半干旱沙质草地植物群落构建对降水的响应
景家琪 , 1 , 2 , 4 , 刘新平 , 1 , 2 , 4 , 何玉惠 3 , 丰洁 5 , 胡鸿姣 1 , 2 , 4 , 徐远志 1 , 2 , 4
1.中国科学院西北生态环境资源研究院,干旱区生态安全与可持续发展全国重点实验室,甘肃 兰州 730000
2.中国科学院西北生态环境资源研究院,奈曼沙漠化研究站,甘肃 兰州 730000
3.中国科学院西北生态环境资源研究院,兰州生态农业试验研究站,甘肃 兰州 730000
4.中国科学院大学,北京 100049
5.通辽市林业和草原科学研究所,内蒙古 通辽 028000
Responces of plant community construction in semi-arid sandy grassland to precipitation changes
Jing Jiaqi , 1 , 2 , 4 , Liu Xinping , 1 , 2 , 4 , He Yuhui 3 , Feng Jie 5 , Hu Hongjiao 1 , 2 , 4 , Xu Yuanzhi 1 , 2 , 4
1.State Key Laboratory of Ecological Safety and Sustainable Development in Arid Lands /, Northwest Institute of Eco-Environment and Resources,Chinese Academy of Sciences,Lanzhou 730000,China
2.Naiman Desertification Research Station /, Northwest Institute of Eco-Environment and Resources,Chinese Academy of Sciences,Lanzhou 730000,China
3.Lanzhou Ecological Agriculture Experimental Research Station, Northwest Institute of Eco-Environment and Resources,Chinese Academy of Sciences,Lanzhou 730000,China
4.University of Chinese Academy of Sciences,Beijing 100049,China
5.Tongliao Forestry and Grassland Science Research Institute,Tongliao 028000,Inner Mongolia,China
通讯作者: 刘新平(E-mail: liuxinping@lzb.ac.cn )
收稿日期: 2024-12-19
修回日期: 2025-03-27
基金资助:
内蒙古自治区防沙治沙科技创新重大示范工程“揭榜挂帅”项目 . 2024JBGS0005-1 中国科学院区域发展青年学者项目 干旱区生态安全与可持续发展全国重点实验室交叉创新团队项目 . E451890201 内蒙古自治区科技计划项目 . 2022YFHH0063
Received: 2024-12-19
Revised: 2025-03-27
摘要
降水是影响半干旱沙质草地植被生长发育和系统稳定维持的关键因子。为探明降水变化如何影响固沙植物群落构建及其生产力,本文整合分析了2014—2023年的降水和半干旱沙质草地植被长期定位监测数据,研究了降水变化对植物群落结构组成、物种多样性和生产力的影响。结果表明:(1)在40%的年降水变异内,多年生草本植物糙隐子草(Cleistogenes squarrosa )和二裂委陵菜(Potentilla bifurca )、一年生草本植物狗尾草(Setaria viridis )和尖头叶藜(Chenopodium acuminatum )及半灌木兴安胡枝子(Lespedeza bicolor )在10年间均为建群种,不受降水变化的影响,而在50%的年降水变异内,草本植物白草(Pennisetum centrasiaticum )和猪毛蒿(Artemisia scoparia )连续5年成为群落优势种,其余物种则随着降水变化呈现年际波动;(2)沙质草地植物群落多样性指数随降水增加而降低,群落丰富度、优势度和均匀度相对稳定,而功能群均匀度离散程度较高且总体上升;(3)沙质草地一年生草本植物和多年生草本植物的生物量与当年7月及生长季降水显著正相关,且二者地上生物量呈现增减互补的变化趋势,这一特征是维持系统生产力稳定的权衡对策,尤其在2018年和2022年表现尤为明显。降水直接决定半干旱沙质草地植物群落构建及其功能群多样性,可为预测半干旱沙质草地未来降水变化情景下的固沙植被群落演替,以及沙地生态系统植被稳定性维持和植被资源化利用提供理论支持。
关键词:
降水变化 群落构建 生物多样性 地上生物量
Abstract
Precipitation directly affects the growth and development of vegetation, as well as the maintenance of system stability, in semi-arid sandy grasslands. To undersdand how changes in precipitation iinfluence the construction and productivity of sand-fixing plant communities, this study analyzes long-term monitoring data of precipitation and vegetation in semi-arid sandy grasslands from 2014 to 2023. It explores the impacts of precipitation changes on plant community structure, species diversity, and productivity. The results show that: (1) Within a 40% variation in annual precipitation, the perennial herbs Cleistogenes squarrosa and Potentilla bifurca , the panicum miliaceum Setaria viridis and Chenopodium acuminatum , as well as the subshrub Lespedeza bicolor were all constructive species over the 10-years. Within a 50% variation in annual precipitation, herbs Pennisetum centrasiaticum and Artemisia scoparia became the dominant species in the community for five consecutive years, while the remaining species fluctuated with the precipitation variation. (2) The diversity index of plant communities in sandy grasslands decreased with increasing precipitation. In contrast, community richness, dominance, and evenness were relatively stable. However, functional group evenness was highly dispersed and generally increased. (3) The biomass of Panicum miliaceum and perennial herbs in sandy grasslands was significantly positively correlated with the precipitation in July of that year and the growing season. Their aboveground biomass exhibited a complementary trend of increase and decrease, which seves as a trade-off mechanism to maintain the stability of system productivity, especially in 2018 and 2022. This study demonstrates that changes in precipitation directly determine the plant community construction and functional group diversity in semi-arid sandy grasslands. It provides theoretical support for predicting the succession of sand-fixing vegetation communities under future precipitation change scenarios, as well as for maintaining vegetation stability and utilizing vegetation resources in sandy ecosystems.
Keywords:
precipitation changes community construction biodiversity above-ground biomass
本文引用格式
景家琪, 刘新平, 何玉惠, 丰洁, 胡鸿姣, 徐远志. 半干旱沙质草地植物群落构建对降水的响应 . 中国沙漠 [J], 2025, 45(4): 314-323 doi:10.7522/j.issn.1000-694X.2025.00067
Jing Jiaqi, Liu Xinping, He Yuhui, Feng Jie, Hu Hongjiao, Xu Yuanzhi. Responces of plant community construction in semi-arid sandy grassland to precipitation changes . Journal of Desert Research
0 引言
半干旱沙质草地是典型草原向荒漠过渡的旱生化草地生态系统[1 ] ,是干旱半干旱地区陆地生态系统的主体部分[2 ] ,也是中国重要的草地生态系统[3 ] 。半干旱沙质草地极易受全球变化影响,且生态系统功能和稳定性较差[4 ] 。降水是影响干旱半干旱地区沙质草地生态系统中植被生长的主要限制因子[5 ] 。固沙植被在维持生态系统结构稳定中发挥着至关重要的作用,对降水变化的响应极为敏感[6 -7 ] 。此外,降水也是制约沙质草地植被生产力和植被恢复的关键环境因子[8 ] 。近年来由于气候变化和人类活动共同作用,中国旱区出现土地退化、水分流失和生物多样性丧失等问题[9 ] 。因此,研究降水变化对沙质草地植被特征及其群落构建的影响,对于植被恢复重建和稳定维持具有重要意义。
植物生物量是生态系统生产力的重要指征[10 -11 ] ,但降水变化与生产力之间的关系仍存在争议。降水增加会显著提高内蒙古典型草原植物群落地上生物量[12 ] ,但在科尔沁固定沙地[13 ] 和黄土高原草原-荒漠带[14 ] ,降水增加过度反而会降低植物群落生物量。同时,草原生态系统生产力对极端干旱和极端湿润处理的响应并非对称[15 ] ,极端干旱处理下草原生产力的降低显著大于极端湿润处理下生产力的增加,且年降水量与之的线性增加关系随气候由干旱向湿润逐渐减弱[16 ] 。此外,半干旱沙地植被群落构建还与降水的季节分配密切相关[17 ] ,尤其是生长季初期的降水直接决定了沙质草地固沙植被幼苗建植和物种组成。因此,由于草地类型、降水量和降水时间等非生物因素的不同,草原生态系统植物生物量对降水变化的响应还存在不确定性。
近年来,植物群落构建机制的研究逐渐成为生态学领域的热点。黄土高原腹地人工林下草本层群落构建机制的研究表明,生态位分化和中性过程的共同作用是影响群落构建的关键因素[18 ] 。此外,功能群多样性在群落构建中也发挥着重要作用,其通过影响植物对资源的获取和利用效率,调节群落的稳定性和功能[19 ] 。功能群多样性不仅能够增强植物群落对环境变化的适应能力,还能通过生态位互补和功能冗余机制,提高群落的整体生产力和稳定性[20 ] 。然而,当前研究大多集中在植物群落多样性及其生物量的动态变化上,而关于优势种和共有种的生物量对高变异降水响应的研究相对匮乏。此外,功能群多样性在群落构建中的作用及其对环境变化的响应机制尚未得到充分探讨。另外,这方面的研究目前大多关注植物群落多样性及其生物量的动态变化,而关于优势种和共有种的生物量响应高变异降水的研究相对匮乏。
本研究以科尔沁沙质草地植物群落为研究对象,结合近10年降水数据,分析降水变化对植物群落及功能群生物多样性和地上生物量的影响。研究旨在解决以下科学问题:①降水变化如何影响沙质草地植物群落及功能群的生物多样性和生物量?②优势种和共有种的生物量对高变异降水的响应是否存在差异?③功能群多样性在群落构建中扮演何种角色?从植物种尺度上探究植被与降水之间的关系,以期为半干旱地区沙质草地植被稳定与可持续利用提供理论依据和实践指导,为后续研究内容的展开奠定坚实基础。
1 材料与方法
1.1 研究区概况
研究区为内蒙古奈曼站沙地综合观测场(42°94′N、120°71′E,海拔约为353 m),位于科尔沁沙地南缘的奈曼旗境内。该地属于温带半干旱大陆性气候,年平均气温为5.8~6.4 ℃,多年平均降水量351.7 mm[21 ] ,年际降水变率大且分布不均,其中80%的降水量集中在4—8月。土壤类型主要为风沙土、沙质栗钙,土质疏松,保水能力较差,土壤有机质含量较低。地貌类型为流动沙丘、半固定沙丘、固定沙丘、平缓沙地等[22 ] 。该区草地植被以一年生或者多年生草本植物为主,主要优势植物有狗尾草(Setaria viridis )、尖头叶藜(Chenopodium acuminatum )、白草(Pennisetum centrasiaticum )和猪毛蒿(Artemisia scoparia )等。实验样地选址在科尔沁沙地的典型沙质草地,该区域植被覆盖度较低,受人类干扰较小,能够较好地反映自然条件下植物群落对降水变化的响应。
1.2 数据来源及分析方法
1.2.1 数据来源
2014—2023年降水数据和植被数据来自内蒙古奈曼站的标准气象站和沙质草地长期综合观测场。该观测场始建于1991年,面积约6 hm2 ,1991—1997年开展放牧试验,1997年开始进行围封恢复试验,2005年作为科尔沁沙地沙质草地植被群落分布、土壤类型和土壤水分状况的典型代表样地建成中国生态系统研究网络(CERN)永久沙地综合观测场。观测场位于奈曼站东北方向3 km的平缓沙质草地,在区域内根据草地植物种类调查和地形图随机设置10个用于长期监测的固定样方(样地大小均为1 m×1 m)。观测场长期围封且有人看护,不受外界干扰、地势较为平坦,沙质草地群落优势明显,水土气生等环境要素按照CERN的统一监测规范每年定期进行。观测场外主要是开放沙质草地和草地开垦的旱作农田。该观测场基本代表了科尔沁沙地沙质草地植被群落分布、土壤类型和土壤水分状况,每年植物生长盛期(8月下旬)进行一次植被调查,调查指标包括植物种,各物种的多度、高度(cm)、盖度(%)和地上生物量等。
1.2.2 分析方法
本研究使用IBM SPSS Statistics 27软件,采用单因素方差分析(One-Way ANOVA)来比较各物种重要值以及功能群多度、高度、盖度和地上生物量在年际间的差异,通过比较组间均值的差异,判断不同年份之间是否存在显著性差异。每个指标均包含10个重复测量值,为方差分析提供了充分的样本量,从而增强了统计检验的稳健性和可靠性。稿件中所有的图使用Origin 2024进行绘图。
相对重要值(IV )反映每个物种在群落中的重要性,植物群落α多样性采用Shannon-Wiener指数(H )、Margalef指数(R )、Simpson指数(D )及Pielou指数(J )表征[23 ] 。
I V = ( R d + R c + R h ) 3 (1)
H = - ∑ i = 1 s ( P i l n P i ) (2)
R = ( S - 1 ) l n N (3)
D = 1 - ∑ i = 1 s ( P i ) (4)
J = H l n S (5)
式中:Rd 为相对多度;Rc 为相对盖度;Rh 为相对高度;S 为总物种数量;N 为总个体数量;Pi =Ni /N ,Ni 表示物种i 的个体数量。
2 结果与分析
2.1 沙质草地植物群落特征的变化
研究区十年调查结果显示,沙质草地共记录23种植物,根据生活型可分为多年生草本、一年生草本和半灌木3个功能群。多年生草本包含10种植物,其中糙影子草(Cleistogenes squarrosa )和二裂委陵菜(Potentilla bifurca )为2014—2023年共有种,且二者重要值均在2020年达到最大值;蒙古野韭、米口袋、乳浆大戟三者均只在降水量较高的年份出现过一次且重要值较小。一年生草本包含12种植物,共有种为狗尾草和尖头叶藜,且重要值在2014—2018年间交替达到最高,成为群落优势种。相较于草本植物而言,半灌木仅有兴安胡枝子一种植物,并且为10年间共有种,其重要值在10年间呈现上升趋势,于2023年达到最大值。除共有种成为当年优势种外,白草和猪毛蒿分别在2014年和2019—2023年成为优势种,其中猪毛蒿的重要值在群落中占比较高,约为19.32%(表1 )。
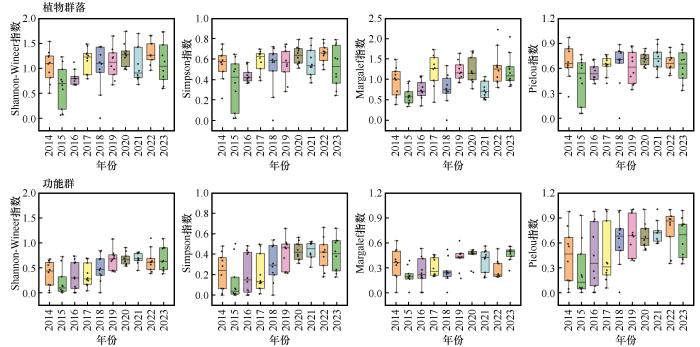
2014—2023年生物多样性指数并无显著差异(图1 )。2014—2023年,植物群落丰富度和优势度在不同年份间呈现波动变化,但整体上没有明显的上升或下降趋势,其指数中位数在10年间相对稳定,但离群值在2015年较为明显;而功能群丰富度和优势度呈现缓慢上升的趋势。植物群落和功能群的多样性指数随着降水增加而逐渐降低,在2016年数值明显低于2017年。植物群落均匀度10年间保持相对稳定,但存在一定的波动,2015年存在极小值;相较于植物群落而言,功能群均匀度离散程度更高,但总体呈现上升趋势。
图1
图1
2014—2023年沙质草地植物群落和功能物种多样性指数
Fig.1
Diversity index of plant communities and functional species in sandy grassland from 2014 to 2023
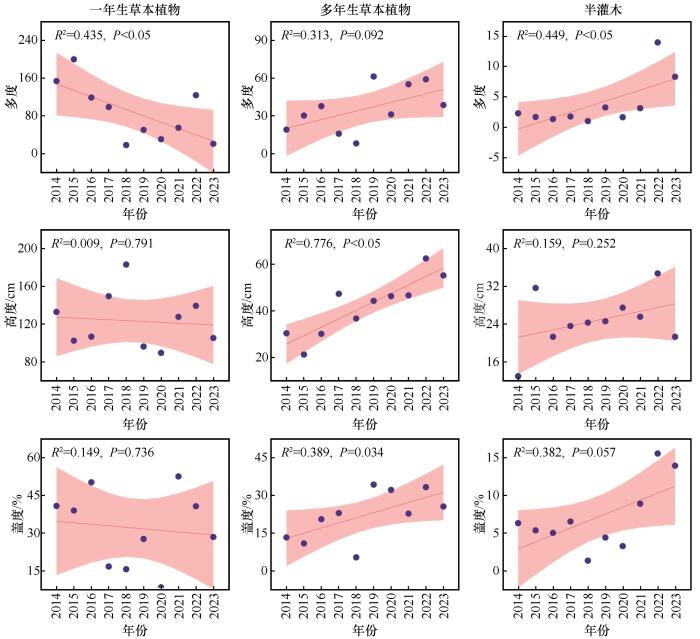
随着观测年限的加长,一年生草本植物的多度、高度、盖度呈现逐渐下降的趋势,而多年生草本植物和半灌木均在逐渐升高。根据单因素方差分析,植物功能群的多度、高度、盖度在不同年份的降水条件下均有差异。一年生草本植物在2015年和2022年的物种丰富度和覆盖度显著高于其他年份(P <0.05),而高度在2016年和2022年达到峰值;多年生草本植物的丰富度和盖度在2018年显著增加,高度在同年达到最大值;半灌木的丰富度、高度和盖度在2014年显著低于其他年份,但在2015年和2022年显著上升,尤其是盖度在2022年达到最高。整体而言,2014—2022年一年生草本植物多度、高度和盖度呈下降趋势,且多度达到显著性水平(图2 ,P< 0.05),多年生草本植物和半灌木呈上升趋势,其中多年生草本植物高度和半灌木多度两个指标达到显著性水平(P< 0.05)。
图2
图2
2014—2023年植物功能群的多度、高度、盖度差异
注: 不同字母揭示了植物群落对环境变化的响应及其在不同年份间的显著性差异(P <0.05)
Fig.2
Comparison of abundance, height and coverage of plant functional groups from 2014 to 2023
2.2 沙质草地植物生物量的动态
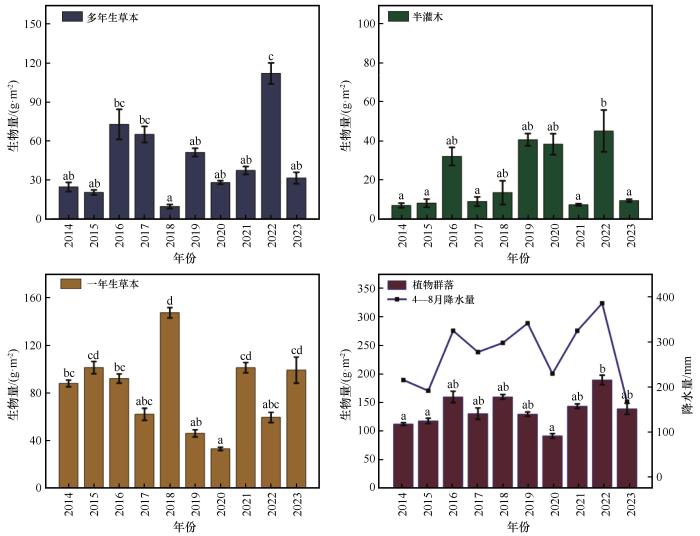
多年生草本植物、半灌木和群落地上生物量在2014—2023年的变化基本一致,而一年生草本植物地上生物量的变化趋势则与之相反(图3 )。其中,2022年植物群落地上生物量显著高于2014年、2015年和2020年(P <0.05),而2022年降水量相较于这3年分别增加了79%、101%和68%。除一年生草本植物外,其余植物的地上生物量均在2016年和2022年数值较大,该趋势与对应年份4—8月的降水相吻合。其中,2018年多年生草本植物的地上生物量达到最小值(8.55 g·m-2 ),而一年生草本植物的地上生物量在这一年达到最大值(147.53 g·m-2 );2022年的结果相反,一年生草本植物地上生物量达到最大值(112.06 g·m-2 ),而多年生草本植物地上生物量降到最小值(59.60 g·m-2 ),且其余年份多年生草本植物与一年生草本植物的地上生物量呈现增减互补的动态变化。
图3
图3
2014—2023年植物功能群地上生物量的变化
Fig.3
Changes of aboveground biomass of plant functional groups from 2014 to 2023
2.3 降水变化与群落构建的关系
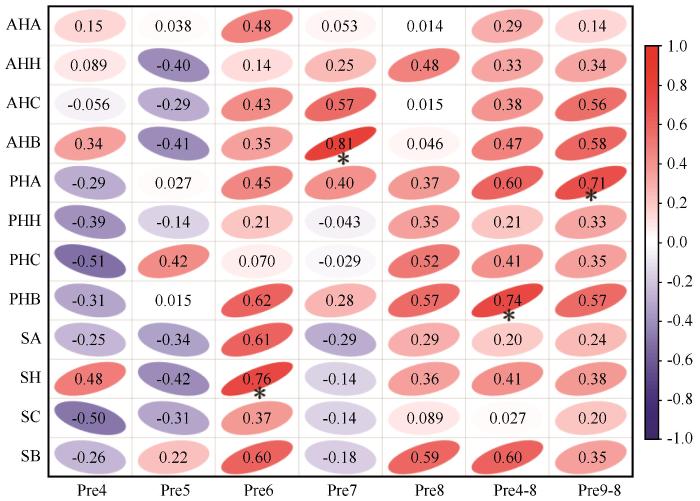
从草地植物群落指标与2014—2023年生长季逐月降水以及全年降水变化的相关分析来看,沙质草地植物群落特征与降水变化存在密切关系(图4 )。一年生草本植物和多年生草本植物的生物量分别与当年7月和生长季降水量显著相关(P <0.05),多年生草本植物的多度与年降水量显著相关(P <0.05),而半灌木的基本特征中,只有高度和当年6月降水量显著相关(P <0.05)。
图4
图4
2014—2023年植被群落特征与降水的相关性
注: AHA:一年生草本植物多度;AHH:一年生草本植物高度;AHC:一年生草本植物盖度;AHB:一年生草本植物生物量;PHA:多年生草本植物多度;PHH:多年生草本植物高度;PHC:多年生草本植物盖度;PHB:多年生草本植物生物量;SA:半灌木多度;SH:半灌木高度;SC:半灌木盖度;SB:半灌木生物量;Pre4:4月降水量;Pre5:5月降水量;Pre6:6月降水量;Pre7:7月降水量;Pre8:8月降水量;Pre4-8:4—8月降水量;Pre9-8:当年9月至次年8月降水量
Fig.4
Correlation between vegetation community characteristics and precipitation from 2014 to 2023
3 讨论
3.1 降水变化对植物群落多样性与生物量的影响
植物群落多样性是衡量物种分布均匀程度和数量的指标,是群落特征的综合体现[24 ] 。本研究结果表明,功能群的丰富度、优势度和均匀度随着降水增加呈现上升趋势,而多样性的变动与之相反。增雨会使植物群落的多样性降低,原因可能是降雨量的增加有助于一年生草本植物萌发并促进生长势,使得杂类草植物多度减少,植物群落中物种多样性由此降低。这一结果与周璐瑶等[25 ] 的研究一致,其指出氮添加与增雨处理相结合显著降低了群落丰富度,进而影响植物群落的多样性。孙岩等[24 ] 发现在沙漠地区增雨可能对某些植物群落的生物量有积极影响,但不会增加群落多样性。这表明降水变化对群落多样性的影响可能因植物群落类型和环境背景而异。同时,在群落多样性减少的状况下,有些植物如糙隐子草、二裂委陵菜、狗尾草、尖头叶藜和兴安胡枝子等受降水变化的影响较小。这与韩佳育[26 ] 、张晶等[27 ] 、杨浩等[28 ] 的研究结果相似,即这些植物对降水变化不敏感且具有较强的适应性。这种现象可能与植物的生态位分化有关,即这些物种在长期进化过程中形成了对干旱或湿润环境的适应策略,从而在降水变化下仍能保持相对稳定的地位[29 ] 。
生物量是衡量沙质草地植物生长发育状况的重要指标,直接反映植物的生长和生产力[30 ] 。本研究结果表明,降水量减少使植物群落地上生物量显著降低,原因可能是为应对干旱带来的危害,植物调节植株高度、叶面积和叶片干物质含量,导致植物地上部分积累的干物质减少,从而降低了植物地上生物量[31 -35 ] 。不同植物或功能群对水分利用策略的差异,导致其竞争能力和优势等级的不同,最终造成植物群落结构和组成比例发生改变。对于一年生草本植物而言,其生物量动态与年降水量变化趋势相反,与多年生草本植物的生物量呈现互补趋势,这是由于降水量减少影响了多年生植物以及半灌木的生长,从而导致其失去竞争力,促进一年生草本植物生长提高地上生物量;一年生草本植物对干旱环境具有一定的适应能力,轻度干旱胁迫对某些一年生草本植物生长影响较小,甚至有一定的促进作用。李慧等[36 ] 指出,组合群落会使植物的生物量和互补效应显著增加,在不同植物组合中,一年生和多年生草本植物的生物量存在互补性,二者组合可以增加总体生物量。美国明尼苏达州长期草地生物多样性试验也印证了我们的观点[37 ] ,即半干旱沙质草地多年生草本植物与一年生草本植物形成互补,共同提高植物群落生物量。这种生物量的互补性可能与植物的生态位分化和资源利用策略有关,未来研究可以进一步探讨其背后的生态机制。
3.2 降水变化对群落构建的影响
干旱半干旱地区草地生态系统稳定性对于降水变化极为敏感,植被作为草地生态系统的主体对降水变化的响应最为显著,而植物群落中不同的功能群对降水动态变化的响应机制却不同[38 -42 ] 。本研究结果表明,一年生草本和多年生草本的生物量分别与当年7月和生长季降水量显著相关。考虑到一年生草本地上生物量是每年8月收获测定,与当年7月降水量显著相关,主要原因是一年生草本植物的生长对短期降水变化非常敏感,前月降水量直接影响次月一年生草本植物的长势。而多年生草本植物的生物量与当年生长季降水量显著相关,是因为多年生草本植物在长期的进化过程中形成了适应干旱等胁迫环境的生理生态特征,对于降水变化敏感度低于一年生草本,其地上生物量动态变化不会被短期降水变化所影响。这与王军峰等[43 ] 在荒漠草原开展野外降水控制试验的结果一致,即一年生草本植物的生物量随短期降水变异波动明显,而多年生草本植物的生物量受整个生长季降水的影响。
降水变化对植物群落构建的影响可以从物种多样性、功能群组成和生态位动态等多个维度进行剖析。首先,降水变化可能通过影响物种的繁殖、生长和存活,进而改变群落的物种组成和多样性。例如,降水的增加可能促进某些物种的生长,但同时也会抑制其他物种的竞争力,导致群落物种多样性降低[44 ] 。降水变化对不同功能群的影响也存在差异。一年生草本植物通常对短期降水变化更为敏感,而多年生草本植物则表现出更强的适应性[45 ] 。这种差异可能导致功能群之间的竞争格局发生变化,进而影响群落的结构和功能[9 ] 。此外,降水变化还可能通过改变植物的生态位宽度和重叠程度,影响物种之间的相互作用和群落的稳定性[23 ] 。例如,降水量的减少可能使植物的生态位更加狭窄,导致物种之间的竞争加剧,而降水量的增加则可能扩大生态位宽度,促进物种共存[45 ] 。不同功能群对水分利用的策略存在差异,导致各功能群植物地上生物量在群落中的比例发生改变。这种生物量分配策略的变化反映了植物群落对降水变化的适应机制。本研究探讨了不同功能群在不同降水条件下的分布和竞争关系。在40%的年降水变异条件下,禾本科植物糙隐子草和豆科植物兴安胡枝子等表现出较强的竞争优势并占据更优生态位,而随着降水变异增加到50%,菊科植物猪毛蒿和禾本科植物白草逐渐成为优势种。这表明降水变化显著影响了功能群之间的竞争格局,进而改变了群落的物种组成和优势度分布,这些变化对群落稳定性和生态系统功能可能产生重要影响。功能群多样性的变化反映了群落对环境胁迫的响应机制,功能群的调整有助于维持生态系统生产力的稳定性。例如,功能群均匀度的离散程度在高降水变异条件下显著增加,这可能削弱了群落的整体稳定性。此外,功能群之间的竞争关系也对生态系统功能产生影响,禾本科植物的生物量与群落总生物量正相关,表明其在生态系统生产力中占据主导地位。
4 结论
降水变化直接决定了半干旱沙质草地植物群落结构与组成,降水增加使功能群植物的丰富度、优势度和均匀度逐渐提高,而多样性则表现出相反的变化趋势,且功能群多样性比群落多样性更容易受降水变化的影响。此外,本研究强调了群落结构对降水变化的敏感性,揭示了功能群在不同降水条件下的群落构建特征。具体而言,不同物种在高变异降水条件下生长状况存在差异,一年生草本、多年生草本以及半灌木中均有作为共有种的植物在40%的年降水变异区间内不受影响,而草本植物白草和猪毛蒿在50%的年降水变异区间内连续5年成为优势种。植物群落通过调整生物量分配策略来适应降水变化,进而影响群落结构和生态系统功能。不同功能群对水分利用的策略存在差异,导致各功能群植物地上生物量在群落中的比例发生改变。当年7月降水量决定了一年生草本植物的生物量,而整个生长季的降水量决定了多年生草本植物的生物量。本研究强调了在半干旱沙质草地生态系统管理中考虑功能群多样性和生物量分配策略的重要性,为沙质草地植被稳定和可持续利用提供理论支持,为应对未来气候变化提供科学依据。
参考文献
View Option
[1]
董晓宇 2000-2017年内蒙古荒漠草原植被物候和净初级生产力对气候变化的响应
[D].西安 :长安大学 , 2020 .
[本文引用: 1]
[2]
韩国栋 ,焦树英 ,毕力格图 ,等 短花针茅草原不同载畜率对植物多样性和草地生产力的影响
[J].生态学报 ,2007 (1 ):182 -188 .
[本文引用: 1]
[3]
晔薷罕 极端干旱对典型草原土壤与植被的影响
[D].北京 :中国农业科学院 ,2022 .
[本文引用: 1]
[4]
赵生龙 ,左小安 ,张铜会 ,等 乌拉特荒漠草原群落物种多样性和生物量关系对放牧强度的响应
[J].干旱区研究 ,2020 ,37 (1 ):168 -177 .
[本文引用: 1]
[5]
罗叙 ,王誉陶 ,张娟 ,等 黄土高原典型草原优势种植物及其根际土壤化学计量对降雨变化的响应
[J].生态学报 ,2022 ,42 (3 ):1002 -1010 .
[本文引用: 1]
[6]
Thomey M L Collins C L Vargasr ,et al Effect of precipitation variability on net primary production and soil respiration in a Chihuahuan desert grassland
[J].Global Change Biology ,2011 ,17 (4 ):1505 -1515 .
[本文引用: 1]
[7]
Baez S Collins S L Pockman W T et al Effects of experimental rainfall manipulations on Chihuahuan desert grass land and shrub land plant communities
[J].Oecologia ,2013 ,172 :1117 -1127 .
[本文引用: 1]
[8]
Rao L E Allen E B Combined effects of precipitation and nitrogen deposition on native and invasive winter annual production in California deserts
[J].Oecologia ,2010 ,162 :1035 -1046 .
[本文引用: 1]
[9]
邢彬彬 ,安慧 ,刘姝萱 ,等 降水变化对荒漠草原植物群落生物量及其权衡关系的影响
[J].草地学报 ,2023 ,31 (10 ):3103 -3113 .
[本文引用: 2]
[10]
Gao Y Z Chen Q Lin S et al Resource manipulation effects on net primary production,biomass allocation and rainuse efficiency of two semiarid grassland sites in Inner Mongolia,China
[J].Oecologia ,2011 ,165 :855 -864 .
[本文引用: 1]
[11]
刁励玮 ,李平 ,刘卫星 ,等 草地生态系统生物量在不同气候及多时间尺度上对氮添加和增雨处理的响应
[J].植物生态学报 ,2018 ,42 (8 ):818 -830 .
[本文引用: 1]
[12]
苗百岭 ,梁存柱 ,史亚博 降水变化对内蒙古典型草原地上生物量的影响
[J].植物生态学报 ,2019 ,43 (7 ):557 -565 .
[本文引用: 1]
[13]
张腊梅 ,刘新平 ,赵学勇 科尔沁固定沙地植被特征对降雨变化的响应
[J].生态学报 ,2014 ,34 (10 ):2737 -2745 .
[本文引用: 1]
[14]
姜基春 ,王国强 ,郭宁 ,等 黄土高原4种植被带草本群落特征及其对降水变化的响应
[J].西北植物学报 ,2019 ,39 (10 ):1861 -1867 .
[本文引用: 1]
[15]
Sun J Zhang B Pan Q et al Non‐linear response of productivity to precipitation extremes in the Inner Mongolia grassland
[J].Functional Ecology ,2023 ,37 (6 ):1663 -1673 .
[本文引用: 1]
[16]
Yang Y Fang J Ma W et al Relationship between variability in aboveground net primary production and precipitation in global grasslands
[J].Geophysical Research Letters ,2008 (35 ):1 -4 .
[本文引用: 1]
[17]
车力木格 ,刘新平 ,何玉惠 半干旱沙地草本植物群落特征对短期降水变化的响应
[J].草业学报 ,2020 ,29 (4 ):19 -28 .
[本文引用: 1]
[18]
施晶晶 ,赵鸣飞 ,王宇航 ,等 黄土高原腹地人工林下草本层群落构建机制
[J].植物生态学报 ,2019 ,43 (9 ):1 -9 .
[本文引用: 1]
[19]
彭书婷 ,王秋雪 ,黄柳菁 植物群落构建研究热点与发展趋势分析
[J].山东林业科技 ,2023 ,53 (6 ):30 -39 .
[本文引用: 1]
[20]
Wagg C Roscher C Weigelt A et al.Biodiversity‐stability relationships strengthen over time in a long‐term grassland experiment
[J].Nature Communications ,2022 ,13 :7752 .
[本文引用: 1]
[21]
刘新平 ,何玉惠 ,赵学勇 ,等 科尔沁沙地奈曼地区降水变化特征分析
[J].水土保持研究 ,2011 ,18 (2 ):155 -158 .
[本文引用: 1]
[22]
左小安 ,赵学勇 ,赵哈林 ,等 科尔沁沙质草地群落物种多样性、生产力与土壤特性的关系
[J].环境科学 ,2007 (5 ):945 -951 .
[本文引用: 1]
[23]
连佳丽 ,陈婧 ,杨雪琴 ,等 荒漠草原植物多样性和微生物多样性对降水变化的响应
[J].生物多样性 ,2024 ,32 (6 ):131 -144 .
[本文引用: 2]
[24]
孙岩 ,何明珠 ,王立 降水控制对荒漠植物群落物种多样性和生物量的影响
[J].生态学报 ,2018 ,38 (7 ):2425 -2433 .
[本文引用: 2]
[25]
周璐瑶 氮添加,增温和降雨改变对黄土高原半干旱草地植物群落多样性及生产力的影响
[D].南京 :南京农业大学 ,2021 .
[本文引用: 1]
[26]
韩佳育 羊草和兴安胡枝子功能性状和种间关系对降水格局变化的响应
[D].长春 :东北师范大学 ,2022 .
[本文引用: 1]
[27]
张晶 ,左小安 ,吕朋 土壤水分和养分对沙质草地优势植物叶片氮回收效率的影响
[J].中国沙漠 ,2024 ,44 (1 ):161 -169 .
[本文引用: 1]
[28]
杨浩 ,罗亚晨 糙隐子草功能性状对氮添加和干旱的响应
[J].植物生态学报 ,2015 ,39 (1 ):32 -42 .
[本文引用: 1]
[29]
朱文艳 ,雷思玲 ,庹清 ,等 基于植物群落抗旱演替模型的生态与环境影响因素作用机理量化分析
[J].理论数学 ,2024 ,14 :1 -11 .
[本文引用: 1]
[30]
毛伟 ,李玉霖 ,崔夺 ,等 沙质草地不同生活史植物的生物量分配对氮素和水分添加的响应
[J].植物生态学报 ,2014 ,38 (2 ):125 -133 .
[本文引用: 1]
[31]
崔鹏 ,黄海霞 ,杨琦琦 裸果木幼苗生物量和抗氧化酶活性对土壤干旱胁迫的响应
[J].中国水土保持科学 ,2020 ,18 (5 ):112 -118 .
[本文引用: 1]
[32]
任昱 ,卢琦 ,吴波 ,等 不同模拟增雨下白刺比叶面积和叶干物质含量的比较
[J].生态学报 ,2015 ,35 (14 ):4707 -4715 .
[33]
李素慧 ,陈金艺 ,王佳敏 ,等 两种一年生草本对喀斯特土壤厚度及供水的生长响应
[J].植物科学学报 ,2021 ,39 (3 ):258 -267 .
[34]
王佳敏 ,宋海燕 ,陈金艺 ,等 多年生黑麦草对干旱胁迫下喀斯特异质生境的生长响应策略
[J].生态学报 ,2020 ,40 (13 ):4566 -4572 .
[35]
Diaz S Cabido M Casanoves F Plant functional traits and environmental filters at a regional scale
[J].Journal of Vegetation Science ,1998 ,9 (1 ):113 -122 .
[本文引用: 1]
[36]
李慧 ,何宜璇 ,斯日木极 ,等 根系生态位差异对生态系统的影响
[J].草业科学 ,2021 ,38 (3 ):501 -513 .
[本文引用: 1]
[37]
Wright I J Reich P B Westoby M Strategy shifts in leaf physiology,structure and nutrient content between species of high‐and low‐rainfall and high‐and low‐nutrient habitats
[J].Functional Ecology ,2001 ,15 (4 ):423 -434 .
[本文引用: 1]
[38]
李卓忆 ,杨庆 ,马柱国 ,等 中国北方干旱半干旱区植被对气候变化和人类活动的响应
[J].大气科学 ,2024 ,48 (3 ):859 -874 .
[本文引用: 1]
[39]
张佳华 ,符淙斌 ,延晓冬 ,等 全球植被叶面积指数对温度和降水的响应研究
[J].地球物理学报 ,2002 ,45 (5 ):631 -637 .
[40]
张清雨 ,赵东升 ,吴绍洪 ,等 基于生态分区的内蒙古地区植被覆盖变化及其影响因素研究
[J].地理科学 ,2013 ,33 (5 ):594 -601 .
[41]
张文强 ,罗格平 ,郑宏伟 ,等 基于随机森林模型的内陆干旱区植被指数变化与驱动力分析:以北天山北坡中段为例
[J].植物生态学报 ,2020 ,44 (11 ):1113 -1126 .
[42]
井光花 ,陈智坤 ,路强强 ,等 半干旱黄土区不同管理措施下草地群落结构对短期氮、水添加的响应
[J].生态学报 ,2021 ,41 (20 ):8192 -8201 .
[本文引用: 1]
[43]
王军锋 ,张丽华 ,赵锐锋 ,等 荒漠草原区不同生活型植物生长对降水变化的响应
[J].应用生态学报 ,2020 ,31 (3 ):778 -786 .
[本文引用: 1]
[44]
Hu Y Li X Y Guo A X et al Species diversity is a strong predictor of ecosystem multifunctionality under altered precipitation in desert steppes
[J].Ecological Indicators ,2022 ,6 :1 -14 .
[本文引用: 1]
[45]
王百竹 ,白建华 ,萨拉 ,等 呼伦贝尔沙化草地不同放牧潜力阶段群落特征
[J].中国沙漠 ,2025 ,45 (2 ):205 -216 .
[本文引用: 2]
2000-2017年内蒙古荒漠草原植被物候和净初级生产力对气候变化的响应
1
2020
... 半干旱沙质草地是典型草原向荒漠过渡的旱生化草地生态系统[1 ] ,是干旱半干旱地区陆地生态系统的主体部分[2 ] ,也是中国重要的草地生态系统[3 ] .半干旱沙质草地极易受全球变化影响,且生态系统功能和稳定性较差[4 ] .降水是影响干旱半干旱地区沙质草地生态系统中植被生长的主要限制因子[5 ] .固沙植被在维持生态系统结构稳定中发挥着至关重要的作用,对降水变化的响应极为敏感[6 -7 ] .此外,降水也是制约沙质草地植被生产力和植被恢复的关键环境因子[8 ] .近年来由于气候变化和人类活动共同作用,中国旱区出现土地退化、水分流失和生物多样性丧失等问题[9 ] .因此,研究降水变化对沙质草地植被特征及其群落构建的影响,对于植被恢复重建和稳定维持具有重要意义. ...
短花针茅草原不同载畜率对植物多样性和草地生产力的影响
1
2007
... 半干旱沙质草地是典型草原向荒漠过渡的旱生化草地生态系统[1 ] ,是干旱半干旱地区陆地生态系统的主体部分[2 ] ,也是中国重要的草地生态系统[3 ] .半干旱沙质草地极易受全球变化影响,且生态系统功能和稳定性较差[4 ] .降水是影响干旱半干旱地区沙质草地生态系统中植被生长的主要限制因子[5 ] .固沙植被在维持生态系统结构稳定中发挥着至关重要的作用,对降水变化的响应极为敏感[6 -7 ] .此外,降水也是制约沙质草地植被生产力和植被恢复的关键环境因子[8 ] .近年来由于气候变化和人类活动共同作用,中国旱区出现土地退化、水分流失和生物多样性丧失等问题[9 ] .因此,研究降水变化对沙质草地植被特征及其群落构建的影响,对于植被恢复重建和稳定维持具有重要意义. ...
极端干旱对典型草原土壤与植被的影响
1
2022
... 半干旱沙质草地是典型草原向荒漠过渡的旱生化草地生态系统[1 ] ,是干旱半干旱地区陆地生态系统的主体部分[2 ] ,也是中国重要的草地生态系统[3 ] .半干旱沙质草地极易受全球变化影响,且生态系统功能和稳定性较差[4 ] .降水是影响干旱半干旱地区沙质草地生态系统中植被生长的主要限制因子[5 ] .固沙植被在维持生态系统结构稳定中发挥着至关重要的作用,对降水变化的响应极为敏感[6 -7 ] .此外,降水也是制约沙质草地植被生产力和植被恢复的关键环境因子[8 ] .近年来由于气候变化和人类活动共同作用,中国旱区出现土地退化、水分流失和生物多样性丧失等问题[9 ] .因此,研究降水变化对沙质草地植被特征及其群落构建的影响,对于植被恢复重建和稳定维持具有重要意义. ...
乌拉特荒漠草原群落物种多样性和生物量关系对放牧强度的响应
1
2020
... 半干旱沙质草地是典型草原向荒漠过渡的旱生化草地生态系统[1 ] ,是干旱半干旱地区陆地生态系统的主体部分[2 ] ,也是中国重要的草地生态系统[3 ] .半干旱沙质草地极易受全球变化影响,且生态系统功能和稳定性较差[4 ] .降水是影响干旱半干旱地区沙质草地生态系统中植被生长的主要限制因子[5 ] .固沙植被在维持生态系统结构稳定中发挥着至关重要的作用,对降水变化的响应极为敏感[6 -7 ] .此外,降水也是制约沙质草地植被生产力和植被恢复的关键环境因子[8 ] .近年来由于气候变化和人类活动共同作用,中国旱区出现土地退化、水分流失和生物多样性丧失等问题[9 ] .因此,研究降水变化对沙质草地植被特征及其群落构建的影响,对于植被恢复重建和稳定维持具有重要意义. ...
黄土高原典型草原优势种植物及其根际土壤化学计量对降雨变化的响应
1
2022
... 半干旱沙质草地是典型草原向荒漠过渡的旱生化草地生态系统[1 ] ,是干旱半干旱地区陆地生态系统的主体部分[2 ] ,也是中国重要的草地生态系统[3 ] .半干旱沙质草地极易受全球变化影响,且生态系统功能和稳定性较差[4 ] .降水是影响干旱半干旱地区沙质草地生态系统中植被生长的主要限制因子[5 ] .固沙植被在维持生态系统结构稳定中发挥着至关重要的作用,对降水变化的响应极为敏感[6 -7 ] .此外,降水也是制约沙质草地植被生产力和植被恢复的关键环境因子[8 ] .近年来由于气候变化和人类活动共同作用,中国旱区出现土地退化、水分流失和生物多样性丧失等问题[9 ] .因此,研究降水变化对沙质草地植被特征及其群落构建的影响,对于植被恢复重建和稳定维持具有重要意义. ...
Effect of precipitation variability on net primary production and soil respiration in a Chihuahuan desert grassland
1
2011
... 半干旱沙质草地是典型草原向荒漠过渡的旱生化草地生态系统[1 ] ,是干旱半干旱地区陆地生态系统的主体部分[2 ] ,也是中国重要的草地生态系统[3 ] .半干旱沙质草地极易受全球变化影响,且生态系统功能和稳定性较差[4 ] .降水是影响干旱半干旱地区沙质草地生态系统中植被生长的主要限制因子[5 ] .固沙植被在维持生态系统结构稳定中发挥着至关重要的作用,对降水变化的响应极为敏感[6 -7 ] .此外,降水也是制约沙质草地植被生产力和植被恢复的关键环境因子[8 ] .近年来由于气候变化和人类活动共同作用,中国旱区出现土地退化、水分流失和生物多样性丧失等问题[9 ] .因此,研究降水变化对沙质草地植被特征及其群落构建的影响,对于植被恢复重建和稳定维持具有重要意义. ...
Effects of experimental rainfall manipulations on Chihuahuan desert grass land and shrub land plant communities
1
2013
... 半干旱沙质草地是典型草原向荒漠过渡的旱生化草地生态系统[1 ] ,是干旱半干旱地区陆地生态系统的主体部分[2 ] ,也是中国重要的草地生态系统[3 ] .半干旱沙质草地极易受全球变化影响,且生态系统功能和稳定性较差[4 ] .降水是影响干旱半干旱地区沙质草地生态系统中植被生长的主要限制因子[5 ] .固沙植被在维持生态系统结构稳定中发挥着至关重要的作用,对降水变化的响应极为敏感[6 -7 ] .此外,降水也是制约沙质草地植被生产力和植被恢复的关键环境因子[8 ] .近年来由于气候变化和人类活动共同作用,中国旱区出现土地退化、水分流失和生物多样性丧失等问题[9 ] .因此,研究降水变化对沙质草地植被特征及其群落构建的影响,对于植被恢复重建和稳定维持具有重要意义. ...
Combined effects of precipitation and nitrogen deposition on native and invasive winter annual production in California deserts
1
2010
... 半干旱沙质草地是典型草原向荒漠过渡的旱生化草地生态系统[1 ] ,是干旱半干旱地区陆地生态系统的主体部分[2 ] ,也是中国重要的草地生态系统[3 ] .半干旱沙质草地极易受全球变化影响,且生态系统功能和稳定性较差[4 ] .降水是影响干旱半干旱地区沙质草地生态系统中植被生长的主要限制因子[5 ] .固沙植被在维持生态系统结构稳定中发挥着至关重要的作用,对降水变化的响应极为敏感[6 -7 ] .此外,降水也是制约沙质草地植被生产力和植被恢复的关键环境因子[8 ] .近年来由于气候变化和人类活动共同作用,中国旱区出现土地退化、水分流失和生物多样性丧失等问题[9 ] .因此,研究降水变化对沙质草地植被特征及其群落构建的影响,对于植被恢复重建和稳定维持具有重要意义. ...
降水变化对荒漠草原植物群落生物量及其权衡关系的影响
2
2023
... 半干旱沙质草地是典型草原向荒漠过渡的旱生化草地生态系统[1 ] ,是干旱半干旱地区陆地生态系统的主体部分[2 ] ,也是中国重要的草地生态系统[3 ] .半干旱沙质草地极易受全球变化影响,且生态系统功能和稳定性较差[4 ] .降水是影响干旱半干旱地区沙质草地生态系统中植被生长的主要限制因子[5 ] .固沙植被在维持生态系统结构稳定中发挥着至关重要的作用,对降水变化的响应极为敏感[6 -7 ] .此外,降水也是制约沙质草地植被生产力和植被恢复的关键环境因子[8 ] .近年来由于气候变化和人类活动共同作用,中国旱区出现土地退化、水分流失和生物多样性丧失等问题[9 ] .因此,研究降水变化对沙质草地植被特征及其群落构建的影响,对于植被恢复重建和稳定维持具有重要意义. ...
... 降水变化对植物群落构建的影响可以从物种多样性、功能群组成和生态位动态等多个维度进行剖析.首先,降水变化可能通过影响物种的繁殖、生长和存活,进而改变群落的物种组成和多样性.例如,降水的增加可能促进某些物种的生长,但同时也会抑制其他物种的竞争力,导致群落物种多样性降低[44 ] .降水变化对不同功能群的影响也存在差异.一年生草本植物通常对短期降水变化更为敏感,而多年生草本植物则表现出更强的适应性[45 ] .这种差异可能导致功能群之间的竞争格局发生变化,进而影响群落的结构和功能[9 ] .此外,降水变化还可能通过改变植物的生态位宽度和重叠程度,影响物种之间的相互作用和群落的稳定性[23 ] .例如,降水量的减少可能使植物的生态位更加狭窄,导致物种之间的竞争加剧,而降水量的增加则可能扩大生态位宽度,促进物种共存[45 ] .不同功能群对水分利用的策略存在差异,导致各功能群植物地上生物量在群落中的比例发生改变.这种生物量分配策略的变化反映了植物群落对降水变化的适应机制.本研究探讨了不同功能群在不同降水条件下的分布和竞争关系.在40%的年降水变异条件下,禾本科植物糙隐子草和豆科植物兴安胡枝子等表现出较强的竞争优势并占据更优生态位,而随着降水变异增加到50%,菊科植物猪毛蒿和禾本科植物白草逐渐成为优势种.这表明降水变化显著影响了功能群之间的竞争格局,进而改变了群落的物种组成和优势度分布,这些变化对群落稳定性和生态系统功能可能产生重要影响.功能群多样性的变化反映了群落对环境胁迫的响应机制,功能群的调整有助于维持生态系统生产力的稳定性.例如,功能群均匀度的离散程度在高降水变异条件下显著增加,这可能削弱了群落的整体稳定性.此外,功能群之间的竞争关系也对生态系统功能产生影响,禾本科植物的生物量与群落总生物量正相关,表明其在生态系统生产力中占据主导地位. ...
Resource manipulation effects on net primary production,biomass allocation and rainuse efficiency of two semiarid grassland sites in Inner Mongolia,China
1
2011
... 植物生物量是生态系统生产力的重要指征[10 -11 ] ,但降水变化与生产力之间的关系仍存在争议.降水增加会显著提高内蒙古典型草原植物群落地上生物量[12 ] ,但在科尔沁固定沙地[13 ] 和黄土高原草原-荒漠带[14 ] ,降水增加过度反而会降低植物群落生物量.同时,草原生态系统生产力对极端干旱和极端湿润处理的响应并非对称[15 ] ,极端干旱处理下草原生产力的降低显著大于极端湿润处理下生产力的增加,且年降水量与之的线性增加关系随气候由干旱向湿润逐渐减弱[16 ] .此外,半干旱沙地植被群落构建还与降水的季节分配密切相关[17 ] ,尤其是生长季初期的降水直接决定了沙质草地固沙植被幼苗建植和物种组成.因此,由于草地类型、降水量和降水时间等非生物因素的不同,草原生态系统植物生物量对降水变化的响应还存在不确定性. ...
草地生态系统生物量在不同气候及多时间尺度上对氮添加和增雨处理的响应
1
2018
... 植物生物量是生态系统生产力的重要指征[10 -11 ] ,但降水变化与生产力之间的关系仍存在争议.降水增加会显著提高内蒙古典型草原植物群落地上生物量[12 ] ,但在科尔沁固定沙地[13 ] 和黄土高原草原-荒漠带[14 ] ,降水增加过度反而会降低植物群落生物量.同时,草原生态系统生产力对极端干旱和极端湿润处理的响应并非对称[15 ] ,极端干旱处理下草原生产力的降低显著大于极端湿润处理下生产力的增加,且年降水量与之的线性增加关系随气候由干旱向湿润逐渐减弱[16 ] .此外,半干旱沙地植被群落构建还与降水的季节分配密切相关[17 ] ,尤其是生长季初期的降水直接决定了沙质草地固沙植被幼苗建植和物种组成.因此,由于草地类型、降水量和降水时间等非生物因素的不同,草原生态系统植物生物量对降水变化的响应还存在不确定性. ...
降水变化对内蒙古典型草原地上生物量的影响
1
2019
... 植物生物量是生态系统生产力的重要指征[10 -11 ] ,但降水变化与生产力之间的关系仍存在争议.降水增加会显著提高内蒙古典型草原植物群落地上生物量[12 ] ,但在科尔沁固定沙地[13 ] 和黄土高原草原-荒漠带[14 ] ,降水增加过度反而会降低植物群落生物量.同时,草原生态系统生产力对极端干旱和极端湿润处理的响应并非对称[15 ] ,极端干旱处理下草原生产力的降低显著大于极端湿润处理下生产力的增加,且年降水量与之的线性增加关系随气候由干旱向湿润逐渐减弱[16 ] .此外,半干旱沙地植被群落构建还与降水的季节分配密切相关[17 ] ,尤其是生长季初期的降水直接决定了沙质草地固沙植被幼苗建植和物种组成.因此,由于草地类型、降水量和降水时间等非生物因素的不同,草原生态系统植物生物量对降水变化的响应还存在不确定性. ...
科尔沁固定沙地植被特征对降雨变化的响应
1
2014
... 植物生物量是生态系统生产力的重要指征[10 -11 ] ,但降水变化与生产力之间的关系仍存在争议.降水增加会显著提高内蒙古典型草原植物群落地上生物量[12 ] ,但在科尔沁固定沙地[13 ] 和黄土高原草原-荒漠带[14 ] ,降水增加过度反而会降低植物群落生物量.同时,草原生态系统生产力对极端干旱和极端湿润处理的响应并非对称[15 ] ,极端干旱处理下草原生产力的降低显著大于极端湿润处理下生产力的增加,且年降水量与之的线性增加关系随气候由干旱向湿润逐渐减弱[16 ] .此外,半干旱沙地植被群落构建还与降水的季节分配密切相关[17 ] ,尤其是生长季初期的降水直接决定了沙质草地固沙植被幼苗建植和物种组成.因此,由于草地类型、降水量和降水时间等非生物因素的不同,草原生态系统植物生物量对降水变化的响应还存在不确定性. ...
黄土高原4种植被带草本群落特征及其对降水变化的响应
1
2019
... 植物生物量是生态系统生产力的重要指征[10 -11 ] ,但降水变化与生产力之间的关系仍存在争议.降水增加会显著提高内蒙古典型草原植物群落地上生物量[12 ] ,但在科尔沁固定沙地[13 ] 和黄土高原草原-荒漠带[14 ] ,降水增加过度反而会降低植物群落生物量.同时,草原生态系统生产力对极端干旱和极端湿润处理的响应并非对称[15 ] ,极端干旱处理下草原生产力的降低显著大于极端湿润处理下生产力的增加,且年降水量与之的线性增加关系随气候由干旱向湿润逐渐减弱[16 ] .此外,半干旱沙地植被群落构建还与降水的季节分配密切相关[17 ] ,尤其是生长季初期的降水直接决定了沙质草地固沙植被幼苗建植和物种组成.因此,由于草地类型、降水量和降水时间等非生物因素的不同,草原生态系统植物生物量对降水变化的响应还存在不确定性. ...
Non‐linear response of productivity to precipitation extremes in the Inner Mongolia grassland
1
2023
... 植物生物量是生态系统生产力的重要指征[10 -11 ] ,但降水变化与生产力之间的关系仍存在争议.降水增加会显著提高内蒙古典型草原植物群落地上生物量[12 ] ,但在科尔沁固定沙地[13 ] 和黄土高原草原-荒漠带[14 ] ,降水增加过度反而会降低植物群落生物量.同时,草原生态系统生产力对极端干旱和极端湿润处理的响应并非对称[15 ] ,极端干旱处理下草原生产力的降低显著大于极端湿润处理下生产力的增加,且年降水量与之的线性增加关系随气候由干旱向湿润逐渐减弱[16 ] .此外,半干旱沙地植被群落构建还与降水的季节分配密切相关[17 ] ,尤其是生长季初期的降水直接决定了沙质草地固沙植被幼苗建植和物种组成.因此,由于草地类型、降水量和降水时间等非生物因素的不同,草原生态系统植物生物量对降水变化的响应还存在不确定性. ...
Relationship between variability in aboveground net primary production and precipitation in global grasslands
1
2008
... 植物生物量是生态系统生产力的重要指征[10 -11 ] ,但降水变化与生产力之间的关系仍存在争议.降水增加会显著提高内蒙古典型草原植物群落地上生物量[12 ] ,但在科尔沁固定沙地[13 ] 和黄土高原草原-荒漠带[14 ] ,降水增加过度反而会降低植物群落生物量.同时,草原生态系统生产力对极端干旱和极端湿润处理的响应并非对称[15 ] ,极端干旱处理下草原生产力的降低显著大于极端湿润处理下生产力的增加,且年降水量与之的线性增加关系随气候由干旱向湿润逐渐减弱[16 ] .此外,半干旱沙地植被群落构建还与降水的季节分配密切相关[17 ] ,尤其是生长季初期的降水直接决定了沙质草地固沙植被幼苗建植和物种组成.因此,由于草地类型、降水量和降水时间等非生物因素的不同,草原生态系统植物生物量对降水变化的响应还存在不确定性. ...
半干旱沙地草本植物群落特征对短期降水变化的响应
1
2020
... 植物生物量是生态系统生产力的重要指征[10 -11 ] ,但降水变化与生产力之间的关系仍存在争议.降水增加会显著提高内蒙古典型草原植物群落地上生物量[12 ] ,但在科尔沁固定沙地[13 ] 和黄土高原草原-荒漠带[14 ] ,降水增加过度反而会降低植物群落生物量.同时,草原生态系统生产力对极端干旱和极端湿润处理的响应并非对称[15 ] ,极端干旱处理下草原生产力的降低显著大于极端湿润处理下生产力的增加,且年降水量与之的线性增加关系随气候由干旱向湿润逐渐减弱[16 ] .此外,半干旱沙地植被群落构建还与降水的季节分配密切相关[17 ] ,尤其是生长季初期的降水直接决定了沙质草地固沙植被幼苗建植和物种组成.因此,由于草地类型、降水量和降水时间等非生物因素的不同,草原生态系统植物生物量对降水变化的响应还存在不确定性. ...
黄土高原腹地人工林下草本层群落构建机制
1
2019
... 近年来,植物群落构建机制的研究逐渐成为生态学领域的热点.黄土高原腹地人工林下草本层群落构建机制的研究表明,生态位分化和中性过程的共同作用是影响群落构建的关键因素[18 ] .此外,功能群多样性在群落构建中也发挥着重要作用,其通过影响植物对资源的获取和利用效率,调节群落的稳定性和功能[19 ] .功能群多样性不仅能够增强植物群落对环境变化的适应能力,还能通过生态位互补和功能冗余机制,提高群落的整体生产力和稳定性[20 ] .然而,当前研究大多集中在植物群落多样性及其生物量的动态变化上,而关于优势种和共有种的生物量对高变异降水响应的研究相对匮乏.此外,功能群多样性在群落构建中的作用及其对环境变化的响应机制尚未得到充分探讨.另外,这方面的研究目前大多关注植物群落多样性及其生物量的动态变化,而关于优势种和共有种的生物量响应高变异降水的研究相对匮乏. ...
植物群落构建研究热点与发展趋势分析
1
2023
... 近年来,植物群落构建机制的研究逐渐成为生态学领域的热点.黄土高原腹地人工林下草本层群落构建机制的研究表明,生态位分化和中性过程的共同作用是影响群落构建的关键因素[18 ] .此外,功能群多样性在群落构建中也发挥着重要作用,其通过影响植物对资源的获取和利用效率,调节群落的稳定性和功能[19 ] .功能群多样性不仅能够增强植物群落对环境变化的适应能力,还能通过生态位互补和功能冗余机制,提高群落的整体生产力和稳定性[20 ] .然而,当前研究大多集中在植物群落多样性及其生物量的动态变化上,而关于优势种和共有种的生物量对高变异降水响应的研究相对匮乏.此外,功能群多样性在群落构建中的作用及其对环境变化的响应机制尚未得到充分探讨.另外,这方面的研究目前大多关注植物群落多样性及其生物量的动态变化,而关于优势种和共有种的生物量响应高变异降水的研究相对匮乏. ...
et al.Biodiversity‐stability relationships strengthen over time in a long‐term grassland experiment
1
2022
... 近年来,植物群落构建机制的研究逐渐成为生态学领域的热点.黄土高原腹地人工林下草本层群落构建机制的研究表明,生态位分化和中性过程的共同作用是影响群落构建的关键因素[18 ] .此外,功能群多样性在群落构建中也发挥着重要作用,其通过影响植物对资源的获取和利用效率,调节群落的稳定性和功能[19 ] .功能群多样性不仅能够增强植物群落对环境变化的适应能力,还能通过生态位互补和功能冗余机制,提高群落的整体生产力和稳定性[20 ] .然而,当前研究大多集中在植物群落多样性及其生物量的动态变化上,而关于优势种和共有种的生物量对高变异降水响应的研究相对匮乏.此外,功能群多样性在群落构建中的作用及其对环境变化的响应机制尚未得到充分探讨.另外,这方面的研究目前大多关注植物群落多样性及其生物量的动态变化,而关于优势种和共有种的生物量响应高变异降水的研究相对匮乏. ...
科尔沁沙地奈曼地区降水变化特征分析
1
2011
... 研究区为内蒙古奈曼站沙地综合观测场(42°94′N、120°71′E,海拔约为353 m),位于科尔沁沙地南缘的奈曼旗境内.该地属于温带半干旱大陆性气候,年平均气温为5.8~6.4 ℃,多年平均降水量351.7 mm[21 ] ,年际降水变率大且分布不均,其中80%的降水量集中在4—8月.土壤类型主要为风沙土、沙质栗钙,土质疏松,保水能力较差,土壤有机质含量较低.地貌类型为流动沙丘、半固定沙丘、固定沙丘、平缓沙地等[22 ] .该区草地植被以一年生或者多年生草本植物为主,主要优势植物有狗尾草(Setaria viridis )、尖头叶藜(Chenopodium acuminatum )、白草(Pennisetum centrasiaticum )和猪毛蒿(Artemisia scoparia )等.实验样地选址在科尔沁沙地的典型沙质草地,该区域植被覆盖度较低,受人类干扰较小,能够较好地反映自然条件下植物群落对降水变化的响应. ...
科尔沁沙质草地群落物种多样性、生产力与土壤特性的关系
1
2007
... 研究区为内蒙古奈曼站沙地综合观测场(42°94′N、120°71′E,海拔约为353 m),位于科尔沁沙地南缘的奈曼旗境内.该地属于温带半干旱大陆性气候,年平均气温为5.8~6.4 ℃,多年平均降水量351.7 mm[21 ] ,年际降水变率大且分布不均,其中80%的降水量集中在4—8月.土壤类型主要为风沙土、沙质栗钙,土质疏松,保水能力较差,土壤有机质含量较低.地貌类型为流动沙丘、半固定沙丘、固定沙丘、平缓沙地等[22 ] .该区草地植被以一年生或者多年生草本植物为主,主要优势植物有狗尾草(Setaria viridis )、尖头叶藜(Chenopodium acuminatum )、白草(Pennisetum centrasiaticum )和猪毛蒿(Artemisia scoparia )等.实验样地选址在科尔沁沙地的典型沙质草地,该区域植被覆盖度较低,受人类干扰较小,能够较好地反映自然条件下植物群落对降水变化的响应. ...
荒漠草原植物多样性和微生物多样性对降水变化的响应
2
2024
... 相对重要值(IV )反映每个物种在群落中的重要性,植物群落α多样性采用Shannon-Wiener指数(H )、Margalef指数(R )、Simpson指数(D )及Pielou指数(J )表征[23 ] . ...
... 降水变化对植物群落构建的影响可以从物种多样性、功能群组成和生态位动态等多个维度进行剖析.首先,降水变化可能通过影响物种的繁殖、生长和存活,进而改变群落的物种组成和多样性.例如,降水的增加可能促进某些物种的生长,但同时也会抑制其他物种的竞争力,导致群落物种多样性降低[44 ] .降水变化对不同功能群的影响也存在差异.一年生草本植物通常对短期降水变化更为敏感,而多年生草本植物则表现出更强的适应性[45 ] .这种差异可能导致功能群之间的竞争格局发生变化,进而影响群落的结构和功能[9 ] .此外,降水变化还可能通过改变植物的生态位宽度和重叠程度,影响物种之间的相互作用和群落的稳定性[23 ] .例如,降水量的减少可能使植物的生态位更加狭窄,导致物种之间的竞争加剧,而降水量的增加则可能扩大生态位宽度,促进物种共存[45 ] .不同功能群对水分利用的策略存在差异,导致各功能群植物地上生物量在群落中的比例发生改变.这种生物量分配策略的变化反映了植物群落对降水变化的适应机制.本研究探讨了不同功能群在不同降水条件下的分布和竞争关系.在40%的年降水变异条件下,禾本科植物糙隐子草和豆科植物兴安胡枝子等表现出较强的竞争优势并占据更优生态位,而随着降水变异增加到50%,菊科植物猪毛蒿和禾本科植物白草逐渐成为优势种.这表明降水变化显著影响了功能群之间的竞争格局,进而改变了群落的物种组成和优势度分布,这些变化对群落稳定性和生态系统功能可能产生重要影响.功能群多样性的变化反映了群落对环境胁迫的响应机制,功能群的调整有助于维持生态系统生产力的稳定性.例如,功能群均匀度的离散程度在高降水变异条件下显著增加,这可能削弱了群落的整体稳定性.此外,功能群之间的竞争关系也对生态系统功能产生影响,禾本科植物的生物量与群落总生物量正相关,表明其在生态系统生产力中占据主导地位. ...
降水控制对荒漠植物群落物种多样性和生物量的影响
2
2018
... 植物群落多样性是衡量物种分布均匀程度和数量的指标,是群落特征的综合体现[24 ] .本研究结果表明,功能群的丰富度、优势度和均匀度随着降水增加呈现上升趋势,而多样性的变动与之相反.增雨会使植物群落的多样性降低,原因可能是降雨量的增加有助于一年生草本植物萌发并促进生长势,使得杂类草植物多度减少,植物群落中物种多样性由此降低.这一结果与周璐瑶等[25 ] 的研究一致,其指出氮添加与增雨处理相结合显著降低了群落丰富度,进而影响植物群落的多样性.孙岩等[24 ] 发现在沙漠地区增雨可能对某些植物群落的生物量有积极影响,但不会增加群落多样性.这表明降水变化对群落多样性的影响可能因植物群落类型和环境背景而异.同时,在群落多样性减少的状况下,有些植物如糙隐子草、二裂委陵菜、狗尾草、尖头叶藜和兴安胡枝子等受降水变化的影响较小.这与韩佳育[26 ] 、张晶等[27 ] 、杨浩等[28 ] 的研究结果相似,即这些植物对降水变化不敏感且具有较强的适应性.这种现象可能与植物的生态位分化有关,即这些物种在长期进化过程中形成了对干旱或湿润环境的适应策略,从而在降水变化下仍能保持相对稳定的地位[29 ] . ...
... [24 ]发现在沙漠地区增雨可能对某些植物群落的生物量有积极影响,但不会增加群落多样性.这表明降水变化对群落多样性的影响可能因植物群落类型和环境背景而异.同时,在群落多样性减少的状况下,有些植物如糙隐子草、二裂委陵菜、狗尾草、尖头叶藜和兴安胡枝子等受降水变化的影响较小.这与韩佳育[26 ] 、张晶等[27 ] 、杨浩等[28 ] 的研究结果相似,即这些植物对降水变化不敏感且具有较强的适应性.这种现象可能与植物的生态位分化有关,即这些物种在长期进化过程中形成了对干旱或湿润环境的适应策略,从而在降水变化下仍能保持相对稳定的地位[29 ] . ...
氮添加,增温和降雨改变对黄土高原半干旱草地植物群落多样性及生产力的影响
1
2021
... 植物群落多样性是衡量物种分布均匀程度和数量的指标,是群落特征的综合体现[24 ] .本研究结果表明,功能群的丰富度、优势度和均匀度随着降水增加呈现上升趋势,而多样性的变动与之相反.增雨会使植物群落的多样性降低,原因可能是降雨量的增加有助于一年生草本植物萌发并促进生长势,使得杂类草植物多度减少,植物群落中物种多样性由此降低.这一结果与周璐瑶等[25 ] 的研究一致,其指出氮添加与增雨处理相结合显著降低了群落丰富度,进而影响植物群落的多样性.孙岩等[24 ] 发现在沙漠地区增雨可能对某些植物群落的生物量有积极影响,但不会增加群落多样性.这表明降水变化对群落多样性的影响可能因植物群落类型和环境背景而异.同时,在群落多样性减少的状况下,有些植物如糙隐子草、二裂委陵菜、狗尾草、尖头叶藜和兴安胡枝子等受降水变化的影响较小.这与韩佳育[26 ] 、张晶等[27 ] 、杨浩等[28 ] 的研究结果相似,即这些植物对降水变化不敏感且具有较强的适应性.这种现象可能与植物的生态位分化有关,即这些物种在长期进化过程中形成了对干旱或湿润环境的适应策略,从而在降水变化下仍能保持相对稳定的地位[29 ] . ...
羊草和兴安胡枝子功能性状和种间关系对降水格局变化的响应
1
2022
... 植物群落多样性是衡量物种分布均匀程度和数量的指标,是群落特征的综合体现[24 ] .本研究结果表明,功能群的丰富度、优势度和均匀度随着降水增加呈现上升趋势,而多样性的变动与之相反.增雨会使植物群落的多样性降低,原因可能是降雨量的增加有助于一年生草本植物萌发并促进生长势,使得杂类草植物多度减少,植物群落中物种多样性由此降低.这一结果与周璐瑶等[25 ] 的研究一致,其指出氮添加与增雨处理相结合显著降低了群落丰富度,进而影响植物群落的多样性.孙岩等[24 ] 发现在沙漠地区增雨可能对某些植物群落的生物量有积极影响,但不会增加群落多样性.这表明降水变化对群落多样性的影响可能因植物群落类型和环境背景而异.同时,在群落多样性减少的状况下,有些植物如糙隐子草、二裂委陵菜、狗尾草、尖头叶藜和兴安胡枝子等受降水变化的影响较小.这与韩佳育[26 ] 、张晶等[27 ] 、杨浩等[28 ] 的研究结果相似,即这些植物对降水变化不敏感且具有较强的适应性.这种现象可能与植物的生态位分化有关,即这些物种在长期进化过程中形成了对干旱或湿润环境的适应策略,从而在降水变化下仍能保持相对稳定的地位[29 ] . ...
土壤水分和养分对沙质草地优势植物叶片氮回收效率的影响
1
2024
... 植物群落多样性是衡量物种分布均匀程度和数量的指标,是群落特征的综合体现[24 ] .本研究结果表明,功能群的丰富度、优势度和均匀度随着降水增加呈现上升趋势,而多样性的变动与之相反.增雨会使植物群落的多样性降低,原因可能是降雨量的增加有助于一年生草本植物萌发并促进生长势,使得杂类草植物多度减少,植物群落中物种多样性由此降低.这一结果与周璐瑶等[25 ] 的研究一致,其指出氮添加与增雨处理相结合显著降低了群落丰富度,进而影响植物群落的多样性.孙岩等[24 ] 发现在沙漠地区增雨可能对某些植物群落的生物量有积极影响,但不会增加群落多样性.这表明降水变化对群落多样性的影响可能因植物群落类型和环境背景而异.同时,在群落多样性减少的状况下,有些植物如糙隐子草、二裂委陵菜、狗尾草、尖头叶藜和兴安胡枝子等受降水变化的影响较小.这与韩佳育[26 ] 、张晶等[27 ] 、杨浩等[28 ] 的研究结果相似,即这些植物对降水变化不敏感且具有较强的适应性.这种现象可能与植物的生态位分化有关,即这些物种在长期进化过程中形成了对干旱或湿润环境的适应策略,从而在降水变化下仍能保持相对稳定的地位[29 ] . ...
糙隐子草功能性状对氮添加和干旱的响应
1
2015
... 植物群落多样性是衡量物种分布均匀程度和数量的指标,是群落特征的综合体现[24 ] .本研究结果表明,功能群的丰富度、优势度和均匀度随着降水增加呈现上升趋势,而多样性的变动与之相反.增雨会使植物群落的多样性降低,原因可能是降雨量的增加有助于一年生草本植物萌发并促进生长势,使得杂类草植物多度减少,植物群落中物种多样性由此降低.这一结果与周璐瑶等[25 ] 的研究一致,其指出氮添加与增雨处理相结合显著降低了群落丰富度,进而影响植物群落的多样性.孙岩等[24 ] 发现在沙漠地区增雨可能对某些植物群落的生物量有积极影响,但不会增加群落多样性.这表明降水变化对群落多样性的影响可能因植物群落类型和环境背景而异.同时,在群落多样性减少的状况下,有些植物如糙隐子草、二裂委陵菜、狗尾草、尖头叶藜和兴安胡枝子等受降水变化的影响较小.这与韩佳育[26 ] 、张晶等[27 ] 、杨浩等[28 ] 的研究结果相似,即这些植物对降水变化不敏感且具有较强的适应性.这种现象可能与植物的生态位分化有关,即这些物种在长期进化过程中形成了对干旱或湿润环境的适应策略,从而在降水变化下仍能保持相对稳定的地位[29 ] . ...
基于植物群落抗旱演替模型的生态与环境影响因素作用机理量化分析
1
2024
... 植物群落多样性是衡量物种分布均匀程度和数量的指标,是群落特征的综合体现[24 ] .本研究结果表明,功能群的丰富度、优势度和均匀度随着降水增加呈现上升趋势,而多样性的变动与之相反.增雨会使植物群落的多样性降低,原因可能是降雨量的增加有助于一年生草本植物萌发并促进生长势,使得杂类草植物多度减少,植物群落中物种多样性由此降低.这一结果与周璐瑶等[25 ] 的研究一致,其指出氮添加与增雨处理相结合显著降低了群落丰富度,进而影响植物群落的多样性.孙岩等[24 ] 发现在沙漠地区增雨可能对某些植物群落的生物量有积极影响,但不会增加群落多样性.这表明降水变化对群落多样性的影响可能因植物群落类型和环境背景而异.同时,在群落多样性减少的状况下,有些植物如糙隐子草、二裂委陵菜、狗尾草、尖头叶藜和兴安胡枝子等受降水变化的影响较小.这与韩佳育[26 ] 、张晶等[27 ] 、杨浩等[28 ] 的研究结果相似,即这些植物对降水变化不敏感且具有较强的适应性.这种现象可能与植物的生态位分化有关,即这些物种在长期进化过程中形成了对干旱或湿润环境的适应策略,从而在降水变化下仍能保持相对稳定的地位[29 ] . ...
沙质草地不同生活史植物的生物量分配对氮素和水分添加的响应
1
2014
... 生物量是衡量沙质草地植物生长发育状况的重要指标,直接反映植物的生长和生产力[30 ] .本研究结果表明,降水量减少使植物群落地上生物量显著降低,原因可能是为应对干旱带来的危害,植物调节植株高度、叶面积和叶片干物质含量,导致植物地上部分积累的干物质减少,从而降低了植物地上生物量[31 -35 ] .不同植物或功能群对水分利用策略的差异,导致其竞争能力和优势等级的不同,最终造成植物群落结构和组成比例发生改变.对于一年生草本植物而言,其生物量动态与年降水量变化趋势相反,与多年生草本植物的生物量呈现互补趋势,这是由于降水量减少影响了多年生植物以及半灌木的生长,从而导致其失去竞争力,促进一年生草本植物生长提高地上生物量;一年生草本植物对干旱环境具有一定的适应能力,轻度干旱胁迫对某些一年生草本植物生长影响较小,甚至有一定的促进作用.李慧等[36 ] 指出,组合群落会使植物的生物量和互补效应显著增加,在不同植物组合中,一年生和多年生草本植物的生物量存在互补性,二者组合可以增加总体生物量.美国明尼苏达州长期草地生物多样性试验也印证了我们的观点[37 ] ,即半干旱沙质草地多年生草本植物与一年生草本植物形成互补,共同提高植物群落生物量.这种生物量的互补性可能与植物的生态位分化和资源利用策略有关,未来研究可以进一步探讨其背后的生态机制. ...
裸果木幼苗生物量和抗氧化酶活性对土壤干旱胁迫的响应
1
2020
... 生物量是衡量沙质草地植物生长发育状况的重要指标,直接反映植物的生长和生产力[30 ] .本研究结果表明,降水量减少使植物群落地上生物量显著降低,原因可能是为应对干旱带来的危害,植物调节植株高度、叶面积和叶片干物质含量,导致植物地上部分积累的干物质减少,从而降低了植物地上生物量[31 -35 ] .不同植物或功能群对水分利用策略的差异,导致其竞争能力和优势等级的不同,最终造成植物群落结构和组成比例发生改变.对于一年生草本植物而言,其生物量动态与年降水量变化趋势相反,与多年生草本植物的生物量呈现互补趋势,这是由于降水量减少影响了多年生植物以及半灌木的生长,从而导致其失去竞争力,促进一年生草本植物生长提高地上生物量;一年生草本植物对干旱环境具有一定的适应能力,轻度干旱胁迫对某些一年生草本植物生长影响较小,甚至有一定的促进作用.李慧等[36 ] 指出,组合群落会使植物的生物量和互补效应显著增加,在不同植物组合中,一年生和多年生草本植物的生物量存在互补性,二者组合可以增加总体生物量.美国明尼苏达州长期草地生物多样性试验也印证了我们的观点[37 ] ,即半干旱沙质草地多年生草本植物与一年生草本植物形成互补,共同提高植物群落生物量.这种生物量的互补性可能与植物的生态位分化和资源利用策略有关,未来研究可以进一步探讨其背后的生态机制. ...
不同模拟增雨下白刺比叶面积和叶干物质含量的比较
0
2015
两种一年生草本对喀斯特土壤厚度及供水的生长响应
0
2021
多年生黑麦草对干旱胁迫下喀斯特异质生境的生长响应策略
0
2020
Plant functional traits and environmental filters at a regional scale
1
1998
... 生物量是衡量沙质草地植物生长发育状况的重要指标,直接反映植物的生长和生产力[30 ] .本研究结果表明,降水量减少使植物群落地上生物量显著降低,原因可能是为应对干旱带来的危害,植物调节植株高度、叶面积和叶片干物质含量,导致植物地上部分积累的干物质减少,从而降低了植物地上生物量[31 -35 ] .不同植物或功能群对水分利用策略的差异,导致其竞争能力和优势等级的不同,最终造成植物群落结构和组成比例发生改变.对于一年生草本植物而言,其生物量动态与年降水量变化趋势相反,与多年生草本植物的生物量呈现互补趋势,这是由于降水量减少影响了多年生植物以及半灌木的生长,从而导致其失去竞争力,促进一年生草本植物生长提高地上生物量;一年生草本植物对干旱环境具有一定的适应能力,轻度干旱胁迫对某些一年生草本植物生长影响较小,甚至有一定的促进作用.李慧等[36 ] 指出,组合群落会使植物的生物量和互补效应显著增加,在不同植物组合中,一年生和多年生草本植物的生物量存在互补性,二者组合可以增加总体生物量.美国明尼苏达州长期草地生物多样性试验也印证了我们的观点[37 ] ,即半干旱沙质草地多年生草本植物与一年生草本植物形成互补,共同提高植物群落生物量.这种生物量的互补性可能与植物的生态位分化和资源利用策略有关,未来研究可以进一步探讨其背后的生态机制. ...
根系生态位差异对生态系统的影响
1
2021
... 生物量是衡量沙质草地植物生长发育状况的重要指标,直接反映植物的生长和生产力[30 ] .本研究结果表明,降水量减少使植物群落地上生物量显著降低,原因可能是为应对干旱带来的危害,植物调节植株高度、叶面积和叶片干物质含量,导致植物地上部分积累的干物质减少,从而降低了植物地上生物量[31 -35 ] .不同植物或功能群对水分利用策略的差异,导致其竞争能力和优势等级的不同,最终造成植物群落结构和组成比例发生改变.对于一年生草本植物而言,其生物量动态与年降水量变化趋势相反,与多年生草本植物的生物量呈现互补趋势,这是由于降水量减少影响了多年生植物以及半灌木的生长,从而导致其失去竞争力,促进一年生草本植物生长提高地上生物量;一年生草本植物对干旱环境具有一定的适应能力,轻度干旱胁迫对某些一年生草本植物生长影响较小,甚至有一定的促进作用.李慧等[36 ] 指出,组合群落会使植物的生物量和互补效应显著增加,在不同植物组合中,一年生和多年生草本植物的生物量存在互补性,二者组合可以增加总体生物量.美国明尼苏达州长期草地生物多样性试验也印证了我们的观点[37 ] ,即半干旱沙质草地多年生草本植物与一年生草本植物形成互补,共同提高植物群落生物量.这种生物量的互补性可能与植物的生态位分化和资源利用策略有关,未来研究可以进一步探讨其背后的生态机制. ...
Strategy shifts in leaf physiology,structure and nutrient content between species of high‐and low‐rainfall and high‐and low‐nutrient habitats
1
2001
... 生物量是衡量沙质草地植物生长发育状况的重要指标,直接反映植物的生长和生产力[30 ] .本研究结果表明,降水量减少使植物群落地上生物量显著降低,原因可能是为应对干旱带来的危害,植物调节植株高度、叶面积和叶片干物质含量,导致植物地上部分积累的干物质减少,从而降低了植物地上生物量[31 -35 ] .不同植物或功能群对水分利用策略的差异,导致其竞争能力和优势等级的不同,最终造成植物群落结构和组成比例发生改变.对于一年生草本植物而言,其生物量动态与年降水量变化趋势相反,与多年生草本植物的生物量呈现互补趋势,这是由于降水量减少影响了多年生植物以及半灌木的生长,从而导致其失去竞争力,促进一年生草本植物生长提高地上生物量;一年生草本植物对干旱环境具有一定的适应能力,轻度干旱胁迫对某些一年生草本植物生长影响较小,甚至有一定的促进作用.李慧等[36 ] 指出,组合群落会使植物的生物量和互补效应显著增加,在不同植物组合中,一年生和多年生草本植物的生物量存在互补性,二者组合可以增加总体生物量.美国明尼苏达州长期草地生物多样性试验也印证了我们的观点[37 ] ,即半干旱沙质草地多年生草本植物与一年生草本植物形成互补,共同提高植物群落生物量.这种生物量的互补性可能与植物的生态位分化和资源利用策略有关,未来研究可以进一步探讨其背后的生态机制. ...
中国北方干旱半干旱区植被对气候变化和人类活动的响应
1
2024
... 干旱半干旱地区草地生态系统稳定性对于降水变化极为敏感,植被作为草地生态系统的主体对降水变化的响应最为显著,而植物群落中不同的功能群对降水动态变化的响应机制却不同[38 -42 ] .本研究结果表明,一年生草本和多年生草本的生物量分别与当年7月和生长季降水量显著相关.考虑到一年生草本地上生物量是每年8月收获测定,与当年7月降水量显著相关,主要原因是一年生草本植物的生长对短期降水变化非常敏感,前月降水量直接影响次月一年生草本植物的长势.而多年生草本植物的生物量与当年生长季降水量显著相关,是因为多年生草本植物在长期的进化过程中形成了适应干旱等胁迫环境的生理生态特征,对于降水变化敏感度低于一年生草本,其地上生物量动态变化不会被短期降水变化所影响.这与王军峰等[43 ] 在荒漠草原开展野外降水控制试验的结果一致,即一年生草本植物的生物量随短期降水变异波动明显,而多年生草本植物的生物量受整个生长季降水的影响. ...
全球植被叶面积指数对温度和降水的响应研究
0
2002
基于生态分区的内蒙古地区植被覆盖变化及其影响因素研究
0
2013
基于随机森林模型的内陆干旱区植被指数变化与驱动力分析:以北天山北坡中段为例
0
2020
半干旱黄土区不同管理措施下草地群落结构对短期氮、水添加的响应
1
2021
... 干旱半干旱地区草地生态系统稳定性对于降水变化极为敏感,植被作为草地生态系统的主体对降水变化的响应最为显著,而植物群落中不同的功能群对降水动态变化的响应机制却不同[38 -42 ] .本研究结果表明,一年生草本和多年生草本的生物量分别与当年7月和生长季降水量显著相关.考虑到一年生草本地上生物量是每年8月收获测定,与当年7月降水量显著相关,主要原因是一年生草本植物的生长对短期降水变化非常敏感,前月降水量直接影响次月一年生草本植物的长势.而多年生草本植物的生物量与当年生长季降水量显著相关,是因为多年生草本植物在长期的进化过程中形成了适应干旱等胁迫环境的生理生态特征,对于降水变化敏感度低于一年生草本,其地上生物量动态变化不会被短期降水变化所影响.这与王军峰等[43 ] 在荒漠草原开展野外降水控制试验的结果一致,即一年生草本植物的生物量随短期降水变异波动明显,而多年生草本植物的生物量受整个生长季降水的影响. ...
荒漠草原区不同生活型植物生长对降水变化的响应
1
2020
... 干旱半干旱地区草地生态系统稳定性对于降水变化极为敏感,植被作为草地生态系统的主体对降水变化的响应最为显著,而植物群落中不同的功能群对降水动态变化的响应机制却不同[38 -42 ] .本研究结果表明,一年生草本和多年生草本的生物量分别与当年7月和生长季降水量显著相关.考虑到一年生草本地上生物量是每年8月收获测定,与当年7月降水量显著相关,主要原因是一年生草本植物的生长对短期降水变化非常敏感,前月降水量直接影响次月一年生草本植物的长势.而多年生草本植物的生物量与当年生长季降水量显著相关,是因为多年生草本植物在长期的进化过程中形成了适应干旱等胁迫环境的生理生态特征,对于降水变化敏感度低于一年生草本,其地上生物量动态变化不会被短期降水变化所影响.这与王军峰等[43 ] 在荒漠草原开展野外降水控制试验的结果一致,即一年生草本植物的生物量随短期降水变异波动明显,而多年生草本植物的生物量受整个生长季降水的影响. ...
Species diversity is a strong predictor of ecosystem multifunctionality under altered precipitation in desert steppes
1
2022
... 降水变化对植物群落构建的影响可以从物种多样性、功能群组成和生态位动态等多个维度进行剖析.首先,降水变化可能通过影响物种的繁殖、生长和存活,进而改变群落的物种组成和多样性.例如,降水的增加可能促进某些物种的生长,但同时也会抑制其他物种的竞争力,导致群落物种多样性降低[44 ] .降水变化对不同功能群的影响也存在差异.一年生草本植物通常对短期降水变化更为敏感,而多年生草本植物则表现出更强的适应性[45 ] .这种差异可能导致功能群之间的竞争格局发生变化,进而影响群落的结构和功能[9 ] .此外,降水变化还可能通过改变植物的生态位宽度和重叠程度,影响物种之间的相互作用和群落的稳定性[23 ] .例如,降水量的减少可能使植物的生态位更加狭窄,导致物种之间的竞争加剧,而降水量的增加则可能扩大生态位宽度,促进物种共存[45 ] .不同功能群对水分利用的策略存在差异,导致各功能群植物地上生物量在群落中的比例发生改变.这种生物量分配策略的变化反映了植物群落对降水变化的适应机制.本研究探讨了不同功能群在不同降水条件下的分布和竞争关系.在40%的年降水变异条件下,禾本科植物糙隐子草和豆科植物兴安胡枝子等表现出较强的竞争优势并占据更优生态位,而随着降水变异增加到50%,菊科植物猪毛蒿和禾本科植物白草逐渐成为优势种.这表明降水变化显著影响了功能群之间的竞争格局,进而改变了群落的物种组成和优势度分布,这些变化对群落稳定性和生态系统功能可能产生重要影响.功能群多样性的变化反映了群落对环境胁迫的响应机制,功能群的调整有助于维持生态系统生产力的稳定性.例如,功能群均匀度的离散程度在高降水变异条件下显著增加,这可能削弱了群落的整体稳定性.此外,功能群之间的竞争关系也对生态系统功能产生影响,禾本科植物的生物量与群落总生物量正相关,表明其在生态系统生产力中占据主导地位. ...
呼伦贝尔沙化草地不同放牧潜力阶段群落特征
2
2025
... 降水变化对植物群落构建的影响可以从物种多样性、功能群组成和生态位动态等多个维度进行剖析.首先,降水变化可能通过影响物种的繁殖、生长和存活,进而改变群落的物种组成和多样性.例如,降水的增加可能促进某些物种的生长,但同时也会抑制其他物种的竞争力,导致群落物种多样性降低[44 ] .降水变化对不同功能群的影响也存在差异.一年生草本植物通常对短期降水变化更为敏感,而多年生草本植物则表现出更强的适应性[45 ] .这种差异可能导致功能群之间的竞争格局发生变化,进而影响群落的结构和功能[9 ] .此外,降水变化还可能通过改变植物的生态位宽度和重叠程度,影响物种之间的相互作用和群落的稳定性[23 ] .例如,降水量的减少可能使植物的生态位更加狭窄,导致物种之间的竞争加剧,而降水量的增加则可能扩大生态位宽度,促进物种共存[45 ] .不同功能群对水分利用的策略存在差异,导致各功能群植物地上生物量在群落中的比例发生改变.这种生物量分配策略的变化反映了植物群落对降水变化的适应机制.本研究探讨了不同功能群在不同降水条件下的分布和竞争关系.在40%的年降水变异条件下,禾本科植物糙隐子草和豆科植物兴安胡枝子等表现出较强的竞争优势并占据更优生态位,而随着降水变异增加到50%,菊科植物猪毛蒿和禾本科植物白草逐渐成为优势种.这表明降水变化显著影响了功能群之间的竞争格局,进而改变了群落的物种组成和优势度分布,这些变化对群落稳定性和生态系统功能可能产生重要影响.功能群多样性的变化反映了群落对环境胁迫的响应机制,功能群的调整有助于维持生态系统生产力的稳定性.例如,功能群均匀度的离散程度在高降水变异条件下显著增加,这可能削弱了群落的整体稳定性.此外,功能群之间的竞争关系也对生态系统功能产生影响,禾本科植物的生物量与群落总生物量正相关,表明其在生态系统生产力中占据主导地位. ...
... [45 ].不同功能群对水分利用的策略存在差异,导致各功能群植物地上生物量在群落中的比例发生改变.这种生物量分配策略的变化反映了植物群落对降水变化的适应机制.本研究探讨了不同功能群在不同降水条件下的分布和竞争关系.在40%的年降水变异条件下,禾本科植物糙隐子草和豆科植物兴安胡枝子等表现出较强的竞争优势并占据更优生态位,而随着降水变异增加到50%,菊科植物猪毛蒿和禾本科植物白草逐渐成为优势种.这表明降水变化显著影响了功能群之间的竞争格局,进而改变了群落的物种组成和优势度分布,这些变化对群落稳定性和生态系统功能可能产生重要影响.功能群多样性的变化反映了群落对环境胁迫的响应机制,功能群的调整有助于维持生态系统生产力的稳定性.例如,功能群均匀度的离散程度在高降水变异条件下显著增加,这可能削弱了群落的整体稳定性.此外,功能群之间的竞争关系也对生态系统功能产生影响,禾本科植物的生物量与群落总生物量正相关,表明其在生态系统生产力中占据主导地位. ...




 甘公网安备 62010202000688号
甘公网安备 62010202000688号
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}