科尔沁沙质草地关键植物种群动态对降水变化的响应
刘新平 , 1 , 2 , 胡鸿姣 1 , 2 , 4 , 何玉惠 1 , 3 , 徐远志 1 , 2 , 4 , 景家琪 1 , 2 , 4 , 张尧 1 , 2 , 4
1.中国科学院西北生态环境资源研究院,干旱区生态安全与可持续发展全国重点实验室,甘肃 兰州 730000
2.中国科学院西北生态环境资源研究院,奈曼沙漠化研究站,甘肃 兰州 730000
3.中国科学院西北生态环境资源研究院,兰州生态农业试验研究站,甘肃 兰州 730000
4.中国科学院大学,北京 100049
Response of key plant populations dynamics to precipitation changes in Horqin Sandy Grassland
Liu Xinping , 1 , 2 , Hu Hongjiao 1 , 2 , 4 , He Yuhui 1 , 3 , Xu Yuanzhi 1 , 2 , 4 , Jing Jiaqi 1 , 2 , 4 , Zhang Yao 1 , 2 , 4
1.State Key Laboratory of Ecological Safety and Sustainable Development in Arid Lands, Northwest Institute of Eco-Environment and Resources,Chinese Academy of Sciences,Lanzhou 730000,China
2.Naiman Desertification Research Station, Northwest Institute of Eco-Environment and Resources,Chinese Academy of Sciences,Lanzhou 730000,China
3.Lanzhou Ecological Agriculture Experimental Research Station, Northwest Institute of Eco-Environment and Resources,Chinese Academy of Sciences,Lanzhou 730000,China
4.University of Chinese Academy of Sciences,Beijing 100049,China
收稿日期: 2025-04-25
修回日期: 2025-06-03
基金资助:
内蒙古自治区防沙治沙科技创新重大示范工程“揭榜挂帅”项目 . 2024JBGS0005-1 中国科学院区域发展青年学者项目 干旱区生态安全与可持续发展全国重点实验室交叉创新团队项目 . E451890201
Received: 2025-04-25
Revised: 2025-06-03
作者简介 About authors
刘新平(1978—),男,甘肃瓜州人,研究员,博士生导师,主要从事生态水文学与恢复生态学研究E-mail:liuxinping@lzb.ac.cn
, E-mail:liuxinping@lzb.ac.cn
摘要
干旱半干旱区陆地生态系统中,关键植物种群的动态变化是植被结构调整的核心驱动力,也是植被响应降水变化的生态基础。本研究以中国北方半干旱区科尔沁沙质草地植被为对象,通过降水变化控制实验,解析关键种群对降水变化的响应路径及其生态功能分异特征,进而识别植被结构的种群水平配置规律。结果表明:降水减少驱动多年生优势植物种被一年生植物替代,如糙隐子草(Cleistogenes squarrosa )的重要值和生态位宽度可由0.21和5.06降低至0,而降水增加仅微调各优势种的优势度。降水变化下,植被指示种功能分异明显:降水增加以多年生植物为主,如糙隐子草指示值可达0.25;降水减少以半灌木为主,如兴安胡枝子(Lespedeza bicolor )指示值可达0.23;阶段性干旱则为猪毛菜(Salsola collina )等一年生植物,指示值为0.39。植被泛化种对阶段性干旱较为敏感,植被特化种对极端降水变化较为敏感,二者生态位收缩均触发中性类群物种数占比增加以填补空缺。具有高重要值、高指示性、广生态位或特殊生态位的关键种均可对降水处理组间差异具有重要贡献。这些关键种群结构特征对降水的极端变化、减少及春季变化会更敏感。本研究强调结合降水格局与对应关键缓冲植物种来优化植被空间配置的可行性,可为半干旱沙质草地植被响应气候变化的适应性管理提供理论支撑。
关键词:
降水格局 植物种群 优势种 指示种 生态位
Abstract
In arid and semi-arid terrestrial ecosystems, the dynamics of key plant populations constitute a central driver of vegetation structural reorganization, and serve as the ecological foundation for vegetation responses to precipitation changes. This study focuses on the Horqin Sandy Grassland vegetation of northern China's semi-arid area. Through controlled experiments manipulating precipitation regimes, we investigated the response pathways of diverse key populations to precipitation changes and their characteristics of ecological functional differentiation, thereby identifying the population-level configuration patterns of vegetation structure. Results demonstrated that: precipitation reduction drove the replacement of perennial dominant species (e.g., Cleistogenes squarrosa , with its importance value and ecological niche breadth declining from 0.21 and 5.06 to near zero, respectively) by annual plants, while precipitation increase only moderately adjusted the dominance hierarchy among species. Indicator species exhibited distinct functional differentiation under precipitation changes: perennials (e.g., Cleistogenes squarrosa with an indicator value of 0.25) dominated under increased precipitation, semi-shrubs (e.g., Lespedeza davurica with an indicator value of 0.23) prevailed under decreased precipitation, and annuals (e.g., Salsola collina with an indicator value of 0.39) proliferated under seasonal droughts. Generalist species showed heightened sensitivity to seasonal droughts, whereas specialists responded markedly to extreme precipitation fluctuations. Niche contraction in both groups triggered an species percentage increase in neutral taxa to fill ecological vacancies. Key populations characterized by high-importance values, strong indicator properties, broad ecological niches, or specialized niches can significantly contributed to inter-treatment differences. Notably, these key populations showed higher sensitivity to extreme precipitation changes, negative precipitation anomalies, and spring-season precipitation changes. This work underscores the feasibility of optimizing vegetation configurations by integrating precipitation regimes with their corresponding buffering species, providing theoretical foundations for adaptive management of sandy grassland ecosystems in semi-arid regions.
Keywords:
precipitation regime plant population dominant species indicator species ecological niche
本文引用格式
刘新平, 胡鸿姣, 何玉惠, 徐远志, 景家琪, 张尧. 科尔沁沙质草地关键植物种群动态对降水变化的响应 . 中国沙漠 [J], 2025, 45(4): 43-56 doi:10.7522/j.issn.1000-694X.2025.00073
Liu Xinping, Hu Hongjiao, He Yuhui, Xu Yuanzhi, Jing Jiaqi, Zhang Yao. Response of key plant populations dynamics to precipitation changes in Horqin Sandy Grassland . Journal of Desert Research
0 引言
全球气候变化驱动陆地生态系统水文格局重构,核心表现为降水时空异质性加剧,极端干旱与强降水事件频率、强度和季节分配呈现同步分异趋势;中高纬度干旱半干旱区域降水变率尤为显著,呈现出干的地方越干、湿的地方越湿的格局[1 ] 。降水作为陆地生态系统中最重要的水分输入形式,其格局变化可改变水分可利用性阈值,显著调控植被群落的结构与功能[2 ] 。尤其是在降水限制型生态系统中,植物种群对水分波动的敏感性构成生态系统稳定性与恢复力的核心调控因子[3 ] 。半干旱沙质草地作为典型的脆弱生态系统,基质特性贫瘠,生态阈值狭窄,其关键植物种群功能性状调整与关键种替代机制已成为维系系统弹性的最后防线[4 ] :短期水分波动可能触发种群竞争优势的重新分配,而长期降水变化则可能驱动群落演替方向的根本性转变[5 ] 。在此背景下,解析半干旱沙质草地关键植物种群对降水变化的响应策略,不仅是预测生态系统碳-水耦合过程的理论基石,也是制定适应性管理策略的科学前提。
重要值、指示值和生态位宽度作为表征植物种群响应环境变化的3个核心指标,分别从物种竞争优势、环境适应特征和资源利用策略3个维度揭示降水驱动的生态过程,其内在关联与作用差异是解析植被结构重组机制的关键科学问题。降水变化可通过改变植物相对盖度、高度和多度,显著影响物种重要值排序和关键种群组成[6 ] 。例如在温带草原中,多年生禾草的重要值通常随降水减少而下降,一年生草本则通过快速生活史策略占据生态位空缺[7 ] 。指示值方法揭示植物对特定环境条件的指示性,为识别关键指示种提供了方法:C4 植物在干旱处理中指示性显著提升,而C3 植物则更适应降水增加环境[8 ] 。生态位宽度作为物种资源利用能力的表征,其动态变化反映种间竞争与环境筛选的协同作用。其中,泛化种通常具有较宽的生态位,能够在多种环境条件下生存,而特化种则对特定环境条件具有高度依赖性,并且极端干旱可导致泛化种生态位收缩,而适度降水增加则可促进特化种的功能分化[9 ] 。尽管已有大量研究关注降水变化对植被的影响,但仍存在3个关键不足:首先,超一半的野外控制实验仅设置常态化降水梯度,难以捕捉极端气候事件的生态效应[10 ] 。其次,现有研究多关注降水量的年际变化,却忽视季节性干旱对植物生活史策略的差异化影响。例如,生长季初期干旱可能通过延迟物候期影响一年生植物的萌发时间,而生长季盛期则直接抑制光合产物的积累[11 ] 。另外,当前研究多侧重于群落水平的单一笼统结构性特征指标(如物种多样性)分析,往往割裂物种重要值、指示值及生态位宽度这3个指标的内在联系,导致关键种替代机制的解析存在偏差[12 ] 。重要值揭示优势种动态,指示值筛选环境敏感种,生态位宽度量化种间竞争与适应策略。三者结合可避免单一指标的局限性,例如,高重要值物种可能缺乏环境指示性,而特化种虽指示性强但贡献度低[12 ] 。
科尔沁沙地位于中国北方农牧交错带典型生态脆弱区,其独特的沙质基质与高降水变率为解析上述问题提供了模式生态系统[11 ,13 ] 。本研究基于科尔沁沙质草地植被,通过7年的降水模拟实验(涉及降水减少和增加、适度和极端、阶段性和连续性的降水变化格局),引入物种重要值、指示值及生态位宽度等多维种群特征指标,分析不同降水变化格局对植被种群结构特征的影响,旨在揭示以下科学问题:①降水变化如何通过优势种响应或更替驱动植被结构调整;②不同降水格局下指示种有何特征差异;③特化种与泛化种的生态位动态如何响应降水变化;④哪些关键种对植被结构响应降水格局转变具有显著贡献,通过以上研究以期为旱区脆弱生态系统植被恢复和管理保护提供理论依据和实践指导。
1 材料与方法
1.1 研究区概况
本研究在中国北部内蒙古东部科尔沁沙地(42°20′—44°50′N、117°15′—123°43′E,海拔约377 m)中南部的中国科学院奈曼沙漠化研究站开展。该区域被划定为半干旱农牧交错区,属温带大陆性半干旱气候。年平均气温约6.4 ℃,生长季4—9月日平均气温大多高于10 ℃。多年平均降水量约351.7 mm,其中生长季降水量占全年的67.5%~85.3%,年均潜在蒸发量1 935 mm。年降水量波动为-30%~30%,极端情况下可达60%(参照 1981—2023年科尔沁沙地气象站降水记录);春夏季降水变率较大,最长连续28天无降水[14 ] 。地貌景观以各类沙丘为特征,土壤由沙(98.2%)、粉砂和黏土(1.8%)构成,物理性质均一,质地粗匀,结构疏松。植被主要由一年生草本植物组成,如大果虫实(Corispermum macrocarpum )、狗尾草(Setaria viridis )和针茅(Stipa capillata )。随着植被恢复进程的推进,多年生植物和禾本科植物的种类及数量所增加,例如猪毛蒿(Artemisia scoparia )、糙隐子草(Cleistogenes squarrosa )和砂引草(Tournefortia sibirica )[15 ] 。
1.2 研究方法
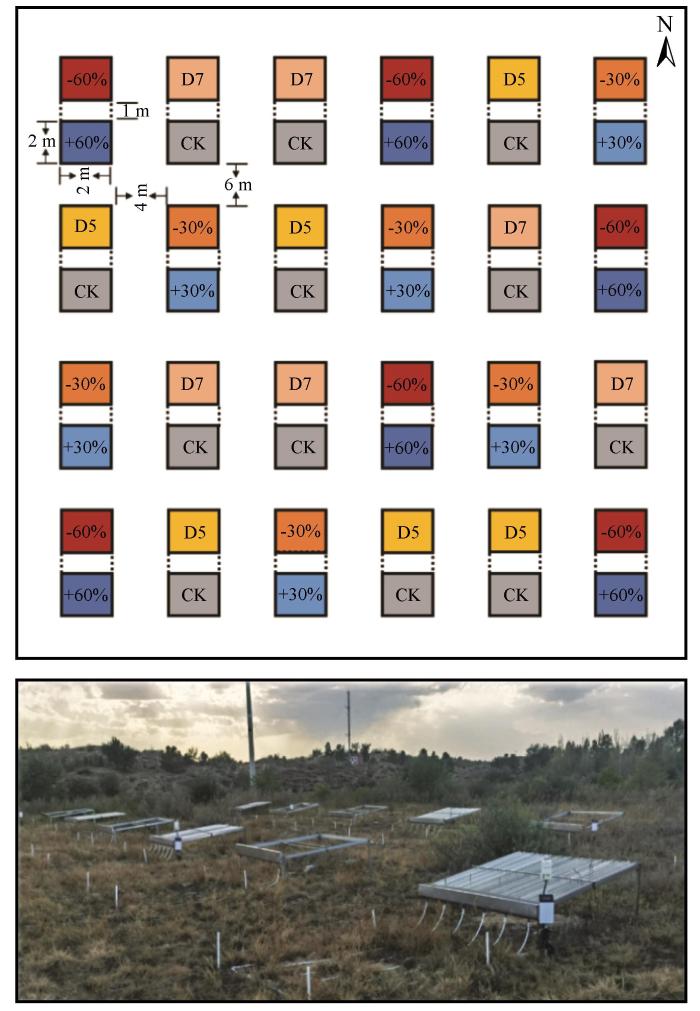
以中国科学院奈曼沙漠化研究站内长期封育的典型沙质草地为研究样地(0.23 km2 ),于2016年建立沙质草地降水增减控制试验场(图1 )。依据研究区降水背景及植被对各季节降水变化的敏感性差异,共设置7个降水处理,分别为降水增加60%(+60%P)和降水减少60%(-60%P),模拟极端湿润和极端干旱事件;5月干旱(D5)和7月干旱(D7),模拟春季和夏季阶段性干旱事件;降水增加30%(+30%P)和降水减少30%(-30%P),模拟适度连续性降水变化事件;以及保持自然降水条件作为基准对照组(CK)。除对照处理设置12个重复外,其余增减降水处理各设置6个重复,共48个处理小区,试验设计为完全随机区组设计。野外降水增减控制试验装置由奈曼站自主研发[16 ] ,对每年自然降水量进行梯度控制,其中降水减少区和降水增加区大小均为2 m×2 m,两者间缓冲区为2 m×2 m。
图1
图1
野外增减雨控制试验场布局及示意图
Fig.1
Field layout of precipitation experiment and schematic diagram of precipitation manipulation device
本研究基于2023年植被生物量高峰期(8月中旬)开展实地植被调查,解析降水变化处理7年后的影响。针对各降水增加/降水减少处理小区,采用对角布设法规范设置2个0.5 m×0.5 m植被调查样方;调查内容包括植物的种类、总株数、分物种株数、总盖度(%)、分物种盖度(%)、分物种自然高度(cm)等。
1.3 统计分析
物种重要值是研究某个物种在群落中的重要性的综合数量指标。基于样方调查数据,采用下列公式计算表示群落中某物种的相对重要值(Relative importance value,RIV)[17 ] 。基于不同降水条件下植物群落内各物种的相对重要值,基于欧氏距离(Euclidean distance)进行分层聚类分析,筛选出重要值排名明显靠前的物种作为各降水处理下的优势种。
R I V = ( C r + H r + D r ) / 3
式中:C r 为相对盖度(%),为某物种盖度除以群落总盖度;H r 为相对高度(%),为某物种所有个体高度之和除以群落所有个体高度之和;D r 为相对多度(%),为某物种总个体数除以群落总个体数。
基于不同降水处理下各植物种群重要值,使用labdsv::indval函数计算各种群指示值(IndVal指数)并筛选指示种[18 ] ;使用vegan::simper函数筛选组间差异贡献显著的物种[19 ] ;使用spaa::niche.width函数[20 ] 和EcolUtils::spec.gen函数定义泛化种、特化种和中性类群[20 ] 。
通过以上数据分析筛选出优势种、指示种、泛化种、特化种及组间差异贡献种均界定为植被响应降水变化的关键种群。本研究所有统计分析均基于R软件(版本4.3.1;R Core Team, 2023)完成,显著性水平设为P <0.05。
2 结果与分析
2.1 沙质草地植物重要值特征
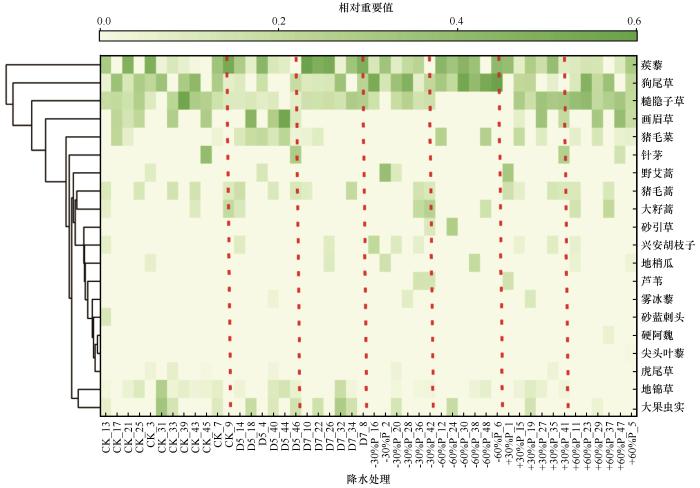
自然降水条件下,植物群落优势种群组成及相应重要值依次为糙隐子草(0.21,多年生)、狗尾草(0.21,一年生)和蒺藜(0.20,一年生;表1 ,图2 )。4种降水减少处理下,蒺藜的重要值排名均提前,甚至在7月干旱处理下成为重要值最大的物种,而糙隐子草重要值排名则均后移,甚至在5月干旱和降水减少60%处理下退出优势种群行列。其中,在降水减少60%处理下,狗尾草和蒺藜为优势种群,两者重要值均明显较高。5月干旱处理下,优势种群中各植物种的重要值排序变动最大,表现为与自然降水条件下的排序完全相反,优势种包括画眉草、蒺藜和猪毛菜;在7月干旱处理下,蒺藜成为最优势种,重要值提升至0.42;在降水减少30%处理下,狗尾草成为最优势种,重要值提升至0.29。降水增加30%处理下,多年生植物中重要值最高的为糙隐子草,一年生植物中重要值最高的为蒺藜,其次为画眉草;降水增加60%处理下,多年生糙隐子草依然为最优势种,其次为一年生狗尾草。总之,降水变化可通过改变优势种群的种类和/或重要值来影响植被种群结构特征,其中生长季初期干旱(5月干旱)和极端干旱(降水减少60%)的影响较突出,主要表现为多年生草本植物优势度降低,一年生草本植物优势度提高;而降水增加的影响较小,无论是适度增加还是极端增加(表1 ,图2 )。
图2
图2
不同降水条件下各植物种重要值的层次聚类
注: CK为自然降水条件;D5和D7分别代表5月干旱和7月干旱;+30%P、-30%P、+60%P和-60%P分别代表全年降水增加30%、减少30%、增加60%和减少60%
Fig.2
Hierarchical clustering of plant species importance values across different precipitation regimes
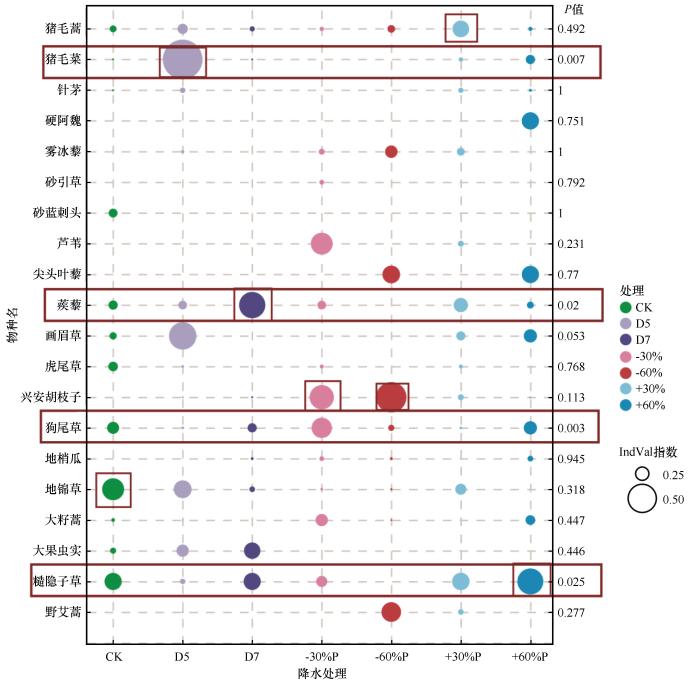
2.2 植物指示值特征
不同增减降水处理下群落中植物种的指示值与相对重要值并无绝对一致性;不同降水变化处理下植物群落具有不同的指示种,同一处理下各植物种的指示性具有明显差异;不同处理下各物种的IndVal指示值均在0.5以下(图3 )。共筛选出4种显著指示种(P <0.05),包括糙隐子草(多年生禾草)、狗尾草(一年生禾草)、蒺藜(一年生杂类草)、猪毛菜(一年生杂类草)。各降水处理指示值均值范围为0.069(CK)~0.099(-30%P),其中5月干旱和降水减少60%处理的指示值离散度较高(标准差分别为0.105和0.089)。降水增加60%处理下,指示种糙隐子草(IndVal=0.254)和硬阿魏(IndVal=0.167)的指示值升高;而降水减少60%处理下指示种兴安胡枝子(IndVal=0.295)指示值明显提升。指示种狗尾草在降水减少30%处理下指示值达0.198,明显高于对照组;降水增加30%处理下指示种为猪毛蒿(IndVal=0.168)。5月干旱处理下,指示种猪毛菜(IndVal=0.386, P =0.007)及画眉草(IndVal=0.267)占据主导,而7月干旱处理下指示种蒺藜(IndVal=0.256)指示值明显提升。降水增加组群落平均指示值为0.085,低于降水减少组的0.098和阶段性干旱组的0.096。此外,降水增加组的指示种均为多年生植物,降水减少组的指示种均为半灌木,阶段性干旱组的指示种均为一年生植物。
图3
图3
不同降水条件下各植物种指示值
注: CK为自然降水条件;D5和D7分别代表5月干旱和7月干旱;+30%P、-30%P、+60%P和-60%P分别代表全年降水增加30%、减少30%、增加60%和减少60%
Fig.3
Indicator values (IndVals) of plant species under different precipitation regimes
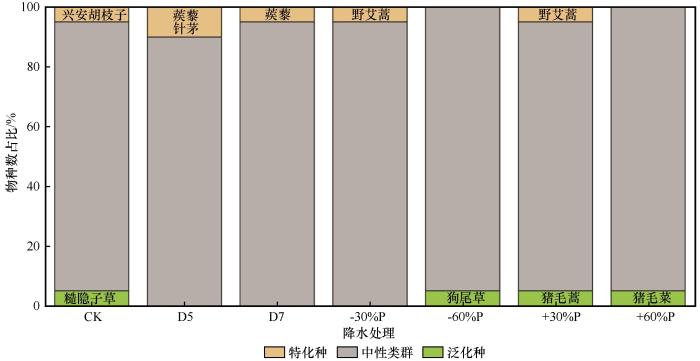
2.3 植物生态位特征
特化种和泛化种在群落中的物种数占比较小,两者种类和物种数占比均受降水变化的影响,并不总是同时存在于植物群落中(图4 ,表2 )。2种阶段性干旱和降水减少30%处理削弱了泛化种在群落结构中的地位,从而降低中性类群的资源竞争力;二种极端降水变化处理削弱了特化种在群落结构中的地位,从而提高中性类群的资源竞争力;降水增加30%处理虽然保持群落中泛化种和特化种的组成地位,并且仍保持均为多年生植物,但是改变了二者的组成物种。自然降水条件下,群落的平均生态位宽度为3.24,特化种为兴安胡枝子,其生态位宽度为2.51,泛化种为糙隐子草,其生态位宽度为5.06。5月干旱、7月干旱和降水减少30%处理下,群落的平均生态位宽度分别为2.15、2.49和1.97,特化种分别为蒺藜+针茅、蒺藜、野艾蒿,生态位宽度分别为3.53+2.5、4.86、2.66。降水减少60%和降水增加60%处理下,群落的平均生态位宽度分别为1.95和2.10,泛化种分别为狗尾草、猪毛菜(均为一年生植物),生态位宽度分别为3.32和1.78。降水增加30%处理下,群落的平均生态位宽度为1.93,特化种和泛化种分别为野艾蒿(2.57)和猪毛蒿(1.54)。
图4
图4
不同降水条件下植被的泛化种和特化种占比
注: CK为自然降水条件;D5和D7分别代表5月干旱和7月干旱;+30%P、-30%P、+60%P和-60%P分别代表全年降水增加30%、减少30%、增加60%和减少60%
Fig.4
Proportion of generalist species and specialist species under different precipitation regimes
2.4 各植物种对降水变化组间差异的贡献特征
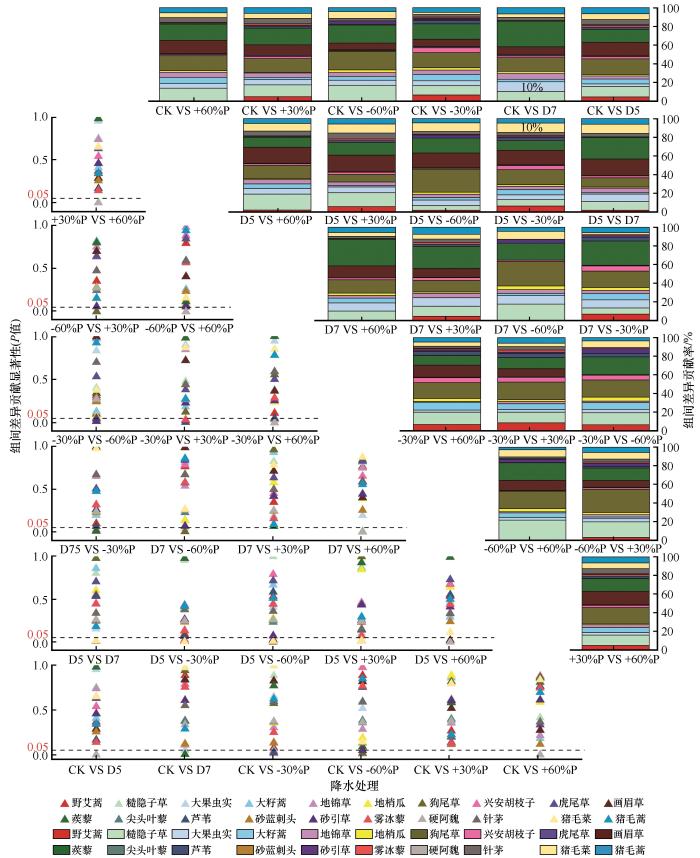
高重要值物种通常具有较高的降水处理组间差异贡献度,但不一定贡献显著;而对组间差异贡献度显著的物种通常具有特殊生态位或具有较高指示性。狗尾草、蒺藜、糙隐子草、画眉草对不同降水增减处理群落组成结构差异的贡献度普遍较高,但贡献度均普遍在20%以下(图5 ,表3 )。如表3 所列,狗尾草在多个处理中均表现出显著的组间差异贡献度(贡献度范围:12.67%~17.82%;P <0.05);特别是在极端降水处理(+60%P和-60%P )下,其贡献度显著升高(P <0.05)。蒺藜同样在多个处理下均表现出显著贡献度(贡献度范围:14.50%~16%;P <0.05),尤其是在极端降水和阶段性干旱处理下(表3 )。糙隐子草在极端降水处理(+60%P和-60%P)下贡献度较高(贡献度范围:9.74%~14.68%,P <0.05),而在适度降水变化处理下贡献度较低。画眉草在D5处理下贡献显著(贡献度范围:10.9%~11.46%;P <0.05),在适度降水变化处理(+30%P和-30%P)下的贡献度中等(P >0.05)。如图5 所示,对照组与降水减少30%处理组间的差异主要由狗尾草(贡献度9.74%,P =0.575)、蒺藜(贡献度9.78%,P =0.772)贡献。对照与降水减少60%处理组间的差异主要由狗尾草(P =0.018)、蒺藜(P =0.089)和糙隐子草(P =0.006)贡献。其他处理组间的主要贡献种如下:降水增加30%与对照组间的差异主要由狗尾草(P =0.817)和蒺藜(P =0.582)贡献;降水增加60%与对照组间的差异主要由狗尾草(P =0.811)和蒺藜(P =0.858)贡献;春季干旱与对照组间的差异主要由狗尾草和蒺藜贡献;夏季干旱与对照组间的差异主要由狗尾草和蒺藜贡献。
图5
图5
各植物种对不同降水处理组差异的贡献度占比(右上角)及显著性(左下角)
注: CK为自然降水条件;D5和D7分别代表5月干旱和7月干旱;+30%P、-30%P、+60%P和-60%P分别代表全年降水增加30%、减少30%、增加60%和减少60%
Fig.5
Contribution percentage of plant species to inter-treatment dissimilarities (upper right) and corresponding significance levels
3 讨论
3.1 降水变化对优势种重要值动态的影响
动优势种群组成与重要值排序的方向性调整,从而对植物群落结构产生深刻影响。本研究中,自然降水条件下,多年生禾草糙隐子草与一年生禾草狗尾草共同占据优势地位,构成科尔沁沙质草地植物群落核心架构。然而,当降水减少时,糙隐子草重要值骤降,其生态位甚至可被一年生植物,如猪毛菜、大果虫实等取代。糙隐子草作为泛化种,依赖广泛的资源获取能力维持优势,其生态位宽度在稳定环境中(CK)显著高于一年生植物猪毛菜,但在极端干旱下(-60%P),其资源适应能力受限,生态位宽度骤降[21 ] 。相比之下,猪毛菜等一年生植物的生态位宽度也有下降,但其快速生长周期和种子繁殖策略更
适应极端环境下的资源波动,而非依赖广谱资源利用能力,因而其能在水分脉冲中占据短暂资源窗口。这表明生态位宽度并非唯一适应指标,生活史策略的权衡同样关键,体现出更强的环境筛选适应性[22 ] 。物种更替具有显著的物候期依赖性。例如7月干旱仅使糙隐子草重要值下降,且未触发关键种替代,而5月干旱却促使糙隐子草退出关键种行列。这种差异可能源于植物生活史策略的阶段性约束:5月正值糙隐子草分蘖高峰期,此时水分亏缺直接抑制其繁殖能力[23 ] ,而7月干旱主要影响其生殖分配,对种群建立的冲击较小[24 ] 。此外,物种更替同样验证了生态弹性阈值理论[25 ] ,当水分胁迫超过临界值,系统将进入以一年生植物为主导的演替新稳态;本研究中为极端干旱(降水减少60%)导致多年生禾草优势的不可逆丧失,提升了一年生植物的关键地位。与温带草原的研究不同,本研究发现适度和极端降水增加(降水增加30%和60%)均并未显著改变关键种组成,仅引起重要值排序微调(如画眉草重要值提升、狗尾草重要值下降)。这可能与沙质土壤的高渗透性有关:短期内过量降水难以被土壤持留,导致水分利用效率未显著提升[26 ] 。这一现象与降水增益促进多样性的经典认知不符,揭示在沙质生境中,降水增加的生态效应存在基质依赖性[27 ] 。在降水增加处理下,糙隐子草重要值仍保持最高,表明多年生植物在湿润条件下具有更强的竞争力。这可能与多年生植物的深根系和较高的水分利用效率有关,使其能够在长期湿润条件下更有效地利用土壤水分[28 ] 。
3.2 降水变化对指示种功能分异的影响
指示种筛选体现环境胁迫对植物功能性状的定向选择。IndVal值与相对重要值的非一致性表明,指示种选择不仅依赖丰度,还与其对特定生境的生态专一性相关。例如,兴安胡枝子在降水减少30%处理下虽非优势种,但其对贫瘠土壤的适应能力使其成为关键指示物种。自然降水条件下指示种地锦草在干旱处理中被猪毛菜取代,而降水增加组则出现糙隐子草和猪毛蒿作为指示种。这一结果表明干旱强化了耐旱性状(如C4 光合途径、肉质叶片)的选择,而降水增加则筛选出具有高水分利用率的物种[29 ] 。指示种生活型与降水处理呈现显著关联:降水增加组由多年生禾草主导,降水减少组以半灌木为主,而阶段性干旱组则由一年生杂类草占据优势。这种格局揭示了降水梯度对植物功能性状的筛选作用。例如,降水增加60%处理下糙隐子草显著指示性印证了其作为保守型策略者在湿润环境中的竞争优势[30 ] ;而降水减少组兴安胡枝子的指示性则凸显了半灌木在干旱胁迫下通过木质化结构降低蒸腾损耗的生存策略[31 ] ;一年生植物采用快速生长-繁殖的r策略应对短期资源波动[32 ] 。另外,极端处理组(降水减少60%和降水增加60%)较低的指示值均值与高离散度并存,表明半干旱系统在极端降水下的脆弱性。当水分阈值超过植物适应能力边界时,原本稳定的种间互作网络可能瓦解,导致指示种特异性下降[33 ] 。
3.3 降水变化对生态位结构的影响
生态位宽度变化揭示物种资源利用策略的环境敏感性差异。泛化种和特化种在群落中比例变化也反映了群落对降水变化的适应能力。自然降水条件下,糙隐子草(泛化种)与兴安胡枝子(特化种)形成互补的生态位格局,而降水减少60%处理使泛化种(猪毛菜)的生态位宽度下降,同时特化种(野艾蒿)消失。这表明泛化种在极端环境条件下具有更强的适应能力,能够通过调整资源利用策略维持群落的稳定性。这一现象可能由两种机制驱动,一是资源竞争失衡:干旱导致土壤水分与养分空间异质性增强,泛化种因无法在多维资源梯度中保持竞争力而衰退[34 ] ;二是环境筛选强化:极端干旱形成强烈的非生物过滤,淘汰了依赖稳定水分供给的特化种,使适应脉冲式资源供应的中性类群(如一年生草本)占据优势[35 ] 。降水增加30%处理虽未改变泛化种/特化种的组成地位,但导致其物种替换(野艾蒿→猪毛蒿)。这表明适度水分增加可能通过改变种间互作(如互利共生或竞争排除)而非直接环境筛选驱动生态位重组[36 ] 。此外,中性类群在特化种/泛化种受抑时竞争优势提升,表明非优势种通过填补生态位空缺维持系统功能的“保险效应”,当环境压力超过物种适应阈值时,具有广泛耐受性的中性物种可通过生态位漂移占据空缺资源[37 ] 。
3.4 植被结构变化的高贡献物种的多维解析
综合多维指标可见,优势种更替是植被结构响应降水变化的直观表现,指示种功能分异反映环境筛选的方向性,而生态位动态则揭示种间资源竞争本质。例如,降水减少60%处理下,糙隐子草(优势种)重要值骤降与其生态位宽度缩窄同步发生,而猪毛菜(指示种)通过快速占据生态位空缺成为新优势种,三者共同印证了资源适应能力下降→生态位释放→先锋种入侵的替代路径。多维指标协同分析,能更全面地揭示降水变化驱动植被结构调整的生态机制。广生态位物种或高重要值物种(如狗尾草、蒺藜、糙隐子草和画眉草)对降水处理组间差异的高贡献度特征(普遍P >0.05)提示其通过非显著性广谱参与在群落缓冲降水波动中发挥核心调控功能;而特殊生态位物种(如兴安胡枝子)的局部显著性贡献则表明狭生态位特化种可通过关键节点突破驱动差异格局。这表明旱区植被恢复应优先保育具有跨梯度适应力的缓冲物种,而非单纯依赖优势种[38 ] 。首先,狗尾草作为一年生禾草,在极端降水处理中贡献度显著提升,其广域贡献度可能与其快速资源捕获策略相关[39 ] 。尽管狗尾草在多个处理组间差异贡献度较高,但其显著性波动范围较大,这可能反映其作为“机会主义种”在不同处理中生态位宽度的动态调整[39 ] 。与之类似,蒺藜作为一年生杂类草,可能通过抑制竞争者水分获取和增强干旱期水分存储能力实现生态位占领[40 ] 。其次,糙隐子草作为多年生禾草,在极端降水处理中贡献度显著,但在适度降水变化中贡献度下降。这种双峰响应模式可能与其深层根系和休眠芽库策略相关:在极端湿润条件下,其分蘖能力增强[41 ] ;在极端干旱时,则通过地上部分生物量再分配维持生存[41 -42 ] 。相比之下,画眉草多在D5处理中贡献度显著,表明其早春萌发特性可能使其在季节性干旱前期占据水分利用窗口期[10 ] 。研究还发现,极端处理组间差异主要由多物种协同贡献(如极端干旱处理中狗尾草、蒺藜和糙隐子草联合贡献),而适度降水变化处理则依赖单一物种主导(如D5处理中画眉草贡献度较高),这表明降水扰动强度与群落响应模式间存在非线性关系:当降水偏离历史波动阈值(如±30%P)时,物种互作网络可能从补偿性响应转向竞争性替代[43 ] 。
4 结论
科尔沁沙质草地植被可通过关键种群的动态更替与生态位重构响应降水变化,其响应程度具有增减雨非对称性、季节差异性以及功能类群依赖性。降水增加可通过调整重要值排序和提升指示性强化多年生植物如糙隐子草的关键地位以维持植被结构,而降水减少则可促使一年生先锋种如蒺藜的优势地位提高。特化种的生态位宽度易受阶段性干旱和适度降水减少的影响而缩小,泛化种的则是易受极端降水变化的影响而缩小;两者的生态位调整均会促使中性类群生态位的补偿性扩张。广生态位物种和优势种如狗尾草、蒺藜对降水变化组间差异的非显著高贡献度,以及特殊生态位物种和指示种如兴安胡枝子对降水处理组间差异的显著低贡献度,均凸显了各类关键种群在植被结构缓冲降水波动中的枢纽作用。值得注意的是,相较于降水增加和降水变化总量,降水减少及其变化时机对植被关键种群结构更具干扰性。因此,半干旱区沙质草地植被恢复应优先保育具有跨降水梯度适应力的关键缓冲物种,而非单纯依赖优势种,并要考虑将返青期降水监测纳入早期预警体系,从而增强旱区植被结构的气候韧性。
参考文献
View Option
[1]
Knapp A K Hoover D L Wilcox K R et al Characterizing differences in precipitation regimes of extreme wet and dry years:implications for climate change experiments
[J].Global Change Biology ,2015 ,21 (7 ):2624 -2633 .
[本文引用: 1]
[2]
张宏史 ,郑秋红 半干旱地区天然草地灌丛化与土壤异质性关系研究进展
[J].植物生态学报 ,2001 ,25 (3 ):366 -370 .
[本文引用: 1]
[3]
Susanne S Osvaldo E S Hierarchy of responses to resource pulses in arid and semi-arid ecosystems
[J].Oecologia ,2004 ,141 (2 ):211 -220 .
[本文引用: 1]
[4]
王明明 ,刘新平 ,何玉惠 ,等 科尔沁沙质草地生物量积累过程对降水变化的响应模拟
[J].生态学报 ,2020 ,40 (11 ):3656 -3665 .
[本文引用: 1]
[5]
Luo Y Q Zhao X Y Zuo X A et al Plant responses to warming and increased precipitation in three categories of dune stabilization in northeastern China
[J].Ecological Research ,2017 ,32 (6 ):887 -898 .
[本文引用: 1]
[6]
张腊梅 ,刘新平 ,赵学勇 ,等 科尔沁固定沙地植被特征对降雨变化的响应
[J].生态学报 ,2014 ,34 (10 ):2737 -2745 .
[本文引用: 1]
[7]
Song J Liang Z H Li X G et al Precipitation changes alter plant dominant species and functional groups by changing soil salinity in a coastal salt marsh
[J].Journal of Environmental Management ,2024 ,368 :122235 .
[本文引用: 1]
[8]
Kirk R W Zheng S Gherardi G L et al Asymmetric responses of primary productivity to precipitation extremes:a synthesis of grassland precipitation manipulation experiments
[J].Global Change Biology ,2017 ,23 (10 ):4376 -4385 .
[本文引用: 1]
[9]
Pan S Y Zhang W L Li Y et al Unveiling novel perspectives on niche differentiation and plasticity in rhizosphere phosphorus forms of submerged macrophytes with different stoichiometric homeostasis
[J].Water Research ,2023 ,246 :120679 .
[本文引用: 1]
[10]
Yue X F Zhang T H Li Y Q Effects of rainfall regime during the growing season on the annual plant communities in semiarid sandy land,Northeast China
[J].Global Ecology and Conservation ,2023 ,43 :e02456 .
[本文引用: 2]
[11]
苗百岭 ,梁存柱 ,史亚博 ,等 降水变化对内蒙古典型草原地上生物量的影响
[J].植物生态学报 ,2019 ,43 (7 ):557 -565 .
[本文引用: 2]
[12]
Hautier Y Tilman D Isbell F et al Anthropogenic environmental changes affect ecosystem stability via biodiversity
[J].Science ,2015 ,348 (6232 ):336 -340 .
[本文引用: 2]
[13]
何玉惠 ,刘新平 ,胡鸿姣 ,等 降水和放牧对沙质草地优势植物出苗和存活的影响
[J].中国沙漠 ,2025 ,45 (1 ):103 -111 .
[本文引用: 1]
[14]
Liu X P He Y H Sun S S et al Restoration of sand-stabilizing vegetation reduces deep percolation of precipitation in semi-arid sandy lands,Northern China
[J].Catena ,2022 ,208 :105728 .
[本文引用: 1]
[15]
Liu X P Luo Y Q Cheng L et al Effect of root and mycelia on fine root decomposition and release of carbon and nitrogen under artemisia halodendron in a semi-arid sandy grassland in China
[J].Frontiers in Plant Science ,2021 ,12 :698054 .
[本文引用: 1]
[16]
刘新平 ,赵学勇 ,何玉惠 ,等 一种野外增减雨试验装置
:201220126834.3 [P].2012-11-21 .
[本文引用: 1]
[17]
郭倩 草地群落植物种等级划分及重要值方法改进
[D].兰州 :兰州大学 ,2017 .
[本文引用: 1]
[18]
Pei L Wu Z Qian Y Q et al Plant community stability,indicator species and their driving factors at a gradient of grazing intensity in an alpine meadow
[J].Ecological Indicators ,2024 ,162 :112012 .
[本文引用: 1]
[19]
Sun R B Chen Y Han W X et al Different contribution of species sorting and exogenous species immigration from manure to soil fungal diversity and community assemblage under long-term fertilization
[J].Soil Biology & Biochemistry ,2020 ,151 :108049 .
[本文引用: 1]
[20]
Zhang J Zhang B G Liu Y Q et al Distinct large-scale biogeographic patterns of fungal communities in bulk soil and soybean rhizosphere in China
[J].Science of the Total Environment ,2018 ,644 :791 -800 .
[本文引用: 2]
[21]
万凌凡 ,刘国华 ,樊辉 ,等 青藏高原东南部森林群落生态位特征与物种多样性的影响因素
[J].生态学报 ,2024 ,44 (13 ):5658 -5668 .
[本文引用: 1]
[22]
Huxman T E Snyder K A Tissue D et al Precipitation pulses and carbon fluxes in semiarid and arid ecosystems
[J].Oecologia ,2004 ,141 (2 ):211 -220 .
[本文引用: 1]
[23]
甄博 ,郭相平 ,陆红飞 ,等 分蘖期旱涝交替胁迫对水稻生理指标的影响
[J].灌溉排水学报 ,2017 ,36 (5 ):36 -40 .
[本文引用: 1]
[24]
Cai F Zhang Y S Mi N et al Maize (Zea mays L.) physiological responses to drought and rewatering,and the associations with water stress degree
[J].Agricultural Water Management ,2020 ,241 :106379 .
[本文引用: 1]
[25]
龚金玉 ,马致远 ,胡琛 ,等 综合生态系统管理的生态阈值理论研究进展
[J].生态学报 ,2024 ,44 (22 ):10499 -10511 .
[本文引用: 1]
[26]
洪光宇 ,王晓江 ,苏庆溥 ,等 毛乌素沙地流动沙丘土壤水分模拟及渗漏特征
[J].中国沙漠 ,2023 ,43 (2 ):288 -298 .
[本文引用: 1]
[27]
Zuo X A Zhao S L Cheng H et al Functional diversity response to geographic and experimental precipitation gradients varies with plant community type
[J].Functional Ecology ,2021 ,35 (9 ):2119 -2132 .
[本文引用: 1]
[28]
Huot C Zhou Y Philp J N M et al Root depth development in tropical perennial forage grasses is related to root angle,root diameter and leaf area
[J].Plant and Soil ,2020 ,456 (1/2 ):145 -158 .
[本文引用: 1]
[29]
Shan C Dongdong D Yubo L et al Genome-wide association study presents insights into the genetic architecture of drought tolerance in maize seedlings under field water-deficit conditions
[J].Frontiers in Plant Science ,2023 ,14 :1165582 .
[本文引用: 1]
[30]
游旭 ,杨浩 糙隐子草生态学研究进展
[J].草地学报 ,2016 ,24 (4 ):726 -730 .
[本文引用: 1]
[31]
孙启忠 ,韩桂荣 ,李志勇 ,等 科尔沁沙地达乌里胡枝子生物量研究
[J].中国草地 ,2001 ,23 (4 ):22 -27 .
[本文引用: 1]
[32]
郭文婷 ,王国华 ,缑倩倩 ,等 河西走廊荒漠绿洲过渡带3种典型一年生藜科植物构件生长及生物量分配特征
[J].草业学报 ,2022 ,31 (2 ):25 -38 .
[本文引用: 1]
[33]
刘菊红 ,王忠武 ,韩国栋 重度放牧对荒漠草原主要植物种间关系及群落稳定性的影响
[J].生态学杂志 ,2019 ,38 (9 ):2595 -2602 .
[本文引用: 1]
[34]
王晓悦 ,许艺馨 ,李春环 ,等 长期降水量变化下荒漠草原植物生物量、多样性的变化及其影响因素
[J].植物生态学报 ,2023 ,47 (4 ):479 -490 .
[本文引用: 1]
[35]
Sun J X Zhou H B Cheng H N et al Distinct strategies of the habitat generalists and specialists in the Arctic sediments: assembly processes,co-occurrence patterns,and environmental implications
[J].Marine Pollution Bulletin ,2024 ,205 :116603 .
[本文引用: 1]
[36]
Yu Y Penuelas J Sardans J et al Water addition but not reduction alters plant biomass-diversity relationship
[J].Global Change Biology ,2024 ,30 (7 ):e17430 .
[本文引用: 1]
[37]
李周园 ,叶小洲 ,王少鹏 生态系统稳定性及其与生物多样性的关系
[J].植物生态学报 ,2021 ,45 (10 ):1127 -1139 .
[本文引用: 1]
[38]
党承林 植物群落的冗余结构:对生态系统稳定性的一种解释
[J].生态学报 ,1998 ,18 (6 ):103 -110 .
[本文引用: 1]
[39]
丁潇然 狗尾草叶水力系统与光合生理沿降水梯度带的调整与协同
[D].杭州 :浙江农林大学 ,2022 .
[本文引用: 2]
[40]
王坤芳 ,彭爽 ,纪明山 入侵植物少花蒺藜草及其伴生植物的光合特性研究
[J].河北农业大学学报 ,2016 ,39 (1 ):43 -48 .
[本文引用: 1]
[41]
秦洁 ,鲍雅静 ,李政海 ,等 退化草原糙隐子草根系特征及地上高度对水分梯度的响应
[J].中国草地学报 ,2015 ,37 (3 ):80 -86 .
[本文引用: 2]
[42]
杨浩 ,罗亚晨 糙隐子草功能性状对氮添加和干旱的响应
[J].植物生态学报 ,2015 ,39 (1 ):32 -42 .
[本文引用: 1]
[43]
Yu S Y Wang W S Liang H X et al Characterizing the nonlinear dynamics of hydrological systems based on global recurrence analysis
[J].Journal of Hydrology ,2025 ,654 :132817 .
[本文引用: 1]
Characterizing differences in precipitation regimes of extreme wet and dry years:implications for climate change experiments
1
2015
... 全球气候变化驱动陆地生态系统水文格局重构,核心表现为降水时空异质性加剧,极端干旱与强降水事件频率、强度和季节分配呈现同步分异趋势;中高纬度干旱半干旱区域降水变率尤为显著,呈现出干的地方越干、湿的地方越湿的格局[1 ] .降水作为陆地生态系统中最重要的水分输入形式,其格局变化可改变水分可利用性阈值,显著调控植被群落的结构与功能[2 ] .尤其是在降水限制型生态系统中,植物种群对水分波动的敏感性构成生态系统稳定性与恢复力的核心调控因子[3 ] .半干旱沙质草地作为典型的脆弱生态系统,基质特性贫瘠,生态阈值狭窄,其关键植物种群功能性状调整与关键种替代机制已成为维系系统弹性的最后防线[4 ] :短期水分波动可能触发种群竞争优势的重新分配,而长期降水变化则可能驱动群落演替方向的根本性转变[5 ] .在此背景下,解析半干旱沙质草地关键植物种群对降水变化的响应策略,不仅是预测生态系统碳-水耦合过程的理论基石,也是制定适应性管理策略的科学前提. ...
半干旱地区天然草地灌丛化与土壤异质性关系研究进展
1
2001
... 全球气候变化驱动陆地生态系统水文格局重构,核心表现为降水时空异质性加剧,极端干旱与强降水事件频率、强度和季节分配呈现同步分异趋势;中高纬度干旱半干旱区域降水变率尤为显著,呈现出干的地方越干、湿的地方越湿的格局[1 ] .降水作为陆地生态系统中最重要的水分输入形式,其格局变化可改变水分可利用性阈值,显著调控植被群落的结构与功能[2 ] .尤其是在降水限制型生态系统中,植物种群对水分波动的敏感性构成生态系统稳定性与恢复力的核心调控因子[3 ] .半干旱沙质草地作为典型的脆弱生态系统,基质特性贫瘠,生态阈值狭窄,其关键植物种群功能性状调整与关键种替代机制已成为维系系统弹性的最后防线[4 ] :短期水分波动可能触发种群竞争优势的重新分配,而长期降水变化则可能驱动群落演替方向的根本性转变[5 ] .在此背景下,解析半干旱沙质草地关键植物种群对降水变化的响应策略,不仅是预测生态系统碳-水耦合过程的理论基石,也是制定适应性管理策略的科学前提. ...
Hierarchy of responses to resource pulses in arid and semi-arid ecosystems
1
2004
... 全球气候变化驱动陆地生态系统水文格局重构,核心表现为降水时空异质性加剧,极端干旱与强降水事件频率、强度和季节分配呈现同步分异趋势;中高纬度干旱半干旱区域降水变率尤为显著,呈现出干的地方越干、湿的地方越湿的格局[1 ] .降水作为陆地生态系统中最重要的水分输入形式,其格局变化可改变水分可利用性阈值,显著调控植被群落的结构与功能[2 ] .尤其是在降水限制型生态系统中,植物种群对水分波动的敏感性构成生态系统稳定性与恢复力的核心调控因子[3 ] .半干旱沙质草地作为典型的脆弱生态系统,基质特性贫瘠,生态阈值狭窄,其关键植物种群功能性状调整与关键种替代机制已成为维系系统弹性的最后防线[4 ] :短期水分波动可能触发种群竞争优势的重新分配,而长期降水变化则可能驱动群落演替方向的根本性转变[5 ] .在此背景下,解析半干旱沙质草地关键植物种群对降水变化的响应策略,不仅是预测生态系统碳-水耦合过程的理论基石,也是制定适应性管理策略的科学前提. ...
科尔沁沙质草地生物量积累过程对降水变化的响应模拟
1
2020
... 全球气候变化驱动陆地生态系统水文格局重构,核心表现为降水时空异质性加剧,极端干旱与强降水事件频率、强度和季节分配呈现同步分异趋势;中高纬度干旱半干旱区域降水变率尤为显著,呈现出干的地方越干、湿的地方越湿的格局[1 ] .降水作为陆地生态系统中最重要的水分输入形式,其格局变化可改变水分可利用性阈值,显著调控植被群落的结构与功能[2 ] .尤其是在降水限制型生态系统中,植物种群对水分波动的敏感性构成生态系统稳定性与恢复力的核心调控因子[3 ] .半干旱沙质草地作为典型的脆弱生态系统,基质特性贫瘠,生态阈值狭窄,其关键植物种群功能性状调整与关键种替代机制已成为维系系统弹性的最后防线[4 ] :短期水分波动可能触发种群竞争优势的重新分配,而长期降水变化则可能驱动群落演替方向的根本性转变[5 ] .在此背景下,解析半干旱沙质草地关键植物种群对降水变化的响应策略,不仅是预测生态系统碳-水耦合过程的理论基石,也是制定适应性管理策略的科学前提. ...
Plant responses to warming and increased precipitation in three categories of dune stabilization in northeastern China
1
2017
... 全球气候变化驱动陆地生态系统水文格局重构,核心表现为降水时空异质性加剧,极端干旱与强降水事件频率、强度和季节分配呈现同步分异趋势;中高纬度干旱半干旱区域降水变率尤为显著,呈现出干的地方越干、湿的地方越湿的格局[1 ] .降水作为陆地生态系统中最重要的水分输入形式,其格局变化可改变水分可利用性阈值,显著调控植被群落的结构与功能[2 ] .尤其是在降水限制型生态系统中,植物种群对水分波动的敏感性构成生态系统稳定性与恢复力的核心调控因子[3 ] .半干旱沙质草地作为典型的脆弱生态系统,基质特性贫瘠,生态阈值狭窄,其关键植物种群功能性状调整与关键种替代机制已成为维系系统弹性的最后防线[4 ] :短期水分波动可能触发种群竞争优势的重新分配,而长期降水变化则可能驱动群落演替方向的根本性转变[5 ] .在此背景下,解析半干旱沙质草地关键植物种群对降水变化的响应策略,不仅是预测生态系统碳-水耦合过程的理论基石,也是制定适应性管理策略的科学前提. ...
科尔沁固定沙地植被特征对降雨变化的响应
1
2014
... 重要值、指示值和生态位宽度作为表征植物种群响应环境变化的3个核心指标,分别从物种竞争优势、环境适应特征和资源利用策略3个维度揭示降水驱动的生态过程,其内在关联与作用差异是解析植被结构重组机制的关键科学问题.降水变化可通过改变植物相对盖度、高度和多度,显著影响物种重要值排序和关键种群组成[6 ] .例如在温带草原中,多年生禾草的重要值通常随降水减少而下降,一年生草本则通过快速生活史策略占据生态位空缺[7 ] .指示值方法揭示植物对特定环境条件的指示性,为识别关键指示种提供了方法:C4 植物在干旱处理中指示性显著提升,而C3 植物则更适应降水增加环境[8 ] .生态位宽度作为物种资源利用能力的表征,其动态变化反映种间竞争与环境筛选的协同作用.其中,泛化种通常具有较宽的生态位,能够在多种环境条件下生存,而特化种则对特定环境条件具有高度依赖性,并且极端干旱可导致泛化种生态位收缩,而适度降水增加则可促进特化种的功能分化[9 ] .尽管已有大量研究关注降水变化对植被的影响,但仍存在3个关键不足:首先,超一半的野外控制实验仅设置常态化降水梯度,难以捕捉极端气候事件的生态效应[10 ] .其次,现有研究多关注降水量的年际变化,却忽视季节性干旱对植物生活史策略的差异化影响.例如,生长季初期干旱可能通过延迟物候期影响一年生植物的萌发时间,而生长季盛期则直接抑制光合产物的积累[11 ] .另外,当前研究多侧重于群落水平的单一笼统结构性特征指标(如物种多样性)分析,往往割裂物种重要值、指示值及生态位宽度这3个指标的内在联系,导致关键种替代机制的解析存在偏差[12 ] .重要值揭示优势种动态,指示值筛选环境敏感种,生态位宽度量化种间竞争与适应策略.三者结合可避免单一指标的局限性,例如,高重要值物种可能缺乏环境指示性,而特化种虽指示性强但贡献度低[12 ] . ...
Precipitation changes alter plant dominant species and functional groups by changing soil salinity in a coastal salt marsh
1
2024
... 重要值、指示值和生态位宽度作为表征植物种群响应环境变化的3个核心指标,分别从物种竞争优势、环境适应特征和资源利用策略3个维度揭示降水驱动的生态过程,其内在关联与作用差异是解析植被结构重组机制的关键科学问题.降水变化可通过改变植物相对盖度、高度和多度,显著影响物种重要值排序和关键种群组成[6 ] .例如在温带草原中,多年生禾草的重要值通常随降水减少而下降,一年生草本则通过快速生活史策略占据生态位空缺[7 ] .指示值方法揭示植物对特定环境条件的指示性,为识别关键指示种提供了方法:C4 植物在干旱处理中指示性显著提升,而C3 植物则更适应降水增加环境[8 ] .生态位宽度作为物种资源利用能力的表征,其动态变化反映种间竞争与环境筛选的协同作用.其中,泛化种通常具有较宽的生态位,能够在多种环境条件下生存,而特化种则对特定环境条件具有高度依赖性,并且极端干旱可导致泛化种生态位收缩,而适度降水增加则可促进特化种的功能分化[9 ] .尽管已有大量研究关注降水变化对植被的影响,但仍存在3个关键不足:首先,超一半的野外控制实验仅设置常态化降水梯度,难以捕捉极端气候事件的生态效应[10 ] .其次,现有研究多关注降水量的年际变化,却忽视季节性干旱对植物生活史策略的差异化影响.例如,生长季初期干旱可能通过延迟物候期影响一年生植物的萌发时间,而生长季盛期则直接抑制光合产物的积累[11 ] .另外,当前研究多侧重于群落水平的单一笼统结构性特征指标(如物种多样性)分析,往往割裂物种重要值、指示值及生态位宽度这3个指标的内在联系,导致关键种替代机制的解析存在偏差[12 ] .重要值揭示优势种动态,指示值筛选环境敏感种,生态位宽度量化种间竞争与适应策略.三者结合可避免单一指标的局限性,例如,高重要值物种可能缺乏环境指示性,而特化种虽指示性强但贡献度低[12 ] . ...
Asymmetric responses of primary productivity to precipitation extremes:a synthesis of grassland precipitation manipulation experiments
1
2017
... 重要值、指示值和生态位宽度作为表征植物种群响应环境变化的3个核心指标,分别从物种竞争优势、环境适应特征和资源利用策略3个维度揭示降水驱动的生态过程,其内在关联与作用差异是解析植被结构重组机制的关键科学问题.降水变化可通过改变植物相对盖度、高度和多度,显著影响物种重要值排序和关键种群组成[6 ] .例如在温带草原中,多年生禾草的重要值通常随降水减少而下降,一年生草本则通过快速生活史策略占据生态位空缺[7 ] .指示值方法揭示植物对特定环境条件的指示性,为识别关键指示种提供了方法:C4 植物在干旱处理中指示性显著提升,而C3 植物则更适应降水增加环境[8 ] .生态位宽度作为物种资源利用能力的表征,其动态变化反映种间竞争与环境筛选的协同作用.其中,泛化种通常具有较宽的生态位,能够在多种环境条件下生存,而特化种则对特定环境条件具有高度依赖性,并且极端干旱可导致泛化种生态位收缩,而适度降水增加则可促进特化种的功能分化[9 ] .尽管已有大量研究关注降水变化对植被的影响,但仍存在3个关键不足:首先,超一半的野外控制实验仅设置常态化降水梯度,难以捕捉极端气候事件的生态效应[10 ] .其次,现有研究多关注降水量的年际变化,却忽视季节性干旱对植物生活史策略的差异化影响.例如,生长季初期干旱可能通过延迟物候期影响一年生植物的萌发时间,而生长季盛期则直接抑制光合产物的积累[11 ] .另外,当前研究多侧重于群落水平的单一笼统结构性特征指标(如物种多样性)分析,往往割裂物种重要值、指示值及生态位宽度这3个指标的内在联系,导致关键种替代机制的解析存在偏差[12 ] .重要值揭示优势种动态,指示值筛选环境敏感种,生态位宽度量化种间竞争与适应策略.三者结合可避免单一指标的局限性,例如,高重要值物种可能缺乏环境指示性,而特化种虽指示性强但贡献度低[12 ] . ...
Unveiling novel perspectives on niche differentiation and plasticity in rhizosphere phosphorus forms of submerged macrophytes with different stoichiometric homeostasis
1
2023
... 重要值、指示值和生态位宽度作为表征植物种群响应环境变化的3个核心指标,分别从物种竞争优势、环境适应特征和资源利用策略3个维度揭示降水驱动的生态过程,其内在关联与作用差异是解析植被结构重组机制的关键科学问题.降水变化可通过改变植物相对盖度、高度和多度,显著影响物种重要值排序和关键种群组成[6 ] .例如在温带草原中,多年生禾草的重要值通常随降水减少而下降,一年生草本则通过快速生活史策略占据生态位空缺[7 ] .指示值方法揭示植物对特定环境条件的指示性,为识别关键指示种提供了方法:C4 植物在干旱处理中指示性显著提升,而C3 植物则更适应降水增加环境[8 ] .生态位宽度作为物种资源利用能力的表征,其动态变化反映种间竞争与环境筛选的协同作用.其中,泛化种通常具有较宽的生态位,能够在多种环境条件下生存,而特化种则对特定环境条件具有高度依赖性,并且极端干旱可导致泛化种生态位收缩,而适度降水增加则可促进特化种的功能分化[9 ] .尽管已有大量研究关注降水变化对植被的影响,但仍存在3个关键不足:首先,超一半的野外控制实验仅设置常态化降水梯度,难以捕捉极端气候事件的生态效应[10 ] .其次,现有研究多关注降水量的年际变化,却忽视季节性干旱对植物生活史策略的差异化影响.例如,生长季初期干旱可能通过延迟物候期影响一年生植物的萌发时间,而生长季盛期则直接抑制光合产物的积累[11 ] .另外,当前研究多侧重于群落水平的单一笼统结构性特征指标(如物种多样性)分析,往往割裂物种重要值、指示值及生态位宽度这3个指标的内在联系,导致关键种替代机制的解析存在偏差[12 ] .重要值揭示优势种动态,指示值筛选环境敏感种,生态位宽度量化种间竞争与适应策略.三者结合可避免单一指标的局限性,例如,高重要值物种可能缺乏环境指示性,而特化种虽指示性强但贡献度低[12 ] . ...
Effects of rainfall regime during the growing season on the annual plant communities in semiarid sandy land,Northeast China
2
2023
... 重要值、指示值和生态位宽度作为表征植物种群响应环境变化的3个核心指标,分别从物种竞争优势、环境适应特征和资源利用策略3个维度揭示降水驱动的生态过程,其内在关联与作用差异是解析植被结构重组机制的关键科学问题.降水变化可通过改变植物相对盖度、高度和多度,显著影响物种重要值排序和关键种群组成[6 ] .例如在温带草原中,多年生禾草的重要值通常随降水减少而下降,一年生草本则通过快速生活史策略占据生态位空缺[7 ] .指示值方法揭示植物对特定环境条件的指示性,为识别关键指示种提供了方法:C4 植物在干旱处理中指示性显著提升,而C3 植物则更适应降水增加环境[8 ] .生态位宽度作为物种资源利用能力的表征,其动态变化反映种间竞争与环境筛选的协同作用.其中,泛化种通常具有较宽的生态位,能够在多种环境条件下生存,而特化种则对特定环境条件具有高度依赖性,并且极端干旱可导致泛化种生态位收缩,而适度降水增加则可促进特化种的功能分化[9 ] .尽管已有大量研究关注降水变化对植被的影响,但仍存在3个关键不足:首先,超一半的野外控制实验仅设置常态化降水梯度,难以捕捉极端气候事件的生态效应[10 ] .其次,现有研究多关注降水量的年际变化,却忽视季节性干旱对植物生活史策略的差异化影响.例如,生长季初期干旱可能通过延迟物候期影响一年生植物的萌发时间,而生长季盛期则直接抑制光合产物的积累[11 ] .另外,当前研究多侧重于群落水平的单一笼统结构性特征指标(如物种多样性)分析,往往割裂物种重要值、指示值及生态位宽度这3个指标的内在联系,导致关键种替代机制的解析存在偏差[12 ] .重要值揭示优势种动态,指示值筛选环境敏感种,生态位宽度量化种间竞争与适应策略.三者结合可避免单一指标的局限性,例如,高重要值物种可能缺乏环境指示性,而特化种虽指示性强但贡献度低[12 ] . ...
... 综合多维指标可见,优势种更替是植被结构响应降水变化的直观表现,指示种功能分异反映环境筛选的方向性,而生态位动态则揭示种间资源竞争本质.例如,降水减少60%处理下,糙隐子草(优势种)重要值骤降与其生态位宽度缩窄同步发生,而猪毛菜(指示种)通过快速占据生态位空缺成为新优势种,三者共同印证了资源适应能力下降→生态位释放→先锋种入侵的替代路径.多维指标协同分析,能更全面地揭示降水变化驱动植被结构调整的生态机制.广生态位物种或高重要值物种(如狗尾草、蒺藜、糙隐子草和画眉草)对降水处理组间差异的高贡献度特征(普遍P >0.05)提示其通过非显著性广谱参与在群落缓冲降水波动中发挥核心调控功能;而特殊生态位物种(如兴安胡枝子)的局部显著性贡献则表明狭生态位特化种可通过关键节点突破驱动差异格局.这表明旱区植被恢复应优先保育具有跨梯度适应力的缓冲物种,而非单纯依赖优势种[38 ] .首先,狗尾草作为一年生禾草,在极端降水处理中贡献度显著提升,其广域贡献度可能与其快速资源捕获策略相关[39 ] .尽管狗尾草在多个处理组间差异贡献度较高,但其显著性波动范围较大,这可能反映其作为“机会主义种”在不同处理中生态位宽度的动态调整[39 ] .与之类似,蒺藜作为一年生杂类草,可能通过抑制竞争者水分获取和增强干旱期水分存储能力实现生态位占领[40 ] .其次,糙隐子草作为多年生禾草,在极端降水处理中贡献度显著,但在适度降水变化中贡献度下降.这种双峰响应模式可能与其深层根系和休眠芽库策略相关:在极端湿润条件下,其分蘖能力增强[41 ] ;在极端干旱时,则通过地上部分生物量再分配维持生存[41 -42 ] .相比之下,画眉草多在D5处理中贡献度显著,表明其早春萌发特性可能使其在季节性干旱前期占据水分利用窗口期[10 ] .研究还发现,极端处理组间差异主要由多物种协同贡献(如极端干旱处理中狗尾草、蒺藜和糙隐子草联合贡献),而适度降水变化处理则依赖单一物种主导(如D5处理中画眉草贡献度较高),这表明降水扰动强度与群落响应模式间存在非线性关系:当降水偏离历史波动阈值(如±30%P)时,物种互作网络可能从补偿性响应转向竞争性替代[43 ] . ...
降水变化对内蒙古典型草原地上生物量的影响
2
2019
... 重要值、指示值和生态位宽度作为表征植物种群响应环境变化的3个核心指标,分别从物种竞争优势、环境适应特征和资源利用策略3个维度揭示降水驱动的生态过程,其内在关联与作用差异是解析植被结构重组机制的关键科学问题.降水变化可通过改变植物相对盖度、高度和多度,显著影响物种重要值排序和关键种群组成[6 ] .例如在温带草原中,多年生禾草的重要值通常随降水减少而下降,一年生草本则通过快速生活史策略占据生态位空缺[7 ] .指示值方法揭示植物对特定环境条件的指示性,为识别关键指示种提供了方法:C4 植物在干旱处理中指示性显著提升,而C3 植物则更适应降水增加环境[8 ] .生态位宽度作为物种资源利用能力的表征,其动态变化反映种间竞争与环境筛选的协同作用.其中,泛化种通常具有较宽的生态位,能够在多种环境条件下生存,而特化种则对特定环境条件具有高度依赖性,并且极端干旱可导致泛化种生态位收缩,而适度降水增加则可促进特化种的功能分化[9 ] .尽管已有大量研究关注降水变化对植被的影响,但仍存在3个关键不足:首先,超一半的野外控制实验仅设置常态化降水梯度,难以捕捉极端气候事件的生态效应[10 ] .其次,现有研究多关注降水量的年际变化,却忽视季节性干旱对植物生活史策略的差异化影响.例如,生长季初期干旱可能通过延迟物候期影响一年生植物的萌发时间,而生长季盛期则直接抑制光合产物的积累[11 ] .另外,当前研究多侧重于群落水平的单一笼统结构性特征指标(如物种多样性)分析,往往割裂物种重要值、指示值及生态位宽度这3个指标的内在联系,导致关键种替代机制的解析存在偏差[12 ] .重要值揭示优势种动态,指示值筛选环境敏感种,生态位宽度量化种间竞争与适应策略.三者结合可避免单一指标的局限性,例如,高重要值物种可能缺乏环境指示性,而特化种虽指示性强但贡献度低[12 ] . ...
... 科尔沁沙地位于中国北方农牧交错带典型生态脆弱区,其独特的沙质基质与高降水变率为解析上述问题提供了模式生态系统[11 ,13 ] .本研究基于科尔沁沙质草地植被,通过7年的降水模拟实验(涉及降水减少和增加、适度和极端、阶段性和连续性的降水变化格局),引入物种重要值、指示值及生态位宽度等多维种群特征指标,分析不同降水变化格局对植被种群结构特征的影响,旨在揭示以下科学问题:①降水变化如何通过优势种响应或更替驱动植被结构调整;②不同降水格局下指示种有何特征差异;③特化种与泛化种的生态位动态如何响应降水变化;④哪些关键种对植被结构响应降水格局转变具有显著贡献,通过以上研究以期为旱区脆弱生态系统植被恢复和管理保护提供理论依据和实践指导. ...
Anthropogenic environmental changes affect ecosystem stability via biodiversity
2
2015
... 重要值、指示值和生态位宽度作为表征植物种群响应环境变化的3个核心指标,分别从物种竞争优势、环境适应特征和资源利用策略3个维度揭示降水驱动的生态过程,其内在关联与作用差异是解析植被结构重组机制的关键科学问题.降水变化可通过改变植物相对盖度、高度和多度,显著影响物种重要值排序和关键种群组成[6 ] .例如在温带草原中,多年生禾草的重要值通常随降水减少而下降,一年生草本则通过快速生活史策略占据生态位空缺[7 ] .指示值方法揭示植物对特定环境条件的指示性,为识别关键指示种提供了方法:C4 植物在干旱处理中指示性显著提升,而C3 植物则更适应降水增加环境[8 ] .生态位宽度作为物种资源利用能力的表征,其动态变化反映种间竞争与环境筛选的协同作用.其中,泛化种通常具有较宽的生态位,能够在多种环境条件下生存,而特化种则对特定环境条件具有高度依赖性,并且极端干旱可导致泛化种生态位收缩,而适度降水增加则可促进特化种的功能分化[9 ] .尽管已有大量研究关注降水变化对植被的影响,但仍存在3个关键不足:首先,超一半的野外控制实验仅设置常态化降水梯度,难以捕捉极端气候事件的生态效应[10 ] .其次,现有研究多关注降水量的年际变化,却忽视季节性干旱对植物生活史策略的差异化影响.例如,生长季初期干旱可能通过延迟物候期影响一年生植物的萌发时间,而生长季盛期则直接抑制光合产物的积累[11 ] .另外,当前研究多侧重于群落水平的单一笼统结构性特征指标(如物种多样性)分析,往往割裂物种重要值、指示值及生态位宽度这3个指标的内在联系,导致关键种替代机制的解析存在偏差[12 ] .重要值揭示优势种动态,指示值筛选环境敏感种,生态位宽度量化种间竞争与适应策略.三者结合可避免单一指标的局限性,例如,高重要值物种可能缺乏环境指示性,而特化种虽指示性强但贡献度低[12 ] . ...
... [12 ]. ...
降水和放牧对沙质草地优势植物出苗和存活的影响
1
2025
... 科尔沁沙地位于中国北方农牧交错带典型生态脆弱区,其独特的沙质基质与高降水变率为解析上述问题提供了模式生态系统[11 ,13 ] .本研究基于科尔沁沙质草地植被,通过7年的降水模拟实验(涉及降水减少和增加、适度和极端、阶段性和连续性的降水变化格局),引入物种重要值、指示值及生态位宽度等多维种群特征指标,分析不同降水变化格局对植被种群结构特征的影响,旨在揭示以下科学问题:①降水变化如何通过优势种响应或更替驱动植被结构调整;②不同降水格局下指示种有何特征差异;③特化种与泛化种的生态位动态如何响应降水变化;④哪些关键种对植被结构响应降水格局转变具有显著贡献,通过以上研究以期为旱区脆弱生态系统植被恢复和管理保护提供理论依据和实践指导. ...
Restoration of sand-stabilizing vegetation reduces deep percolation of precipitation in semi-arid sandy lands,Northern China
1
2022
... 本研究在中国北部内蒙古东部科尔沁沙地(42°20′—44°50′N、117°15′—123°43′E,海拔约377 m)中南部的中国科学院奈曼沙漠化研究站开展.该区域被划定为半干旱农牧交错区,属温带大陆性半干旱气候.年平均气温约6.4 ℃,生长季4—9月日平均气温大多高于10 ℃.多年平均降水量约351.7 mm,其中生长季降水量占全年的67.5%~85.3%,年均潜在蒸发量1 935 mm.年降水量波动为-30%~30%,极端情况下可达60%(参照 1981—2023年科尔沁沙地气象站降水记录);春夏季降水变率较大,最长连续28天无降水[14 ] .地貌景观以各类沙丘为特征,土壤由沙(98.2%)、粉砂和黏土(1.8%)构成,物理性质均一,质地粗匀,结构疏松.植被主要由一年生草本植物组成,如大果虫实(Corispermum macrocarpum )、狗尾草(Setaria viridis )和针茅(Stipa capillata ).随着植被恢复进程的推进,多年生植物和禾本科植物的种类及数量所增加,例如猪毛蒿(Artemisia scoparia )、糙隐子草(Cleistogenes squarrosa )和砂引草(Tournefortia sibirica )[15 ] . ...
Effect of root and mycelia on fine root decomposition and release of carbon and nitrogen under artemisia halodendron in a semi-arid sandy grassland in China
1
2021
... 本研究在中国北部内蒙古东部科尔沁沙地(42°20′—44°50′N、117°15′—123°43′E,海拔约377 m)中南部的中国科学院奈曼沙漠化研究站开展.该区域被划定为半干旱农牧交错区,属温带大陆性半干旱气候.年平均气温约6.4 ℃,生长季4—9月日平均气温大多高于10 ℃.多年平均降水量约351.7 mm,其中生长季降水量占全年的67.5%~85.3%,年均潜在蒸发量1 935 mm.年降水量波动为-30%~30%,极端情况下可达60%(参照 1981—2023年科尔沁沙地气象站降水记录);春夏季降水变率较大,最长连续28天无降水[14 ] .地貌景观以各类沙丘为特征,土壤由沙(98.2%)、粉砂和黏土(1.8%)构成,物理性质均一,质地粗匀,结构疏松.植被主要由一年生草本植物组成,如大果虫实(Corispermum macrocarpum )、狗尾草(Setaria viridis )和针茅(Stipa capillata ).随着植被恢复进程的推进,多年生植物和禾本科植物的种类及数量所增加,例如猪毛蒿(Artemisia scoparia )、糙隐子草(Cleistogenes squarrosa )和砂引草(Tournefortia sibirica )[15 ] . ...
一种野外增减雨试验装置
1
... 以中国科学院奈曼沙漠化研究站内长期封育的典型沙质草地为研究样地(0.23 km2 ),于2016年建立沙质草地降水增减控制试验场(图1 ).依据研究区降水背景及植被对各季节降水变化的敏感性差异,共设置7个降水处理,分别为降水增加60%(+60%P)和降水减少60%(-60%P),模拟极端湿润和极端干旱事件;5月干旱(D5)和7月干旱(D7),模拟春季和夏季阶段性干旱事件;降水增加30%(+30%P)和降水减少30%(-30%P),模拟适度连续性降水变化事件;以及保持自然降水条件作为基准对照组(CK).除对照处理设置12个重复外,其余增减降水处理各设置6个重复,共48个处理小区,试验设计为完全随机区组设计.野外降水增减控制试验装置由奈曼站自主研发[16 ] ,对每年自然降水量进行梯度控制,其中降水减少区和降水增加区大小均为2 m×2 m,两者间缓冲区为2 m×2 m. ...
草地群落植物种等级划分及重要值方法改进
1
2017
... 物种重要值是研究某个物种在群落中的重要性的综合数量指标.基于样方调查数据,采用下列公式计算表示群落中某物种的相对重要值(Relative importance value,RIV)[17 ] .基于不同降水条件下植物群落内各物种的相对重要值,基于欧氏距离(Euclidean distance)进行分层聚类分析,筛选出重要值排名明显靠前的物种作为各降水处理下的优势种. ...
Plant community stability,indicator species and their driving factors at a gradient of grazing intensity in an alpine meadow
1
2024
... 基于不同降水处理下各植物种群重要值,使用labdsv::indval函数计算各种群指示值(IndVal指数)并筛选指示种[18 ] ;使用vegan::simper函数筛选组间差异贡献显著的物种[19 ] ;使用spaa::niche.width函数[20 ] 和EcolUtils::spec.gen函数定义泛化种、特化种和中性类群[20 ] . ...
Different contribution of species sorting and exogenous species immigration from manure to soil fungal diversity and community assemblage under long-term fertilization
1
2020
... 基于不同降水处理下各植物种群重要值,使用labdsv::indval函数计算各种群指示值(IndVal指数)并筛选指示种[18 ] ;使用vegan::simper函数筛选组间差异贡献显著的物种[19 ] ;使用spaa::niche.width函数[20 ] 和EcolUtils::spec.gen函数定义泛化种、特化种和中性类群[20 ] . ...
Distinct large-scale biogeographic patterns of fungal communities in bulk soil and soybean rhizosphere in China
2
2018
... 基于不同降水处理下各植物种群重要值,使用labdsv::indval函数计算各种群指示值(IndVal指数)并筛选指示种[18 ] ;使用vegan::simper函数筛选组间差异贡献显著的物种[19 ] ;使用spaa::niche.width函数[20 ] 和EcolUtils::spec.gen函数定义泛化种、特化种和中性类群[20 ] . ...
... [20 ]. ...
青藏高原东南部森林群落生态位特征与物种多样性的影响因素
1
2024
... 动优势种群组成与重要值排序的方向性调整,从而对植物群落结构产生深刻影响.本研究中,自然降水条件下,多年生禾草糙隐子草与一年生禾草狗尾草共同占据优势地位,构成科尔沁沙质草地植物群落核心架构.然而,当降水减少时,糙隐子草重要值骤降,其生态位甚至可被一年生植物,如猪毛菜、大果虫实等取代.糙隐子草作为泛化种,依赖广泛的资源获取能力维持优势,其生态位宽度在稳定环境中(CK)显著高于一年生植物猪毛菜,但在极端干旱下(-60%P),其资源适应能力受限,生态位宽度骤降[21 ] .相比之下,猪毛菜等一年生植物的生态位宽度也有下降,但其快速生长周期和种子繁殖策略更 ...
Precipitation pulses and carbon fluxes in semiarid and arid ecosystems
1
2004
... 适应极端环境下的资源波动,而非依赖广谱资源利用能力,因而其能在水分脉冲中占据短暂资源窗口.这表明生态位宽度并非唯一适应指标,生活史策略的权衡同样关键,体现出更强的环境筛选适应性[22 ] .物种更替具有显著的物候期依赖性.例如7月干旱仅使糙隐子草重要值下降,且未触发关键种替代,而5月干旱却促使糙隐子草退出关键种行列.这种差异可能源于植物生活史策略的阶段性约束:5月正值糙隐子草分蘖高峰期,此时水分亏缺直接抑制其繁殖能力[23 ] ,而7月干旱主要影响其生殖分配,对种群建立的冲击较小[24 ] .此外,物种更替同样验证了生态弹性阈值理论[25 ] ,当水分胁迫超过临界值,系统将进入以一年生植物为主导的演替新稳态;本研究中为极端干旱(降水减少60%)导致多年生禾草优势的不可逆丧失,提升了一年生植物的关键地位.与温带草原的研究不同,本研究发现适度和极端降水增加(降水增加30%和60%)均并未显著改变关键种组成,仅引起重要值排序微调(如画眉草重要值提升、狗尾草重要值下降).这可能与沙质土壤的高渗透性有关:短期内过量降水难以被土壤持留,导致水分利用效率未显著提升[26 ] .这一现象与降水增益促进多样性的经典认知不符,揭示在沙质生境中,降水增加的生态效应存在基质依赖性[27 ] .在降水增加处理下,糙隐子草重要值仍保持最高,表明多年生植物在湿润条件下具有更强的竞争力.这可能与多年生植物的深根系和较高的水分利用效率有关,使其能够在长期湿润条件下更有效地利用土壤水分[28 ] . ...
分蘖期旱涝交替胁迫对水稻生理指标的影响
1
2017
... 适应极端环境下的资源波动,而非依赖广谱资源利用能力,因而其能在水分脉冲中占据短暂资源窗口.这表明生态位宽度并非唯一适应指标,生活史策略的权衡同样关键,体现出更强的环境筛选适应性[22 ] .物种更替具有显著的物候期依赖性.例如7月干旱仅使糙隐子草重要值下降,且未触发关键种替代,而5月干旱却促使糙隐子草退出关键种行列.这种差异可能源于植物生活史策略的阶段性约束:5月正值糙隐子草分蘖高峰期,此时水分亏缺直接抑制其繁殖能力[23 ] ,而7月干旱主要影响其生殖分配,对种群建立的冲击较小[24 ] .此外,物种更替同样验证了生态弹性阈值理论[25 ] ,当水分胁迫超过临界值,系统将进入以一年生植物为主导的演替新稳态;本研究中为极端干旱(降水减少60%)导致多年生禾草优势的不可逆丧失,提升了一年生植物的关键地位.与温带草原的研究不同,本研究发现适度和极端降水增加(降水增加30%和60%)均并未显著改变关键种组成,仅引起重要值排序微调(如画眉草重要值提升、狗尾草重要值下降).这可能与沙质土壤的高渗透性有关:短期内过量降水难以被土壤持留,导致水分利用效率未显著提升[26 ] .这一现象与降水增益促进多样性的经典认知不符,揭示在沙质生境中,降水增加的生态效应存在基质依赖性[27 ] .在降水增加处理下,糙隐子草重要值仍保持最高,表明多年生植物在湿润条件下具有更强的竞争力.这可能与多年生植物的深根系和较高的水分利用效率有关,使其能够在长期湿润条件下更有效地利用土壤水分[28 ] . ...
Maize (Zea mays L.) physiological responses to drought and rewatering,and the associations with water stress degree
1
2020
... 适应极端环境下的资源波动,而非依赖广谱资源利用能力,因而其能在水分脉冲中占据短暂资源窗口.这表明生态位宽度并非唯一适应指标,生活史策略的权衡同样关键,体现出更强的环境筛选适应性[22 ] .物种更替具有显著的物候期依赖性.例如7月干旱仅使糙隐子草重要值下降,且未触发关键种替代,而5月干旱却促使糙隐子草退出关键种行列.这种差异可能源于植物生活史策略的阶段性约束:5月正值糙隐子草分蘖高峰期,此时水分亏缺直接抑制其繁殖能力[23 ] ,而7月干旱主要影响其生殖分配,对种群建立的冲击较小[24 ] .此外,物种更替同样验证了生态弹性阈值理论[25 ] ,当水分胁迫超过临界值,系统将进入以一年生植物为主导的演替新稳态;本研究中为极端干旱(降水减少60%)导致多年生禾草优势的不可逆丧失,提升了一年生植物的关键地位.与温带草原的研究不同,本研究发现适度和极端降水增加(降水增加30%和60%)均并未显著改变关键种组成,仅引起重要值排序微调(如画眉草重要值提升、狗尾草重要值下降).这可能与沙质土壤的高渗透性有关:短期内过量降水难以被土壤持留,导致水分利用效率未显著提升[26 ] .这一现象与降水增益促进多样性的经典认知不符,揭示在沙质生境中,降水增加的生态效应存在基质依赖性[27 ] .在降水增加处理下,糙隐子草重要值仍保持最高,表明多年生植物在湿润条件下具有更强的竞争力.这可能与多年生植物的深根系和较高的水分利用效率有关,使其能够在长期湿润条件下更有效地利用土壤水分[28 ] . ...
综合生态系统管理的生态阈值理论研究进展
1
2024
... 适应极端环境下的资源波动,而非依赖广谱资源利用能力,因而其能在水分脉冲中占据短暂资源窗口.这表明生态位宽度并非唯一适应指标,生活史策略的权衡同样关键,体现出更强的环境筛选适应性[22 ] .物种更替具有显著的物候期依赖性.例如7月干旱仅使糙隐子草重要值下降,且未触发关键种替代,而5月干旱却促使糙隐子草退出关键种行列.这种差异可能源于植物生活史策略的阶段性约束:5月正值糙隐子草分蘖高峰期,此时水分亏缺直接抑制其繁殖能力[23 ] ,而7月干旱主要影响其生殖分配,对种群建立的冲击较小[24 ] .此外,物种更替同样验证了生态弹性阈值理论[25 ] ,当水分胁迫超过临界值,系统将进入以一年生植物为主导的演替新稳态;本研究中为极端干旱(降水减少60%)导致多年生禾草优势的不可逆丧失,提升了一年生植物的关键地位.与温带草原的研究不同,本研究发现适度和极端降水增加(降水增加30%和60%)均并未显著改变关键种组成,仅引起重要值排序微调(如画眉草重要值提升、狗尾草重要值下降).这可能与沙质土壤的高渗透性有关:短期内过量降水难以被土壤持留,导致水分利用效率未显著提升[26 ] .这一现象与降水增益促进多样性的经典认知不符,揭示在沙质生境中,降水增加的生态效应存在基质依赖性[27 ] .在降水增加处理下,糙隐子草重要值仍保持最高,表明多年生植物在湿润条件下具有更强的竞争力.这可能与多年生植物的深根系和较高的水分利用效率有关,使其能够在长期湿润条件下更有效地利用土壤水分[28 ] . ...
毛乌素沙地流动沙丘土壤水分模拟及渗漏特征
1
2023
... 适应极端环境下的资源波动,而非依赖广谱资源利用能力,因而其能在水分脉冲中占据短暂资源窗口.这表明生态位宽度并非唯一适应指标,生活史策略的权衡同样关键,体现出更强的环境筛选适应性[22 ] .物种更替具有显著的物候期依赖性.例如7月干旱仅使糙隐子草重要值下降,且未触发关键种替代,而5月干旱却促使糙隐子草退出关键种行列.这种差异可能源于植物生活史策略的阶段性约束:5月正值糙隐子草分蘖高峰期,此时水分亏缺直接抑制其繁殖能力[23 ] ,而7月干旱主要影响其生殖分配,对种群建立的冲击较小[24 ] .此外,物种更替同样验证了生态弹性阈值理论[25 ] ,当水分胁迫超过临界值,系统将进入以一年生植物为主导的演替新稳态;本研究中为极端干旱(降水减少60%)导致多年生禾草优势的不可逆丧失,提升了一年生植物的关键地位.与温带草原的研究不同,本研究发现适度和极端降水增加(降水增加30%和60%)均并未显著改变关键种组成,仅引起重要值排序微调(如画眉草重要值提升、狗尾草重要值下降).这可能与沙质土壤的高渗透性有关:短期内过量降水难以被土壤持留,导致水分利用效率未显著提升[26 ] .这一现象与降水增益促进多样性的经典认知不符,揭示在沙质生境中,降水增加的生态效应存在基质依赖性[27 ] .在降水增加处理下,糙隐子草重要值仍保持最高,表明多年生植物在湿润条件下具有更强的竞争力.这可能与多年生植物的深根系和较高的水分利用效率有关,使其能够在长期湿润条件下更有效地利用土壤水分[28 ] . ...
Functional diversity response to geographic and experimental precipitation gradients varies with plant community type
1
2021
... 适应极端环境下的资源波动,而非依赖广谱资源利用能力,因而其能在水分脉冲中占据短暂资源窗口.这表明生态位宽度并非唯一适应指标,生活史策略的权衡同样关键,体现出更强的环境筛选适应性[22 ] .物种更替具有显著的物候期依赖性.例如7月干旱仅使糙隐子草重要值下降,且未触发关键种替代,而5月干旱却促使糙隐子草退出关键种行列.这种差异可能源于植物生活史策略的阶段性约束:5月正值糙隐子草分蘖高峰期,此时水分亏缺直接抑制其繁殖能力[23 ] ,而7月干旱主要影响其生殖分配,对种群建立的冲击较小[24 ] .此外,物种更替同样验证了生态弹性阈值理论[25 ] ,当水分胁迫超过临界值,系统将进入以一年生植物为主导的演替新稳态;本研究中为极端干旱(降水减少60%)导致多年生禾草优势的不可逆丧失,提升了一年生植物的关键地位.与温带草原的研究不同,本研究发现适度和极端降水增加(降水增加30%和60%)均并未显著改变关键种组成,仅引起重要值排序微调(如画眉草重要值提升、狗尾草重要值下降).这可能与沙质土壤的高渗透性有关:短期内过量降水难以被土壤持留,导致水分利用效率未显著提升[26 ] .这一现象与降水增益促进多样性的经典认知不符,揭示在沙质生境中,降水增加的生态效应存在基质依赖性[27 ] .在降水增加处理下,糙隐子草重要值仍保持最高,表明多年生植物在湿润条件下具有更强的竞争力.这可能与多年生植物的深根系和较高的水分利用效率有关,使其能够在长期湿润条件下更有效地利用土壤水分[28 ] . ...
Root depth development in tropical perennial forage grasses is related to root angle,root diameter and leaf area
1
2020
... 适应极端环境下的资源波动,而非依赖广谱资源利用能力,因而其能在水分脉冲中占据短暂资源窗口.这表明生态位宽度并非唯一适应指标,生活史策略的权衡同样关键,体现出更强的环境筛选适应性[22 ] .物种更替具有显著的物候期依赖性.例如7月干旱仅使糙隐子草重要值下降,且未触发关键种替代,而5月干旱却促使糙隐子草退出关键种行列.这种差异可能源于植物生活史策略的阶段性约束:5月正值糙隐子草分蘖高峰期,此时水分亏缺直接抑制其繁殖能力[23 ] ,而7月干旱主要影响其生殖分配,对种群建立的冲击较小[24 ] .此外,物种更替同样验证了生态弹性阈值理论[25 ] ,当水分胁迫超过临界值,系统将进入以一年生植物为主导的演替新稳态;本研究中为极端干旱(降水减少60%)导致多年生禾草优势的不可逆丧失,提升了一年生植物的关键地位.与温带草原的研究不同,本研究发现适度和极端降水增加(降水增加30%和60%)均并未显著改变关键种组成,仅引起重要值排序微调(如画眉草重要值提升、狗尾草重要值下降).这可能与沙质土壤的高渗透性有关:短期内过量降水难以被土壤持留,导致水分利用效率未显著提升[26 ] .这一现象与降水增益促进多样性的经典认知不符,揭示在沙质生境中,降水增加的生态效应存在基质依赖性[27 ] .在降水增加处理下,糙隐子草重要值仍保持最高,表明多年生植物在湿润条件下具有更强的竞争力.这可能与多年生植物的深根系和较高的水分利用效率有关,使其能够在长期湿润条件下更有效地利用土壤水分[28 ] . ...
Genome-wide association study presents insights into the genetic architecture of drought tolerance in maize seedlings under field water-deficit conditions
1
2023
... 指示种筛选体现环境胁迫对植物功能性状的定向选择.IndVal值与相对重要值的非一致性表明,指示种选择不仅依赖丰度,还与其对特定生境的生态专一性相关.例如,兴安胡枝子在降水减少30%处理下虽非优势种,但其对贫瘠土壤的适应能力使其成为关键指示物种.自然降水条件下指示种地锦草在干旱处理中被猪毛菜取代,而降水增加组则出现糙隐子草和猪毛蒿作为指示种.这一结果表明干旱强化了耐旱性状(如C4 光合途径、肉质叶片)的选择,而降水增加则筛选出具有高水分利用率的物种[29 ] .指示种生活型与降水处理呈现显著关联:降水增加组由多年生禾草主导,降水减少组以半灌木为主,而阶段性干旱组则由一年生杂类草占据优势.这种格局揭示了降水梯度对植物功能性状的筛选作用.例如,降水增加60%处理下糙隐子草显著指示性印证了其作为保守型策略者在湿润环境中的竞争优势[30 ] ;而降水减少组兴安胡枝子的指示性则凸显了半灌木在干旱胁迫下通过木质化结构降低蒸腾损耗的生存策略[31 ] ;一年生植物采用快速生长-繁殖的r策略应对短期资源波动[32 ] .另外,极端处理组(降水减少60%和降水增加60%)较低的指示值均值与高离散度并存,表明半干旱系统在极端降水下的脆弱性.当水分阈值超过植物适应能力边界时,原本稳定的种间互作网络可能瓦解,导致指示种特异性下降[33 ] . ...
糙隐子草生态学研究进展
1
2016
... 指示种筛选体现环境胁迫对植物功能性状的定向选择.IndVal值与相对重要值的非一致性表明,指示种选择不仅依赖丰度,还与其对特定生境的生态专一性相关.例如,兴安胡枝子在降水减少30%处理下虽非优势种,但其对贫瘠土壤的适应能力使其成为关键指示物种.自然降水条件下指示种地锦草在干旱处理中被猪毛菜取代,而降水增加组则出现糙隐子草和猪毛蒿作为指示种.这一结果表明干旱强化了耐旱性状(如C4 光合途径、肉质叶片)的选择,而降水增加则筛选出具有高水分利用率的物种[29 ] .指示种生活型与降水处理呈现显著关联:降水增加组由多年生禾草主导,降水减少组以半灌木为主,而阶段性干旱组则由一年生杂类草占据优势.这种格局揭示了降水梯度对植物功能性状的筛选作用.例如,降水增加60%处理下糙隐子草显著指示性印证了其作为保守型策略者在湿润环境中的竞争优势[30 ] ;而降水减少组兴安胡枝子的指示性则凸显了半灌木在干旱胁迫下通过木质化结构降低蒸腾损耗的生存策略[31 ] ;一年生植物采用快速生长-繁殖的r策略应对短期资源波动[32 ] .另外,极端处理组(降水减少60%和降水增加60%)较低的指示值均值与高离散度并存,表明半干旱系统在极端降水下的脆弱性.当水分阈值超过植物适应能力边界时,原本稳定的种间互作网络可能瓦解,导致指示种特异性下降[33 ] . ...
科尔沁沙地达乌里胡枝子生物量研究
1
2001
... 指示种筛选体现环境胁迫对植物功能性状的定向选择.IndVal值与相对重要值的非一致性表明,指示种选择不仅依赖丰度,还与其对特定生境的生态专一性相关.例如,兴安胡枝子在降水减少30%处理下虽非优势种,但其对贫瘠土壤的适应能力使其成为关键指示物种.自然降水条件下指示种地锦草在干旱处理中被猪毛菜取代,而降水增加组则出现糙隐子草和猪毛蒿作为指示种.这一结果表明干旱强化了耐旱性状(如C4 光合途径、肉质叶片)的选择,而降水增加则筛选出具有高水分利用率的物种[29 ] .指示种生活型与降水处理呈现显著关联:降水增加组由多年生禾草主导,降水减少组以半灌木为主,而阶段性干旱组则由一年生杂类草占据优势.这种格局揭示了降水梯度对植物功能性状的筛选作用.例如,降水增加60%处理下糙隐子草显著指示性印证了其作为保守型策略者在湿润环境中的竞争优势[30 ] ;而降水减少组兴安胡枝子的指示性则凸显了半灌木在干旱胁迫下通过木质化结构降低蒸腾损耗的生存策略[31 ] ;一年生植物采用快速生长-繁殖的r策略应对短期资源波动[32 ] .另外,极端处理组(降水减少60%和降水增加60%)较低的指示值均值与高离散度并存,表明半干旱系统在极端降水下的脆弱性.当水分阈值超过植物适应能力边界时,原本稳定的种间互作网络可能瓦解,导致指示种特异性下降[33 ] . ...
河西走廊荒漠绿洲过渡带3种典型一年生藜科植物构件生长及生物量分配特征
1
2022
... 指示种筛选体现环境胁迫对植物功能性状的定向选择.IndVal值与相对重要值的非一致性表明,指示种选择不仅依赖丰度,还与其对特定生境的生态专一性相关.例如,兴安胡枝子在降水减少30%处理下虽非优势种,但其对贫瘠土壤的适应能力使其成为关键指示物种.自然降水条件下指示种地锦草在干旱处理中被猪毛菜取代,而降水增加组则出现糙隐子草和猪毛蒿作为指示种.这一结果表明干旱强化了耐旱性状(如C4 光合途径、肉质叶片)的选择,而降水增加则筛选出具有高水分利用率的物种[29 ] .指示种生活型与降水处理呈现显著关联:降水增加组由多年生禾草主导,降水减少组以半灌木为主,而阶段性干旱组则由一年生杂类草占据优势.这种格局揭示了降水梯度对植物功能性状的筛选作用.例如,降水增加60%处理下糙隐子草显著指示性印证了其作为保守型策略者在湿润环境中的竞争优势[30 ] ;而降水减少组兴安胡枝子的指示性则凸显了半灌木在干旱胁迫下通过木质化结构降低蒸腾损耗的生存策略[31 ] ;一年生植物采用快速生长-繁殖的r策略应对短期资源波动[32 ] .另外,极端处理组(降水减少60%和降水增加60%)较低的指示值均值与高离散度并存,表明半干旱系统在极端降水下的脆弱性.当水分阈值超过植物适应能力边界时,原本稳定的种间互作网络可能瓦解,导致指示种特异性下降[33 ] . ...
重度放牧对荒漠草原主要植物种间关系及群落稳定性的影响
1
2019
... 指示种筛选体现环境胁迫对植物功能性状的定向选择.IndVal值与相对重要值的非一致性表明,指示种选择不仅依赖丰度,还与其对特定生境的生态专一性相关.例如,兴安胡枝子在降水减少30%处理下虽非优势种,但其对贫瘠土壤的适应能力使其成为关键指示物种.自然降水条件下指示种地锦草在干旱处理中被猪毛菜取代,而降水增加组则出现糙隐子草和猪毛蒿作为指示种.这一结果表明干旱强化了耐旱性状(如C4 光合途径、肉质叶片)的选择,而降水增加则筛选出具有高水分利用率的物种[29 ] .指示种生活型与降水处理呈现显著关联:降水增加组由多年生禾草主导,降水减少组以半灌木为主,而阶段性干旱组则由一年生杂类草占据优势.这种格局揭示了降水梯度对植物功能性状的筛选作用.例如,降水增加60%处理下糙隐子草显著指示性印证了其作为保守型策略者在湿润环境中的竞争优势[30 ] ;而降水减少组兴安胡枝子的指示性则凸显了半灌木在干旱胁迫下通过木质化结构降低蒸腾损耗的生存策略[31 ] ;一年生植物采用快速生长-繁殖的r策略应对短期资源波动[32 ] .另外,极端处理组(降水减少60%和降水增加60%)较低的指示值均值与高离散度并存,表明半干旱系统在极端降水下的脆弱性.当水分阈值超过植物适应能力边界时,原本稳定的种间互作网络可能瓦解,导致指示种特异性下降[33 ] . ...
长期降水量变化下荒漠草原植物生物量、多样性的变化及其影响因素
1
2023
... 生态位宽度变化揭示物种资源利用策略的环境敏感性差异.泛化种和特化种在群落中比例变化也反映了群落对降水变化的适应能力.自然降水条件下,糙隐子草(泛化种)与兴安胡枝子(特化种)形成互补的生态位格局,而降水减少60%处理使泛化种(猪毛菜)的生态位宽度下降,同时特化种(野艾蒿)消失.这表明泛化种在极端环境条件下具有更强的适应能力,能够通过调整资源利用策略维持群落的稳定性.这一现象可能由两种机制驱动,一是资源竞争失衡:干旱导致土壤水分与养分空间异质性增强,泛化种因无法在多维资源梯度中保持竞争力而衰退[34 ] ;二是环境筛选强化:极端干旱形成强烈的非生物过滤,淘汰了依赖稳定水分供给的特化种,使适应脉冲式资源供应的中性类群(如一年生草本)占据优势[35 ] .降水增加30%处理虽未改变泛化种/特化种的组成地位,但导致其物种替换(野艾蒿→猪毛蒿).这表明适度水分增加可能通过改变种间互作(如互利共生或竞争排除)而非直接环境筛选驱动生态位重组[36 ] .此外,中性类群在特化种/泛化种受抑时竞争优势提升,表明非优势种通过填补生态位空缺维持系统功能的“保险效应”,当环境压力超过物种适应阈值时,具有广泛耐受性的中性物种可通过生态位漂移占据空缺资源[37 ] . ...
Distinct strategies of the habitat generalists and specialists in the Arctic sediments: assembly processes,co-occurrence patterns,and environmental implications
1
2024
... 生态位宽度变化揭示物种资源利用策略的环境敏感性差异.泛化种和特化种在群落中比例变化也反映了群落对降水变化的适应能力.自然降水条件下,糙隐子草(泛化种)与兴安胡枝子(特化种)形成互补的生态位格局,而降水减少60%处理使泛化种(猪毛菜)的生态位宽度下降,同时特化种(野艾蒿)消失.这表明泛化种在极端环境条件下具有更强的适应能力,能够通过调整资源利用策略维持群落的稳定性.这一现象可能由两种机制驱动,一是资源竞争失衡:干旱导致土壤水分与养分空间异质性增强,泛化种因无法在多维资源梯度中保持竞争力而衰退[34 ] ;二是环境筛选强化:极端干旱形成强烈的非生物过滤,淘汰了依赖稳定水分供给的特化种,使适应脉冲式资源供应的中性类群(如一年生草本)占据优势[35 ] .降水增加30%处理虽未改变泛化种/特化种的组成地位,但导致其物种替换(野艾蒿→猪毛蒿).这表明适度水分增加可能通过改变种间互作(如互利共生或竞争排除)而非直接环境筛选驱动生态位重组[36 ] .此外,中性类群在特化种/泛化种受抑时竞争优势提升,表明非优势种通过填补生态位空缺维持系统功能的“保险效应”,当环境压力超过物种适应阈值时,具有广泛耐受性的中性物种可通过生态位漂移占据空缺资源[37 ] . ...
Water addition but not reduction alters plant biomass-diversity relationship
1
2024
... 生态位宽度变化揭示物种资源利用策略的环境敏感性差异.泛化种和特化种在群落中比例变化也反映了群落对降水变化的适应能力.自然降水条件下,糙隐子草(泛化种)与兴安胡枝子(特化种)形成互补的生态位格局,而降水减少60%处理使泛化种(猪毛菜)的生态位宽度下降,同时特化种(野艾蒿)消失.这表明泛化种在极端环境条件下具有更强的适应能力,能够通过调整资源利用策略维持群落的稳定性.这一现象可能由两种机制驱动,一是资源竞争失衡:干旱导致土壤水分与养分空间异质性增强,泛化种因无法在多维资源梯度中保持竞争力而衰退[34 ] ;二是环境筛选强化:极端干旱形成强烈的非生物过滤,淘汰了依赖稳定水分供给的特化种,使适应脉冲式资源供应的中性类群(如一年生草本)占据优势[35 ] .降水增加30%处理虽未改变泛化种/特化种的组成地位,但导致其物种替换(野艾蒿→猪毛蒿).这表明适度水分增加可能通过改变种间互作(如互利共生或竞争排除)而非直接环境筛选驱动生态位重组[36 ] .此外,中性类群在特化种/泛化种受抑时竞争优势提升,表明非优势种通过填补生态位空缺维持系统功能的“保险效应”,当环境压力超过物种适应阈值时,具有广泛耐受性的中性物种可通过生态位漂移占据空缺资源[37 ] . ...
生态系统稳定性及其与生物多样性的关系
1
2021
... 生态位宽度变化揭示物种资源利用策略的环境敏感性差异.泛化种和特化种在群落中比例变化也反映了群落对降水变化的适应能力.自然降水条件下,糙隐子草(泛化种)与兴安胡枝子(特化种)形成互补的生态位格局,而降水减少60%处理使泛化种(猪毛菜)的生态位宽度下降,同时特化种(野艾蒿)消失.这表明泛化种在极端环境条件下具有更强的适应能力,能够通过调整资源利用策略维持群落的稳定性.这一现象可能由两种机制驱动,一是资源竞争失衡:干旱导致土壤水分与养分空间异质性增强,泛化种因无法在多维资源梯度中保持竞争力而衰退[34 ] ;二是环境筛选强化:极端干旱形成强烈的非生物过滤,淘汰了依赖稳定水分供给的特化种,使适应脉冲式资源供应的中性类群(如一年生草本)占据优势[35 ] .降水增加30%处理虽未改变泛化种/特化种的组成地位,但导致其物种替换(野艾蒿→猪毛蒿).这表明适度水分增加可能通过改变种间互作(如互利共生或竞争排除)而非直接环境筛选驱动生态位重组[36 ] .此外,中性类群在特化种/泛化种受抑时竞争优势提升,表明非优势种通过填补生态位空缺维持系统功能的“保险效应”,当环境压力超过物种适应阈值时,具有广泛耐受性的中性物种可通过生态位漂移占据空缺资源[37 ] . ...
植物群落的冗余结构:对生态系统稳定性的一种解释
1
1998
... 综合多维指标可见,优势种更替是植被结构响应降水变化的直观表现,指示种功能分异反映环境筛选的方向性,而生态位动态则揭示种间资源竞争本质.例如,降水减少60%处理下,糙隐子草(优势种)重要值骤降与其生态位宽度缩窄同步发生,而猪毛菜(指示种)通过快速占据生态位空缺成为新优势种,三者共同印证了资源适应能力下降→生态位释放→先锋种入侵的替代路径.多维指标协同分析,能更全面地揭示降水变化驱动植被结构调整的生态机制.广生态位物种或高重要值物种(如狗尾草、蒺藜、糙隐子草和画眉草)对降水处理组间差异的高贡献度特征(普遍P >0.05)提示其通过非显著性广谱参与在群落缓冲降水波动中发挥核心调控功能;而特殊生态位物种(如兴安胡枝子)的局部显著性贡献则表明狭生态位特化种可通过关键节点突破驱动差异格局.这表明旱区植被恢复应优先保育具有跨梯度适应力的缓冲物种,而非单纯依赖优势种[38 ] .首先,狗尾草作为一年生禾草,在极端降水处理中贡献度显著提升,其广域贡献度可能与其快速资源捕获策略相关[39 ] .尽管狗尾草在多个处理组间差异贡献度较高,但其显著性波动范围较大,这可能反映其作为“机会主义种”在不同处理中生态位宽度的动态调整[39 ] .与之类似,蒺藜作为一年生杂类草,可能通过抑制竞争者水分获取和增强干旱期水分存储能力实现生态位占领[40 ] .其次,糙隐子草作为多年生禾草,在极端降水处理中贡献度显著,但在适度降水变化中贡献度下降.这种双峰响应模式可能与其深层根系和休眠芽库策略相关:在极端湿润条件下,其分蘖能力增强[41 ] ;在极端干旱时,则通过地上部分生物量再分配维持生存[41 -42 ] .相比之下,画眉草多在D5处理中贡献度显著,表明其早春萌发特性可能使其在季节性干旱前期占据水分利用窗口期[10 ] .研究还发现,极端处理组间差异主要由多物种协同贡献(如极端干旱处理中狗尾草、蒺藜和糙隐子草联合贡献),而适度降水变化处理则依赖单一物种主导(如D5处理中画眉草贡献度较高),这表明降水扰动强度与群落响应模式间存在非线性关系:当降水偏离历史波动阈值(如±30%P)时,物种互作网络可能从补偿性响应转向竞争性替代[43 ] . ...
狗尾草叶水力系统与光合生理沿降水梯度带的调整与协同
2
2022
... 综合多维指标可见,优势种更替是植被结构响应降水变化的直观表现,指示种功能分异反映环境筛选的方向性,而生态位动态则揭示种间资源竞争本质.例如,降水减少60%处理下,糙隐子草(优势种)重要值骤降与其生态位宽度缩窄同步发生,而猪毛菜(指示种)通过快速占据生态位空缺成为新优势种,三者共同印证了资源适应能力下降→生态位释放→先锋种入侵的替代路径.多维指标协同分析,能更全面地揭示降水变化驱动植被结构调整的生态机制.广生态位物种或高重要值物种(如狗尾草、蒺藜、糙隐子草和画眉草)对降水处理组间差异的高贡献度特征(普遍P >0.05)提示其通过非显著性广谱参与在群落缓冲降水波动中发挥核心调控功能;而特殊生态位物种(如兴安胡枝子)的局部显著性贡献则表明狭生态位特化种可通过关键节点突破驱动差异格局.这表明旱区植被恢复应优先保育具有跨梯度适应力的缓冲物种,而非单纯依赖优势种[38 ] .首先,狗尾草作为一年生禾草,在极端降水处理中贡献度显著提升,其广域贡献度可能与其快速资源捕获策略相关[39 ] .尽管狗尾草在多个处理组间差异贡献度较高,但其显著性波动范围较大,这可能反映其作为“机会主义种”在不同处理中生态位宽度的动态调整[39 ] .与之类似,蒺藜作为一年生杂类草,可能通过抑制竞争者水分获取和增强干旱期水分存储能力实现生态位占领[40 ] .其次,糙隐子草作为多年生禾草,在极端降水处理中贡献度显著,但在适度降水变化中贡献度下降.这种双峰响应模式可能与其深层根系和休眠芽库策略相关:在极端湿润条件下,其分蘖能力增强[41 ] ;在极端干旱时,则通过地上部分生物量再分配维持生存[41 -42 ] .相比之下,画眉草多在D5处理中贡献度显著,表明其早春萌发特性可能使其在季节性干旱前期占据水分利用窗口期[10 ] .研究还发现,极端处理组间差异主要由多物种协同贡献(如极端干旱处理中狗尾草、蒺藜和糙隐子草联合贡献),而适度降水变化处理则依赖单一物种主导(如D5处理中画眉草贡献度较高),这表明降水扰动强度与群落响应模式间存在非线性关系:当降水偏离历史波动阈值(如±30%P)时,物种互作网络可能从补偿性响应转向竞争性替代[43 ] . ...
... [39 ].与之类似,蒺藜作为一年生杂类草,可能通过抑制竞争者水分获取和增强干旱期水分存储能力实现生态位占领[40 ] .其次,糙隐子草作为多年生禾草,在极端降水处理中贡献度显著,但在适度降水变化中贡献度下降.这种双峰响应模式可能与其深层根系和休眠芽库策略相关:在极端湿润条件下,其分蘖能力增强[41 ] ;在极端干旱时,则通过地上部分生物量再分配维持生存[41 -42 ] .相比之下,画眉草多在D5处理中贡献度显著,表明其早春萌发特性可能使其在季节性干旱前期占据水分利用窗口期[10 ] .研究还发现,极端处理组间差异主要由多物种协同贡献(如极端干旱处理中狗尾草、蒺藜和糙隐子草联合贡献),而适度降水变化处理则依赖单一物种主导(如D5处理中画眉草贡献度较高),这表明降水扰动强度与群落响应模式间存在非线性关系:当降水偏离历史波动阈值(如±30%P)时,物种互作网络可能从补偿性响应转向竞争性替代[43 ] . ...
入侵植物少花蒺藜草及其伴生植物的光合特性研究
1
2016
... 综合多维指标可见,优势种更替是植被结构响应降水变化的直观表现,指示种功能分异反映环境筛选的方向性,而生态位动态则揭示种间资源竞争本质.例如,降水减少60%处理下,糙隐子草(优势种)重要值骤降与其生态位宽度缩窄同步发生,而猪毛菜(指示种)通过快速占据生态位空缺成为新优势种,三者共同印证了资源适应能力下降→生态位释放→先锋种入侵的替代路径.多维指标协同分析,能更全面地揭示降水变化驱动植被结构调整的生态机制.广生态位物种或高重要值物种(如狗尾草、蒺藜、糙隐子草和画眉草)对降水处理组间差异的高贡献度特征(普遍P >0.05)提示其通过非显著性广谱参与在群落缓冲降水波动中发挥核心调控功能;而特殊生态位物种(如兴安胡枝子)的局部显著性贡献则表明狭生态位特化种可通过关键节点突破驱动差异格局.这表明旱区植被恢复应优先保育具有跨梯度适应力的缓冲物种,而非单纯依赖优势种[38 ] .首先,狗尾草作为一年生禾草,在极端降水处理中贡献度显著提升,其广域贡献度可能与其快速资源捕获策略相关[39 ] .尽管狗尾草在多个处理组间差异贡献度较高,但其显著性波动范围较大,这可能反映其作为“机会主义种”在不同处理中生态位宽度的动态调整[39 ] .与之类似,蒺藜作为一年生杂类草,可能通过抑制竞争者水分获取和增强干旱期水分存储能力实现生态位占领[40 ] .其次,糙隐子草作为多年生禾草,在极端降水处理中贡献度显著,但在适度降水变化中贡献度下降.这种双峰响应模式可能与其深层根系和休眠芽库策略相关:在极端湿润条件下,其分蘖能力增强[41 ] ;在极端干旱时,则通过地上部分生物量再分配维持生存[41 -42 ] .相比之下,画眉草多在D5处理中贡献度显著,表明其早春萌发特性可能使其在季节性干旱前期占据水分利用窗口期[10 ] .研究还发现,极端处理组间差异主要由多物种协同贡献(如极端干旱处理中狗尾草、蒺藜和糙隐子草联合贡献),而适度降水变化处理则依赖单一物种主导(如D5处理中画眉草贡献度较高),这表明降水扰动强度与群落响应模式间存在非线性关系:当降水偏离历史波动阈值(如±30%P)时,物种互作网络可能从补偿性响应转向竞争性替代[43 ] . ...
退化草原糙隐子草根系特征及地上高度对水分梯度的响应
2
2015
... 综合多维指标可见,优势种更替是植被结构响应降水变化的直观表现,指示种功能分异反映环境筛选的方向性,而生态位动态则揭示种间资源竞争本质.例如,降水减少60%处理下,糙隐子草(优势种)重要值骤降与其生态位宽度缩窄同步发生,而猪毛菜(指示种)通过快速占据生态位空缺成为新优势种,三者共同印证了资源适应能力下降→生态位释放→先锋种入侵的替代路径.多维指标协同分析,能更全面地揭示降水变化驱动植被结构调整的生态机制.广生态位物种或高重要值物种(如狗尾草、蒺藜、糙隐子草和画眉草)对降水处理组间差异的高贡献度特征(普遍P >0.05)提示其通过非显著性广谱参与在群落缓冲降水波动中发挥核心调控功能;而特殊生态位物种(如兴安胡枝子)的局部显著性贡献则表明狭生态位特化种可通过关键节点突破驱动差异格局.这表明旱区植被恢复应优先保育具有跨梯度适应力的缓冲物种,而非单纯依赖优势种[38 ] .首先,狗尾草作为一年生禾草,在极端降水处理中贡献度显著提升,其广域贡献度可能与其快速资源捕获策略相关[39 ] .尽管狗尾草在多个处理组间差异贡献度较高,但其显著性波动范围较大,这可能反映其作为“机会主义种”在不同处理中生态位宽度的动态调整[39 ] .与之类似,蒺藜作为一年生杂类草,可能通过抑制竞争者水分获取和增强干旱期水分存储能力实现生态位占领[40 ] .其次,糙隐子草作为多年生禾草,在极端降水处理中贡献度显著,但在适度降水变化中贡献度下降.这种双峰响应模式可能与其深层根系和休眠芽库策略相关:在极端湿润条件下,其分蘖能力增强[41 ] ;在极端干旱时,则通过地上部分生物量再分配维持生存[41 -42 ] .相比之下,画眉草多在D5处理中贡献度显著,表明其早春萌发特性可能使其在季节性干旱前期占据水分利用窗口期[10 ] .研究还发现,极端处理组间差异主要由多物种协同贡献(如极端干旱处理中狗尾草、蒺藜和糙隐子草联合贡献),而适度降水变化处理则依赖单一物种主导(如D5处理中画眉草贡献度较高),这表明降水扰动强度与群落响应模式间存在非线性关系:当降水偏离历史波动阈值(如±30%P)时,物种互作网络可能从补偿性响应转向竞争性替代[43 ] . ...
... [41 -42 ].相比之下,画眉草多在D5处理中贡献度显著,表明其早春萌发特性可能使其在季节性干旱前期占据水分利用窗口期[10 ] .研究还发现,极端处理组间差异主要由多物种协同贡献(如极端干旱处理中狗尾草、蒺藜和糙隐子草联合贡献),而适度降水变化处理则依赖单一物种主导(如D5处理中画眉草贡献度较高),这表明降水扰动强度与群落响应模式间存在非线性关系:当降水偏离历史波动阈值(如±30%P)时,物种互作网络可能从补偿性响应转向竞争性替代[43 ] . ...
糙隐子草功能性状对氮添加和干旱的响应
1
2015
... 综合多维指标可见,优势种更替是植被结构响应降水变化的直观表现,指示种功能分异反映环境筛选的方向性,而生态位动态则揭示种间资源竞争本质.例如,降水减少60%处理下,糙隐子草(优势种)重要值骤降与其生态位宽度缩窄同步发生,而猪毛菜(指示种)通过快速占据生态位空缺成为新优势种,三者共同印证了资源适应能力下降→生态位释放→先锋种入侵的替代路径.多维指标协同分析,能更全面地揭示降水变化驱动植被结构调整的生态机制.广生态位物种或高重要值物种(如狗尾草、蒺藜、糙隐子草和画眉草)对降水处理组间差异的高贡献度特征(普遍P >0.05)提示其通过非显著性广谱参与在群落缓冲降水波动中发挥核心调控功能;而特殊生态位物种(如兴安胡枝子)的局部显著性贡献则表明狭生态位特化种可通过关键节点突破驱动差异格局.这表明旱区植被恢复应优先保育具有跨梯度适应力的缓冲物种,而非单纯依赖优势种[38 ] .首先,狗尾草作为一年生禾草,在极端降水处理中贡献度显著提升,其广域贡献度可能与其快速资源捕获策略相关[39 ] .尽管狗尾草在多个处理组间差异贡献度较高,但其显著性波动范围较大,这可能反映其作为“机会主义种”在不同处理中生态位宽度的动态调整[39 ] .与之类似,蒺藜作为一年生杂类草,可能通过抑制竞争者水分获取和增强干旱期水分存储能力实现生态位占领[40 ] .其次,糙隐子草作为多年生禾草,在极端降水处理中贡献度显著,但在适度降水变化中贡献度下降.这种双峰响应模式可能与其深层根系和休眠芽库策略相关:在极端湿润条件下,其分蘖能力增强[41 ] ;在极端干旱时,则通过地上部分生物量再分配维持生存[41 -42 ] .相比之下,画眉草多在D5处理中贡献度显著,表明其早春萌发特性可能使其在季节性干旱前期占据水分利用窗口期[10 ] .研究还发现,极端处理组间差异主要由多物种协同贡献(如极端干旱处理中狗尾草、蒺藜和糙隐子草联合贡献),而适度降水变化处理则依赖单一物种主导(如D5处理中画眉草贡献度较高),这表明降水扰动强度与群落响应模式间存在非线性关系:当降水偏离历史波动阈值(如±30%P)时,物种互作网络可能从补偿性响应转向竞争性替代[43 ] . ...
Characterizing the nonlinear dynamics of hydrological systems based on global recurrence analysis
1
2025
... 综合多维指标可见,优势种更替是植被结构响应降水变化的直观表现,指示种功能分异反映环境筛选的方向性,而生态位动态则揭示种间资源竞争本质.例如,降水减少60%处理下,糙隐子草(优势种)重要值骤降与其生态位宽度缩窄同步发生,而猪毛菜(指示种)通过快速占据生态位空缺成为新优势种,三者共同印证了资源适应能力下降→生态位释放→先锋种入侵的替代路径.多维指标协同分析,能更全面地揭示降水变化驱动植被结构调整的生态机制.广生态位物种或高重要值物种(如狗尾草、蒺藜、糙隐子草和画眉草)对降水处理组间差异的高贡献度特征(普遍P >0.05)提示其通过非显著性广谱参与在群落缓冲降水波动中发挥核心调控功能;而特殊生态位物种(如兴安胡枝子)的局部显著性贡献则表明狭生态位特化种可通过关键节点突破驱动差异格局.这表明旱区植被恢复应优先保育具有跨梯度适应力的缓冲物种,而非单纯依赖优势种[38 ] .首先,狗尾草作为一年生禾草,在极端降水处理中贡献度显著提升,其广域贡献度可能与其快速资源捕获策略相关[39 ] .尽管狗尾草在多个处理组间差异贡献度较高,但其显著性波动范围较大,这可能反映其作为“机会主义种”在不同处理中生态位宽度的动态调整[39 ] .与之类似,蒺藜作为一年生杂类草,可能通过抑制竞争者水分获取和增强干旱期水分存储能力实现生态位占领[40 ] .其次,糙隐子草作为多年生禾草,在极端降水处理中贡献度显著,但在适度降水变化中贡献度下降.这种双峰响应模式可能与其深层根系和休眠芽库策略相关:在极端湿润条件下,其分蘖能力增强[41 ] ;在极端干旱时,则通过地上部分生物量再分配维持生存[41 -42 ] .相比之下,画眉草多在D5处理中贡献度显著,表明其早春萌发特性可能使其在季节性干旱前期占据水分利用窗口期[10 ] .研究还发现,极端处理组间差异主要由多物种协同贡献(如极端干旱处理中狗尾草、蒺藜和糙隐子草联合贡献),而适度降水变化处理则依赖单一物种主导(如D5处理中画眉草贡献度较高),这表明降水扰动强度与群落响应模式间存在非线性关系:当降水偏离历史波动阈值(如±30%P)时,物种互作网络可能从补偿性响应转向竞争性替代[43 ] . ...




 甘公网安备 62010202000688号
甘公网安备 62010202000688号
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}