


0 引言
巴丹吉林沙漠位于中国内蒙古自治区阿拉善盟境内,地理上以雅布赖山以西、北大山以北、弱水以东、拐子湖以南及39°30′—42°00′N、98°30′—104°00′E作为划分依据,呈西北低东南高趋势,总面积达4.7万km2,是中国第三大沙漠[1],主要生境为沙漠与戈壁,生态环境脆弱。目前巴丹吉林沙漠仍面临着气候变化和流动沙丘的威胁,因此加强对当地的生态治理及监管尤为重要。
拟步甲昆虫作为沙漠环境中广布的地表甲虫,对沙漠生态系统重要性体现在维护当地生态系统的稳定性、研究当地的物种多样性和生态系统运作模式[2]、指示当地生态环境变化、反映该地区气候变化情况等方面,其中以物种多样性与环境因子关系研究较多。如张大治等[3]认为白芨滩拟步甲的物种多样性可指示破碎化生态环境治理及评估状况;任嘉隆等[4]通过气候与地表节肢动物聚集情况的探究,说明昆虫群落聚集状况反映了当地的气候变化。拟步甲群落时空分布的研究不仅能加深理解该生态系统的结构、功能,揭示生态因子影响其分布格局的机制,还能为评估生态系统健康状况、预测环境变化趋势提供宝贵的生物信息[5]。关于群落时空分布的研究有杨贵军等[6]对小尺度环境下的拟步甲生态位及种间关系进行研究,加深了对贺兰山生态系统的理解;汤新杰[7]研究北祁连山拟步甲分布状况和驱动要素揭示了生态因子对拟步甲分布格局的影响。此外,拟步甲昆虫作为指示性昆虫,其种群数量、种类组成及分布变化能够敏感地反映出当地生态环境的微小变动,为环境监测和生态保护提供重要依据[8-12]。
尽管地表甲虫的空间分布及其驱动要素近年来已受到学者们的广泛关注,但针对荒漠生态系统的研究仍然相对有限。本研究以栅格法对巴丹吉林沙漠边缘进行样地划分,采用主动采集的方法捕获拟步甲成虫并基于地理统计空间分析软件对巴丹吉林沙漠边缘拟步甲物种组成和分布状况进行研究,探究影响其分布状况的因素,为进一步研究巴丹吉林沙漠边缘生态保护提供相应的理论依据。
1 研究区概况及研究方法
1.1 研究区概况
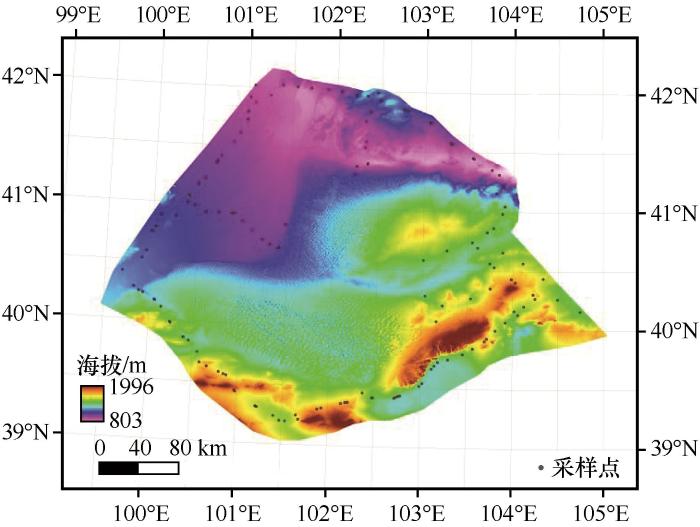
采样点涵盖巴丹吉林沙漠边缘除无人区以外地带及毗邻地带,涉及该地区所有生境,整体海拔800~2 000 m(图1)。巴丹吉林沙漠边缘地区属大陆性气候,年降水量50~60 mm,年均气温7~8 ℃,降水集中在5—7月,昼夜温差大。研究区处于风沙土带及灰棕漠土带,土壤类型可以分为风沙土、灰漠土、灰棕漠土、灰色草甸土和内陆盐土[13],流动沙丘地10~15 km沙山的砂粒度组成中,粗砂占0.33%~15.04%,中砂占6.36%~50.77%,细砂占21.6%~52.0%,极细砂占16.47%~38.67%,粉砂占0.33%~3.9%[14]。研究区内植被以白刺(Nitraria tangutorum)、梭梭(Haloxylon ammodendron)、柠条(Caragana korshinskii)、霸王(Zygophyllum xanthoxylum)等低矮灌木和骆驼蓬(Peganum harmala)、芨芨草(Neotrinia splendens)等部分草本及胡杨(Populus euphratica)、红柳(Tamarix ramosissima)为主。
图1
1.2 数据采集
按经度×纬度(30′×30′)对巴丹吉林沙漠边缘及周边地区划分栅格,每栅格设2个样点,样点面积为1 km2。分别于2023年7月11—17日、2024年5月6—12日和2024年7月24日至8月1日进行3次拟步甲成虫主动采集工作,调查每样点拟步甲成虫的种类和数量。①振落法:使用工具对灌木、低矮乔木枝条进行敲打,受到惊吓的拟步甲会掉落于地面上,此时使用徒手采集。②筛网法:沿植物根际周围用工具取枯落物和沙土放于筛网,多次抖动后取出拟步甲。③搜集法:搜索石下、植物根际、地表、沙面上活动的拟步甲。
1.3 数据分析
1.3.1 物种组成
对每样点拟步甲成虫的种类和数量进行统计,个体数占比大于等于10%者为优势种,个体数占比在1%~10%为常见种,个体数占比小于1%者为稀有种。
1.3.2 拟步甲空间分布格局
利用DEM高程数据对巴丹吉林沙漠地形进行区别,按巴丹吉林沙漠地图及其地表生态将巴丹吉林沙漠划分为戈壁和沙漠2种生境,并将采样点物种信息置于相应的生态环境中统计物种组成,计算Shannon多样性指数、Simpson指数、Pielou均匀度指数、Margalef丰富度指数等6项α多样性指数,并利用Past 4.1.7对不同生境下拟步甲群落的各项指数进行显著性检验。
采用普通克里金空间插值分析依据协方差函数对巴丹吉林沙漠有限区域内拟步甲物种数量和个体数量进行无偏最优估计。本研究采用地理统计分析工具ArcGIS对研究区内拟步甲群落种类数和物种数量进行普通克里金法空间插值分析预测,通过空间局部估计绘制巴丹吉林沙漠拟步甲科昆虫空间分布格局图,分析不同区域拟步甲群落空间格局分布特征。
1.3.3 影响群落分布的驱动要素
对所提取的19种生态环境因子数据通过主成分分析(Principal Components Analysis)进行数据富集,并进行旋转处理,筛选出特征值大于1的环境因子作为主要影响拟步甲群落分布的因子。
采用消除趋势对应分析(detrended correspondence analysis,DCA)确定排序轴的梯度长度(lengths of gradient),并依据LGA值的大小选择适宜的排序分析方法。由于本文DCA计算出的LGA(5.42)>3,故采用基于单峰型模型的典范对应分析(Canonical correspondence analysis,CCA)来确定拟步甲昆虫分布与驱动要素之间的关系。将2种地形数据和筛选后获得的环境数据进行对数转换,使其正态化,通过典范对应分析阐明巴丹吉林沙漠拟步甲群落分布和环境因子之间的关系。
主成分分析采用SPSS 27软件,典范对应分析采用Canoco 5.0软件。
2 结果与分析
2.1 物种组成
经鉴定及统计汇总,巴丹吉林沙漠边缘地区共计采集拟步甲成虫15 664只,隶属于2亚科5族17属70种(表1)。其中波氏东鳖甲(16.30%)、谢氏宽漠王(13.45%)、尖尾东鳖甲(10.32%)为该地区的优势种,克氏扁漠甲(7.59%)、拱背扁漠甲(7.34%)、纳氏东鳖甲(6.19%)、暗色扁漠甲(4.40%)、鄂漠王(3.65%)、库氏东鳖甲(3.49%)、小丽东鳖甲(3.38%)、粗壮真土甲(2.56%)、莱氏脊漠甲(2.21%)、磨光东鳖甲(1.78%)、紫奇扁漠甲(1.71%)、克蒙小鳖甲(1.59%)、凹缝东鳖甲(1.53%)、居维扁漠甲(1.19%)、无边东鳖甲(1.07%)为该地区的常见种。
表1 巴丹吉林沙漠边缘拟步甲群落物种组成
Table 1
| 族 | 属 | 种 | 沙漠+戈壁 | 沙漠环境 | 戈壁环境 | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| 数量 | 百分比/%a | 数量 | 百分比/%b | 数量 | 百分比/%c | |||||
| 砚甲族(Akidini) | 砚甲属(Cyphogenia) | 中华砚甲(C. chinensis) | 16 | 0.10 | 8 | 0.19 | 8 | 0.07 | ||
| 漠甲族(Pimeliini) | 漠王属(Platyope) | 鄂漠王(P. ordossica) | 572 | 3.65 | 146 | 3.42 | 426 | 3.74 | ||
| 蒙古漠王(P. mongolica) | 19 | 0.12 | 0 | 0.00 | 19 | 0.17 | ||||
| 大瘤漠王(P. granulate) | 6 | 0.04 | 0 | 0.00 | 6 | 0.05 | ||||
| 角漠甲属(Trigonocnera) | 突角漠甲(T. pseudopimelia pseudopimelia) | 3 | 0.02 | 1 | 0.02 | 2 | 0.02 | |||
| 宽漠王属(Mantichorula) | 谢氏宽漠王(M. semenowi) | 2 107 | 13.45 | 707 | 16.55 | 1 400 | 12.29 | |||
| 宽漠王(M. grandis) | 20 | 0.13 | 17 | 0.40 | 3 | 0.03 | ||||
| 胖漠甲属(Trigonoscelis) | 光滑胖漠甲(T. (C.) sublaevigata sublaevigata) | 2 | 0.01 | 0 | 0.00 | 2 | 0.02 | |||
| 脊漠甲属(Pterocoma) | 洛氏脊漠甲(P. (M.) loczyi) | 118 | 0.75 | 31 | 0.73 | 87 | 0.76 | |||
| 莱氏脊漠甲(P. (M.) reitteri) | 346 | 2.21 | 39 | 0.91 | 307 | 2.69 | ||||
| 埃氏脊漠甲(P. (M.) amandana edmundi) | 57 | 0.36 | 6 | 0.14 | 51 | 0.45 | ||||
| 扁漠甲属(Sternotrigon) | 白毛扁漠甲(S. albipulis) | 6 | 0.04 | 0 | 0.00 | 6 | 0.05 | |||
| 拱背扁漠甲(S. grandis) | 1 149 | 7.34 | 327 | 7.66 | 822 | 7.21 | ||||
| 紫奇扁漠甲(S. zichyi) | 268 | 1.71 | 47 | 1.10 | 221 | 1.94 | ||||
| 克氏扁漠甲(S. kraatzi) | 1 189 | 7.59 | 162 | 3.79 | 1 027 | 9.01 | ||||
| 暗色扁漠甲(S. opaea) | 689 | 4.40 | 154 | 3.61 | 535 | 4.70 | ||||
| 多毛扁漠甲(S. setosa setosa) | 144 | 0.92 | 36 | 0.84 | 108 | 0.95 | ||||
| 居维扁漠甲(S. setosa juvencus) | 186 | 1.19 | 4 | 0.09 | 182 | 1.60 | ||||
| 宽漠甲属(Sternoplax) | 磨光宽漠甲(S.opaca) | 6 | 0.04 | 0 | 0.00 | 6 | 0.05 | |||
| 鳖甲族(Tentyriini) | 东鳖甲属(Anatolica) | 伪东鳖甲(A. pseudiduna) | 132 | 0.84 | 0 | 0.00 | 132 | 1.16 | ||
| 凹缝东鳖甲(A. suturalis) | 240 | 1.53 | 90 | 2.11 | 150 | 1.32 | ||||
| 波氏东鳖甲(A. potanini) | 2 554 | 16.30 | 1 140 | 26.69 | 1 414 | 12.41 | ||||
| 奇异东鳖甲(A. paradoxa) | 103 | 0.66 | 10 | 0.23 | 93 | 0.82 | ||||
| 平颊东鳖甲(A. dashidorzsi temporalis) | 20 | 0.13 | 8 | 0.19 | 12 | 0.11 | ||||
| 尖尾东鳖甲(A. mucronata) | 1 617 | 10.32 | 383 | 8.97 | 1 234 | 10.83 | ||||
| 小东鳖甲(A. minima) | 2 | 0.01 | 2 | 0.05 | 0 | 0.00 | ||||
| 无边东鳖甲(A. immarginata) | 167 | 1.07 | 5 | 0.12 | 162 | 1.42 | ||||
| 小丽东鳖甲(A. amoenula) | 530 | 3.38 | 162 | 3.79 | 368 | 3.23 | ||||
| 库氏东鳖甲(A. kulzeri) | 546 | 3.49 | 110 | 2.58 | 436 | 3.83 | ||||
| 纳氏东鳖甲(A. nureti) | 969 | 6.19 | 142 | 3.32 | 827 | 7.26 | ||||
| 全边东鳖甲(A. integra) | 9 | 0.06 | 1 | 0.02 | 8 | 0.07 | ||||
| 纵凹东鳖甲(A. externecostata) | 94 | 0.60 | 12 | 0.28 | 82 | 0.72 | ||||
| 宽突东鳖甲(A. sternalis) | 52 | 0.33 | 39 | 0.91 | 13 | 0.11 | ||||
| 磨光东鳖甲(A. polita polita) | 279 | 1.78 | 221 | 5.17 | 58 | 0.51 | ||||
| 宽腹东鳖甲(A. gravidula) | 2 | 0.01 | 0 | 0.00 | 2 | 0.02 | ||||
| 突颊东鳖甲(A. tsendsureni) | 6 | 0.04 | 0 | 0.00 | 6 | 0.05 | ||||
| 鳖甲族(Tentyriini) | 小鳖甲属(Microdera) | 山丹小鳖甲(M. (M.) shandanana) | 109 | 0.70 | 2 | 0.05 | 107 | 0.94 | ||
| 克蒙小鳖甲(M. (M.) mongolica kozlovi) | 249 | 1.59 | 32 | 0.75 | 217 | 1.90 | ||||
| 宽颈小鳖甲(M. (M.) laticollis) | 90 | 0.57 | 5 | 0.12 | 85 | 0.75 | ||||
| 球胸小鳖甲(M. (D.) globata) | 22 | 0.14 | 0 | 0.00 | 22 | 0.19 | ||||
| 阿小鳖甲(M. (D.) kraatzi alashanica) | 9 | 0.06 | 0 | 0.00 | 9 | 0.08 | ||||
| 蒙古小鳖甲(M. (M.) mongolica mongolica) | 4 | 0.03 | 0 | 0.00 | 4 | 0.04 | ||||
| 克小鳖甲(M. (D.) kraatzi kraatzi) | 2 | 0.01 | 1 | 0.02 | 1 | 0.01 | ||||
| 姬小鳖甲(M. (D.) elegans) | 35 | 0.22 | 0 | 0.00 | 35 | 0.31 | ||||
| 胸鳖甲属(Colposcelis) | 三沟胸鳖甲(C. (S.) trisulcata) | 7 | 0.04 | 6 | 0.14 | 1 | 0.01 | |||
| 隆胸鳖甲(C. (S.) montivaga) | 16 | 0.10 | 0 | 0.00 | 16 | 0.14 | ||||
| 福氏胸鳖甲(C. (S.) forsteri) | 3 | 0.02 | 0 | 0.00 | 3 | 0.03 | ||||
| 琵甲族(Blaptini) | 琵甲属(Blaps) | 异距琵甲(B. (B.) kiritshenkoi) | 13 | 0.08 | 9 | 0.21 | 4 | 0.04 | ||
| 叉尾琵甲(B. (B.) furcala) | 7 | 0.04 | 0 | 0.00 | 7 | 0.06 | ||||
| 戈壁琵甲(B. (B.) gobiensis) | 4 | 0.03 | 0 | 0.00 | 4 | 0.04 | ||||
| 皱纹琵甲(B. (B.) rugosa) | 17 | 0.11 | 2 | 0.05 | 15 | 0.13 | ||||
| 拟步行琵甲(B. (B.) caraboides caraboides) | 13 | 0.08 | 9 | 0.21 | 4 | 0.04 | ||||
| 弯齿琵甲(B. femoralis) | 31 | 0.20 | 0 | 0.00 | 31 | 0.27 | ||||
| 达氏琵甲(B. (B.) davidis) | 1 | 0.01 | 0 | 0.00 | 12 | 0.11 | ||||
| 土甲族(Opatrini) | 真土甲属(Eumylada) | 粗壮真土甲(E. glandulosa) | 401 | 2.56 | 94 | 2.20 | 307 | 2.69 | ||
| 同点真土甲(E. punctifera) | 59 | 0.38 | 29 | 0.68 | 30 | 0.26 | ||||
| 奥氏真土甲(E. oberbergeri) | 14 | 0.09 | 0 | 0.00 | 14 | 0.12 | ||||
| 鄂真土甲(E.ordosana) | 64 | 0.41 | 33 | 0.77 | 31 | 0.27 | ||||
| 伪坚土甲属(Scleropatrum) | 粗背伪坚土甲(S. horridum horridum) | 3 | 0.02 | 0 | 0.00 | 3 | 0.03 | |||
| 瘤翅伪坚土甲(S. tuberculatum) | 1 | 0.01 | 0 | 0.00 | 1 | 0.01 | ||||
| 笨土甲属(Penthicus) | 阿笨土甲(P. (M.) alashanicus) | 2 | 0.01 | 0 | 0.00 | 2 | 0.02 | |||
| 钝突笨土甲(P. (M.) nojonicus) | 14 | 0.09 | 0 | 0.00 | 14 | 0.12 | ||||
| 福笨土甲(P. (M.) frater) | 1 | 0.01 | 1 | 0.02 | 0 | 0.00 | ||||
| 钝角笨土甲(P. (M.) obtusangulus) | 1 | 0.01 | 0 | 0.00 | 1 | 0.01 | ||||
| 阿土甲属(Anatrum) | 山丹阿土甲(A. shandanicum) | 65 | 0.41 | 0 | 0.00 | 65 | 0.57 | |||
| 漠土甲属(Melanesthes) | 蒙古漠土甲(M. (M.) mongolica) | 2 | 0.01 | 1 | 0.02 | 36 | 0.32 | |||
| 景泰漠土甲(M. (M.) jintaiensis) | 129 | 0.82 | 35 | 0.82 | 94 | 0.83 | ||||
| 达氏漠土甲(M. (M.) davadshamsi) | 12 | 0.08 | 1 | 0.02 | 0 | 0.00 | ||||
| 纤毛漠土甲(M. (M.) ciliata) | 2 | 0.01 | 0 | 0.00 | 2 | 0.02 | ||||
| 蒙南漠土甲(M. (M.) jenseni meridionalis) | 37 | 0.24 | 1 | 0.02 | 1 | 0.01 | ||||
| 合计 | 15 664 | 100.00 | 4 271 | 100.00 | 11 393 | 100.00 | ||||
2.2 拟步甲族、属组成
由表2可知,该地区拟步甲在族、属分类阶元上存在明显差异。在族级阶元上,巴丹吉林沙漠以漠甲族(41.18%)和土甲族(29.41%)为主。在属级阶元上以扁漠甲属(10.00%)、东鳖甲属(24.29%)、小鳖甲属(11.43%)以及琵甲属(10.00%)为主。由此可见,从属级阶元上来说巴丹吉林沙漠边缘拟步甲群落以鳖甲族为主要构成类群,漠甲族为次要构成类群。
表2 巴丹吉林沙漠边缘拟步甲群落族、属数量及百分比
Table 2
| 族 | 数量 | 百分比/%a | 属 | 数量 | 百分比/%b |
|---|---|---|---|---|---|
| 砚甲族 | 1 | 5.88 | 砚甲属 | 1 | 1.43 |
| 漠甲族 | 7 | 41.18 | 漠王属 | 3 | 4.29 |
| 角漠甲属 | 1 | 1.43 | |||
| 宽漠王属 | 2 | 2.86 | |||
| 胖漠甲属 | 1 | 1.43 | |||
| 脊漠甲属 | 3 | 4.29 | |||
| 扁漠甲属 | 7 | 10.00 | |||
| 宽漠甲属 | 1 | 1.43 | |||
| 鳖甲族 | 3 | 17.65 | 东鳖甲属 | 17 | 24.29 |
| 小鳖甲属 | 8 | 11.43 | |||
| 胸鳖甲属 | 3 | 4.29 | |||
| 琵甲族 | 1 | 5.88 | 琵甲属 | 7 | 10.00 |
| 土甲族 | 5 | 29.41 | 真土甲属 | 4 | 5.71 |
| 伪坚土甲属 | 2 | 2.86 | |||
| 笨土甲属 | 4 | 5.71 | |||
| 阿土甲属 | 1 | 1.43 | |||
| 漠土甲属 | 5 | 7.14 | |||
| 共计 | 17 | 70 | 100.00 |
2.3 物种丰度变化与地域差异
对表1生境中的拟步甲群落构成分析可知,戈壁环境中的拟步甲群落丰度最高,沙漠环境拟步甲群落丰度最低。其中沙漠生境中波氏东鳖甲、谢氏宽漠王2种拟步甲占据绝对优势地位;戈壁环境中以谢氏宽漠王、波氏东鳖甲、克氏扁漠甲、拱背扁漠甲、尖尾东鳖甲占绝对优势地位。克氏扁漠甲、拱背扁漠甲等扁漠甲属拟步甲昆虫和尖尾东鳖甲、纳氏东鳖甲等东鳖甲属拟步甲昆虫在不同生境中均有分布,说明扁漠甲属和东鳖甲属拟步甲昆虫具有更强的环境适应性。
2.4 空间分布格局
2.4.1 物种多样性
表3表明Simpson指数、Shannon指数、PieLou均匀度指数均为戈壁环境>沙漠环境,说明戈壁环境中拟步甲群落的物种多样性和分布的均匀程度最高,沙漠环境最差。由于这3项指数受个体数量影响较大,且相互影响,因此选用Margalef丰富度指数、Fisher-alpha指数及Berger-Parker指数进行进一步分析。Margalef丰富度指数结果表明物种多样性戈壁环境>沙漠环境。Berger-Parker指数表明优势种在戈壁生境中的生态地位突出。Fisher-alpha指数结果表明戈壁环境的物种多样性和均匀程度优于沙漠环境。整体结果表明戈壁环境拟步甲的群落构成更加丰富、稳定,沙漠环境拟步甲群落分布更为散乱。
表3 巴丹吉林沙漠边缘3种生境拟步甲科昆虫α多样性指数
Table 3
| 沙漠环境 | 戈壁环境 | |
|---|---|---|
| Simpson_1-D | 0.8285±0.0113a | 0.8368±0.1231b |
| Shannon_H | 2.138±0.1719a | 2.4327±0.7135b |
| Evenness_e^H/S | 0.4036±0.1586a | 0.363±0.0598a |
| Margalef | 3.1843±1.1336a | 4.2557±2.4904b |
| Fisher-alpha | 4.176±1.5313a | 5.7267±3.8681b |
| Berger-Parker | 0.3059±0.0565a | 0.3116±0.1698b |
2.4.2 普通克里金空间插值
普通克里金空间插值表明,巴丹吉林沙漠边缘拟步甲群落多集中分布于西北及南部区域中,形成两个重要的核心分布区域。整体上看巴丹吉林沙漠边缘拟步甲群落多分布于边缘区域,物种数量及个体数量由边缘区域向核心区域逐渐减少,北部区域内拟步甲群落的分布状况较为稀疏。
如图2所示,该地区拟步甲群落物种数量呈明显聚集分布,东北、西南、东南地区为3个主要聚集区。高密度与低密度区域镶嵌分布,中心区域出现间断不规则低密度斑块,西部、南部、东部均有少量低密度斑块镶嵌其中。从整体看,拟步甲群落物种数量分布呈边缘向内依次递减的趋势,西侧拟步甲物种数量高于东侧。
图2
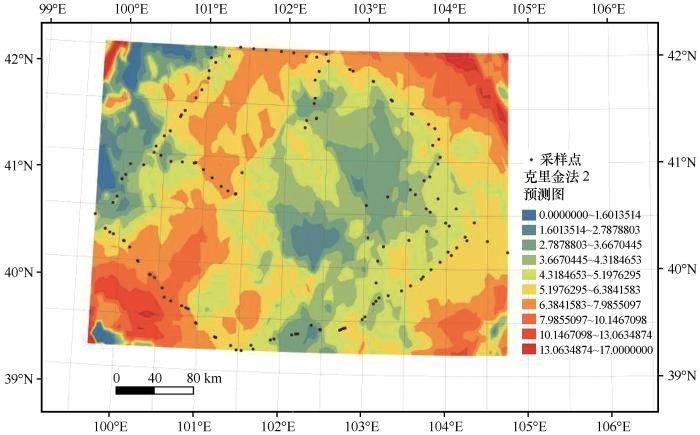
图2
巴丹吉林沙漠边缘物种数量分布预测
Fig.2
Predicted distribution of the number of species in Badain Jaran Desert margin
个体数量整体由南向北依次递减,北部和中部核心地区有低密度斑块出现,西北侧以及东南侧区域为拟步甲数量分布高密度区域(图3)。巴丹吉林沙漠边缘拟步甲群落个体数量为20~200只,部分生境中的拟步甲数量在气温、湿度等适宜条件下可达500头以上。除北部地区外,沙漠边缘地区拟步甲
图3
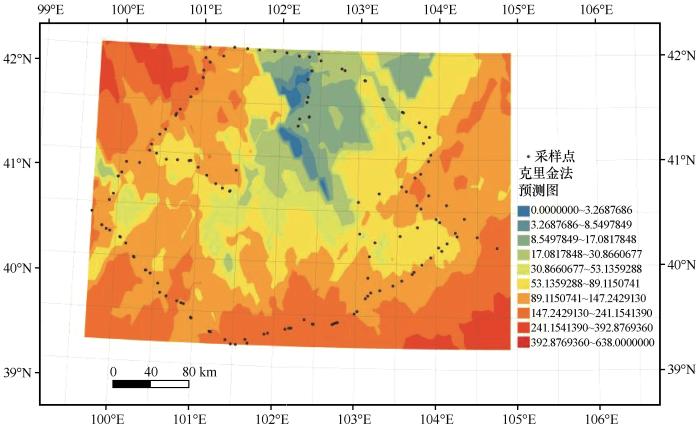
图3
巴丹吉林沙漠边缘个体数量分布预测
Fig.3
Predicted distribution of species populations in Badain Jaran Desert margin
数量远高于沙漠核心地区。
2.5 群落分布与环境因子的关系
主成分分析结果见表4,前4个特征值大于1的环境因子所代表的原始信息量依次为64.37%、12.36%、11.87%和5.32%,共反映总信息量的93.92%,说明年平均气温、最冷月份的最低气温、年气温范围和最潮湿季度的平均气温是影响巴丹吉林沙漠边缘拟步甲物种多样性分布格局的主要因子。
表4 巴丹吉林沙漠边缘拟步甲多样性分布格局影响因素的主成分分析
Table 4
| 成分 | 初始特征值 | 提取载荷平方和 | 旋转载荷平方和 | ||||||
|---|---|---|---|---|---|---|---|---|---|
| 总计 | 方差/% | 累积/% | 总计 | 方差/% | 累积/% | 总计 | 方差/% | 累积/% | |
| 年气温范围 | 12.23 | 64.37 | 64.37 | 12.23 | 64.37 | 64.37 | 11.19 | 58.88 | 58.88 |
| 年平均气温 | 2.35 | 12.36 | 76.73 | 2.35 | 12.36 | 76.73 | 2.54 | 13.38 | 72.26 |
| 最冷月份的最低气温 | 2.25 | 11.87 | 88.59 | 2.25 | 11.87 | 88.59 | 2.21 | 11.61 | 83.86 |
| 最潮湿季度的平均气温 | 1.01 | 5.32 | 93.92 | 1.01 | 5.32 | 93.92 | 1.91 | 10.06 | 93.92 |
| 年降水量 | 0.71 | 3.72 | 97.63 | ||||||
| 最潮湿月份的降水量 | 0.31 | 1.63 | 99.27 | ||||||
| 最干燥季度的平均气温 | 0.07 | 0.35 | 99.61 | ||||||
| 最热季度的平均气温 | 0.04 | 0.19 | 99.80 | ||||||
| 最热季度的平均气温 | 0.02 | 0.10 | 99.90 | ||||||
| 平均日潮差 | 0.01 | 0.07 | 99.96 | ||||||
| 等温性 | 0.00 | 0.01 | 99.98 | ||||||
| 气温季节性 | 0.00 | 0.01 | 99.99 | ||||||
| 最热月份的最高气温 | 0.00 | 0.01 | 99.99 | ||||||
| 最干燥月份的降水量 | 0.00 | 0.00 | 100.00 | ||||||
| 降水季节性 | 0.00 | 0.00 | 100.00 | ||||||
| 最湿季降水量 | 0.00 | 0.00 | 100.00 | ||||||
| 最干燥季度的降水量 | 0.00 | 0.00 | 100.00 | ||||||
| 最暖季度的降水量 | 0.00 | 0.00 | 100.00 | ||||||
| 最冷季度的降水量 | 0.00 | 0.00 | 100.00 | ||||||
巴丹吉林沙漠边缘拟步甲分布状况与影响因子排序结果见表5和图4。前4个排序轴的特征值分别为0.24、0.15、0.11、0.10,物种-环境关系方差累计贡献率为77.07%,其中前两轴累计贡献率为49.99%,结果表明前两轴能较好地反映出研究区内拟步甲科昆虫分布与环境因子的关系。年气温范围对巴丹吉林沙漠边缘拟步甲的分布状况解释值最大,其次分别是年平均气温、最冷月份的最低气温,海拔、坡位、坡向3种地形因子对拟步甲群落分布影响较小。从物种分布情况来看,叉尾琵甲与粗背伪坚土甲(sp66,sp69)的分布状况受海拔因素影响较大,与其他影响因子相关性较小。突颊东鳖甲和光滑胖漠甲(sp46,sp47)受年气温范围影响较大,居维扁漠甲与宽腹东鳖甲受年平均气温影响较大,其余物种分布状况受多个因素影响。
表5 影响巴丹吉林沙漠边缘拟步甲分布状况因子的CCA 排序分析参数
Table 5
| 参数 | Axis 1 | Axis 2 | Axis 3 | Axis 4 |
|---|---|---|---|---|
| 特征值 | 0.24 | 0.15 | 0.11 | 0.10 |
| 累积解释变异 | 2.11 | 3.48 | 4.46 | 5.36 |
| 伪典型相关 | 0.68 | 0.62 | 0.56 | 0.55 |
| 累积解释拟合变异 | 30.26 | 49.99 | 64.08 | 77.07 |
图4
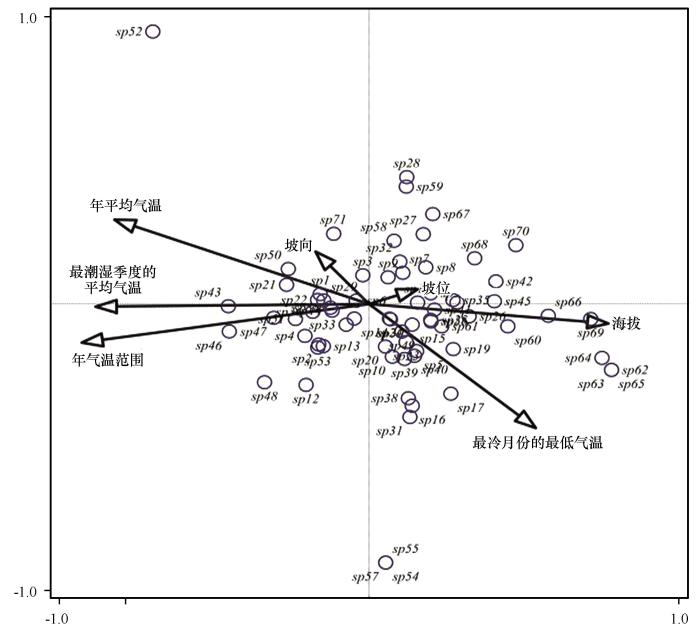
图4
巴丹吉林沙漠边缘拟步甲分布状况与生境因子的排序
Fig.4
Distribution status and ranking of habitat factors of the proposed steppe beetle in Badain Jaran Desert margin
3 讨论
3次采集共涉及拟步甲2亚科5族17属70种,其中波氏东鳖甲,尖尾东鳖甲和谢氏宽漠王为该地区的优势种。与孙晓杰[19]结果相比,本文所涉及的种类与其基本一致,表明拟步甲群落的分布状况受其生存环境限制,特有种分布较为局限,不同物种在地理分布状况与自然气候和地理环境等环境因素呈现一致性。他富源等[24]和徐志昭[25]发现鳖甲属、土甲属及漠甲属均为其研究区拟步甲群落的主要构成类群,且优势种均隶属于上述属,与本文的研究结果一致,说明了拟步甲群落组成对荒漠环境的选择具有趋向性。不同生境群落组成和多样性差异可能与不同属种拟步甲昆虫的生理生化活动、周期性活动节律以及分布地生态环境等有关[26-28]。在不同生境共有种中,波氏东鳖甲在沙漠生境拟步甲群落占比远高于戈壁生境,说明当生境向沙漠环境变迁时,波氏东鳖甲在该生境拟步甲物种构成占比会增加,与刘新民等[29]的研究结果一致;克氏扁漠甲等扁漠甲属及纳氏东鳖甲、库氏东鳖甲等部分东鳖甲属在戈壁环境拟步甲群落中占比更高,表明该类拟步甲昆虫具有更强的环境适应性。琵甲属、土甲属拟步甲昆虫在戈壁环境中的物种数量占比更高,说明该类拟步甲昆虫更适合生存于环境丰富的生境。
α多样性指数整体结果表明戈壁环境拟步甲的群落构成更加丰富、稳定,沙漠环境拟步甲群落分布更为散乱,且优势种在种群构成方面占突出地位,说明生境丰富的程度对拟步甲群落的结构和功能的多样性具有支持作用,这与贾雨桥等[30]的研究结果一致。普通克里金空间插值结果表明,巴丹吉林沙漠边缘西侧区域拟步甲物种数0~5,说明该地区生态环境较差,无法支持更大规模的拟步甲群落生存,与贺达汉等[31]的研究结果一致。西侧鼎新镇、巴音陶海苏木及南部阿拉善右旗、雅布赖镇为城镇范围,其范围内拟步甲种类数目较少,说明人为活动因素对拟步甲分布影响极大,与何卫强[32]研究结果一致。东北、西南地区为山脉地区,该区域内生境复杂程度高且物种数目占已鉴定结果的86%,说明层次分明、复杂多样的生态环境中拟步甲物种数量会增多,与王怡等[33]的研究结果一致。中部沙漠腹地拟步甲个体数量少于周边沙漠地区,或是由于沙漠核心地区昼夜温差大,气温变化强烈,空气对流现象严重,近地面风速较大,沙丘表面沙土易被扰动,进而影响拟步甲昆虫的正常活动。另外,该区域内日照时间相对较长,水分蒸发量大和地表植被稀少且单一,使得拟步甲昆虫适宜生活环境受到限制。巴丹吉林沙漠边缘西侧拟步甲物种数量高于东侧,形成此现象的原因可能是西侧生态保护区、生态景区等高植被盖度环境多于东侧,生态环境的多样性提高了拟步甲群落的物种丰富度,与他人研究结果一致[34-36]。北部区域的拟步甲物种数量和个体数量最少,造成此现象出现的原因是,北部区域戈壁破碎环境及京新高速、丹阿线两条重要运输路线等人为因素严重影响了拟步甲在此生境中的数量。
典范对应分析结果表明拟步甲群落昆虫分布状况与影响因子之间的相关性能较好地对应,排序图显示气温对拟步甲昆虫分布影响较大,与他人研究结果一致[37-39]。年气温范围、年平均气温与海拔、坡位、坡向明显负相关,说明该地区地形因子是通过降低地区气温从而改变拟步甲群落构成和数量,与他人研究结果一致[40-41]。最冷月份的最低气温与海拔、坡位、坡向正相关,也从侧面印证了这一点。由于巴丹吉林沙漠边缘地理环境限制,风向呈西北至东南,风积沙方向较为一致且海拔梯度变化较小,太阳辐射强度较均匀,植被生长状况一致,因此该地区环境具有一定的稳定性和缓冲外界生态因子变化的能力,进而对于海拔、坡位、坡向这3类地形因子的变化不敏感,这可能也是导致巴丹吉林沙漠边缘拟步甲群落分布和地形因子之间的相关性分析未达到显著性水平的原因[42-44]。此外,本研究发现巴丹吉林沙漠边缘拟步甲昆虫分布具有明显的空间分布特征,且拟步甲群落物种数量和个体数量分布状况受气温变化影响显著。
4 结论
巴丹吉林沙漠边缘拟步甲群落集中分布于西北及南部区域,形成两个核心分布区,且多分布于沙漠边缘区域,物种数量及个体数量由边缘向核心逐渐减少,北部区域内群落分布较稀疏。年气温范围对巴丹吉林沙漠边缘拟步甲的分布状况影响最大。
参考文献
Trophic structure and the role of predation in shaping hot desert communities
[J].
Experimental effects of habitat fragmentation on old-field canopy insects:community,guild and species responses
[J].
Effects of landscape structure and habitat type on a plant-herbivore-parasitoid community
[J].
沙坡头地区人工固沙植被演替中大型土壤动物生物指示作用研究
[J].
小兴安岭阔叶红松林昆虫群落时空格局及其对气温的响应
[D].
帽儿山红松人工林鞘翅目成虫群落小尺度空间异质性变化特征
[J].
中国漠甲亚科分类与地理分布(鞘翅目:拟步甲科)
[D].
World Clim 2:new 1km spatial resolution climate surfaces for global land areas
[J].
全球变化对土壤动物多样性的影响
[J].
古尔班通古特沙漠梭梭林象甲科昆虫分布及其动态研究
[J].
Spatiotemporal variations of tenebrionid beetles (Coleoptera:Tenebrionidae) in the gobi desert.Northwest China
[J].
地形微生境对区域温度变化的缓冲作用
[J].
Adaptive trends of darkling beetles (Col.Tenebrionidae) on environmental gradients on the island of Tenerife (Canary Islands)
[J].
流动沙地灌丛内外生境中土壤动物群落结构研究
[J].
Factors influencing microhabitat partitioning in arid-land darkling beetles (Tenebrionidae):temperature and water conservation
[J].
Small-scale habitat use and assemblage structure of ground-dwelling beetles in a Patagonian shrub steppe
[J].
Effects of habitat complexity on the abundance,species richness and size of darkling beetles (Tenebrionidae) in artificial vegetation
[J].
Seasonal preference of darkling beetles (Tenebrionidae) for shrub vegetation due to high temperatures,not predation or food availability
[J].
Factors that affect the assembly of ground-dwelling beetles at small scales in primary mixed broadleaved-Korean pine forests in North-east China
[J].

 甘公网安备 62010202000688号
甘公网安备 62010202000688号
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}