资源获取和分配上的协调性.干旱区植物通过地上与地下部分的形态互补、资源分配动态调整及生理信号联动,实现了水分获取、运输与消耗的高效协调[85].这种根冠协同机制是其适应极端干旱的核心策略,也为人工植被恢复中的物种选择和生态管理提供了理论依据. ...
黑河下游柽柳根系水力提升对林分蒸散的贡献
1
2017
... 固沙灌木地上和地下部分在生长发育过程中存在密切协调关系,是植物适应干旱环境、维持自身稳定的关键[122].干旱区植物根系通常具有深根性和广布性.例如白梭梭和梭梭的垂直根深逾5 m,可至地下水层吸收水分[123].柽柳主根和侧根均极为发达,主根可伸至地下水层,最深10 m以上[124].此外,一些植物如沙拐枣根系多为水平分布,但在地下水较深的地区,其根系也能垂直向下发展以获取水分[125].以上根系的形态和分布使得植物能够有效地利用土壤水分资源.长期干旱时植物通过落叶(如梭梭)、茎干休眠等方式减少地上部分的水分消耗,同时维持根系活力(如沙拐枣根系持续生长寻找水源).植物地上和地下部分在生理功能上也表现为协调关系[126].植物通过冠层截留降水,减少土壤水分蒸发.同时,植物根系能够将截留水分吸收并输送到地上部分,支持植物蒸腾作用和光合作用.固沙灌木冠层截留和树干径流在水分再分配中起着重要作用,能够提高根际区水分供应[55].植物在资源分配上也表现出对干旱环境的适应性,根据环境条件调整地上和地下部分的生物量分配,以优化对光、水和养分等资源的利用[127].干旱条件下植物可能会将更多的资源分配到根系,以增强水分获取能力.同时,植物叶片和细根在碳、氮、磷等元素的分配中也存在协同关系.西北干旱区荒漠植物叶片和细根在碳、氮、磷含量及其计量比上表现出显著的正相关关系[128],这表明植物在完成正常的生命活动时,地上与地下部分的养分分配存在协同作用.深根系植物(如梭梭)主根深入地下水源,地上部分冠层稀疏(减少蒸腾面积),形成高而疏的形态,减少水分竞争.浅根系植物(如沙柳)侧根广泛分布于表层,冠层密集,依赖小降水后对表层水分的快速利用.植物地上和地下部分之间存在信息流和信号传递的协调机制[129].植物可通过木质部和韧皮部传递激素、压力波和电信号等,以协调地上和地下部分的生长和发育.信号传递能够帮助植物快速响应环境变化,如干旱胁迫下气孔调节和根系水分吸收调整.柠条和油蒿细根生长量和死亡量分别受土壤水分和温度共同影响,且不同年代植物对土壤水分利用深度和比例存在差异[53].柠条主要利用60、100 cm的土壤水分,二者占全部水源的75%左右;而油蒿主要利用10、20 cm的土壤水分,二者占全部水源的80%左右.随着固沙植被年限增加,植物对土壤表层水分利用比例逐渐减少,这是植物为了适应深层土壤水分相对稳定环境而做出的生理调整,也反映了植物地上和地下部分在资源获取和分配上的协调性.干旱区植物通过地上与地下部分的形态互补、资源分配动态调整及生理信号联动,实现了水分获取、运输与消耗的高效协调[85].这种根冠协同机制是其适应极端干旱的核心策略,也为人工植被恢复中的物种选择和生态管理提供了理论依据. ...
Root distribution of three dominant desert shrubs and their water uptake dynamics
1
2017
... 固沙灌木地上和地下部分在生长发育过程中存在密切协调关系,是植物适应干旱环境、维持自身稳定的关键[122].干旱区植物根系通常具有深根性和广布性.例如白梭梭和梭梭的垂直根深逾5 m,可至地下水层吸收水分[123].柽柳主根和侧根均极为发达,主根可伸至地下水层,最深10 m以上[124].此外,一些植物如沙拐枣根系多为水平分布,但在地下水较深的地区,其根系也能垂直向下发展以获取水分[125].以上根系的形态和分布使得植物能够有效地利用土壤水分资源.长期干旱时植物通过落叶(如梭梭)、茎干休眠等方式减少地上部分的水分消耗,同时维持根系活力(如沙拐枣根系持续生长寻找水源).植物地上和地下部分在生理功能上也表现为协调关系[126].植物通过冠层截留降水,减少土壤水分蒸发.同时,植物根系能够将截留水分吸收并输送到地上部分,支持植物蒸腾作用和光合作用.固沙灌木冠层截留和树干径流在水分再分配中起着重要作用,能够提高根际区水分供应[55].植物在资源分配上也表现出对干旱环境的适应性,根据环境条件调整地上和地下部分的生物量分配,以优化对光、水和养分等资源的利用[127].干旱条件下植物可能会将更多的资源分配到根系,以增强水分获取能力.同时,植物叶片和细根在碳、氮、磷等元素的分配中也存在协同关系.西北干旱区荒漠植物叶片和细根在碳、氮、磷含量及其计量比上表现出显著的正相关关系[128],这表明植物在完成正常的生命活动时,地上与地下部分的养分分配存在协同作用.深根系植物(如梭梭)主根深入地下水源,地上部分冠层稀疏(减少蒸腾面积),形成高而疏的形态,减少水分竞争.浅根系植物(如沙柳)侧根广泛分布于表层,冠层密集,依赖小降水后对表层水分的快速利用.植物地上和地下部分之间存在信息流和信号传递的协调机制[129].植物可通过木质部和韧皮部传递激素、压力波和电信号等,以协调地上和地下部分的生长和发育.信号传递能够帮助植物快速响应环境变化,如干旱胁迫下气孔调节和根系水分吸收调整.柠条和油蒿细根生长量和死亡量分别受土壤水分和温度共同影响,且不同年代植物对土壤水分利用深度和比例存在差异[53].柠条主要利用60、100 cm的土壤水分,二者占全部水源的75%左右;而油蒿主要利用10、20 cm的土壤水分,二者占全部水源的80%左右.随着固沙植被年限增加,植物对土壤表层水分利用比例逐渐减少,这是植物为了适应深层土壤水分相对稳定环境而做出的生理调整,也反映了植物地上和地下部分在资源获取和分配上的协调性.干旱区植物通过地上与地下部分的形态互补、资源分配动态调整及生理信号联动,实现了水分获取、运输与消耗的高效协调[85].这种根冠协同机制是其适应极端干旱的核心策略,也为人工植被恢复中的物种选择和生态管理提供了理论依据. ...
荒漠植物性状权衡策略及功能多样性研究进展
1
2022
... 固沙灌木地上和地下部分在生长发育过程中存在密切协调关系,是植物适应干旱环境、维持自身稳定的关键[122].干旱区植物根系通常具有深根性和广布性.例如白梭梭和梭梭的垂直根深逾5 m,可至地下水层吸收水分[123].柽柳主根和侧根均极为发达,主根可伸至地下水层,最深10 m以上[124].此外,一些植物如沙拐枣根系多为水平分布,但在地下水较深的地区,其根系也能垂直向下发展以获取水分[125].以上根系的形态和分布使得植物能够有效地利用土壤水分资源.长期干旱时植物通过落叶(如梭梭)、茎干休眠等方式减少地上部分的水分消耗,同时维持根系活力(如沙拐枣根系持续生长寻找水源).植物地上和地下部分在生理功能上也表现为协调关系[126].植物通过冠层截留降水,减少土壤水分蒸发.同时,植物根系能够将截留水分吸收并输送到地上部分,支持植物蒸腾作用和光合作用.固沙灌木冠层截留和树干径流在水分再分配中起着重要作用,能够提高根际区水分供应[55].植物在资源分配上也表现出对干旱环境的适应性,根据环境条件调整地上和地下部分的生物量分配,以优化对光、水和养分等资源的利用[127].干旱条件下植物可能会将更多的资源分配到根系,以增强水分获取能力.同时,植物叶片和细根在碳、氮、磷等元素的分配中也存在协同关系.西北干旱区荒漠植物叶片和细根在碳、氮、磷含量及其计量比上表现出显著的正相关关系[128],这表明植物在完成正常的生命活动时,地上与地下部分的养分分配存在协同作用.深根系植物(如梭梭)主根深入地下水源,地上部分冠层稀疏(减少蒸腾面积),形成高而疏的形态,减少水分竞争.浅根系植物(如沙柳)侧根广泛分布于表层,冠层密集,依赖小降水后对表层水分的快速利用.植物地上和地下部分之间存在信息流和信号传递的协调机制[129].植物可通过木质部和韧皮部传递激素、压力波和电信号等,以协调地上和地下部分的生长和发育.信号传递能够帮助植物快速响应环境变化,如干旱胁迫下气孔调节和根系水分吸收调整.柠条和油蒿细根生长量和死亡量分别受土壤水分和温度共同影响,且不同年代植物对土壤水分利用深度和比例存在差异[53].柠条主要利用60、100 cm的土壤水分,二者占全部水源的75%左右;而油蒿主要利用10、20 cm的土壤水分,二者占全部水源的80%左右.随着固沙植被年限增加,植物对土壤表层水分利用比例逐渐减少,这是植物为了适应深层土壤水分相对稳定环境而做出的生理调整,也反映了植物地上和地下部分在资源获取和分配上的协调性.干旱区植物通过地上与地下部分的形态互补、资源分配动态调整及生理信号联动,实现了水分获取、运输与消耗的高效协调[85].这种根冠协同机制是其适应极端干旱的核心策略,也为人工植被恢复中的物种选择和生态管理提供了理论依据. ...
荒漠草地4种灌木生物量分配特征
1
2013
... 固沙灌木地上和地下部分在生长发育过程中存在密切协调关系,是植物适应干旱环境、维持自身稳定的关键[122].干旱区植物根系通常具有深根性和广布性.例如白梭梭和梭梭的垂直根深逾5 m,可至地下水层吸收水分[123].柽柳主根和侧根均极为发达,主根可伸至地下水层,最深10 m以上[124].此外,一些植物如沙拐枣根系多为水平分布,但在地下水较深的地区,其根系也能垂直向下发展以获取水分[125].以上根系的形态和分布使得植物能够有效地利用土壤水分资源.长期干旱时植物通过落叶(如梭梭)、茎干休眠等方式减少地上部分的水分消耗,同时维持根系活力(如沙拐枣根系持续生长寻找水源).植物地上和地下部分在生理功能上也表现为协调关系[126].植物通过冠层截留降水,减少土壤水分蒸发.同时,植物根系能够将截留水分吸收并输送到地上部分,支持植物蒸腾作用和光合作用.固沙灌木冠层截留和树干径流在水分再分配中起着重要作用,能够提高根际区水分供应[55].植物在资源分配上也表现出对干旱环境的适应性,根据环境条件调整地上和地下部分的生物量分配,以优化对光、水和养分等资源的利用[127].干旱条件下植物可能会将更多的资源分配到根系,以增强水分获取能力.同时,植物叶片和细根在碳、氮、磷等元素的分配中也存在协同关系.西北干旱区荒漠植物叶片和细根在碳、氮、磷含量及其计量比上表现出显著的正相关关系[128],这表明植物在完成正常的生命活动时,地上与地下部分的养分分配存在协同作用.深根系植物(如梭梭)主根深入地下水源,地上部分冠层稀疏(减少蒸腾面积),形成高而疏的形态,减少水分竞争.浅根系植物(如沙柳)侧根广泛分布于表层,冠层密集,依赖小降水后对表层水分的快速利用.植物地上和地下部分之间存在信息流和信号传递的协调机制[129].植物可通过木质部和韧皮部传递激素、压力波和电信号等,以协调地上和地下部分的生长和发育.信号传递能够帮助植物快速响应环境变化,如干旱胁迫下气孔调节和根系水分吸收调整.柠条和油蒿细根生长量和死亡量分别受土壤水分和温度共同影响,且不同年代植物对土壤水分利用深度和比例存在差异[53].柠条主要利用60、100 cm的土壤水分,二者占全部水源的75%左右;而油蒿主要利用10、20 cm的土壤水分,二者占全部水源的80%左右.随着固沙植被年限增加,植物对土壤表层水分利用比例逐渐减少,这是植物为了适应深层土壤水分相对稳定环境而做出的生理调整,也反映了植物地上和地下部分在资源获取和分配上的协调性.干旱区植物通过地上与地下部分的形态互补、资源分配动态调整及生理信号联动,实现了水分获取、运输与消耗的高效协调[85].这种根冠协同机制是其适应极端干旱的核心策略,也为人工植被恢复中的物种选择和生态管理提供了理论依据. ...
西北干旱区荒漠植物叶片和细根碳、氮、磷化学计量特征
1
2024
... 固沙灌木地上和地下部分在生长发育过程中存在密切协调关系,是植物适应干旱环境、维持自身稳定的关键[122].干旱区植物根系通常具有深根性和广布性.例如白梭梭和梭梭的垂直根深逾5 m,可至地下水层吸收水分[123].柽柳主根和侧根均极为发达,主根可伸至地下水层,最深10 m以上[124].此外,一些植物如沙拐枣根系多为水平分布,但在地下水较深的地区,其根系也能垂直向下发展以获取水分[125].以上根系的形态和分布使得植物能够有效地利用土壤水分资源.长期干旱时植物通过落叶(如梭梭)、茎干休眠等方式减少地上部分的水分消耗,同时维持根系活力(如沙拐枣根系持续生长寻找水源).植物地上和地下部分在生理功能上也表现为协调关系[126].植物通过冠层截留降水,减少土壤水分蒸发.同时,植物根系能够将截留水分吸收并输送到地上部分,支持植物蒸腾作用和光合作用.固沙灌木冠层截留和树干径流在水分再分配中起着重要作用,能够提高根际区水分供应[55].植物在资源分配上也表现出对干旱环境的适应性,根据环境条件调整地上和地下部分的生物量分配,以优化对光、水和养分等资源的利用[127].干旱条件下植物可能会将更多的资源分配到根系,以增强水分获取能力.同时,植物叶片和细根在碳、氮、磷等元素的分配中也存在协同关系.西北干旱区荒漠植物叶片和细根在碳、氮、磷含量及其计量比上表现出显著的正相关关系[128],这表明植物在完成正常的生命活动时,地上与地下部分的养分分配存在协同作用.深根系植物(如梭梭)主根深入地下水源,地上部分冠层稀疏(减少蒸腾面积),形成高而疏的形态,减少水分竞争.浅根系植物(如沙柳)侧根广泛分布于表层,冠层密集,依赖小降水后对表层水分的快速利用.植物地上和地下部分之间存在信息流和信号传递的协调机制[129].植物可通过木质部和韧皮部传递激素、压力波和电信号等,以协调地上和地下部分的生长和发育.信号传递能够帮助植物快速响应环境变化,如干旱胁迫下气孔调节和根系水分吸收调整.柠条和油蒿细根生长量和死亡量分别受土壤水分和温度共同影响,且不同年代植物对土壤水分利用深度和比例存在差异[53].柠条主要利用60、100 cm的土壤水分,二者占全部水源的75%左右;而油蒿主要利用10、20 cm的土壤水分,二者占全部水源的80%左右.随着固沙植被年限增加,植物对土壤表层水分利用比例逐渐减少,这是植物为了适应深层土壤水分相对稳定环境而做出的生理调整,也反映了植物地上和地下部分在资源获取和分配上的协调性.干旱区植物通过地上与地下部分的形态互补、资源分配动态调整及生理信号联动,实现了水分获取、运输与消耗的高效协调[85].这种根冠协同机制是其适应极端干旱的核心策略,也为人工植被恢复中的物种选择和生态管理提供了理论依据. ...
Interactions between above and below ground plant structures:mechanisms and ecosystem services
1
2022
... 固沙灌木地上和地下部分在生长发育过程中存在密切协调关系,是植物适应干旱环境、维持自身稳定的关键[122].干旱区植物根系通常具有深根性和广布性.例如白梭梭和梭梭的垂直根深逾5 m,可至地下水层吸收水分[123].柽柳主根和侧根均极为发达,主根可伸至地下水层,最深10 m以上[124].此外,一些植物如沙拐枣根系多为水平分布,但在地下水较深的地区,其根系也能垂直向下发展以获取水分[125].以上根系的形态和分布使得植物能够有效地利用土壤水分资源.长期干旱时植物通过落叶(如梭梭)、茎干休眠等方式减少地上部分的水分消耗,同时维持根系活力(如沙拐枣根系持续生长寻找水源).植物地上和地下部分在生理功能上也表现为协调关系[126].植物通过冠层截留降水,减少土壤水分蒸发.同时,植物根系能够将截留水分吸收并输送到地上部分,支持植物蒸腾作用和光合作用.固沙灌木冠层截留和树干径流在水分再分配中起着重要作用,能够提高根际区水分供应[55].植物在资源分配上也表现出对干旱环境的适应性,根据环境条件调整地上和地下部分的生物量分配,以优化对光、水和养分等资源的利用[127].干旱条件下植物可能会将更多的资源分配到根系,以增强水分获取能力.同时,植物叶片和细根在碳、氮、磷等元素的分配中也存在协同关系.西北干旱区荒漠植物叶片和细根在碳、氮、磷含量及其计量比上表现出显著的正相关关系[128],这表明植物在完成正常的生命活动时,地上与地下部分的养分分配存在协同作用.深根系植物(如梭梭)主根深入地下水源,地上部分冠层稀疏(减少蒸腾面积),形成高而疏的形态,减少水分竞争.浅根系植物(如沙柳)侧根广泛分布于表层,冠层密集,依赖小降水后对表层水分的快速利用.植物地上和地下部分之间存在信息流和信号传递的协调机制[129].植物可通过木质部和韧皮部传递激素、压力波和电信号等,以协调地上和地下部分的生长和发育.信号传递能够帮助植物快速响应环境变化,如干旱胁迫下气孔调节和根系水分吸收调整.柠条和油蒿细根生长量和死亡量分别受土壤水分和温度共同影响,且不同年代植物对土壤水分利用深度和比例存在差异[53].柠条主要利用60、100 cm的土壤水分,二者占全部水源的75%左右;而油蒿主要利用10、20 cm的土壤水分,二者占全部水源的80%左右.随着固沙植被年限增加,植物对土壤表层水分利用比例逐渐减少,这是植物为了适应深层土壤水分相对稳定环境而做出的生理调整,也反映了植物地上和地下部分在资源获取和分配上的协调性.干旱区植物通过地上与地下部分的形态互补、资源分配动态调整及生理信号联动,实现了水分获取、运输与消耗的高效协调[85].这种根冠协同机制是其适应极端干旱的核心策略,也为人工植被恢复中的物种选择和生态管理提供了理论依据. ...
干旱半干旱地区人工固沙灌木林生态系统演变特征
1
2005
... 植被和土壤之间协同演替关系是沙漠生态系统稳定性的基础[2].腾格里沙漠随着固沙植被建立,植被在从人工系统向自然系统的演变中经历了覆盖度从增加到减少的自疏过程、从灌木到半灌木再到草本的变化,土壤从流动风沙土向钙积旱成土发育[130].固沙灌木与土壤之间存在着复杂的互馈关系,这种关系在维持生态系统稳定性方面发挥着关键作用.一方面,固沙植物通过根系生长、凋落物分解等过程影响土壤的物理和化学性质[19,131].例如植物根系在生长过程中分泌有机物质,增加土壤有机碳含量,改善土壤结构和保水能力[132];同时,根系周转和凋落物分解也为土壤提供丰富养分,促进土壤肥力提升.另一方面,土壤水分和养分状况又反作用于植物的生长和发育.适宜的土壤水分条件能够促进植物根系扩展和吸收功能,提高植物水分利用效率和抗旱能力;而土壤养分则直接影响植物生长速度、生物量积累以及繁殖能力[133].不同演替阶段柠条的水分利用特征表明:演替早期,土壤水分含量决定柠条的水分利用来源,演替中后期,其决定因子分别为植物的基径和土壤黏粉粒含量[134]. ...
沙坡头地区凋落物分解及其对土壤微生物群落的影响
1
2022
... 植被和土壤之间协同演替关系是沙漠生态系统稳定性的基础[2].腾格里沙漠随着固沙植被建立,植被在从人工系统向自然系统的演变中经历了覆盖度从增加到减少的自疏过程、从灌木到半灌木再到草本的变化,土壤从流动风沙土向钙积旱成土发育[130].固沙灌木与土壤之间存在着复杂的互馈关系,这种关系在维持生态系统稳定性方面发挥着关键作用.一方面,固沙植物通过根系生长、凋落物分解等过程影响土壤的物理和化学性质[19,131].例如植物根系在生长过程中分泌有机物质,增加土壤有机碳含量,改善土壤结构和保水能力[132];同时,根系周转和凋落物分解也为土壤提供丰富养分,促进土壤肥力提升.另一方面,土壤水分和养分状况又反作用于植物的生长和发育.适宜的土壤水分条件能够促进植物根系扩展和吸收功能,提高植物水分利用效率和抗旱能力;而土壤养分则直接影响植物生长速度、生物量积累以及繁殖能力[133].不同演替阶段柠条的水分利用特征表明:演替早期,土壤水分含量决定柠条的水分利用来源,演替中后期,其决定因子分别为植物的基径和土壤黏粉粒含量[134]. ...
Selecting suitable shrub and herb species to revegetation from the perspective of root exudates:an implication for ecological restoration of desertification in an alpine meadow of the Eastern Tibetan Plateau
1
2022
... 植被和土壤之间协同演替关系是沙漠生态系统稳定性的基础[2].腾格里沙漠随着固沙植被建立,植被在从人工系统向自然系统的演变中经历了覆盖度从增加到减少的自疏过程、从灌木到半灌木再到草本的变化,土壤从流动风沙土向钙积旱成土发育[130].固沙灌木与土壤之间存在着复杂的互馈关系,这种关系在维持生态系统稳定性方面发挥着关键作用.一方面,固沙植物通过根系生长、凋落物分解等过程影响土壤的物理和化学性质[19,131].例如植物根系在生长过程中分泌有机物质,增加土壤有机碳含量,改善土壤结构和保水能力[132];同时,根系周转和凋落物分解也为土壤提供丰富养分,促进土壤肥力提升.另一方面,土壤水分和养分状况又反作用于植物的生长和发育.适宜的土壤水分条件能够促进植物根系扩展和吸收功能,提高植物水分利用效率和抗旱能力;而土壤养分则直接影响植物生长速度、生物量积累以及繁殖能力[133].不同演替阶段柠条的水分利用特征表明:演替早期,土壤水分含量决定柠条的水分利用来源,演替中后期,其决定因子分别为植物的基径和土壤黏粉粒含量[134]. ...
腾格里沙漠人工固沙植被中油蒿的生长及生物量分配动态
1
2009
... 植被和土壤之间协同演替关系是沙漠生态系统稳定性的基础[2].腾格里沙漠随着固沙植被建立,植被在从人工系统向自然系统的演变中经历了覆盖度从增加到减少的自疏过程、从灌木到半灌木再到草本的变化,土壤从流动风沙土向钙积旱成土发育[130].固沙灌木与土壤之间存在着复杂的互馈关系,这种关系在维持生态系统稳定性方面发挥着关键作用.一方面,固沙植物通过根系生长、凋落物分解等过程影响土壤的物理和化学性质[19,131].例如植物根系在生长过程中分泌有机物质,增加土壤有机碳含量,改善土壤结构和保水能力[132];同时,根系周转和凋落物分解也为土壤提供丰富养分,促进土壤肥力提升.另一方面,土壤水分和养分状况又反作用于植物的生长和发育.适宜的土壤水分条件能够促进植物根系扩展和吸收功能,提高植物水分利用效率和抗旱能力;而土壤养分则直接影响植物生长速度、生物量积累以及繁殖能力[133].不同演替阶段柠条的水分利用特征表明:演替早期,土壤水分含量决定柠条的水分利用来源,演替中后期,其决定因子分别为植物的基径和土壤黏粉粒含量[134]. ...
Vegetation and soil succession determine the water-use strategy of Caragana korshinskii in a chronosequence of revegetated areas in Ningxia,Northwest China
1
2022
... 植被和土壤之间协同演替关系是沙漠生态系统稳定性的基础[2].腾格里沙漠随着固沙植被建立,植被在从人工系统向自然系统的演变中经历了覆盖度从增加到减少的自疏过程、从灌木到半灌木再到草本的变化,土壤从流动风沙土向钙积旱成土发育[130].固沙灌木与土壤之间存在着复杂的互馈关系,这种关系在维持生态系统稳定性方面发挥着关键作用.一方面,固沙植物通过根系生长、凋落物分解等过程影响土壤的物理和化学性质[19,131].例如植物根系在生长过程中分泌有机物质,增加土壤有机碳含量,改善土壤结构和保水能力[132];同时,根系周转和凋落物分解也为土壤提供丰富养分,促进土壤肥力提升.另一方面,土壤水分和养分状况又反作用于植物的生长和发育.适宜的土壤水分条件能够促进植物根系扩展和吸收功能,提高植物水分利用效率和抗旱能力;而土壤养分则直接影响植物生长速度、生物量积累以及繁殖能力[133].不同演替阶段柠条的水分利用特征表明:演替早期,土壤水分含量决定柠条的水分利用来源,演替中后期,其决定因子分别为植物的基径和土壤黏粉粒含量[134]. ...



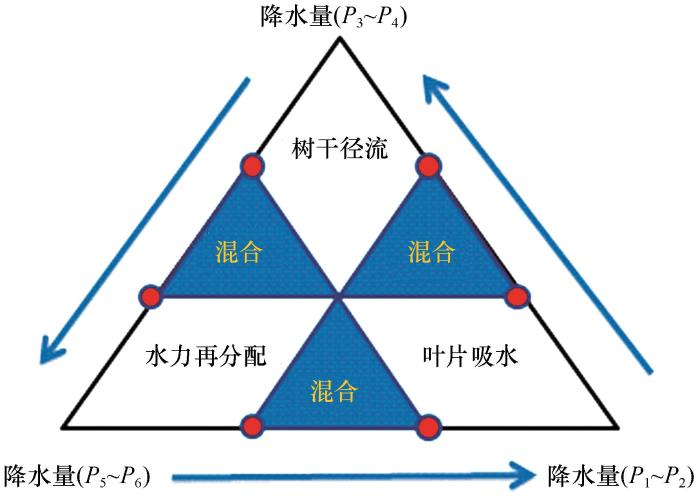


 甘公网安备 62010202000688号
甘公网安备 62010202000688号
{kind=link}
{kind=link}
{kind=link}
{kind=link}