Climate Change 2023:Synthesis Report.Contribution of Working Groups I,II and III to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change
2
2023
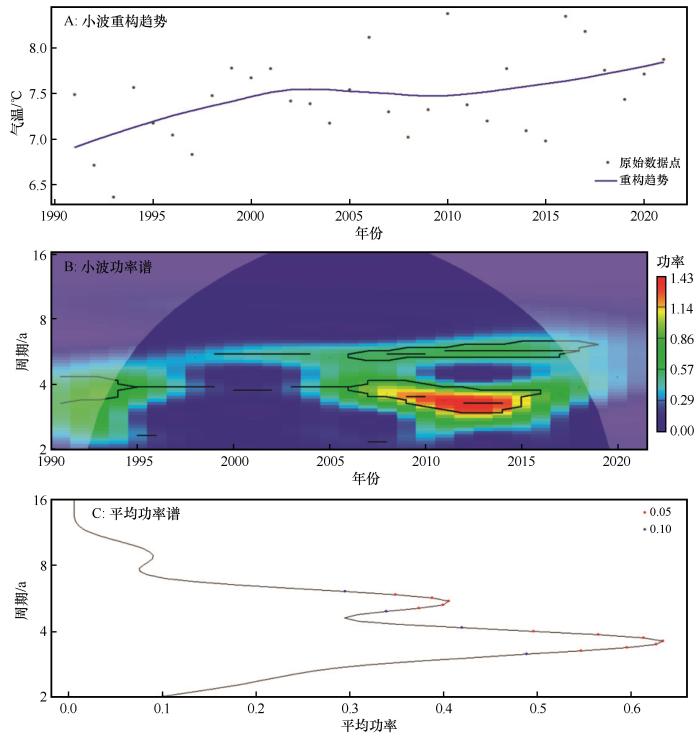
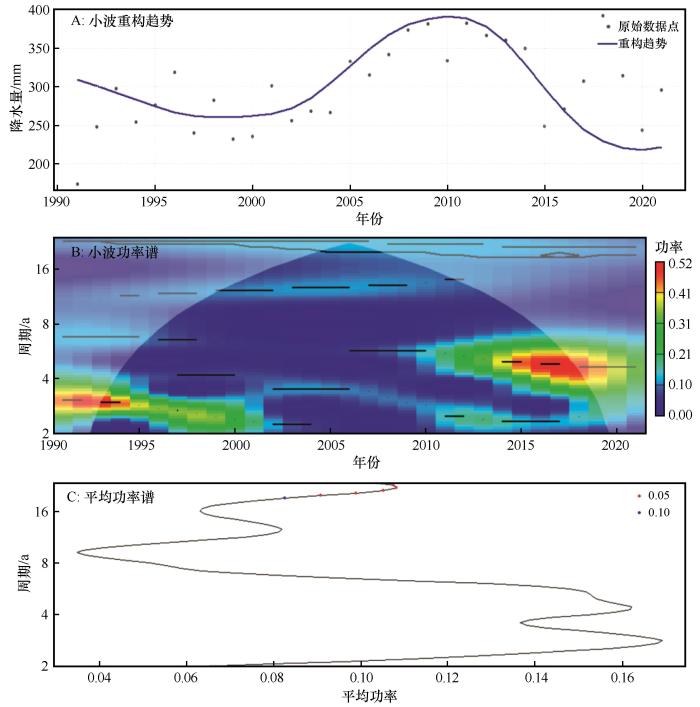
... 2011—2020年全球表面温度较1850—1900年平均水平上升约1.1 ℃,全球变暖趋势日益加剧[1].气温每升高1 ℃,饱和水汽压增加约7%,大气水汽含量随之上升,从而导致全球极端降水事件的频率、强度和持续时间普遍增强[2].在中国西北干旱区,1961—2020年气温以约0.33 ℃/10a的速率持续升高;自2000年以来,降水以29 mm/10a的速率显著增加,呈现明显的暖湿化特征[3].持续的气候扰动显著抑制植被的光合效率[4-5],削弱生态系统生产力并改变生态系统结构,致使生态系统稳定性的不确定性提高[1],从而对生态系统的服务功能、区域的生态环境和社会经济的可持续发展构成系统性挑战.因此,探究气候变化对生态系统稳定性的影响及其驱动机制,对提升生态系统应对气候变化的能力,保障生态环境和社会经济可持续发展具有重要意义. ...
... [1],从而对生态系统的服务功能、区域的生态环境和社会经济的可持续发展构成系统性挑战.因此,探究气候变化对生态系统稳定性的影响及其驱动机制,对提升生态系统应对气候变化的能力,保障生态环境和社会经济可持续发展具有重要意义. ...
Emergent constraints on future extreme precipitation intensification:from global to continental scales
1
2023
... 2011—2020年全球表面温度较1850—1900年平均水平上升约1.1 ℃,全球变暖趋势日益加剧[1].气温每升高1 ℃,饱和水汽压增加约7%,大气水汽含量随之上升,从而导致全球极端降水事件的频率、强度和持续时间普遍增强[2].在中国西北干旱区,1961—2020年气温以约0.33 ℃/10a的速率持续升高;自2000年以来,降水以29 mm/10a的速率显著增加,呈现明显的暖湿化特征[3].持续的气候扰动显著抑制植被的光合效率[4-5],削弱生态系统生产力并改变生态系统结构,致使生态系统稳定性的不确定性提高[1],从而对生态系统的服务功能、区域的生态环境和社会经济的可持续发展构成系统性挑战.因此,探究气候变化对生态系统稳定性的影响及其驱动机制,对提升生态系统应对气候变化的能力,保障生态环境和社会经济可持续发展具有重要意义. ...
The eastward expansion of the climate humidification trend in Northwest China and the synergistic influences on the circulation mechanism
1
2022
... 2011—2020年全球表面温度较1850—1900年平均水平上升约1.1 ℃,全球变暖趋势日益加剧[1].气温每升高1 ℃,饱和水汽压增加约7%,大气水汽含量随之上升,从而导致全球极端降水事件的频率、强度和持续时间普遍增强[2].在中国西北干旱区,1961—2020年气温以约0.33 ℃/10a的速率持续升高;自2000年以来,降水以29 mm/10a的速率显著增加,呈现明显的暖湿化特征[3].持续的气候扰动显著抑制植被的光合效率[4-5],削弱生态系统生产力并改变生态系统结构,致使生态系统稳定性的不确定性提高[1],从而对生态系统的服务功能、区域的生态环境和社会经济的可持续发展构成系统性挑战.因此,探究气候变化对生态系统稳定性的影响及其驱动机制,对提升生态系统应对气候变化的能力,保障生态环境和社会经济可持续发展具有重要意义. ...
Climate changes and photosynthesis
2
2016
... 2011—2020年全球表面温度较1850—1900年平均水平上升约1.1 ℃,全球变暖趋势日益加剧[1].气温每升高1 ℃,饱和水汽压增加约7%,大气水汽含量随之上升,从而导致全球极端降水事件的频率、强度和持续时间普遍增强[2].在中国西北干旱区,1961—2020年气温以约0.33 ℃/10a的速率持续升高;自2000年以来,降水以29 mm/10a的速率显著增加,呈现明显的暖湿化特征[3].持续的气候扰动显著抑制植被的光合效率[4-5],削弱生态系统生产力并改变生态系统结构,致使生态系统稳定性的不确定性提高[1],从而对生态系统的服务功能、区域的生态环境和社会经济的可持续发展构成系统性挑战.因此,探究气候变化对生态系统稳定性的影响及其驱动机制,对提升生态系统应对气候变化的能力,保障生态环境和社会经济可持续发展具有重要意义. ...
... 在研究时段内,回归结果显示气候变暖促进了乔木林生态系统稳定性的提升.而气温升高对生态系统稳定性的正效应可能与光合作用增强和生长季延长有关.温度上升在一定范围内可以提高光合作用效率和生产力,进而提升森林的抵抗力和恢复力[52].Wang等[53]利用北美洲的森林永久样地的研究同样证明,气温升高能够促进树木的生长,提高森林的生产力,从而增强生态系统稳定性,但白云杉(Picea glauca)、山杨(Populus tremuloides)、杰克松(Pinus banksiana)的生产力在夏季最高温分别到达23.4、27.0、26.2 ℃时开始下降,说明升温对生态系统稳定性的影响温度存在阈值.过度升温可能导致干旱胁迫和呼吸消耗增加,从而削弱稳定性[4].Ma等[54]在青藏高原高寒草地的试验中发现,气候变暖显著降低了该地区生态系统的时间稳定性.Zhao等[55]通过评估温度升高背景下的19个浮游生物食物网的稳定性,发现温度升高会降低其生态稳定性,这意味着生态系统稳定性在不同群落中对气温变化的响应可能存在差异.本研究中气温变化对生态系统稳定性的影响为“正效应”,这可能是气温变化仍处于乔木林生态系统可承受温度的适宜区间,而未来持续升温是否对生态系统稳定性造成负面影响还需进一步研究. ...
Northern ecosystem productivity reduced by Rossby-wave-driven hot-dry conditions
1
2025
... 2011—2020年全球表面温度较1850—1900年平均水平上升约1.1 ℃,全球变暖趋势日益加剧[1].气温每升高1 ℃,饱和水汽压增加约7%,大气水汽含量随之上升,从而导致全球极端降水事件的频率、强度和持续时间普遍增强[2].在中国西北干旱区,1961—2020年气温以约0.33 ℃/10a的速率持续升高;自2000年以来,降水以29 mm/10a的速率显著增加,呈现明显的暖湿化特征[3].持续的气候扰动显著抑制植被的光合效率[4-5],削弱生态系统生产力并改变生态系统结构,致使生态系统稳定性的不确定性提高[1],从而对生态系统的服务功能、区域的生态环境和社会经济的可持续发展构成系统性挑战.因此,探究气候变化对生态系统稳定性的影响及其驱动机制,对提升生态系统应对气候变化的能力,保障生态环境和社会经济可持续发展具有重要意义. ...
Stability and diversity of ecosystems
1
2007
... 生态系统稳定性是指生态系统在长期发展与演化过程中维持或恢复其结构与功能相对稳定的能力[6-8].近年来,生态系统稳定性主要集中在研究方法和影响机制的研究上.在研究方法上,建立了以抵抗力、恢复力与时间稳定性为核心的多维度量化框架[9],并提出了综合评价法、试验观测法和遥感分析法等评估生态系统稳定性方法.综合评价法通过构建多维度指标体系,并结合层次分析法[10-12]、模糊综合评价[13]、主成分分析[14]等方法进行评估,能全面评估生态系统的多维状况,但指标选取和权重设定较为主观;试验观测法依托野外调查和控制试验,利用物种组成[15]、生物多样性[16-17]、种群动态[18]等数据解释生态系统的抵抗力与恢复机制,适合小时空尺度机理研究;遥感分析法则利用归一化植被指数(Normalized Difference Vegetation Index,NDVI)、增强型植被指数(Enhanced Vegetation Index,EVI)、太阳诱导叶绿素荧光(Solar-Induced Chlorophyll Fluorescence,SIF)、总初级生产力(Gross Primary Productivity,GPP)、叶面积指数(Leaf Area Index,LAI)等指标表征生态系统稳定性[19-20],覆盖范围广、时间序列完整,适合揭示大尺度生态系统稳定性的格局和趋势.另外,探讨了气候、生物、人类活动对生态系统稳定性的影响.其中气候因子是最核心的外部驱动力[8,21].气候变暖会改变物种组成与群落结构,进而影响种间作用与生态系统稳定性[22];长期的水分胁迫则会导致植被代谢受限、光合效率下降,从而降低生态系统的抵抗力和恢复力[21,23-24].生物多样性则通过调节群落功能波动与养分循环增强生态系统稳定性[25-26],但物种同步性和种群变异性的增加可能导致生态系统稳定性下降[27].人类活动(如大规模的土地利用变化和森林砍伐)则导致生境丧失、生态功能退化,从而削弱生态系统的稳定性[28-29],而合理放牧等适度干扰又能缓解种间竞争,提升和恢复群落的稳定性[30-31].尽管相关研究取得了很大的进展,但是生态系统稳定性的概念和度量方法仍不统一,多重扰动的交互效应与稳定性下降区的精准识别仍缺乏系统性的探索,区域间的差异比较研究相对匮乏[32].典型区域尤其是干旱区高寒山地生态系统,由于自然环境复杂、观测条件受限,现有研究对其生态系统稳定性的变化特征及其驱动机制仍缺乏系统认识,亟需对相关问题进行系统分析. ...
Unifying the concepts of stability and resilience in ecology
0
2021
Global climate regulates dimensions of terrestrial ecosystem stability
2
2023
... 生态系统稳定性是指生态系统在长期发展与演化过程中维持或恢复其结构与功能相对稳定的能力[6-8].近年来,生态系统稳定性主要集中在研究方法和影响机制的研究上.在研究方法上,建立了以抵抗力、恢复力与时间稳定性为核心的多维度量化框架[9],并提出了综合评价法、试验观测法和遥感分析法等评估生态系统稳定性方法.综合评价法通过构建多维度指标体系,并结合层次分析法[10-12]、模糊综合评价[13]、主成分分析[14]等方法进行评估,能全面评估生态系统的多维状况,但指标选取和权重设定较为主观;试验观测法依托野外调查和控制试验,利用物种组成[15]、生物多样性[16-17]、种群动态[18]等数据解释生态系统的抵抗力与恢复机制,适合小时空尺度机理研究;遥感分析法则利用归一化植被指数(Normalized Difference Vegetation Index,NDVI)、增强型植被指数(Enhanced Vegetation Index,EVI)、太阳诱导叶绿素荧光(Solar-Induced Chlorophyll Fluorescence,SIF)、总初级生产力(Gross Primary Productivity,GPP)、叶面积指数(Leaf Area Index,LAI)等指标表征生态系统稳定性[19-20],覆盖范围广、时间序列完整,适合揭示大尺度生态系统稳定性的格局和趋势.另外,探讨了气候、生物、人类活动对生态系统稳定性的影响.其中气候因子是最核心的外部驱动力[8,21].气候变暖会改变物种组成与群落结构,进而影响种间作用与生态系统稳定性[22];长期的水分胁迫则会导致植被代谢受限、光合效率下降,从而降低生态系统的抵抗力和恢复力[21,23-24].生物多样性则通过调节群落功能波动与养分循环增强生态系统稳定性[25-26],但物种同步性和种群变异性的增加可能导致生态系统稳定性下降[27].人类活动(如大规模的土地利用变化和森林砍伐)则导致生境丧失、生态功能退化,从而削弱生态系统的稳定性[28-29],而合理放牧等适度干扰又能缓解种间竞争,提升和恢复群落的稳定性[30-31].尽管相关研究取得了很大的进展,但是生态系统稳定性的概念和度量方法仍不统一,多重扰动的交互效应与稳定性下降区的精准识别仍缺乏系统性的探索,区域间的差异比较研究相对匮乏[32].典型区域尤其是干旱区高寒山地生态系统,由于自然环境复杂、观测条件受限,现有研究对其生态系统稳定性的变化特征及其驱动机制仍缺乏系统认识,亟需对相关问题进行系统分析. ...
... [8,21].气候变暖会改变物种组成与群落结构,进而影响种间作用与生态系统稳定性[22];长期的水分胁迫则会导致植被代谢受限、光合效率下降,从而降低生态系统的抵抗力和恢复力[21,23-24].生物多样性则通过调节群落功能波动与养分循环增强生态系统稳定性[25-26],但物种同步性和种群变异性的增加可能导致生态系统稳定性下降[27].人类活动(如大规模的土地利用变化和森林砍伐)则导致生境丧失、生态功能退化,从而削弱生态系统的稳定性[28-29],而合理放牧等适度干扰又能缓解种间竞争,提升和恢复群落的稳定性[30-31].尽管相关研究取得了很大的进展,但是生态系统稳定性的概念和度量方法仍不统一,多重扰动的交互效应与稳定性下降区的精准识别仍缺乏系统性的探索,区域间的差异比较研究相对匮乏[32].典型区域尤其是干旱区高寒山地生态系统,由于自然环境复杂、观测条件受限,现有研究对其生态系统稳定性的变化特征及其驱动机制仍缺乏系统认识,亟需对相关问题进行系统分析. ...
陆地生态系统稳定性空间格局及影响机制研究综述
1
2021
... 生态系统稳定性是指生态系统在长期发展与演化过程中维持或恢复其结构与功能相对稳定的能力[6-8].近年来,生态系统稳定性主要集中在研究方法和影响机制的研究上.在研究方法上,建立了以抵抗力、恢复力与时间稳定性为核心的多维度量化框架[9],并提出了综合评价法、试验观测法和遥感分析法等评估生态系统稳定性方法.综合评价法通过构建多维度指标体系,并结合层次分析法[10-12]、模糊综合评价[13]、主成分分析[14]等方法进行评估,能全面评估生态系统的多维状况,但指标选取和权重设定较为主观;试验观测法依托野外调查和控制试验,利用物种组成[15]、生物多样性[16-17]、种群动态[18]等数据解释生态系统的抵抗力与恢复机制,适合小时空尺度机理研究;遥感分析法则利用归一化植被指数(Normalized Difference Vegetation Index,NDVI)、增强型植被指数(Enhanced Vegetation Index,EVI)、太阳诱导叶绿素荧光(Solar-Induced Chlorophyll Fluorescence,SIF)、总初级生产力(Gross Primary Productivity,GPP)、叶面积指数(Leaf Area Index,LAI)等指标表征生态系统稳定性[19-20],覆盖范围广、时间序列完整,适合揭示大尺度生态系统稳定性的格局和趋势.另外,探讨了气候、生物、人类活动对生态系统稳定性的影响.其中气候因子是最核心的外部驱动力[8,21].气候变暖会改变物种组成与群落结构,进而影响种间作用与生态系统稳定性[22];长期的水分胁迫则会导致植被代谢受限、光合效率下降,从而降低生态系统的抵抗力和恢复力[21,23-24].生物多样性则通过调节群落功能波动与养分循环增强生态系统稳定性[25-26],但物种同步性和种群变异性的增加可能导致生态系统稳定性下降[27].人类活动(如大规模的土地利用变化和森林砍伐)则导致生境丧失、生态功能退化,从而削弱生态系统的稳定性[28-29],而合理放牧等适度干扰又能缓解种间竞争,提升和恢复群落的稳定性[30-31].尽管相关研究取得了很大的进展,但是生态系统稳定性的概念和度量方法仍不统一,多重扰动的交互效应与稳定性下降区的精准识别仍缺乏系统性的探索,区域间的差异比较研究相对匮乏[32].典型区域尤其是干旱区高寒山地生态系统,由于自然环境复杂、观测条件受限,现有研究对其生态系统稳定性的变化特征及其驱动机制仍缺乏系统认识,亟需对相关问题进行系统分析. ...
鄱阳湖生态系统空间结构与湿地功能分析及稳定性评价
1
2016
... 生态系统稳定性是指生态系统在长期发展与演化过程中维持或恢复其结构与功能相对稳定的能力[6-8].近年来,生态系统稳定性主要集中在研究方法和影响机制的研究上.在研究方法上,建立了以抵抗力、恢复力与时间稳定性为核心的多维度量化框架[9],并提出了综合评价法、试验观测法和遥感分析法等评估生态系统稳定性方法.综合评价法通过构建多维度指标体系,并结合层次分析法[10-12]、模糊综合评价[13]、主成分分析[14]等方法进行评估,能全面评估生态系统的多维状况,但指标选取和权重设定较为主观;试验观测法依托野外调查和控制试验,利用物种组成[15]、生物多样性[16-17]、种群动态[18]等数据解释生态系统的抵抗力与恢复机制,适合小时空尺度机理研究;遥感分析法则利用归一化植被指数(Normalized Difference Vegetation Index,NDVI)、增强型植被指数(Enhanced Vegetation Index,EVI)、太阳诱导叶绿素荧光(Solar-Induced Chlorophyll Fluorescence,SIF)、总初级生产力(Gross Primary Productivity,GPP)、叶面积指数(Leaf Area Index,LAI)等指标表征生态系统稳定性[19-20],覆盖范围广、时间序列完整,适合揭示大尺度生态系统稳定性的格局和趋势.另外,探讨了气候、生物、人类活动对生态系统稳定性的影响.其中气候因子是最核心的外部驱动力[8,21].气候变暖会改变物种组成与群落结构,进而影响种间作用与生态系统稳定性[22];长期的水分胁迫则会导致植被代谢受限、光合效率下降,从而降低生态系统的抵抗力和恢复力[21,23-24].生物多样性则通过调节群落功能波动与养分循环增强生态系统稳定性[25-26],但物种同步性和种群变异性的增加可能导致生态系统稳定性下降[27].人类活动(如大规模的土地利用变化和森林砍伐)则导致生境丧失、生态功能退化,从而削弱生态系统的稳定性[28-29],而合理放牧等适度干扰又能缓解种间竞争,提升和恢复群落的稳定性[30-31].尽管相关研究取得了很大的进展,但是生态系统稳定性的概念和度量方法仍不统一,多重扰动的交互效应与稳定性下降区的精准识别仍缺乏系统性的探索,区域间的差异比较研究相对匮乏[32].典型区域尤其是干旱区高寒山地生态系统,由于自然环境复杂、观测条件受限,现有研究对其生态系统稳定性的变化特征及其驱动机制仍缺乏系统认识,亟需对相关问题进行系统分析. ...
Wetland ecosystem stability evaluation by using Analytical Hierarchy Process (AHP) approach in Yinchuan Plain,China
0
2013
吉林龙湾泥炭沼泽湿地生态系统稳定性研究
1
2021
... 生态系统稳定性是指生态系统在长期发展与演化过程中维持或恢复其结构与功能相对稳定的能力[6-8].近年来,生态系统稳定性主要集中在研究方法和影响机制的研究上.在研究方法上,建立了以抵抗力、恢复力与时间稳定性为核心的多维度量化框架[9],并提出了综合评价法、试验观测法和遥感分析法等评估生态系统稳定性方法.综合评价法通过构建多维度指标体系,并结合层次分析法[10-12]、模糊综合评价[13]、主成分分析[14]等方法进行评估,能全面评估生态系统的多维状况,但指标选取和权重设定较为主观;试验观测法依托野外调查和控制试验,利用物种组成[15]、生物多样性[16-17]、种群动态[18]等数据解释生态系统的抵抗力与恢复机制,适合小时空尺度机理研究;遥感分析法则利用归一化植被指数(Normalized Difference Vegetation Index,NDVI)、增强型植被指数(Enhanced Vegetation Index,EVI)、太阳诱导叶绿素荧光(Solar-Induced Chlorophyll Fluorescence,SIF)、总初级生产力(Gross Primary Productivity,GPP)、叶面积指数(Leaf Area Index,LAI)等指标表征生态系统稳定性[19-20],覆盖范围广、时间序列完整,适合揭示大尺度生态系统稳定性的格局和趋势.另外,探讨了气候、生物、人类活动对生态系统稳定性的影响.其中气候因子是最核心的外部驱动力[8,21].气候变暖会改变物种组成与群落结构,进而影响种间作用与生态系统稳定性[22];长期的水分胁迫则会导致植被代谢受限、光合效率下降,从而降低生态系统的抵抗力和恢复力[21,23-24].生物多样性则通过调节群落功能波动与养分循环增强生态系统稳定性[25-26],但物种同步性和种群变异性的增加可能导致生态系统稳定性下降[27].人类活动(如大规模的土地利用变化和森林砍伐)则导致生境丧失、生态功能退化,从而削弱生态系统的稳定性[28-29],而合理放牧等适度干扰又能缓解种间竞争,提升和恢复群落的稳定性[30-31].尽管相关研究取得了很大的进展,但是生态系统稳定性的概念和度量方法仍不统一,多重扰动的交互效应与稳定性下降区的精准识别仍缺乏系统性的探索,区域间的差异比较研究相对匮乏[32].典型区域尤其是干旱区高寒山地生态系统,由于自然环境复杂、观测条件受限,现有研究对其生态系统稳定性的变化特征及其驱动机制仍缺乏系统认识,亟需对相关问题进行系统分析. ...
长白山云冷杉林不同演替阶段群落稳定性
1
2015
... 生态系统稳定性是指生态系统在长期发展与演化过程中维持或恢复其结构与功能相对稳定的能力[6-8].近年来,生态系统稳定性主要集中在研究方法和影响机制的研究上.在研究方法上,建立了以抵抗力、恢复力与时间稳定性为核心的多维度量化框架[9],并提出了综合评价法、试验观测法和遥感分析法等评估生态系统稳定性方法.综合评价法通过构建多维度指标体系,并结合层次分析法[10-12]、模糊综合评价[13]、主成分分析[14]等方法进行评估,能全面评估生态系统的多维状况,但指标选取和权重设定较为主观;试验观测法依托野外调查和控制试验,利用物种组成[15]、生物多样性[16-17]、种群动态[18]等数据解释生态系统的抵抗力与恢复机制,适合小时空尺度机理研究;遥感分析法则利用归一化植被指数(Normalized Difference Vegetation Index,NDVI)、增强型植被指数(Enhanced Vegetation Index,EVI)、太阳诱导叶绿素荧光(Solar-Induced Chlorophyll Fluorescence,SIF)、总初级生产力(Gross Primary Productivity,GPP)、叶面积指数(Leaf Area Index,LAI)等指标表征生态系统稳定性[19-20],覆盖范围广、时间序列完整,适合揭示大尺度生态系统稳定性的格局和趋势.另外,探讨了气候、生物、人类活动对生态系统稳定性的影响.其中气候因子是最核心的外部驱动力[8,21].气候变暖会改变物种组成与群落结构,进而影响种间作用与生态系统稳定性[22];长期的水分胁迫则会导致植被代谢受限、光合效率下降,从而降低生态系统的抵抗力和恢复力[21,23-24].生物多样性则通过调节群落功能波动与养分循环增强生态系统稳定性[25-26],但物种同步性和种群变异性的增加可能导致生态系统稳定性下降[27].人类活动(如大规模的土地利用变化和森林砍伐)则导致生境丧失、生态功能退化,从而削弱生态系统的稳定性[28-29],而合理放牧等适度干扰又能缓解种间竞争,提升和恢复群落的稳定性[30-31].尽管相关研究取得了很大的进展,但是生态系统稳定性的概念和度量方法仍不统一,多重扰动的交互效应与稳定性下降区的精准识别仍缺乏系统性的探索,区域间的差异比较研究相对匮乏[32].典型区域尤其是干旱区高寒山地生态系统,由于自然环境复杂、观测条件受限,现有研究对其生态系统稳定性的变化特征及其驱动机制仍缺乏系统认识,亟需对相关问题进行系统分析. ...
秦岭松栎混交林群落的稳定性
1
2015
... 生态系统稳定性是指生态系统在长期发展与演化过程中维持或恢复其结构与功能相对稳定的能力[6-8].近年来,生态系统稳定性主要集中在研究方法和影响机制的研究上.在研究方法上,建立了以抵抗力、恢复力与时间稳定性为核心的多维度量化框架[9],并提出了综合评价法、试验观测法和遥感分析法等评估生态系统稳定性方法.综合评价法通过构建多维度指标体系,并结合层次分析法[10-12]、模糊综合评价[13]、主成分分析[14]等方法进行评估,能全面评估生态系统的多维状况,但指标选取和权重设定较为主观;试验观测法依托野外调查和控制试验,利用物种组成[15]、生物多样性[16-17]、种群动态[18]等数据解释生态系统的抵抗力与恢复机制,适合小时空尺度机理研究;遥感分析法则利用归一化植被指数(Normalized Difference Vegetation Index,NDVI)、增强型植被指数(Enhanced Vegetation Index,EVI)、太阳诱导叶绿素荧光(Solar-Induced Chlorophyll Fluorescence,SIF)、总初级生产力(Gross Primary Productivity,GPP)、叶面积指数(Leaf Area Index,LAI)等指标表征生态系统稳定性[19-20],覆盖范围广、时间序列完整,适合揭示大尺度生态系统稳定性的格局和趋势.另外,探讨了气候、生物、人类活动对生态系统稳定性的影响.其中气候因子是最核心的外部驱动力[8,21].气候变暖会改变物种组成与群落结构,进而影响种间作用与生态系统稳定性[22];长期的水分胁迫则会导致植被代谢受限、光合效率下降,从而降低生态系统的抵抗力和恢复力[21,23-24].生物多样性则通过调节群落功能波动与养分循环增强生态系统稳定性[25-26],但物种同步性和种群变异性的增加可能导致生态系统稳定性下降[27].人类活动(如大规模的土地利用变化和森林砍伐)则导致生境丧失、生态功能退化,从而削弱生态系统的稳定性[28-29],而合理放牧等适度干扰又能缓解种间竞争,提升和恢复群落的稳定性[30-31].尽管相关研究取得了很大的进展,但是生态系统稳定性的概念和度量方法仍不统一,多重扰动的交互效应与稳定性下降区的精准识别仍缺乏系统性的探索,区域间的差异比较研究相对匮乏[32].典型区域尤其是干旱区高寒山地生态系统,由于自然环境复杂、观测条件受限,现有研究对其生态系统稳定性的变化特征及其驱动机制仍缺乏系统认识,亟需对相关问题进行系统分析. ...
刈割、施肥对高寒草甸物种多样性与生态系统功能关系的影响及群落稳定性机制
1
2013
... 生态系统稳定性是指生态系统在长期发展与演化过程中维持或恢复其结构与功能相对稳定的能力[6-8].近年来,生态系统稳定性主要集中在研究方法和影响机制的研究上.在研究方法上,建立了以抵抗力、恢复力与时间稳定性为核心的多维度量化框架[9],并提出了综合评价法、试验观测法和遥感分析法等评估生态系统稳定性方法.综合评价法通过构建多维度指标体系,并结合层次分析法[10-12]、模糊综合评价[13]、主成分分析[14]等方法进行评估,能全面评估生态系统的多维状况,但指标选取和权重设定较为主观;试验观测法依托野外调查和控制试验,利用物种组成[15]、生物多样性[16-17]、种群动态[18]等数据解释生态系统的抵抗力与恢复机制,适合小时空尺度机理研究;遥感分析法则利用归一化植被指数(Normalized Difference Vegetation Index,NDVI)、增强型植被指数(Enhanced Vegetation Index,EVI)、太阳诱导叶绿素荧光(Solar-Induced Chlorophyll Fluorescence,SIF)、总初级生产力(Gross Primary Productivity,GPP)、叶面积指数(Leaf Area Index,LAI)等指标表征生态系统稳定性[19-20],覆盖范围广、时间序列完整,适合揭示大尺度生态系统稳定性的格局和趋势.另外,探讨了气候、生物、人类活动对生态系统稳定性的影响.其中气候因子是最核心的外部驱动力[8,21].气候变暖会改变物种组成与群落结构,进而影响种间作用与生态系统稳定性[22];长期的水分胁迫则会导致植被代谢受限、光合效率下降,从而降低生态系统的抵抗力和恢复力[21,23-24].生物多样性则通过调节群落功能波动与养分循环增强生态系统稳定性[25-26],但物种同步性和种群变异性的增加可能导致生态系统稳定性下降[27].人类活动(如大规模的土地利用变化和森林砍伐)则导致生境丧失、生态功能退化,从而削弱生态系统的稳定性[28-29],而合理放牧等适度干扰又能缓解种间竞争,提升和恢复群落的稳定性[30-31].尽管相关研究取得了很大的进展,但是生态系统稳定性的概念和度量方法仍不统一,多重扰动的交互效应与稳定性下降区的精准识别仍缺乏系统性的探索,区域间的差异比较研究相对匮乏[32].典型区域尤其是干旱区高寒山地生态系统,由于自然环境复杂、观测条件受限,现有研究对其生态系统稳定性的变化特征及其驱动机制仍缺乏系统认识,亟需对相关问题进行系统分析. ...
Biodiversity and ecosystem stability in a decade-long grassland experiment
1
2006
... 生态系统稳定性是指生态系统在长期发展与演化过程中维持或恢复其结构与功能相对稳定的能力[6-8].近年来,生态系统稳定性主要集中在研究方法和影响机制的研究上.在研究方法上,建立了以抵抗力、恢复力与时间稳定性为核心的多维度量化框架[9],并提出了综合评价法、试验观测法和遥感分析法等评估生态系统稳定性方法.综合评价法通过构建多维度指标体系,并结合层次分析法[10-12]、模糊综合评价[13]、主成分分析[14]等方法进行评估,能全面评估生态系统的多维状况,但指标选取和权重设定较为主观;试验观测法依托野外调查和控制试验,利用物种组成[15]、生物多样性[16-17]、种群动态[18]等数据解释生态系统的抵抗力与恢复机制,适合小时空尺度机理研究;遥感分析法则利用归一化植被指数(Normalized Difference Vegetation Index,NDVI)、增强型植被指数(Enhanced Vegetation Index,EVI)、太阳诱导叶绿素荧光(Solar-Induced Chlorophyll Fluorescence,SIF)、总初级生产力(Gross Primary Productivity,GPP)、叶面积指数(Leaf Area Index,LAI)等指标表征生态系统稳定性[19-20],覆盖范围广、时间序列完整,适合揭示大尺度生态系统稳定性的格局和趋势.另外,探讨了气候、生物、人类活动对生态系统稳定性的影响.其中气候因子是最核心的外部驱动力[8,21].气候变暖会改变物种组成与群落结构,进而影响种间作用与生态系统稳定性[22];长期的水分胁迫则会导致植被代谢受限、光合效率下降,从而降低生态系统的抵抗力和恢复力[21,23-24].生物多样性则通过调节群落功能波动与养分循环增强生态系统稳定性[25-26],但物种同步性和种群变异性的增加可能导致生态系统稳定性下降[27].人类活动(如大规模的土地利用变化和森林砍伐)则导致生境丧失、生态功能退化,从而削弱生态系统的稳定性[28-29],而合理放牧等适度干扰又能缓解种间竞争,提升和恢复群落的稳定性[30-31].尽管相关研究取得了很大的进展,但是生态系统稳定性的概念和度量方法仍不统一,多重扰动的交互效应与稳定性下降区的精准识别仍缺乏系统性的探索,区域间的差异比较研究相对匮乏[32].典型区域尤其是干旱区高寒山地生态系统,由于自然环境复杂、观测条件受限,现有研究对其生态系统稳定性的变化特征及其驱动机制仍缺乏系统认识,亟需对相关问题进行系统分析. ...
Environmental conditions are the dominant factor influencing stability of terrestrial ecosystems on the Tibetan Plateau
1
2023
... 生态系统稳定性是指生态系统在长期发展与演化过程中维持或恢复其结构与功能相对稳定的能力[6-8].近年来,生态系统稳定性主要集中在研究方法和影响机制的研究上.在研究方法上,建立了以抵抗力、恢复力与时间稳定性为核心的多维度量化框架[9],并提出了综合评价法、试验观测法和遥感分析法等评估生态系统稳定性方法.综合评价法通过构建多维度指标体系,并结合层次分析法[10-12]、模糊综合评价[13]、主成分分析[14]等方法进行评估,能全面评估生态系统的多维状况,但指标选取和权重设定较为主观;试验观测法依托野外调查和控制试验,利用物种组成[15]、生物多样性[16-17]、种群动态[18]等数据解释生态系统的抵抗力与恢复机制,适合小时空尺度机理研究;遥感分析法则利用归一化植被指数(Normalized Difference Vegetation Index,NDVI)、增强型植被指数(Enhanced Vegetation Index,EVI)、太阳诱导叶绿素荧光(Solar-Induced Chlorophyll Fluorescence,SIF)、总初级生产力(Gross Primary Productivity,GPP)、叶面积指数(Leaf Area Index,LAI)等指标表征生态系统稳定性[19-20],覆盖范围广、时间序列完整,适合揭示大尺度生态系统稳定性的格局和趋势.另外,探讨了气候、生物、人类活动对生态系统稳定性的影响.其中气候因子是最核心的外部驱动力[8,21].气候变暖会改变物种组成与群落结构,进而影响种间作用与生态系统稳定性[22];长期的水分胁迫则会导致植被代谢受限、光合效率下降,从而降低生态系统的抵抗力和恢复力[21,23-24].生物多样性则通过调节群落功能波动与养分循环增强生态系统稳定性[25-26],但物种同步性和种群变异性的增加可能导致生态系统稳定性下降[27].人类活动(如大规模的土地利用变化和森林砍伐)则导致生境丧失、生态功能退化,从而削弱生态系统的稳定性[28-29],而合理放牧等适度干扰又能缓解种间竞争,提升和恢复群落的稳定性[30-31].尽管相关研究取得了很大的进展,但是生态系统稳定性的概念和度量方法仍不统一,多重扰动的交互效应与稳定性下降区的精准识别仍缺乏系统性的探索,区域间的差异比较研究相对匮乏[32].典型区域尤其是干旱区高寒山地生态系统,由于自然环境复杂、观测条件受限,现有研究对其生态系统稳定性的变化特征及其驱动机制仍缺乏系统认识,亟需对相关问题进行系统分析. ...
The impact of diffusive migration on ecosystem stability
1
2015
... 生态系统稳定性是指生态系统在长期发展与演化过程中维持或恢复其结构与功能相对稳定的能力[6-8].近年来,生态系统稳定性主要集中在研究方法和影响机制的研究上.在研究方法上,建立了以抵抗力、恢复力与时间稳定性为核心的多维度量化框架[9],并提出了综合评价法、试验观测法和遥感分析法等评估生态系统稳定性方法.综合评价法通过构建多维度指标体系,并结合层次分析法[10-12]、模糊综合评价[13]、主成分分析[14]等方法进行评估,能全面评估生态系统的多维状况,但指标选取和权重设定较为主观;试验观测法依托野外调查和控制试验,利用物种组成[15]、生物多样性[16-17]、种群动态[18]等数据解释生态系统的抵抗力与恢复机制,适合小时空尺度机理研究;遥感分析法则利用归一化植被指数(Normalized Difference Vegetation Index,NDVI)、增强型植被指数(Enhanced Vegetation Index,EVI)、太阳诱导叶绿素荧光(Solar-Induced Chlorophyll Fluorescence,SIF)、总初级生产力(Gross Primary Productivity,GPP)、叶面积指数(Leaf Area Index,LAI)等指标表征生态系统稳定性[19-20],覆盖范围广、时间序列完整,适合揭示大尺度生态系统稳定性的格局和趋势.另外,探讨了气候、生物、人类活动对生态系统稳定性的影响.其中气候因子是最核心的外部驱动力[8,21].气候变暖会改变物种组成与群落结构,进而影响种间作用与生态系统稳定性[22];长期的水分胁迫则会导致植被代谢受限、光合效率下降,从而降低生态系统的抵抗力和恢复力[21,23-24].生物多样性则通过调节群落功能波动与养分循环增强生态系统稳定性[25-26],但物种同步性和种群变异性的增加可能导致生态系统稳定性下降[27].人类活动(如大规模的土地利用变化和森林砍伐)则导致生境丧失、生态功能退化,从而削弱生态系统的稳定性[28-29],而合理放牧等适度干扰又能缓解种间竞争,提升和恢复群落的稳定性[30-31].尽管相关研究取得了很大的进展,但是生态系统稳定性的概念和度量方法仍不统一,多重扰动的交互效应与稳定性下降区的精准识别仍缺乏系统性的探索,区域间的差异比较研究相对匮乏[32].典型区域尤其是干旱区高寒山地生态系统,由于自然环境复杂、观测条件受限,现有研究对其生态系统稳定性的变化特征及其驱动机制仍缺乏系统认识,亟需对相关问题进行系统分析. ...
Phylotype diversity within soil fungal functional groups drives ecosystem stability
1
2022
... 生态系统稳定性是指生态系统在长期发展与演化过程中维持或恢复其结构与功能相对稳定的能力[6-8].近年来,生态系统稳定性主要集中在研究方法和影响机制的研究上.在研究方法上,建立了以抵抗力、恢复力与时间稳定性为核心的多维度量化框架[9],并提出了综合评价法、试验观测法和遥感分析法等评估生态系统稳定性方法.综合评价法通过构建多维度指标体系,并结合层次分析法[10-12]、模糊综合评价[13]、主成分分析[14]等方法进行评估,能全面评估生态系统的多维状况,但指标选取和权重设定较为主观;试验观测法依托野外调查和控制试验,利用物种组成[15]、生物多样性[16-17]、种群动态[18]等数据解释生态系统的抵抗力与恢复机制,适合小时空尺度机理研究;遥感分析法则利用归一化植被指数(Normalized Difference Vegetation Index,NDVI)、增强型植被指数(Enhanced Vegetation Index,EVI)、太阳诱导叶绿素荧光(Solar-Induced Chlorophyll Fluorescence,SIF)、总初级生产力(Gross Primary Productivity,GPP)、叶面积指数(Leaf Area Index,LAI)等指标表征生态系统稳定性[19-20],覆盖范围广、时间序列完整,适合揭示大尺度生态系统稳定性的格局和趋势.另外,探讨了气候、生物、人类活动对生态系统稳定性的影响.其中气候因子是最核心的外部驱动力[8,21].气候变暖会改变物种组成与群落结构,进而影响种间作用与生态系统稳定性[22];长期的水分胁迫则会导致植被代谢受限、光合效率下降,从而降低生态系统的抵抗力和恢复力[21,23-24].生物多样性则通过调节群落功能波动与养分循环增强生态系统稳定性[25-26],但物种同步性和种群变异性的增加可能导致生态系统稳定性下降[27].人类活动(如大规模的土地利用变化和森林砍伐)则导致生境丧失、生态功能退化,从而削弱生态系统的稳定性[28-29],而合理放牧等适度干扰又能缓解种间竞争,提升和恢复群落的稳定性[30-31].尽管相关研究取得了很大的进展,但是生态系统稳定性的概念和度量方法仍不统一,多重扰动的交互效应与稳定性下降区的精准识别仍缺乏系统性的探索,区域间的差异比较研究相对匮乏[32].典型区域尤其是干旱区高寒山地生态系统,由于自然环境复杂、观测条件受限,现有研究对其生态系统稳定性的变化特征及其驱动机制仍缺乏系统认识,亟需对相关问题进行系统分析. ...
Soil biota diversity and plant diversity both contributed to ecosystem stability in grasslands
1
2023
... 生态系统稳定性是指生态系统在长期发展与演化过程中维持或恢复其结构与功能相对稳定的能力[6-8].近年来,生态系统稳定性主要集中在研究方法和影响机制的研究上.在研究方法上,建立了以抵抗力、恢复力与时间稳定性为核心的多维度量化框架[9],并提出了综合评价法、试验观测法和遥感分析法等评估生态系统稳定性方法.综合评价法通过构建多维度指标体系,并结合层次分析法[10-12]、模糊综合评价[13]、主成分分析[14]等方法进行评估,能全面评估生态系统的多维状况,但指标选取和权重设定较为主观;试验观测法依托野外调查和控制试验,利用物种组成[15]、生物多样性[16-17]、种群动态[18]等数据解释生态系统的抵抗力与恢复机制,适合小时空尺度机理研究;遥感分析法则利用归一化植被指数(Normalized Difference Vegetation Index,NDVI)、增强型植被指数(Enhanced Vegetation Index,EVI)、太阳诱导叶绿素荧光(Solar-Induced Chlorophyll Fluorescence,SIF)、总初级生产力(Gross Primary Productivity,GPP)、叶面积指数(Leaf Area Index,LAI)等指标表征生态系统稳定性[19-20],覆盖范围广、时间序列完整,适合揭示大尺度生态系统稳定性的格局和趋势.另外,探讨了气候、生物、人类活动对生态系统稳定性的影响.其中气候因子是最核心的外部驱动力[8,21].气候变暖会改变物种组成与群落结构,进而影响种间作用与生态系统稳定性[22];长期的水分胁迫则会导致植被代谢受限、光合效率下降,从而降低生态系统的抵抗力和恢复力[21,23-24].生物多样性则通过调节群落功能波动与养分循环增强生态系统稳定性[25-26],但物种同步性和种群变异性的增加可能导致生态系统稳定性下降[27].人类活动(如大规模的土地利用变化和森林砍伐)则导致生境丧失、生态功能退化,从而削弱生态系统的稳定性[28-29],而合理放牧等适度干扰又能缓解种间竞争,提升和恢复群落的稳定性[30-31].尽管相关研究取得了很大的进展,但是生态系统稳定性的概念和度量方法仍不统一,多重扰动的交互效应与稳定性下降区的精准识别仍缺乏系统性的探索,区域间的差异比较研究相对匮乏[32].典型区域尤其是干旱区高寒山地生态系统,由于自然环境复杂、观测条件受限,现有研究对其生态系统稳定性的变化特征及其驱动机制仍缺乏系统认识,亟需对相关问题进行系统分析. ...
High ecosystem stability of evergreen broadleaf forests under severe droughts
2
2019
... 生态系统稳定性是指生态系统在长期发展与演化过程中维持或恢复其结构与功能相对稳定的能力[6-8].近年来,生态系统稳定性主要集中在研究方法和影响机制的研究上.在研究方法上,建立了以抵抗力、恢复力与时间稳定性为核心的多维度量化框架[9],并提出了综合评价法、试验观测法和遥感分析法等评估生态系统稳定性方法.综合评价法通过构建多维度指标体系,并结合层次分析法[10-12]、模糊综合评价[13]、主成分分析[14]等方法进行评估,能全面评估生态系统的多维状况,但指标选取和权重设定较为主观;试验观测法依托野外调查和控制试验,利用物种组成[15]、生物多样性[16-17]、种群动态[18]等数据解释生态系统的抵抗力与恢复机制,适合小时空尺度机理研究;遥感分析法则利用归一化植被指数(Normalized Difference Vegetation Index,NDVI)、增强型植被指数(Enhanced Vegetation Index,EVI)、太阳诱导叶绿素荧光(Solar-Induced Chlorophyll Fluorescence,SIF)、总初级生产力(Gross Primary Productivity,GPP)、叶面积指数(Leaf Area Index,LAI)等指标表征生态系统稳定性[19-20],覆盖范围广、时间序列完整,适合揭示大尺度生态系统稳定性的格局和趋势.另外,探讨了气候、生物、人类活动对生态系统稳定性的影响.其中气候因子是最核心的外部驱动力[8,21].气候变暖会改变物种组成与群落结构,进而影响种间作用与生态系统稳定性[22];长期的水分胁迫则会导致植被代谢受限、光合效率下降,从而降低生态系统的抵抗力和恢复力[21,23-24].生物多样性则通过调节群落功能波动与养分循环增强生态系统稳定性[25-26],但物种同步性和种群变异性的增加可能导致生态系统稳定性下降[27].人类活动(如大规模的土地利用变化和森林砍伐)则导致生境丧失、生态功能退化,从而削弱生态系统的稳定性[28-29],而合理放牧等适度干扰又能缓解种间竞争,提升和恢复群落的稳定性[30-31].尽管相关研究取得了很大的进展,但是生态系统稳定性的概念和度量方法仍不统一,多重扰动的交互效应与稳定性下降区的精准识别仍缺乏系统性的探索,区域间的差异比较研究相对匮乏[32].典型区域尤其是干旱区高寒山地生态系统,由于自然环境复杂、观测条件受限,现有研究对其生态系统稳定性的变化特征及其驱动机制仍缺乏系统认识,亟需对相关问题进行系统分析. ...
... [21,23-24].生物多样性则通过调节群落功能波动与养分循环增强生态系统稳定性[25-26],但物种同步性和种群变异性的增加可能导致生态系统稳定性下降[27].人类活动(如大规模的土地利用变化和森林砍伐)则导致生境丧失、生态功能退化,从而削弱生态系统的稳定性[28-29],而合理放牧等适度干扰又能缓解种间竞争,提升和恢复群落的稳定性[30-31].尽管相关研究取得了很大的进展,但是生态系统稳定性的概念和度量方法仍不统一,多重扰动的交互效应与稳定性下降区的精准识别仍缺乏系统性的探索,区域间的差异比较研究相对匮乏[32].典型区域尤其是干旱区高寒山地生态系统,由于自然环境复杂、观测条件受限,现有研究对其生态系统稳定性的变化特征及其驱动机制仍缺乏系统认识,亟需对相关问题进行系统分析. ...
Multiple dimensions of climate change and their implications for biodiversity
1
2014
... 生态系统稳定性是指生态系统在长期发展与演化过程中维持或恢复其结构与功能相对稳定的能力[6-8].近年来,生态系统稳定性主要集中在研究方法和影响机制的研究上.在研究方法上,建立了以抵抗力、恢复力与时间稳定性为核心的多维度量化框架[9],并提出了综合评价法、试验观测法和遥感分析法等评估生态系统稳定性方法.综合评价法通过构建多维度指标体系,并结合层次分析法[10-12]、模糊综合评价[13]、主成分分析[14]等方法进行评估,能全面评估生态系统的多维状况,但指标选取和权重设定较为主观;试验观测法依托野外调查和控制试验,利用物种组成[15]、生物多样性[16-17]、种群动态[18]等数据解释生态系统的抵抗力与恢复机制,适合小时空尺度机理研究;遥感分析法则利用归一化植被指数(Normalized Difference Vegetation Index,NDVI)、增强型植被指数(Enhanced Vegetation Index,EVI)、太阳诱导叶绿素荧光(Solar-Induced Chlorophyll Fluorescence,SIF)、总初级生产力(Gross Primary Productivity,GPP)、叶面积指数(Leaf Area Index,LAI)等指标表征生态系统稳定性[19-20],覆盖范围广、时间序列完整,适合揭示大尺度生态系统稳定性的格局和趋势.另外,探讨了气候、生物、人类活动对生态系统稳定性的影响.其中气候因子是最核心的外部驱动力[8,21].气候变暖会改变物种组成与群落结构,进而影响种间作用与生态系统稳定性[22];长期的水分胁迫则会导致植被代谢受限、光合效率下降,从而降低生态系统的抵抗力和恢复力[21,23-24].生物多样性则通过调节群落功能波动与养分循环增强生态系统稳定性[25-26],但物种同步性和种群变异性的增加可能导致生态系统稳定性下降[27].人类活动(如大规模的土地利用变化和森林砍伐)则导致生境丧失、生态功能退化,从而削弱生态系统的稳定性[28-29],而合理放牧等适度干扰又能缓解种间竞争,提升和恢复群落的稳定性[30-31].尽管相关研究取得了很大的进展,但是生态系统稳定性的概念和度量方法仍不统一,多重扰动的交互效应与稳定性下降区的精准识别仍缺乏系统性的探索,区域间的差异比较研究相对匮乏[32].典型区域尤其是干旱区高寒山地生态系统,由于自然环境复杂、观测条件受限,现有研究对其生态系统稳定性的变化特征及其驱动机制仍缺乏系统认识,亟需对相关问题进行系统分析. ...
A framework for the resilience of seagrass ecosystems
1
2015
... 生态系统稳定性是指生态系统在长期发展与演化过程中维持或恢复其结构与功能相对稳定的能力[6-8].近年来,生态系统稳定性主要集中在研究方法和影响机制的研究上.在研究方法上,建立了以抵抗力、恢复力与时间稳定性为核心的多维度量化框架[9],并提出了综合评价法、试验观测法和遥感分析法等评估生态系统稳定性方法.综合评价法通过构建多维度指标体系,并结合层次分析法[10-12]、模糊综合评价[13]、主成分分析[14]等方法进行评估,能全面评估生态系统的多维状况,但指标选取和权重设定较为主观;试验观测法依托野外调查和控制试验,利用物种组成[15]、生物多样性[16-17]、种群动态[18]等数据解释生态系统的抵抗力与恢复机制,适合小时空尺度机理研究;遥感分析法则利用归一化植被指数(Normalized Difference Vegetation Index,NDVI)、增强型植被指数(Enhanced Vegetation Index,EVI)、太阳诱导叶绿素荧光(Solar-Induced Chlorophyll Fluorescence,SIF)、总初级生产力(Gross Primary Productivity,GPP)、叶面积指数(Leaf Area Index,LAI)等指标表征生态系统稳定性[19-20],覆盖范围广、时间序列完整,适合揭示大尺度生态系统稳定性的格局和趋势.另外,探讨了气候、生物、人类活动对生态系统稳定性的影响.其中气候因子是最核心的外部驱动力[8,21].气候变暖会改变物种组成与群落结构,进而影响种间作用与生态系统稳定性[22];长期的水分胁迫则会导致植被代谢受限、光合效率下降,从而降低生态系统的抵抗力和恢复力[21,23-24].生物多样性则通过调节群落功能波动与养分循环增强生态系统稳定性[25-26],但物种同步性和种群变异性的增加可能导致生态系统稳定性下降[27].人类活动(如大规模的土地利用变化和森林砍伐)则导致生境丧失、生态功能退化,从而削弱生态系统的稳定性[28-29],而合理放牧等适度干扰又能缓解种间竞争,提升和恢复群落的稳定性[30-31].尽管相关研究取得了很大的进展,但是生态系统稳定性的概念和度量方法仍不统一,多重扰动的交互效应与稳定性下降区的精准识别仍缺乏系统性的探索,区域间的差异比较研究相对匮乏[32].典型区域尤其是干旱区高寒山地生态系统,由于自然环境复杂、观测条件受限,现有研究对其生态系统稳定性的变化特征及其驱动机制仍缺乏系统认识,亟需对相关问题进行系统分析. ...
拔节期干旱胁迫下冬小麦品种间产量及生理响应的差异
1
2020
... 生态系统稳定性是指生态系统在长期发展与演化过程中维持或恢复其结构与功能相对稳定的能力[6-8].近年来,生态系统稳定性主要集中在研究方法和影响机制的研究上.在研究方法上,建立了以抵抗力、恢复力与时间稳定性为核心的多维度量化框架[9],并提出了综合评价法、试验观测法和遥感分析法等评估生态系统稳定性方法.综合评价法通过构建多维度指标体系,并结合层次分析法[10-12]、模糊综合评价[13]、主成分分析[14]等方法进行评估,能全面评估生态系统的多维状况,但指标选取和权重设定较为主观;试验观测法依托野外调查和控制试验,利用物种组成[15]、生物多样性[16-17]、种群动态[18]等数据解释生态系统的抵抗力与恢复机制,适合小时空尺度机理研究;遥感分析法则利用归一化植被指数(Normalized Difference Vegetation Index,NDVI)、增强型植被指数(Enhanced Vegetation Index,EVI)、太阳诱导叶绿素荧光(Solar-Induced Chlorophyll Fluorescence,SIF)、总初级生产力(Gross Primary Productivity,GPP)、叶面积指数(Leaf Area Index,LAI)等指标表征生态系统稳定性[19-20],覆盖范围广、时间序列完整,适合揭示大尺度生态系统稳定性的格局和趋势.另外,探讨了气候、生物、人类活动对生态系统稳定性的影响.其中气候因子是最核心的外部驱动力[8,21].气候变暖会改变物种组成与群落结构,进而影响种间作用与生态系统稳定性[22];长期的水分胁迫则会导致植被代谢受限、光合效率下降,从而降低生态系统的抵抗力和恢复力[21,23-24].生物多样性则通过调节群落功能波动与养分循环增强生态系统稳定性[25-26],但物种同步性和种群变异性的增加可能导致生态系统稳定性下降[27].人类活动(如大规模的土地利用变化和森林砍伐)则导致生境丧失、生态功能退化,从而削弱生态系统的稳定性[28-29],而合理放牧等适度干扰又能缓解种间竞争,提升和恢复群落的稳定性[30-31].尽管相关研究取得了很大的进展,但是生态系统稳定性的概念和度量方法仍不统一,多重扰动的交互效应与稳定性下降区的精准识别仍缺乏系统性的探索,区域间的差异比较研究相对匮乏[32].典型区域尤其是干旱区高寒山地生态系统,由于自然环境复杂、观测条件受限,现有研究对其生态系统稳定性的变化特征及其驱动机制仍缺乏系统认识,亟需对相关问题进行系统分析. ...
Ecological realism and mechanisms by which diversity begets stability
1
2009
... 生态系统稳定性是指生态系统在长期发展与演化过程中维持或恢复其结构与功能相对稳定的能力[6-8].近年来,生态系统稳定性主要集中在研究方法和影响机制的研究上.在研究方法上,建立了以抵抗力、恢复力与时间稳定性为核心的多维度量化框架[9],并提出了综合评价法、试验观测法和遥感分析法等评估生态系统稳定性方法.综合评价法通过构建多维度指标体系,并结合层次分析法[10-12]、模糊综合评价[13]、主成分分析[14]等方法进行评估,能全面评估生态系统的多维状况,但指标选取和权重设定较为主观;试验观测法依托野外调查和控制试验,利用物种组成[15]、生物多样性[16-17]、种群动态[18]等数据解释生态系统的抵抗力与恢复机制,适合小时空尺度机理研究;遥感分析法则利用归一化植被指数(Normalized Difference Vegetation Index,NDVI)、增强型植被指数(Enhanced Vegetation Index,EVI)、太阳诱导叶绿素荧光(Solar-Induced Chlorophyll Fluorescence,SIF)、总初级生产力(Gross Primary Productivity,GPP)、叶面积指数(Leaf Area Index,LAI)等指标表征生态系统稳定性[19-20],覆盖范围广、时间序列完整,适合揭示大尺度生态系统稳定性的格局和趋势.另外,探讨了气候、生物、人类活动对生态系统稳定性的影响.其中气候因子是最核心的外部驱动力[8,21].气候变暖会改变物种组成与群落结构,进而影响种间作用与生态系统稳定性[22];长期的水分胁迫则会导致植被代谢受限、光合效率下降,从而降低生态系统的抵抗力和恢复力[21,23-24].生物多样性则通过调节群落功能波动与养分循环增强生态系统稳定性[25-26],但物种同步性和种群变异性的增加可能导致生态系统稳定性下降[27].人类活动(如大规模的土地利用变化和森林砍伐)则导致生境丧失、生态功能退化,从而削弱生态系统的稳定性[28-29],而合理放牧等适度干扰又能缓解种间竞争,提升和恢复群落的稳定性[30-31].尽管相关研究取得了很大的进展,但是生态系统稳定性的概念和度量方法仍不统一,多重扰动的交互效应与稳定性下降区的精准识别仍缺乏系统性的探索,区域间的差异比较研究相对匮乏[32].典型区域尤其是干旱区高寒山地生态系统,由于自然环境复杂、观测条件受限,现有研究对其生态系统稳定性的变化特征及其驱动机制仍缺乏系统认识,亟需对相关问题进行系统分析. ...
Consistent effects of biodiversity loss on multifunctionality across contrasting ecosystems
1
2018
... 生态系统稳定性是指生态系统在长期发展与演化过程中维持或恢复其结构与功能相对稳定的能力[6-8].近年来,生态系统稳定性主要集中在研究方法和影响机制的研究上.在研究方法上,建立了以抵抗力、恢复力与时间稳定性为核心的多维度量化框架[9],并提出了综合评价法、试验观测法和遥感分析法等评估生态系统稳定性方法.综合评价法通过构建多维度指标体系,并结合层次分析法[10-12]、模糊综合评价[13]、主成分分析[14]等方法进行评估,能全面评估生态系统的多维状况,但指标选取和权重设定较为主观;试验观测法依托野外调查和控制试验,利用物种组成[15]、生物多样性[16-17]、种群动态[18]等数据解释生态系统的抵抗力与恢复机制,适合小时空尺度机理研究;遥感分析法则利用归一化植被指数(Normalized Difference Vegetation Index,NDVI)、增强型植被指数(Enhanced Vegetation Index,EVI)、太阳诱导叶绿素荧光(Solar-Induced Chlorophyll Fluorescence,SIF)、总初级生产力(Gross Primary Productivity,GPP)、叶面积指数(Leaf Area Index,LAI)等指标表征生态系统稳定性[19-20],覆盖范围广、时间序列完整,适合揭示大尺度生态系统稳定性的格局和趋势.另外,探讨了气候、生物、人类活动对生态系统稳定性的影响.其中气候因子是最核心的外部驱动力[8,21].气候变暖会改变物种组成与群落结构,进而影响种间作用与生态系统稳定性[22];长期的水分胁迫则会导致植被代谢受限、光合效率下降,从而降低生态系统的抵抗力和恢复力[21,23-24].生物多样性则通过调节群落功能波动与养分循环增强生态系统稳定性[25-26],但物种同步性和种群变异性的增加可能导致生态系统稳定性下降[27].人类活动(如大规模的土地利用变化和森林砍伐)则导致生境丧失、生态功能退化,从而削弱生态系统的稳定性[28-29],而合理放牧等适度干扰又能缓解种间竞争,提升和恢复群落的稳定性[30-31].尽管相关研究取得了很大的进展,但是生态系统稳定性的概念和度量方法仍不统一,多重扰动的交互效应与稳定性下降区的精准识别仍缺乏系统性的探索,区域间的差异比较研究相对匮乏[32].典型区域尤其是干旱区高寒山地生态系统,由于自然环境复杂、观测条件受限,现有研究对其生态系统稳定性的变化特征及其驱动机制仍缺乏系统认识,亟需对相关问题进行系统分析. ...
Nutrient-induced shifts of dominant species reduce ecosystem stability via increases in species synchrony and population variability
1
2019
... 生态系统稳定性是指生态系统在长期发展与演化过程中维持或恢复其结构与功能相对稳定的能力[6-8].近年来,生态系统稳定性主要集中在研究方法和影响机制的研究上.在研究方法上,建立了以抵抗力、恢复力与时间稳定性为核心的多维度量化框架[9],并提出了综合评价法、试验观测法和遥感分析法等评估生态系统稳定性方法.综合评价法通过构建多维度指标体系,并结合层次分析法[10-12]、模糊综合评价[13]、主成分分析[14]等方法进行评估,能全面评估生态系统的多维状况,但指标选取和权重设定较为主观;试验观测法依托野外调查和控制试验,利用物种组成[15]、生物多样性[16-17]、种群动态[18]等数据解释生态系统的抵抗力与恢复机制,适合小时空尺度机理研究;遥感分析法则利用归一化植被指数(Normalized Difference Vegetation Index,NDVI)、增强型植被指数(Enhanced Vegetation Index,EVI)、太阳诱导叶绿素荧光(Solar-Induced Chlorophyll Fluorescence,SIF)、总初级生产力(Gross Primary Productivity,GPP)、叶面积指数(Leaf Area Index,LAI)等指标表征生态系统稳定性[19-20],覆盖范围广、时间序列完整,适合揭示大尺度生态系统稳定性的格局和趋势.另外,探讨了气候、生物、人类活动对生态系统稳定性的影响.其中气候因子是最核心的外部驱动力[8,21].气候变暖会改变物种组成与群落结构,进而影响种间作用与生态系统稳定性[22];长期的水分胁迫则会导致植被代谢受限、光合效率下降,从而降低生态系统的抵抗力和恢复力[21,23-24].生物多样性则通过调节群落功能波动与养分循环增强生态系统稳定性[25-26],但物种同步性和种群变异性的增加可能导致生态系统稳定性下降[27].人类活动(如大规模的土地利用变化和森林砍伐)则导致生境丧失、生态功能退化,从而削弱生态系统的稳定性[28-29],而合理放牧等适度干扰又能缓解种间竞争,提升和恢复群落的稳定性[30-31].尽管相关研究取得了很大的进展,但是生态系统稳定性的概念和度量方法仍不统一,多重扰动的交互效应与稳定性下降区的精准识别仍缺乏系统性的探索,区域间的差异比较研究相对匮乏[32].典型区域尤其是干旱区高寒山地生态系统,由于自然环境复杂、观测条件受限,现有研究对其生态系统稳定性的变化特征及其驱动机制仍缺乏系统认识,亟需对相关问题进行系统分析. ...
退化森林生态系统恢复评价研究综述
1
2010
... 生态系统稳定性是指生态系统在长期发展与演化过程中维持或恢复其结构与功能相对稳定的能力[6-8].近年来,生态系统稳定性主要集中在研究方法和影响机制的研究上.在研究方法上,建立了以抵抗力、恢复力与时间稳定性为核心的多维度量化框架[9],并提出了综合评价法、试验观测法和遥感分析法等评估生态系统稳定性方法.综合评价法通过构建多维度指标体系,并结合层次分析法[10-12]、模糊综合评价[13]、主成分分析[14]等方法进行评估,能全面评估生态系统的多维状况,但指标选取和权重设定较为主观;试验观测法依托野外调查和控制试验,利用物种组成[15]、生物多样性[16-17]、种群动态[18]等数据解释生态系统的抵抗力与恢复机制,适合小时空尺度机理研究;遥感分析法则利用归一化植被指数(Normalized Difference Vegetation Index,NDVI)、增强型植被指数(Enhanced Vegetation Index,EVI)、太阳诱导叶绿素荧光(Solar-Induced Chlorophyll Fluorescence,SIF)、总初级生产力(Gross Primary Productivity,GPP)、叶面积指数(Leaf Area Index,LAI)等指标表征生态系统稳定性[19-20],覆盖范围广、时间序列完整,适合揭示大尺度生态系统稳定性的格局和趋势.另外,探讨了气候、生物、人类活动对生态系统稳定性的影响.其中气候因子是最核心的外部驱动力[8,21].气候变暖会改变物种组成与群落结构,进而影响种间作用与生态系统稳定性[22];长期的水分胁迫则会导致植被代谢受限、光合效率下降,从而降低生态系统的抵抗力和恢复力[21,23-24].生物多样性则通过调节群落功能波动与养分循环增强生态系统稳定性[25-26],但物种同步性和种群变异性的增加可能导致生态系统稳定性下降[27].人类活动(如大规模的土地利用变化和森林砍伐)则导致生境丧失、生态功能退化,从而削弱生态系统的稳定性[28-29],而合理放牧等适度干扰又能缓解种间竞争,提升和恢复群落的稳定性[30-31].尽管相关研究取得了很大的进展,但是生态系统稳定性的概念和度量方法仍不统一,多重扰动的交互效应与稳定性下降区的精准识别仍缺乏系统性的探索,区域间的差异比较研究相对匮乏[32].典型区域尤其是干旱区高寒山地生态系统,由于自然环境复杂、观测条件受限,现有研究对其生态系统稳定性的变化特征及其驱动机制仍缺乏系统认识,亟需对相关问题进行系统分析. ...
A pantropical analysis of the impacts of forest degradation and conversion on local temperature
1
2017
... 生态系统稳定性是指生态系统在长期发展与演化过程中维持或恢复其结构与功能相对稳定的能力[6-8].近年来,生态系统稳定性主要集中在研究方法和影响机制的研究上.在研究方法上,建立了以抵抗力、恢复力与时间稳定性为核心的多维度量化框架[9],并提出了综合评价法、试验观测法和遥感分析法等评估生态系统稳定性方法.综合评价法通过构建多维度指标体系,并结合层次分析法[10-12]、模糊综合评价[13]、主成分分析[14]等方法进行评估,能全面评估生态系统的多维状况,但指标选取和权重设定较为主观;试验观测法依托野外调查和控制试验,利用物种组成[15]、生物多样性[16-17]、种群动态[18]等数据解释生态系统的抵抗力与恢复机制,适合小时空尺度机理研究;遥感分析法则利用归一化植被指数(Normalized Difference Vegetation Index,NDVI)、增强型植被指数(Enhanced Vegetation Index,EVI)、太阳诱导叶绿素荧光(Solar-Induced Chlorophyll Fluorescence,SIF)、总初级生产力(Gross Primary Productivity,GPP)、叶面积指数(Leaf Area Index,LAI)等指标表征生态系统稳定性[19-20],覆盖范围广、时间序列完整,适合揭示大尺度生态系统稳定性的格局和趋势.另外,探讨了气候、生物、人类活动对生态系统稳定性的影响.其中气候因子是最核心的外部驱动力[8,21].气候变暖会改变物种组成与群落结构,进而影响种间作用与生态系统稳定性[22];长期的水分胁迫则会导致植被代谢受限、光合效率下降,从而降低生态系统的抵抗力和恢复力[21,23-24].生物多样性则通过调节群落功能波动与养分循环增强生态系统稳定性[25-26],但物种同步性和种群变异性的增加可能导致生态系统稳定性下降[27].人类活动(如大规模的土地利用变化和森林砍伐)则导致生境丧失、生态功能退化,从而削弱生态系统的稳定性[28-29],而合理放牧等适度干扰又能缓解种间竞争,提升和恢复群落的稳定性[30-31].尽管相关研究取得了很大的进展,但是生态系统稳定性的概念和度量方法仍不统一,多重扰动的交互效应与稳定性下降区的精准识别仍缺乏系统性的探索,区域间的差异比较研究相对匮乏[32].典型区域尤其是干旱区高寒山地生态系统,由于自然环境复杂、观测条件受限,现有研究对其生态系统稳定性的变化特征及其驱动机制仍缺乏系统认识,亟需对相关问题进行系统分析. ...
放牧对草地的作用
1
2006
... 生态系统稳定性是指生态系统在长期发展与演化过程中维持或恢复其结构与功能相对稳定的能力[6-8].近年来,生态系统稳定性主要集中在研究方法和影响机制的研究上.在研究方法上,建立了以抵抗力、恢复力与时间稳定性为核心的多维度量化框架[9],并提出了综合评价法、试验观测法和遥感分析法等评估生态系统稳定性方法.综合评价法通过构建多维度指标体系,并结合层次分析法[10-12]、模糊综合评价[13]、主成分分析[14]等方法进行评估,能全面评估生态系统的多维状况,但指标选取和权重设定较为主观;试验观测法依托野外调查和控制试验,利用物种组成[15]、生物多样性[16-17]、种群动态[18]等数据解释生态系统的抵抗力与恢复机制,适合小时空尺度机理研究;遥感分析法则利用归一化植被指数(Normalized Difference Vegetation Index,NDVI)、增强型植被指数(Enhanced Vegetation Index,EVI)、太阳诱导叶绿素荧光(Solar-Induced Chlorophyll Fluorescence,SIF)、总初级生产力(Gross Primary Productivity,GPP)、叶面积指数(Leaf Area Index,LAI)等指标表征生态系统稳定性[19-20],覆盖范围广、时间序列完整,适合揭示大尺度生态系统稳定性的格局和趋势.另外,探讨了气候、生物、人类活动对生态系统稳定性的影响.其中气候因子是最核心的外部驱动力[8,21].气候变暖会改变物种组成与群落结构,进而影响种间作用与生态系统稳定性[22];长期的水分胁迫则会导致植被代谢受限、光合效率下降,从而降低生态系统的抵抗力和恢复力[21,23-24].生物多样性则通过调节群落功能波动与养分循环增强生态系统稳定性[25-26],但物种同步性和种群变异性的增加可能导致生态系统稳定性下降[27].人类活动(如大规模的土地利用变化和森林砍伐)则导致生境丧失、生态功能退化,从而削弱生态系统的稳定性[28-29],而合理放牧等适度干扰又能缓解种间竞争,提升和恢复群落的稳定性[30-31].尽管相关研究取得了很大的进展,但是生态系统稳定性的概念和度量方法仍不统一,多重扰动的交互效应与稳定性下降区的精准识别仍缺乏系统性的探索,区域间的差异比较研究相对匮乏[32].典型区域尤其是干旱区高寒山地生态系统,由于自然环境复杂、观测条件受限,现有研究对其生态系统稳定性的变化特征及其驱动机制仍缺乏系统认识,亟需对相关问题进行系统分析. ...
两种放牧模式下高寒草甸群落植被构成及稳定性特征
1
2019
... 生态系统稳定性是指生态系统在长期发展与演化过程中维持或恢复其结构与功能相对稳定的能力[6-8].近年来,生态系统稳定性主要集中在研究方法和影响机制的研究上.在研究方法上,建立了以抵抗力、恢复力与时间稳定性为核心的多维度量化框架[9],并提出了综合评价法、试验观测法和遥感分析法等评估生态系统稳定性方法.综合评价法通过构建多维度指标体系,并结合层次分析法[10-12]、模糊综合评价[13]、主成分分析[14]等方法进行评估,能全面评估生态系统的多维状况,但指标选取和权重设定较为主观;试验观测法依托野外调查和控制试验,利用物种组成[15]、生物多样性[16-17]、种群动态[18]等数据解释生态系统的抵抗力与恢复机制,适合小时空尺度机理研究;遥感分析法则利用归一化植被指数(Normalized Difference Vegetation Index,NDVI)、增强型植被指数(Enhanced Vegetation Index,EVI)、太阳诱导叶绿素荧光(Solar-Induced Chlorophyll Fluorescence,SIF)、总初级生产力(Gross Primary Productivity,GPP)、叶面积指数(Leaf Area Index,LAI)等指标表征生态系统稳定性[19-20],覆盖范围广、时间序列完整,适合揭示大尺度生态系统稳定性的格局和趋势.另外,探讨了气候、生物、人类活动对生态系统稳定性的影响.其中气候因子是最核心的外部驱动力[8,21].气候变暖会改变物种组成与群落结构,进而影响种间作用与生态系统稳定性[22];长期的水分胁迫则会导致植被代谢受限、光合效率下降,从而降低生态系统的抵抗力和恢复力[21,23-24].生物多样性则通过调节群落功能波动与养分循环增强生态系统稳定性[25-26],但物种同步性和种群变异性的增加可能导致生态系统稳定性下降[27].人类活动(如大规模的土地利用变化和森林砍伐)则导致生境丧失、生态功能退化,从而削弱生态系统的稳定性[28-29],而合理放牧等适度干扰又能缓解种间竞争,提升和恢复群落的稳定性[30-31].尽管相关研究取得了很大的进展,但是生态系统稳定性的概念和度量方法仍不统一,多重扰动的交互效应与稳定性下降区的精准识别仍缺乏系统性的探索,区域间的差异比较研究相对匮乏[32].典型区域尤其是干旱区高寒山地生态系统,由于自然环境复杂、观测条件受限,现有研究对其生态系统稳定性的变化特征及其驱动机制仍缺乏系统认识,亟需对相关问题进行系统分析. ...
生态系统稳定性概念、评估方法及影响因素
1
2025
... 生态系统稳定性是指生态系统在长期发展与演化过程中维持或恢复其结构与功能相对稳定的能力[6-8].近年来,生态系统稳定性主要集中在研究方法和影响机制的研究上.在研究方法上,建立了以抵抗力、恢复力与时间稳定性为核心的多维度量化框架[9],并提出了综合评价法、试验观测法和遥感分析法等评估生态系统稳定性方法.综合评价法通过构建多维度指标体系,并结合层次分析法[10-12]、模糊综合评价[13]、主成分分析[14]等方法进行评估,能全面评估生态系统的多维状况,但指标选取和权重设定较为主观;试验观测法依托野外调查和控制试验,利用物种组成[15]、生物多样性[16-17]、种群动态[18]等数据解释生态系统的抵抗力与恢复机制,适合小时空尺度机理研究;遥感分析法则利用归一化植被指数(Normalized Difference Vegetation Index,NDVI)、增强型植被指数(Enhanced Vegetation Index,EVI)、太阳诱导叶绿素荧光(Solar-Induced Chlorophyll Fluorescence,SIF)、总初级生产力(Gross Primary Productivity,GPP)、叶面积指数(Leaf Area Index,LAI)等指标表征生态系统稳定性[19-20],覆盖范围广、时间序列完整,适合揭示大尺度生态系统稳定性的格局和趋势.另外,探讨了气候、生物、人类活动对生态系统稳定性的影响.其中气候因子是最核心的外部驱动力[8,21].气候变暖会改变物种组成与群落结构,进而影响种间作用与生态系统稳定性[22];长期的水分胁迫则会导致植被代谢受限、光合效率下降,从而降低生态系统的抵抗力和恢复力[21,23-24].生物多样性则通过调节群落功能波动与养分循环增强生态系统稳定性[25-26],但物种同步性和种群变异性的增加可能导致生态系统稳定性下降[27].人类活动(如大规模的土地利用变化和森林砍伐)则导致生境丧失、生态功能退化,从而削弱生态系统的稳定性[28-29],而合理放牧等适度干扰又能缓解种间竞争,提升和恢复群落的稳定性[30-31].尽管相关研究取得了很大的进展,但是生态系统稳定性的概念和度量方法仍不统一,多重扰动的交互效应与稳定性下降区的精准识别仍缺乏系统性的探索,区域间的差异比较研究相对匮乏[32].典型区域尤其是干旱区高寒山地生态系统,由于自然环境复杂、观测条件受限,现有研究对其生态系统稳定性的变化特征及其驱动机制仍缺乏系统认识,亟需对相关问题进行系统分析. ...
祁连山不同林地类型土壤特性及其水源涵养功能
1
2010
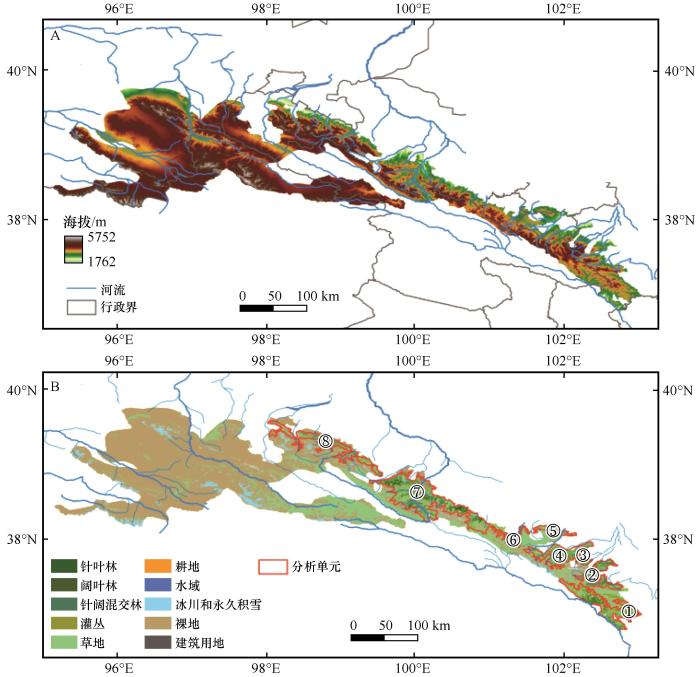
... 祁连山是中国西北干旱区生态安全的重要屏障,特别是其乔木林生态系统起着核心作用.祁连山的乔木林生态系统主要分布在黑河、石羊河和疏勒河的河川径流形成区,承担着涵养水源、调节径流的关键功能[33],在水土保持、区域气候调节[34]和固碳增汇[35]中也发挥着重要作用,素有“绿色水库”之称.气候变化将改变乔木林生态系统的结构、组成和功能[36-37],进而可能降低其生态系统稳定性,削弱其涵养水源的功能,加剧水资源短缺[38],从而威胁河西走廊生态环境的安全和社会经济的可持续发展.因此,选择祁连山国家公园为研究对象,开展气候变化对乔木林生态系统稳定性影响的研究,以揭示祁连山乔木林生态系统稳定性的时空变化规律,探究气候变化对其生态系统稳定性的影响机制,将对建立科学的干旱区山地乔木林生态系统应对气候变化的管理策略有重要意义. ...
干旱半干旱区山地森林的水分调节功能
1
2001
... 祁连山是中国西北干旱区生态安全的重要屏障,特别是其乔木林生态系统起着核心作用.祁连山的乔木林生态系统主要分布在黑河、石羊河和疏勒河的河川径流形成区,承担着涵养水源、调节径流的关键功能[33],在水土保持、区域气候调节[34]和固碳增汇[35]中也发挥着重要作用,素有“绿色水库”之称.气候变化将改变乔木林生态系统的结构、组成和功能[36-37],进而可能降低其生态系统稳定性,削弱其涵养水源的功能,加剧水资源短缺[38],从而威胁河西走廊生态环境的安全和社会经济的可持续发展.因此,选择祁连山国家公园为研究对象,开展气候变化对乔木林生态系统稳定性影响的研究,以揭示祁连山乔木林生态系统稳定性的时空变化规律,探究气候变化对其生态系统稳定性的影响机制,将对建立科学的干旱区山地乔木林生态系统应对气候变化的管理策略有重要意义. ...
中国森林乔木林碳储量及其固碳潜力预测
1
2018
... 祁连山是中国西北干旱区生态安全的重要屏障,特别是其乔木林生态系统起着核心作用.祁连山的乔木林生态系统主要分布在黑河、石羊河和疏勒河的河川径流形成区,承担着涵养水源、调节径流的关键功能[33],在水土保持、区域气候调节[34]和固碳增汇[35]中也发挥着重要作用,素有“绿色水库”之称.气候变化将改变乔木林生态系统的结构、组成和功能[36-37],进而可能降低其生态系统稳定性,削弱其涵养水源的功能,加剧水资源短缺[38],从而威胁河西走廊生态环境的安全和社会经济的可持续发展.因此,选择祁连山国家公园为研究对象,开展气候变化对乔木林生态系统稳定性影响的研究,以揭示祁连山乔木林生态系统稳定性的时空变化规律,探究气候变化对其生态系统稳定性的影响机制,将对建立科学的干旱区山地乔木林生态系统应对气候变化的管理策略有重要意义. ...
Impacts of climatic and edaphic factors on the diversity,structure and biomass of species-poor and structurally-complex forests
1
2020
... 祁连山是中国西北干旱区生态安全的重要屏障,特别是其乔木林生态系统起着核心作用.祁连山的乔木林生态系统主要分布在黑河、石羊河和疏勒河的河川径流形成区,承担着涵养水源、调节径流的关键功能[33],在水土保持、区域气候调节[34]和固碳增汇[35]中也发挥着重要作用,素有“绿色水库”之称.气候变化将改变乔木林生态系统的结构、组成和功能[36-37],进而可能降低其生态系统稳定性,削弱其涵养水源的功能,加剧水资源短缺[38],从而威胁河西走廊生态环境的安全和社会经济的可持续发展.因此,选择祁连山国家公园为研究对象,开展气候变化对乔木林生态系统稳定性影响的研究,以揭示祁连山乔木林生态系统稳定性的时空变化规律,探究气候变化对其生态系统稳定性的影响机制,将对建立科学的干旱区山地乔木林生态系统应对气候变化的管理策略有重要意义. ...
Global patterns and climatic controls of forest structural complexity
1
2021
... 祁连山是中国西北干旱区生态安全的重要屏障,特别是其乔木林生态系统起着核心作用.祁连山的乔木林生态系统主要分布在黑河、石羊河和疏勒河的河川径流形成区,承担着涵养水源、调节径流的关键功能[33],在水土保持、区域气候调节[34]和固碳增汇[35]中也发挥着重要作用,素有“绿色水库”之称.气候变化将改变乔木林生态系统的结构、组成和功能[36-37],进而可能降低其生态系统稳定性,削弱其涵养水源的功能,加剧水资源短缺[38],从而威胁河西走廊生态环境的安全和社会经济的可持续发展.因此,选择祁连山国家公园为研究对象,开展气候变化对乔木林生态系统稳定性影响的研究,以揭示祁连山乔木林生态系统稳定性的时空变化规律,探究气候变化对其生态系统稳定性的影响机制,将对建立科学的干旱区山地乔木林生态系统应对气候变化的管理策略有重要意义. ...
Trees,forests and water:cool insights for a hot world
1
2017
... 祁连山是中国西北干旱区生态安全的重要屏障,特别是其乔木林生态系统起着核心作用.祁连山的乔木林生态系统主要分布在黑河、石羊河和疏勒河的河川径流形成区,承担着涵养水源、调节径流的关键功能[33],在水土保持、区域气候调节[34]和固碳增汇[35]中也发挥着重要作用,素有“绿色水库”之称.气候变化将改变乔木林生态系统的结构、组成和功能[36-37],进而可能降低其生态系统稳定性,削弱其涵养水源的功能,加剧水资源短缺[38],从而威胁河西走廊生态环境的安全和社会经济的可持续发展.因此,选择祁连山国家公园为研究对象,开展气候变化对乔木林生态系统稳定性影响的研究,以揭示祁连山乔木林生态系统稳定性的时空变化规律,探究气候变化对其生态系统稳定性的影响机制,将对建立科学的干旱区山地乔木林生态系统应对气候变化的管理策略有重要意义. ...
祁连山国家公园土地利用类型数据集(1990-2020)
1
2023
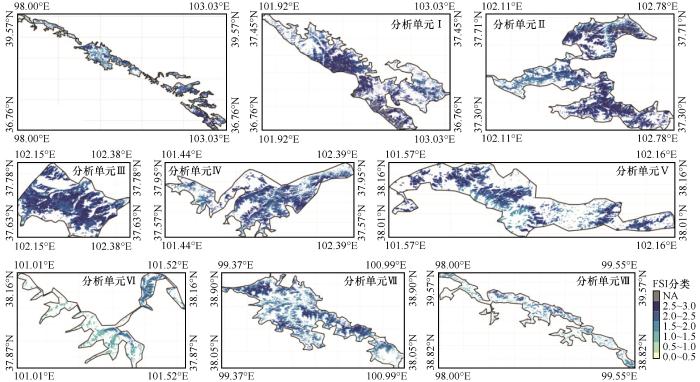
... 首先,基于遥感解译的土地利用数据[39],采用8邻接原则将乔木林像元聚合为斑块.然后,采用空间距离阈值法将相距小于1 000 m的乔木林斑块合并,并进一步以流域范围为生态边界,将合并后的斑块整合为初始分析单元.最后,结合地形特征和土地利用背景信息,对整合后的初始分析单元的边界进行优化,剔除过小和孤立斑块,确保分析单元在空间上相对完整、功能上相对一致,形成了空间尺度较大、生态功能明确的乔木林景观分析单元Ⅰ~Ⅷ.其次,利用R语言工具计算香农多样性指数(Shannon's Diversity Index, SHDI)、辛普森多样性指数(Simpson's Diversity Index, SIDI)、香农均匀度指数(Shannon's Evenness Index, SHEI)、景观形状指数(Landscape Shape Index, LSI)、平均斑块周长(Mean Patch Perimeter, PERIM_MN)、边界密度(Edge Density, ED)、斑块数(Number of Patches, NP)、斑块密度(Patch Density, PD)、平均斑块面积(Mean Patch Area, AREA_MN)、最大斑块指数(Largest Patch Index, LPI)、平均最近邻距离(Mean Euclidean Nearest Neighbor Distance,ENN_MN)、平均连续度指数(Mean Contiguity Index,CONTIG_MN)等景观格局指数[40],以定量分析分析单元的景观格局特征. ...
Empirical patterns of the effects of changing scale on landscape metrics
1
2002
... 首先,基于遥感解译的土地利用数据[39],采用8邻接原则将乔木林像元聚合为斑块.然后,采用空间距离阈值法将相距小于1 000 m的乔木林斑块合并,并进一步以流域范围为生态边界,将合并后的斑块整合为初始分析单元.最后,结合地形特征和土地利用背景信息,对整合后的初始分析单元的边界进行优化,剔除过小和孤立斑块,确保分析单元在空间上相对完整、功能上相对一致,形成了空间尺度较大、生态功能明确的乔木林景观分析单元Ⅰ~Ⅷ.其次,利用R语言工具计算香农多样性指数(Shannon's Diversity Index, SHDI)、辛普森多样性指数(Simpson's Diversity Index, SIDI)、香农均匀度指数(Shannon's Evenness Index, SHEI)、景观形状指数(Landscape Shape Index, LSI)、平均斑块周长(Mean Patch Perimeter, PERIM_MN)、边界密度(Edge Density, ED)、斑块数(Number of Patches, NP)、斑块密度(Patch Density, PD)、平均斑块面积(Mean Patch Area, AREA_MN)、最大斑块指数(Largest Patch Index, LPI)、平均最近邻距离(Mean Euclidean Nearest Neighbor Distance,ENN_MN)、平均连续度指数(Mean Contiguity Index,CONTIG_MN)等景观格局指数[40],以定量分析分析单元的景观格局特征. ...
Detecting dryland degradation using Time Series Segmentation and Residual Trend analysis (TSS-RESTREND)
1
2017
... 气候弹性指数(Climate Elasticity, CE)是指 NDVI 变化中未能由气温和降水的线性关系解释的波动,相对于气候因子总体变异性的比值,该指数可用于反映植被变化中气候因子之外的综合扰动效应[41-43].基于以上定义,在给定的滑动时间窗口(5 a)内建立 NDVI 与气温、降水之间的线性回归方程,并基于回归残差构建气候弹性指数. ...
RESTREND-based assessment of factors affecting vegetation dynamics on the Mongolian Plateau
0
2021
Ecological restoration policy should pay more attention to the high productivity grasslands
1
2021
... 气候弹性指数(Climate Elasticity, CE)是指 NDVI 变化中未能由气温和降水的线性关系解释的波动,相对于气候因子总体变异性的比值,该指数可用于反映植被变化中气候因子之外的综合扰动效应[41-43].基于以上定义,在给定的滑动时间窗口(5 a)内建立 NDVI 与气温、降水之间的线性回归方程,并基于回归残差构建气候弹性指数. ...
VDCALC framework:a comprehensive approach for assessing vegetation dynamics
1
2024
... 式中:CVi,j,t 表示像元(i,j)在第t个滑动时间窗口内的NDVI变异系数;σi,j,t 和μi,j,t 分别为该时间窗口内的NDVI的标准差与均值.CV值越小,说明植被波动性越低,生态系统越稳定[44-45]. ...
The complexity and stability of ecosystems
1
1984
... 式中:CVi,j,t 表示像元(i,j)在第t个滑动时间窗口内的NDVI变异系数;σi,j,t 和μi,j,t 分别为该时间窗口内的NDVI的标准差与均值.CV值越小,说明植被波动性越低,生态系统越稳定[44-45]. ...
Integrating landsat NDVI data with climate and anthropogenic factors reveals drivers of vegetation dynamics in the semi-arid basin of western China
1
2025
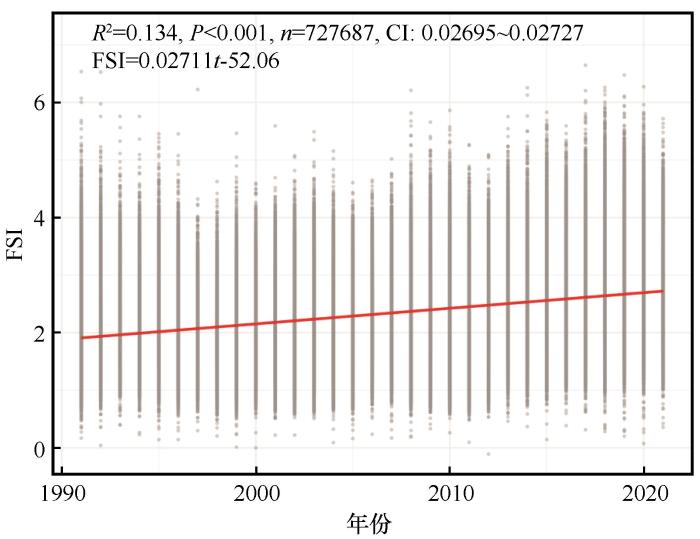
... 研究区内98.2%的乔木林生态系统多年NDVI变异系数小于0.223,该阈值可作为稳定性分级的重要参考[46-47].为增强CV值在低值区的区分度,对乔木林生态系统的时间稳定性进行负对数转换,从而定义乔木林生态系统的森林稳定性指数(Forest Stability Index, FSI)为: ...
Driving mechanisms of productivity stability vary with selective harvesting intensities in a mixed broad-leaved Korean pine forest
1
2023
... 研究区内98.2%的乔木林生态系统多年NDVI变异系数小于0.223,该阈值可作为稳定性分级的重要参考[46-47].为增强CV值在低值区的区分度,对乔木林生态系统的时间稳定性进行负对数转换,从而定义乔木林生态系统的森林稳定性指数(Forest Stability Index, FSI)为: ...
祁连山国家公园植被时空变化及其对人类活动的响应
1
2023
... 本研究结果表明,1991—2021年祁连山国家公园乔木林生态系统中极高稳定性面积占比77.25%,且乔木林生态系统整体的FSI以0.027 a⁻¹(α=0.05,tα=±1.96<335.92,n=727 687)的速率显著提升,这一趋势与区域内其他生态系统功能的研究一致.李玉辰等[48]指出,祁连山国家公园的NDVI和NPP整体上升,其中87.29%的区域NPP显著提高,生态系统服务功能(如生境质量、土壤保持、水源涵养和碳储存等)亦显著增强[49-50].Zhang等[51]基于NDVI计算青藏高原植被覆盖度的研究也表明,2000—2015年青藏高原60.30%的面积上植被覆盖度呈增加趋势,且显著增加区域主要分布在青藏高原北部. ...
Ecological conservation pattern based on ecosystem services in the Qilian Mountains,Northwest China
1
2023
... 本研究结果表明,1991—2021年祁连山国家公园乔木林生态系统中极高稳定性面积占比77.25%,且乔木林生态系统整体的FSI以0.027 a⁻¹(α=0.05,tα=±1.96<335.92,n=727 687)的速率显著提升,这一趋势与区域内其他生态系统功能的研究一致.李玉辰等[48]指出,祁连山国家公园的NDVI和NPP整体上升,其中87.29%的区域NPP显著提高,生态系统服务功能(如生境质量、土壤保持、水源涵养和碳储存等)亦显著增强[49-50].Zhang等[51]基于NDVI计算青藏高原植被覆盖度的研究也表明,2000—2015年青藏高原60.30%的面积上植被覆盖度呈增加趋势,且显著增加区域主要分布在青藏高原北部. ...
2000-2020年祁连山生态系统服务时空分异研究
1
2023
... 本研究结果表明,1991—2021年祁连山国家公园乔木林生态系统中极高稳定性面积占比77.25%,且乔木林生态系统整体的FSI以0.027 a⁻¹(α=0.05,tα=±1.96<335.92,n=727 687)的速率显著提升,这一趋势与区域内其他生态系统功能的研究一致.李玉辰等[48]指出,祁连山国家公园的NDVI和NPP整体上升,其中87.29%的区域NPP显著提高,生态系统服务功能(如生境质量、土壤保持、水源涵养和碳储存等)亦显著增强[49-50].Zhang等[51]基于NDVI计算青藏高原植被覆盖度的研究也表明,2000—2015年青藏高原60.30%的面积上植被覆盖度呈增加趋势,且显著增加区域主要分布在青藏高原北部. ...
Vegetation change and its relationship with climate factors and elevation on the Tibetan Plateau
1
2019
... 本研究结果表明,1991—2021年祁连山国家公园乔木林生态系统中极高稳定性面积占比77.25%,且乔木林生态系统整体的FSI以0.027 a⁻¹(α=0.05,tα=±1.96<335.92,n=727 687)的速率显著提升,这一趋势与区域内其他生态系统功能的研究一致.李玉辰等[48]指出,祁连山国家公园的NDVI和NPP整体上升,其中87.29%的区域NPP显著提高,生态系统服务功能(如生境质量、土壤保持、水源涵养和碳储存等)亦显著增强[49-50].Zhang等[51]基于NDVI计算青藏高原植被覆盖度的研究也表明,2000—2015年青藏高原60.30%的面积上植被覆盖度呈增加趋势,且显著增加区域主要分布在青藏高原北部. ...
Temperature response of photosynthesis in C3,C4,and CAM plants:temperature acclimation and temperature adaptation
1
2014
... 在研究时段内,回归结果显示气候变暖促进了乔木林生态系统稳定性的提升.而气温升高对生态系统稳定性的正效应可能与光合作用增强和生长季延长有关.温度上升在一定范围内可以提高光合作用效率和生产力,进而提升森林的抵抗力和恢复力[52].Wang等[53]利用北美洲的森林永久样地的研究同样证明,气温升高能够促进树木的生长,提高森林的生产力,从而增强生态系统稳定性,但白云杉(Picea glauca)、山杨(Populus tremuloides)、杰克松(Pinus banksiana)的生产力在夏季最高温分别到达23.4、27.0、26.2 ℃时开始下降,说明升温对生态系统稳定性的影响温度存在阈值.过度升温可能导致干旱胁迫和呼吸消耗增加,从而削弱稳定性[4].Ma等[54]在青藏高原高寒草地的试验中发现,气候变暖显著降低了该地区生态系统的时间稳定性.Zhao等[55]通过评估温度升高背景下的19个浮游生物食物网的稳定性,发现温度升高会降低其生态稳定性,这意味着生态系统稳定性在不同群落中对气温变化的响应可能存在差异.本研究中气温变化对生态系统稳定性的影响为“正效应”,这可能是气温变化仍处于乔木林生态系统可承受温度的适宜区间,而未来持续升温是否对生态系统稳定性造成负面影响还需进一步研究. ...
Warming-induced tree growth may help offset increasing disturbance across the Canadian boreal forest
1
2023
... 在研究时段内,回归结果显示气候变暖促进了乔木林生态系统稳定性的提升.而气温升高对生态系统稳定性的正效应可能与光合作用增强和生长季延长有关.温度上升在一定范围内可以提高光合作用效率和生产力,进而提升森林的抵抗力和恢复力[52].Wang等[53]利用北美洲的森林永久样地的研究同样证明,气温升高能够促进树木的生长,提高森林的生产力,从而增强生态系统稳定性,但白云杉(Picea glauca)、山杨(Populus tremuloides)、杰克松(Pinus banksiana)的生产力在夏季最高温分别到达23.4、27.0、26.2 ℃时开始下降,说明升温对生态系统稳定性的影响温度存在阈值.过度升温可能导致干旱胁迫和呼吸消耗增加,从而削弱稳定性[4].Ma等[54]在青藏高原高寒草地的试验中发现,气候变暖显著降低了该地区生态系统的时间稳定性.Zhao等[55]通过评估温度升高背景下的19个浮游生物食物网的稳定性,发现温度升高会降低其生态稳定性,这意味着生态系统稳定性在不同群落中对气温变化的响应可能存在差异.本研究中气温变化对生态系统稳定性的影响为“正效应”,这可能是气温变化仍处于乔木林生态系统可承受温度的适宜区间,而未来持续升温是否对生态系统稳定性造成负面影响还需进一步研究. ...
Climate warming reduces the temporal stability of plant community biomass production
1
2017
... 在研究时段内,回归结果显示气候变暖促进了乔木林生态系统稳定性的提升.而气温升高对生态系统稳定性的正效应可能与光合作用增强和生长季延长有关.温度上升在一定范围内可以提高光合作用效率和生产力,进而提升森林的抵抗力和恢复力[52].Wang等[53]利用北美洲的森林永久样地的研究同样证明,气温升高能够促进树木的生长,提高森林的生产力,从而增强生态系统稳定性,但白云杉(Picea glauca)、山杨(Populus tremuloides)、杰克松(Pinus banksiana)的生产力在夏季最高温分别到达23.4、27.0、26.2 ℃时开始下降,说明升温对生态系统稳定性的影响温度存在阈值.过度升温可能导致干旱胁迫和呼吸消耗增加,从而削弱稳定性[4].Ma等[54]在青藏高原高寒草地的试验中发现,气候变暖显著降低了该地区生态系统的时间稳定性.Zhao等[55]通过评估温度升高背景下的19个浮游生物食物网的稳定性,发现温度升高会降低其生态稳定性,这意味着生态系统稳定性在不同群落中对气温变化的响应可能存在差异.本研究中气温变化对生态系统稳定性的影响为“正效应”,这可能是气温变化仍处于乔木林生态系统可承受温度的适宜区间,而未来持续升温是否对生态系统稳定性造成负面影响还需进一步研究. ...
Relationships of temperature and biodiversity with stability of natural aquatic food webs
1
2023
... 在研究时段内,回归结果显示气候变暖促进了乔木林生态系统稳定性的提升.而气温升高对生态系统稳定性的正效应可能与光合作用增强和生长季延长有关.温度上升在一定范围内可以提高光合作用效率和生产力,进而提升森林的抵抗力和恢复力[52].Wang等[53]利用北美洲的森林永久样地的研究同样证明,气温升高能够促进树木的生长,提高森林的生产力,从而增强生态系统稳定性,但白云杉(Picea glauca)、山杨(Populus tremuloides)、杰克松(Pinus banksiana)的生产力在夏季最高温分别到达23.4、27.0、26.2 ℃时开始下降,说明升温对生态系统稳定性的影响温度存在阈值.过度升温可能导致干旱胁迫和呼吸消耗增加,从而削弱稳定性[4].Ma等[54]在青藏高原高寒草地的试验中发现,气候变暖显著降低了该地区生态系统的时间稳定性.Zhao等[55]通过评估温度升高背景下的19个浮游生物食物网的稳定性,发现温度升高会降低其生态稳定性,这意味着生态系统稳定性在不同群落中对气温变化的响应可能存在差异.本研究中气温变化对生态系统稳定性的影响为“正效应”,这可能是气温变化仍处于乔木林生态系统可承受温度的适宜区间,而未来持续升温是否对生态系统稳定性造成负面影响还需进一步研究. ...
Divergent driving mechanisms of community temporal stability in China's drylands
1
2024
... 本研究结果显示,在分析单元Ⅱ、Ⅲ、Ⅳ、Ⅴ、Ⅶ、Ⅷ中降水量的增加促进了生态系统稳定性的提升.这与现有研究结论一致,即在干旱与半干旱地区,降水量增加通过提高植被的平均生产力、物种异步性等,进而增强群落的时间稳定性[56-58].Liu等[59]通过对青藏高原高寒草地生态系统的79个样地开展实地调查获得的植物物种多样性、年净初级生产力和土壤理化性质的研究,指出降水量的增加通过提高物种多样性和增强地下生态系统多功能性,间接促进群落生产力的提升,从而提高生态系统稳定性.Smith等[60]的全球尺度研究也表明,植被稳定性随着水分可利用性的提高而增强.然而在分析单元Ⅵ中,降水量对生态系统稳定性的影响相对微弱;在分析单元Ⅰ中,降水量增加对生态系统稳定性反而表现为负效应(-0.26).这一现象表明,降水对稳定性的作用并非单向度的正效应,而是受到区域背景水热条件的调控[61].在本研究中,单元Ⅰ位于祁连山国家公园的最东端,其年降水量高于中西部地区,在这一相对湿润的本底条件,额外的降水可能带来多重负面影响.其一,持续增加的降水可能造成土壤过湿甚至渍涝,导致根系缺氧、病原滋生与个体抗逆性的下降[62-63];其二,降水量的持续增加还会加剧土壤侵蚀和养分流失,从而削弱群落维持稳定性的能力[64];其三,降水量增加可能伴随云量增加、辐射减弱,从而导致区域光照条件恶化,限制光合作用,降低植被生产力、削弱生态系统稳定性[65]. ...
Temporal variation characteristics in the association between climate and vegetation in Northwest China
0
2024
Precipitation variability has a weak but significant stabilizing effect on community structure
1
2024
... 本研究结果显示,在分析单元Ⅱ、Ⅲ、Ⅳ、Ⅴ、Ⅶ、Ⅷ中降水量的增加促进了生态系统稳定性的提升.这与现有研究结论一致,即在干旱与半干旱地区,降水量增加通过提高植被的平均生产力、物种异步性等,进而增强群落的时间稳定性[56-58].Liu等[59]通过对青藏高原高寒草地生态系统的79个样地开展实地调查获得的植物物种多样性、年净初级生产力和土壤理化性质的研究,指出降水量的增加通过提高物种多样性和增强地下生态系统多功能性,间接促进群落生产力的提升,从而提高生态系统稳定性.Smith等[60]的全球尺度研究也表明,植被稳定性随着水分可利用性的提高而增强.然而在分析单元Ⅵ中,降水量对生态系统稳定性的影响相对微弱;在分析单元Ⅰ中,降水量增加对生态系统稳定性反而表现为负效应(-0.26).这一现象表明,降水对稳定性的作用并非单向度的正效应,而是受到区域背景水热条件的调控[61].在本研究中,单元Ⅰ位于祁连山国家公园的最东端,其年降水量高于中西部地区,在这一相对湿润的本底条件,额外的降水可能带来多重负面影响.其一,持续增加的降水可能造成土壤过湿甚至渍涝,导致根系缺氧、病原滋生与个体抗逆性的下降[62-63];其二,降水量的持续增加还会加剧土壤侵蚀和养分流失,从而削弱群落维持稳定性的能力[64];其三,降水量增加可能伴随云量增加、辐射减弱,从而导致区域光照条件恶化,限制光合作用,降低植被生产力、削弱生态系统稳定性[65]. ...
Precipitation mediates the effects of species diversity and belowground ecosystem multifunctionality on community stability across alpine grasslands,Qinghai-Xizang Plateau
1
2025
... 本研究结果显示,在分析单元Ⅱ、Ⅲ、Ⅳ、Ⅴ、Ⅶ、Ⅷ中降水量的增加促进了生态系统稳定性的提升.这与现有研究结论一致,即在干旱与半干旱地区,降水量增加通过提高植被的平均生产力、物种异步性等,进而增强群落的时间稳定性[56-58].Liu等[59]通过对青藏高原高寒草地生态系统的79个样地开展实地调查获得的植物物种多样性、年净初级生产力和土壤理化性质的研究,指出降水量的增加通过提高物种多样性和增强地下生态系统多功能性,间接促进群落生产力的提升,从而提高生态系统稳定性.Smith等[60]的全球尺度研究也表明,植被稳定性随着水分可利用性的提高而增强.然而在分析单元Ⅵ中,降水量对生态系统稳定性的影响相对微弱;在分析单元Ⅰ中,降水量增加对生态系统稳定性反而表现为负效应(-0.26).这一现象表明,降水对稳定性的作用并非单向度的正效应,而是受到区域背景水热条件的调控[61].在本研究中,单元Ⅰ位于祁连山国家公园的最东端,其年降水量高于中西部地区,在这一相对湿润的本底条件,额外的降水可能带来多重负面影响.其一,持续增加的降水可能造成土壤过湿甚至渍涝,导致根系缺氧、病原滋生与个体抗逆性的下降[62-63];其二,降水量的持续增加还会加剧土壤侵蚀和养分流失,从而削弱群落维持稳定性的能力[64];其三,降水量增加可能伴随云量增加、辐射减弱,从而导致区域光照条件恶化,限制光合作用,降低植被生产力、削弱生态系统稳定性[65]. ...
Global vegetation resilience linked to water availability and variability
1
2023
... 本研究结果显示,在分析单元Ⅱ、Ⅲ、Ⅳ、Ⅴ、Ⅶ、Ⅷ中降水量的增加促进了生态系统稳定性的提升.这与现有研究结论一致,即在干旱与半干旱地区,降水量增加通过提高植被的平均生产力、物种异步性等,进而增强群落的时间稳定性[56-58].Liu等[59]通过对青藏高原高寒草地生态系统的79个样地开展实地调查获得的植物物种多样性、年净初级生产力和土壤理化性质的研究,指出降水量的增加通过提高物种多样性和增强地下生态系统多功能性,间接促进群落生产力的提升,从而提高生态系统稳定性.Smith等[60]的全球尺度研究也表明,植被稳定性随着水分可利用性的提高而增强.然而在分析单元Ⅵ中,降水量对生态系统稳定性的影响相对微弱;在分析单元Ⅰ中,降水量增加对生态系统稳定性反而表现为负效应(-0.26).这一现象表明,降水对稳定性的作用并非单向度的正效应,而是受到区域背景水热条件的调控[61].在本研究中,单元Ⅰ位于祁连山国家公园的最东端,其年降水量高于中西部地区,在这一相对湿润的本底条件,额外的降水可能带来多重负面影响.其一,持续增加的降水可能造成土壤过湿甚至渍涝,导致根系缺氧、病原滋生与个体抗逆性的下降[62-63];其二,降水量的持续增加还会加剧土壤侵蚀和养分流失,从而削弱群落维持稳定性的能力[64];其三,降水量增加可能伴随云量增加、辐射减弱,从而导致区域光照条件恶化,限制光合作用,降低植被生产力、削弱生态系统稳定性[65]. ...
Vegetation response to changes in climate across different climate zones in China
1
2023
... 本研究结果显示,在分析单元Ⅱ、Ⅲ、Ⅳ、Ⅴ、Ⅶ、Ⅷ中降水量的增加促进了生态系统稳定性的提升.这与现有研究结论一致,即在干旱与半干旱地区,降水量增加通过提高植被的平均生产力、物种异步性等,进而增强群落的时间稳定性[56-58].Liu等[59]通过对青藏高原高寒草地生态系统的79个样地开展实地调查获得的植物物种多样性、年净初级生产力和土壤理化性质的研究,指出降水量的增加通过提高物种多样性和增强地下生态系统多功能性,间接促进群落生产力的提升,从而提高生态系统稳定性.Smith等[60]的全球尺度研究也表明,植被稳定性随着水分可利用性的提高而增强.然而在分析单元Ⅵ中,降水量对生态系统稳定性的影响相对微弱;在分析单元Ⅰ中,降水量增加对生态系统稳定性反而表现为负效应(-0.26).这一现象表明,降水对稳定性的作用并非单向度的正效应,而是受到区域背景水热条件的调控[61].在本研究中,单元Ⅰ位于祁连山国家公园的最东端,其年降水量高于中西部地区,在这一相对湿润的本底条件,额外的降水可能带来多重负面影响.其一,持续增加的降水可能造成土壤过湿甚至渍涝,导致根系缺氧、病原滋生与个体抗逆性的下降[62-63];其二,降水量的持续增加还会加剧土壤侵蚀和养分流失,从而削弱群落维持稳定性的能力[64];其三,降水量增加可能伴随云量增加、辐射减弱,从而导致区域光照条件恶化,限制光合作用,降低植被生产力、削弱生态系统稳定性[65]. ...
A review of soil waterlogging impacts,mechanisms,and adaptive strategies
1
2025
... 本研究结果显示,在分析单元Ⅱ、Ⅲ、Ⅳ、Ⅴ、Ⅶ、Ⅷ中降水量的增加促进了生态系统稳定性的提升.这与现有研究结论一致,即在干旱与半干旱地区,降水量增加通过提高植被的平均生产力、物种异步性等,进而增强群落的时间稳定性[56-58].Liu等[59]通过对青藏高原高寒草地生态系统的79个样地开展实地调查获得的植物物种多样性、年净初级生产力和土壤理化性质的研究,指出降水量的增加通过提高物种多样性和增强地下生态系统多功能性,间接促进群落生产力的提升,从而提高生态系统稳定性.Smith等[60]的全球尺度研究也表明,植被稳定性随着水分可利用性的提高而增强.然而在分析单元Ⅵ中,降水量对生态系统稳定性的影响相对微弱;在分析单元Ⅰ中,降水量增加对生态系统稳定性反而表现为负效应(-0.26).这一现象表明,降水对稳定性的作用并非单向度的正效应,而是受到区域背景水热条件的调控[61].在本研究中,单元Ⅰ位于祁连山国家公园的最东端,其年降水量高于中西部地区,在这一相对湿润的本底条件,额外的降水可能带来多重负面影响.其一,持续增加的降水可能造成土壤过湿甚至渍涝,导致根系缺氧、病原滋生与个体抗逆性的下降[62-63];其二,降水量的持续增加还会加剧土壤侵蚀和养分流失,从而削弱群落维持稳定性的能力[64];其三,降水量增加可能伴随云量增加、辐射减弱,从而导致区域光照条件恶化,限制光合作用,降低植被生产力、削弱生态系统稳定性[65]. ...
Waterlogging of soil induces diverging rates of senescence in Svalbard reindeer forage plants
1
2025
... 本研究结果显示,在分析单元Ⅱ、Ⅲ、Ⅳ、Ⅴ、Ⅶ、Ⅷ中降水量的增加促进了生态系统稳定性的提升.这与现有研究结论一致,即在干旱与半干旱地区,降水量增加通过提高植被的平均生产力、物种异步性等,进而增强群落的时间稳定性[56-58].Liu等[59]通过对青藏高原高寒草地生态系统的79个样地开展实地调查获得的植物物种多样性、年净初级生产力和土壤理化性质的研究,指出降水量的增加通过提高物种多样性和增强地下生态系统多功能性,间接促进群落生产力的提升,从而提高生态系统稳定性.Smith等[60]的全球尺度研究也表明,植被稳定性随着水分可利用性的提高而增强.然而在分析单元Ⅵ中,降水量对生态系统稳定性的影响相对微弱;在分析单元Ⅰ中,降水量增加对生态系统稳定性反而表现为负效应(-0.26).这一现象表明,降水对稳定性的作用并非单向度的正效应,而是受到区域背景水热条件的调控[61].在本研究中,单元Ⅰ位于祁连山国家公园的最东端,其年降水量高于中西部地区,在这一相对湿润的本底条件,额外的降水可能带来多重负面影响.其一,持续增加的降水可能造成土壤过湿甚至渍涝,导致根系缺氧、病原滋生与个体抗逆性的下降[62-63];其二,降水量的持续增加还会加剧土壤侵蚀和养分流失,从而削弱群落维持稳定性的能力[64];其三,降水量增加可能伴随云量增加、辐射减弱,从而导致区域光照条件恶化,限制光合作用,降低植被生产力、削弱生态系统稳定性[65]. ...
Increased precipitation weakens the positive effect of vegetation greening on erosion
1
2023
... 本研究结果显示,在分析单元Ⅱ、Ⅲ、Ⅳ、Ⅴ、Ⅶ、Ⅷ中降水量的增加促进了生态系统稳定性的提升.这与现有研究结论一致,即在干旱与半干旱地区,降水量增加通过提高植被的平均生产力、物种异步性等,进而增强群落的时间稳定性[56-58].Liu等[59]通过对青藏高原高寒草地生态系统的79个样地开展实地调查获得的植物物种多样性、年净初级生产力和土壤理化性质的研究,指出降水量的增加通过提高物种多样性和增强地下生态系统多功能性,间接促进群落生产力的提升,从而提高生态系统稳定性.Smith等[60]的全球尺度研究也表明,植被稳定性随着水分可利用性的提高而增强.然而在分析单元Ⅵ中,降水量对生态系统稳定性的影响相对微弱;在分析单元Ⅰ中,降水量增加对生态系统稳定性反而表现为负效应(-0.26).这一现象表明,降水对稳定性的作用并非单向度的正效应,而是受到区域背景水热条件的调控[61].在本研究中,单元Ⅰ位于祁连山国家公园的最东端,其年降水量高于中西部地区,在这一相对湿润的本底条件,额外的降水可能带来多重负面影响.其一,持续增加的降水可能造成土壤过湿甚至渍涝,导致根系缺氧、病原滋生与个体抗逆性的下降[62-63];其二,降水量的持续增加还会加剧土壤侵蚀和养分流失,从而削弱群落维持稳定性的能力[64];其三,降水量增加可能伴随云量增加、辐射减弱,从而导致区域光照条件恶化,限制光合作用,降低植被生产力、削弱生态系统稳定性[65]. ...
Effect of diffuse fraction on gross primary productivity and light use efficiency in a warm-temperate mixed plantation
1
2022
... 本研究结果显示,在分析单元Ⅱ、Ⅲ、Ⅳ、Ⅴ、Ⅶ、Ⅷ中降水量的增加促进了生态系统稳定性的提升.这与现有研究结论一致,即在干旱与半干旱地区,降水量增加通过提高植被的平均生产力、物种异步性等,进而增强群落的时间稳定性[56-58].Liu等[59]通过对青藏高原高寒草地生态系统的79个样地开展实地调查获得的植物物种多样性、年净初级生产力和土壤理化性质的研究,指出降水量的增加通过提高物种多样性和增强地下生态系统多功能性,间接促进群落生产力的提升,从而提高生态系统稳定性.Smith等[60]的全球尺度研究也表明,植被稳定性随着水分可利用性的提高而增强.然而在分析单元Ⅵ中,降水量对生态系统稳定性的影响相对微弱;在分析单元Ⅰ中,降水量增加对生态系统稳定性反而表现为负效应(-0.26).这一现象表明,降水对稳定性的作用并非单向度的正效应,而是受到区域背景水热条件的调控[61].在本研究中,单元Ⅰ位于祁连山国家公园的最东端,其年降水量高于中西部地区,在这一相对湿润的本底条件,额外的降水可能带来多重负面影响.其一,持续增加的降水可能造成土壤过湿甚至渍涝,导致根系缺氧、病原滋生与个体抗逆性的下降[62-63];其二,降水量的持续增加还会加剧土壤侵蚀和养分流失,从而削弱群落维持稳定性的能力[64];其三,降水量增加可能伴随云量增加、辐射减弱,从而导致区域光照条件恶化,限制光合作用,降低植被生产力、削弱生态系统稳定性[65]. ...
Landscape heterogeneity buffers biodiversity of simulated meta-food-webs under global change through rescue and drainage effects
1
2021
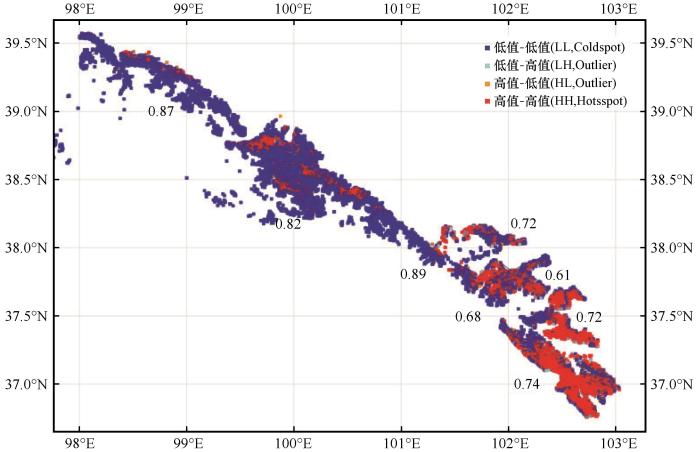
... 本研究发现,气候弹性指数在所有分析单元中对乔木林生态系统稳定性的影响均为负效应,其CE值每增加0.10,FSI值为-1.00~-1.75.这一结果表明,FSI除受到气温和降水的影响外,还受到非气候因子的制约.分析单元Ⅶ和Ⅷ中CE值增加对FSI影响的负效应最强,这些区域景观破碎化程度较高、空间异质性较低,说明景观条件差可能放大了非气候扰动对FSI的影响[66].相比之下,在景观多样性高、异质性强的分析单元(Ⅰ、Ⅱ、Ⅲ),CE 虽仍表现为负效应,但数值较小(-1.00~-1.25),表明景观结构较好时可以在一定程度上缓冲非气候扰动的不利影响[67]. ...
An analysis on the spatial heterogeneity characteristics of landscape ecological risk in Qilian Mountain National Park
1
2024
... 本研究发现,气候弹性指数在所有分析单元中对乔木林生态系统稳定性的影响均为负效应,其CE值每增加0.10,FSI值为-1.00~-1.75.这一结果表明,FSI除受到气温和降水的影响外,还受到非气候因子的制约.分析单元Ⅶ和Ⅷ中CE值增加对FSI影响的负效应最强,这些区域景观破碎化程度较高、空间异质性较低,说明景观条件差可能放大了非气候扰动对FSI的影响[66].相比之下,在景观多样性高、异质性强的分析单元(Ⅰ、Ⅱ、Ⅲ),CE 虽仍表现为负效应,但数值较小(-1.00~-1.25),表明景观结构较好时可以在一定程度上缓冲非气候扰动的不利影响[67]. ...
Responses of terrestrial ecosystems to temperature and precipitation change:a meta-analysis of experimental manipulation
1
2011
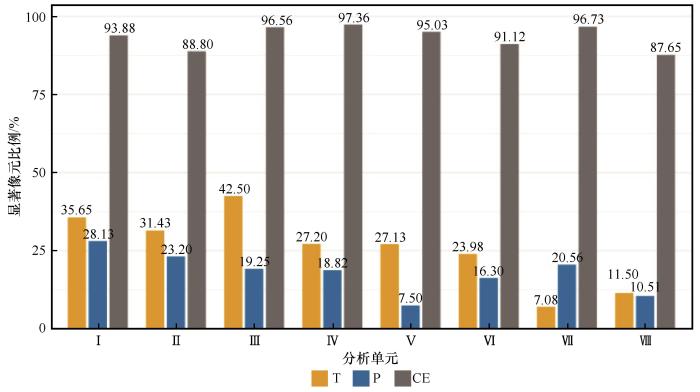
... 本研究结果显示,气温与降水对乔木林生态系统稳定性的独立解释力相对有限,其与FSI显著相关的像元比例分别为7.08%~42.50%和7.50%~28.13%,对FSI的贡献率仅为5.00%~13.00%和4.00%~10.00%,与已有研究结果一致,即单一气候因子往往难以充分解释生态系统过程的主要变异[68-70].Li等[71]的研究进一步显示,全球中高纬地区春季复合暖干(CWD)、温度和降水对夏季植被变异的显著相关面积分别为27.00%、15.00%和24.00%,也表明了复合指标的解释力通常优于单因子.本研究中的气候弹性指数(CE)正是利用残差法构建的用于代理非气候因素的综合指标,其与FSI的显著相关的像元比例为87.65%~97.36%,对FSI的贡献率为33.00%~47.00%,明显高于气温的或降水的.多元回归模型的R²为0.41~0.56,表明气温、降水和CE等3类因子能够在一定程度上解释乔木林生态系统稳定性的变化,但仍有44%~59%的未解释部分.这是因为CE仅是对非气候因素的近似度量,不足以完全涵盖植被结构、土壤环境、地形格局和人类活动等因素[72-73].因此,未来研究应纳入更多直接表征非气候因素的指标,以进一步提升模型对生态系统稳定性变异机制的解释能力. ...
Quantitative assessment of spatiotemporal variations and drivers of gross primary productivity in tropical ecosystems at higher resolution
0
2024
Plant functional diversity shapes soil respiration response to soil moisture availability
1
2025
... 本研究结果显示,气温与降水对乔木林生态系统稳定性的独立解释力相对有限,其与FSI显著相关的像元比例分别为7.08%~42.50%和7.50%~28.13%,对FSI的贡献率仅为5.00%~13.00%和4.00%~10.00%,与已有研究结果一致,即单一气候因子往往难以充分解释生态系统过程的主要变异[68-70].Li等[71]的研究进一步显示,全球中高纬地区春季复合暖干(CWD)、温度和降水对夏季植被变异的显著相关面积分别为27.00%、15.00%和24.00%,也表明了复合指标的解释力通常优于单因子.本研究中的气候弹性指数(CE)正是利用残差法构建的用于代理非气候因素的综合指标,其与FSI的显著相关的像元比例为87.65%~97.36%,对FSI的贡献率为33.00%~47.00%,明显高于气温的或降水的.多元回归模型的R²为0.41~0.56,表明气温、降水和CE等3类因子能够在一定程度上解释乔木林生态系统稳定性的变化,但仍有44%~59%的未解释部分.这是因为CE仅是对非气候因素的近似度量,不足以完全涵盖植被结构、土壤环境、地形格局和人类活动等因素[72-73].因此,未来研究应纳入更多直接表征非气候因素的指标,以进一步提升模型对生态系统稳定性变异机制的解释能力. ...
Regional asymmetry in the response of global vegetation growth to springtime compound climate events
1
2022
... 本研究结果显示,气温与降水对乔木林生态系统稳定性的独立解释力相对有限,其与FSI显著相关的像元比例分别为7.08%~42.50%和7.50%~28.13%,对FSI的贡献率仅为5.00%~13.00%和4.00%~10.00%,与已有研究结果一致,即单一气候因子往往难以充分解释生态系统过程的主要变异[68-70].Li等[71]的研究进一步显示,全球中高纬地区春季复合暖干(CWD)、温度和降水对夏季植被变异的显著相关面积分别为27.00%、15.00%和24.00%,也表明了复合指标的解释力通常优于单因子.本研究中的气候弹性指数(CE)正是利用残差法构建的用于代理非气候因素的综合指标,其与FSI的显著相关的像元比例为87.65%~97.36%,对FSI的贡献率为33.00%~47.00%,明显高于气温的或降水的.多元回归模型的R²为0.41~0.56,表明气温、降水和CE等3类因子能够在一定程度上解释乔木林生态系统稳定性的变化,但仍有44%~59%的未解释部分.这是因为CE仅是对非气候因素的近似度量,不足以完全涵盖植被结构、土壤环境、地形格局和人类活动等因素[72-73].因此,未来研究应纳入更多直接表征非气候因素的指标,以进一步提升模型对生态系统稳定性变异机制的解释能力. ...
Phenology-based residual trend analysis of MODIS-NDVI time series for assessing human-induced land degradation
1
2018
... 本研究结果显示,气温与降水对乔木林生态系统稳定性的独立解释力相对有限,其与FSI显著相关的像元比例分别为7.08%~42.50%和7.50%~28.13%,对FSI的贡献率仅为5.00%~13.00%和4.00%~10.00%,与已有研究结果一致,即单一气候因子往往难以充分解释生态系统过程的主要变异[68-70].Li等[71]的研究进一步显示,全球中高纬地区春季复合暖干(CWD)、温度和降水对夏季植被变异的显著相关面积分别为27.00%、15.00%和24.00%,也表明了复合指标的解释力通常优于单因子.本研究中的气候弹性指数(CE)正是利用残差法构建的用于代理非气候因素的综合指标,其与FSI的显著相关的像元比例为87.65%~97.36%,对FSI的贡献率为33.00%~47.00%,明显高于气温的或降水的.多元回归模型的R²为0.41~0.56,表明气温、降水和CE等3类因子能够在一定程度上解释乔木林生态系统稳定性的变化,但仍有44%~59%的未解释部分.这是因为CE仅是对非气候因素的近似度量,不足以完全涵盖植被结构、土壤环境、地形格局和人类活动等因素[72-73].因此,未来研究应纳入更多直接表征非气候因素的指标,以进一步提升模型对生态系统稳定性变异机制的解释能力. ...
Impacts of climate change and human activities on different degraded grassland based on NDVI
1
2022
... 本研究结果显示,气温与降水对乔木林生态系统稳定性的独立解释力相对有限,其与FSI显著相关的像元比例分别为7.08%~42.50%和7.50%~28.13%,对FSI的贡献率仅为5.00%~13.00%和4.00%~10.00%,与已有研究结果一致,即单一气候因子往往难以充分解释生态系统过程的主要变异[68-70].Li等[71]的研究进一步显示,全球中高纬地区春季复合暖干(CWD)、温度和降水对夏季植被变异的显著相关面积分别为27.00%、15.00%和24.00%,也表明了复合指标的解释力通常优于单因子.本研究中的气候弹性指数(CE)正是利用残差法构建的用于代理非气候因素的综合指标,其与FSI的显著相关的像元比例为87.65%~97.36%,对FSI的贡献率为33.00%~47.00%,明显高于气温的或降水的.多元回归模型的R²为0.41~0.56,表明气温、降水和CE等3类因子能够在一定程度上解释乔木林生态系统稳定性的变化,但仍有44%~59%的未解释部分.这是因为CE仅是对非气候因素的近似度量,不足以完全涵盖植被结构、土壤环境、地形格局和人类活动等因素[72-73].因此,未来研究应纳入更多直接表征非气候因素的指标,以进一步提升模型对生态系统稳定性变异机制的解释能力. ...




 甘公网安备 62010202000688号
甘公网安备 62010202000688号
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}