Nebkhas play important roles in desertification control and biodiversity protection in arid and semi-arid regions of China
2
2020
... 荒漠生态系统是全球陆地生态系统的重要组成部分,约占地球陆地面积的35%,其典型特征包括极度水分匮乏、土壤贫瘠、盐渍化以及强烈的风沙活动等,这些极端条件对植物的生长和生存构成了严峻挑战[1 -2 ] .生活于此环境的沙生灌木在长期适应过程中,逐渐成为了荒漠生态系统的主要植被类型,并演化出了一系列独特的生存策略来适应和克服水分和养分的限制,其中根系超强的可塑性是其适应极端生境的核心策略[3 ] .因此,研究沙生灌木的根系适应性为理解干旱区地下生态过程提供了关键视角[4 ] . ...
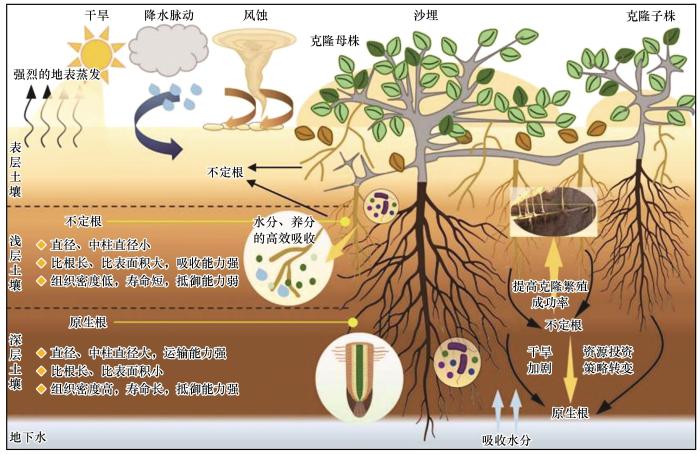
... 荒漠生态系统中强烈的风沙活动是驱动植物根系生长和发育的主要因素,当风沙流受植物阻拦时,沙物质在植物周围堆积,形成干旱-半干旱荒漠区常见的地貌类型——灌丛沙堆[1 ,56 -57 ] .这个过程使灌丛植被遭受一定程度的沙埋胁迫.不定根的生长可增强植物对浅层水分和养分的吸收能力以应对短期沙埋,维持植物的生理功能[53 ,58 ] .在沙埋干扰下,沙埋界面频繁的风沙活动及沙粒跃迁、撞击和摩擦,可使植物将资源重新分配至土壤表层的不定根和垂直茎上.不定根细胞排列更加紧密,木质化程度提高,从而增强了抗磨损能力和寿命,形成对干旱环境的适应性策略[8 ] .不定根的发育是沙生灌木适应风沙生境的重要策略,能够很好地缓解沙埋导致的资源胁迫. ...
Effect of nitrogen application on the sensitivity of desert shrub community productivity to precipitation in Central Asia
1
2022
... 荒漠生态系统是全球陆地生态系统的重要组成部分,约占地球陆地面积的35%,其典型特征包括极度水分匮乏、土壤贫瘠、盐渍化以及强烈的风沙活动等,这些极端条件对植物的生长和生存构成了严峻挑战[1 -2 ] .生活于此环境的沙生灌木在长期适应过程中,逐渐成为了荒漠生态系统的主要植被类型,并演化出了一系列独特的生存策略来适应和克服水分和养分的限制,其中根系超强的可塑性是其适应极端生境的核心策略[3 ] .因此,研究沙生灌木的根系适应性为理解干旱区地下生态过程提供了关键视角[4 ] . ...
Harnessing root architecture to address global challenges
1
2022
... 荒漠生态系统是全球陆地生态系统的重要组成部分,约占地球陆地面积的35%,其典型特征包括极度水分匮乏、土壤贫瘠、盐渍化以及强烈的风沙活动等,这些极端条件对植物的生长和生存构成了严峻挑战[1 -2 ] .生活于此环境的沙生灌木在长期适应过程中,逐渐成为了荒漠生态系统的主要植被类型,并演化出了一系列独特的生存策略来适应和克服水分和养分的限制,其中根系超强的可塑性是其适应极端生境的核心策略[3 ] .因此,研究沙生灌木的根系适应性为理解干旱区地下生态过程提供了关键视角[4 ] . ...
Patterns of root architecture adaptation of a phreatophytic perennial desert plant in a hyperarid desert
1
2013
... 荒漠生态系统是全球陆地生态系统的重要组成部分,约占地球陆地面积的35%,其典型特征包括极度水分匮乏、土壤贫瘠、盐渍化以及强烈的风沙活动等,这些极端条件对植物的生长和生存构成了严峻挑战[1 -2 ] .生活于此环境的沙生灌木在长期适应过程中,逐渐成为了荒漠生态系统的主要植被类型,并演化出了一系列独特的生存策略来适应和克服水分和养分的限制,其中根系超强的可塑性是其适应极端生境的核心策略[3 ] .因此,研究沙生灌木的根系适应性为理解干旱区地下生态过程提供了关键视角[4 ] . ...
The physiology of adventitious roots
4
2016
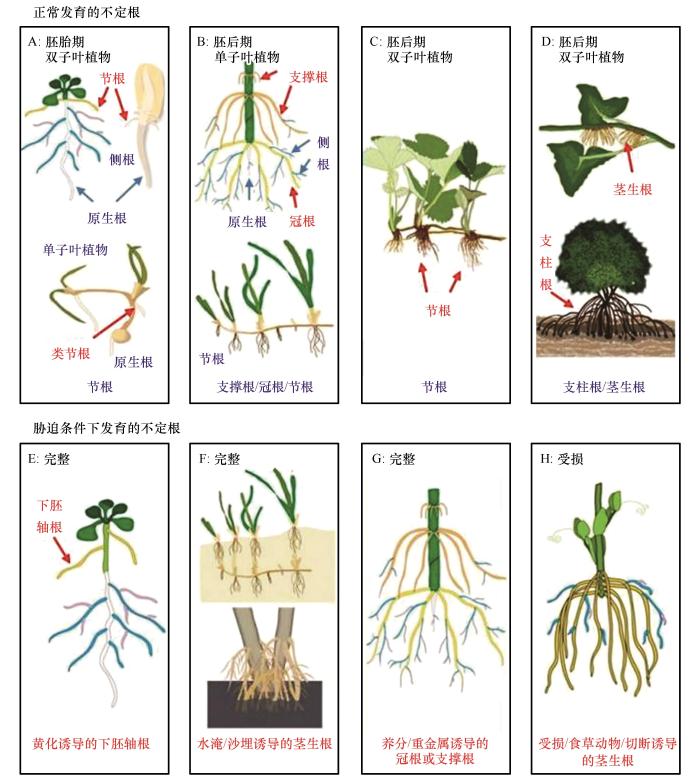
... 不定根(Adventitious roots)是由植物非根组织发育的根系,通常在植物的正常发育过程中(谷物的冠根和草莓的节根)或者在胁迫条件下(水淹、机械伤害、水分及养分胁迫等)产生[5 ] (图1 ).原生根(Initial roots)由植物胚胎根发育而来,不定根和原生根共同组成了植物的根系系统,保证了植物的固定及资源吸收利用[5 -6 ] .许多荒漠灌木在遭遇沙埋、损伤等干扰后,能从被埋的茎干上产生大量的不定根以应对各种复杂的环境[7 -8 ] .我们前期的大量野外调查也发现,中国西北荒漠区常见的白刺(Nitraria tangutorum )、柠条锦鸡儿(Caragana korshinskii )、霸王(Zygophyllum xanthoxylum )、红砂(Reaumuria songarica )、沙拐枣(Calligonum mongolicum )和柽柳(Tamarix chinensis )等沙生植物在沙埋后均可发育大量不定根(图2 ).这些不定根在缓解植物水分亏缺、增强养分捕获及促进克隆分株定植等方面发挥着关键作用[9 -11 ] .在干旱和半干旱地区,荒漠植物根系这种特有的生长和发育模式是其长期适应地带性水热条件的结果,也是荒漠植物生长发育及群落演变的关键,其构型和功能直接决定了植物对地下资源的获取能力,进而影响植物的生长发育、竞争能力及环境适应性[12 ] . ...
... [5 -6 ].许多荒漠灌木在遭遇沙埋、损伤等干扰后,能从被埋的茎干上产生大量的不定根以应对各种复杂的环境[7 -8 ] .我们前期的大量野外调查也发现,中国西北荒漠区常见的白刺(Nitraria tangutorum )、柠条锦鸡儿(Caragana korshinskii )、霸王(Zygophyllum xanthoxylum )、红砂(Reaumuria songarica )、沙拐枣(Calligonum mongolicum )和柽柳(Tamarix chinensis )等沙生植物在沙埋后均可发育大量不定根(图2 ).这些不定根在缓解植物水分亏缺、增强养分捕获及促进克隆分株定植等方面发挥着关键作用[9 -11 ] .在干旱和半干旱地区,荒漠植物根系这种特有的生长和发育模式是其长期适应地带性水热条件的结果,也是荒漠植物生长发育及群落演变的关键,其构型和功能直接决定了植物对地下资源的获取能力,进而影响植物的生长发育、竞争能力及环境适应性[12 ] . ...
... [
5 ]
Schematic diagram illustrating adventitious roots formation<sup>[<xref ref-type="bibr" rid="R5">5</xref>]</sup> Fig.1 ![]()
图2 中国北方主要沙区常见沙生灌木被埋茎上发育的不定根 Adventitious roots developed on buried stems of common sand-fixing shrubs in major sandy regions of Northern China Fig.2 ![]()
以往关于植物根系的研究,主要涉及原生根的分布格局、构型特征、水分吸收来源及其对资源异质性的响应[13 -14 ] .此外,干旱区与非干旱区植物的根系生态策略及功能存在显著分异.在干旱区,强烈的蒸发与贫瘠的土壤促使植物将更多碳资源分配给根系,特别是通过发展不定根来增强水分吸收和运输能力,以应对干旱、养分匮乏及沙埋等胁迫.相比之下,非干旱区水分供应稳定,植物倾向于将光合产物优先分配至地上部分,以促进生长与繁殖,其不定根主要承担营养繁殖的功能.大量研究揭示了荒漠植物通过发展深根系以利用深层土壤水或地下水[15 -16 ] ,或通过调整根系构型(如由叉状分支转向鱼尾形分支)来应对水分和养分的空间异质性[17 -18 ] .例如,在河西走廊戈壁区和风沙区,红砂和白刺两种荒漠植物根系分支结构复杂,次级分支较多,根系分支均接近叉状分支模式,但随着降水量逐年减少,红砂根系构型向鱼尾形分支模式发展,以降低根系内部对营养物质的竞争,提高根系对贫瘠土壤水分和养分的吸收效率,从而适应干旱贫瘠的土壤环境[19 ] .而白刺主要增加侧根分支数目、根长及根系连接长度,加强扩展能力来应对干旱环境[20 ] .可见,不同物种根系构型及对资源异质性的响应策略是不同的.不同物种根系水分利用策略也有不同,沙拐枣根系吸收水分主要来源于深层土壤水(约26%)和地下水(超过30%),而白刺由于其二态性根系结构特征[21 ] ,在强降水事件后根系主要利用浅层土壤水分(超过40%),降水稀少时则依靠地下水.在浅层土壤中,白刺根系生物量大于沙拐枣,在深层土壤中则相反,说明二者根系具有不同的水分利用策略[22 ] . ...
... [
5 ]
Fig.1 ![]()
图2 中国北方主要沙区常见沙生灌木被埋茎上发育的不定根 Adventitious roots developed on buried stems of common sand-fixing shrubs in major sandy regions of Northern China Fig.2 ![]()
以往关于植物根系的研究,主要涉及原生根的分布格局、构型特征、水分吸收来源及其对资源异质性的响应[13 -14 ] .此外,干旱区与非干旱区植物的根系生态策略及功能存在显著分异.在干旱区,强烈的蒸发与贫瘠的土壤促使植物将更多碳资源分配给根系,特别是通过发展不定根来增强水分吸收和运输能力,以应对干旱、养分匮乏及沙埋等胁迫.相比之下,非干旱区水分供应稳定,植物倾向于将光合产物优先分配至地上部分,以促进生长与繁殖,其不定根主要承担营养繁殖的功能.大量研究揭示了荒漠植物通过发展深根系以利用深层土壤水或地下水[15 -16 ] ,或通过调整根系构型(如由叉状分支转向鱼尾形分支)来应对水分和养分的空间异质性[17 -18 ] .例如,在河西走廊戈壁区和风沙区,红砂和白刺两种荒漠植物根系分支结构复杂,次级分支较多,根系分支均接近叉状分支模式,但随着降水量逐年减少,红砂根系构型向鱼尾形分支模式发展,以降低根系内部对营养物质的竞争,提高根系对贫瘠土壤水分和养分的吸收效率,从而适应干旱贫瘠的土壤环境[19 ] .而白刺主要增加侧根分支数目、根长及根系连接长度,加强扩展能力来应对干旱环境[20 ] .可见,不同物种根系构型及对资源异质性的响应策略是不同的.不同物种根系水分利用策略也有不同,沙拐枣根系吸收水分主要来源于深层土壤水(约26%)和地下水(超过30%),而白刺由于其二态性根系结构特征[21 ] ,在强降水事件后根系主要利用浅层土壤水分(超过40%),降水稀少时则依靠地下水.在浅层土壤中,白刺根系生物量大于沙拐枣,在深层土壤中则相反,说明二者根系具有不同的水分利用策略[22 ] . ...
Adventitious roots and lateral roots:similarities and differences
2
2014
... 不定根(Adventitious roots)是由植物非根组织发育的根系,通常在植物的正常发育过程中(谷物的冠根和草莓的节根)或者在胁迫条件下(水淹、机械伤害、水分及养分胁迫等)产生[5 ] (图1 ).原生根(Initial roots)由植物胚胎根发育而来,不定根和原生根共同组成了植物的根系系统,保证了植物的固定及资源吸收利用[5 -6 ] .许多荒漠灌木在遭遇沙埋、损伤等干扰后,能从被埋的茎干上产生大量的不定根以应对各种复杂的环境[7 -8 ] .我们前期的大量野外调查也发现,中国西北荒漠区常见的白刺(Nitraria tangutorum )、柠条锦鸡儿(Caragana korshinskii )、霸王(Zygophyllum xanthoxylum )、红砂(Reaumuria songarica )、沙拐枣(Calligonum mongolicum )和柽柳(Tamarix chinensis )等沙生植物在沙埋后均可发育大量不定根(图2 ).这些不定根在缓解植物水分亏缺、增强养分捕获及促进克隆分株定植等方面发挥着关键作用[9 -11 ] .在干旱和半干旱地区,荒漠植物根系这种特有的生长和发育模式是其长期适应地带性水热条件的结果,也是荒漠植物生长发育及群落演变的关键,其构型和功能直接决定了植物对地下资源的获取能力,进而影响植物的生长发育、竞争能力及环境适应性[12 ] . ...
... Regulation factors related to adventitious roots
Table 1 调控机制 调控因子 功能及作用机制 参考文献 植物激素 生长素(IAA) 调控基因表达,促进细胞分裂分化,诱导根原基形成,促使不定根产生 [26 -28 ] 细胞分裂素(CK) 与生长素拮抗,高浓度抑制根原基形成 [26 -27 ,31 ] 乙烯(ET) 诱导皮层细胞死亡,形成通气组织,帮助根原基细胞产生,但可能在不定根诱导后期负向调控 [26 ,30 -31 ] 独脚金内酯(SL) 促进单子叶植物不定根伸长,植物受养分胁迫或机械损伤时,调控根系发育 [31 ] 茉莉酸(JA) 负向调控,过表达茉莉酸的负向调节因子可以促进植物产生更长、更多的不定根 [26 ,31 ] 脱落酸(ABA) 对水稻冠根生长有抑制作用 [27 ,31 ] 赤霉素(GA) 通常抑制不定根发育,但其和乙烯一起可促进皮层细胞死亡,促进不定根出现 [26 -27 ,31 ] 转录因子 WOX家族(WOX10/WOX11) 受生长素调控,建立根原基细胞 [32 -34 ] LBD家族(AtLBD18) 调控不定根的发生 [32 ] ARF家族(AtARF4/AtARF6) 参与木质部中柱鞘细胞的启动 [6 ,35 ] OsPLT家族(OsPLT1/OsPLT2) OsPLT1正向调控不定根和侧根的起始和生长,OsPLT2控制侧根的形成但不影响不定根的发育 [36 ] miR156 维持植物的高可塑性,使植物保持较强的根系再生能力,以应对不确定的环境资源 [37 ] 蛋白质 过氧化物酶 诱导不定根形成,参与植物的各种生理过程,如抗氧化防御和木质素的形成等 [38 ] 多酚氧化酶 主要集中在根原基形成区,影响细胞分裂分化和根原基的启动 [39 ] 吲哚乙酸氧化酶 氧化和降解IAA,调节内源激素的含量,帮助触发和启动植物根原基 [40 ]
<strong>2</strong> 不定根的形态特征 植物不定根的分布、形态及构型特征由遗传信息和生境条件共同决定[41 -42 ] .不同生活型植物之间不定根的分布和构型存在显著差异,同时,复杂的荒漠生境进一步加剧了根系形态的多样性. ...
Adventitious roots are key to the development of nebkhas in extremely arid regions
4
2019
... 不定根(Adventitious roots)是由植物非根组织发育的根系,通常在植物的正常发育过程中(谷物的冠根和草莓的节根)或者在胁迫条件下(水淹、机械伤害、水分及养分胁迫等)产生[5 ] (图1 ).原生根(Initial roots)由植物胚胎根发育而来,不定根和原生根共同组成了植物的根系系统,保证了植物的固定及资源吸收利用[5 -6 ] .许多荒漠灌木在遭遇沙埋、损伤等干扰后,能从被埋的茎干上产生大量的不定根以应对各种复杂的环境[7 -8 ] .我们前期的大量野外调查也发现,中国西北荒漠区常见的白刺(Nitraria tangutorum )、柠条锦鸡儿(Caragana korshinskii )、霸王(Zygophyllum xanthoxylum )、红砂(Reaumuria songarica )、沙拐枣(Calligonum mongolicum )和柽柳(Tamarix chinensis )等沙生植物在沙埋后均可发育大量不定根(图2 ).这些不定根在缓解植物水分亏缺、增强养分捕获及促进克隆分株定植等方面发挥着关键作用[9 -11 ] .在干旱和半干旱地区,荒漠植物根系这种特有的生长和发育模式是其长期适应地带性水热条件的结果,也是荒漠植物生长发育及群落演变的关键,其构型和功能直接决定了植物对地下资源的获取能力,进而影响植物的生长发育、竞争能力及环境适应性[12 ] . ...
... 相较于原生根,不定根具有更高的水分吸收效率,其以“低成本”的构建方式使植物以相同的碳消耗快速生长出更长的根系,从而在小降水事件或短暂的水分脉动出现时,能够迅速延伸至新的湿润土壤区域来获取水分[23 ] .此外,不定根根系形态特征还与土壤养分含量密切相关.研究表明,不定根的根直径随着土壤有机碳含量的增加而增大[7 ,24 ] .同时,荒漠克隆植物,如沙拐枣和沙鞭(Psammochloa villosa )等在土壤表层发育大量不定根可以极大地提高克隆分株的存活能力[9 ,11 ] .总之,不定根的存在和发育在植物的资源吸收利用效率、胁迫适应性和无性繁殖等方面均有重要意义.尽管如此,目前对荒漠灌木不定根的研究仍处于起步阶段,其形成的分子和基因调控机制、分布规律、对资源吸收的贡献及其增强植物适应性的内在机理等一系列问题仍缺乏深入理解.因此,本文系统梳理了不定根的发育机制、形态特征和生态功能等方面的研究成果,并对其在荒漠地下生态学研究中的未来发展趋势提出展望.通过该综述,以期丰富荒漠地下生态学研究理论体系,为干旱区荒漠植被建植和管理及生态恢复提供科学依据. ...
... 沙埋也是限制荒漠植物不定根生长和分布的关键因素,与根系形态特征紧密相关[51 ] .沙埋促进灌丛“肥岛效应”形成,造成沙堆土壤和丘间低地土壤养分分布不均[52 ] .沙埋导致植物产生不定根且不定根根径、长度及生物量与土壤有机碳、氮、磷含量显著正相关[44 ,52 ] .在沙埋干扰下,许多沙生灌木被埋枝叶通过分解纤维素产生可溶性糖和淀粉,为不定根生长提供能量[53 ] ,且不定根生物量占比随着沙埋深度的加剧而增大,如沙鞭、白刺等荒漠植物[7 -8 , 11 ] . ...
... 在极端生境中,沙生灌木有性繁殖成功率较低,营养繁殖成为部分荒漠植物种群维持与扩张的重要方式.荒漠区常见的沙拐枣和沙鞭等植物在营养繁殖过程中,克隆分株会产生大量不定根,这些不定根对表层土壤水分和养分的高效吸收有助于克隆分株存活,直接影响无性系分株的定植与生长发育,对极端生境下植物的自然更新和种群的维持扩张至关重要[11 ,69 ] .研究表明,在沙丘生境中,白刺在遭遇沙埋后可快速萌发并长出不定根,不定根细根生物量(87.1%)显著高于原生根细根生物量(12.9%).在成熟灌丛沙堆中,不定根氮和钾含量显著高于原生根,且沙堆内土壤水分高于丘间低地,并被不定根吸收[7 ] .相比之下,种子萌发形成的实生苗由于原生根根系不发达,幼苗难以度过干旱期,制约了沙拐枣种群的自然更新.综上,不定根的产生促进了荒漠克隆植物的生长和繁殖,是干旱生境中克隆分株存活率远高于实生苗的重要原因[9 ,11 ] . ...
Adventitious root production and plastic resource allocation to biomass determine burial tolerance in woody plants from central Canadian coastal dunes
3
2006
... 不定根(Adventitious roots)是由植物非根组织发育的根系,通常在植物的正常发育过程中(谷物的冠根和草莓的节根)或者在胁迫条件下(水淹、机械伤害、水分及养分胁迫等)产生[5 ] (图1 ).原生根(Initial roots)由植物胚胎根发育而来,不定根和原生根共同组成了植物的根系系统,保证了植物的固定及资源吸收利用[5 -6 ] .许多荒漠灌木在遭遇沙埋、损伤等干扰后,能从被埋的茎干上产生大量的不定根以应对各种复杂的环境[7 -8 ] .我们前期的大量野外调查也发现,中国西北荒漠区常见的白刺(Nitraria tangutorum )、柠条锦鸡儿(Caragana korshinskii )、霸王(Zygophyllum xanthoxylum )、红砂(Reaumuria songarica )、沙拐枣(Calligonum mongolicum )和柽柳(Tamarix chinensis )等沙生植物在沙埋后均可发育大量不定根(图2 ).这些不定根在缓解植物水分亏缺、增强养分捕获及促进克隆分株定植等方面发挥着关键作用[9 -11 ] .在干旱和半干旱地区,荒漠植物根系这种特有的生长和发育模式是其长期适应地带性水热条件的结果,也是荒漠植物生长发育及群落演变的关键,其构型和功能直接决定了植物对地下资源的获取能力,进而影响植物的生长发育、竞争能力及环境适应性[12 ] . ...
... 沙埋也是限制荒漠植物不定根生长和分布的关键因素,与根系形态特征紧密相关[51 ] .沙埋促进灌丛“肥岛效应”形成,造成沙堆土壤和丘间低地土壤养分分布不均[52 ] .沙埋导致植物产生不定根且不定根根径、长度及生物量与土壤有机碳、氮、磷含量显著正相关[44 ,52 ] .在沙埋干扰下,许多沙生灌木被埋枝叶通过分解纤维素产生可溶性糖和淀粉,为不定根生长提供能量[53 ] ,且不定根生物量占比随着沙埋深度的加剧而增大,如沙鞭、白刺等荒漠植物[7 -8 , 11 ] . ...
... 荒漠生态系统中强烈的风沙活动是驱动植物根系生长和发育的主要因素,当风沙流受植物阻拦时,沙物质在植物周围堆积,形成干旱-半干旱荒漠区常见的地貌类型——灌丛沙堆[1 ,56 -57 ] .这个过程使灌丛植被遭受一定程度的沙埋胁迫.不定根的生长可增强植物对浅层水分和养分的吸收能力以应对短期沙埋,维持植物的生理功能[53 ,58 ] .在沙埋干扰下,沙埋界面频繁的风沙活动及沙粒跃迁、撞击和摩擦,可使植物将资源重新分配至土壤表层的不定根和垂直茎上.不定根细胞排列更加紧密,木质化程度提高,从而增强了抗磨损能力和寿命,形成对干旱环境的适应性策略[8 ] .不定根的发育是沙生灌木适应风沙生境的重要策略,能够很好地缓解沙埋导致的资源胁迫. ...
Burial depth and diameter of the rhizome fragments affect the regenerative capacity of a clonal shrub
3
2015
... 不定根(Adventitious roots)是由植物非根组织发育的根系,通常在植物的正常发育过程中(谷物的冠根和草莓的节根)或者在胁迫条件下(水淹、机械伤害、水分及养分胁迫等)产生[5 ] (图1 ).原生根(Initial roots)由植物胚胎根发育而来,不定根和原生根共同组成了植物的根系系统,保证了植物的固定及资源吸收利用[5 -6 ] .许多荒漠灌木在遭遇沙埋、损伤等干扰后,能从被埋的茎干上产生大量的不定根以应对各种复杂的环境[7 -8 ] .我们前期的大量野外调查也发现,中国西北荒漠区常见的白刺(Nitraria tangutorum )、柠条锦鸡儿(Caragana korshinskii )、霸王(Zygophyllum xanthoxylum )、红砂(Reaumuria songarica )、沙拐枣(Calligonum mongolicum )和柽柳(Tamarix chinensis )等沙生植物在沙埋后均可发育大量不定根(图2 ).这些不定根在缓解植物水分亏缺、增强养分捕获及促进克隆分株定植等方面发挥着关键作用[9 -11 ] .在干旱和半干旱地区,荒漠植物根系这种特有的生长和发育模式是其长期适应地带性水热条件的结果,也是荒漠植物生长发育及群落演变的关键,其构型和功能直接决定了植物对地下资源的获取能力,进而影响植物的生长发育、竞争能力及环境适应性[12 ] . ...
... 相较于原生根,不定根具有更高的水分吸收效率,其以“低成本”的构建方式使植物以相同的碳消耗快速生长出更长的根系,从而在小降水事件或短暂的水分脉动出现时,能够迅速延伸至新的湿润土壤区域来获取水分[23 ] .此外,不定根根系形态特征还与土壤养分含量密切相关.研究表明,不定根的根直径随着土壤有机碳含量的增加而增大[7 ,24 ] .同时,荒漠克隆植物,如沙拐枣和沙鞭(Psammochloa villosa )等在土壤表层发育大量不定根可以极大地提高克隆分株的存活能力[9 ,11 ] .总之,不定根的存在和发育在植物的资源吸收利用效率、胁迫适应性和无性繁殖等方面均有重要意义.尽管如此,目前对荒漠灌木不定根的研究仍处于起步阶段,其形成的分子和基因调控机制、分布规律、对资源吸收的贡献及其增强植物适应性的内在机理等一系列问题仍缺乏深入理解.因此,本文系统梳理了不定根的发育机制、形态特征和生态功能等方面的研究成果,并对其在荒漠地下生态学研究中的未来发展趋势提出展望.通过该综述,以期丰富荒漠地下生态学研究理论体系,为干旱区荒漠植被建植和管理及生态恢复提供科学依据. ...
... 在极端生境中,沙生灌木有性繁殖成功率较低,营养繁殖成为部分荒漠植物种群维持与扩张的重要方式.荒漠区常见的沙拐枣和沙鞭等植物在营养繁殖过程中,克隆分株会产生大量不定根,这些不定根对表层土壤水分和养分的高效吸收有助于克隆分株存活,直接影响无性系分株的定植与生长发育,对极端生境下植物的自然更新和种群的维持扩张至关重要[11 ,69 ] .研究表明,在沙丘生境中,白刺在遭遇沙埋后可快速萌发并长出不定根,不定根细根生物量(87.1%)显著高于原生根细根生物量(12.9%).在成熟灌丛沙堆中,不定根氮和钾含量显著高于原生根,且沙堆内土壤水分高于丘间低地,并被不定根吸收[7 ] .相比之下,种子萌发形成的实生苗由于原生根根系不发达,幼苗难以度过干旱期,制约了沙拐枣种群的自然更新.综上,不定根的产生促进了荒漠克隆植物的生长和繁殖,是干旱生境中克隆分株存活率远高于实生苗的重要原因[9 ,11 ] . ...
Root system origin and structure influence planting shock of black spruce seedlings in boreal microsites
0
2019
Fixing active sand dune by native grasses in the desert of Northwest China
5
2024
... 不定根(Adventitious roots)是由植物非根组织发育的根系,通常在植物的正常发育过程中(谷物的冠根和草莓的节根)或者在胁迫条件下(水淹、机械伤害、水分及养分胁迫等)产生[5 ] (图1 ).原生根(Initial roots)由植物胚胎根发育而来,不定根和原生根共同组成了植物的根系系统,保证了植物的固定及资源吸收利用[5 -6 ] .许多荒漠灌木在遭遇沙埋、损伤等干扰后,能从被埋的茎干上产生大量的不定根以应对各种复杂的环境[7 -8 ] .我们前期的大量野外调查也发现,中国西北荒漠区常见的白刺(Nitraria tangutorum )、柠条锦鸡儿(Caragana korshinskii )、霸王(Zygophyllum xanthoxylum )、红砂(Reaumuria songarica )、沙拐枣(Calligonum mongolicum )和柽柳(Tamarix chinensis )等沙生植物在沙埋后均可发育大量不定根(图2 ).这些不定根在缓解植物水分亏缺、增强养分捕获及促进克隆分株定植等方面发挥着关键作用[9 -11 ] .在干旱和半干旱地区,荒漠植物根系这种特有的生长和发育模式是其长期适应地带性水热条件的结果,也是荒漠植物生长发育及群落演变的关键,其构型和功能直接决定了植物对地下资源的获取能力,进而影响植物的生长发育、竞争能力及环境适应性[12 ] . ...
... 相较于原生根,不定根具有更高的水分吸收效率,其以“低成本”的构建方式使植物以相同的碳消耗快速生长出更长的根系,从而在小降水事件或短暂的水分脉动出现时,能够迅速延伸至新的湿润土壤区域来获取水分[23 ] .此外,不定根根系形态特征还与土壤养分含量密切相关.研究表明,不定根的根直径随着土壤有机碳含量的增加而增大[7 ,24 ] .同时,荒漠克隆植物,如沙拐枣和沙鞭(Psammochloa villosa )等在土壤表层发育大量不定根可以极大地提高克隆分株的存活能力[9 ,11 ] .总之,不定根的存在和发育在植物的资源吸收利用效率、胁迫适应性和无性繁殖等方面均有重要意义.尽管如此,目前对荒漠灌木不定根的研究仍处于起步阶段,其形成的分子和基因调控机制、分布规律、对资源吸收的贡献及其增强植物适应性的内在机理等一系列问题仍缺乏深入理解.因此,本文系统梳理了不定根的发育机制、形态特征和生态功能等方面的研究成果,并对其在荒漠地下生态学研究中的未来发展趋势提出展望.通过该综述,以期丰富荒漠地下生态学研究理论体系,为干旱区荒漠植被建植和管理及生态恢复提供科学依据. ...
... 沙埋也是限制荒漠植物不定根生长和分布的关键因素,与根系形态特征紧密相关[51 ] .沙埋促进灌丛“肥岛效应”形成,造成沙堆土壤和丘间低地土壤养分分布不均[52 ] .沙埋导致植物产生不定根且不定根根径、长度及生物量与土壤有机碳、氮、磷含量显著正相关[44 ,52 ] .在沙埋干扰下,许多沙生灌木被埋枝叶通过分解纤维素产生可溶性糖和淀粉,为不定根生长提供能量[53 ] ,且不定根生物量占比随着沙埋深度的加剧而增大,如沙鞭、白刺等荒漠植物[7 -8 , 11 ] . ...
... 在极端生境中,沙生灌木有性繁殖成功率较低,营养繁殖成为部分荒漠植物种群维持与扩张的重要方式.荒漠区常见的沙拐枣和沙鞭等植物在营养繁殖过程中,克隆分株会产生大量不定根,这些不定根对表层土壤水分和养分的高效吸收有助于克隆分株存活,直接影响无性系分株的定植与生长发育,对极端生境下植物的自然更新和种群的维持扩张至关重要[11 ,69 ] .研究表明,在沙丘生境中,白刺在遭遇沙埋后可快速萌发并长出不定根,不定根细根生物量(87.1%)显著高于原生根细根生物量(12.9%).在成熟灌丛沙堆中,不定根氮和钾含量显著高于原生根,且沙堆内土壤水分高于丘间低地,并被不定根吸收[7 ] .相比之下,种子萌发形成的实生苗由于原生根根系不发达,幼苗难以度过干旱期,制约了沙拐枣种群的自然更新.综上,不定根的产生促进了荒漠克隆植物的生长和繁殖,是干旱生境中克隆分株存活率远高于实生苗的重要原因[9 ,11 ] . ...
... ,11 ]. ...
Root traits as drivers of plant and ecosystem functioning: current understanding,pitfalls and future research needs
1
2021
... 不定根(Adventitious roots)是由植物非根组织发育的根系,通常在植物的正常发育过程中(谷物的冠根和草莓的节根)或者在胁迫条件下(水淹、机械伤害、水分及养分胁迫等)产生[5 ] (图1 ).原生根(Initial roots)由植物胚胎根发育而来,不定根和原生根共同组成了植物的根系系统,保证了植物的固定及资源吸收利用[5 -6 ] .许多荒漠灌木在遭遇沙埋、损伤等干扰后,能从被埋的茎干上产生大量的不定根以应对各种复杂的环境[7 -8 ] .我们前期的大量野外调查也发现,中国西北荒漠区常见的白刺(Nitraria tangutorum )、柠条锦鸡儿(Caragana korshinskii )、霸王(Zygophyllum xanthoxylum )、红砂(Reaumuria songarica )、沙拐枣(Calligonum mongolicum )和柽柳(Tamarix chinensis )等沙生植物在沙埋后均可发育大量不定根(图2 ).这些不定根在缓解植物水分亏缺、增强养分捕获及促进克隆分株定植等方面发挥着关键作用[9 -11 ] .在干旱和半干旱地区,荒漠植物根系这种特有的生长和发育模式是其长期适应地带性水热条件的结果,也是荒漠植物生长发育及群落演变的关键,其构型和功能直接决定了植物对地下资源的获取能力,进而影响植物的生长发育、竞争能力及环境适应性[12 ] . ...
河西走廊中部两种荒漠植物根系构型特征
1
2013
... 以往关于植物根系的研究,主要涉及原生根的分布格局、构型特征、水分吸收来源及其对资源异质性的响应[13 -14 ] .此外,干旱区与非干旱区植物的根系生态策略及功能存在显著分异.在干旱区,强烈的蒸发与贫瘠的土壤促使植物将更多碳资源分配给根系,特别是通过发展不定根来增强水分吸收和运输能力,以应对干旱、养分匮乏及沙埋等胁迫.相比之下,非干旱区水分供应稳定,植物倾向于将光合产物优先分配至地上部分,以促进生长与繁殖,其不定根主要承担营养繁殖的功能.大量研究揭示了荒漠植物通过发展深根系以利用深层土壤水或地下水[15 -16 ] ,或通过调整根系构型(如由叉状分支转向鱼尾形分支)来应对水分和养分的空间异质性[17 -18 ] .例如,在河西走廊戈壁区和风沙区,红砂和白刺两种荒漠植物根系分支结构复杂,次级分支较多,根系分支均接近叉状分支模式,但随着降水量逐年减少,红砂根系构型向鱼尾形分支模式发展,以降低根系内部对营养物质的竞争,提高根系对贫瘠土壤水分和养分的吸收效率,从而适应干旱贫瘠的土壤环境[19 ] .而白刺主要增加侧根分支数目、根长及根系连接长度,加强扩展能力来应对干旱环境[20 ] .可见,不同物种根系构型及对资源异质性的响应策略是不同的.不同物种根系水分利用策略也有不同,沙拐枣根系吸收水分主要来源于深层土壤水(约26%)和地下水(超过30%),而白刺由于其二态性根系结构特征[21 ] ,在强降水事件后根系主要利用浅层土壤水分(超过40%),降水稀少时则依靠地下水.在浅层土壤中,白刺根系生物量大于沙拐枣,在深层土壤中则相反,说明二者根系具有不同的水分利用策略[22 ] . ...
Plant root mechanisms and their effects on carbon and nutrient accumulation in desert ecosystems under changes in land use and climate
1
2024
... 以往关于植物根系的研究,主要涉及原生根的分布格局、构型特征、水分吸收来源及其对资源异质性的响应[13 -14 ] .此外,干旱区与非干旱区植物的根系生态策略及功能存在显著分异.在干旱区,强烈的蒸发与贫瘠的土壤促使植物将更多碳资源分配给根系,特别是通过发展不定根来增强水分吸收和运输能力,以应对干旱、养分匮乏及沙埋等胁迫.相比之下,非干旱区水分供应稳定,植物倾向于将光合产物优先分配至地上部分,以促进生长与繁殖,其不定根主要承担营养繁殖的功能.大量研究揭示了荒漠植物通过发展深根系以利用深层土壤水或地下水[15 -16 ] ,或通过调整根系构型(如由叉状分支转向鱼尾形分支)来应对水分和养分的空间异质性[17 -18 ] .例如,在河西走廊戈壁区和风沙区,红砂和白刺两种荒漠植物根系分支结构复杂,次级分支较多,根系分支均接近叉状分支模式,但随着降水量逐年减少,红砂根系构型向鱼尾形分支模式发展,以降低根系内部对营养物质的竞争,提高根系对贫瘠土壤水分和养分的吸收效率,从而适应干旱贫瘠的土壤环境[19 ] .而白刺主要增加侧根分支数目、根长及根系连接长度,加强扩展能力来应对干旱环境[20 ] .可见,不同物种根系构型及对资源异质性的响应策略是不同的.不同物种根系水分利用策略也有不同,沙拐枣根系吸收水分主要来源于深层土壤水(约26%)和地下水(超过30%),而白刺由于其二态性根系结构特征[21 ] ,在强降水事件后根系主要利用浅层土壤水分(超过40%),降水稀少时则依靠地下水.在浅层土壤中,白刺根系生物量大于沙拐枣,在深层土壤中则相反,说明二者根系具有不同的水分利用策略[22 ] . ...
Water-use strategy of three central Asian desert shrubs and their responses to rain pulse events
1
2006
... 以往关于植物根系的研究,主要涉及原生根的分布格局、构型特征、水分吸收来源及其对资源异质性的响应[13 -14 ] .此外,干旱区与非干旱区植物的根系生态策略及功能存在显著分异.在干旱区,强烈的蒸发与贫瘠的土壤促使植物将更多碳资源分配给根系,特别是通过发展不定根来增强水分吸收和运输能力,以应对干旱、养分匮乏及沙埋等胁迫.相比之下,非干旱区水分供应稳定,植物倾向于将光合产物优先分配至地上部分,以促进生长与繁殖,其不定根主要承担营养繁殖的功能.大量研究揭示了荒漠植物通过发展深根系以利用深层土壤水或地下水[15 -16 ] ,或通过调整根系构型(如由叉状分支转向鱼尾形分支)来应对水分和养分的空间异质性[17 -18 ] .例如,在河西走廊戈壁区和风沙区,红砂和白刺两种荒漠植物根系分支结构复杂,次级分支较多,根系分支均接近叉状分支模式,但随着降水量逐年减少,红砂根系构型向鱼尾形分支模式发展,以降低根系内部对营养物质的竞争,提高根系对贫瘠土壤水分和养分的吸收效率,从而适应干旱贫瘠的土壤环境[19 ] .而白刺主要增加侧根分支数目、根长及根系连接长度,加强扩展能力来应对干旱环境[20 ] .可见,不同物种根系构型及对资源异质性的响应策略是不同的.不同物种根系水分利用策略也有不同,沙拐枣根系吸收水分主要来源于深层土壤水(约26%)和地下水(超过30%),而白刺由于其二态性根系结构特征[21 ] ,在强降水事件后根系主要利用浅层土壤水分(超过40%),降水稀少时则依靠地下水.在浅层土壤中,白刺根系生物量大于沙拐枣,在深层土壤中则相反,说明二者根系具有不同的水分利用策略[22 ] . ...
Stable oxygen isotopes reveal distinct water use patterns of two Haloxylon species in the Gurbantonggut Desert
1
2015
... 以往关于植物根系的研究,主要涉及原生根的分布格局、构型特征、水分吸收来源及其对资源异质性的响应[13 -14 ] .此外,干旱区与非干旱区植物的根系生态策略及功能存在显著分异.在干旱区,强烈的蒸发与贫瘠的土壤促使植物将更多碳资源分配给根系,特别是通过发展不定根来增强水分吸收和运输能力,以应对干旱、养分匮乏及沙埋等胁迫.相比之下,非干旱区水分供应稳定,植物倾向于将光合产物优先分配至地上部分,以促进生长与繁殖,其不定根主要承担营养繁殖的功能.大量研究揭示了荒漠植物通过发展深根系以利用深层土壤水或地下水[15 -16 ] ,或通过调整根系构型(如由叉状分支转向鱼尾形分支)来应对水分和养分的空间异质性[17 -18 ] .例如,在河西走廊戈壁区和风沙区,红砂和白刺两种荒漠植物根系分支结构复杂,次级分支较多,根系分支均接近叉状分支模式,但随着降水量逐年减少,红砂根系构型向鱼尾形分支模式发展,以降低根系内部对营养物质的竞争,提高根系对贫瘠土壤水分和养分的吸收效率,从而适应干旱贫瘠的土壤环境[19 ] .而白刺主要增加侧根分支数目、根长及根系连接长度,加强扩展能力来应对干旱环境[20 ] .可见,不同物种根系构型及对资源异质性的响应策略是不同的.不同物种根系水分利用策略也有不同,沙拐枣根系吸收水分主要来源于深层土壤水(约26%)和地下水(超过30%),而白刺由于其二态性根系结构特征[21 ] ,在强降水事件后根系主要利用浅层土壤水分(超过40%),降水稀少时则依靠地下水.在浅层土壤中,白刺根系生物量大于沙拐枣,在深层土壤中则相反,说明二者根系具有不同的水分利用策略[22 ] . ...
塔克拉玛干沙漠腹地3种植物根系构型及其生境适应策略
1
2008
... 以往关于植物根系的研究,主要涉及原生根的分布格局、构型特征、水分吸收来源及其对资源异质性的响应[13 -14 ] .此外,干旱区与非干旱区植物的根系生态策略及功能存在显著分异.在干旱区,强烈的蒸发与贫瘠的土壤促使植物将更多碳资源分配给根系,特别是通过发展不定根来增强水分吸收和运输能力,以应对干旱、养分匮乏及沙埋等胁迫.相比之下,非干旱区水分供应稳定,植物倾向于将光合产物优先分配至地上部分,以促进生长与繁殖,其不定根主要承担营养繁殖的功能.大量研究揭示了荒漠植物通过发展深根系以利用深层土壤水或地下水[15 -16 ] ,或通过调整根系构型(如由叉状分支转向鱼尾形分支)来应对水分和养分的空间异质性[17 -18 ] .例如,在河西走廊戈壁区和风沙区,红砂和白刺两种荒漠植物根系分支结构复杂,次级分支较多,根系分支均接近叉状分支模式,但随着降水量逐年减少,红砂根系构型向鱼尾形分支模式发展,以降低根系内部对营养物质的竞争,提高根系对贫瘠土壤水分和养分的吸收效率,从而适应干旱贫瘠的土壤环境[19 ] .而白刺主要增加侧根分支数目、根长及根系连接长度,加强扩展能力来应对干旱环境[20 ] .可见,不同物种根系构型及对资源异质性的响应策略是不同的.不同物种根系水分利用策略也有不同,沙拐枣根系吸收水分主要来源于深层土壤水(约26%)和地下水(超过30%),而白刺由于其二态性根系结构特征[21 ] ,在强降水事件后根系主要利用浅层土壤水分(超过40%),降水稀少时则依靠地下水.在浅层土壤中,白刺根系生物量大于沙拐枣,在深层土壤中则相反,说明二者根系具有不同的水分利用策略[22 ] . ...
红砂根系构型对干旱的生态适应
1
2012
... 以往关于植物根系的研究,主要涉及原生根的分布格局、构型特征、水分吸收来源及其对资源异质性的响应[13 -14 ] .此外,干旱区与非干旱区植物的根系生态策略及功能存在显著分异.在干旱区,强烈的蒸发与贫瘠的土壤促使植物将更多碳资源分配给根系,特别是通过发展不定根来增强水分吸收和运输能力,以应对干旱、养分匮乏及沙埋等胁迫.相比之下,非干旱区水分供应稳定,植物倾向于将光合产物优先分配至地上部分,以促进生长与繁殖,其不定根主要承担营养繁殖的功能.大量研究揭示了荒漠植物通过发展深根系以利用深层土壤水或地下水[15 -16 ] ,或通过调整根系构型(如由叉状分支转向鱼尾形分支)来应对水分和养分的空间异质性[17 -18 ] .例如,在河西走廊戈壁区和风沙区,红砂和白刺两种荒漠植物根系分支结构复杂,次级分支较多,根系分支均接近叉状分支模式,但随着降水量逐年减少,红砂根系构型向鱼尾形分支模式发展,以降低根系内部对营养物质的竞争,提高根系对贫瘠土壤水分和养分的吸收效率,从而适应干旱贫瘠的土壤环境[19 ] .而白刺主要增加侧根分支数目、根长及根系连接长度,加强扩展能力来应对干旱环境[20 ] .可见,不同物种根系构型及对资源异质性的响应策略是不同的.不同物种根系水分利用策略也有不同,沙拐枣根系吸收水分主要来源于深层土壤水(约26%)和地下水(超过30%),而白刺由于其二态性根系结构特征[21 ] ,在强降水事件后根系主要利用浅层土壤水分(超过40%),降水稀少时则依靠地下水.在浅层土壤中,白刺根系生物量大于沙拐枣,在深层土壤中则相反,说明二者根系具有不同的水分利用策略[22 ] . ...
额济纳荒漠植物红砂(Reaumuria songarica )根系构型、叶片生理指标变化的研究
1
2022
... 以往关于植物根系的研究,主要涉及原生根的分布格局、构型特征、水分吸收来源及其对资源异质性的响应[13 -14 ] .此外,干旱区与非干旱区植物的根系生态策略及功能存在显著分异.在干旱区,强烈的蒸发与贫瘠的土壤促使植物将更多碳资源分配给根系,特别是通过发展不定根来增强水分吸收和运输能力,以应对干旱、养分匮乏及沙埋等胁迫.相比之下,非干旱区水分供应稳定,植物倾向于将光合产物优先分配至地上部分,以促进生长与繁殖,其不定根主要承担营养繁殖的功能.大量研究揭示了荒漠植物通过发展深根系以利用深层土壤水或地下水[15 -16 ] ,或通过调整根系构型(如由叉状分支转向鱼尾形分支)来应对水分和养分的空间异质性[17 -18 ] .例如,在河西走廊戈壁区和风沙区,红砂和白刺两种荒漠植物根系分支结构复杂,次级分支较多,根系分支均接近叉状分支模式,但随着降水量逐年减少,红砂根系构型向鱼尾形分支模式发展,以降低根系内部对营养物质的竞争,提高根系对贫瘠土壤水分和养分的吸收效率,从而适应干旱贫瘠的土壤环境[19 ] .而白刺主要增加侧根分支数目、根长及根系连接长度,加强扩展能力来应对干旱环境[20 ] .可见,不同物种根系构型及对资源异质性的响应策略是不同的.不同物种根系水分利用策略也有不同,沙拐枣根系吸收水分主要来源于深层土壤水(约26%)和地下水(超过30%),而白刺由于其二态性根系结构特征[21 ] ,在强降水事件后根系主要利用浅层土壤水分(超过40%),降水稀少时则依靠地下水.在浅层土壤中,白刺根系生物量大于沙拐枣,在深层土壤中则相反,说明二者根系具有不同的水分利用策略[22 ] . ...
西北典型荒漠植物根系形态结构和功能及抗旱生理研究
1
2013
... 以往关于植物根系的研究,主要涉及原生根的分布格局、构型特征、水分吸收来源及其对资源异质性的响应[13 -14 ] .此外,干旱区与非干旱区植物的根系生态策略及功能存在显著分异.在干旱区,强烈的蒸发与贫瘠的土壤促使植物将更多碳资源分配给根系,特别是通过发展不定根来增强水分吸收和运输能力,以应对干旱、养分匮乏及沙埋等胁迫.相比之下,非干旱区水分供应稳定,植物倾向于将光合产物优先分配至地上部分,以促进生长与繁殖,其不定根主要承担营养繁殖的功能.大量研究揭示了荒漠植物通过发展深根系以利用深层土壤水或地下水[15 -16 ] ,或通过调整根系构型(如由叉状分支转向鱼尾形分支)来应对水分和养分的空间异质性[17 -18 ] .例如,在河西走廊戈壁区和风沙区,红砂和白刺两种荒漠植物根系分支结构复杂,次级分支较多,根系分支均接近叉状分支模式,但随着降水量逐年减少,红砂根系构型向鱼尾形分支模式发展,以降低根系内部对营养物质的竞争,提高根系对贫瘠土壤水分和养分的吸收效率,从而适应干旱贫瘠的土壤环境[19 ] .而白刺主要增加侧根分支数目、根长及根系连接长度,加强扩展能力来应对干旱环境[20 ] .可见,不同物种根系构型及对资源异质性的响应策略是不同的.不同物种根系水分利用策略也有不同,沙拐枣根系吸收水分主要来源于深层土壤水(约26%)和地下水(超过30%),而白刺由于其二态性根系结构特征[21 ] ,在强降水事件后根系主要利用浅层土壤水分(超过40%),降水稀少时则依靠地下水.在浅层土壤中,白刺根系生物量大于沙拐枣,在深层土壤中则相反,说明二者根系具有不同的水分利用策略[22 ] . ...
Root distribution of Nitraria sibirica with seasonally varying water sources in a desert habitat
2
2015
... 以往关于植物根系的研究,主要涉及原生根的分布格局、构型特征、水分吸收来源及其对资源异质性的响应[13 -14 ] .此外,干旱区与非干旱区植物的根系生态策略及功能存在显著分异.在干旱区,强烈的蒸发与贫瘠的土壤促使植物将更多碳资源分配给根系,特别是通过发展不定根来增强水分吸收和运输能力,以应对干旱、养分匮乏及沙埋等胁迫.相比之下,非干旱区水分供应稳定,植物倾向于将光合产物优先分配至地上部分,以促进生长与繁殖,其不定根主要承担营养繁殖的功能.大量研究揭示了荒漠植物通过发展深根系以利用深层土壤水或地下水[15 -16 ] ,或通过调整根系构型(如由叉状分支转向鱼尾形分支)来应对水分和养分的空间异质性[17 -18 ] .例如,在河西走廊戈壁区和风沙区,红砂和白刺两种荒漠植物根系分支结构复杂,次级分支较多,根系分支均接近叉状分支模式,但随着降水量逐年减少,红砂根系构型向鱼尾形分支模式发展,以降低根系内部对营养物质的竞争,提高根系对贫瘠土壤水分和养分的吸收效率,从而适应干旱贫瘠的土壤环境[19 ] .而白刺主要增加侧根分支数目、根长及根系连接长度,加强扩展能力来应对干旱环境[20 ] .可见,不同物种根系构型及对资源异质性的响应策略是不同的.不同物种根系水分利用策略也有不同,沙拐枣根系吸收水分主要来源于深层土壤水(约26%)和地下水(超过30%),而白刺由于其二态性根系结构特征[21 ] ,在强降水事件后根系主要利用浅层土壤水分(超过40%),降水稀少时则依靠地下水.在浅层土壤中,白刺根系生物量大于沙拐枣,在深层土壤中则相反,说明二者根系具有不同的水分利用策略[22 ] . ...
... 不定根的形态和构型与植物水分的获取途径密切相关,是其生态适应性的最直观体现.在轻度干旱胁迫下,植物会增加不定根的数量来提高水分获取效率[45 -46 ] ,且不定根的直径与土壤水分显著正相关,表明当短暂的降水发生时,植物会增加根系直径以增强水分运输效率[47 -48 ] .研究表明,沙生灌木白刺根系呈二态性分布[21 ] ,即在降水较为充足的季节或月份,白刺利用不定根吸收小降水事件补充的浅层土壤水;在降水较少或浅层土壤水匮乏时,转而利用原生根吸收由较大降水事件补给的深层土壤水分及地下水以满足其正常生长发育需求[49 ] .这是由于在干旱-半干旱生态系统中,降水事件具有高度不确定性和脉冲性,浅层土壤水分动态波动剧烈.植物可以通过快速产生不定根,来高效捕获降水后浅层土壤的短暂水分.然而,强烈的地表蒸发使得这些水分迅速散失,导致不定根的功能窗口期很短.当浅层资源难以持续供给时,植物则将资源投入转向深层根系构建,以获取更稳定的深层土壤水或地下水,从而保障长期的生存与繁殖[50 ] . ...
Hydrological niche separation between two coexisting shrubs in an extremely arid region
1
2025
... 以往关于植物根系的研究,主要涉及原生根的分布格局、构型特征、水分吸收来源及其对资源异质性的响应[13 -14 ] .此外,干旱区与非干旱区植物的根系生态策略及功能存在显著分异.在干旱区,强烈的蒸发与贫瘠的土壤促使植物将更多碳资源分配给根系,特别是通过发展不定根来增强水分吸收和运输能力,以应对干旱、养分匮乏及沙埋等胁迫.相比之下,非干旱区水分供应稳定,植物倾向于将光合产物优先分配至地上部分,以促进生长与繁殖,其不定根主要承担营养繁殖的功能.大量研究揭示了荒漠植物通过发展深根系以利用深层土壤水或地下水[15 -16 ] ,或通过调整根系构型(如由叉状分支转向鱼尾形分支)来应对水分和养分的空间异质性[17 -18 ] .例如,在河西走廊戈壁区和风沙区,红砂和白刺两种荒漠植物根系分支结构复杂,次级分支较多,根系分支均接近叉状分支模式,但随着降水量逐年减少,红砂根系构型向鱼尾形分支模式发展,以降低根系内部对营养物质的竞争,提高根系对贫瘠土壤水分和养分的吸收效率,从而适应干旱贫瘠的土壤环境[19 ] .而白刺主要增加侧根分支数目、根长及根系连接长度,加强扩展能力来应对干旱环境[20 ] .可见,不同物种根系构型及对资源异质性的响应策略是不同的.不同物种根系水分利用策略也有不同,沙拐枣根系吸收水分主要来源于深层土壤水(约26%)和地下水(超过30%),而白刺由于其二态性根系结构特征[21 ] ,在强降水事件后根系主要利用浅层土壤水分(超过40%),降水稀少时则依靠地下水.在浅层土壤中,白刺根系生物量大于沙拐枣,在深层土壤中则相反,说明二者根系具有不同的水分利用策略[22 ] . ...
Steep,cheap and deep: an ideotype to optimize water and N acquisition by maize root systems
2
2013
... 相较于原生根,不定根具有更高的水分吸收效率,其以“低成本”的构建方式使植物以相同的碳消耗快速生长出更长的根系,从而在小降水事件或短暂的水分脉动出现时,能够迅速延伸至新的湿润土壤区域来获取水分[23 ] .此外,不定根根系形态特征还与土壤养分含量密切相关.研究表明,不定根的根直径随着土壤有机碳含量的增加而增大[7 ,24 ] .同时,荒漠克隆植物,如沙拐枣和沙鞭(Psammochloa villosa )等在土壤表层发育大量不定根可以极大地提高克隆分株的存活能力[9 ,11 ] .总之,不定根的存在和发育在植物的资源吸收利用效率、胁迫适应性和无性繁殖等方面均有重要意义.尽管如此,目前对荒漠灌木不定根的研究仍处于起步阶段,其形成的分子和基因调控机制、分布规律、对资源吸收的贡献及其增强植物适应性的内在机理等一系列问题仍缺乏深入理解.因此,本文系统梳理了不定根的发育机制、形态特征和生态功能等方面的研究成果,并对其在荒漠地下生态学研究中的未来发展趋势提出展望.通过该综述,以期丰富荒漠地下生态学研究理论体系,为干旱区荒漠植被建植和管理及生态恢复提供科学依据. ...
... 土壤养分对根系发育的影响和水分胁迫相似[61 -62 ] .在养分胁迫条件下,植物会增加不定根的数量来提高对表层磷的吸收和利用,且不定根根长与土壤总磷含量显著正相关,这是由于磷在土壤中移动缓慢,易与土壤颗粒结合,有效性低且分布不均,因此植物通过增加根系长度和密度,同时发育根毛来扩大吸收面积,从而增强对磷的吸收能力[63 ] .此外,植物通过增加不定根的生长角度(重力响应度)来探索深层土壤中的氮[23 ,45 ,64 -65 ] .在盐碱胁迫下,仙人掌(Echinocactus platyacanthus )幼苗仍能持续发育不定根,表明不定根有助于增强植物的抗逆性[66 ] .安宁等[44 ] 的研究也表明,在低水分条件下,氮磷添加可提高不定根的比根长和比表面积,而在水分充足条件下,氮磷添加反而抑制了不定根的形态特征变化,说明荒漠植物根系具有较强的可塑性,不同物种对水分和养分变化的响应存在显著差异.因此,不定根的存在有助于植物适应贫瘠或养分异质性较大的土壤生境,在低养分条件下,不定根的养分吸收能力高于原生根,而在高养分条件下,二者差异不明显[67 ] .总之,不定根的发育极大地提高了植物在胁迫生境中的养分获取能力,是植物适应极端生境的重要策略[68 ] . ...
Response mechanisms of adventitious root architectural characteristics of Nitraria tangutorum shrubs to soil nutrients in nabkha
1
2022
... 相较于原生根,不定根具有更高的水分吸收效率,其以“低成本”的构建方式使植物以相同的碳消耗快速生长出更长的根系,从而在小降水事件或短暂的水分脉动出现时,能够迅速延伸至新的湿润土壤区域来获取水分[23 ] .此外,不定根根系形态特征还与土壤养分含量密切相关.研究表明,不定根的根直径随着土壤有机碳含量的增加而增大[7 ,24 ] .同时,荒漠克隆植物,如沙拐枣和沙鞭(Psammochloa villosa )等在土壤表层发育大量不定根可以极大地提高克隆分株的存活能力[9 ,11 ] .总之,不定根的存在和发育在植物的资源吸收利用效率、胁迫适应性和无性繁殖等方面均有重要意义.尽管如此,目前对荒漠灌木不定根的研究仍处于起步阶段,其形成的分子和基因调控机制、分布规律、对资源吸收的贡献及其增强植物适应性的内在机理等一系列问题仍缺乏深入理解.因此,本文系统梳理了不定根的发育机制、形态特征和生态功能等方面的研究成果,并对其在荒漠地下生态学研究中的未来发展趋势提出展望.通过该综述,以期丰富荒漠地下生态学研究理论体系,为干旱区荒漠植被建植和管理及生态恢复提供科学依据. ...
Uncovering protein regulation during adventitious root formation in Platycladus orientalis cuttings
1
2024
... 不定根作为植物根系的重要组成部分,其生长和发育受环境因素(光照、水分、养分和温度等)以及植物自身特性(树种、激素、基因、蛋白质、转录因子和插穗年龄等)的共同调控[25 ] .不定根的形成通常经历诱导期、启动期和表达期,分别对应根原基细胞的诱导产生、分裂分化与不定根的伸长突破.在诱导期,根原基细胞感知多种信号刺激,部分基因开启表达,植物代谢能力增强,诱导下胚轴不定根根原基产生;在启动期,根原基细胞持续分裂分化,分生组织快速形成,根原基发育加快;在表达期,根原基继续发育生长至伸出表皮层,与原组织中的维管束相连,至此不定根的形成结束[26 -27 ] . ...
植物不定根生成的生理及分子调控机制研究进展
9
2024
... 不定根作为植物根系的重要组成部分,其生长和发育受环境因素(光照、水分、养分和温度等)以及植物自身特性(树种、激素、基因、蛋白质、转录因子和插穗年龄等)的共同调控[25 ] .不定根的形成通常经历诱导期、启动期和表达期,分别对应根原基细胞的诱导产生、分裂分化与不定根的伸长突破.在诱导期,根原基细胞感知多种信号刺激,部分基因开启表达,植物代谢能力增强,诱导下胚轴不定根根原基产生;在启动期,根原基细胞持续分裂分化,分生组织快速形成,根原基发育加快;在表达期,根原基继续发育生长至伸出表皮层,与原组织中的维管束相连,至此不定根的形成结束[26 -27 ] . ...
... 生长素(IAA)在不定根发生过程中起核心调控作用,主要负责诱导根原基的形成[28 ] .在不定根的形成过程中,3-吲哚丁酸(IBA)可以转化为具有活性的IAA[29 ] .此外,植物其余多种激素参与不定根的发育调控.例如细胞分裂素(CK)与生长素呈拮抗作用,抑制CK的表达可促进根原基细胞产生与不定根的伸长和突破[26 ] ;乙烯(ET)可通过调节基因表达,诱导皮层细胞死亡,帮助根原基细胞产生[30 ] .赤霉素(GA)在植物不定根发生的不同阶段可起到抑制或促进作用[26 ] ;茉莉酸(JA)对不定根进行负向调节,植物可通过调节OsJJAZ1/EG2基因来抑制茉莉酸的表达,从而促进根原基细胞的产生[31 ] .不定根的形成不仅是植物的生理响应过程,也受遗传基因严格控制.当植物体内激素积累时,相关基因会快速做出响应,发挥重要作用[32 -33 ] .在植物激素与基因调控不定根形成的过程中,多种关键蛋白质也参与不定根的形成和发育. ...
... [26 ];茉莉酸(JA)对不定根进行负向调节,植物可通过调节OsJJAZ1/EG2基因来抑制茉莉酸的表达,从而促进根原基细胞的产生[31 ] .不定根的形成不仅是植物的生理响应过程,也受遗传基因严格控制.当植物体内激素积累时,相关基因会快速做出响应,发挥重要作用[32 -33 ] .在植物激素与基因调控不定根形成的过程中,多种关键蛋白质也参与不定根的形成和发育. ...
... 不定根的形成是一个多因子协同调控的复杂生物学过程[26 -28 ,30 -40 ] (表1 ).然而,与水稻、玉米和桉树等经济作物和经济林树种相比,沙生灌木在不定根的形成能力和调控机制方面具有显著差异.作为干旱、半干旱区荒漠生态系统的主要植被类型,沙生灌木长期处于沙埋、水分和养分等环境胁迫条件下,通常需要依靠快速形成不定根来维持水分和养分的吸收与固定,因此,其不定根的形成条件和发育机制等可能与上述经济作物存在显著差异,具有一定的特殊性.然而,现有关于不定根形成机制的研究主要集中于桉树、水稻、玉米等经济物种,针对沙生灌木的系统研究相对匮乏.对于沙生灌木在沙埋、干旱等特殊生境中,不定根诱导过程中激素动态、关键基因及蛋白质调控网络仍缺乏深入了解,还存在一系列问题待我们深入探讨.例如,沙生灌木中激素互作对不定根形成的调控路径是否不同于经济林和作物,是否存在特殊的IBA-IAA转化机制,是否具有响应沙地环境的特异转录因子和蛋白质表达模式.未来亟需进一步系统研究,以揭示沙生灌木独特的胁迫适应机制并为干旱区植被恢复与育种提供理论依据. ...
... Regulation factors related to adventitious roots
Table 1 调控机制 调控因子 功能及作用机制 参考文献 植物激素 生长素(IAA) 调控基因表达,促进细胞分裂分化,诱导根原基形成,促使不定根产生 [26 -28 ] 细胞分裂素(CK) 与生长素拮抗,高浓度抑制根原基形成 [26 -27 ,31 ] 乙烯(ET) 诱导皮层细胞死亡,形成通气组织,帮助根原基细胞产生,但可能在不定根诱导后期负向调控 [26 ,30 -31 ] 独脚金内酯(SL) 促进单子叶植物不定根伸长,植物受养分胁迫或机械损伤时,调控根系发育 [31 ] 茉莉酸(JA) 负向调控,过表达茉莉酸的负向调节因子可以促进植物产生更长、更多的不定根 [26 ,31 ] 脱落酸(ABA) 对水稻冠根生长有抑制作用 [27 ,31 ] 赤霉素(GA) 通常抑制不定根发育,但其和乙烯一起可促进皮层细胞死亡,促进不定根出现 [26 -27 ,31 ] 转录因子 WOX家族(WOX10/WOX11) 受生长素调控,建立根原基细胞 [32 -34 ] LBD家族(AtLBD18) 调控不定根的发生 [32 ] ARF家族(AtARF4/AtARF6) 参与木质部中柱鞘细胞的启动 [6 ,35 ] OsPLT家族(OsPLT1/OsPLT2) OsPLT1正向调控不定根和侧根的起始和生长,OsPLT2控制侧根的形成但不影响不定根的发育 [36 ] miR156 维持植物的高可塑性,使植物保持较强的根系再生能力,以应对不确定的环境资源 [37 ] 蛋白质 过氧化物酶 诱导不定根形成,参与植物的各种生理过程,如抗氧化防御和木质素的形成等 [38 ] 多酚氧化酶 主要集中在根原基形成区,影响细胞分裂分化和根原基的启动 [39 ] 吲哚乙酸氧化酶 氧化和降解IAA,调节内源激素的含量,帮助触发和启动植物根原基 [40 ]
<strong>2</strong> 不定根的形态特征 植物不定根的分布、形态及构型特征由遗传信息和生境条件共同决定[41 -42 ] .不同生活型植物之间不定根的分布和构型存在显著差异,同时,复杂的荒漠生境进一步加剧了根系形态的多样性. ...
... [
26 -
27 ,
31 ]
乙烯(ET) 诱导皮层细胞死亡,形成通气组织,帮助根原基细胞产生,但可能在不定根诱导后期负向调控 [26 ,30 -31 ] 独脚金内酯(SL) 促进单子叶植物不定根伸长,植物受养分胁迫或机械损伤时,调控根系发育 [31 ] 茉莉酸(JA) 负向调控,过表达茉莉酸的负向调节因子可以促进植物产生更长、更多的不定根 [26 ,31 ] 脱落酸(ABA) 对水稻冠根生长有抑制作用 [27 ,31 ] 赤霉素(GA) 通常抑制不定根发育,但其和乙烯一起可促进皮层细胞死亡,促进不定根出现 [26 -27 ,31 ] 转录因子 WOX家族(WOX10/WOX11) 受生长素调控,建立根原基细胞 [32 -34 ] LBD家族(AtLBD18) 调控不定根的发生 [32 ] ARF家族(AtARF4/AtARF6) 参与木质部中柱鞘细胞的启动 [6 ,35 ] OsPLT家族(OsPLT1/OsPLT2) OsPLT1正向调控不定根和侧根的起始和生长,OsPLT2控制侧根的形成但不影响不定根的发育 [36 ] miR156 维持植物的高可塑性,使植物保持较强的根系再生能力,以应对不确定的环境资源 [37 ] 蛋白质 过氧化物酶 诱导不定根形成,参与植物的各种生理过程,如抗氧化防御和木质素的形成等 [38 ] 多酚氧化酶 主要集中在根原基形成区,影响细胞分裂分化和根原基的启动 [39 ] 吲哚乙酸氧化酶 氧化和降解IAA,调节内源激素的含量,帮助触发和启动植物根原基 [40 ] <strong>2</strong> 不定根的形态特征 植物不定根的分布、形态及构型特征由遗传信息和生境条件共同决定[41 -42 ] .不同生活型植物之间不定根的分布和构型存在显著差异,同时,复杂的荒漠生境进一步加剧了根系形态的多样性. ...
... [
26 ,
30 -
31 ]
独脚金内酯(SL) 促进单子叶植物不定根伸长,植物受养分胁迫或机械损伤时,调控根系发育 [31 ] 茉莉酸(JA) 负向调控,过表达茉莉酸的负向调节因子可以促进植物产生更长、更多的不定根 [26 ,31 ] 脱落酸(ABA) 对水稻冠根生长有抑制作用 [27 ,31 ] 赤霉素(GA) 通常抑制不定根发育,但其和乙烯一起可促进皮层细胞死亡,促进不定根出现 [26 -27 ,31 ] 转录因子 WOX家族(WOX10/WOX11) 受生长素调控,建立根原基细胞 [32 -34 ] LBD家族(AtLBD18) 调控不定根的发生 [32 ] ARF家族(AtARF4/AtARF6) 参与木质部中柱鞘细胞的启动 [6 ,35 ] OsPLT家族(OsPLT1/OsPLT2) OsPLT1正向调控不定根和侧根的起始和生长,OsPLT2控制侧根的形成但不影响不定根的发育 [36 ] miR156 维持植物的高可塑性,使植物保持较强的根系再生能力,以应对不确定的环境资源 [37 ] 蛋白质 过氧化物酶 诱导不定根形成,参与植物的各种生理过程,如抗氧化防御和木质素的形成等 [38 ] 多酚氧化酶 主要集中在根原基形成区,影响细胞分裂分化和根原基的启动 [39 ] 吲哚乙酸氧化酶 氧化和降解IAA,调节内源激素的含量,帮助触发和启动植物根原基 [40 ] <strong>2</strong> 不定根的形态特征 植物不定根的分布、形态及构型特征由遗传信息和生境条件共同决定[41 -42 ] .不同生活型植物之间不定根的分布和构型存在显著差异,同时,复杂的荒漠生境进一步加剧了根系形态的多样性. ...
... [
26 ,
31 ]
脱落酸(ABA) 对水稻冠根生长有抑制作用 [27 ,31 ] 赤霉素(GA) 通常抑制不定根发育,但其和乙烯一起可促进皮层细胞死亡,促进不定根出现 [26 -27 ,31 ] 转录因子 WOX家族(WOX10/WOX11) 受生长素调控,建立根原基细胞 [32 -34 ] LBD家族(AtLBD18) 调控不定根的发生 [32 ] ARF家族(AtARF4/AtARF6) 参与木质部中柱鞘细胞的启动 [6 ,35 ] OsPLT家族(OsPLT1/OsPLT2) OsPLT1正向调控不定根和侧根的起始和生长,OsPLT2控制侧根的形成但不影响不定根的发育 [36 ] miR156 维持植物的高可塑性,使植物保持较强的根系再生能力,以应对不确定的环境资源 [37 ] 蛋白质 过氧化物酶 诱导不定根形成,参与植物的各种生理过程,如抗氧化防御和木质素的形成等 [38 ] 多酚氧化酶 主要集中在根原基形成区,影响细胞分裂分化和根原基的启动 [39 ] 吲哚乙酸氧化酶 氧化和降解IAA,调节内源激素的含量,帮助触发和启动植物根原基 [40 ] <strong>2</strong> 不定根的形态特征 植物不定根的分布、形态及构型特征由遗传信息和生境条件共同决定[41 -42 ] .不同生活型植物之间不定根的分布和构型存在显著差异,同时,复杂的荒漠生境进一步加剧了根系形态的多样性. ...
... [
26 -
27 ,
31 ]
转录因子 WOX家族(WOX10/WOX11) 受生长素调控,建立根原基细胞 [32 -34 ] LBD家族(AtLBD18) 调控不定根的发生 [32 ] ARF家族(AtARF4/AtARF6) 参与木质部中柱鞘细胞的启动 [6 ,35 ] OsPLT家族(OsPLT1/OsPLT2) OsPLT1正向调控不定根和侧根的起始和生长,OsPLT2控制侧根的形成但不影响不定根的发育 [36 ] miR156 维持植物的高可塑性,使植物保持较强的根系再生能力,以应对不确定的环境资源 [37 ] 蛋白质 过氧化物酶 诱导不定根形成,参与植物的各种生理过程,如抗氧化防御和木质素的形成等 [38 ] 多酚氧化酶 主要集中在根原基形成区,影响细胞分裂分化和根原基的启动 [39 ] 吲哚乙酸氧化酶 氧化和降解IAA,调节内源激素的含量,帮助触发和启动植物根原基 [40 ] <strong>2</strong> 不定根的形态特征 植物不定根的分布、形态及构型特征由遗传信息和生境条件共同决定[41 -42 ] .不同生活型植物之间不定根的分布和构型存在显著差异,同时,复杂的荒漠生境进一步加剧了根系形态的多样性. ...
木本植物不定根发生机制研究进展
4
2022
... 不定根作为植物根系的重要组成部分,其生长和发育受环境因素(光照、水分、养分和温度等)以及植物自身特性(树种、激素、基因、蛋白质、转录因子和插穗年龄等)的共同调控[25 ] .不定根的形成通常经历诱导期、启动期和表达期,分别对应根原基细胞的诱导产生、分裂分化与不定根的伸长突破.在诱导期,根原基细胞感知多种信号刺激,部分基因开启表达,植物代谢能力增强,诱导下胚轴不定根根原基产生;在启动期,根原基细胞持续分裂分化,分生组织快速形成,根原基发育加快;在表达期,根原基继续发育生长至伸出表皮层,与原组织中的维管束相连,至此不定根的形成结束[26 -27 ] . ...
... Regulation factors related to adventitious roots
Table 1 调控机制 调控因子 功能及作用机制 参考文献 植物激素 生长素(IAA) 调控基因表达,促进细胞分裂分化,诱导根原基形成,促使不定根产生 [26 -28 ] 细胞分裂素(CK) 与生长素拮抗,高浓度抑制根原基形成 [26 -27 ,31 ] 乙烯(ET) 诱导皮层细胞死亡,形成通气组织,帮助根原基细胞产生,但可能在不定根诱导后期负向调控 [26 ,30 -31 ] 独脚金内酯(SL) 促进单子叶植物不定根伸长,植物受养分胁迫或机械损伤时,调控根系发育 [31 ] 茉莉酸(JA) 负向调控,过表达茉莉酸的负向调节因子可以促进植物产生更长、更多的不定根 [26 ,31 ] 脱落酸(ABA) 对水稻冠根生长有抑制作用 [27 ,31 ] 赤霉素(GA) 通常抑制不定根发育,但其和乙烯一起可促进皮层细胞死亡,促进不定根出现 [26 -27 ,31 ] 转录因子 WOX家族(WOX10/WOX11) 受生长素调控,建立根原基细胞 [32 -34 ] LBD家族(AtLBD18) 调控不定根的发生 [32 ] ARF家族(AtARF4/AtARF6) 参与木质部中柱鞘细胞的启动 [6 ,35 ] OsPLT家族(OsPLT1/OsPLT2) OsPLT1正向调控不定根和侧根的起始和生长,OsPLT2控制侧根的形成但不影响不定根的发育 [36 ] miR156 维持植物的高可塑性,使植物保持较强的根系再生能力,以应对不确定的环境资源 [37 ] 蛋白质 过氧化物酶 诱导不定根形成,参与植物的各种生理过程,如抗氧化防御和木质素的形成等 [38 ] 多酚氧化酶 主要集中在根原基形成区,影响细胞分裂分化和根原基的启动 [39 ] 吲哚乙酸氧化酶 氧化和降解IAA,调节内源激素的含量,帮助触发和启动植物根原基 [40 ]
<strong>2</strong> 不定根的形态特征 植物不定根的分布、形态及构型特征由遗传信息和生境条件共同决定[41 -42 ] .不同生活型植物之间不定根的分布和构型存在显著差异,同时,复杂的荒漠生境进一步加剧了根系形态的多样性. ...
... [
27 ,
31 ]
赤霉素(GA) 通常抑制不定根发育,但其和乙烯一起可促进皮层细胞死亡,促进不定根出现 [26 -27 ,31 ] 转录因子 WOX家族(WOX10/WOX11) 受生长素调控,建立根原基细胞 [32 -34 ] LBD家族(AtLBD18) 调控不定根的发生 [32 ] ARF家族(AtARF4/AtARF6) 参与木质部中柱鞘细胞的启动 [6 ,35 ] OsPLT家族(OsPLT1/OsPLT2) OsPLT1正向调控不定根和侧根的起始和生长,OsPLT2控制侧根的形成但不影响不定根的发育 [36 ] miR156 维持植物的高可塑性,使植物保持较强的根系再生能力,以应对不确定的环境资源 [37 ] 蛋白质 过氧化物酶 诱导不定根形成,参与植物的各种生理过程,如抗氧化防御和木质素的形成等 [38 ] 多酚氧化酶 主要集中在根原基形成区,影响细胞分裂分化和根原基的启动 [39 ] 吲哚乙酸氧化酶 氧化和降解IAA,调节内源激素的含量,帮助触发和启动植物根原基 [40 ] <strong>2</strong> 不定根的形态特征 植物不定根的分布、形态及构型特征由遗传信息和生境条件共同决定[41 -42 ] .不同生活型植物之间不定根的分布和构型存在显著差异,同时,复杂的荒漠生境进一步加剧了根系形态的多样性. ...
... -
27 ,
31 ]
转录因子 WOX家族(WOX10/WOX11) 受生长素调控,建立根原基细胞 [32 -34 ] LBD家族(AtLBD18) 调控不定根的发生 [32 ] ARF家族(AtARF4/AtARF6) 参与木质部中柱鞘细胞的启动 [6 ,35 ] OsPLT家族(OsPLT1/OsPLT2) OsPLT1正向调控不定根和侧根的起始和生长,OsPLT2控制侧根的形成但不影响不定根的发育 [36 ] miR156 维持植物的高可塑性,使植物保持较强的根系再生能力,以应对不确定的环境资源 [37 ] 蛋白质 过氧化物酶 诱导不定根形成,参与植物的各种生理过程,如抗氧化防御和木质素的形成等 [38 ] 多酚氧化酶 主要集中在根原基形成区,影响细胞分裂分化和根原基的启动 [39 ] 吲哚乙酸氧化酶 氧化和降解IAA,调节内源激素的含量,帮助触发和启动植物根原基 [40 ] <strong>2</strong> 不定根的形态特征 植物不定根的分布、形态及构型特征由遗传信息和生境条件共同决定[41 -42 ] .不同生活型植物之间不定根的分布和构型存在显著差异,同时,复杂的荒漠生境进一步加剧了根系形态的多样性. ...
Auxin is a central player in the hormone cross-talks that control adventitious rooting
3
2014
... 生长素(IAA)在不定根发生过程中起核心调控作用,主要负责诱导根原基的形成[28 ] .在不定根的形成过程中,3-吲哚丁酸(IBA)可以转化为具有活性的IAA[29 ] .此外,植物其余多种激素参与不定根的发育调控.例如细胞分裂素(CK)与生长素呈拮抗作用,抑制CK的表达可促进根原基细胞产生与不定根的伸长和突破[26 ] ;乙烯(ET)可通过调节基因表达,诱导皮层细胞死亡,帮助根原基细胞产生[30 ] .赤霉素(GA)在植物不定根发生的不同阶段可起到抑制或促进作用[26 ] ;茉莉酸(JA)对不定根进行负向调节,植物可通过调节OsJJAZ1/EG2基因来抑制茉莉酸的表达,从而促进根原基细胞的产生[31 ] .不定根的形成不仅是植物的生理响应过程,也受遗传基因严格控制.当植物体内激素积累时,相关基因会快速做出响应,发挥重要作用[32 -33 ] .在植物激素与基因调控不定根形成的过程中,多种关键蛋白质也参与不定根的形成和发育. ...
... 不定根的形成是一个多因子协同调控的复杂生物学过程[26 -28 ,30 -40 ] (表1 ).然而,与水稻、玉米和桉树等经济作物和经济林树种相比,沙生灌木在不定根的形成能力和调控机制方面具有显著差异.作为干旱、半干旱区荒漠生态系统的主要植被类型,沙生灌木长期处于沙埋、水分和养分等环境胁迫条件下,通常需要依靠快速形成不定根来维持水分和养分的吸收与固定,因此,其不定根的形成条件和发育机制等可能与上述经济作物存在显著差异,具有一定的特殊性.然而,现有关于不定根形成机制的研究主要集中于桉树、水稻、玉米等经济物种,针对沙生灌木的系统研究相对匮乏.对于沙生灌木在沙埋、干旱等特殊生境中,不定根诱导过程中激素动态、关键基因及蛋白质调控网络仍缺乏深入了解,还存在一系列问题待我们深入探讨.例如,沙生灌木中激素互作对不定根形成的调控路径是否不同于经济林和作物,是否存在特殊的IBA-IAA转化机制,是否具有响应沙地环境的特异转录因子和蛋白质表达模式.未来亟需进一步系统研究,以揭示沙生灌木独特的胁迫适应机制并为干旱区植被恢复与育种提供理论依据. ...
... Regulation factors related to adventitious roots
Table 1 调控机制 调控因子 功能及作用机制 参考文献 植物激素 生长素(IAA) 调控基因表达,促进细胞分裂分化,诱导根原基形成,促使不定根产生 [26 -28 ] 细胞分裂素(CK) 与生长素拮抗,高浓度抑制根原基形成 [26 -27 ,31 ] 乙烯(ET) 诱导皮层细胞死亡,形成通气组织,帮助根原基细胞产生,但可能在不定根诱导后期负向调控 [26 ,30 -31 ] 独脚金内酯(SL) 促进单子叶植物不定根伸长,植物受养分胁迫或机械损伤时,调控根系发育 [31 ] 茉莉酸(JA) 负向调控,过表达茉莉酸的负向调节因子可以促进植物产生更长、更多的不定根 [26 ,31 ] 脱落酸(ABA) 对水稻冠根生长有抑制作用 [27 ,31 ] 赤霉素(GA) 通常抑制不定根发育,但其和乙烯一起可促进皮层细胞死亡,促进不定根出现 [26 -27 ,31 ] 转录因子 WOX家族(WOX10/WOX11) 受生长素调控,建立根原基细胞 [32 -34 ] LBD家族(AtLBD18) 调控不定根的发生 [32 ] ARF家族(AtARF4/AtARF6) 参与木质部中柱鞘细胞的启动 [6 ,35 ] OsPLT家族(OsPLT1/OsPLT2) OsPLT1正向调控不定根和侧根的起始和生长,OsPLT2控制侧根的形成但不影响不定根的发育 [36 ] miR156 维持植物的高可塑性,使植物保持较强的根系再生能力,以应对不确定的环境资源 [37 ] 蛋白质 过氧化物酶 诱导不定根形成,参与植物的各种生理过程,如抗氧化防御和木质素的形成等 [38 ] 多酚氧化酶 主要集中在根原基形成区,影响细胞分裂分化和根原基的启动 [39 ] 吲哚乙酸氧化酶 氧化和降解IAA,调节内源激素的含量,帮助触发和启动植物根原基 [40 ]
<strong>2</strong> 不定根的形态特征 植物不定根的分布、形态及构型特征由遗传信息和生境条件共同决定[41 -42 ] .不同生活型植物之间不定根的分布和构型存在显著差异,同时,复杂的荒漠生境进一步加剧了根系形态的多样性. ...
Factors governing cellular reprogramming competence in Arabidopsis adventitious root formation
1
2024
... 生长素(IAA)在不定根发生过程中起核心调控作用,主要负责诱导根原基的形成[28 ] .在不定根的形成过程中,3-吲哚丁酸(IBA)可以转化为具有活性的IAA[29 ] .此外,植物其余多种激素参与不定根的发育调控.例如细胞分裂素(CK)与生长素呈拮抗作用,抑制CK的表达可促进根原基细胞产生与不定根的伸长和突破[26 ] ;乙烯(ET)可通过调节基因表达,诱导皮层细胞死亡,帮助根原基细胞产生[30 ] .赤霉素(GA)在植物不定根发生的不同阶段可起到抑制或促进作用[26 ] ;茉莉酸(JA)对不定根进行负向调节,植物可通过调节OsJJAZ1/EG2基因来抑制茉莉酸的表达,从而促进根原基细胞的产生[31 ] .不定根的形成不仅是植物的生理响应过程,也受遗传基因严格控制.当植物体内激素积累时,相关基因会快速做出响应,发挥重要作用[32 -33 ] .在植物激素与基因调控不定根形成的过程中,多种关键蛋白质也参与不定根的形成和发育. ...
When stress and development go hand in hand:main hormonal controls of adventitious rooting in cuttings
3
2013
... 生长素(IAA)在不定根发生过程中起核心调控作用,主要负责诱导根原基的形成[28 ] .在不定根的形成过程中,3-吲哚丁酸(IBA)可以转化为具有活性的IAA[29 ] .此外,植物其余多种激素参与不定根的发育调控.例如细胞分裂素(CK)与生长素呈拮抗作用,抑制CK的表达可促进根原基细胞产生与不定根的伸长和突破[26 ] ;乙烯(ET)可通过调节基因表达,诱导皮层细胞死亡,帮助根原基细胞产生[30 ] .赤霉素(GA)在植物不定根发生的不同阶段可起到抑制或促进作用[26 ] ;茉莉酸(JA)对不定根进行负向调节,植物可通过调节OsJJAZ1/EG2基因来抑制茉莉酸的表达,从而促进根原基细胞的产生[31 ] .不定根的形成不仅是植物的生理响应过程,也受遗传基因严格控制.当植物体内激素积累时,相关基因会快速做出响应,发挥重要作用[32 -33 ] .在植物激素与基因调控不定根形成的过程中,多种关键蛋白质也参与不定根的形成和发育. ...
... 不定根的形成是一个多因子协同调控的复杂生物学过程[26 -28 ,30 -40 ] (表1 ).然而,与水稻、玉米和桉树等经济作物和经济林树种相比,沙生灌木在不定根的形成能力和调控机制方面具有显著差异.作为干旱、半干旱区荒漠生态系统的主要植被类型,沙生灌木长期处于沙埋、水分和养分等环境胁迫条件下,通常需要依靠快速形成不定根来维持水分和养分的吸收与固定,因此,其不定根的形成条件和发育机制等可能与上述经济作物存在显著差异,具有一定的特殊性.然而,现有关于不定根形成机制的研究主要集中于桉树、水稻、玉米等经济物种,针对沙生灌木的系统研究相对匮乏.对于沙生灌木在沙埋、干旱等特殊生境中,不定根诱导过程中激素动态、关键基因及蛋白质调控网络仍缺乏深入了解,还存在一系列问题待我们深入探讨.例如,沙生灌木中激素互作对不定根形成的调控路径是否不同于经济林和作物,是否存在特殊的IBA-IAA转化机制,是否具有响应沙地环境的特异转录因子和蛋白质表达模式.未来亟需进一步系统研究,以揭示沙生灌木独特的胁迫适应机制并为干旱区植被恢复与育种提供理论依据. ...
... Regulation factors related to adventitious roots
Table 1 调控机制 调控因子 功能及作用机制 参考文献 植物激素 生长素(IAA) 调控基因表达,促进细胞分裂分化,诱导根原基形成,促使不定根产生 [26 -28 ] 细胞分裂素(CK) 与生长素拮抗,高浓度抑制根原基形成 [26 -27 ,31 ] 乙烯(ET) 诱导皮层细胞死亡,形成通气组织,帮助根原基细胞产生,但可能在不定根诱导后期负向调控 [26 ,30 -31 ] 独脚金内酯(SL) 促进单子叶植物不定根伸长,植物受养分胁迫或机械损伤时,调控根系发育 [31 ] 茉莉酸(JA) 负向调控,过表达茉莉酸的负向调节因子可以促进植物产生更长、更多的不定根 [26 ,31 ] 脱落酸(ABA) 对水稻冠根生长有抑制作用 [27 ,31 ] 赤霉素(GA) 通常抑制不定根发育,但其和乙烯一起可促进皮层细胞死亡,促进不定根出现 [26 -27 ,31 ] 转录因子 WOX家族(WOX10/WOX11) 受生长素调控,建立根原基细胞 [32 -34 ] LBD家族(AtLBD18) 调控不定根的发生 [32 ] ARF家族(AtARF4/AtARF6) 参与木质部中柱鞘细胞的启动 [6 ,35 ] OsPLT家族(OsPLT1/OsPLT2) OsPLT1正向调控不定根和侧根的起始和生长,OsPLT2控制侧根的形成但不影响不定根的发育 [36 ] miR156 维持植物的高可塑性,使植物保持较强的根系再生能力,以应对不确定的环境资源 [37 ] 蛋白质 过氧化物酶 诱导不定根形成,参与植物的各种生理过程,如抗氧化防御和木质素的形成等 [38 ] 多酚氧化酶 主要集中在根原基形成区,影响细胞分裂分化和根原基的启动 [39 ] 吲哚乙酸氧化酶 氧化和降解IAA,调节内源激素的含量,帮助触发和启动植物根原基 [40 ]
<strong>2</strong> 不定根的形态特征 植物不定根的分布、形态及构型特征由遗传信息和生境条件共同决定[41 -42 ] .不同生活型植物之间不定根的分布和构型存在显著差异,同时,复杂的荒漠生境进一步加剧了根系形态的多样性. ...
Genetic and hormonal blueprint of shoot-borne adventitious root development in rice and maize
7
2022
... 生长素(IAA)在不定根发生过程中起核心调控作用,主要负责诱导根原基的形成[28 ] .在不定根的形成过程中,3-吲哚丁酸(IBA)可以转化为具有活性的IAA[29 ] .此外,植物其余多种激素参与不定根的发育调控.例如细胞分裂素(CK)与生长素呈拮抗作用,抑制CK的表达可促进根原基细胞产生与不定根的伸长和突破[26 ] ;乙烯(ET)可通过调节基因表达,诱导皮层细胞死亡,帮助根原基细胞产生[30 ] .赤霉素(GA)在植物不定根发生的不同阶段可起到抑制或促进作用[26 ] ;茉莉酸(JA)对不定根进行负向调节,植物可通过调节OsJJAZ1/EG2基因来抑制茉莉酸的表达,从而促进根原基细胞的产生[31 ] .不定根的形成不仅是植物的生理响应过程,也受遗传基因严格控制.当植物体内激素积累时,相关基因会快速做出响应,发挥重要作用[32 -33 ] .在植物激素与基因调控不定根形成的过程中,多种关键蛋白质也参与不定根的形成和发育. ...
... Regulation factors related to adventitious roots
Table 1 调控机制 调控因子 功能及作用机制 参考文献 植物激素 生长素(IAA) 调控基因表达,促进细胞分裂分化,诱导根原基形成,促使不定根产生 [26 -28 ] 细胞分裂素(CK) 与生长素拮抗,高浓度抑制根原基形成 [26 -27 ,31 ] 乙烯(ET) 诱导皮层细胞死亡,形成通气组织,帮助根原基细胞产生,但可能在不定根诱导后期负向调控 [26 ,30 -31 ] 独脚金内酯(SL) 促进单子叶植物不定根伸长,植物受养分胁迫或机械损伤时,调控根系发育 [31 ] 茉莉酸(JA) 负向调控,过表达茉莉酸的负向调节因子可以促进植物产生更长、更多的不定根 [26 ,31 ] 脱落酸(ABA) 对水稻冠根生长有抑制作用 [27 ,31 ] 赤霉素(GA) 通常抑制不定根发育,但其和乙烯一起可促进皮层细胞死亡,促进不定根出现 [26 -27 ,31 ] 转录因子 WOX家族(WOX10/WOX11) 受生长素调控,建立根原基细胞 [32 -34 ] LBD家族(AtLBD18) 调控不定根的发生 [32 ] ARF家族(AtARF4/AtARF6) 参与木质部中柱鞘细胞的启动 [6 ,35 ] OsPLT家族(OsPLT1/OsPLT2) OsPLT1正向调控不定根和侧根的起始和生长,OsPLT2控制侧根的形成但不影响不定根的发育 [36 ] miR156 维持植物的高可塑性,使植物保持较强的根系再生能力,以应对不确定的环境资源 [37 ] 蛋白质 过氧化物酶 诱导不定根形成,参与植物的各种生理过程,如抗氧化防御和木质素的形成等 [38 ] 多酚氧化酶 主要集中在根原基形成区,影响细胞分裂分化和根原基的启动 [39 ] 吲哚乙酸氧化酶 氧化和降解IAA,调节内源激素的含量,帮助触发和启动植物根原基 [40 ]
<strong>2</strong> 不定根的形态特征 植物不定根的分布、形态及构型特征由遗传信息和生境条件共同决定[41 -42 ] .不同生活型植物之间不定根的分布和构型存在显著差异,同时,复杂的荒漠生境进一步加剧了根系形态的多样性. ...
... -
31 ]
独脚金内酯(SL) 促进单子叶植物不定根伸长,植物受养分胁迫或机械损伤时,调控根系发育 [31 ] 茉莉酸(JA) 负向调控,过表达茉莉酸的负向调节因子可以促进植物产生更长、更多的不定根 [26 ,31 ] 脱落酸(ABA) 对水稻冠根生长有抑制作用 [27 ,31 ] 赤霉素(GA) 通常抑制不定根发育,但其和乙烯一起可促进皮层细胞死亡,促进不定根出现 [26 -27 ,31 ] 转录因子 WOX家族(WOX10/WOX11) 受生长素调控,建立根原基细胞 [32 -34 ] LBD家族(AtLBD18) 调控不定根的发生 [32 ] ARF家族(AtARF4/AtARF6) 参与木质部中柱鞘细胞的启动 [6 ,35 ] OsPLT家族(OsPLT1/OsPLT2) OsPLT1正向调控不定根和侧根的起始和生长,OsPLT2控制侧根的形成但不影响不定根的发育 [36 ] miR156 维持植物的高可塑性,使植物保持较强的根系再生能力,以应对不确定的环境资源 [37 ] 蛋白质 过氧化物酶 诱导不定根形成,参与植物的各种生理过程,如抗氧化防御和木质素的形成等 [38 ] 多酚氧化酶 主要集中在根原基形成区,影响细胞分裂分化和根原基的启动 [39 ] 吲哚乙酸氧化酶 氧化和降解IAA,调节内源激素的含量,帮助触发和启动植物根原基 [40 ] <strong>2</strong> 不定根的形态特征 植物不定根的分布、形态及构型特征由遗传信息和生境条件共同决定[41 -42 ] .不同生活型植物之间不定根的分布和构型存在显著差异,同时,复杂的荒漠生境进一步加剧了根系形态的多样性. ...
... [
31 ]
茉莉酸(JA) 负向调控,过表达茉莉酸的负向调节因子可以促进植物产生更长、更多的不定根 [26 ,31 ] 脱落酸(ABA) 对水稻冠根生长有抑制作用 [27 ,31 ] 赤霉素(GA) 通常抑制不定根发育,但其和乙烯一起可促进皮层细胞死亡,促进不定根出现 [26 -27 ,31 ] 转录因子 WOX家族(WOX10/WOX11) 受生长素调控,建立根原基细胞 [32 -34 ] LBD家族(AtLBD18) 调控不定根的发生 [32 ] ARF家族(AtARF4/AtARF6) 参与木质部中柱鞘细胞的启动 [6 ,35 ] OsPLT家族(OsPLT1/OsPLT2) OsPLT1正向调控不定根和侧根的起始和生长,OsPLT2控制侧根的形成但不影响不定根的发育 [36 ] miR156 维持植物的高可塑性,使植物保持较强的根系再生能力,以应对不确定的环境资源 [37 ] 蛋白质 过氧化物酶 诱导不定根形成,参与植物的各种生理过程,如抗氧化防御和木质素的形成等 [38 ] 多酚氧化酶 主要集中在根原基形成区,影响细胞分裂分化和根原基的启动 [39 ] 吲哚乙酸氧化酶 氧化和降解IAA,调节内源激素的含量,帮助触发和启动植物根原基 [40 ] <strong>2</strong> 不定根的形态特征 植物不定根的分布、形态及构型特征由遗传信息和生境条件共同决定[41 -42 ] .不同生活型植物之间不定根的分布和构型存在显著差异,同时,复杂的荒漠生境进一步加剧了根系形态的多样性. ...
... ,
31 ]
脱落酸(ABA) 对水稻冠根生长有抑制作用 [27 ,31 ] 赤霉素(GA) 通常抑制不定根发育,但其和乙烯一起可促进皮层细胞死亡,促进不定根出现 [26 -27 ,31 ] 转录因子 WOX家族(WOX10/WOX11) 受生长素调控,建立根原基细胞 [32 -34 ] LBD家族(AtLBD18) 调控不定根的发生 [32 ] ARF家族(AtARF4/AtARF6) 参与木质部中柱鞘细胞的启动 [6 ,35 ] OsPLT家族(OsPLT1/OsPLT2) OsPLT1正向调控不定根和侧根的起始和生长,OsPLT2控制侧根的形成但不影响不定根的发育 [36 ] miR156 维持植物的高可塑性,使植物保持较强的根系再生能力,以应对不确定的环境资源 [37 ] 蛋白质 过氧化物酶 诱导不定根形成,参与植物的各种生理过程,如抗氧化防御和木质素的形成等 [38 ] 多酚氧化酶 主要集中在根原基形成区,影响细胞分裂分化和根原基的启动 [39 ] 吲哚乙酸氧化酶 氧化和降解IAA,调节内源激素的含量,帮助触发和启动植物根原基 [40 ] <strong>2</strong> 不定根的形态特征 植物不定根的分布、形态及构型特征由遗传信息和生境条件共同决定[41 -42 ] .不同生活型植物之间不定根的分布和构型存在显著差异,同时,复杂的荒漠生境进一步加剧了根系形态的多样性. ...
... ,
31 ]
赤霉素(GA) 通常抑制不定根发育,但其和乙烯一起可促进皮层细胞死亡,促进不定根出现 [26 -27 ,31 ] 转录因子 WOX家族(WOX10/WOX11) 受生长素调控,建立根原基细胞 [32 -34 ] LBD家族(AtLBD18) 调控不定根的发生 [32 ] ARF家族(AtARF4/AtARF6) 参与木质部中柱鞘细胞的启动 [6 ,35 ] OsPLT家族(OsPLT1/OsPLT2) OsPLT1正向调控不定根和侧根的起始和生长,OsPLT2控制侧根的形成但不影响不定根的发育 [36 ] miR156 维持植物的高可塑性,使植物保持较强的根系再生能力,以应对不确定的环境资源 [37 ] 蛋白质 过氧化物酶 诱导不定根形成,参与植物的各种生理过程,如抗氧化防御和木质素的形成等 [38 ] 多酚氧化酶 主要集中在根原基形成区,影响细胞分裂分化和根原基的启动 [39 ] 吲哚乙酸氧化酶 氧化和降解IAA,调节内源激素的含量,帮助触发和启动植物根原基 [40 ] <strong>2</strong> 不定根的形态特征 植物不定根的分布、形态及构型特征由遗传信息和生境条件共同决定[41 -42 ] .不同生活型植物之间不定根的分布和构型存在显著差异,同时,复杂的荒漠生境进一步加剧了根系形态的多样性. ...
... ,
31 ]
转录因子 WOX家族(WOX10/WOX11) 受生长素调控,建立根原基细胞 [32 -34 ] LBD家族(AtLBD18) 调控不定根的发生 [32 ] ARF家族(AtARF4/AtARF6) 参与木质部中柱鞘细胞的启动 [6 ,35 ] OsPLT家族(OsPLT1/OsPLT2) OsPLT1正向调控不定根和侧根的起始和生长,OsPLT2控制侧根的形成但不影响不定根的发育 [36 ] miR156 维持植物的高可塑性,使植物保持较强的根系再生能力,以应对不确定的环境资源 [37 ] 蛋白质 过氧化物酶 诱导不定根形成,参与植物的各种生理过程,如抗氧化防御和木质素的形成等 [38 ] 多酚氧化酶 主要集中在根原基形成区,影响细胞分裂分化和根原基的启动 [39 ] 吲哚乙酸氧化酶 氧化和降解IAA,调节内源激素的含量,帮助触发和启动植物根原基 [40 ] <strong>2</strong> 不定根的形态特征 植物不定根的分布、形态及构型特征由遗传信息和生境条件共同决定[41 -42 ] .不同生活型植物之间不定根的分布和构型存在显著差异,同时,复杂的荒漠生境进一步加剧了根系形态的多样性. ...
WOX11 and 12 are involved in the first-step cell fate transition during de novo root organogenesis in Arabidopsis
3
2014
... 生长素(IAA)在不定根发生过程中起核心调控作用,主要负责诱导根原基的形成[28 ] .在不定根的形成过程中,3-吲哚丁酸(IBA)可以转化为具有活性的IAA[29 ] .此外,植物其余多种激素参与不定根的发育调控.例如细胞分裂素(CK)与生长素呈拮抗作用,抑制CK的表达可促进根原基细胞产生与不定根的伸长和突破[26 ] ;乙烯(ET)可通过调节基因表达,诱导皮层细胞死亡,帮助根原基细胞产生[30 ] .赤霉素(GA)在植物不定根发生的不同阶段可起到抑制或促进作用[26 ] ;茉莉酸(JA)对不定根进行负向调节,植物可通过调节OsJJAZ1/EG2基因来抑制茉莉酸的表达,从而促进根原基细胞的产生[31 ] .不定根的形成不仅是植物的生理响应过程,也受遗传基因严格控制.当植物体内激素积累时,相关基因会快速做出响应,发挥重要作用[32 -33 ] .在植物激素与基因调控不定根形成的过程中,多种关键蛋白质也参与不定根的形成和发育. ...
... Regulation factors related to adventitious roots
Table 1 调控机制 调控因子 功能及作用机制 参考文献 植物激素 生长素(IAA) 调控基因表达,促进细胞分裂分化,诱导根原基形成,促使不定根产生 [26 -28 ] 细胞分裂素(CK) 与生长素拮抗,高浓度抑制根原基形成 [26 -27 ,31 ] 乙烯(ET) 诱导皮层细胞死亡,形成通气组织,帮助根原基细胞产生,但可能在不定根诱导后期负向调控 [26 ,30 -31 ] 独脚金内酯(SL) 促进单子叶植物不定根伸长,植物受养分胁迫或机械损伤时,调控根系发育 [31 ] 茉莉酸(JA) 负向调控,过表达茉莉酸的负向调节因子可以促进植物产生更长、更多的不定根 [26 ,31 ] 脱落酸(ABA) 对水稻冠根生长有抑制作用 [27 ,31 ] 赤霉素(GA) 通常抑制不定根发育,但其和乙烯一起可促进皮层细胞死亡,促进不定根出现 [26 -27 ,31 ] 转录因子 WOX家族(WOX10/WOX11) 受生长素调控,建立根原基细胞 [32 -34 ] LBD家族(AtLBD18) 调控不定根的发生 [32 ] ARF家族(AtARF4/AtARF6) 参与木质部中柱鞘细胞的启动 [6 ,35 ] OsPLT家族(OsPLT1/OsPLT2) OsPLT1正向调控不定根和侧根的起始和生长,OsPLT2控制侧根的形成但不影响不定根的发育 [36 ] miR156 维持植物的高可塑性,使植物保持较强的根系再生能力,以应对不确定的环境资源 [37 ] 蛋白质 过氧化物酶 诱导不定根形成,参与植物的各种生理过程,如抗氧化防御和木质素的形成等 [38 ] 多酚氧化酶 主要集中在根原基形成区,影响细胞分裂分化和根原基的启动 [39 ] 吲哚乙酸氧化酶 氧化和降解IAA,调节内源激素的含量,帮助触发和启动植物根原基 [40 ]
<strong>2</strong> 不定根的形态特征 植物不定根的分布、形态及构型特征由遗传信息和生境条件共同决定[41 -42 ] .不同生活型植物之间不定根的分布和构型存在显著差异,同时,复杂的荒漠生境进一步加剧了根系形态的多样性. ...
... [
32 ]
ARF家族(AtARF4/AtARF6) 参与木质部中柱鞘细胞的启动 [6 ,35 ] OsPLT家族(OsPLT1/OsPLT2) OsPLT1正向调控不定根和侧根的起始和生长,OsPLT2控制侧根的形成但不影响不定根的发育 [36 ] miR156 维持植物的高可塑性,使植物保持较强的根系再生能力,以应对不确定的环境资源 [37 ] 蛋白质 过氧化物酶 诱导不定根形成,参与植物的各种生理过程,如抗氧化防御和木质素的形成等 [38 ] 多酚氧化酶 主要集中在根原基形成区,影响细胞分裂分化和根原基的启动 [39 ] 吲哚乙酸氧化酶 氧化和降解IAA,调节内源激素的含量,帮助触发和启动植物根原基 [40 ] <strong>2</strong> 不定根的形态特征 植物不定根的分布、形态及构型特征由遗传信息和生境条件共同决定[41 -42 ] .不同生活型植物之间不定根的分布和构型存在显著差异,同时,复杂的荒漠生境进一步加剧了根系形态的多样性. ...
The interaction between rice ERF3 and WOX11 promotes crown root development by regulating gene expression involved in cytokinin signaling
1
2015
... 生长素(IAA)在不定根发生过程中起核心调控作用,主要负责诱导根原基的形成[28 ] .在不定根的形成过程中,3-吲哚丁酸(IBA)可以转化为具有活性的IAA[29 ] .此外,植物其余多种激素参与不定根的发育调控.例如细胞分裂素(CK)与生长素呈拮抗作用,抑制CK的表达可促进根原基细胞产生与不定根的伸长和突破[26 ] ;乙烯(ET)可通过调节基因表达,诱导皮层细胞死亡,帮助根原基细胞产生[30 ] .赤霉素(GA)在植物不定根发生的不同阶段可起到抑制或促进作用[26 ] ;茉莉酸(JA)对不定根进行负向调节,植物可通过调节OsJJAZ1/EG2基因来抑制茉莉酸的表达,从而促进根原基细胞的产生[31 ] .不定根的形成不仅是植物的生理响应过程,也受遗传基因严格控制.当植物体内激素积累时,相关基因会快速做出响应,发挥重要作用[32 -33 ] .在植物激素与基因调控不定根形成的过程中,多种关键蛋白质也参与不定根的形成和发育. ...
Genetic control of adventitious rooting traits in bi-parental pedigree of Eucalyptus tereticornis ?×?E. camaldulensis
1
2021
... Regulation factors related to adventitious roots
Table 1 调控机制 调控因子 功能及作用机制 参考文献 植物激素 生长素(IAA) 调控基因表达,促进细胞分裂分化,诱导根原基形成,促使不定根产生 [26 -28 ] 细胞分裂素(CK) 与生长素拮抗,高浓度抑制根原基形成 [26 -27 ,31 ] 乙烯(ET) 诱导皮层细胞死亡,形成通气组织,帮助根原基细胞产生,但可能在不定根诱导后期负向调控 [26 ,30 -31 ] 独脚金内酯(SL) 促进单子叶植物不定根伸长,植物受养分胁迫或机械损伤时,调控根系发育 [31 ] 茉莉酸(JA) 负向调控,过表达茉莉酸的负向调节因子可以促进植物产生更长、更多的不定根 [26 ,31 ] 脱落酸(ABA) 对水稻冠根生长有抑制作用 [27 ,31 ] 赤霉素(GA) 通常抑制不定根发育,但其和乙烯一起可促进皮层细胞死亡,促进不定根出现 [26 -27 ,31 ] 转录因子 WOX家族(WOX10/WOX11) 受生长素调控,建立根原基细胞 [32 -34 ] LBD家族(AtLBD18) 调控不定根的发生 [32 ] ARF家族(AtARF4/AtARF6) 参与木质部中柱鞘细胞的启动 [6 ,35 ] OsPLT家族(OsPLT1/OsPLT2) OsPLT1正向调控不定根和侧根的起始和生长,OsPLT2控制侧根的形成但不影响不定根的发育 [36 ] miR156 维持植物的高可塑性,使植物保持较强的根系再生能力,以应对不确定的环境资源 [37 ] 蛋白质 过氧化物酶 诱导不定根形成,参与植物的各种生理过程,如抗氧化防御和木质素的形成等 [38 ] 多酚氧化酶 主要集中在根原基形成区,影响细胞分裂分化和根原基的启动 [39 ] 吲哚乙酸氧化酶 氧化和降解IAA,调节内源激素的含量,帮助触发和启动植物根原基 [40 ]
<strong>2</strong> 不定根的形态特征 植物不定根的分布、形态及构型特征由遗传信息和生境条件共同决定[41 -42 ] .不同生活型植物之间不定根的分布和构型存在显著差异,同时,复杂的荒漠生境进一步加剧了根系形态的多样性. ...
ARL1,a LOB-domain protein required for adventitious root formation in rice
1
2005
... Regulation factors related to adventitious roots
Table 1 调控机制 调控因子 功能及作用机制 参考文献 植物激素 生长素(IAA) 调控基因表达,促进细胞分裂分化,诱导根原基形成,促使不定根产生 [26 -28 ] 细胞分裂素(CK) 与生长素拮抗,高浓度抑制根原基形成 [26 -27 ,31 ] 乙烯(ET) 诱导皮层细胞死亡,形成通气组织,帮助根原基细胞产生,但可能在不定根诱导后期负向调控 [26 ,30 -31 ] 独脚金内酯(SL) 促进单子叶植物不定根伸长,植物受养分胁迫或机械损伤时,调控根系发育 [31 ] 茉莉酸(JA) 负向调控,过表达茉莉酸的负向调节因子可以促进植物产生更长、更多的不定根 [26 ,31 ] 脱落酸(ABA) 对水稻冠根生长有抑制作用 [27 ,31 ] 赤霉素(GA) 通常抑制不定根发育,但其和乙烯一起可促进皮层细胞死亡,促进不定根出现 [26 -27 ,31 ] 转录因子 WOX家族(WOX10/WOX11) 受生长素调控,建立根原基细胞 [32 -34 ] LBD家族(AtLBD18) 调控不定根的发生 [32 ] ARF家族(AtARF4/AtARF6) 参与木质部中柱鞘细胞的启动 [6 ,35 ] OsPLT家族(OsPLT1/OsPLT2) OsPLT1正向调控不定根和侧根的起始和生长,OsPLT2控制侧根的形成但不影响不定根的发育 [36 ] miR156 维持植物的高可塑性,使植物保持较强的根系再生能力,以应对不确定的环境资源 [37 ] 蛋白质 过氧化物酶 诱导不定根形成,参与植物的各种生理过程,如抗氧化防御和木质素的形成等 [38 ] 多酚氧化酶 主要集中在根原基形成区,影响细胞分裂分化和根原基的启动 [39 ] 吲哚乙酸氧化酶 氧化和降解IAA,调节内源激素的含量,帮助触发和启动植物根原基 [40 ]
<strong>2</strong> 不定根的形态特征 植物不定根的分布、形态及构型特征由遗传信息和生境条件共同决定[41 -42 ] .不同生活型植物之间不定根的分布和构型存在显著差异,同时,复杂的荒漠生境进一步加剧了根系形态的多样性. ...
Species-specific function of conserved regulators in orchestrating rice root architecture
1
2022
... Regulation factors related to adventitious roots
Table 1 调控机制 调控因子 功能及作用机制 参考文献 植物激素 生长素(IAA) 调控基因表达,促进细胞分裂分化,诱导根原基形成,促使不定根产生 [26 -28 ] 细胞分裂素(CK) 与生长素拮抗,高浓度抑制根原基形成 [26 -27 ,31 ] 乙烯(ET) 诱导皮层细胞死亡,形成通气组织,帮助根原基细胞产生,但可能在不定根诱导后期负向调控 [26 ,30 -31 ] 独脚金内酯(SL) 促进单子叶植物不定根伸长,植物受养分胁迫或机械损伤时,调控根系发育 [31 ] 茉莉酸(JA) 负向调控,过表达茉莉酸的负向调节因子可以促进植物产生更长、更多的不定根 [26 ,31 ] 脱落酸(ABA) 对水稻冠根生长有抑制作用 [27 ,31 ] 赤霉素(GA) 通常抑制不定根发育,但其和乙烯一起可促进皮层细胞死亡,促进不定根出现 [26 -27 ,31 ] 转录因子 WOX家族(WOX10/WOX11) 受生长素调控,建立根原基细胞 [32 -34 ] LBD家族(AtLBD18) 调控不定根的发生 [32 ] ARF家族(AtARF4/AtARF6) 参与木质部中柱鞘细胞的启动 [6 ,35 ] OsPLT家族(OsPLT1/OsPLT2) OsPLT1正向调控不定根和侧根的起始和生长,OsPLT2控制侧根的形成但不影响不定根的发育 [36 ] miR156 维持植物的高可塑性,使植物保持较强的根系再生能力,以应对不确定的环境资源 [37 ] 蛋白质 过氧化物酶 诱导不定根形成,参与植物的各种生理过程,如抗氧化防御和木质素的形成等 [38 ] 多酚氧化酶 主要集中在根原基形成区,影响细胞分裂分化和根原基的启动 [39 ] 吲哚乙酸氧化酶 氧化和降解IAA,调节内源激素的含量,帮助触发和启动植物根原基 [40 ]
<strong>2</strong> 不定根的形态特征 植物不定根的分布、形态及构型特征由遗传信息和生境条件共同决定[41 -42 ] .不同生活型植物之间不定根的分布和构型存在显著差异,同时,复杂的荒漠生境进一步加剧了根系形态的多样性. ...
Understanding of adventitious root formation: What can we learn from comparative genetics
1
2020
... Regulation factors related to adventitious roots
Table 1 调控机制 调控因子 功能及作用机制 参考文献 植物激素 生长素(IAA) 调控基因表达,促进细胞分裂分化,诱导根原基形成,促使不定根产生 [26 -28 ] 细胞分裂素(CK) 与生长素拮抗,高浓度抑制根原基形成 [26 -27 ,31 ] 乙烯(ET) 诱导皮层细胞死亡,形成通气组织,帮助根原基细胞产生,但可能在不定根诱导后期负向调控 [26 ,30 -31 ] 独脚金内酯(SL) 促进单子叶植物不定根伸长,植物受养分胁迫或机械损伤时,调控根系发育 [31 ] 茉莉酸(JA) 负向调控,过表达茉莉酸的负向调节因子可以促进植物产生更长、更多的不定根 [26 ,31 ] 脱落酸(ABA) 对水稻冠根生长有抑制作用 [27 ,31 ] 赤霉素(GA) 通常抑制不定根发育,但其和乙烯一起可促进皮层细胞死亡,促进不定根出现 [26 -27 ,31 ] 转录因子 WOX家族(WOX10/WOX11) 受生长素调控,建立根原基细胞 [32 -34 ] LBD家族(AtLBD18) 调控不定根的发生 [32 ] ARF家族(AtARF4/AtARF6) 参与木质部中柱鞘细胞的启动 [6 ,35 ] OsPLT家族(OsPLT1/OsPLT2) OsPLT1正向调控不定根和侧根的起始和生长,OsPLT2控制侧根的形成但不影响不定根的发育 [36 ] miR156 维持植物的高可塑性,使植物保持较强的根系再生能力,以应对不确定的环境资源 [37 ] 蛋白质 过氧化物酶 诱导不定根形成,参与植物的各种生理过程,如抗氧化防御和木质素的形成等 [38 ] 多酚氧化酶 主要集中在根原基形成区,影响细胞分裂分化和根原基的启动 [39 ] 吲哚乙酸氧化酶 氧化和降解IAA,调节内源激素的含量,帮助触发和启动植物根原基 [40 ]
<strong>2</strong> 不定根的形态特征 植物不定根的分布、形态及构型特征由遗传信息和生境条件共同决定[41 -42 ] .不同生活型植物之间不定根的分布和构型存在显著差异,同时,复杂的荒漠生境进一步加剧了根系形态的多样性. ...
Practical uses of peroxidase activity as a predictive marker of rooting performance of micropropagated shoots
1
1992
... Regulation factors related to adventitious roots
Table 1 调控机制 调控因子 功能及作用机制 参考文献 植物激素 生长素(IAA) 调控基因表达,促进细胞分裂分化,诱导根原基形成,促使不定根产生 [26 -28 ] 细胞分裂素(CK) 与生长素拮抗,高浓度抑制根原基形成 [26 -27 ,31 ] 乙烯(ET) 诱导皮层细胞死亡,形成通气组织,帮助根原基细胞产生,但可能在不定根诱导后期负向调控 [26 ,30 -31 ] 独脚金内酯(SL) 促进单子叶植物不定根伸长,植物受养分胁迫或机械损伤时,调控根系发育 [31 ] 茉莉酸(JA) 负向调控,过表达茉莉酸的负向调节因子可以促进植物产生更长、更多的不定根 [26 ,31 ] 脱落酸(ABA) 对水稻冠根生长有抑制作用 [27 ,31 ] 赤霉素(GA) 通常抑制不定根发育,但其和乙烯一起可促进皮层细胞死亡,促进不定根出现 [26 -27 ,31 ] 转录因子 WOX家族(WOX10/WOX11) 受生长素调控,建立根原基细胞 [32 -34 ] LBD家族(AtLBD18) 调控不定根的发生 [32 ] ARF家族(AtARF4/AtARF6) 参与木质部中柱鞘细胞的启动 [6 ,35 ] OsPLT家族(OsPLT1/OsPLT2) OsPLT1正向调控不定根和侧根的起始和生长,OsPLT2控制侧根的形成但不影响不定根的发育 [36 ] miR156 维持植物的高可塑性,使植物保持较强的根系再生能力,以应对不确定的环境资源 [37 ] 蛋白质 过氧化物酶 诱导不定根形成,参与植物的各种生理过程,如抗氧化防御和木质素的形成等 [38 ] 多酚氧化酶 主要集中在根原基形成区,影响细胞分裂分化和根原基的启动 [39 ] 吲哚乙酸氧化酶 氧化和降解IAA,调节内源激素的含量,帮助触发和启动植物根原基 [40 ]
<strong>2</strong> 不定根的形态特征 植物不定根的分布、形态及构型特征由遗传信息和生境条件共同决定[41 -42 ] .不同生活型植物之间不定根的分布和构型存在显著差异,同时,复杂的荒漠生境进一步加剧了根系形态的多样性. ...
Rooting of herbaceous minicuttings of peach rootstock under effect of indolebutyric acid (IBA)
1
2015
... Regulation factors related to adventitious roots
Table 1 调控机制 调控因子 功能及作用机制 参考文献 植物激素 生长素(IAA) 调控基因表达,促进细胞分裂分化,诱导根原基形成,促使不定根产生 [26 -28 ] 细胞分裂素(CK) 与生长素拮抗,高浓度抑制根原基形成 [26 -27 ,31 ] 乙烯(ET) 诱导皮层细胞死亡,形成通气组织,帮助根原基细胞产生,但可能在不定根诱导后期负向调控 [26 ,30 -31 ] 独脚金内酯(SL) 促进单子叶植物不定根伸长,植物受养分胁迫或机械损伤时,调控根系发育 [31 ] 茉莉酸(JA) 负向调控,过表达茉莉酸的负向调节因子可以促进植物产生更长、更多的不定根 [26 ,31 ] 脱落酸(ABA) 对水稻冠根生长有抑制作用 [27 ,31 ] 赤霉素(GA) 通常抑制不定根发育,但其和乙烯一起可促进皮层细胞死亡,促进不定根出现 [26 -27 ,31 ] 转录因子 WOX家族(WOX10/WOX11) 受生长素调控,建立根原基细胞 [32 -34 ] LBD家族(AtLBD18) 调控不定根的发生 [32 ] ARF家族(AtARF4/AtARF6) 参与木质部中柱鞘细胞的启动 [6 ,35 ] OsPLT家族(OsPLT1/OsPLT2) OsPLT1正向调控不定根和侧根的起始和生长,OsPLT2控制侧根的形成但不影响不定根的发育 [36 ] miR156 维持植物的高可塑性,使植物保持较强的根系再生能力,以应对不确定的环境资源 [37 ] 蛋白质 过氧化物酶 诱导不定根形成,参与植物的各种生理过程,如抗氧化防御和木质素的形成等 [38 ] 多酚氧化酶 主要集中在根原基形成区,影响细胞分裂分化和根原基的启动 [39 ] 吲哚乙酸氧化酶 氧化和降解IAA,调节内源激素的含量,帮助触发和启动植物根原基 [40 ]
<strong>2</strong> 不定根的形态特征 植物不定根的分布、形态及构型特征由遗传信息和生境条件共同决定[41 -42 ] .不同生活型植物之间不定根的分布和构型存在显著差异,同时,复杂的荒漠生境进一步加剧了根系形态的多样性. ...
Differential rooting and sprouting behaviour of two Jatropha species and associated physiological and biochemical changes
2
2005
... 不定根的形成是一个多因子协同调控的复杂生物学过程[26 -28 ,30 -40 ] (表1 ).然而,与水稻、玉米和桉树等经济作物和经济林树种相比,沙生灌木在不定根的形成能力和调控机制方面具有显著差异.作为干旱、半干旱区荒漠生态系统的主要植被类型,沙生灌木长期处于沙埋、水分和养分等环境胁迫条件下,通常需要依靠快速形成不定根来维持水分和养分的吸收与固定,因此,其不定根的形成条件和发育机制等可能与上述经济作物存在显著差异,具有一定的特殊性.然而,现有关于不定根形成机制的研究主要集中于桉树、水稻、玉米等经济物种,针对沙生灌木的系统研究相对匮乏.对于沙生灌木在沙埋、干旱等特殊生境中,不定根诱导过程中激素动态、关键基因及蛋白质调控网络仍缺乏深入了解,还存在一系列问题待我们深入探讨.例如,沙生灌木中激素互作对不定根形成的调控路径是否不同于经济林和作物,是否存在特殊的IBA-IAA转化机制,是否具有响应沙地环境的特异转录因子和蛋白质表达模式.未来亟需进一步系统研究,以揭示沙生灌木独特的胁迫适应机制并为干旱区植被恢复与育种提供理论依据. ...
... Regulation factors related to adventitious roots
Table 1 调控机制 调控因子 功能及作用机制 参考文献 植物激素 生长素(IAA) 调控基因表达,促进细胞分裂分化,诱导根原基形成,促使不定根产生 [26 -28 ] 细胞分裂素(CK) 与生长素拮抗,高浓度抑制根原基形成 [26 -27 ,31 ] 乙烯(ET) 诱导皮层细胞死亡,形成通气组织,帮助根原基细胞产生,但可能在不定根诱导后期负向调控 [26 ,30 -31 ] 独脚金内酯(SL) 促进单子叶植物不定根伸长,植物受养分胁迫或机械损伤时,调控根系发育 [31 ] 茉莉酸(JA) 负向调控,过表达茉莉酸的负向调节因子可以促进植物产生更长、更多的不定根 [26 ,31 ] 脱落酸(ABA) 对水稻冠根生长有抑制作用 [27 ,31 ] 赤霉素(GA) 通常抑制不定根发育,但其和乙烯一起可促进皮层细胞死亡,促进不定根出现 [26 -27 ,31 ] 转录因子 WOX家族(WOX10/WOX11) 受生长素调控,建立根原基细胞 [32 -34 ] LBD家族(AtLBD18) 调控不定根的发生 [32 ] ARF家族(AtARF4/AtARF6) 参与木质部中柱鞘细胞的启动 [6 ,35 ] OsPLT家族(OsPLT1/OsPLT2) OsPLT1正向调控不定根和侧根的起始和生长,OsPLT2控制侧根的形成但不影响不定根的发育 [36 ] miR156 维持植物的高可塑性,使植物保持较强的根系再生能力,以应对不确定的环境资源 [37 ] 蛋白质 过氧化物酶 诱导不定根形成,参与植物的各种生理过程,如抗氧化防御和木质素的形成等 [38 ] 多酚氧化酶 主要集中在根原基形成区,影响细胞分裂分化和根原基的启动 [39 ] 吲哚乙酸氧化酶 氧化和降解IAA,调节内源激素的含量,帮助触发和启动植物根原基 [40 ]
<strong>2</strong> 不定根的形态特征 植物不定根的分布、形态及构型特征由遗传信息和生境条件共同决定[41 -42 ] .不同生活型植物之间不定根的分布和构型存在显著差异,同时,复杂的荒漠生境进一步加剧了根系形态的多样性. ...
Quantifying root extracts and exudates of sedge and shrub in relation to root morphology
1
2017
... 植物不定根的分布、形态及构型特征由遗传信息和生境条件共同决定[41 -42 ] .不同生活型植物之间不定根的分布和构型存在显著差异,同时,复杂的荒漠生境进一步加剧了根系形态的多样性. ...
Root-soil physical and biotic interactions with a focus on tree root systems:a review
1
2018
... 植物不定根的分布、形态及构型特征由遗传信息和生境条件共同决定[41 -42 ] .不同生活型植物之间不定根的分布和构型存在显著差异,同时,复杂的荒漠生境进一步加剧了根系形态的多样性. ...
Using anatomical traits to understand root functions across root orders of herbaceous species in a temperate steppe
1
2022
... 根系解剖学研究可以揭示植物根系形态与功能之间的紧密联系,皮层通过直接吸收或借助菌根真菌间接吸收养分和水分,其厚度可反映根系的资源吸收能力;中柱负责水分和养分的向上运输和光合产物的向下运输,其直径表征根系的运输能力[43 ] .安宁等[44 ] 通过解剖不定根的结构发现,沙生灌木不定根直径和中柱面积均随根序的增加而增大,即不定根的运输能力均随根序的增加而加强.此外,不定根和原生根根系解剖结构上存在显著差异.不定根的直径、中柱直径和根组织密度均小于原生根,但比根长和比表面积显著高于原生根,这使得不定根能在较低生物量投资下实现水分和养分的快速吸收,然而由于不定根直径小,根组织密度低,木质化程度较弱,其在机械胁迫、干旱沙埋或病原菌侵染等环境变化下的防御性和寿命均弱于原生根.相反,原生根具有较大的直径和中柱直径,根组织密度更高,其结构更为坚固,运输组织发达,有利于维持长期的物质运输与结构稳定性.可见,不定根这种“快速周转型”的资源获取策略,能够优先满足短期资源需求,但同时也牺牲了长期结构稳定性.而原生根则通过“防御保守型”策略,将有限资源分配给结构性特征而非代谢活性组织,使其在持续胁迫下维持功能完整性. ...
不定根发育对典型荒漠灌木白刺光合作用的影响
3
2025
... 根系解剖学研究可以揭示植物根系形态与功能之间的紧密联系,皮层通过直接吸收或借助菌根真菌间接吸收养分和水分,其厚度可反映根系的资源吸收能力;中柱负责水分和养分的向上运输和光合产物的向下运输,其直径表征根系的运输能力[43 ] .安宁等[44 ] 通过解剖不定根的结构发现,沙生灌木不定根直径和中柱面积均随根序的增加而增大,即不定根的运输能力均随根序的增加而加强.此外,不定根和原生根根系解剖结构上存在显著差异.不定根的直径、中柱直径和根组织密度均小于原生根,但比根长和比表面积显著高于原生根,这使得不定根能在较低生物量投资下实现水分和养分的快速吸收,然而由于不定根直径小,根组织密度低,木质化程度较弱,其在机械胁迫、干旱沙埋或病原菌侵染等环境变化下的防御性和寿命均弱于原生根.相反,原生根具有较大的直径和中柱直径,根组织密度更高,其结构更为坚固,运输组织发达,有利于维持长期的物质运输与结构稳定性.可见,不定根这种“快速周转型”的资源获取策略,能够优先满足短期资源需求,但同时也牺牲了长期结构稳定性.而原生根则通过“防御保守型”策略,将有限资源分配给结构性特征而非代谢活性组织,使其在持续胁迫下维持功能完整性. ...
... 沙埋也是限制荒漠植物不定根生长和分布的关键因素,与根系形态特征紧密相关[51 ] .沙埋促进灌丛“肥岛效应”形成,造成沙堆土壤和丘间低地土壤养分分布不均[52 ] .沙埋导致植物产生不定根且不定根根径、长度及生物量与土壤有机碳、氮、磷含量显著正相关[44 ,52 ] .在沙埋干扰下,许多沙生灌木被埋枝叶通过分解纤维素产生可溶性糖和淀粉,为不定根生长提供能量[53 ] ,且不定根生物量占比随着沙埋深度的加剧而增大,如沙鞭、白刺等荒漠植物[7 -8 , 11 ] . ...
... 土壤养分对根系发育的影响和水分胁迫相似[61 -62 ] .在养分胁迫条件下,植物会增加不定根的数量来提高对表层磷的吸收和利用,且不定根根长与土壤总磷含量显著正相关,这是由于磷在土壤中移动缓慢,易与土壤颗粒结合,有效性低且分布不均,因此植物通过增加根系长度和密度,同时发育根毛来扩大吸收面积,从而增强对磷的吸收能力[63 ] .此外,植物通过增加不定根的生长角度(重力响应度)来探索深层土壤中的氮[23 ,45 ,64 -65 ] .在盐碱胁迫下,仙人掌(Echinocactus platyacanthus )幼苗仍能持续发育不定根,表明不定根有助于增强植物的抗逆性[66 ] .安宁等[44 ] 的研究也表明,在低水分条件下,氮磷添加可提高不定根的比根长和比表面积,而在水分充足条件下,氮磷添加反而抑制了不定根的形态特征变化,说明荒漠植物根系具有较强的可塑性,不同物种对水分和养分变化的响应存在显著差异.因此,不定根的存在有助于植物适应贫瘠或养分异质性较大的土壤生境,在低养分条件下,不定根的养分吸收能力高于原生根,而在高养分条件下,二者差异不明显[67 ] .总之,不定根的发育极大地提高了植物在胁迫生境中的养分获取能力,是植物适应极端生境的重要策略[68 ] . ...
Contribution of adventitious vs initial roots to growth and physiology of black spruce seedlings
4
2019
... 不定根的形态和构型与植物水分的获取途径密切相关,是其生态适应性的最直观体现.在轻度干旱胁迫下,植物会增加不定根的数量来提高水分获取效率[45 -46 ] ,且不定根的直径与土壤水分显著正相关,表明当短暂的降水发生时,植物会增加根系直径以增强水分运输效率[47 -48 ] .研究表明,沙生灌木白刺根系呈二态性分布[21 ] ,即在降水较为充足的季节或月份,白刺利用不定根吸收小降水事件补充的浅层土壤水;在降水较少或浅层土壤水匮乏时,转而利用原生根吸收由较大降水事件补给的深层土壤水分及地下水以满足其正常生长发育需求[49 ] .这是由于在干旱-半干旱生态系统中,降水事件具有高度不确定性和脉冲性,浅层土壤水分动态波动剧烈.植物可以通过快速产生不定根,来高效捕获降水后浅层土壤的短暂水分.然而,强烈的地表蒸发使得这些水分迅速散失,导致不定根的功能窗口期很短.当浅层资源难以持续供给时,植物则将资源投入转向深层根系构建,以获取更稳定的深层土壤水或地下水,从而保障长期的生存与繁殖[50 ] . ...
... 关于水分对荒漠植物根系生长发育影响的研究已有很多.研究表明,在同一生境中,有不定根生长的幼苗受干旱胁迫的程度远低于无不定根的幼苗[45 -46 ] .在干旱胁迫下,不定根的存在增强了幼苗的水分吸收能力,使其净光合速率和水势均显著高于无不定根幼苗,提高了幼苗的存活率并增加了干旱生境中植物自然更新的可能性[46 ] .Pernot等[45 ] 在研究黑云杉(Picea mariana )幼苗时发现,具有不定根的幼苗在干旱条件下的净光合速率比仅具有原生根的幼苗高15%~25%,株高相对生长率提高24%.也有研究表明,不定根的水力导度和水通道蛋白表达能力是原生根的3~4倍,导致其水分吸收能力明显高于原生根[59 ] .除参与植物水分吸收和利用外,不定根还可调控土壤水文过程.例如,大量的不定根可促进沙堆土壤优先流发育,加速降水由表层向深层入渗,增强了白刺灌丛对极端干旱生境的适应能力[60 ] .因此,在一定程度的干旱胁迫下,不定根的存在可缓解水分胁迫对植物的负面影响,增强沙生植物对干旱胁迫的适应能力. ...
... [45 ]在研究黑云杉(Picea mariana )幼苗时发现,具有不定根的幼苗在干旱条件下的净光合速率比仅具有原生根的幼苗高15%~25%,株高相对生长率提高24%.也有研究表明,不定根的水力导度和水通道蛋白表达能力是原生根的3~4倍,导致其水分吸收能力明显高于原生根[59 ] .除参与植物水分吸收和利用外,不定根还可调控土壤水文过程.例如,大量的不定根可促进沙堆土壤优先流发育,加速降水由表层向深层入渗,增强了白刺灌丛对极端干旱生境的适应能力[60 ] .因此,在一定程度的干旱胁迫下,不定根的存在可缓解水分胁迫对植物的负面影响,增强沙生植物对干旱胁迫的适应能力. ...
... 土壤养分对根系发育的影响和水分胁迫相似[61 -62 ] .在养分胁迫条件下,植物会增加不定根的数量来提高对表层磷的吸收和利用,且不定根根长与土壤总磷含量显著正相关,这是由于磷在土壤中移动缓慢,易与土壤颗粒结合,有效性低且分布不均,因此植物通过增加根系长度和密度,同时发育根毛来扩大吸收面积,从而增强对磷的吸收能力[63 ] .此外,植物通过增加不定根的生长角度(重力响应度)来探索深层土壤中的氮[23 ,45 ,64 -65 ] .在盐碱胁迫下,仙人掌(Echinocactus platyacanthus )幼苗仍能持续发育不定根,表明不定根有助于增强植物的抗逆性[66 ] .安宁等[44 ] 的研究也表明,在低水分条件下,氮磷添加可提高不定根的比根长和比表面积,而在水分充足条件下,氮磷添加反而抑制了不定根的形态特征变化,说明荒漠植物根系具有较强的可塑性,不同物种对水分和养分变化的响应存在显著差异.因此,不定根的存在有助于植物适应贫瘠或养分异质性较大的土壤生境,在低养分条件下,不定根的养分吸收能力高于原生根,而在高养分条件下,二者差异不明显[67 ] .总之,不定根的发育极大地提高了植物在胁迫生境中的养分获取能力,是植物适应极端生境的重要策略[68 ] . ...
Early root growth plasticity in seedlings of three Mediterranean woody species
3
2007
... 不定根的形态和构型与植物水分的获取途径密切相关,是其生态适应性的最直观体现.在轻度干旱胁迫下,植物会增加不定根的数量来提高水分获取效率[45 -46 ] ,且不定根的直径与土壤水分显著正相关,表明当短暂的降水发生时,植物会增加根系直径以增强水分运输效率[47 -48 ] .研究表明,沙生灌木白刺根系呈二态性分布[21 ] ,即在降水较为充足的季节或月份,白刺利用不定根吸收小降水事件补充的浅层土壤水;在降水较少或浅层土壤水匮乏时,转而利用原生根吸收由较大降水事件补给的深层土壤水分及地下水以满足其正常生长发育需求[49 ] .这是由于在干旱-半干旱生态系统中,降水事件具有高度不确定性和脉冲性,浅层土壤水分动态波动剧烈.植物可以通过快速产生不定根,来高效捕获降水后浅层土壤的短暂水分.然而,强烈的地表蒸发使得这些水分迅速散失,导致不定根的功能窗口期很短.当浅层资源难以持续供给时,植物则将资源投入转向深层根系构建,以获取更稳定的深层土壤水或地下水,从而保障长期的生存与繁殖[50 ] . ...
... 关于水分对荒漠植物根系生长发育影响的研究已有很多.研究表明,在同一生境中,有不定根生长的幼苗受干旱胁迫的程度远低于无不定根的幼苗[45 -46 ] .在干旱胁迫下,不定根的存在增强了幼苗的水分吸收能力,使其净光合速率和水势均显著高于无不定根幼苗,提高了幼苗的存活率并增加了干旱生境中植物自然更新的可能性[46 ] .Pernot等[45 ] 在研究黑云杉(Picea mariana )幼苗时发现,具有不定根的幼苗在干旱条件下的净光合速率比仅具有原生根的幼苗高15%~25%,株高相对生长率提高24%.也有研究表明,不定根的水力导度和水通道蛋白表达能力是原生根的3~4倍,导致其水分吸收能力明显高于原生根[59 ] .除参与植物水分吸收和利用外,不定根还可调控土壤水文过程.例如,大量的不定根可促进沙堆土壤优先流发育,加速降水由表层向深层入渗,增强了白刺灌丛对极端干旱生境的适应能力[60 ] .因此,在一定程度的干旱胁迫下,不定根的存在可缓解水分胁迫对植物的负面影响,增强沙生植物对干旱胁迫的适应能力. ...
... [46 ].Pernot等[45 ] 在研究黑云杉(Picea mariana )幼苗时发现,具有不定根的幼苗在干旱条件下的净光合速率比仅具有原生根的幼苗高15%~25%,株高相对生长率提高24%.也有研究表明,不定根的水力导度和水通道蛋白表达能力是原生根的3~4倍,导致其水分吸收能力明显高于原生根[59 ] .除参与植物水分吸收和利用外,不定根还可调控土壤水文过程.例如,大量的不定根可促进沙堆土壤优先流发育,加速降水由表层向深层入渗,增强了白刺灌丛对极端干旱生境的适应能力[60 ] .因此,在一定程度的干旱胁迫下,不定根的存在可缓解水分胁迫对植物的负面影响,增强沙生植物对干旱胁迫的适应能力. ...
Influence of root system characteristics on black spruce seedling responses to limiting conditions
1
2019
... 不定根的形态和构型与植物水分的获取途径密切相关,是其生态适应性的最直观体现.在轻度干旱胁迫下,植物会增加不定根的数量来提高水分获取效率[45 -46 ] ,且不定根的直径与土壤水分显著正相关,表明当短暂的降水发生时,植物会增加根系直径以增强水分运输效率[47 -48 ] .研究表明,沙生灌木白刺根系呈二态性分布[21 ] ,即在降水较为充足的季节或月份,白刺利用不定根吸收小降水事件补充的浅层土壤水;在降水较少或浅层土壤水匮乏时,转而利用原生根吸收由较大降水事件补给的深层土壤水分及地下水以满足其正常生长发育需求[49 ] .这是由于在干旱-半干旱生态系统中,降水事件具有高度不确定性和脉冲性,浅层土壤水分动态波动剧烈.植物可以通过快速产生不定根,来高效捕获降水后浅层土壤的短暂水分.然而,强烈的地表蒸发使得这些水分迅速散失,导致不定根的功能窗口期很短.当浅层资源难以持续供给时,植物则将资源投入转向深层根系构建,以获取更稳定的深层土壤水或地下水,从而保障长期的生存与繁殖[50 ] . ...
Friendly neighbours: hydraulic redistribution accounts for one quarter of water used by neighbouring drought stressed tree saplings
1
2021
... 不定根的形态和构型与植物水分的获取途径密切相关,是其生态适应性的最直观体现.在轻度干旱胁迫下,植物会增加不定根的数量来提高水分获取效率[45 -46 ] ,且不定根的直径与土壤水分显著正相关,表明当短暂的降水发生时,植物会增加根系直径以增强水分运输效率[47 -48 ] .研究表明,沙生灌木白刺根系呈二态性分布[21 ] ,即在降水较为充足的季节或月份,白刺利用不定根吸收小降水事件补充的浅层土壤水;在降水较少或浅层土壤水匮乏时,转而利用原生根吸收由较大降水事件补给的深层土壤水分及地下水以满足其正常生长发育需求[49 ] .这是由于在干旱-半干旱生态系统中,降水事件具有高度不确定性和脉冲性,浅层土壤水分动态波动剧烈.植物可以通过快速产生不定根,来高效捕获降水后浅层土壤的短暂水分.然而,强烈的地表蒸发使得这些水分迅速散失,导致不定根的功能窗口期很短.当浅层资源难以持续供给时,植物则将资源投入转向深层根系构建,以获取更稳定的深层土壤水或地下水,从而保障长期的生存与繁殖[50 ] . ...
Water use trade-offs and optimal adaptations to pulse-driven arid ecosystems
1
2001
... 不定根的形态和构型与植物水分的获取途径密切相关,是其生态适应性的最直观体现.在轻度干旱胁迫下,植物会增加不定根的数量来提高水分获取效率[45 -46 ] ,且不定根的直径与土壤水分显著正相关,表明当短暂的降水发生时,植物会增加根系直径以增强水分运输效率[47 -48 ] .研究表明,沙生灌木白刺根系呈二态性分布[21 ] ,即在降水较为充足的季节或月份,白刺利用不定根吸收小降水事件补充的浅层土壤水;在降水较少或浅层土壤水匮乏时,转而利用原生根吸收由较大降水事件补给的深层土壤水分及地下水以满足其正常生长发育需求[49 ] .这是由于在干旱-半干旱生态系统中,降水事件具有高度不确定性和脉冲性,浅层土壤水分动态波动剧烈.植物可以通过快速产生不定根,来高效捕获降水后浅层土壤的短暂水分.然而,强烈的地表蒸发使得这些水分迅速散失,导致不定根的功能窗口期很短.当浅层资源难以持续供给时,植物则将资源投入转向深层根系构建,以获取更稳定的深层土壤水或地下水,从而保障长期的生存与繁殖[50 ] . ...
Biomass allocation to leaves,stems and roots:meta-analyses of interspecific variation and environmental control
1
2012
... 不定根的形态和构型与植物水分的获取途径密切相关,是其生态适应性的最直观体现.在轻度干旱胁迫下,植物会增加不定根的数量来提高水分获取效率[45 -46 ] ,且不定根的直径与土壤水分显著正相关,表明当短暂的降水发生时,植物会增加根系直径以增强水分运输效率[47 -48 ] .研究表明,沙生灌木白刺根系呈二态性分布[21 ] ,即在降水较为充足的季节或月份,白刺利用不定根吸收小降水事件补充的浅层土壤水;在降水较少或浅层土壤水匮乏时,转而利用原生根吸收由较大降水事件补给的深层土壤水分及地下水以满足其正常生长发育需求[49 ] .这是由于在干旱-半干旱生态系统中,降水事件具有高度不确定性和脉冲性,浅层土壤水分动态波动剧烈.植物可以通过快速产生不定根,来高效捕获降水后浅层土壤的短暂水分.然而,强烈的地表蒸发使得这些水分迅速散失,导致不定根的功能窗口期很短.当浅层资源难以持续供给时,植物则将资源投入转向深层根系构建,以获取更稳定的深层土壤水或地下水,从而保障长期的生存与繁殖[50 ] . ...
Variability in amount and frequency of water supply affects roots but not growth of arid shrubs
1
2009
... 沙埋也是限制荒漠植物不定根生长和分布的关键因素,与根系形态特征紧密相关[51 ] .沙埋促进灌丛“肥岛效应”形成,造成沙堆土壤和丘间低地土壤养分分布不均[52 ] .沙埋导致植物产生不定根且不定根根径、长度及生物量与土壤有机碳、氮、磷含量显著正相关[44 ,52 ] .在沙埋干扰下,许多沙生灌木被埋枝叶通过分解纤维素产生可溶性糖和淀粉,为不定根生长提供能量[53 ] ,且不定根生物量占比随着沙埋深度的加剧而增大,如沙鞭、白刺等荒漠植物[7 -8 , 11 ] . ...
白刺(Nitraria tangutorum )灌丛不定根构型特征及生物量分配模式
2
2022
... 沙埋也是限制荒漠植物不定根生长和分布的关键因素,与根系形态特征紧密相关[51 ] .沙埋促进灌丛“肥岛效应”形成,造成沙堆土壤和丘间低地土壤养分分布不均[52 ] .沙埋导致植物产生不定根且不定根根径、长度及生物量与土壤有机碳、氮、磷含量显著正相关[44 ,52 ] .在沙埋干扰下,许多沙生灌木被埋枝叶通过分解纤维素产生可溶性糖和淀粉,为不定根生长提供能量[53 ] ,且不定根生物量占比随着沙埋深度的加剧而增大,如沙鞭、白刺等荒漠植物[7 -8 , 11 ] . ...
... ,52 ].在沙埋干扰下,许多沙生灌木被埋枝叶通过分解纤维素产生可溶性糖和淀粉,为不定根生长提供能量[53 ] ,且不定根生物量占比随着沙埋深度的加剧而增大,如沙鞭、白刺等荒漠植物[7 -8 , 11 ] . ...
小叶锦鸡儿抗沙埋生长与抗氧化酶及同工酶变化的关系
2
2015
... 沙埋也是限制荒漠植物不定根生长和分布的关键因素,与根系形态特征紧密相关[51 ] .沙埋促进灌丛“肥岛效应”形成,造成沙堆土壤和丘间低地土壤养分分布不均[52 ] .沙埋导致植物产生不定根且不定根根径、长度及生物量与土壤有机碳、氮、磷含量显著正相关[44 ,52 ] .在沙埋干扰下,许多沙生灌木被埋枝叶通过分解纤维素产生可溶性糖和淀粉,为不定根生长提供能量[53 ] ,且不定根生物量占比随着沙埋深度的加剧而增大,如沙鞭、白刺等荒漠植物[7 -8 , 11 ] . ...
... 荒漠生态系统中强烈的风沙活动是驱动植物根系生长和发育的主要因素,当风沙流受植物阻拦时,沙物质在植物周围堆积,形成干旱-半干旱荒漠区常见的地貌类型——灌丛沙堆[1 ,56 -57 ] .这个过程使灌丛植被遭受一定程度的沙埋胁迫.不定根的生长可增强植物对浅层水分和养分的吸收能力以应对短期沙埋,维持植物的生理功能[53 ,58 ] .在沙埋干扰下,沙埋界面频繁的风沙活动及沙粒跃迁、撞击和摩擦,可使植物将资源重新分配至土壤表层的不定根和垂直茎上.不定根细胞排列更加紧密,木质化程度提高,从而增强了抗磨损能力和寿命,形成对干旱环境的适应性策略[8 ] .不定根的发育是沙生灌木适应风沙生境的重要策略,能够很好地缓解沙埋导致的资源胁迫. ...
达乌里胡枝子根系形态特征对土壤水分变化的响应
1
2014
... 总之,沙生灌木会通过产生不定根并不断调整其形态与构型,实现对浅层土壤水分和养分的快速吸收和固定,从而适应干旱、沙埋等胁迫生境.因此,根系形态特征可作为表征植物环境适应能力的重要指标[54 -55 ] . ...
白羊草根系形态特征对土壤水分阶段变化的响应
1
2016
... 总之,沙生灌木会通过产生不定根并不断调整其形态与构型,实现对浅层土壤水分和养分的快速吸收和固定,从而适应干旱、沙埋等胁迫生境.因此,根系形态特征可作为表征植物环境适应能力的重要指标[54 -55 ] . ...
A mechanism of self-organization in a desert with phytogenic mounds
1
2016
... 荒漠生态系统中强烈的风沙活动是驱动植物根系生长和发育的主要因素,当风沙流受植物阻拦时,沙物质在植物周围堆积,形成干旱-半干旱荒漠区常见的地貌类型——灌丛沙堆[1 ,56 -57 ] .这个过程使灌丛植被遭受一定程度的沙埋胁迫.不定根的生长可增强植物对浅层水分和养分的吸收能力以应对短期沙埋,维持植物的生理功能[53 ,58 ] .在沙埋干扰下,沙埋界面频繁的风沙活动及沙粒跃迁、撞击和摩擦,可使植物将资源重新分配至土壤表层的不定根和垂直茎上.不定根细胞排列更加紧密,木质化程度提高,从而增强了抗磨损能力和寿命,形成对干旱环境的适应性策略[8 ] .不定根的发育是沙生灌木适应风沙生境的重要策略,能够很好地缓解沙埋导致的资源胁迫. ...
干旱地区灌丛沙堆研究现状与展望
1
2010
... 荒漠生态系统中强烈的风沙活动是驱动植物根系生长和发育的主要因素,当风沙流受植物阻拦时,沙物质在植物周围堆积,形成干旱-半干旱荒漠区常见的地貌类型——灌丛沙堆[1 ,56 -57 ] .这个过程使灌丛植被遭受一定程度的沙埋胁迫.不定根的生长可增强植物对浅层水分和养分的吸收能力以应对短期沙埋,维持植物的生理功能[53 ,58 ] .在沙埋干扰下,沙埋界面频繁的风沙活动及沙粒跃迁、撞击和摩擦,可使植物将资源重新分配至土壤表层的不定根和垂直茎上.不定根细胞排列更加紧密,木质化程度提高,从而增强了抗磨损能力和寿命,形成对干旱环境的适应性策略[8 ] .不定根的发育是沙生灌木适应风沙生境的重要策略,能够很好地缓解沙埋导致的资源胁迫. ...
半干旱风沙区黄柳幼苗生长发育对沙埋的响应
1
2012
... 荒漠生态系统中强烈的风沙活动是驱动植物根系生长和发育的主要因素,当风沙流受植物阻拦时,沙物质在植物周围堆积,形成干旱-半干旱荒漠区常见的地貌类型——灌丛沙堆[1 ,56 -57 ] .这个过程使灌丛植被遭受一定程度的沙埋胁迫.不定根的生长可增强植物对浅层水分和养分的吸收能力以应对短期沙埋,维持植物的生理功能[53 ,58 ] .在沙埋干扰下,沙埋界面频繁的风沙活动及沙粒跃迁、撞击和摩擦,可使植物将资源重新分配至土壤表层的不定根和垂直茎上.不定根细胞排列更加紧密,木质化程度提高,从而增强了抗磨损能力和寿命,形成对干旱环境的适应性策略[8 ] .不定根的发育是沙生灌木适应风沙生境的重要策略,能够很好地缓解沙埋导致的资源胁迫. ...
Aquaporin-facilitated water uptake in barley (Hordeum vulgare L.) roots
1
2011
... 关于水分对荒漠植物根系生长发育影响的研究已有很多.研究表明,在同一生境中,有不定根生长的幼苗受干旱胁迫的程度远低于无不定根的幼苗[45 -46 ] .在干旱胁迫下,不定根的存在增强了幼苗的水分吸收能力,使其净光合速率和水势均显著高于无不定根幼苗,提高了幼苗的存活率并增加了干旱生境中植物自然更新的可能性[46 ] .Pernot等[45 ] 在研究黑云杉(Picea mariana )幼苗时发现,具有不定根的幼苗在干旱条件下的净光合速率比仅具有原生根的幼苗高15%~25%,株高相对生长率提高24%.也有研究表明,不定根的水力导度和水通道蛋白表达能力是原生根的3~4倍,导致其水分吸收能力明显高于原生根[59 ] .除参与植物水分吸收和利用外,不定根还可调控土壤水文过程.例如,大量的不定根可促进沙堆土壤优先流发育,加速降水由表层向深层入渗,增强了白刺灌丛对极端干旱生境的适应能力[60 ] .因此,在一定程度的干旱胁迫下,不定根的存在可缓解水分胁迫对植物的负面影响,增强沙生植物对干旱胁迫的适应能力. ...
Preferential flow in soils is key to the development of nebkhas in water limited regions
1
2025
... 关于水分对荒漠植物根系生长发育影响的研究已有很多.研究表明,在同一生境中,有不定根生长的幼苗受干旱胁迫的程度远低于无不定根的幼苗[45 -46 ] .在干旱胁迫下,不定根的存在增强了幼苗的水分吸收能力,使其净光合速率和水势均显著高于无不定根幼苗,提高了幼苗的存活率并增加了干旱生境中植物自然更新的可能性[46 ] .Pernot等[45 ] 在研究黑云杉(Picea mariana )幼苗时发现,具有不定根的幼苗在干旱条件下的净光合速率比仅具有原生根的幼苗高15%~25%,株高相对生长率提高24%.也有研究表明,不定根的水力导度和水通道蛋白表达能力是原生根的3~4倍,导致其水分吸收能力明显高于原生根[59 ] .除参与植物水分吸收和利用外,不定根还可调控土壤水文过程.例如,大量的不定根可促进沙堆土壤优先流发育,加速降水由表层向深层入渗,增强了白刺灌丛对极端干旱生境的适应能力[60 ] .因此,在一定程度的干旱胁迫下,不定根的存在可缓解水分胁迫对植物的负面影响,增强沙生植物对干旱胁迫的适应能力. ...
土壤水分胁迫对红砂幼苗细根形态和功能特征的影响
1
2014
... 土壤养分对根系发育的影响和水分胁迫相似[61 -62 ] .在养分胁迫条件下,植物会增加不定根的数量来提高对表层磷的吸收和利用,且不定根根长与土壤总磷含量显著正相关,这是由于磷在土壤中移动缓慢,易与土壤颗粒结合,有效性低且分布不均,因此植物通过增加根系长度和密度,同时发育根毛来扩大吸收面积,从而增强对磷的吸收能力[63 ] .此外,植物通过增加不定根的生长角度(重力响应度)来探索深层土壤中的氮[23 ,45 ,64 -65 ] .在盐碱胁迫下,仙人掌(Echinocactus platyacanthus )幼苗仍能持续发育不定根,表明不定根有助于增强植物的抗逆性[66 ] .安宁等[44 ] 的研究也表明,在低水分条件下,氮磷添加可提高不定根的比根长和比表面积,而在水分充足条件下,氮磷添加反而抑制了不定根的形态特征变化,说明荒漠植物根系具有较强的可塑性,不同物种对水分和养分变化的响应存在显著差异.因此,不定根的存在有助于植物适应贫瘠或养分异质性较大的土壤生境,在低养分条件下,不定根的养分吸收能力高于原生根,而在高养分条件下,二者差异不明显[67 ] .总之,不定根的发育极大地提高了植物在胁迫生境中的养分获取能力,是植物适应极端生境的重要策略[68 ] . ...
武功山芒根系垂直分布及其与土壤养分的关系
1
2017
... 土壤养分对根系发育的影响和水分胁迫相似[61 -62 ] .在养分胁迫条件下,植物会增加不定根的数量来提高对表层磷的吸收和利用,且不定根根长与土壤总磷含量显著正相关,这是由于磷在土壤中移动缓慢,易与土壤颗粒结合,有效性低且分布不均,因此植物通过增加根系长度和密度,同时发育根毛来扩大吸收面积,从而增强对磷的吸收能力[63 ] .此外,植物通过增加不定根的生长角度(重力响应度)来探索深层土壤中的氮[23 ,45 ,64 -65 ] .在盐碱胁迫下,仙人掌(Echinocactus platyacanthus )幼苗仍能持续发育不定根,表明不定根有助于增强植物的抗逆性[66 ] .安宁等[44 ] 的研究也表明,在低水分条件下,氮磷添加可提高不定根的比根长和比表面积,而在水分充足条件下,氮磷添加反而抑制了不定根的形态特征变化,说明荒漠植物根系具有较强的可塑性,不同物种对水分和养分变化的响应存在显著差异.因此,不定根的存在有助于植物适应贫瘠或养分异质性较大的土壤生境,在低养分条件下,不定根的养分吸收能力高于原生根,而在高养分条件下,二者差异不明显[67 ] .总之,不定根的发育极大地提高了植物在胁迫生境中的养分获取能力,是植物适应极端生境的重要策略[68 ] . ...
Environmental control of root system biology
1
2016
... 土壤养分对根系发育的影响和水分胁迫相似[61 -62 ] .在养分胁迫条件下,植物会增加不定根的数量来提高对表层磷的吸收和利用,且不定根根长与土壤总磷含量显著正相关,这是由于磷在土壤中移动缓慢,易与土壤颗粒结合,有效性低且分布不均,因此植物通过增加根系长度和密度,同时发育根毛来扩大吸收面积,从而增强对磷的吸收能力[63 ] .此外,植物通过增加不定根的生长角度(重力响应度)来探索深层土壤中的氮[23 ,45 ,64 -65 ] .在盐碱胁迫下,仙人掌(Echinocactus platyacanthus )幼苗仍能持续发育不定根,表明不定根有助于增强植物的抗逆性[66 ] .安宁等[44 ] 的研究也表明,在低水分条件下,氮磷添加可提高不定根的比根长和比表面积,而在水分充足条件下,氮磷添加反而抑制了不定根的形态特征变化,说明荒漠植物根系具有较强的可塑性,不同物种对水分和养分变化的响应存在显著差异.因此,不定根的存在有助于植物适应贫瘠或养分异质性较大的土壤生境,在低养分条件下,不定根的养分吸收能力高于原生根,而在高养分条件下,二者差异不明显[67 ] .总之,不定根的发育极大地提高了植物在胁迫生境中的养分获取能力,是植物适应极端生境的重要策略[68 ] . ...
Genotypic variation for root traits of maize (Zea mays L.) from the Purhepecha Plateau under contrasting phosphorus availability
1
2011
... 土壤养分对根系发育的影响和水分胁迫相似[61 -62 ] .在养分胁迫条件下,植物会增加不定根的数量来提高对表层磷的吸收和利用,且不定根根长与土壤总磷含量显著正相关,这是由于磷在土壤中移动缓慢,易与土壤颗粒结合,有效性低且分布不均,因此植物通过增加根系长度和密度,同时发育根毛来扩大吸收面积,从而增强对磷的吸收能力[63 ] .此外,植物通过增加不定根的生长角度(重力响应度)来探索深层土壤中的氮[23 ,45 ,64 -65 ] .在盐碱胁迫下,仙人掌(Echinocactus platyacanthus )幼苗仍能持续发育不定根,表明不定根有助于增强植物的抗逆性[66 ] .安宁等[44 ] 的研究也表明,在低水分条件下,氮磷添加可提高不定根的比根长和比表面积,而在水分充足条件下,氮磷添加反而抑制了不定根的形态特征变化,说明荒漠植物根系具有较强的可塑性,不同物种对水分和养分变化的响应存在显著差异.因此,不定根的存在有助于植物适应贫瘠或养分异质性较大的土壤生境,在低养分条件下,不定根的养分吸收能力高于原生根,而在高养分条件下,二者差异不明显[67 ] .总之,不定根的发育极大地提高了植物在胁迫生境中的养分获取能力,是植物适应极端生境的重要策略[68 ] . ...
草原化荒漠植被草本层片植物对人工施加氮素的响应
1
2010
... 土壤养分对根系发育的影响和水分胁迫相似[61 -62 ] .在养分胁迫条件下,植物会增加不定根的数量来提高对表层磷的吸收和利用,且不定根根长与土壤总磷含量显著正相关,这是由于磷在土壤中移动缓慢,易与土壤颗粒结合,有效性低且分布不均,因此植物通过增加根系长度和密度,同时发育根毛来扩大吸收面积,从而增强对磷的吸收能力[63 ] .此外,植物通过增加不定根的生长角度(重力响应度)来探索深层土壤中的氮[23 ,45 ,64 -65 ] .在盐碱胁迫下,仙人掌(Echinocactus platyacanthus )幼苗仍能持续发育不定根,表明不定根有助于增强植物的抗逆性[66 ] .安宁等[44 ] 的研究也表明,在低水分条件下,氮磷添加可提高不定根的比根长和比表面积,而在水分充足条件下,氮磷添加反而抑制了不定根的形态特征变化,说明荒漠植物根系具有较强的可塑性,不同物种对水分和养分变化的响应存在显著差异.因此,不定根的存在有助于植物适应贫瘠或养分异质性较大的土壤生境,在低养分条件下,不定根的养分吸收能力高于原生根,而在高养分条件下,二者差异不明显[67 ] .总之,不定根的发育极大地提高了植物在胁迫生境中的养分获取能力,是植物适应极端生境的重要策略[68 ] . ...
Persistent adventitious and basal root development during salt stress tolerance in Echinocactus platyacanthus (Cactaceae) seedlings
1
2021
... 土壤养分对根系发育的影响和水分胁迫相似[61 -62 ] .在养分胁迫条件下,植物会增加不定根的数量来提高对表层磷的吸收和利用,且不定根根长与土壤总磷含量显著正相关,这是由于磷在土壤中移动缓慢,易与土壤颗粒结合,有效性低且分布不均,因此植物通过增加根系长度和密度,同时发育根毛来扩大吸收面积,从而增强对磷的吸收能力[63 ] .此外,植物通过增加不定根的生长角度(重力响应度)来探索深层土壤中的氮[23 ,45 ,64 -65 ] .在盐碱胁迫下,仙人掌(Echinocactus platyacanthus )幼苗仍能持续发育不定根,表明不定根有助于增强植物的抗逆性[66 ] .安宁等[44 ] 的研究也表明,在低水分条件下,氮磷添加可提高不定根的比根长和比表面积,而在水分充足条件下,氮磷添加反而抑制了不定根的形态特征变化,说明荒漠植物根系具有较强的可塑性,不同物种对水分和养分变化的响应存在显著差异.因此,不定根的存在有助于植物适应贫瘠或养分异质性较大的土壤生境,在低养分条件下,不定根的养分吸收能力高于原生根,而在高养分条件下,二者差异不明显[67 ] .总之,不定根的发育极大地提高了植物在胁迫生境中的养分获取能力,是植物适应极端生境的重要策略[68 ] . ...
Phenotypic plasticity of the maize root system in response to heterogeneous nitrogen availability
1
2014
... 土壤养分对根系发育的影响和水分胁迫相似[61 -62 ] .在养分胁迫条件下,植物会增加不定根的数量来提高对表层磷的吸收和利用,且不定根根长与土壤总磷含量显著正相关,这是由于磷在土壤中移动缓慢,易与土壤颗粒结合,有效性低且分布不均,因此植物通过增加根系长度和密度,同时发育根毛来扩大吸收面积,从而增强对磷的吸收能力[63 ] .此外,植物通过增加不定根的生长角度(重力响应度)来探索深层土壤中的氮[23 ,45 ,64 -65 ] .在盐碱胁迫下,仙人掌(Echinocactus platyacanthus )幼苗仍能持续发育不定根,表明不定根有助于增强植物的抗逆性[66 ] .安宁等[44 ] 的研究也表明,在低水分条件下,氮磷添加可提高不定根的比根长和比表面积,而在水分充足条件下,氮磷添加反而抑制了不定根的形态特征变化,说明荒漠植物根系具有较强的可塑性,不同物种对水分和养分变化的响应存在显著差异.因此,不定根的存在有助于植物适应贫瘠或养分异质性较大的土壤生境,在低养分条件下,不定根的养分吸收能力高于原生根,而在高养分条件下,二者差异不明显[67 ] .总之,不定根的发育极大地提高了植物在胁迫生境中的养分获取能力,是植物适应极端生境的重要策略[68 ] . ...
Shrub growth improves morphological features of nebkhas:a case study of Nitraria tangutorum in the Tengger Desert
1
2024
... 土壤养分对根系发育的影响和水分胁迫相似[61 -62 ] .在养分胁迫条件下,植物会增加不定根的数量来提高对表层磷的吸收和利用,且不定根根长与土壤总磷含量显著正相关,这是由于磷在土壤中移动缓慢,易与土壤颗粒结合,有效性低且分布不均,因此植物通过增加根系长度和密度,同时发育根毛来扩大吸收面积,从而增强对磷的吸收能力[63 ] .此外,植物通过增加不定根的生长角度(重力响应度)来探索深层土壤中的氮[23 ,45 ,64 -65 ] .在盐碱胁迫下,仙人掌(Echinocactus platyacanthus )幼苗仍能持续发育不定根,表明不定根有助于增强植物的抗逆性[66 ] .安宁等[44 ] 的研究也表明,在低水分条件下,氮磷添加可提高不定根的比根长和比表面积,而在水分充足条件下,氮磷添加反而抑制了不定根的形态特征变化,说明荒漠植物根系具有较强的可塑性,不同物种对水分和养分变化的响应存在显著差异.因此,不定根的存在有助于植物适应贫瘠或养分异质性较大的土壤生境,在低养分条件下,不定根的养分吸收能力高于原生根,而在高养分条件下,二者差异不明显[67 ] .总之,不定根的发育极大地提高了植物在胁迫生境中的养分获取能力,是植物适应极端生境的重要策略[68 ] . ...
Belowground plant functional ecology: towards an integrated perspective
1
2018
... 在极端生境中,沙生灌木有性繁殖成功率较低,营养繁殖成为部分荒漠植物种群维持与扩张的重要方式.荒漠区常见的沙拐枣和沙鞭等植物在营养繁殖过程中,克隆分株会产生大量不定根,这些不定根对表层土壤水分和养分的高效吸收有助于克隆分株存活,直接影响无性系分株的定植与生长发育,对极端生境下植物的自然更新和种群的维持扩张至关重要[11 ,69 ] .研究表明,在沙丘生境中,白刺在遭遇沙埋后可快速萌发并长出不定根,不定根细根生物量(87.1%)显著高于原生根细根生物量(12.9%).在成熟灌丛沙堆中,不定根氮和钾含量显著高于原生根,且沙堆内土壤水分高于丘间低地,并被不定根吸收[7 ] .相比之下,种子萌发形成的实生苗由于原生根根系不发达,幼苗难以度过干旱期,制约了沙拐枣种群的自然更新.综上,不定根的产生促进了荒漠克隆植物的生长和繁殖,是干旱生境中克隆分株存活率远高于实生苗的重要原因[9 ,11 ] . ...





 甘公网安备 62010202000688号
甘公网安备 62010202000688号
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}