Grassland soil carbon sequestration:current understanding,challenges,and solutions
1
2022
... 草地是最大的陆地生态系统,具有固碳、水土保持、维持生物多样性、促进畜牧业发展和提供文化服务等多种重要生态功能[1].中国草地面积约4亿hm2,约占国土面积的42%[2].然而,由于气候变化(如干旱化)和不合理土地利用[3],草地沙漠化是中国乃至全球旱区面临的重要生态环境问题.据估计,约2.5亿发展中国家人口直接受到沙漠化的影响[4].因此,世界许多国家致力于沙漠化防治,如何恢复沙漠化草地,并维持草地生态系统稳定性成为当前面临的重要问题. ...
1
1996
... 草地是最大的陆地生态系统,具有固碳、水土保持、维持生物多样性、促进畜牧业发展和提供文化服务等多种重要生态功能[1].中国草地面积约4亿hm2,约占国土面积的42%[2].然而,由于气候变化(如干旱化)和不合理土地利用[3],草地沙漠化是中国乃至全球旱区面临的重要生态环境问题.据估计,约2.5亿发展中国家人口直接受到沙漠化的影响[4].因此,世界许多国家致力于沙漠化防治,如何恢复沙漠化草地,并维持草地生态系统稳定性成为当前面临的重要问题. ...
Global desertification:drivers and feedbacks
1
2013
... 草地是最大的陆地生态系统,具有固碳、水土保持、维持生物多样性、促进畜牧业发展和提供文化服务等多种重要生态功能[1].中国草地面积约4亿hm2,约占国土面积的42%[2].然而,由于气候变化(如干旱化)和不合理土地利用[3],草地沙漠化是中国乃至全球旱区面临的重要生态环境问题.据估计,约2.5亿发展中国家人口直接受到沙漠化的影响[4].因此,世界许多国家致力于沙漠化防治,如何恢复沙漠化草地,并维持草地生态系统稳定性成为当前面临的重要问题. ...
Global desertification:building a science for dryland development
1
2007
... 草地是最大的陆地生态系统,具有固碳、水土保持、维持生物多样性、促进畜牧业发展和提供文化服务等多种重要生态功能[1].中国草地面积约4亿hm2,约占国土面积的42%[2].然而,由于气候变化(如干旱化)和不合理土地利用[3],草地沙漠化是中国乃至全球旱区面临的重要生态环境问题.据估计,约2.5亿发展中国家人口直接受到沙漠化的影响[4].因此,世界许多国家致力于沙漠化防治,如何恢复沙漠化草地,并维持草地生态系统稳定性成为当前面临的重要问题. ...
Grazing exclusion effect on soil and vegetation properties in Imam Kandi Rangelands,Iran
1
2013
... 围封通过排除人类活动干扰,依靠自然演替和更新恢复受损生态系统,具有效率高、成本低、已恢复生态系统稳定性强、生态系统服务高等优势,是退化草地恢复广泛采用的方法.现有研究表明围封有利于植被群落恢复演替、提高土壤肥力[5].围封使退化典型草原[6]、荒漠草原[7]、沙质草地[8]、高寒草地[9]植物群落高度、地上生物量显著增加.但对物种多样性的影响则存在差异,这可能与草地类型、气候、生物地理格局、退化程度等存在差异有关[10].例如,短期(6年)围封显著增加轻度、中度退化荒漠草原的物种多样性,但重度退化荒漠草原围封后物种多样性降低[11].围封未改变荒漠草原优势建群种的优势度[12];但在典型草原中物种多样性随围封年限先降后增,群落优势物种依次沿杂类草、多年生禾草、杂类草3个阶段演替[6].与放牧相比,围封10年沙质草地可显著增加物种多样性、增强群落稳定性,并改善群落结构和功能[8];但围封20年后物种多样性与不同程度放牧样地差异不显著[13].严重沙漠化草地围封恢复主要有3个阶段:①狗尾草、马唐(Digitaria sanguinalis)等定殖与消失,②多年生根茎禾草为优势物种,③多年生丛生禾草为优势物种[14].尽管大量研究报道了围封对植物群落的影响,且围封年限对植物群落和土壤肥力恢复产生重要影响[15],但针对严重沙漠化草地长期围封对植物群落的影响研究有限,其驱动机制仍不清楚. ...
温带草原围封对植物群落的影响
2
2023
... 围封通过排除人类活动干扰,依靠自然演替和更新恢复受损生态系统,具有效率高、成本低、已恢复生态系统稳定性强、生态系统服务高等优势,是退化草地恢复广泛采用的方法.现有研究表明围封有利于植被群落恢复演替、提高土壤肥力[5].围封使退化典型草原[6]、荒漠草原[7]、沙质草地[8]、高寒草地[9]植物群落高度、地上生物量显著增加.但对物种多样性的影响则存在差异,这可能与草地类型、气候、生物地理格局、退化程度等存在差异有关[10].例如,短期(6年)围封显著增加轻度、中度退化荒漠草原的物种多样性,但重度退化荒漠草原围封后物种多样性降低[11].围封未改变荒漠草原优势建群种的优势度[12];但在典型草原中物种多样性随围封年限先降后增,群落优势物种依次沿杂类草、多年生禾草、杂类草3个阶段演替[6].与放牧相比,围封10年沙质草地可显著增加物种多样性、增强群落稳定性,并改善群落结构和功能[8];但围封20年后物种多样性与不同程度放牧样地差异不显著[13].严重沙漠化草地围封恢复主要有3个阶段:①狗尾草、马唐(Digitaria sanguinalis)等定殖与消失,②多年生根茎禾草为优势物种,③多年生丛生禾草为优势物种[14].尽管大量研究报道了围封对植物群落的影响,且围封年限对植物群落和土壤肥力恢复产生重要影响[15],但针对严重沙漠化草地长期围封对植物群落的影响研究有限,其驱动机制仍不清楚. ...
... [6].与放牧相比,围封10年沙质草地可显著增加物种多样性、增强群落稳定性,并改善群落结构和功能[8];但围封20年后物种多样性与不同程度放牧样地差异不显著[13].严重沙漠化草地围封恢复主要有3个阶段:①狗尾草、马唐(Digitaria sanguinalis)等定殖与消失,②多年生根茎禾草为优势物种,③多年生丛生禾草为优势物种[14].尽管大量研究报道了围封对植物群落的影响,且围封年限对植物群落和土壤肥力恢复产生重要影响[15],但针对严重沙漠化草地长期围封对植物群落的影响研究有限,其驱动机制仍不清楚. ...
围封10年对荒漠草原群落物种多样性与优势种空间分布格局的影响
1
2024
... 围封通过排除人类活动干扰,依靠自然演替和更新恢复受损生态系统,具有效率高、成本低、已恢复生态系统稳定性强、生态系统服务高等优势,是退化草地恢复广泛采用的方法.现有研究表明围封有利于植被群落恢复演替、提高土壤肥力[5].围封使退化典型草原[6]、荒漠草原[7]、沙质草地[8]、高寒草地[9]植物群落高度、地上生物量显著增加.但对物种多样性的影响则存在差异,这可能与草地类型、气候、生物地理格局、退化程度等存在差异有关[10].例如,短期(6年)围封显著增加轻度、中度退化荒漠草原的物种多样性,但重度退化荒漠草原围封后物种多样性降低[11].围封未改变荒漠草原优势建群种的优势度[12];但在典型草原中物种多样性随围封年限先降后增,群落优势物种依次沿杂类草、多年生禾草、杂类草3个阶段演替[6].与放牧相比,围封10年沙质草地可显著增加物种多样性、增强群落稳定性,并改善群落结构和功能[8];但围封20年后物种多样性与不同程度放牧样地差异不显著[13].严重沙漠化草地围封恢复主要有3个阶段:①狗尾草、马唐(Digitaria sanguinalis)等定殖与消失,②多年生根茎禾草为优势物种,③多年生丛生禾草为优势物种[14].尽管大量研究报道了围封对植物群落的影响,且围封年限对植物群落和土壤肥力恢复产生重要影响[15],但针对严重沙漠化草地长期围封对植物群落的影响研究有限,其驱动机制仍不清楚. ...
围封对退化沙质草地植物群落的影响
4
2018
... 围封通过排除人类活动干扰,依靠自然演替和更新恢复受损生态系统,具有效率高、成本低、已恢复生态系统稳定性强、生态系统服务高等优势,是退化草地恢复广泛采用的方法.现有研究表明围封有利于植被群落恢复演替、提高土壤肥力[5].围封使退化典型草原[6]、荒漠草原[7]、沙质草地[8]、高寒草地[9]植物群落高度、地上生物量显著增加.但对物种多样性的影响则存在差异,这可能与草地类型、气候、生物地理格局、退化程度等存在差异有关[10].例如,短期(6年)围封显著增加轻度、中度退化荒漠草原的物种多样性,但重度退化荒漠草原围封后物种多样性降低[11].围封未改变荒漠草原优势建群种的优势度[12];但在典型草原中物种多样性随围封年限先降后增,群落优势物种依次沿杂类草、多年生禾草、杂类草3个阶段演替[6].与放牧相比,围封10年沙质草地可显著增加物种多样性、增强群落稳定性,并改善群落结构和功能[8];但围封20年后物种多样性与不同程度放牧样地差异不显著[13].严重沙漠化草地围封恢复主要有3个阶段:①狗尾草、马唐(Digitaria sanguinalis)等定殖与消失,②多年生根茎禾草为优势物种,③多年生丛生禾草为优势物种[14].尽管大量研究报道了围封对植物群落的影响,且围封年限对植物群落和土壤肥力恢复产生重要影响[15],但针对严重沙漠化草地长期围封对植物群落的影响研究有限,其驱动机制仍不清楚. ...
... [8];但围封20年后物种多样性与不同程度放牧样地差异不显著[13].严重沙漠化草地围封恢复主要有3个阶段:①狗尾草、马唐(Digitaria sanguinalis)等定殖与消失,②多年生根茎禾草为优势物种,③多年生丛生禾草为优势物种[14].尽管大量研究报道了围封对植物群落的影响,且围封年限对植物群落和土壤肥力恢复产生重要影响[15],但针对严重沙漠化草地长期围封对植物群落的影响研究有限,其驱动机制仍不清楚. ...
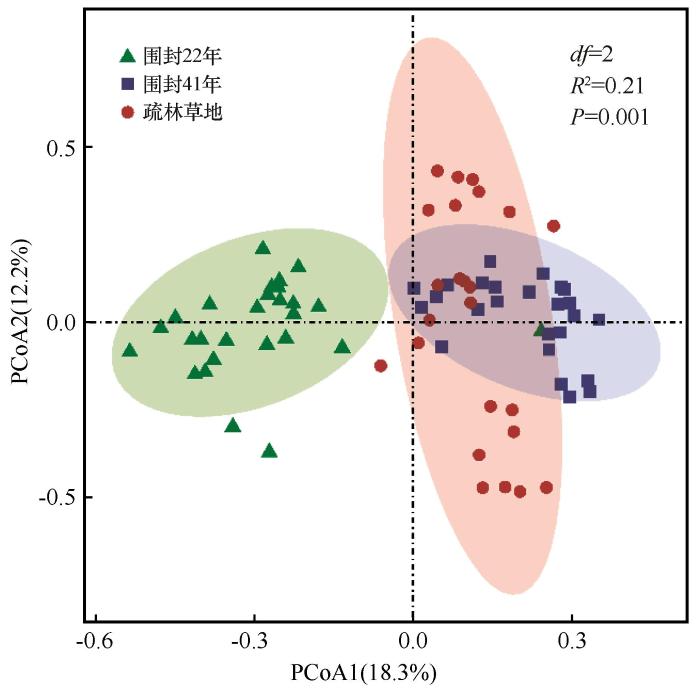
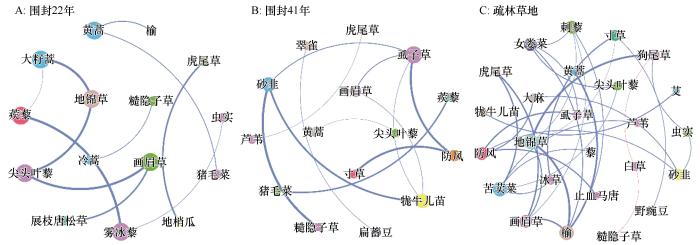
... 共现网络分析表明,围封后植物群落共现模式发生明显改变(图5).随围封年限增加,尽管网络节点数未发生改变,但平均连通度增加,表明植物群落网络结构复杂度增加[24].上述结果与先前报道的长期围封使植物群落组成多样化、复杂化的结果一致[8].关键种与群落其他物种的相互作用最为强烈,对群落稳定性、多样性具有重要作用.网络分析结果表明围封后植物物种间以正相关关系为主(图5),表明关键种对其他物种的分布和数量具有积极影响,从而指示群落演替[25].与优势物种不同,关键种以矮小杂类草(如虱子草、牻牛儿苗、砂韭)为主,主要是由于随围封年限增加,植物群落盖度、高度均显著增加(图2),以上物种在空间、光照不足的条件下更具优势,从而占据不同的生态位,并削弱种间竞争,实现群落结构的稳定. ...
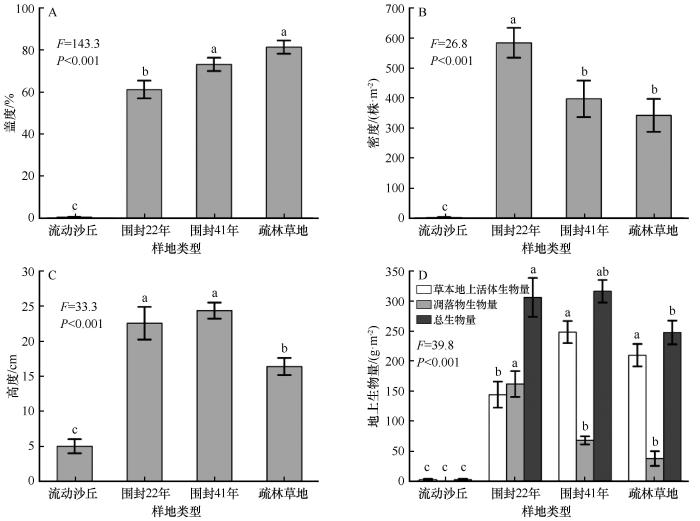
... 现有研究表明,荒漠草地的最佳围封年限为9~12 a[26].但本研究结果与此年限相差甚远.在科尔沁沙地,严重沙漠化草地围封41年后,尽管植物群落数量特征与疏林草地接近,地上生物量、生物多样性甚至高于后者,但植物群落结构、优势物种组成、网络关系与疏林草地相比仍存在较大差异(图4~5).长期围封后,优势植株个体生长高大,占据主要生存空间,加剧种间竞争,限制矮小植株生长[14].相反,一些研究报道了放牧反而有利于缓和竞争、促进种群密度增加[27-29].另一方面,土壤结构改善和肥力恢复对植被恢复至关重要,但现有研究表明,土壤养分恢复通常需要数十年甚至上百年的时间[19,30].这表明严重沙漠化土地植被自然恢复过程漫长且缓慢.尽管很难将流动沙丘恢复到与疏林草地相当的状况,但进一步确定可持续围封的最佳年限是有必要的,围封时间过长会引发对草地生态系统可持续的担忧[8,28]. ...
围栏封育对藏北高寒草地植物多样性与生态系统多功能性的影响
1
2025
... 围封通过排除人类活动干扰,依靠自然演替和更新恢复受损生态系统,具有效率高、成本低、已恢复生态系统稳定性强、生态系统服务高等优势,是退化草地恢复广泛采用的方法.现有研究表明围封有利于植被群落恢复演替、提高土壤肥力[5].围封使退化典型草原[6]、荒漠草原[7]、沙质草地[8]、高寒草地[9]植物群落高度、地上生物量显著增加.但对物种多样性的影响则存在差异,这可能与草地类型、气候、生物地理格局、退化程度等存在差异有关[10].例如,短期(6年)围封显著增加轻度、中度退化荒漠草原的物种多样性,但重度退化荒漠草原围封后物种多样性降低[11].围封未改变荒漠草原优势建群种的优势度[12];但在典型草原中物种多样性随围封年限先降后增,群落优势物种依次沿杂类草、多年生禾草、杂类草3个阶段演替[6].与放牧相比,围封10年沙质草地可显著增加物种多样性、增强群落稳定性,并改善群落结构和功能[8];但围封20年后物种多样性与不同程度放牧样地差异不显著[13].严重沙漠化草地围封恢复主要有3个阶段:①狗尾草、马唐(Digitaria sanguinalis)等定殖与消失,②多年生根茎禾草为优势物种,③多年生丛生禾草为优势物种[14].尽管大量研究报道了围封对植物群落的影响,且围封年限对植物群落和土壤肥力恢复产生重要影响[15],但针对严重沙漠化草地长期围封对植物群落的影响研究有限,其驱动机制仍不清楚. ...
Effects of grazing exclusion on vegetation community characteristics over 22 years in the Zoige alpine meadows from China
1
2023
... 围封通过排除人类活动干扰,依靠自然演替和更新恢复受损生态系统,具有效率高、成本低、已恢复生态系统稳定性强、生态系统服务高等优势,是退化草地恢复广泛采用的方法.现有研究表明围封有利于植被群落恢复演替、提高土壤肥力[5].围封使退化典型草原[6]、荒漠草原[7]、沙质草地[8]、高寒草地[9]植物群落高度、地上生物量显著增加.但对物种多样性的影响则存在差异,这可能与草地类型、气候、生物地理格局、退化程度等存在差异有关[10].例如,短期(6年)围封显著增加轻度、中度退化荒漠草原的物种多样性,但重度退化荒漠草原围封后物种多样性降低[11].围封未改变荒漠草原优势建群种的优势度[12];但在典型草原中物种多样性随围封年限先降后增,群落优势物种依次沿杂类草、多年生禾草、杂类草3个阶段演替[6].与放牧相比,围封10年沙质草地可显著增加物种多样性、增强群落稳定性,并改善群落结构和功能[8];但围封20年后物种多样性与不同程度放牧样地差异不显著[13].严重沙漠化草地围封恢复主要有3个阶段:①狗尾草、马唐(Digitaria sanguinalis)等定殖与消失,②多年生根茎禾草为优势物种,③多年生丛生禾草为优势物种[14].尽管大量研究报道了围封对植物群落的影响,且围封年限对植物群落和土壤肥力恢复产生重要影响[15],但针对严重沙漠化草地长期围封对植物群落的影响研究有限,其驱动机制仍不清楚. ...
围封对不同退化程度荒漠草原植物群落和土壤的影响
1
2023
... 围封通过排除人类活动干扰,依靠自然演替和更新恢复受损生态系统,具有效率高、成本低、已恢复生态系统稳定性强、生态系统服务高等优势,是退化草地恢复广泛采用的方法.现有研究表明围封有利于植被群落恢复演替、提高土壤肥力[5].围封使退化典型草原[6]、荒漠草原[7]、沙质草地[8]、高寒草地[9]植物群落高度、地上生物量显著增加.但对物种多样性的影响则存在差异,这可能与草地类型、气候、生物地理格局、退化程度等存在差异有关[10].例如,短期(6年)围封显著增加轻度、中度退化荒漠草原的物种多样性,但重度退化荒漠草原围封后物种多样性降低[11].围封未改变荒漠草原优势建群种的优势度[12];但在典型草原中物种多样性随围封年限先降后增,群落优势物种依次沿杂类草、多年生禾草、杂类草3个阶段演替[6].与放牧相比,围封10年沙质草地可显著增加物种多样性、增强群落稳定性,并改善群落结构和功能[8];但围封20年后物种多样性与不同程度放牧样地差异不显著[13].严重沙漠化草地围封恢复主要有3个阶段:①狗尾草、马唐(Digitaria sanguinalis)等定殖与消失,②多年生根茎禾草为优势物种,③多年生丛生禾草为优势物种[14].尽管大量研究报道了围封对植物群落的影响,且围封年限对植物群落和土壤肥力恢复产生重要影响[15],但针对严重沙漠化草地长期围封对植物群落的影响研究有限,其驱动机制仍不清楚. ...
围封对荒漠草原典型植物群落特征及多样性的影响
2
2025
... 围封通过排除人类活动干扰,依靠自然演替和更新恢复受损生态系统,具有效率高、成本低、已恢复生态系统稳定性强、生态系统服务高等优势,是退化草地恢复广泛采用的方法.现有研究表明围封有利于植被群落恢复演替、提高土壤肥力[5].围封使退化典型草原[6]、荒漠草原[7]、沙质草地[8]、高寒草地[9]植物群落高度、地上生物量显著增加.但对物种多样性的影响则存在差异,这可能与草地类型、气候、生物地理格局、退化程度等存在差异有关[10].例如,短期(6年)围封显著增加轻度、中度退化荒漠草原的物种多样性,但重度退化荒漠草原围封后物种多样性降低[11].围封未改变荒漠草原优势建群种的优势度[12];但在典型草原中物种多样性随围封年限先降后增,群落优势物种依次沿杂类草、多年生禾草、杂类草3个阶段演替[6].与放牧相比,围封10年沙质草地可显著增加物种多样性、增强群落稳定性,并改善群落结构和功能[8];但围封20年后物种多样性与不同程度放牧样地差异不显著[13].严重沙漠化草地围封恢复主要有3个阶段:①狗尾草、马唐(Digitaria sanguinalis)等定殖与消失,②多年生根茎禾草为优势物种,③多年生丛生禾草为优势物种[14].尽管大量研究报道了围封对植物群落的影响,且围封年限对植物群落和土壤肥力恢复产生重要影响[15],但针对严重沙漠化草地长期围封对植物群落的影响研究有限,其驱动机制仍不清楚. ...
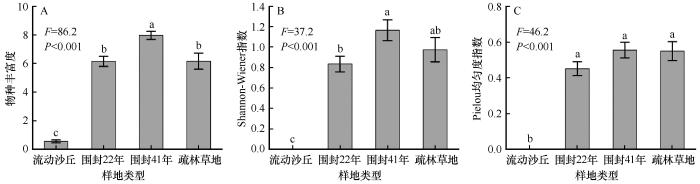
... 长期围封使植物群落由一年生草本沙蓬逐渐向以狗尾草为主的一年生草本植物(围封22年)和以多年生根茎性、丛生性禾草和蒿属植物为主(围封41年)的多年生草本植物群落变化.草本植物物种多样性和生物量也显著增加,甚至略高于疏林草地,这与先前观测到的结果基本一致[13-14],而与沙漠化过程相反[20].围封在一定程度上排除了人类活动干扰,一年生植物大多属于机会主义者,具有较高的水分和养分利用效率,容易在恢复早期形成竞争优势[13,21].随围封进行,多年生植物逐渐定殖,一方面植被盖度和养分输入增加,土壤侵蚀和蒸发减少,为多年生植物生长创造了条件[22].另一方面,由于其深根性[13],可以从深层土壤中获取养分和水分,从而更具优势.与王益涛等[12]报道的围封恢复后优势物种无变化的结果不同,在本研究中,随围封年限增加,植物群落优势物种由一年生逐渐向多年生植物转变,且围封41年后优势程度相当的物种数量增加.这可能与围封年限和恢复的起始对照(即退化程度)有关,前者仅围封4年.相反,本研究未观察到其他研究[23]报道的退化草地围封后灌木优势度明显增加的结果. ...
放牧强度对科尔沁沙地沙质草地植被的影响
4
2016
... 围封通过排除人类活动干扰,依靠自然演替和更新恢复受损生态系统,具有效率高、成本低、已恢复生态系统稳定性强、生态系统服务高等优势,是退化草地恢复广泛采用的方法.现有研究表明围封有利于植被群落恢复演替、提高土壤肥力[5].围封使退化典型草原[6]、荒漠草原[7]、沙质草地[8]、高寒草地[9]植物群落高度、地上生物量显著增加.但对物种多样性的影响则存在差异,这可能与草地类型、气候、生物地理格局、退化程度等存在差异有关[10].例如,短期(6年)围封显著增加轻度、中度退化荒漠草原的物种多样性,但重度退化荒漠草原围封后物种多样性降低[11].围封未改变荒漠草原优势建群种的优势度[12];但在典型草原中物种多样性随围封年限先降后增,群落优势物种依次沿杂类草、多年生禾草、杂类草3个阶段演替[6].与放牧相比,围封10年沙质草地可显著增加物种多样性、增强群落稳定性,并改善群落结构和功能[8];但围封20年后物种多样性与不同程度放牧样地差异不显著[13].严重沙漠化草地围封恢复主要有3个阶段:①狗尾草、马唐(Digitaria sanguinalis)等定殖与消失,②多年生根茎禾草为优势物种,③多年生丛生禾草为优势物种[14].尽管大量研究报道了围封对植物群落的影响,且围封年限对植物群落和土壤肥力恢复产生重要影响[15],但针对严重沙漠化草地长期围封对植物群落的影响研究有限,其驱动机制仍不清楚. ...
... 长期围封使植物群落由一年生草本沙蓬逐渐向以狗尾草为主的一年生草本植物(围封22年)和以多年生根茎性、丛生性禾草和蒿属植物为主(围封41年)的多年生草本植物群落变化.草本植物物种多样性和生物量也显著增加,甚至略高于疏林草地,这与先前观测到的结果基本一致[13-14],而与沙漠化过程相反[20].围封在一定程度上排除了人类活动干扰,一年生植物大多属于机会主义者,具有较高的水分和养分利用效率,容易在恢复早期形成竞争优势[13,21].随围封进行,多年生植物逐渐定殖,一方面植被盖度和养分输入增加,土壤侵蚀和蒸发减少,为多年生植物生长创造了条件[22].另一方面,由于其深根性[13],可以从深层土壤中获取养分和水分,从而更具优势.与王益涛等[12]报道的围封恢复后优势物种无变化的结果不同,在本研究中,随围封年限增加,植物群落优势物种由一年生逐渐向多年生植物转变,且围封41年后优势程度相当的物种数量增加.这可能与围封年限和恢复的起始对照(即退化程度)有关,前者仅围封4年.相反,本研究未观察到其他研究[23]报道的退化草地围封后灌木优势度明显增加的结果. ...
... [13,21].随围封进行,多年生植物逐渐定殖,一方面植被盖度和养分输入增加,土壤侵蚀和蒸发减少,为多年生植物生长创造了条件[22].另一方面,由于其深根性[13],可以从深层土壤中获取养分和水分,从而更具优势.与王益涛等[12]报道的围封恢复后优势物种无变化的结果不同,在本研究中,随围封年限增加,植物群落优势物种由一年生逐渐向多年生植物转变,且围封41年后优势程度相当的物种数量增加.这可能与围封年限和恢复的起始对照(即退化程度)有关,前者仅围封4年.相反,本研究未观察到其他研究[23]报道的退化草地围封后灌木优势度明显增加的结果. ...
... [13],可以从深层土壤中获取养分和水分,从而更具优势.与王益涛等[12]报道的围封恢复后优势物种无变化的结果不同,在本研究中,随围封年限增加,植物群落优势物种由一年生逐渐向多年生植物转变,且围封41年后优势程度相当的物种数量增加.这可能与围封年限和恢复的起始对照(即退化程度)有关,前者仅围封4年.相反,本研究未观察到其他研究[23]报道的退化草地围封后灌木优势度明显增加的结果. ...
我国北方沙漠化生态系统修复案例
3
2024
... 围封通过排除人类活动干扰,依靠自然演替和更新恢复受损生态系统,具有效率高、成本低、已恢复生态系统稳定性强、生态系统服务高等优势,是退化草地恢复广泛采用的方法.现有研究表明围封有利于植被群落恢复演替、提高土壤肥力[5].围封使退化典型草原[6]、荒漠草原[7]、沙质草地[8]、高寒草地[9]植物群落高度、地上生物量显著增加.但对物种多样性的影响则存在差异,这可能与草地类型、气候、生物地理格局、退化程度等存在差异有关[10].例如,短期(6年)围封显著增加轻度、中度退化荒漠草原的物种多样性,但重度退化荒漠草原围封后物种多样性降低[11].围封未改变荒漠草原优势建群种的优势度[12];但在典型草原中物种多样性随围封年限先降后增,群落优势物种依次沿杂类草、多年生禾草、杂类草3个阶段演替[6].与放牧相比,围封10年沙质草地可显著增加物种多样性、增强群落稳定性,并改善群落结构和功能[8];但围封20年后物种多样性与不同程度放牧样地差异不显著[13].严重沙漠化草地围封恢复主要有3个阶段:①狗尾草、马唐(Digitaria sanguinalis)等定殖与消失,②多年生根茎禾草为优势物种,③多年生丛生禾草为优势物种[14].尽管大量研究报道了围封对植物群落的影响,且围封年限对植物群落和土壤肥力恢复产生重要影响[15],但针对严重沙漠化草地长期围封对植物群落的影响研究有限,其驱动机制仍不清楚. ...
... 长期围封使植物群落由一年生草本沙蓬逐渐向以狗尾草为主的一年生草本植物(围封22年)和以多年生根茎性、丛生性禾草和蒿属植物为主(围封41年)的多年生草本植物群落变化.草本植物物种多样性和生物量也显著增加,甚至略高于疏林草地,这与先前观测到的结果基本一致[13-14],而与沙漠化过程相反[20].围封在一定程度上排除了人类活动干扰,一年生植物大多属于机会主义者,具有较高的水分和养分利用效率,容易在恢复早期形成竞争优势[13,21].随围封进行,多年生植物逐渐定殖,一方面植被盖度和养分输入增加,土壤侵蚀和蒸发减少,为多年生植物生长创造了条件[22].另一方面,由于其深根性[13],可以从深层土壤中获取养分和水分,从而更具优势.与王益涛等[12]报道的围封恢复后优势物种无变化的结果不同,在本研究中,随围封年限增加,植物群落优势物种由一年生逐渐向多年生植物转变,且围封41年后优势程度相当的物种数量增加.这可能与围封年限和恢复的起始对照(即退化程度)有关,前者仅围封4年.相反,本研究未观察到其他研究[23]报道的退化草地围封后灌木优势度明显增加的结果. ...
... 现有研究表明,荒漠草地的最佳围封年限为9~12 a[26].但本研究结果与此年限相差甚远.在科尔沁沙地,严重沙漠化草地围封41年后,尽管植物群落数量特征与疏林草地接近,地上生物量、生物多样性甚至高于后者,但植物群落结构、优势物种组成、网络关系与疏林草地相比仍存在较大差异(图4~5).长期围封后,优势植株个体生长高大,占据主要生存空间,加剧种间竞争,限制矮小植株生长[14].相反,一些研究报道了放牧反而有利于缓和竞争、促进种群密度增加[27-29].另一方面,土壤结构改善和肥力恢复对植被恢复至关重要,但现有研究表明,土壤养分恢复通常需要数十年甚至上百年的时间[19,30].这表明严重沙漠化土地植被自然恢复过程漫长且缓慢.尽管很难将流动沙丘恢复到与疏林草地相当的状况,但进一步确定可持续围封的最佳年限是有必要的,围封时间过长会引发对草地生态系统可持续的担忧[8,28]. ...
Vegetation characteristics and soil properties in grazing exclusion areas of the Inner Mongolia desert steppe
1
2023
... 围封通过排除人类活动干扰,依靠自然演替和更新恢复受损生态系统,具有效率高、成本低、已恢复生态系统稳定性强、生态系统服务高等优势,是退化草地恢复广泛采用的方法.现有研究表明围封有利于植被群落恢复演替、提高土壤肥力[5].围封使退化典型草原[6]、荒漠草原[7]、沙质草地[8]、高寒草地[9]植物群落高度、地上生物量显著增加.但对物种多样性的影响则存在差异,这可能与草地类型、气候、生物地理格局、退化程度等存在差异有关[10].例如,短期(6年)围封显著增加轻度、中度退化荒漠草原的物种多样性,但重度退化荒漠草原围封后物种多样性降低[11].围封未改变荒漠草原优势建群种的优势度[12];但在典型草原中物种多样性随围封年限先降后增,群落优势物种依次沿杂类草、多年生禾草、杂类草3个阶段演替[6].与放牧相比,围封10年沙质草地可显著增加物种多样性、增强群落稳定性,并改善群落结构和功能[8];但围封20年后物种多样性与不同程度放牧样地差异不显著[13].严重沙漠化草地围封恢复主要有3个阶段:①狗尾草、马唐(Digitaria sanguinalis)等定殖与消失,②多年生根茎禾草为优势物种,③多年生丛生禾草为优势物种[14].尽管大量研究报道了围封对植物群落的影响,且围封年限对植物群落和土壤肥力恢复产生重要影响[15],但针对严重沙漠化草地长期围封对植物群落的影响研究有限,其驱动机制仍不清楚. ...
科尔沁沙地沙漠化土地恢复面临的挑战
1
2009
... 科尔沁沙地原为水草丰美的疏林草原,但由于人口快速增长,过度开垦、放牧、樵采导致严重沙漠化,到20世纪80年代沙漠化面积超过6万km2,位居四大沙地之首[16].科尔沁沙地是最早开始沙漠化草地围封恢复试验的地区之一.经过数十年的生态治理,该区域率先实现了沙漠化整体逆转、退化生态系统加速恢复[17].因此,本文以科尔沁沙地严重沙漠化草地(流动沙丘)长期围封下植物群落特征为主要研究对象,通过时空替代法,分析长期围封对植物群落数量特征、结构、多样性、种间网络共现模式的影响及其驱动因子,以期为该区域沙漠化防治策略制定和优化提供科学依据. ...
科尔沁沙地防沙治沙实践与生态可持续修复浅议
1
2024
... 科尔沁沙地原为水草丰美的疏林草原,但由于人口快速增长,过度开垦、放牧、樵采导致严重沙漠化,到20世纪80年代沙漠化面积超过6万km2,位居四大沙地之首[16].科尔沁沙地是最早开始沙漠化草地围封恢复试验的地区之一.经过数十年的生态治理,该区域率先实现了沙漠化整体逆转、退化生态系统加速恢复[17].因此,本文以科尔沁沙地严重沙漠化草地(流动沙丘)长期围封下植物群落特征为主要研究对象,通过时空替代法,分析长期围封对植物群落数量特征、结构、多样性、种间网络共现模式的影响及其驱动因子,以期为该区域沙漠化防治策略制定和优化提供科学依据. ...
Accumulation of carbon and nitrogen in the plant-soil system after afforestation of active sand dunes in China's Horqin Sandy Land
1
2013
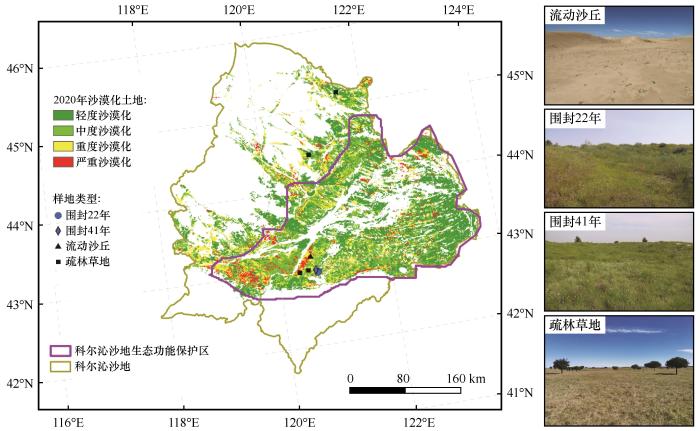
... 科尔沁沙地位于内蒙古自治区东南部(41.69°—46.00°N、117.82°—123.70°E,图1),总面积约12万km2;其中,科尔沁沙地防风固沙生态功能保护区约5.4万km2.研究区为温带大陆性半干旱气候,属于半干旱-半湿润气候过渡区.2000—2024年平均气温为7.3 ℃,平均降水量315 mm(数据来源于中国科学院奈曼沙漠化研究站气象观测数据).土壤类型主要有栗钙土、风沙土、潮土、黑钙土等.植物群落原为以禾草和稀疏分布的木本植物组成的疏林草地为主[18],由于沙漠化,优势草本植物逐渐变为一年生沙生植物.截至2020年,沙漠化土地面积约4.66万km2,主要分布于中、东部地区(图1). ...
Effects of grazing exclusion on carbon sequestration and the associated vegetation and soil characteristics at a semi-arid desertified sandy site in Inner Mongolia,Northern China
2
2012
... 野外调查依托中国科学院奈曼沙漠化研究站长期监测样地开展.根据已有研究基础[19],本研究选取流动沙丘围封22、41年的恢复样地为主要研究对象,并以流动沙丘(严重沙漠化,等效恢复0年)和疏林草地(近顶级群落)为对照.野外调查于2022年7月底集中进行.每类样地设置至少3个重复(图1),每个样地随机设置3个20 m×20 m大样方,间距大于50 m,且均位于阳坡以尽可能较少坡向的影响.大样方内沿对角线以10 m为间隔,设置3个1 m×1 m小样方.小样方内开展植物群落调查,记录物种组成、高度、盖度、株数等,然后收集地上活体生物量和凋落物生物量.随后,小样方内利用直径2.5 cm的土钻以九点法采集0~20 cm土壤样品.同时,利用体积100 cm3的环刀采集原状土两份,用于测定土壤容重、含水量、饱和持水量、田间持水量.期间,记录样地位置、地形等信息(表1). ...
... 现有研究表明,荒漠草地的最佳围封年限为9~12 a[26].但本研究结果与此年限相差甚远.在科尔沁沙地,严重沙漠化草地围封41年后,尽管植物群落数量特征与疏林草地接近,地上生物量、生物多样性甚至高于后者,但植物群落结构、优势物种组成、网络关系与疏林草地相比仍存在较大差异(图4~5).长期围封后,优势植株个体生长高大,占据主要生存空间,加剧种间竞争,限制矮小植株生长[14].相反,一些研究报道了放牧反而有利于缓和竞争、促进种群密度增加[27-29].另一方面,土壤结构改善和肥力恢复对植被恢复至关重要,但现有研究表明,土壤养分恢复通常需要数十年甚至上百年的时间[19,30].这表明严重沙漠化土地植被自然恢复过程漫长且缓慢.尽管很难将流动沙丘恢复到与疏林草地相当的状况,但进一步确定可持续围封的最佳年限是有必要的,围封时间过长会引发对草地生态系统可持续的担忧[8,28]. ...
Response of the plant-soil system to desertification in the Hulun Buir Sandy Land, China
2
2023
... 利用公式(1)~(3)[20]计算植物物种重要值(IV)、Shannon-Wiener多样性指数(H)、Pielou均匀度指数(J). ...
... 长期围封使植物群落由一年生草本沙蓬逐渐向以狗尾草为主的一年生草本植物(围封22年)和以多年生根茎性、丛生性禾草和蒿属植物为主(围封41年)的多年生草本植物群落变化.草本植物物种多样性和生物量也显著增加,甚至略高于疏林草地,这与先前观测到的结果基本一致[13-14],而与沙漠化过程相反[20].围封在一定程度上排除了人类活动干扰,一年生植物大多属于机会主义者,具有较高的水分和养分利用效率,容易在恢复早期形成竞争优势[13,21].随围封进行,多年生植物逐渐定殖,一方面植被盖度和养分输入增加,土壤侵蚀和蒸发减少,为多年生植物生长创造了条件[22].另一方面,由于其深根性[13],可以从深层土壤中获取养分和水分,从而更具优势.与王益涛等[12]报道的围封恢复后优势物种无变化的结果不同,在本研究中,随围封年限增加,植物群落优势物种由一年生逐渐向多年生植物转变,且围封41年后优势程度相当的物种数量增加.这可能与围封年限和恢复的起始对照(即退化程度)有关,前者仅围封4年.相反,本研究未观察到其他研究[23]报道的退化草地围封后灌木优势度明显增加的结果. ...
围封年限对内蒙古灌丛化草原小叶锦鸡儿灌丛结构及群落种间关联的影响
1
2021
... 长期围封使植物群落由一年生草本沙蓬逐渐向以狗尾草为主的一年生草本植物(围封22年)和以多年生根茎性、丛生性禾草和蒿属植物为主(围封41年)的多年生草本植物群落变化.草本植物物种多样性和生物量也显著增加,甚至略高于疏林草地,这与先前观测到的结果基本一致[13-14],而与沙漠化过程相反[20].围封在一定程度上排除了人类活动干扰,一年生植物大多属于机会主义者,具有较高的水分和养分利用效率,容易在恢复早期形成竞争优势[13,21].随围封进行,多年生植物逐渐定殖,一方面植被盖度和养分输入增加,土壤侵蚀和蒸发减少,为多年生植物生长创造了条件[22].另一方面,由于其深根性[13],可以从深层土壤中获取养分和水分,从而更具优势.与王益涛等[12]报道的围封恢复后优势物种无变化的结果不同,在本研究中,随围封年限增加,植物群落优势物种由一年生逐渐向多年生植物转变,且围封41年后优势程度相当的物种数量增加.这可能与围封年限和恢复的起始对照(即退化程度)有关,前者仅围封4年.相反,本研究未观察到其他研究[23]报道的退化草地围封后灌木优势度明显增加的结果. ...
科尔沁沙地围封和造林的土壤有机碳固存效应及其影响机制
2
2024
... 长期围封使植物群落由一年生草本沙蓬逐渐向以狗尾草为主的一年生草本植物(围封22年)和以多年生根茎性、丛生性禾草和蒿属植物为主(围封41年)的多年生草本植物群落变化.草本植物物种多样性和生物量也显著增加,甚至略高于疏林草地,这与先前观测到的结果基本一致[13-14],而与沙漠化过程相反[20].围封在一定程度上排除了人类活动干扰,一年生植物大多属于机会主义者,具有较高的水分和养分利用效率,容易在恢复早期形成竞争优势[13,21].随围封进行,多年生植物逐渐定殖,一方面植被盖度和养分输入增加,土壤侵蚀和蒸发减少,为多年生植物生长创造了条件[22].另一方面,由于其深根性[13],可以从深层土壤中获取养分和水分,从而更具优势.与王益涛等[12]报道的围封恢复后优势物种无变化的结果不同,在本研究中,随围封年限增加,植物群落优势物种由一年生逐渐向多年生植物转变,且围封41年后优势程度相当的物种数量增加.这可能与围封年限和恢复的起始对照(即退化程度)有关,前者仅围封4年.相反,本研究未观察到其他研究[23]报道的退化草地围封后灌木优势度明显增加的结果. ...
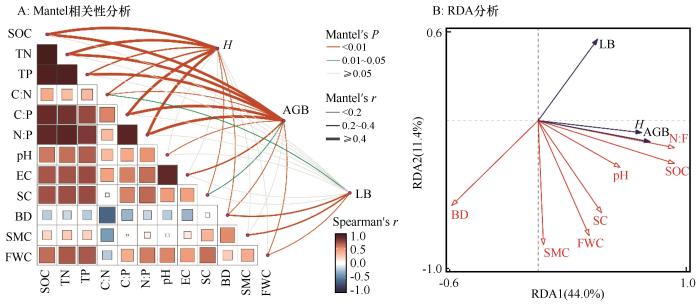
... Mantel检验和RDA结果均表明,植物群落多样性和生物量主要与土壤养分状况有关(图6),其中SOC含量是最关键的影响因素(表3).这与诸多研究结果一致[31-32],而与物种多样性、生物量主要受N∶P(即二者的化学计量平衡)调控的结果略有差异[33].一方面,SOC与TN、TP(因共线性被剔除)具有较强的相关性,指示土壤为植物生长提供能量和养分的能力.通常,植物群落物种多样性与土壤养分具有较强正相关关系,土壤养分供应决定了土壤能支持的植物总量,并主导群落结构和功能的变化[34].另一方面,在经历严重沙漠化后,土壤有机物(氮也主要以有机形式储存于土壤中)随土壤黏粉粒等细颗粒物质流失而大量损失[35].相比之下,植物多样性与TP的关系稍弱(图6A).磷属于沉积性元素,主要来源于风化作用[36-37],在退化和恢复过程中变化幅度相对较小[22].然而,植物地上活体生物量主要受N∶P的驱动,且二者呈显著的正相关关系(图6),表明在围封恢复过程中植物群落生产力可能受氮限制. ...
呼伦贝尔沙化草地围封效应及生物多样性变化研究
1
2008
... 长期围封使植物群落由一年生草本沙蓬逐渐向以狗尾草为主的一年生草本植物(围封22年)和以多年生根茎性、丛生性禾草和蒿属植物为主(围封41年)的多年生草本植物群落变化.草本植物物种多样性和生物量也显著增加,甚至略高于疏林草地,这与先前观测到的结果基本一致[13-14],而与沙漠化过程相反[20].围封在一定程度上排除了人类活动干扰,一年生植物大多属于机会主义者,具有较高的水分和养分利用效率,容易在恢复早期形成竞争优势[13,21].随围封进行,多年生植物逐渐定殖,一方面植被盖度和养分输入增加,土壤侵蚀和蒸发减少,为多年生植物生长创造了条件[22].另一方面,由于其深根性[13],可以从深层土壤中获取养分和水分,从而更具优势.与王益涛等[12]报道的围封恢复后优势物种无变化的结果不同,在本研究中,随围封年限增加,植物群落优势物种由一年生逐渐向多年生植物转变,且围封41年后优势程度相当的物种数量增加.这可能与围封年限和恢复的起始对照(即退化程度)有关,前者仅围封4年.相反,本研究未观察到其他研究[23]报道的退化草地围封后灌木优势度明显增加的结果. ...
科尔沁沙质草地植物群落对增温的响应
1
2024
... 共现网络分析表明,围封后植物群落共现模式发生明显改变(图5).随围封年限增加,尽管网络节点数未发生改变,但平均连通度增加,表明植物群落网络结构复杂度增加[24].上述结果与先前报道的长期围封使植物群落组成多样化、复杂化的结果一致[8].关键种与群落其他物种的相互作用最为强烈,对群落稳定性、多样性具有重要作用.网络分析结果表明围封后植物物种间以正相关关系为主(图5),表明关键种对其他物种的分布和数量具有积极影响,从而指示群落演替[25].与优势物种不同,关键种以矮小杂类草(如虱子草、牻牛儿苗、砂韭)为主,主要是由于随围封年限增加,植物群落盖度、高度均显著增加(图2),以上物种在空间、光照不足的条件下更具优势,从而占据不同的生态位,并削弱种间竞争,实现群落结构的稳定. ...
不同放牧方式对高寒草甸植物群落关键种的影响
1
2024
... 共现网络分析表明,围封后植物群落共现模式发生明显改变(图5).随围封年限增加,尽管网络节点数未发生改变,但平均连通度增加,表明植物群落网络结构复杂度增加[24].上述结果与先前报道的长期围封使植物群落组成多样化、复杂化的结果一致[8].关键种与群落其他物种的相互作用最为强烈,对群落稳定性、多样性具有重要作用.网络分析结果表明围封后植物物种间以正相关关系为主(图5),表明关键种对其他物种的分布和数量具有积极影响,从而指示群落演替[25].与优势物种不同,关键种以矮小杂类草(如虱子草、牻牛儿苗、砂韭)为主,主要是由于随围封年限增加,植物群落盖度、高度均显著增加(图2),以上物种在空间、光照不足的条件下更具优势,从而占据不同的生态位,并削弱种间竞争,实现群落结构的稳定. ...
基于植物群落特征的内蒙古荒漠草原适宜围封年限对比评价
1
2021
... 现有研究表明,荒漠草地的最佳围封年限为9~12 a[26].但本研究结果与此年限相差甚远.在科尔沁沙地,严重沙漠化草地围封41年后,尽管植物群落数量特征与疏林草地接近,地上生物量、生物多样性甚至高于后者,但植物群落结构、优势物种组成、网络关系与疏林草地相比仍存在较大差异(图4~5).长期围封后,优势植株个体生长高大,占据主要生存空间,加剧种间竞争,限制矮小植株生长[14].相反,一些研究报道了放牧反而有利于缓和竞争、促进种群密度增加[27-29].另一方面,土壤结构改善和肥力恢复对植被恢复至关重要,但现有研究表明,土壤养分恢复通常需要数十年甚至上百年的时间[19,30].这表明严重沙漠化土地植被自然恢复过程漫长且缓慢.尽管很难将流动沙丘恢复到与疏林草地相当的状况,但进一步确定可持续围封的最佳年限是有必要的,围封时间过长会引发对草地生态系统可持续的担忧[8,28]. ...
封育和放牧对高寒草甸植被群落特征和土壤特性的影响
1
2021
... 现有研究表明,荒漠草地的最佳围封年限为9~12 a[26].但本研究结果与此年限相差甚远.在科尔沁沙地,严重沙漠化草地围封41年后,尽管植物群落数量特征与疏林草地接近,地上生物量、生物多样性甚至高于后者,但植物群落结构、优势物种组成、网络关系与疏林草地相比仍存在较大差异(图4~5).长期围封后,优势植株个体生长高大,占据主要生存空间,加剧种间竞争,限制矮小植株生长[14].相反,一些研究报道了放牧反而有利于缓和竞争、促进种群密度增加[27-29].另一方面,土壤结构改善和肥力恢复对植被恢复至关重要,但现有研究表明,土壤养分恢复通常需要数十年甚至上百年的时间[19,30].这表明严重沙漠化土地植被自然恢复过程漫长且缓慢.尽管很难将流动沙丘恢复到与疏林草地相当的状况,但进一步确定可持续围封的最佳年限是有必要的,围封时间过长会引发对草地生态系统可持续的担忧[8,28]. ...
禁牧对退化草地恢复的作用
1
2015
... 现有研究表明,荒漠草地的最佳围封年限为9~12 a[26].但本研究结果与此年限相差甚远.在科尔沁沙地,严重沙漠化草地围封41年后,尽管植物群落数量特征与疏林草地接近,地上生物量、生物多样性甚至高于后者,但植物群落结构、优势物种组成、网络关系与疏林草地相比仍存在较大差异(图4~5).长期围封后,优势植株个体生长高大,占据主要生存空间,加剧种间竞争,限制矮小植株生长[14].相反,一些研究报道了放牧反而有利于缓和竞争、促进种群密度增加[27-29].另一方面,土壤结构改善和肥力恢复对植被恢复至关重要,但现有研究表明,土壤养分恢复通常需要数十年甚至上百年的时间[19,30].这表明严重沙漠化土地植被自然恢复过程漫长且缓慢.尽管很难将流动沙丘恢复到与疏林草地相当的状况,但进一步确定可持续围封的最佳年限是有必要的,围封时间过长会引发对草地生态系统可持续的担忧[8,28]. ...
不同放牧模式对祁连山高寒草甸植物群落特征的影响
1
2023
... 现有研究表明,荒漠草地的最佳围封年限为9~12 a[26].但本研究结果与此年限相差甚远.在科尔沁沙地,严重沙漠化草地围封41年后,尽管植物群落数量特征与疏林草地接近,地上生物量、生物多样性甚至高于后者,但植物群落结构、优势物种组成、网络关系与疏林草地相比仍存在较大差异(图4~5).长期围封后,优势植株个体生长高大,占据主要生存空间,加剧种间竞争,限制矮小植株生长[14].相反,一些研究报道了放牧反而有利于缓和竞争、促进种群密度增加[27-29].另一方面,土壤结构改善和肥力恢复对植被恢复至关重要,但现有研究表明,土壤养分恢复通常需要数十年甚至上百年的时间[19,30].这表明严重沙漠化土地植被自然恢复过程漫长且缓慢.尽管很难将流动沙丘恢复到与疏林草地相当的状况,但进一步确定可持续围封的最佳年限是有必要的,围封时间过长会引发对草地生态系统可持续的担忧[8,28]. ...
Recovery of topsoil physicochemical properties in revegetated sites in the sand-burial ecosystems of the Tengger Desert, Northern China
1
2007
... 现有研究表明,荒漠草地的最佳围封年限为9~12 a[26].但本研究结果与此年限相差甚远.在科尔沁沙地,严重沙漠化草地围封41年后,尽管植物群落数量特征与疏林草地接近,地上生物量、生物多样性甚至高于后者,但植物群落结构、优势物种组成、网络关系与疏林草地相比仍存在较大差异(图4~5).长期围封后,优势植株个体生长高大,占据主要生存空间,加剧种间竞争,限制矮小植株生长[14].相反,一些研究报道了放牧反而有利于缓和竞争、促进种群密度增加[27-29].另一方面,土壤结构改善和肥力恢复对植被恢复至关重要,但现有研究表明,土壤养分恢复通常需要数十年甚至上百年的时间[19,30].这表明严重沙漠化土地植被自然恢复过程漫长且缓慢.尽管很难将流动沙丘恢复到与疏林草地相当的状况,但进一步确定可持续围封的最佳年限是有必要的,围封时间过长会引发对草地生态系统可持续的担忧[8,28]. ...
Plant community C∶N∶P stoichiometry is mediated by soil nutrients and plant functional groups during grassland desertification
1
2021
... Mantel检验和RDA结果均表明,植物群落多样性和生物量主要与土壤养分状况有关(图6),其中SOC含量是最关键的影响因素(表3).这与诸多研究结果一致[31-32],而与物种多样性、生物量主要受N∶P(即二者的化学计量平衡)调控的结果略有差异[33].一方面,SOC与TN、TP(因共线性被剔除)具有较强的相关性,指示土壤为植物生长提供能量和养分的能力.通常,植物群落物种多样性与土壤养分具有较强正相关关系,土壤养分供应决定了土壤能支持的植物总量,并主导群落结构和功能的变化[34].另一方面,在经历严重沙漠化后,土壤有机物(氮也主要以有机形式储存于土壤中)随土壤黏粉粒等细颗粒物质流失而大量损失[35].相比之下,植物多样性与TP的关系稍弱(图6A).磷属于沉积性元素,主要来源于风化作用[36-37],在退化和恢复过程中变化幅度相对较小[22].然而,植物地上活体生物量主要受N∶P的驱动,且二者呈显著的正相关关系(图6),表明在围封恢复过程中植物群落生产力可能受氮限制. ...
Impact of desertification on soil and plant nutrient stoichiometry in a desert grassland
1
2019
... Mantel检验和RDA结果均表明,植物群落多样性和生物量主要与土壤养分状况有关(图6),其中SOC含量是最关键的影响因素(表3).这与诸多研究结果一致[31-32],而与物种多样性、生物量主要受N∶P(即二者的化学计量平衡)调控的结果略有差异[33].一方面,SOC与TN、TP(因共线性被剔除)具有较强的相关性,指示土壤为植物生长提供能量和养分的能力.通常,植物群落物种多样性与土壤养分具有较强正相关关系,土壤养分供应决定了土壤能支持的植物总量,并主导群落结构和功能的变化[34].另一方面,在经历严重沙漠化后,土壤有机物(氮也主要以有机形式储存于土壤中)随土壤黏粉粒等细颗粒物质流失而大量损失[35].相比之下,植物多样性与TP的关系稍弱(图6A).磷属于沉积性元素,主要来源于风化作用[36-37],在退化和恢复过程中变化幅度相对较小[22].然而,植物地上活体生物量主要受N∶P的驱动,且二者呈显著的正相关关系(图6),表明在围封恢复过程中植物群落生产力可能受氮限制. ...
Nitrogen:phosphorous supply ratio and allometry in five alpine plant species
1
2016
... Mantel检验和RDA结果均表明,植物群落多样性和生物量主要与土壤养分状况有关(图6),其中SOC含量是最关键的影响因素(表3).这与诸多研究结果一致[31-32],而与物种多样性、生物量主要受N∶P(即二者的化学计量平衡)调控的结果略有差异[33].一方面,SOC与TN、TP(因共线性被剔除)具有较强的相关性,指示土壤为植物生长提供能量和养分的能力.通常,植物群落物种多样性与土壤养分具有较强正相关关系,土壤养分供应决定了土壤能支持的植物总量,并主导群落结构和功能的变化[34].另一方面,在经历严重沙漠化后,土壤有机物(氮也主要以有机形式储存于土壤中)随土壤黏粉粒等细颗粒物质流失而大量损失[35].相比之下,植物多样性与TP的关系稍弱(图6A).磷属于沉积性元素,主要来源于风化作用[36-37],在退化和恢复过程中变化幅度相对较小[22].然而,植物地上活体生物量主要受N∶P的驱动,且二者呈显著的正相关关系(图6),表明在围封恢复过程中植物群落生产力可能受氮限制. ...
Variations in stand structure and diversity along a soil fertility gradient in a Brazilian Savanna (Cerrado) in Southern Mato Grosso
1
2013
... Mantel检验和RDA结果均表明,植物群落多样性和生物量主要与土壤养分状况有关(图6),其中SOC含量是最关键的影响因素(表3).这与诸多研究结果一致[31-32],而与物种多样性、生物量主要受N∶P(即二者的化学计量平衡)调控的结果略有差异[33].一方面,SOC与TN、TP(因共线性被剔除)具有较强的相关性,指示土壤为植物生长提供能量和养分的能力.通常,植物群落物种多样性与土壤养分具有较强正相关关系,土壤养分供应决定了土壤能支持的植物总量,并主导群落结构和功能的变化[34].另一方面,在经历严重沙漠化后,土壤有机物(氮也主要以有机形式储存于土壤中)随土壤黏粉粒等细颗粒物质流失而大量损失[35].相比之下,植物多样性与TP的关系稍弱(图6A).磷属于沉积性元素,主要来源于风化作用[36-37],在退化和恢复过程中变化幅度相对较小[22].然而,植物地上活体生物量主要受N∶P的驱动,且二者呈显著的正相关关系(图6),表明在围封恢复过程中植物群落生产力可能受氮限制. ...
Effects of desertification on soil organic C and N content in sandy farmland and grassland of Inner Mongolia
1
2009
... Mantel检验和RDA结果均表明,植物群落多样性和生物量主要与土壤养分状况有关(图6),其中SOC含量是最关键的影响因素(表3).这与诸多研究结果一致[31-32],而与物种多样性、生物量主要受N∶P(即二者的化学计量平衡)调控的结果略有差异[33].一方面,SOC与TN、TP(因共线性被剔除)具有较强的相关性,指示土壤为植物生长提供能量和养分的能力.通常,植物群落物种多样性与土壤养分具有较强正相关关系,土壤养分供应决定了土壤能支持的植物总量,并主导群落结构和功能的变化[34].另一方面,在经历严重沙漠化后,土壤有机物(氮也主要以有机形式储存于土壤中)随土壤黏粉粒等细颗粒物质流失而大量损失[35].相比之下,植物多样性与TP的关系稍弱(图6A).磷属于沉积性元素,主要来源于风化作用[36-37],在退化和恢复过程中变化幅度相对较小[22].然而,植物地上活体生物量主要受N∶P的驱动,且二者呈显著的正相关关系(图6),表明在围封恢复过程中植物群落生产力可能受氮限制. ...
Soil and fine roots ecological stoichiometry in different vegetation restoration stages in a karst area,Southwest China
1
2019
... Mantel检验和RDA结果均表明,植物群落多样性和生物量主要与土壤养分状况有关(图6),其中SOC含量是最关键的影响因素(表3).这与诸多研究结果一致[31-32],而与物种多样性、生物量主要受N∶P(即二者的化学计量平衡)调控的结果略有差异[33].一方面,SOC与TN、TP(因共线性被剔除)具有较强的相关性,指示土壤为植物生长提供能量和养分的能力.通常,植物群落物种多样性与土壤养分具有较强正相关关系,土壤养分供应决定了土壤能支持的植物总量,并主导群落结构和功能的变化[34].另一方面,在经历严重沙漠化后,土壤有机物(氮也主要以有机形式储存于土壤中)随土壤黏粉粒等细颗粒物质流失而大量损失[35].相比之下,植物多样性与TP的关系稍弱(图6A).磷属于沉积性元素,主要来源于风化作用[36-37],在退化和恢复过程中变化幅度相对较小[22].然而,植物地上活体生物量主要受N∶P的驱动,且二者呈显著的正相关关系(图6),表明在围封恢复过程中植物群落生产力可能受氮限制. ...
Effects of farmland conversion on the stoichiometry of carbon, nitrogen, and phosphorus in soil aggregates on the Loess Plateau of China
1
2019
... Mantel检验和RDA结果均表明,植物群落多样性和生物量主要与土壤养分状况有关(图6),其中SOC含量是最关键的影响因素(表3).这与诸多研究结果一致[31-32],而与物种多样性、生物量主要受N∶P(即二者的化学计量平衡)调控的结果略有差异[33].一方面,SOC与TN、TP(因共线性被剔除)具有较强的相关性,指示土壤为植物生长提供能量和养分的能力.通常,植物群落物种多样性与土壤养分具有较强正相关关系,土壤养分供应决定了土壤能支持的植物总量,并主导群落结构和功能的变化[34].另一方面,在经历严重沙漠化后,土壤有机物(氮也主要以有机形式储存于土壤中)随土壤黏粉粒等细颗粒物质流失而大量损失[35].相比之下,植物多样性与TP的关系稍弱(图6A).磷属于沉积性元素,主要来源于风化作用[36-37],在退化和恢复过程中变化幅度相对较小[22].然而,植物地上活体生物量主要受N∶P的驱动,且二者呈显著的正相关关系(图6),表明在围封恢复过程中植物群落生产力可能受氮限制. ...
Impact of tropical land-use change on soil organic carbon stocks-a meta-analysis
1
2011
... 土壤物理性质对植物群落也有重要影响,其中土壤容重和含水量是主要的影响因素.土壤容重与植物群落特征呈负相关关系,容重越大,土壤越紧实,从而限制土壤养分积累[38]、植物根系生长及其对养分的吸收[39].对于半干旱区沙地生态系统而言,水分可利用性是初级生产力的重要限制因子[40-41].土壤含水量增加有利于养分在土壤中的迁移、扩散和保持能力,增强生态系统生物多样性和恢复力[42-43]. ...
Heterogeneity of leaf stoichiometry of different life forms along environmental transects in typical ecologically fragile areas of China
1
2024
... 土壤物理性质对植物群落也有重要影响,其中土壤容重和含水量是主要的影响因素.土壤容重与植物群落特征呈负相关关系,容重越大,土壤越紧实,从而限制土壤养分积累[38]、植物根系生长及其对养分的吸收[39].对于半干旱区沙地生态系统而言,水分可利用性是初级生产力的重要限制因子[40-41].土壤含水量增加有利于养分在土壤中的迁移、扩散和保持能力,增强生态系统生物多样性和恢复力[42-43]. ...
Higher precipitation strengthens the microbial interactions in semi-arid grassland soils
1
2018
... 土壤物理性质对植物群落也有重要影响,其中土壤容重和含水量是主要的影响因素.土壤容重与植物群落特征呈负相关关系,容重越大,土壤越紧实,从而限制土壤养分积累[38]、植物根系生长及其对养分的吸收[39].对于半干旱区沙地生态系统而言,水分可利用性是初级生产力的重要限制因子[40-41].土壤含水量增加有利于养分在土壤中的迁移、扩散和保持能力,增强生态系统生物多样性和恢复力[42-43]. ...
宁夏草原植物叶片氮磷化学计量特征及其驱动因素
1
2019
... 土壤物理性质对植物群落也有重要影响,其中土壤容重和含水量是主要的影响因素.土壤容重与植物群落特征呈负相关关系,容重越大,土壤越紧实,从而限制土壤养分积累[38]、植物根系生长及其对养分的吸收[39].对于半干旱区沙地生态系统而言,水分可利用性是初级生产力的重要限制因子[40-41].土壤含水量增加有利于养分在土壤中的迁移、扩散和保持能力,增强生态系统生物多样性和恢复力[42-43]. ...
Changes of plant N∶P stoichiometry across a 3000-km aridity transect in grasslands of Northern China
1
2019
... 土壤物理性质对植物群落也有重要影响,其中土壤容重和含水量是主要的影响因素.土壤容重与植物群落特征呈负相关关系,容重越大,土壤越紧实,从而限制土壤养分积累[38]、植物根系生长及其对养分的吸收[39].对于半干旱区沙地生态系统而言,水分可利用性是初级生产力的重要限制因子[40-41].土壤含水量增加有利于养分在土壤中的迁移、扩散和保持能力,增强生态系统生物多样性和恢复力[42-43]. ...
Coherent responses of terrestrial C∶N stoichiometry to drought across plants,soil,and microorganisms in forests and grasslands
1
2020
... 土壤物理性质对植物群落也有重要影响,其中土壤容重和含水量是主要的影响因素.土壤容重与植物群落特征呈负相关关系,容重越大,土壤越紧实,从而限制土壤养分积累[38]、植物根系生长及其对养分的吸收[39].对于半干旱区沙地生态系统而言,水分可利用性是初级生产力的重要限制因子[40-41].土壤含水量增加有利于养分在土壤中的迁移、扩散和保持能力,增强生态系统生物多样性和恢复力[42-43]. ...




 甘公网安备 62010202000688号
甘公网安备 62010202000688号
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}