


0 引言
荒漠绿洲过渡带是维持绿洲稳定的关键,过渡带退化是绿洲环境退化的根本原因[1-2]。自20世纪80年代,为防止沙漠对绿洲的侵蚀,中国西北地区开展了一系列以人工固沙植被建设为主要措施的生态修复工程[3]。经过40~50 a的建设,原来以流动沙丘为主的沙漠景观演变为结构复杂的半人工半自然的植被生态系统,人工植被主要包括种植耐旱耐盐的乡土灌木如梭梭、柠条、花棒、油蒿等;自然植被主要包括天然入侵的草本植物,例如雾冰藜、小画眉草和白茎盐生草等一年生草本植物[4-5]。这些草本植物的逐步入侵与定居丰富了群落物种组成,推动了群落结构由单一向复杂演变。作为荒漠绿洲过渡带植被生态系统的重要组成部分,林下草本植物在维持生态稳定、调节能量与物质循环以及改善土壤等方面发挥着重要作用,促使生态环境向草原化荒漠的地带性方向发展[6-7]。
一年生植物是荒漠绿洲过渡带人工植被区建立后随着生境改变而天然入侵的,它们具有分布广、盖度大、物种多且逐年显示其优势性的特征。同时,在人工林生态系统林下它们对生境变化又极其敏感[8-9],物种组成、多样性和优势种更替频繁[10]。在短暂的生长时期内,一年生草本植物要经受沙漠地区严酷的自然环境胁迫,它们得以适应恶劣的生境在荒漠中生存、繁衍后代与它们独特的生理、个体生长以及种群和群落适应特征相关。因此,学者们对一年生草本植物的生理和生长特性[11-12]、群落结构[13-14]、物种多样性[15-16]及其与环境因子的关系[17-18]开展了大量研究。张景光等[19]在腾格里沙漠研究发现,一年生植物的覆盖度随着年降水量及其分配状况的变化而呈现出很大的波动,湿润年一年生草本植物覆盖度可以超过40%,而干旱年覆盖度一般只有20%;苏永中等[20]在科尔沁沙地对小叶锦鸡儿人工林下草本群落研究发现,建植初期(5 a)以一年生浅根型植物为主,随着土壤环境条件的改善,到28 a时多年生植物逐渐占据优势,林下草本层群落组成和结构趋于稳定。何文强等[21]在毛乌素沙地南缘对柠条固沙林林下草本群落的研究表明,林下草本植物的Shannon-Wiener和Simpson多样性指数均呈上升趋势,土壤水分和土壤pH对草本植物多样性有显著贡献。因此,不同地区人工林林下草本植物的演替过程、物种多样性及其驱动因素都存在巨大差异。然而,目前从人工林林下优势种更替和群落多样性与生境关系角度分析荒漠绿洲过渡带林下草本植物演变规律的研究仍未见报道。
河西走廊是中国西北主要的绿洲分布与强烈风沙活动重叠区,尤其是在荒漠绿洲过渡带,过度放牧和农田扩张导致大量天然植被受到破坏,风沙侵蚀给绿洲生态环境带来极大的威胁。梭梭(Haloxylon ammodendron)作为河西走廊荒漠绿洲过渡带生态防护林建设的先锋物种,具有耐旱、耐寒、抗风蚀、耐沙埋等特性[22],对维护荒漠绿洲过渡带生态系统的稳定性起到了至关重要的作用。目前关于河西走廊荒漠绿洲过渡带人工梭梭林的研究主要集中在林下土壤理化性质变化[23-24]、土壤微生物变化[25-26]、梭梭种群空间格局演变等[27]等方面。但对处于过渡带特殊环境背景下梭梭林下草本植物多样性、结构及数量动态方面的研究仍有不足,而这正是人工林下植物物种保护、种群和多样性恢复的基础。鉴于此,本研究以河西走廊荒漠绿洲过渡带不同林龄(流动沙丘0、5、10、20、30、40 a)梭梭人工林下草本植物为研究对象,通过调查不同林龄林下土壤理化性质以及草本植物物种组成、结构和多样性演变特征,拟解决以下两个科学问题:①种植40年梭梭林下土壤理化性质、林下草本植物优势种以及物种多样性如何演变?②在演变过程中,优势种更替的主要控制因素?研究结果可以为干旱荒漠地区植被恢复与生态工程建设提供理论依据和实践参考。
1 材料与方法
1.1 研究区概况
研究区位于黑河中游、河西走廊中段的甘肃省临泽县北部荒漠绿洲过渡带(39°20′—39°23′N、100°07′—100°08′E,海拔1 375 m)。属于典型的温带大陆性荒漠气候,年均气温7.6 ℃,年降水量117 mm,多在7—9月,年蒸发量2 390 mm,日照充足,年日照时数3 045 h。风向以西北为主,年均风速3.2 m·s-1,春季风沙活动强烈,最大风速可达21 m·s-1。地貌类型主要为风成地貌,有流动、半流动、半固定、固定沙丘等。其中流动沙丘以3~8 m的高度分布,形态多为新月形或链形。土壤类型以灰棕漠土和风沙土为主,质地粗糙,土壤较贫瘠。地带性植被主要有灌木、小灌木、半灌木等荒漠植物,通常伴有一年生草本植物如沙蓬(Agriophyllum squarrosum)、雾冰藜(Bassia dasyphylla)、白茎盐生草(Halogeton arachnoideus)等,以藜科、禾本科、菊科、柽柳科植物较为常见[28]。
1.2 样地设置与植物调查
于2015年和2023年6—9月进行了两次采样。采用“空间代时间”的方法,以流动沙丘为对照(0 a),选取不同林龄(5、10、20、30、40 a)的人工梭梭林,按林龄划分为5个采样区(A、B、C、D、E区)。每个林龄选择3个样点,样点大小为200 m×200 m,每个样点布设3个灌木样方,样方大小为25 m×25 m,样点位置均为平坦沙地,样方间的距离大于50 m,在A、B、C、D、E区分别布设了7、4、5、7个和5个1 m×1 m的样方,共计28个样方(图1),记录样方内物种数、密度、盖度等信息,地上生物量采用直接取样烘干法来完成。
图1
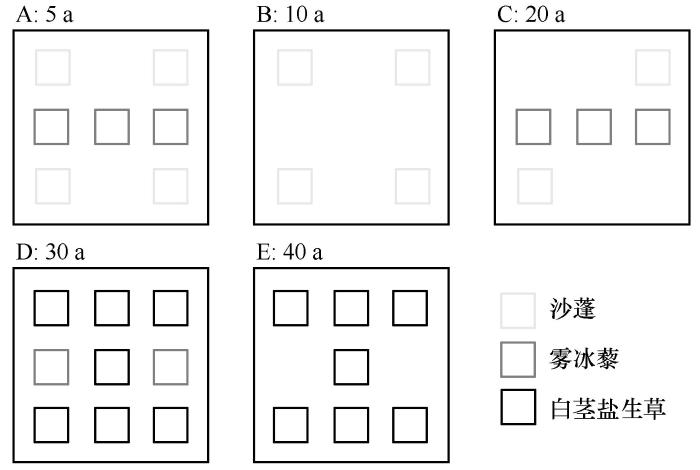
图1
不同林龄梭梭林3种优势一年生草本植物样区
Fig.1
Sample areas of three dominant annual herbaceous species of Haloxylon ammodendron woodland with different ages
1.3 土壤样品采集与测定
在每个灌木样方内随机采集0~10 cm和10~20 cm深度土样3个,混合后装入已编号的铝盒和自封袋中带回。为揭示优势种更替的机制,在环境梯度下A、B、C、D、E区的28个1 m×1 m样方内,分别采用多点混合法按梅花状采集0~10 cm土壤样品,样品同样分装保存,用于土壤理化性状的分析。土壤粒级分析采用湿筛加吸管法,土壤含水量采用烘干法测定,土壤有机质、有机碳含量采用重铬酸钾外加热法测定,土壤总碳(TC)含量采用元素分析仪(PE2400,Pekin EImer公司,美国)测定,利用总碳与有机碳的差值计算无机碳,土壤全氮含量凯氏定氮法测定,土壤全磷含量采用钼锑抗比色法测定,土壤全钾含量采用原子吸收法测定,土壤有效磷含量用碳酸氢钠浸提-钼锑抗比色法测定,土壤速效钾含量用醋酸铵浸提-火焰光度法测定,土壤水解性氮含量采用碱解扩散法测定。pH值采用1∶1的土水比悬液pH计测定,土壤电导率采用1∶5土水比浸提液测定,Ca2+和Na+采用EDTA络合滴定法测定,Cl-采用AgNO3滴定法测定,HCO
表1 不同林龄梭梭人工林土壤粒径及盐离子组成
Table 1
深度 /cm | 林龄 /a | 土壤粒径组成/% | 土壤盐离子含量/(g·kg-1) | |||||
|---|---|---|---|---|---|---|---|---|
| 砂粒(>0.05 mm) | 粉粒(0.002~0.05 mm) | 黏粒(<0.002 mm) | Na+ | Ca2+ | Cl- | HCO | ||
| 0~10 | CK(0) | 92.76±0.55ab | 3.56±0.31c | 3.70±0.27c | 0.01±0.00c | 0.03±0.00b | 0.28±0.06c | 0.17±0.00e |
| 5 | 92.20±0.79ab | 2.72±0.22c | 5.07±0.57b | 0.24±0.01a | 0.05±0.02b | 0.87±0.16b | 0.61±0.08b | |
| 10 | 93.86±0.09a | 2.87±0.26c | 3.27±0.17c | 0.21±0.02a | 0.05±0.01b | 0.78±0.09b | 0.44±0.02c | |
| 20 | 90.25±2.22bc | 5.86±1.53b | 3.89±0.69c | 0.25±0.03a | 0.04±0.01b | 0.69±0.04b | 0.72±0.01a | |
| 30 | 87.40±1.54bc | 6.70±0.33b | 5.91±0.20b | 0.13±0.02b | 0.10±0.02a | 0.87±0.17b | 0.31±0.04d | |
| 40 | 76.74±2.74d | 11.57±1.90a | 11.68±0.84a | 0.22±0.03a | 0.08±0.01a | 1.12±0.18a | 0.74±0.05a | |
| 10~20 | CK(0) | 94.90±0.44a | 1.84±0.34c | 3.25±0.28b | 0.04±0.02b | 0.03±0.00b | 0.26±0.15c | 0.18±0.01b |
| 5 | 93.10±0.50ab | 3.23±1.17bc | 3.67±1.57b | 0.20±0.07b | 0.05±0.02b | 0.57±0.20ab | 0.35±0.08ab | |
| 10 | 92.52±0.24b | 2.68±0.20bc | 4.80±0.31b | 0.08±0.04b | 0.07±0.02b | 0.41±0.07bc | 0.28±0.06ab | |
| 20 | 91.63±2.76b | 3.10±0.19bc | 4.64±3.73b | 0.21±0.05b | 0.07±0.01b | 0.56±0.18ab | 0.40±0.18a | |
| 30 | 91.56±0.11b | 3.73±1.79b | 5.34±0.11b | 0.16±0.07b | 0.06±0.01b | 0.43±0.17bc | 0.29±0.01ab | |
| 40 | 81.98±0.79c | 7.11±0.53a | 10.91±0.31a | 0.65±0.50a | 0.15±0.09a | 0.78±0.13a | 0.38±0.11a | |
1.4 数据处理与分析
1.4.1 林下草本植物多样性计算
根据野外调查数据计算林下草本植物的重要值(important value,IV)和多样性指数,植物多样性选用Margalef丰富度指数、Shannon-Wiener多样性指数、Simpson优势度指数、Pielou均匀性指数来进行评价。各指数计算公式如下[29]:
式中:RC为植物种的相对盖度;RD为相对密度;RF为相对频度。
Margalef丰富度指数(M):
式中:S为物种总数;N为调查样方内所有植物个体总数。
Shannon-Wiener多样性指数(H):
式中:Pi 为S个物种群落中第i个物种的个体数占所有物种总个体数的比例。
Simpson优势度指数(D):
式中:Pi 为S个物种群落中第i个物种的个体数占所有物种总个体数的比例。
Pielou均匀度指数(J):
1.4.2 高斯模型分析
为研究不同林龄3种优势一年生草本植物的生态阈值,利用Matlab14a软件进行曲线拟合和高斯模型回归,将3种优势一年生草本植物的生物量取自然对数后分别与水分、盐度进行一元二次函数拟合,确定得到的一元二次曲线方程是否符合高斯模型,再将一年生草本植物生物量分别与水分、盐度进行高斯模型回归。高斯模型方程[30]为:
式中:y为能够代表一年生草本植物生长状况的一个指标;c为对应指标的最大值;u为相应的生物指标达到最大值时对应的环境因子值,为该指标的耐度。植物的一般生态阈值为[u-2t,u+2t],最适生态阈值为[u-t,u+t]。
本研究采用Excel 2010对数据进行预处理与基础统计;利用SPSS 21.0软件对不同林龄梭梭人工林下土壤理化性质及草本植物物种多样性的差异进行单因素方差分析(One-Way ANOVA),显著性水平设定为P=0.05;基于R软件中“vegan”程序包,采用主坐标分析(Principal Coordinate Analysis,PCoA)和基于Bray-Curtis距离的多元方差分析(PERMANOVA,Adonis检验)比较不同林龄梭梭人工林植物群落组成的差异;利用Canoco5软件进行冗余分析(Redundancy analysis,RDA),探讨不同林龄人工林林下草本层物种多样性指数与土壤理化性质的关系;作图由Origin 2024软件完成。
2 结果与分析
2.1 不同林龄人工梭梭林土壤理化性质变化
不同林龄梭梭人工林土壤理化性质差异显著(图2,P<0.05)。随着林龄的增加,0~10 cm和10~20 cm土壤电导率和养分含量(有机质、全氮、全磷)总体呈显著升高趋势。在0~10 cm土层,土壤水分表现为先升后降的变化趋势,平均含水量由流动沙丘的0.25%逐渐增加,在30 a林龄时显著升高达到峰值1.28%,到40 a林龄时下降至0.92%。土壤pH值7.78~8.86,整体呈碱性,随林龄的增加波动变化。土壤电导率、有机质、全氮、全磷的含量在40 a林龄时达到最大值,分别为224.93 μS·cm-1、10.02 g·kg-1、0.86 g·kg-1、0.85 g·kg-1,显著高于流动沙丘时期。在10~20 cm土层,土壤水分和pH值随林龄的增加呈现先增加后减少再增加再减少的变化趋势。土壤水分含量为1.17%~3.43%,在5 a林龄时达到最大值。土壤pH值为8.06~8.51,仍维持碱性。电导率为44.45~478.19 μS·cm-1,普遍高于表层土层。而土壤养分(有机质、全氮和全磷)的含量均低于0~10 cm土层,但同样在40 a时达到峰值。
图2
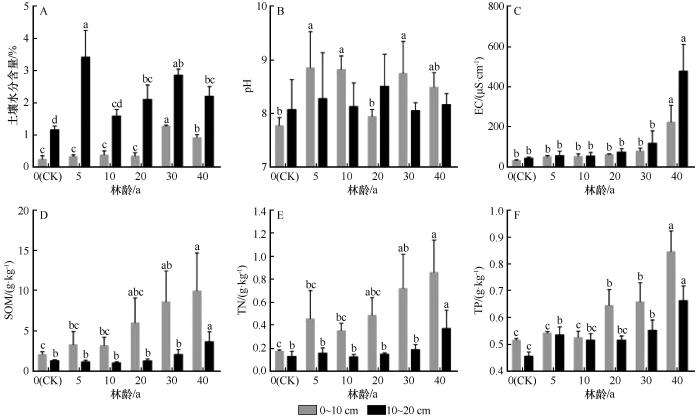
图2
不同林龄人工梭梭林土壤理化性质
Fig.2
Physicochemical properties of soil in artificial Haloxylon ammodendron forests with different ages
2.2 人工梭梭林下草本植物优势种变化
PCoA分析表明,不同林龄间植物群落组成存在显著差异(R2=0.487,P=0.001,图3)。PC1和PC2分别解释了群落变异的47.6%和21.7%,总解释率为69.3%,不同林龄样地沿PC1轴呈连续分布,反映出群落在演变过程中的渐变特征。依据样点分布特征及物种组成变化,将演变过程划分为3个阶段:早期(5~10 a)、中期(20~30 a)和后期(40 a)。
图3
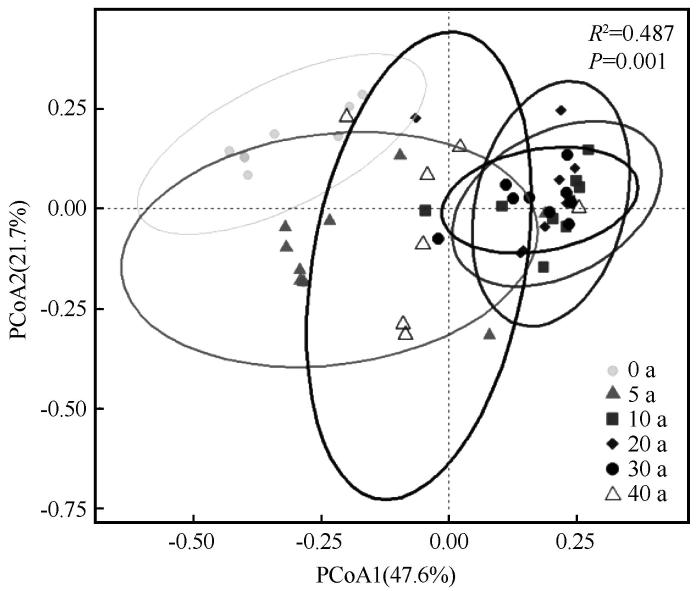
图3
梭梭林不同样地植物群落结构差异的PCoA分析
Fig.3
PCoA analysis of differences in plant community structure among sites of Haloxylonammodendron woodland
图4
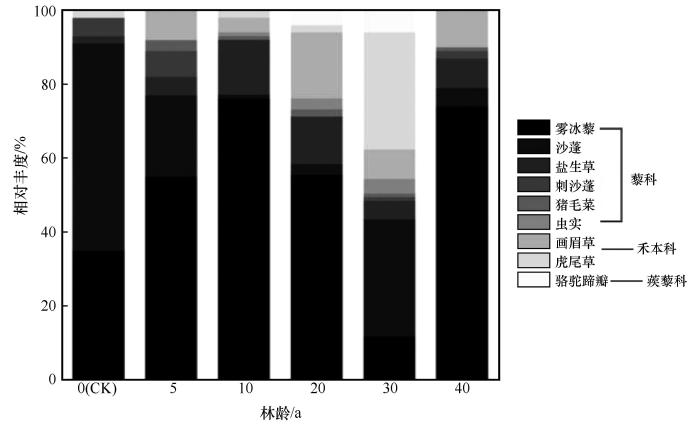
图4
不同林龄梭梭林下草本植物相对丰度
Fig.4
Relative abundance of understory herbaceous plants in Haloxylon ammodendron woodland of different ages
表2 不同林龄梭梭林下物种组成及其重要值
Table 2
| 物种名 | 生活型 | 林龄/a | |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 0(CK) | 5 | 10 | 20 | 30 | 40 | ||||||||||||||
| D | F | IV | D | F | IV | D | F | IV | D | F | IV | D | F | IV | D | F | IV | ||
| 泡泡刺(Nitraria sphaerocarpa) | S | 5.00 | 0.02 | 1.71 | — | — | — | — | — | — | — | — | — | — | — | — | — | — | — |
| 沙拐枣(Calligonum mongolicunl) | S | 3.81 | 0.69 | 8.55 | 2.00 | 0.04 | 1.88 | 1.00 | 0.03 | 0.85 | 1.00 | 0.07 | 1.35 | — | — | — | 3.60 | 0.17 | 4.47 |
| 梭梭(Haloxylon ammodendron) | S | — | — | — | 1.50 | 0.04 | 18.61 | — | — | 1.00 | 0.04 | 7.32 | — | — | — | — | |||
| 骆驼蹄瓣(Zygophyllum fabago) | P | — | — | — | — | — | — | — | — | — | 7.00 | 0.02 | 2.71 | 22.38 | 0.27 | 7.92 | — | — | — |
| 雾冰藜(Bassia dasyphylla) | A | 62.83a | 0.93a | 32.93a | 32.75a | 0.98a | 36.85a | 98.13a | 1.00a | 61.93a | 87.22a | 0.91a | 38.73a | 45.50b | 1.00a | 21.46a | 88.95a | 0.7a | 57.73a |
| 沙蓬(Agriophyllum Squarrosum) | A | 102.21a | 0.84a | 37.57a | 13.11a | 0.62a | 17.58a | 1.00b | 0.03b | 0.79b | 4.33b | 0.07b | 4.38b | 124.00a | 0.07b | 18.89a | 5.57b | 0.23b | 6.60b |
| 白茎盐生草(Halogeton arachnoideus) | A | 3.00b | 0.24b | 3.93b | 2.80b | 0.11b | 7.54b | 18.55a | 0.67a | 21.52a | 20.50a | 0.76a | 18.65a | 20.89b | 0.63a | 12.33b | 9.40b | 0.33a | 16.29a |
| 刺沙蓬(Salsola ruthenica) | A | 9.47b | 0.33b | 11.47b | 4.50b | 0.04b | 3.58b | — | — | — | — | — | — | 2.0b | 0.10b | 2.07b | 3.0a | 0.03b | 1.86b |
| 画眉草(Eragrostis pilosa) | A | — | — | — | 5.0b | 0.02b | 10.34b | 5.0b | 0.20b | 4.55b | 21.0a | 0.07b | 8.61b | 32.33b | 0.20b | 7.43b | 12.60b | 0.17b | 10.52b |
| 猪毛菜(Salsola collina) | A | — | — | — | 1.63b | 0.16b | 3.63b | 1.36b | 0.37b | 7.71b | 2.60b | 0.11b | 6.02b | 3.43b | 0.23b | 5.63b | 1.00b | 0.07b | 2.53b |
| 虎尾草(Chloris virgata) | A | 3.50b | 0.04b | 3.85b | — | — | — | 2.00b | 0.09b | 1.26b | 2.50b | 0.09b | 9.72b | 123.00a | 0.03b | 17.87a | — | — | — |
| 虫实(Corispermum macrocarpum) | A | — | — | — | — | — | — | 1.0b | 0.03b | 1.39b | 4.33b | 0.07b | 2.51b | 16.00b | 0.10b | 6.41b | — | — | — |
| 群落密度 | 189.82b | 63.28b | 128.05b | 151.19b | 389.53a | 124.12b | |||||||||||||
| 灌木层密度 | 8.81a | 3.50b | 1.00b | 2.00b | — | 3.60b | |||||||||||||
| 草本层密度 | 181.01b | 59.78b | 127.05b | 149.49b | 389.53a | 120.52b | |||||||||||||
| 灌木层物种数 | 2 | 2 | 1 | 2 | 0 | 1 | |||||||||||||
| 草本层物种数 | 5 | 6 | 7 | 8 | 9 | 7 | |||||||||||||
| 草本层优势种 | 沙蓬+雾冰藜 | 雾冰藜+沙蓬 | 雾冰藜+白茎盐生草 | 雾冰藜+白茎盐生草 | 雾冰藜+沙蓬 | 雾冰藜+白茎盐生草 | |||||||||||||
2.3 林下草本植物物种多样性
梭梭林不同林龄之间林下草本植物的多样性存在显著差异(P<0.05,图5)。Margalef丰富度、Shannon-Wiener多样性、Simpson优势度和Pielou均匀度均表现出相似的变化规律,即随演变阶段的推进呈现先增加后减少的趋势(图6)。在早期,林下草本植物的Margalef物种丰富度指数较流动沙丘有所上升,10 a林龄时的物种丰富度是流动沙丘时(0 a,0.387)的1.53倍。Shannon-Wiener多样性、Simpson优势度和Pielou均匀度表现为先增加后减少的趋势。进入中期,物种多样性指数呈一定的上升趋势。Margalef丰富度、Shannon-Wiener多样性、Simpson优势度和Pielou均匀度均在30 a达到最大值,分别高于流动沙丘时的52.75%、50.81%、47.03%和19.23%。到后期,林下草本植物的物种多样性有所降低。
图5
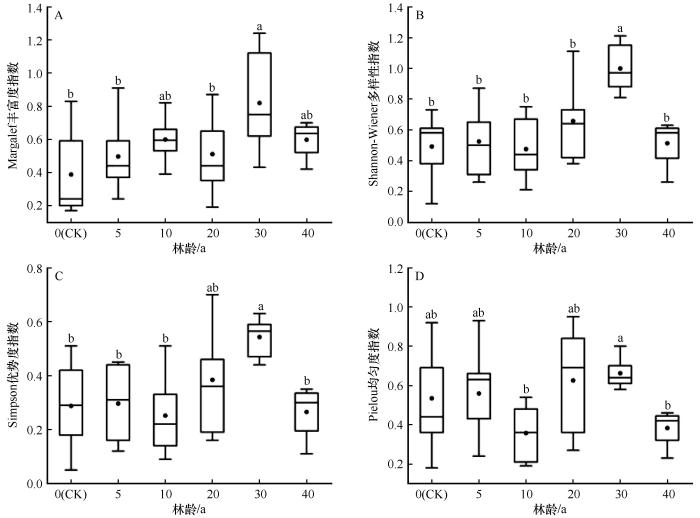
图5
不同林龄梭梭林下草本植物多样性指数
Fig.5
Understory herbaceous plant diversity in Haloxylon ammodendron woodland of different ages
图6
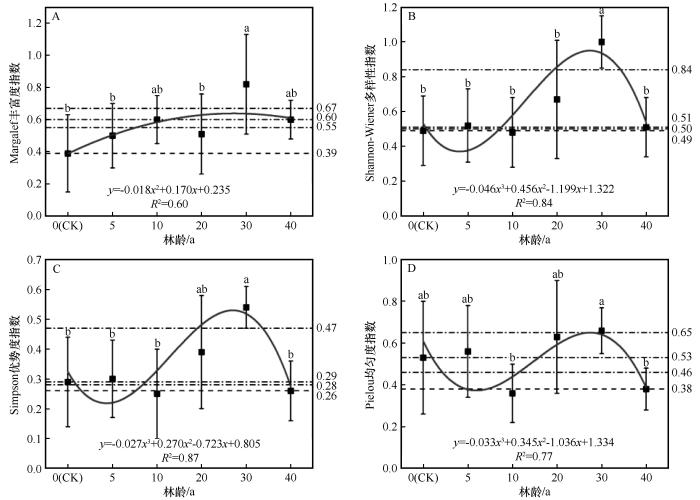
图6
不同林龄梭梭林下草本植物多样性指数变化趋势
Fig.6
Changes in herbaceous plant diversity index under Haloxylon ammodendron woodland with different ages
2.4 不同林龄3种优势一年生草本植物的生态阈值
2.4.1 3种一年生优势植物生物指标间的相关性
3种一年草本植物生物指标间的相关关系如表3所列,一年生草本植物的密度、地上生物量、地下生物量以及总生物量间呈现极显著的正相关关系,所以选用生物量指标分析荒漠绿洲过渡带人工梭梭林林下3种优势一年生草本植物生长最适土壤理化指标的生态阈值。
表3 人工梭梭林下3种优势一年生草本植物生物指标之间的相关性分析
Table 3
| 密度 | 地上生物量 | 地下生物量 | 总生物量 | |
|---|---|---|---|---|
| 密度 | 1 | |||
| 地上生物量 | 0.517** | 1 | ||
| 地下生物量 | 0.641** | 0.908** | 1 | |
| 总生物量 | 0.538** | 0.999** | 0.929** | 1 |
2.4.2 优势一年生草本植物与土壤理化指标之间的相关性
表4 人工梭梭林下3种优势一年生草本植物生物量与土壤理化指标之间的相关关系
Table 4
| 物种 | 土壤理化指标(x) | 拟合曲线方程 | R2 | P |
|---|---|---|---|---|
| 沙蓬 | 电导率 | y=-0.002x2+0.336x-13.049 | 0.589 | 0.045 |
| 含水量 | y=-23.924x2+39.424x-14.142 | 0.637 | 0.029 | |
| pH | y=1.07x-6.994 | 0.018 | 0.715 | |
| 无机碳 | y=-1.257x2+10.198x-19.011 | 0.332 | 0.244 | |
| 有机碳 | y=-0.131x2+1.188x-1.293 | 0.175 | 0.510 | |
| 全氮 | y=10.91x2-10.637x+3.424 | 0.358 | 0.212 | |
| 全磷 | y=598.543x2-314.178x+41.986 | 0.149 | 0.568 | |
| 全钾 | y=0.192x2-6.799x+61.127 | 0.033 | 0.889 | |
| 水解性氮 | y=0.001x2+0.020x-0.083 | 0.317 | 0.264 | |
| 有效磷 | y=0.008x2-0.238x+2.310 | 0.204 | 0.451 | |
| 速效钾 | y=0.003x2-0.520x+25.418 | 0.270 | 0.332 | |
| Cl- | y=-0.002x2+0.556x-39.621 | 0.200 | 0.458 | |
| HCO | y=-0.065x+5.815 | 0.098 | 0.698 | |
| Na+ | y=0.00005667x2-0.009x+1.383 | 0.005 | 0.982 | |
| Ca2+ | y=-0.042x2+0.521x-0.444 | 0.001 | 0.996 | |
| 雾冰藜 | 电导率 | y=-0.001x2+0.174x-6.967 | 0.622 | 0.033 |
| 含水量 | y=-3.101x2+5.13x-1.002 | 0.613 | 0.036 | |
| pH | y=0.598x-3.832 | 0.008 | 0.817 | |
| 无机碳 | y=0.987x2-8.813x+20.069 | 0.736 | 0.018 | |
| 有机碳 | y=0.963x2-5.892x+9.399 | 0.768 | 0.012 | |
| 全氮 | y=-5.467x2+3.755x+0.264 | 0.170 | 0.571 | |
| 全磷 | y=-33.807x2+29.221x-4.956 | 0.302 | 0.341 | |
| 全钾 | y=0.061x2-2.432x+24.419 | 0.716 | 0.023 | |
| 水解性氮 | y=0.001x2+0.009x+0.408 | 0.218 | 0.478 | |
| 有效磷 | y=-0.011x2+0.238x-0.185 | 0.331 | 0.300 | |
| 速效钾 | y=0.092x-4.532 | 0.241 | 0.437 | |
| Cl- | y=0.036x-0.699 | 0.265 | 0.397 | |
| HCO | y=-0.036x+2.57 | 0.286 | 0.363 | |
| Na+ | y=-0.038x+2.434 | 0.508 | 0.119 | |
| Ca2+ | y=-0.026x2+0.166x+0.638 | 0.198 | 0.516 | |
| 白茎盐生草 | 电导率 | y=-0.001x2+0.160x-8.201 | 0.680 | 0.019 |
| 含水量 | y=-9.129x2+15.328x-5.229 | 0.584 | 0.046 | |
| pH | y=-3.281x+25.425 | 0.225 | 0.197 | |
| 无机碳 | y=1.543x2-13.975x+32.159 | 0.506 | 0.121 | |
| 有机碳 | y=-0.133x2+0.943x-0.789 | 0.041 | 0.882 | |
| 全氮 | y=-6.963x2+5.108x-0.053 | 0.028 | 0.919 | |
| 全磷 | y=303.944x2-177.117x+26.423 | 0.100 | 0.729 | |
| 全钾 | y=-0.244x2+8.774x-78.036 | 0.232 | 0.453 | |
| 水解性氮 | y=0.003x2-0.125x+1.916 | 0.029 | 0.915 | |
| 有效磷 | y=0.007x2-0.164x+1.14 | 0.404 | 0.212 | |
| 速效钾 | y=0.027x-0.352 | 0.127 | 0.665 | |
| Cl- | y=-0.00004078x2-0.01x+3.366 | 0.229 | 0.458 | |
| HCO | y=-0.084x+4.098 | 0.429 | 0.186 | |
| Na+ | y=-0.00008733x2+0.023x-0.405 | 0.196 | 0.519 | |
| Ca2+ | y=0.029x2-0.723x+4.911 | 0.140 | 0.637 |
图7
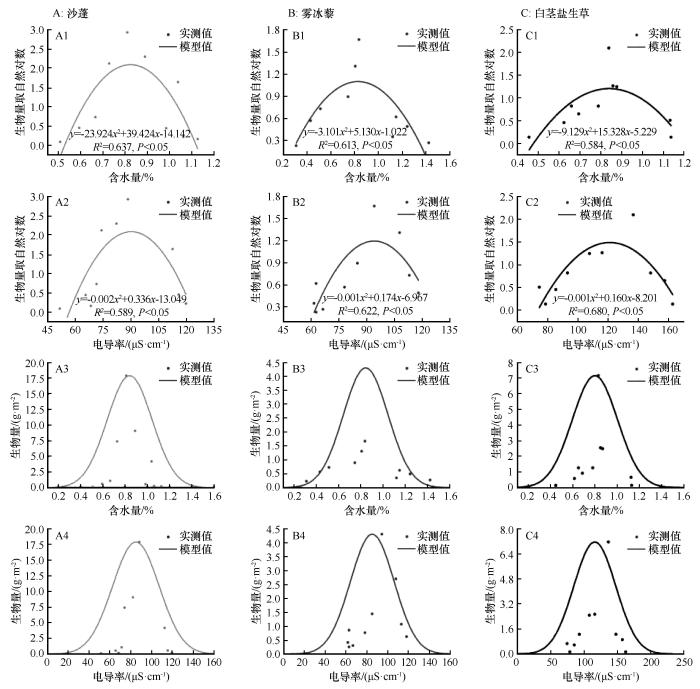
图7
3种优势一年生草本植物水盐响应及模型拟合结果
Fig.7
Water-salt response and model fitting results for three dominant annual herbaceous plants
2.4.3 优势一年生草本植物的土壤水盐生态阈值
土壤水分、电导率过高或过低都会抑制一年生草本植物的生长。根据拟合的一元二次曲线与高斯模型之间的转换关系,可以解出高斯回归方程(表5)。
表5 3种优势一年生草本植物的土壤水盐生态阈值高斯回归方程
Table 5
| 物种 | 拟合结果 | |
|---|---|---|
| 土壤含水量 | 土壤电导率 | |
| 沙蓬 | ||
| 雾冰藜 | ||
| 白茎盐生草 | ||
沙蓬对土壤含水量的最适值u为0.841%,其生态阈值为[0.437%,1.245%],最适生态阈值为[0.639%,1.043%]。当土壤含水量低于0.437%或高于1.245%时,沙蓬的生长将受到显著限制。雾冰藜的最适含水量u为0.856%,生态阈值为[0.153%,1.560%],最适生态阈值为[0.504%,1.201%]。当土壤含水量低于0.153%或高于1.560%时,其生长明显受限。白茎盐生草的最适含水量u为0.807%,生态阈值为[0.399%,1.214%],最适生态阈值为[0.603%,1.010%]。当土壤含水量低于0.399%或高于1.214%时,其生长同样受到限制。3种优势一年生草本植物对土壤含水量的生长最适值依次为:雾冰藜(0.856%)>沙蓬(0.841%)>白茎盐生草(0.807%,图7、图8)。沙蓬对土壤电导率的最适值为84.975 μS·cm-1,生态阈值为[40.279,129.671] μS·cm-1,最适生态阈值为[62.627,107.323] μS·cm-1。雾冰藜的最适电导率为84.967 μS·cm-1,生态阈值为[42.995,126.939] μS·cm-1,最适生态阈值为[63.981,105.953] μS·cm-1。白茎盐生草的最适电导率为116.067 μS·cm-1,其生态阈值为[53.003,179.131] μS·cm-1,最适生态阈值为[84.535,147.599] μS·cm-1。3种优势一年生草本植物对土壤电导率的生长最适值依次为:白茎盐生草(116.067 μS·cm-1)>沙蓬(84.975 μS·cm-1)>雾冰藜(84.967 μS·cm-1)。模型模拟值与实测生物量的比较结果表明,基于模型推导的土壤水分与电导率生态阈值能够有效表征3种优势一年生草本植物在自然生境下的最适生长条件与生态幅,这从生态学角度验证了水盐阈值模型对荒漠地区一年生草本植物水分适应机制的解释效力。
图8
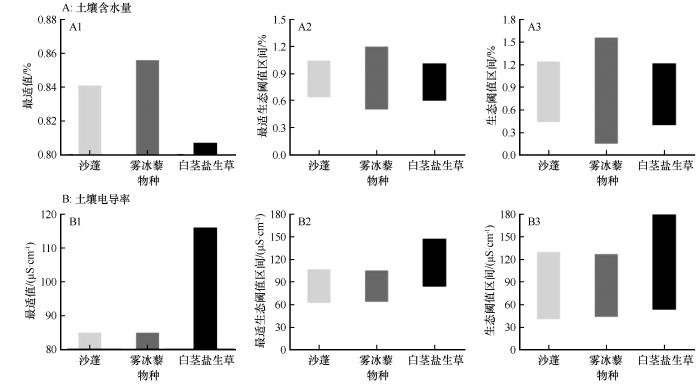
图8
3种优势一年生草本植物水盐生态阈值
Fig.8
Water-salt ecological thresholds of three dominant annual herbaceous plants
2.5 林下草本植物多样性与土壤环境因子的关系
人工梭梭林林下草本植物多样性指数与土壤环境因子之间存在显著相关性(图9)。Margalef丰富度与pH显著正相关,与HCO
图9
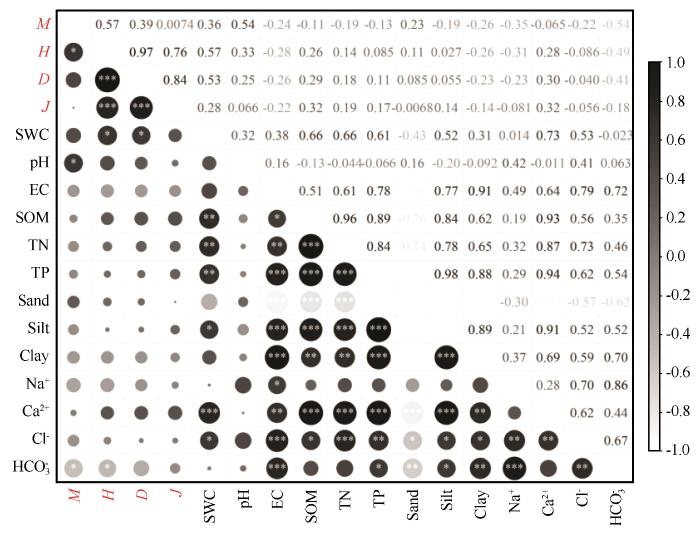
图9
林下草本植物多样性指数与土壤环境因子相关性分析
Fig.9
Correlation analysis between diversity indices of understory herbaceous plants and soil environmental factors
图10
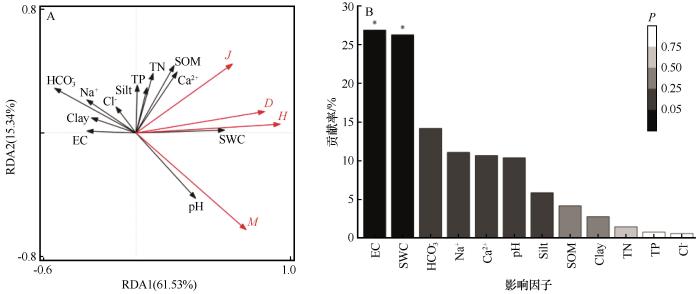
图10
林下草本物种多样性与土壤环境因子冗余分析RDA排序
Fig.10
RDA ordination plot of species diversity in understory vegetation and soil environmental factors
3 讨论
3.1 梭梭人工林下草本植物的演替特征
本研究发现,种植梭梭40 a,林下一年生草本层植物优势类群呈现出由耐旱性向耐盐性转变的趋势。按照优势种和土壤理化性质特征,我们将梭梭林下草本层群落演替划分为3个阶段:一年生草本植物入侵与定居阶段(5~10 a),种群扩张阶段(20~30 a)和优势种竞争与排斥(40 a)阶段。在流动沙丘,风沙活动极为强烈,地表裸露,土壤质地疏松,水分极易蒸发流失。在如此严酷的生境条件下,仅有少数具有耐旱、耐瘠薄特性的先锋植物在丘间低地等微凹地带存活[31],主要由沙蓬、雾冰藜和刺沙蓬等耐风沙一年生草本植物组成,其中沙蓬是一年生草本层的优势种,其重要值为37.57。而在种植梭梭后,地表风蚀得到有效抑制,土壤水分和养分含量明显提高,大量种子和细小颗粒物在人工林下沉积[23],一年生草本植物开始入侵与定居,林下草本植物的Margalef丰富度指数从0.387上升到0.598,而雾冰藜的重要值升为36.85,沙蓬重要值降低到17.58,雾冰藜成为群落的优势种。同时大量枯死的一年生植物形成微型风障加之地形因素,使萌发较晚的幼苗在其保护下能够成活,并形成一小片植丛。在林下草本植物种群扩张阶段,随着定居时间的推移,由最初单株或几株植物形成的植丛不断延伸和扩大,一些幼苗在植丛边缘地带不断拓展其分布范围,使草本植物群落的分布面积扩展。同时,土壤环境条件逐渐稳定,尤其是0~10 cm土层水分和养分含量在30 a时显著增加,土壤水分比流动沙丘土壤水分增加4.18倍,土壤养分含量分别增加3.06、3.10倍和0.28倍,土壤肥力水平提高为其他一年生草本植物侵入与定居创造了条件。林下草本植物的多样性进一步增加,小画眉草、猪毛菜和虎尾草等物种大量分布,群落进入到竞争与排斥(40 a)阶段。在这一阶段,白茎盐生草在林下分布范围不断扩张,成为新的优势物种,其重要值升高到16.29,并与沙生先锋植物(例如沙蓬)对光照、水分、营养元素等资源展开竞争,沙生植物存在范围被边缘化(沙蓬重要值降至6.60)且多样性也有所下降,因此导致一年生草本层植物物种多样性指数也随之下降。
3.2 梭梭人工林下草本层优势种的生态阈值
本研究发现,梭梭林下草本层植物优势种由流动沙丘沙蓬向半固定沙丘(种植梭梭20~30 a)雾冰藜和固定沙丘(40 a)白茎盐生草转变(图11)。在沙丘生境,土壤水分是植物生长、繁殖和自我更新的主要限制因素[32-34]。本研究发现沙蓬在流动沙丘分布范围最广,而雾冰藜和白茎盐生草对土壤含水量要求较高,因此多数出现在半固定和固定沙丘。在种植梭梭后,流动沙丘固定过程土壤-植物系统向地带性方向演变,土壤水分和养分大量表聚引起植物系统向浅层化发生。同时,梭梭作为稀盐盐生植物,从土壤和地下水中吸收大量盐分,其枯落物分解会将盐分重新释放回土壤,导致土壤盐分逐步在表层积聚[35]。土壤盐分积聚导致降水入渗减少,浅层土壤水分降低,梭梭对深层水分50~150 cm的利用增加,最终形成深层土壤干旱层[36]。因此,在种植梭梭40 a,土壤电导率显著上升至224.93 μS·cm-1,含水量稳定在0.92%左右。此时土壤电导率远超沙蓬与雾冰藜的生态阈值上限,而接近白茎盐生草的阈值区间,白茎盐生草逐渐取代沙蓬成为后期优势种。总体来看,林龄增长引起的土壤水分升高与盐分积累是驱动梭梭人工林下草本植物由耐旱性向耐盐性演变的关键生态机制。
图11
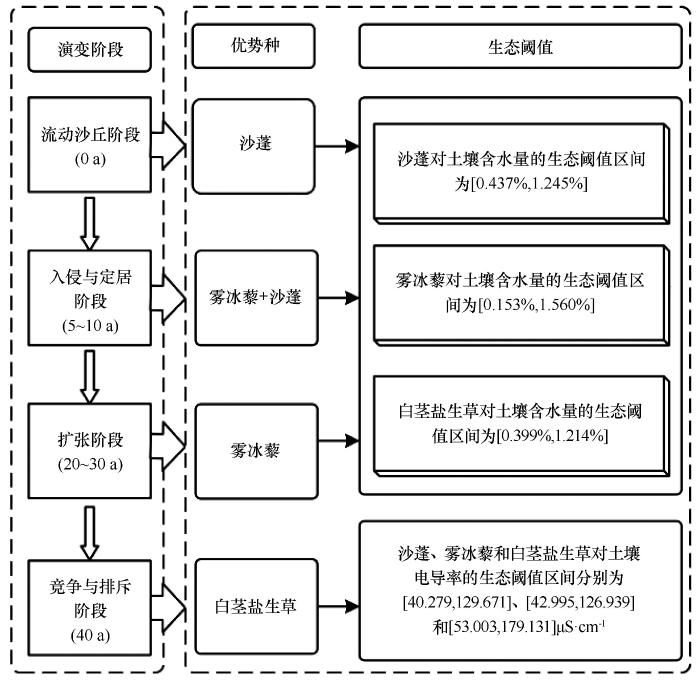
图11
不同演变阶段优势种概念图
Fig.11
Conceptual diagram of dominant species in different evolutionary stages
3.3 林下植物多样性与土壤环境因子的关系
本研究发现,随着梭梭种植年限增加,林下草本植物多样性呈先增加后减少的趋势。这主要是由于梭梭的种植使得原来质地相对均一的流沙发生资源分布空间异质性转变,向梭梭根系附近集聚,促进了土壤成土过程,为沙地一年生植物入侵和定居提供条件。一年生植物通常属于机会主义者,具备较强的资源获取与利用能力,能够在生态恢复初期凭借较高的水分和养分利用效率迅速建立种群并获得竞争优势[11,37-38]。随着林龄的增加,Margalef丰富度、Shannon-Wiener多样性、Simpson优势度及Pielou均匀度指数均在30 a林龄时达到最大值,分别为0.818、0.998、0.543及0.662。而在后期阶段,群落物种组成和多样性指数下降至0.596、0.513、0.265及0.383,这可能与土壤表层黏粉粒含量增加(表1)和盐结皮发生有关。一方面,细小颗粒物沉积导致一个以粉沙和细沙为主的结皮层形成,土体构型发生明显的变化,这使梭梭林土壤-植被系统趋于一个更薄的活动层,土壤水分入渗能力下降,同时也使草本植物种子难以进入土壤积累与保存[39];而盐分的累积也对林下一些非盐生植物的萌发、生长和繁殖产生抑制效应[40]。因此,在梭梭人工林种植40 a后,林下草本层多样性出现波动,其群落稳定性和演变过程仍需要进行更长时间尺度的观测。
4 结论
梭梭林下草本层群落演替划分为3个阶段:一年生草本植物入侵与定居阶段(5~10 a),种群扩张阶段(20~30 a)和优势种竞争与排斥(40 a)阶段。梭梭林下草本植物优势种由流动沙丘(0 a)沙蓬向早期(5~10 a)雾冰藜沙蓬转变,而中期(20~30 a)又由雾冰藜向后期(40 a)白茎盐生草演变。早期和中期优势种表现出较强的耐旱性,后期优势种表现出较强的耐盐性。林下草本植物物种多样性呈先升后降规律,土壤电导率和含水量是影响林下草本植物多样性变化的关键因子。
参考文献
Assessing the effects of China's three-north shelter forest program over 40 years
[J].
Drivers and impacts of changes in China's drylands
[J].
Contrasting patterns of fine-scale herb layer species composition in temperate forests
[J].
Redefining ecosystem multifunctionality
[J].
Effect of soil environment on species diversity of desert plant communities
[J].
荒漠绿洲过渡带梭梭(Haloxylon ammodendron)林建植对表层土壤碳氮磷化学计量特征及储量的长期影响
[J].
Succession of soil bacterial community along a 46-year choronsequence artificial revegetation in an arid oasis-desert ecotone
[J].
Ecological effects of desertification control and desertified land reclamation in an oasis-desert ecotone in an arid region:a case study in Hexi Corridor,Northwest China
[J].
植物干旱胁迫下水分代谢、碳饥饿与死亡机理
[J].
Root biomass distribution of planted Haloxylon ammodendron in a duplex soil in an oasis:desert boundary area
[J].
Effects of desert shrubs on fine-scale spatial patterns of understory vegetation in dry-land
[J].

 甘公网安备 62010202000688号
甘公网安备 62010202000688号
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}