


0 引言
荒漠灌木根系是吸收水分、养分以及适应干旱生境的核心功能器官。细根(直径<2 mm)因比表面积大、代谢活性高,通常被视为水分和养分吸收的主要执行者[3]。细根还表现出高度的形态与生理可塑性。遇脉冲降水后可快速改变生长速率、方向和密度,以占据湿润土层并提高水分吸收速率[4];而在持续干旱下则促使浅层细根衰亡或进入低代谢状态,将有限碳资源优先分配给更木质化的深根或贮存器官,以降低代谢开销[5]。大量野外观测与实验研究表明,多数荒漠植物根系表现出显著的垂直分层格局,浅层细根密集形成网状结构,使植物能快速响应降雨脉冲以利用瞬时水资源;深层根系通常能深入地下数米以利用稳定的深层水源,在旱季承担稳定供水,保障植物长期存活[6-8]。在干旱区植物群落中,根系垂直分布常表现出显著的生态位分异,不同物种通过占据不同深度的土壤水分资源减少直接竞争,从而维持长期共存[9]。这类根系分层结构是荒漠生态系统构建稳定功能群的重要基础,可增强群落在水分波动环境下的整体抵抗力[10]。此外,荒漠灌木细根生长具有明显的季节性和对资源脉冲的响应性,雨季或融雪期为生长旺盛期,旱季则进入生长停滞或缓慢修复状态[11]。
近年来,微根窗作为一种原位、非破坏性的根系观测技术,在根系生态学研究中得到广泛应用。该技术通过在土壤中埋设透明管并定期拍摄根系影像,从而实现对根系生长过程的实时、重复、无损监测。微根窗技术不仅能够追踪根系的生长、凋亡与周转,还可结合降雨、土壤含水量等环境因子的同步观测,直接揭示根系对环境变化的动态响应[12]。同时,观测可在不同深度进行,便于解析根系的垂直分布动态变化。但微根窗方法仍存在一定缺陷,例如管道插入可能改变局部根的生长轨迹;在不同土壤湿润条件下图像质量存在波动,会影响根系识别的准确性[13]。依托微根窗法的大量研究表明,植物细根生长和死亡的深度格局随降水量与季节分布变化而变化,在湿润年份根系分布常出现向下延伸现象,而在干旱年份浅层占优势[14-15];时间尺度研究发现,根系的短期波动与土壤含水率的脉冲存在滞后性,降雨后表层细根会在数日内迅速增生以捕获水分,而中深层根的生长高峰可滞后数周甚至数月[16-17]。
在中国荒漠地区,柠条(Caragana korshinskii)和花棒(Hedysarum scoparium)是重要的固沙和水土保持植物,广泛用于防风固沙、植被恢复和生态修复等工程中。因其深根性、耐旱性和生长适应性,在黄土高原、内蒙古和甘肃等地区得到广泛应用。其根系在土壤中的分布和生长动态对其在干旱地区的生长和生态功能具有重要影响。尽管已有研究关注荒漠植物细根功能,但针对花棒和柠条在垂直梯度上细根分布与动态变化对土壤含水量的响应差异仍缺乏系统解析,二者在不同降水条件下的根系资源分配模式也未能得到关注。因此,探究两个物种细根伸长和分布的季节性和年度变化,以及细根动态对土壤水分变化的响应机制,不仅有利于理解其对于干旱胁迫的适应性策略,同时可为基于物种特性的精准植被配置与修复管理提供理论依据,为解释干旱区灌木群落的共存格局和资源分配提供关键支撑。
1 材料与方法
1.1 研究区概况
研究区位于甘肃河西走廊中部临泽县北部(39°21′N,100°07′E),地处绿洲-荒漠过渡带。试验依托中国科学院临泽内陆河流域研究站开展。该地区属于温带大陆性气候,多年平均降水量117 mm,夏季降水占全年降水的60%~70%。年平均气温为7.6 ℃,极端高温39.1 ℃,极端低温-27 ℃,≥10 ℃的年平均无霜期105 d。地带性土壤为灰棕漠土,并分布盐碱化土壤和风沙土。该区域天然植被的建群种为柠条锦鸡儿和花棒等荒漠灌木,草本层片主要由虫实(Corispermum chinganicum)、雾冰藜(Bassia dasyphylla)、沙蓬(Agriophyllum squarrosum)等物种组成。
1.2 试验设计
本研究选取荒漠区常见的2种灌木——花棒和柠条锦鸡儿作为观测物种,每个物种各选3棵冠幅相近且生长状态良好的植株,设置3个生物学重复可反映个体在自然条件下的基本变异范围,并满足独立样本t 检验和单因素方差分析(ANOVA)计算组内方差的最低要求。距离每棵植株主干50 cm处用配套土钻在所选区域内进行预打孔,钻孔与地面的夹角按照Johnson等[18]建议的与地面倾斜成60°安装。然后将内径5.0 cm、外径5.8 cm、长140 cm的微根窗管插入预打孔中,并用钻出的土填满管壁外侧,以保证管壁与周围土壤紧密接触。微根窗管下端用橡皮帽封死,上端露出地面20 cm,露出部分用黑色胶带缠绕并盖好防水管盖,防止光照射入管内对根系生长造成不利影响,以及避免灰尘、水分进入管内影响摄像头采集根系生长图片。微根管入土根管长约120 cm,垂向监测深度约为103 cm。
1.3 根系动态测定
2022—2024年生长季内,使用CI-600根扫描仪在300 dpi下进行根系拍摄,拍摄时保持曝光一致,以保证图片清晰度,每年拍摄4次。每根微根窗管需要拍摄6次,对应土层垂直深度为0~17、17~34、34~52、52~69、69~86、86~103 cm,共432张图片。用WinRHIZOTron Pro 2009 (Regent, Quebee, Canada)根系分析软件对获得的根系图像进行手动识别分析,得到每张图片中根系的总根长(L)。以单位时间内总根长的变化表示花棒或柠条的细根伸长率(X):
式中:X为花棒或柠条细根伸长率,单位为cm·d-1;ΔL为相邻2次观测期之间根系总长的增加量;T为2次观测的间隔天数。
1.4 土壤含水量
本研究中土壤含水量为土壤体积含水量,由中国科学院临泽内陆河流域研究站自动观测系统获取。该系统采用土壤水分监测系统(ECH2O EC-5土壤含水传感器+EM50数据采集器,METER Group, USA),分别在土壤垂直剖面20、40、60、80、100 cm深度水平埋入土壤水分传感器。数据采集器每1 h自动采集1次数据并存贮,以24 h观测数据的平均值作为每日土壤含水量数据。
1.5 数据分析
采用独立样本t检验对花棒与柠条伸长率差异显著性进行分析,采用单因素方差分析法(ANOVA)分别对两个物种在不同土层深度处的细根伸长率差异显著性进行分析,显著性水平设为0.05。使用Origin Pro 2021软件绘图。
2 结果
2.1 降水及土壤含水量的动态变化特征
2022—2024年降水均呈现不均衡分布(图1)。2022年降水量为109.6 mm,其中6—9月占比高达86%(94.2 mm),并伴随3次集中性强降雨事件(6月20日15 mm,7月14日9.1 mm,7月20日11 mm),根据《水文情报预报规范》可划分为平水年。相比之下,2023年和2024年降水量分别为83.6 mm和78.2 mm,可划分为枯水年,且降水更为分散或偏少,6—9月占比分别为61%(50.7 mm)和78%(60.72 mm)。整体来看,降水量逐年递减,水分条件逐渐趋紧。
图1
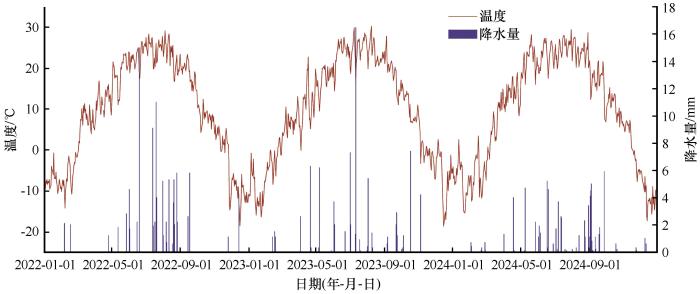
图1
2022—2024年试验样地气象状况
Fig.1
Meteorological conditions at the study site from 2022 to 2024
土壤含水量均呈现出明显的季节性波动,但仅0—40 cm土壤含水量对降水响应显著(P<0.05),且平水年与枯水年之间响应具有差异。在2022年(平水年)生长季集中性降水后,20 cm层处土壤含水量显著提升(P<0.05,图2),如7月20日降雨前后土壤含水量由3.25%升至9.81%,并在生长季维持较高水平;而40 cm层处土壤含水量仅表现出小幅提升,如7月20日降雨前后土壤含水量从4.87%升至5.48 %。在2023年与2024年(枯水年)生长季降雨后,20 cm层处土壤含水量提升幅度明显减小,小降水仅带来有限补给,例如2024年5月10日降雨后仅由1.40%升至1.81 %,且40 cm层处土壤含水量响应不显著。60~100 cm土层处土壤含水量主要受土壤热力过程与地下水补给调控,呈季节性波动,通常在冬季冻结期达到最低值,而在夏季融化且降水集中期达到最高值:三层最低值分别为0.11%、2.71%、3.75%,最高值分别为4.58%、5.81%、7.38%。
图2
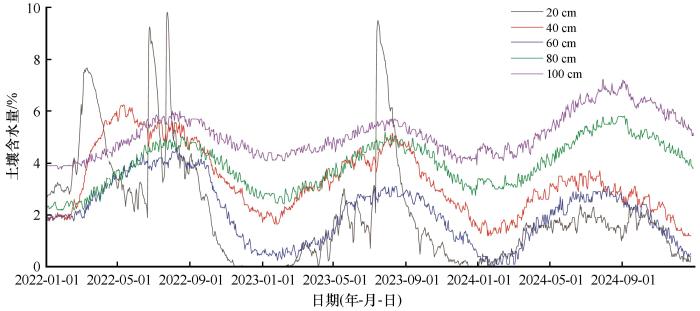
图2
2022—2024年试验样地土壤含水量动态
Fig.2
Dynamics of soil water content in the experimental plot from 2022 to 2024
2.2 细根分布的时空差异
花棒与柠条的细根在103 cm土层内均表现出明显的垂直分层特征,但二者的分布深度与动态变化存在差异。花棒细根主要集中在34~69 cm处,其分布随季节与降水状况呈现一定可塑性。柠条细根分布更深,集中于52~86 cm处且垂直分布较稳定,年内与年际变化幅度较小。
平水年中,两种灌木的细根垂直分布均表现出较明显的年内动态变化(图3)。2022年8月前花棒细根主要分布在52~86 cm土层处,总占比49%~56%,8月后则上移至34~69 cm处,总占比56%。而柠条细根7月主要分布在34~52 cm土层处,占比80%,7月后下移至52~86 cm处,总占比57%。可见,在水分条件较好的平水年,两种物种的细根分布均呈现一定的“中层优势”。所有土层中,34~52 cm和69~86 cm土层处花棒占比变化最大,前者从18%升至30%,后者从33%降至21%,而柠条各土层占比均相对稳定。
图3
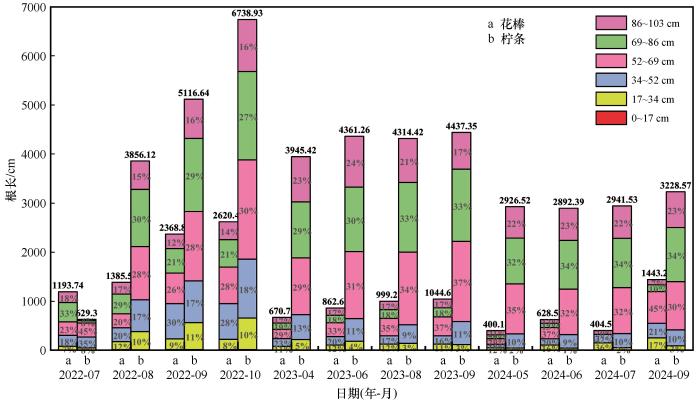
图3
两种荒漠植物不同土层处细根总根长占比动态
Fig.3
Temporal dynamics of the proportional fine‐root length in different soil layers for the two shrub species
枯水年中,花棒细根高度集中于52~69 cm土层处,单层占比在两年内从29%升至45%;同时0~34 cm层占比逐年上升,由11%增至17%;而在2023年69~103 cm深土层细根占比稳定于35%~36%,2024年降至17%,在两年中细根分布呈现出向中下层聚集同时增大浅层占比的趋势。相比之下,柠条在枯水年始终主要分布于52~86 cm处,且各层占比变化幅度小,仅52~69 cm和86~103 cm土层处占比变化较大;52~69 cm土层占比先在2023年从29%升高至37%,再于2024年降至30%;而86~103 cm土层与之相反,2023年从23%降至17%,2024年重新升至23%。总体来看,枯水年花棒更依赖浅-中层土壤水分,而柠条则表现出向更深层延伸的分布格局。
2.3 细根伸长率的时空差异
在3年观测期内,两个物种细根伸长率均在平水年显著高于枯水年(P<0.05,图4)。平水年花棒和柠条的细根在时间和垂直方向上表现出明显的分异生长模式:花棒总体呈先发展浅层后扩展中层的模式,而柠条则主要从深层向中层发展。枯水年两个物种生长重心均集中分布于中深层,但不同年份的层次优势有细微差异。总体上,两种植物在各土层均表现出一定的生长节律性,最大伸长率的时点存在种间和深度上的差异。
图4
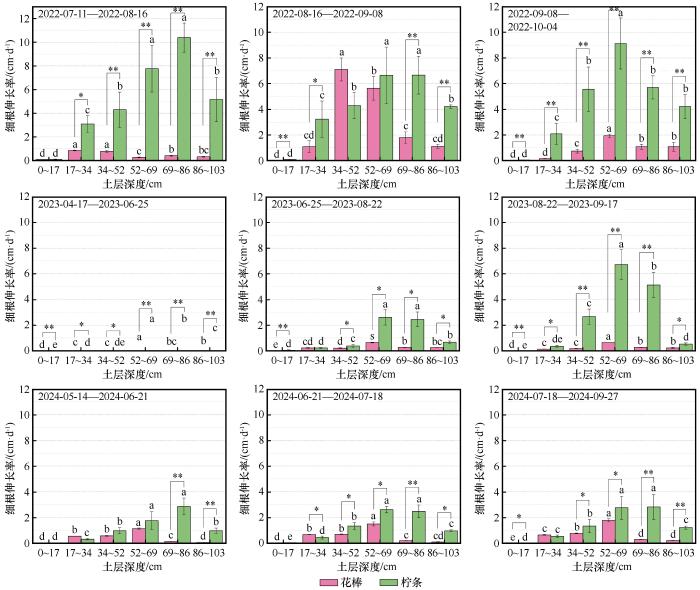
图4
不同土壤深度下两种荒漠植物细根伸长率动态
Fig.4
Fine‐root elongation rate of the two species across soil depths
2022年生长季内,花棒各土层内细根伸长率均表现出随时间先升高后降低的变化规律,且34~69 cm土层在8—9月的细根伸长率显著高于前后月份(P<0.05)。花棒细根伸长率最高值随时间从17~34 cm逐渐降至52~69 cm处,表现出从浅层向中层发展的趋势。而柠条不同土层内细根伸长率随时间具有不同的变化规律,0~34 cm土层内伸长率先升高后降低;34~52 cm土层内伸长率逐渐升高;52~69 cm土层内伸长率先降低后升高;69~103 cm土层内伸长率逐渐降低。柠条细根伸长率最高值随时间从69~86 cm逐渐升至52~69 cm处,表现出从深层向中层发展的趋势。
2023、2024年为枯水年,花棒与柠条细根伸长率均随土层先增大后减小,最高值分别在52~69 cm和56~86 cm土层处。两年中,花棒细根伸长率在52~69 cm土层处始终显著高于其余5个土层(P<0.05),2023年其余土层间伸长率无显著差异,但2024年17~52 cm土层处伸长率显著高于0~17、69~103 cm土层(P<0.05)。2023年柠条细根伸长率在34~86 cm土层处随时间逐渐增大,且8—9月伸长率显著高于前一观测时段(P<0.05);而86~103 cm土层处细根伸长率随时间逐渐降低。2024年柠条各土层的细根伸长率未显示出显著的季节性变化,伸长活动主要维持于52~86 cm。
在3年观测期内,两种植物细根具有一定的生长节律,花棒0~17、17~34 cm处细根伸长率最大值在7—8月,其余4层最大值均在8—9月。柠条34~52、52~69 cm处细根伸长率最大值在9—10月,其余4层最大值在8—9月。
2.4 细根伸长与土壤含水量的关系
浅层土壤能快速响应降雨事件,两个物种在0~17 cm土层处细根伸长率与土壤含水量呈极显著正相关(P<0.01,图5),土壤含水量分别能解释花棒和柠条细根伸长率37%和52%的变异,而在17~34 cm土层处仅柠条的细根伸长率与土壤含水量呈极显著正相关(P<0.01)且能解释41%的变异,表明柠条细根在浅层土壤中能快速响应降雨后土壤含水量的变化,而花棒仅在表层响应降雨,表层以下则对小降雨事件无响应。
图5
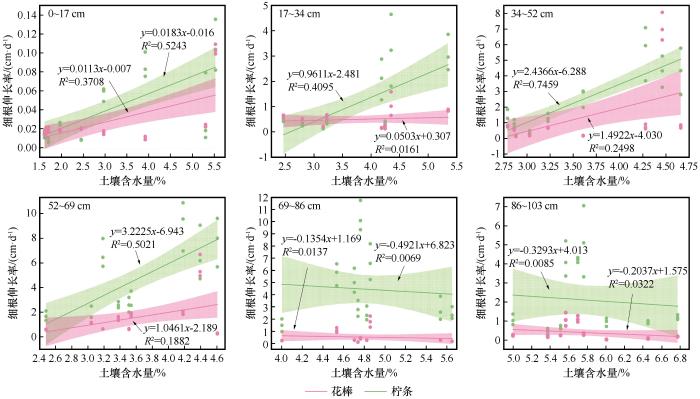
图5
不同土壤深度下两种荒漠植物细根伸长率与土壤含水量的线性关系
Fig.5
Liner relationships between fine-root elongation rates of two shrub species and soil water content at different soil depths
中层土壤含水量随降雨事件无明显变化,但因水分下渗作用,平水年土壤含水量显著高于枯水年(P<0.05)。花棒与柠条在34~52 cm土层处细根伸长率与土壤含水量均呈极显著正相关(P<0.01)土壤含水量分别能解释24%和74%的变异,在52~69 cm土层处分别为显著(P<0.05)和极显著正相关(P<0.01),分别解释19%和50%的变异。表明在中层土壤中两个物种均能响应长期较高的含水量,但花棒响应相对较弱。
由于观测期间深层土壤含水量受地下水补给影响小,季节性波动小,两个物种在深层的细根伸长率与同期深层土壤含水量之间未表现出显著的线性相关关系。
3 讨论
3.1 根系时空变化特征及生态适应
根系的垂直分布是植物在水分与养分受限环境中实现生存与竞争的核心策略,通过将细根分布于不同深度,植物既能快速截留表层降雨脉冲以支持短期生长,也能依靠中深层或地下水维持长时段的水分供给[19]。本研究表明花棒与柠条根系分别主要分布在36~69 cm与52~86 cm土层,呈现显著垂直分层模式。已有柠条根系的研究表明,20~80 cm土层是柠条细根在垂直方向上的主要分布范围,这与本研究一致[20]。平水年,中层相对较高的细根占比与该层在降雨事件后的入渗深度及较长的湿润持续期相符,长期较高的土壤含水量为细根伸长提供稳定的水源;反观浅层,尽管能被短时湿润,但沙土保水性差以及表层土壤水分的高速蒸发使细根的长期生长受限[21],导致浅层根系占比较小。枯水年,花棒浅层细根占比显著高于柠条(P<0.05),表明干旱胁迫下花棒通过在浅中层土壤中形成相对密集的细根网络以迅速捕获降雨后的入渗水分,从而在短期脉冲性降水事件中占据优势;而柠条始终依赖较深的根系获取相对稳定的深层土壤水分,柠条在深层土壤中的细根比例较大,并能在不同季节保持较高的活性。深层土壤含水量虽更稳定,但向深处持续投入大量新生细根的碳成本较高,且深层土壤的物理或气化条件可能抑制细根的高密度分布,从而使深层细根占比较小。
从生态功能角度看,深根策略通常伴随更高的构建与维护碳成本,但能够接触稳定的深层水源或地下水补给,从而在长期干旱期提供持续的水分支持;浅根策略则碳成本较低、更能迅速响应降水脉冲而获益[22]。因此,花棒向浅中层聚集的策略可被视为在脉冲资源环境中通过低成本、高周转的吸收根来快速占领资源的适应性;而柠条较深的根系分配则体现出对稳定供水的投资倾向,增强旱季生存的稳定性。两种策略在群落层面的互补性既可减少直接竞争,又能提高群落对不同水文情景的整体抗逆性。
从时间变化看,资源有效性随季节发生变化,导致细根伸长具有季节性波动[23-24]。在整个生长季,植物细根伸长率的季节变化表现为生长季初期较小、中期逐渐增大、末期又变小的趋势[25]。本研究表明,试验地花棒和柠条细根伸长率的季节变化具有相似的单峰型变化规律;除花棒浅层细根和柠条中层细根峰值出现在7—8月和9—10月外,其余峰值均在8—9月。夏季花棒浅层细根伸长出现高峰可能与土壤含水量和土壤温度有关,一方面土壤温度高使微生物和酶活性增强并加快细根呼吸作用,从而使细根伸长加快,另一方面较为集中的夏季降水使浅层土壤含水量升高,有利于细根伸长[26],使花棒细根在表层快速伸长以捕获雨水。秋季柠条中层细根伸长出现峰值可能是植物已经完成种子繁殖,需要储存足够的碳水化合物维持冬季的生存,并为第二年春季恢复生长做好营养储备[27]。其余土层细根伸长率出现在夏末初秋可能因为地上光合作用在生长期累积的光合产物经过时滞分配到根系,使得根系增长的高峰常在叶片生长高峰之后若干周到数月,这在多项长期微根窗与碳分配研究中被观测到[28]。
3.2 根系动态的年际差异对土壤水分变化的响应
平水年,在包含多次集中降雨事件的7—9月,0~60 cm土层含水量显著上升(P<0.05)。土壤湿度的快速升高不仅直接提供浅中层细根生长所需水分,同时使可溶性养分溶解增加并下渗,从而为浅中层根系出现“脉冲式”伸长提供物质基础[31-32]。而浅中层细根对水分和养分的快速吸收促进植物地上部分生长以及光合速率提高,光合产物的增加促使植物将更多碳分配到细根以支持进一步的水分和养分采集,从而使深层土壤中细根伸长加快。与此相比,枯水年中花棒与柠条各土层细根的伸长率均显著低于平水年(P<0.05),这与枯水年中浅土壤含水量显著偏低的结果一致,说明水分胁迫是限制生长季根系伸长的重要因子。在干旱背景下,细根根长密度、伸长速率与周转率均显著下降,是半干旱和干旱生态系统的普遍规律[33]。浅中层细根以对水分条件敏感的高周转吸收根为主,在降水量较少且分散的2023年,零散降水不足以形成有效水分脉冲,难以维持浅层细根快速伸长,使浅层细根占比下降;而在降水量较多且集中于生长季的2024年,生长季浅层水分条件相对改善,浅层细根生长得以增强,占比提高。
花棒的伸长率在0~17、34~69 cm土层与土壤含水量呈极显著(P<0.01)或显著正相关(P<0.05),但土壤含水量仅能解释19%~37%的变异,表明花棒浅中层细根生长受其他因素的影响较大,例如夏季适当高温可能促进细根的生长[34],多项研究表明物种的细根生产与温度之间存在显著相关性[35-36]。柠条的伸长率在0~69 cm均与土壤水分呈极显著正相关(P<0.01),在更广范围内与土壤水分动态保持紧密耦合,这与孙亚荣等[37]对柠条人工林的研究结果一致。在浅层,土壤含水量能解释柠条细根伸长率41%~52%的变异,中层能解释高达75%的变异,表明土壤含水量是柠条浅中层根系生长的主要影响因素。深层土壤中,花棒和柠条细根伸长率与土壤含水量均无显著相关性,可能由于深层土壤受地下水影响,全年土壤含水量变化幅度小,因此对细根生长无显著影响,深层土壤中土壤温度可能成为细根生长的主要影响因素。
4 结论
2022年两个物种的细根伸长率显著高于2023—2024年,表明降水总量与降水的集中度对根系生长具有强驱动作用。花棒与柠条通过根系分布和生长的动态变化响应土壤含水量的变化。花棒细根主要分布于浅中层土壤且可塑性强,能迅速利用降雨形成的短期水分脉冲并在不同降水条件下通过改变细根深度以适应水分胁迫。柠条细根延伸至中深层且各层细根占比稳定,更依赖稳定的深层水源以保障植株生长。两种根系动态模式体现了荒漠灌木在水分时空异质环境下的功能互补,有助于维持群落的水分利用效率与生态稳定性。基于两个物种根系分布差异,在荒漠植被恢复中可采用带状混交配置,以达到快速截留表层水与长期利用深层水的目标,从而促进地被覆盖与固沙,提高抗旱稳定性与生态稳健性。
本研究仍存在一定局限性。首先,由于未同步获取更精细的根系形态生理指标,如比根长、根毛密度以及根系水导率等参数,因此无法从根系构建成本与水力学性能的层面进一步解析两种灌木在水分获取策略上的差异。其次,本研究未开展稳定同位素(δ¹⁸O、δ²H)追踪实验,因而不能直接定量花棒与柠条在不同生长季节对浅层土壤水、深层土壤水或地下水等不同水源的利用比例。后续研究建议增加根系形态生理指标与同位素追踪实验,以获得更为完整且全面的植物根系适应策略。
参考文献
Plant responses to precipitation in desert ecosystems:integrating functional types,pulses,thresholds,and delays
[J].
Root phenology in a changing climate
[J].
Root distribution of three dominant desert shrubs and their water uptake dynamics
[J].
A starting guide to root ecology: strengthening ecological concepts and standardising root classification, sampling, processing and trait measurements
[J].
Temporal dynamics of fine roots under long‐term exposure to elevated CO2 in the Mojave Desert
[J].
Plant root mechanisms and their effects on carbon and nutrient accumulation in desert ecosystems under changes in land use and climate
[J].
Efficient prediction of profile mean soil water content for hillslope-scale Caragana korshinskii plantation using temporal stability analysis
[J].
Dynamic root growth in response to depth-varying soil moisture availability:a rhizobox study
[J].
Rooting depths, lateral root spreads and below-ground/above-ground allometries of plants in water-limited ecosystems
[J].
Legacies of precipitation fluctuations on primary production: theory and data synthesis
[J].
Root system architecture and its scaling relationships of Reaumuria soongorica in Alxa steppe desert,Northwest China
[J].
Image analysis procedure for the optical scanning of fine-root dynamics: errors depending on the observer and root-viewing window size
[J].
Spatial dynamics of dark septate endophytes in the roots and rhizospheres of Hedysarum scoparium in Northwest China and the influence of edaphic variables
[J].
Fine‐root dynamics vary with soil depth and precipitation in a low‐nutrient tropical forest in the Central Amazonia
[J].
Time‐varying response of fine root growth to soil temperature and soil moisture in cypress and deciduous oak forests
[J].
High frequency root dynamics:sampling and interpretation using replicated robotic minirhizotrons
[J].
Advancing fine root research with minirhizotrons
[J].
Minirhizotron measurements can supplement deep soil coring to evaluate root growth of winter wheat when certain pitfalls are avoided
[J].
Reconstructing plant root area and water uptake profiles
[J].
Predicting fine root lifespan from plant functional traits in temperate trees
[J].
Carbon allocation dynamics in conifers and broadleaved tree species revealed by pulse labeling and mass balance
[J].
Heavily intensified grazing reduces root production in an Inner Mongolia temperate steppe
[J].
Fine-root production dominates response of a deciduous forest to atmospheric CO2 enrichment
[J].
Precipitation-productivity relationships and the duration of precipitation anomalies:an underappreciated dimension of climate change
[J].
Effects of elevated CO2 on fine root dynamics in a Mojave Desert community:a FACE study
[J].
Precipitation pulses and carbon fluxes in semiarid and arid ecosystems
[J].
Water pulses and biogeochemical cycles in arid and semiarid ecosystems
[J].
The role of water stress and soil texture on plant roots anatomy,architecture, and senescence
[J].
Effects of long-term nitrogen addition and precipitation reduction on the fine root dynamics and morphology in a temperate forest
[J].
Fine‐root functional trait responses to experimental warming: a global meta‐analysis
[J].
Variations in biomass, production and respiration of fine roots in a young larch forest
[J].

 甘公网安备 62010202000688号
甘公网安备 62010202000688号
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}