Responses of soil N2O emissions and their abiotic and biotic drivers to altered rainfall regimes and co-occurring wet N deposition in a semi-arid grassland
3
2021
... 氧化亚氮(N2O)是大气中除二氧化碳(CO2)和甲烷(CH4)外的另一种重要的温室气体,其单分子增温潜势约为CO2的300倍[1].有别于其他温室气体的是,N2O还能通过在平流层中发生一系列复杂的化学反应加剧臭氧层的破坏,导致地面紫外辐射增强并威胁人类的健康与生存[2-4].目前,受人类活动和气候变化的影响,高温、干旱和极端降雨等事件导致土壤频繁地经历干旱和再湿润过程[5],频繁的干湿交替会改变土壤水分及养分的可利用性和理化性质,进而影响土壤氮循环过程和温室气体的排放[6-7].同时,化石燃料、资源过度利用等因素导致全球地表温度升高[8],而荒漠区的地表温度据预测将增加更多[9].因此,降水变化和气候变暖均会直接或间接影响荒漠土壤N2O排放过程. ...
... 荒漠生态系统植被稀疏且养分匮乏,气候条件表现为气温高、降水少且蒸发量大等.荒漠土壤中各种生物过程直接或间接地受降水驱动.荒漠区面积广袤,即使土壤环境对气候变化的微弱响应,也会引起温室气体排放量的显著变化.因此,揭示荒漠土壤N2O排放机制是荒漠生态系统研究的热点,目前的研究集中于土壤N2O排放对气候变化、氮添加和放牧的响应,且多从土壤性质和微生物角度解释土壤N2O排放变化[1].除此之外,对土壤N2O产生的硝化和反硝化作用过程开展了大量研究,同时对N2O产生途径的硝酸盐异化还原为铵和非生物过程的研究也逐渐深入.随着分子生态学和同位素示踪理论和技术的发展,从土壤微生物功能基因的微观角度揭示了N2O产生的微生物驱动机理,并量化了土壤氮循环过程对N2O排放的影响及贡献,然而,目前相关技术在荒漠生态系统中的研究偏少.此外,目前N2O排放时间尺度的研究多在几周到数月,长期研究(例如3年以上)的案例较少.由于缺乏对荒漠区土壤N2O排放过程的深入系统研究,因此难以估算其对全球土壤N2O排放的贡献,这为估算全球土壤N2O排放总量及增温潜势带来了很大的不确定性. ...
... 土壤中NH-N和NO-N是硝化细菌和反硝化细菌利用的底物,二者在土壤中的含量是影响N2O排放的关键因素[14].有研究表明,土壤中NH-N和NO-N含量增加将显著促进N2O的排放,使N2O年排放量增加55%~133%[30];同样,高有机氮含量(通过矿化作用将其转化为无机氮)也会促进N2O排放[15].但是在半干旱草原的模拟实验表明土壤氮含量增加对N2O排放的促进作用有限,施加氮肥(NH4NO3)会增加土壤N2O排放量(与不施肥相比约增加30%);土壤NO-N和NH-N含量的增加对N2O排放的促进作用弱于预期,这在很大程度上由植物-微生物对氮素的竞争所致,植物吸收了大部分的氮素[1]. ...
Nitrous oxide (N2O):the dominant ozone-depleting substance emitted in the 21st century
1
2009
... 氧化亚氮(N2O)是大气中除二氧化碳(CO2)和甲烷(CH4)外的另一种重要的温室气体,其单分子增温潜势约为CO2的300倍[1].有别于其他温室气体的是,N2O还能通过在平流层中发生一系列复杂的化学反应加剧臭氧层的破坏,导致地面紫外辐射增强并威胁人类的健康与生存[2-4].目前,受人类活动和气候变化的影响,高温、干旱和极端降雨等事件导致土壤频繁地经历干旱和再湿润过程[5],频繁的干湿交替会改变土壤水分及养分的可利用性和理化性质,进而影响土壤氮循环过程和温室气体的排放[6-7].同时,化石燃料、资源过度利用等因素导致全球地表温度升高[8],而荒漠区的地表温度据预测将增加更多[9].因此,降水变化和气候变暖均会直接或间接影响荒漠土壤N2O排放过程. ...
Legacy effects of soil moisture on microbial community structure and N2O emissions
1
2016
... 荒漠土壤N2O的排放通量和排放量在不同研究中差异较大[10-11].影响荒漠土壤N2O排放的因素复杂,如降水导致土壤水分变化进而影响土壤中细菌和真菌的种类和活性,从而影响土壤中氮素的转化以及N2O排放;气候变暖导致土壤温度的改变,通过影响微生物代谢活动和物质转换来影响N2O排放[3-4,12-13];土壤有机碳和有机氮分别通过影响微生物能量供应和氮矿化速率影响N2O排放[14-16];土壤质地和不同粒径土壤团聚体的养分供给、O2含量以及通气条件等方面的差异,也会导致N2O排放的波动[10,17].因此,亟待深入系统地开展荒漠生态系统土壤N2O排放过程及其影响机理的研究,为准确估算其对全球变化提供数据支撑. ...
Historical soil drainage mediates the response of soil greenhouse gas emissions to intense precipitation events
2
2019
... 氧化亚氮(N2O)是大气中除二氧化碳(CO2)和甲烷(CH4)外的另一种重要的温室气体,其单分子增温潜势约为CO2的300倍[1].有别于其他温室气体的是,N2O还能通过在平流层中发生一系列复杂的化学反应加剧臭氧层的破坏,导致地面紫外辐射增强并威胁人类的健康与生存[2-4].目前,受人类活动和气候变化的影响,高温、干旱和极端降雨等事件导致土壤频繁地经历干旱和再湿润过程[5],频繁的干湿交替会改变土壤水分及养分的可利用性和理化性质,进而影响土壤氮循环过程和温室气体的排放[6-7].同时,化石燃料、资源过度利用等因素导致全球地表温度升高[8],而荒漠区的地表温度据预测将增加更多[9].因此,降水变化和气候变暖均会直接或间接影响荒漠土壤N2O排放过程. ...
... 荒漠土壤N2O的排放通量和排放量在不同研究中差异较大[10-11].影响荒漠土壤N2O排放的因素复杂,如降水导致土壤水分变化进而影响土壤中细菌和真菌的种类和活性,从而影响土壤中氮素的转化以及N2O排放;气候变暖导致土壤温度的改变,通过影响微生物代谢活动和物质转换来影响N2O排放[3-4,12-13];土壤有机碳和有机氮分别通过影响微生物能量供应和氮矿化速率影响N2O排放[14-16];土壤质地和不同粒径土壤团聚体的养分供给、O2含量以及通气条件等方面的差异,也会导致N2O排放的波动[10,17].因此,亟待深入系统地开展荒漠生态系统土壤N2O排放过程及其影响机理的研究,为准确估算其对全球变化提供数据支撑. ...
Land-atmosphere coupling and climate change in Europe
1
2006
... 氧化亚氮(N2O)是大气中除二氧化碳(CO2)和甲烷(CH4)外的另一种重要的温室气体,其单分子增温潜势约为CO2的300倍[1].有别于其他温室气体的是,N2O还能通过在平流层中发生一系列复杂的化学反应加剧臭氧层的破坏,导致地面紫外辐射增强并威胁人类的健康与生存[2-4].目前,受人类活动和气候变化的影响,高温、干旱和极端降雨等事件导致土壤频繁地经历干旱和再湿润过程[5],频繁的干湿交替会改变土壤水分及养分的可利用性和理化性质,进而影响土壤氮循环过程和温室气体的排放[6-7].同时,化石燃料、资源过度利用等因素导致全球地表温度升高[8],而荒漠区的地表温度据预测将增加更多[9].因此,降水变化和气候变暖均会直接或间接影响荒漠土壤N2O排放过程. ...
Decomposition of soil and plant carbon from pasture systems after 9 years of exposure to elevated CO2:impact on C cycling and modeling
1
2004
... 氧化亚氮(N2O)是大气中除二氧化碳(CO2)和甲烷(CH4)外的另一种重要的温室气体,其单分子增温潜势约为CO2的300倍[1].有别于其他温室气体的是,N2O还能通过在平流层中发生一系列复杂的化学反应加剧臭氧层的破坏,导致地面紫外辐射增强并威胁人类的健康与生存[2-4].目前,受人类活动和气候变化的影响,高温、干旱和极端降雨等事件导致土壤频繁地经历干旱和再湿润过程[5],频繁的干湿交替会改变土壤水分及养分的可利用性和理化性质,进而影响土壤氮循环过程和温室气体的排放[6-7].同时,化石燃料、资源过度利用等因素导致全球地表温度升高[8],而荒漠区的地表温度据预测将增加更多[9].因此,降水变化和气候变暖均会直接或间接影响荒漠土壤N2O排放过程. ...
Drying-rewetting effects on N cycling in grassland soils of varying microbial community composition and management intensity in south central Chile
1
2011
... 氧化亚氮(N2O)是大气中除二氧化碳(CO2)和甲烷(CH4)外的另一种重要的温室气体,其单分子增温潜势约为CO2的300倍[1].有别于其他温室气体的是,N2O还能通过在平流层中发生一系列复杂的化学反应加剧臭氧层的破坏,导致地面紫外辐射增强并威胁人类的健康与生存[2-4].目前,受人类活动和气候变化的影响,高温、干旱和极端降雨等事件导致土壤频繁地经历干旱和再湿润过程[5],频繁的干湿交替会改变土壤水分及养分的可利用性和理化性质,进而影响土壤氮循环过程和温室气体的排放[6-7].同时,化石燃料、资源过度利用等因素导致全球地表温度升高[8],而荒漠区的地表温度据预测将增加更多[9].因此,降水变化和气候变暖均会直接或间接影响荒漠土壤N2O排放过程. ...
Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change
1
2021
... 氧化亚氮(N2O)是大气中除二氧化碳(CO2)和甲烷(CH4)外的另一种重要的温室气体,其单分子增温潜势约为CO2的300倍[1].有别于其他温室气体的是,N2O还能通过在平流层中发生一系列复杂的化学反应加剧臭氧层的破坏,导致地面紫外辐射增强并威胁人类的健康与生存[2-4].目前,受人类活动和气候变化的影响,高温、干旱和极端降雨等事件导致土壤频繁地经历干旱和再湿润过程[5],频繁的干湿交替会改变土壤水分及养分的可利用性和理化性质,进而影响土壤氮循环过程和温室气体的排放[6-7].同时,化石燃料、资源过度利用等因素导致全球地表温度升高[8],而荒漠区的地表温度据预测将增加更多[9].因此,降水变化和气候变暖均会直接或间接影响荒漠土壤N2O排放过程. ...
Global warming and changes in risk of concurrent climate extremes:Insights from the 2014 California drought
1
2014
... 氧化亚氮(N2O)是大气中除二氧化碳(CO2)和甲烷(CH4)外的另一种重要的温室气体,其单分子增温潜势约为CO2的300倍[1].有别于其他温室气体的是,N2O还能通过在平流层中发生一系列复杂的化学反应加剧臭氧层的破坏,导致地面紫外辐射增强并威胁人类的健康与生存[2-4].目前,受人类活动和气候变化的影响,高温、干旱和极端降雨等事件导致土壤频繁地经历干旱和再湿润过程[5],频繁的干湿交替会改变土壤水分及养分的可利用性和理化性质,进而影响土壤氮循环过程和温室气体的排放[6-7].同时,化石燃料、资源过度利用等因素导致全球地表温度升高[8],而荒漠区的地表温度据预测将增加更多[9].因此,降水变化和气候变暖均会直接或间接影响荒漠土壤N2O排放过程. ...
模拟增温对荒漠生物土壤结皮-土壤系统CO2、CH4和N2O通量的影响
11
2014
... 荒漠土壤N2O的排放通量和排放量在不同研究中差异较大[10-11].影响荒漠土壤N2O排放的因素复杂,如降水导致土壤水分变化进而影响土壤中细菌和真菌的种类和活性,从而影响土壤中氮素的转化以及N2O排放;气候变暖导致土壤温度的改变,通过影响微生物代谢活动和物质转换来影响N2O排放[3-4,12-13];土壤有机碳和有机氮分别通过影响微生物能量供应和氮矿化速率影响N2O排放[14-16];土壤质地和不同粒径土壤团聚体的养分供给、O2含量以及通气条件等方面的差异,也会导致N2O排放的波动[10,17].因此,亟待深入系统地开展荒漠生态系统土壤N2O排放过程及其影响机理的研究,为准确估算其对全球变化提供数据支撑. ...
... [10,17].因此,亟待深入系统地开展荒漠生态系统土壤N2O排放过程及其影响机理的研究,为准确估算其对全球变化提供数据支撑. ...
... N2O排放量在不同研究中具有明显的差异性,如在古尔班通古特沙漠进行1年和连续3年的野外观测研究表明,土壤N2O年排放量为0.13 kg·hm-2[30,32].然而,在腾格里沙漠东南缘的研究表明,生物土壤结皮(Biological soil crust,BSC)的N2O年排放量为负值(平均为-1.44 kg·hm-2),表现为N2O年吸收[10],这可能主要与结皮性质有关,相对致密且渗水性低的生物结皮不利于N2O向大气扩散,土壤微生物促使N2O向N2转化.不同研究中,土壤N2O排放速率及排放量见表1[10-11,33-43]. ...
... [10-11,33-43]. ...
... N
2O emission fluxes and emissions in different studies
Table 1| 生境类型 | 植被类型 | 土壤类型 | 观测 时间 | 年排放量 /(kg·hm-2) | 生长季排放量 占全年比重/% | 排放通量 /(μg·m-2·h-1) | 参考 文献 |
|---|
| 荒漠 | 藻类 | 结皮土 | 全年 | -0.19 | — | -2.12(均值) | [10] |
| 藻类 | 结皮土 | 季节性 | — | — | -18.5~9.8 | [33] |
| 苔藓 | 结皮土 | 全年 | -3.87 | — | -4.42(均值) | [10] |
| 苔藓 | 结皮土 | 季节性 | — | — | -15.5~4.5 | [33] |
| 混生 | 结皮土 | 全年 | -0.27 | — | -3.12(均值) | [10] |
| 草本 | 砂质土 | 全年 | 0.13 | — | 0~3.46 | [32] |
| 牧草 | 砂质土 | 全年 | 0.13 | 43 | 1.49(均值) | [30] |
| 稀疏灌木,草本 | 砂质土 | 夏季 | — | — | 0~8 | [11] |
| 干旱草原 | 草本 | 砂质土 | 全年 | 0.22 | 53 | -0.7~10.3 | [34] |
| 草本 | 砂质土 | 生长季 | — | — | 8.58(均值) | [35] |
| 半干旱草地 | 草本 | 砂质土 | 生长季 | — | — | -0.14~46.62 | [36] |
| 草本 | 细壤土 | 生长季 | — | — | 0~400 | [37] |
| 半干旱草原 | 草本 | 盐碱土 | 生长季 | 13.1 | 90 | 18.7(均值) | [38] |
| 牧草 | 砂质土 | 生长季 | — | — | 19.8(均值) | [29] |
| 草本 | 砂质土 | 生长季 | — | — | -0.34~0.12 | [39] |
| 草本 | 砂质土 | 生长季 | — | — | 2.8~3.9 | [40] |
| 草本 | 砂质土 | 生长季 | — | — | 3.4~12 | [41] |
| 苔藓 | 结皮土 | 生长季 | — | — | -0.42~0.84 | [42] |
| 草本 | 砂质土 | 生长季 | — | — | 4(均值) | [43] |
“—”代表无相关数据. ...
... [
10]
| 苔藓 | 结皮土 | 季节性 | — | — | -15.5~4.5 | [33] |
| 混生 | 结皮土 | 全年 | -0.27 | — | -3.12(均值) | [10] |
| 草本 | 砂质土 | 全年 | 0.13 | — | 0~3.46 | [32] |
| 牧草 | 砂质土 | 全年 | 0.13 | 43 | 1.49(均值) | [30] |
| 稀疏灌木,草本 | 砂质土 | 夏季 | — | — | 0~8 | [11] |
| 干旱草原 | 草本 | 砂质土 | 全年 | 0.22 | 53 | -0.7~10.3 | [34] |
| 草本 | 砂质土 | 生长季 | — | — | 8.58(均值) | [35] |
| 半干旱草地 | 草本 | 砂质土 | 生长季 | — | — | -0.14~46.62 | [36] |
| 草本 | 细壤土 | 生长季 | — | — | 0~400 | [37] |
| 半干旱草原 | 草本 | 盐碱土 | 生长季 | 13.1 | 90 | 18.7(均值) | [38] |
| 牧草 | 砂质土 | 生长季 | — | — | 19.8(均值) | [29] |
| 草本 | 砂质土 | 生长季 | — | — | -0.34~0.12 | [39] |
| 草本 | 砂质土 | 生长季 | — | — | 2.8~3.9 | [40] |
| 草本 | 砂质土 | 生长季 | — | — | 3.4~12 | [41] |
| 苔藓 | 结皮土 | 生长季 | — | — | -0.42~0.84 | [42] |
| 草本 | 砂质土 | 生长季 | — | — | 4(均值) | [43] |
“—”代表无相关数据. ...
... [
10]
| 草本 | 砂质土 | 全年 | 0.13 | — | 0~3.46 | [32] |
| 牧草 | 砂质土 | 全年 | 0.13 | 43 | 1.49(均值) | [30] |
| 稀疏灌木,草本 | 砂质土 | 夏季 | — | — | 0~8 | [11] |
| 干旱草原 | 草本 | 砂质土 | 全年 | 0.22 | 53 | -0.7~10.3 | [34] |
| 草本 | 砂质土 | 生长季 | — | — | 8.58(均值) | [35] |
| 半干旱草地 | 草本 | 砂质土 | 生长季 | — | — | -0.14~46.62 | [36] |
| 草本 | 细壤土 | 生长季 | — | — | 0~400 | [37] |
| 半干旱草原 | 草本 | 盐碱土 | 生长季 | 13.1 | 90 | 18.7(均值) | [38] |
| 牧草 | 砂质土 | 生长季 | — | — | 19.8(均值) | [29] |
| 草本 | 砂质土 | 生长季 | — | — | -0.34~0.12 | [39] |
| 草本 | 砂质土 | 生长季 | — | — | 2.8~3.9 | [40] |
| 草本 | 砂质土 | 生长季 | — | — | 3.4~12 | [41] |
| 苔藓 | 结皮土 | 生长季 | — | — | -0.42~0.84 | [42] |
| 草本 | 砂质土 | 生长季 | — | — | 4(均值) | [43] |
“—”代表无相关数据. ...
... 荒漠土壤N2O排放具有明显的季节动态.在中国北方荒漠草原为期1年的观测结果表明,N2O排放通量整体表现为春夏季节较高(最大值为14.79 μg· m-2·h-1),冬季排放量较低甚至为负(最低值为-0.93 μg·m-2·h-1)[30],这主要是由于生长季土壤在多次降水脉冲后反复经历干湿循环过程,干旱阶段土壤微生物主要以硝化作用产生N2O;短暂湿润阶段土壤微生物主要以反硝化作用产生N2O,同时适宜的温度也有助于增加微生物活性,因此生长季荒漠土壤产生较多的N2O(占全年的43%~90%)[44];非生长季排放的N2O较少,主要与土壤含水量低、温度较低、微生物活性较低有关,同时冬季表层土壤冰冻或者被积雪覆盖导致N2O无法向大气扩散,故N2O排放减少[10]. ...
... 土壤中不同粒径组成造成土壤通透性差异较大,这对硝化作用和反硝化作用以及N2O扩散速率均有重要影响[55],同时不同粒径土壤团聚体中养分差异对N2O产生也有重要影响[56].荒漠区土壤砂粒含量普遍较高,但在不同生境斑块(如隐花植物、草本和灌木)中砂粒含量有所不同,其中流沙中砂粒含量最高、黏粒含量最低[10,29,57].相关研究表明,黏粒含量高的土壤N2O排放量较高,这主要与黏粒含量高的土壤保水能力强、孔隙度低以及O2含量低,有利于反硝化作用有关[55].此外,不同粒径土壤团聚体对N2O的生成也有不同影响[56],并且N2O排放量的研究结果存在差异[58-59],这主要与不同粒径的土壤团聚体在通透性、营养物质含量和微生物分布等方面不同有关[60]. ...
... 有关不同类型生物土壤结皮的研究表明,藓类结皮、藻类结皮和混生结皮均表现为N2O负通量,即N2O净吸收,吸收量的大小为藓类结皮>混生结皮>藻类结皮[10].也有研究发现,藻类结皮和藓类结皮生长季(4—10月)的吸收通量显著高于非生长季[33].藓类结皮更易产生相对厌氧环境,促进了反硝化作用产生N2O,同时也阻止了N2O通过扩散从土壤进入大气,将N2O转化为N2,因此整体表现为N2O净吸收[10].养分更为贫瘠的风沙土却表现为N2O净排放,即使在土壤最干燥的时期N2O也会排放到大气中[17]. ...
... [10].养分更为贫瘠的风沙土却表现为N2O净排放,即使在土壤最干燥的时期N2O也会排放到大气中[17]. ...
The driving effect of nitrogen-related functional microorganisms under water and nitrogen addition on N2O emission in a temperate desert
3
2021
... 荒漠土壤N2O的排放通量和排放量在不同研究中差异较大[10-11].影响荒漠土壤N2O排放的因素复杂,如降水导致土壤水分变化进而影响土壤中细菌和真菌的种类和活性,从而影响土壤中氮素的转化以及N2O排放;气候变暖导致土壤温度的改变,通过影响微生物代谢活动和物质转换来影响N2O排放[3-4,12-13];土壤有机碳和有机氮分别通过影响微生物能量供应和氮矿化速率影响N2O排放[14-16];土壤质地和不同粒径土壤团聚体的养分供给、O2含量以及通气条件等方面的差异,也会导致N2O排放的波动[10,17].因此,亟待深入系统地开展荒漠生态系统土壤N2O排放过程及其影响机理的研究,为准确估算其对全球变化提供数据支撑. ...
... N2O排放量在不同研究中具有明显的差异性,如在古尔班通古特沙漠进行1年和连续3年的野外观测研究表明,土壤N2O年排放量为0.13 kg·hm-2[30,32].然而,在腾格里沙漠东南缘的研究表明,生物土壤结皮(Biological soil crust,BSC)的N2O年排放量为负值(平均为-1.44 kg·hm-2),表现为N2O年吸收[10],这可能主要与结皮性质有关,相对致密且渗水性低的生物结皮不利于N2O向大气扩散,土壤微生物促使N2O向N2转化.不同研究中,土壤N2O排放速率及排放量见表1[10-11,33-43]. ...
... N
2O emission fluxes and emissions in different studies
Table 1| 生境类型 | 植被类型 | 土壤类型 | 观测 时间 | 年排放量 /(kg·hm-2) | 生长季排放量 占全年比重/% | 排放通量 /(μg·m-2·h-1) | 参考 文献 |
|---|
| 荒漠 | 藻类 | 结皮土 | 全年 | -0.19 | — | -2.12(均值) | [10] |
| 藻类 | 结皮土 | 季节性 | — | — | -18.5~9.8 | [33] |
| 苔藓 | 结皮土 | 全年 | -3.87 | — | -4.42(均值) | [10] |
| 苔藓 | 结皮土 | 季节性 | — | — | -15.5~4.5 | [33] |
| 混生 | 结皮土 | 全年 | -0.27 | — | -3.12(均值) | [10] |
| 草本 | 砂质土 | 全年 | 0.13 | — | 0~3.46 | [32] |
| 牧草 | 砂质土 | 全年 | 0.13 | 43 | 1.49(均值) | [30] |
| 稀疏灌木,草本 | 砂质土 | 夏季 | — | — | 0~8 | [11] |
| 干旱草原 | 草本 | 砂质土 | 全年 | 0.22 | 53 | -0.7~10.3 | [34] |
| 草本 | 砂质土 | 生长季 | — | — | 8.58(均值) | [35] |
| 半干旱草地 | 草本 | 砂质土 | 生长季 | — | — | -0.14~46.62 | [36] |
| 草本 | 细壤土 | 生长季 | — | — | 0~400 | [37] |
| 半干旱草原 | 草本 | 盐碱土 | 生长季 | 13.1 | 90 | 18.7(均值) | [38] |
| 牧草 | 砂质土 | 生长季 | — | — | 19.8(均值) | [29] |
| 草本 | 砂质土 | 生长季 | — | — | -0.34~0.12 | [39] |
| 草本 | 砂质土 | 生长季 | — | — | 2.8~3.9 | [40] |
| 草本 | 砂质土 | 生长季 | — | — | 3.4~12 | [41] |
| 苔藓 | 结皮土 | 生长季 | — | — | -0.42~0.84 | [42] |
| 草本 | 砂质土 | 生长季 | — | — | 4(均值) | [43] |
“—”代表无相关数据. ...
Natural perturbations,drying-wetting and freezing-thawing cycles,and the emission of nitrous oxide,carbon dioxide and methane from farmed organic soils
1
2001
... 荒漠土壤N2O的排放通量和排放量在不同研究中差异较大[10-11].影响荒漠土壤N2O排放的因素复杂,如降水导致土壤水分变化进而影响土壤中细菌和真菌的种类和活性,从而影响土壤中氮素的转化以及N2O排放;气候变暖导致土壤温度的改变,通过影响微生物代谢活动和物质转换来影响N2O排放[3-4,12-13];土壤有机碳和有机氮分别通过影响微生物能量供应和氮矿化速率影响N2O排放[14-16];土壤质地和不同粒径土壤团聚体的养分供给、O2含量以及通气条件等方面的差异,也会导致N2O排放的波动[10,17].因此,亟待深入系统地开展荒漠生态系统土壤N2O排放过程及其影响机理的研究,为准确估算其对全球变化提供数据支撑. ...
Increased soil release of greenhouse gases shrinks terrestrial carbon uptake enhancement under warming
1
2020
... 荒漠土壤N2O的排放通量和排放量在不同研究中差异较大[10-11].影响荒漠土壤N2O排放的因素复杂,如降水导致土壤水分变化进而影响土壤中细菌和真菌的种类和活性,从而影响土壤中氮素的转化以及N2O排放;气候变暖导致土壤温度的改变,通过影响微生物代谢活动和物质转换来影响N2O排放[3-4,12-13];土壤有机碳和有机氮分别通过影响微生物能量供应和氮矿化速率影响N2O排放[14-16];土壤质地和不同粒径土壤团聚体的养分供给、O2含量以及通气条件等方面的差异,也会导致N2O排放的波动[10,17].因此,亟待深入系统地开展荒漠生态系统土壤N2O排放过程及其影响机理的研究,为准确估算其对全球变化提供数据支撑. ...
Biochar mitigates the N2O emissions from acidic soil by increasing the nosZ and nirK gene abundance and soil pH
3
2020
... 荒漠土壤N2O的排放通量和排放量在不同研究中差异较大[10-11].影响荒漠土壤N2O排放的因素复杂,如降水导致土壤水分变化进而影响土壤中细菌和真菌的种类和活性,从而影响土壤中氮素的转化以及N2O排放;气候变暖导致土壤温度的改变,通过影响微生物代谢活动和物质转换来影响N2O排放[3-4,12-13];土壤有机碳和有机氮分别通过影响微生物能量供应和氮矿化速率影响N2O排放[14-16];土壤质地和不同粒径土壤团聚体的养分供给、O2含量以及通气条件等方面的差异,也会导致N2O排放的波动[10,17].因此,亟待深入系统地开展荒漠生态系统土壤N2O排放过程及其影响机理的研究,为准确估算其对全球变化提供数据支撑. ...
... 土壤有机碳是影响异养硝化细菌和反硝化细菌活性的重要因素[14].荒漠土壤有机碳的主要来源为植物凋落物分解、根系分泌和微生物分泌、死亡残体释放[45].自然降水后,雨水促进植物和生物结皮生长,有利于植物根系、生物结皮对土壤有机碳的输入;同时,土壤水势增加使得微生物承受较高的渗透压力,其中一部分微生物死亡释放出有机物质,另一部分通过释放大量易分解含碳化合物(包括小分子含碳化合物和可溶性碳)来适应迅速增加的土壤水势.有机碳可为微生物提供重要碳源[16],有机碳供给增加一方面可加快微生物繁殖,消耗氧气从而形成厌氧环境促进反硝化作用发生,另一方面有机碳作为能量来源,促进微生物新陈代谢和物质循环,同时也为反硝化细菌提供电子从而促进反硝化作用[46]. ...
... 土壤中NH-N和NO-N是硝化细菌和反硝化细菌利用的底物,二者在土壤中的含量是影响N2O排放的关键因素[14].有研究表明,土壤中NH-N和NO-N含量增加将显著促进N2O的排放,使N2O年排放量增加55%~133%[30];同样,高有机氮含量(通过矿化作用将其转化为无机氮)也会促进N2O排放[15].但是在半干旱草原的模拟实验表明土壤氮含量增加对N2O排放的促进作用有限,施加氮肥(NH4NO3)会增加土壤N2O排放量(与不施肥相比约增加30%);土壤NO-N和NH-N含量的增加对N2O排放的促进作用弱于预期,这在很大程度上由植物-微生物对氮素的竞争所致,植物吸收了大部分的氮素[1]. ...
Contrasting effects of different pH-raising materials on N2O emissions in acidic upland soils
2
2021
... 土壤中NH-N和NO-N是硝化细菌和反硝化细菌利用的底物,二者在土壤中的含量是影响N2O排放的关键因素[14].有研究表明,土壤中NH-N和NO-N含量增加将显著促进N2O的排放,使N2O年排放量增加55%~133%[30];同样,高有机氮含量(通过矿化作用将其转化为无机氮)也会促进N2O排放[15].但是在半干旱草原的模拟实验表明土壤氮含量增加对N2O排放的促进作用有限,施加氮肥(NH4NO3)会增加土壤N2O排放量(与不施肥相比约增加30%);土壤NO-N和NH-N含量的增加对N2O排放的促进作用弱于预期,这在很大程度上由植物-微生物对氮素的竞争所致,植物吸收了大部分的氮素[1]. ...
... pH影响参与氮转化的相关过程,如反硝化、自养和异养硝化,这些生物过程都与N2O的产生有关[48].有研究指出,增加土壤pH促进N2O向N2的转化,整体表现为低N2O排放[15],主要原因在于pH影响细菌和真菌活性,大多数最适宜反硝化微生物生长的pH为6.0~8.0的中性环境,在pH较高或较低时反硝化作用速率较低[49-50].同时,pH也会通过调控AOA和AOB的群落结构影响N2O通量,例如AOB在中性、碱性土壤中占主导,而AOA在酸性土壤中占主导[51]. ...
Dry-wet cycles affect nitrous oxide emissions across aquatic-terrestrial interfaces:a mesocosms study
2
2022
... 荒漠土壤N2O的排放通量和排放量在不同研究中差异较大[10-11].影响荒漠土壤N2O排放的因素复杂,如降水导致土壤水分变化进而影响土壤中细菌和真菌的种类和活性,从而影响土壤中氮素的转化以及N2O排放;气候变暖导致土壤温度的改变,通过影响微生物代谢活动和物质转换来影响N2O排放[3-4,12-13];土壤有机碳和有机氮分别通过影响微生物能量供应和氮矿化速率影响N2O排放[14-16];土壤质地和不同粒径土壤团聚体的养分供给、O2含量以及通气条件等方面的差异,也会导致N2O排放的波动[10,17].因此,亟待深入系统地开展荒漠生态系统土壤N2O排放过程及其影响机理的研究,为准确估算其对全球变化提供数据支撑. ...
... 土壤有机碳是影响异养硝化细菌和反硝化细菌活性的重要因素[14].荒漠土壤有机碳的主要来源为植物凋落物分解、根系分泌和微生物分泌、死亡残体释放[45].自然降水后,雨水促进植物和生物结皮生长,有利于植物根系、生物结皮对土壤有机碳的输入;同时,土壤水势增加使得微生物承受较高的渗透压力,其中一部分微生物死亡释放出有机物质,另一部分通过释放大量易分解含碳化合物(包括小分子含碳化合物和可溶性碳)来适应迅速增加的土壤水势.有机碳可为微生物提供重要碳源[16],有机碳供给增加一方面可加快微生物繁殖,消耗氧气从而形成厌氧环境促进反硝化作用发生,另一方面有机碳作为能量来源,促进微生物新陈代谢和物质循环,同时也为反硝化细菌提供电子从而促进反硝化作用[46]. ...
Trace N gas losses and N mineralization in Mojave desert soils exposed to elevated CO2
2
2002
... 荒漠土壤N2O的排放通量和排放量在不同研究中差异较大[10-11].影响荒漠土壤N2O排放的因素复杂,如降水导致土壤水分变化进而影响土壤中细菌和真菌的种类和活性,从而影响土壤中氮素的转化以及N2O排放;气候变暖导致土壤温度的改变,通过影响微生物代谢活动和物质转换来影响N2O排放[3-4,12-13];土壤有机碳和有机氮分别通过影响微生物能量供应和氮矿化速率影响N2O排放[14-16];土壤质地和不同粒径土壤团聚体的养分供给、O2含量以及通气条件等方面的差异,也会导致N2O排放的波动[10,17].因此,亟待深入系统地开展荒漠生态系统土壤N2O排放过程及其影响机理的研究,为准确估算其对全球变化提供数据支撑. ...
... 有关不同类型生物土壤结皮的研究表明,藓类结皮、藻类结皮和混生结皮均表现为N2O负通量,即N2O净吸收,吸收量的大小为藓类结皮>混生结皮>藻类结皮[10].也有研究发现,藻类结皮和藓类结皮生长季(4—10月)的吸收通量显著高于非生长季[33].藓类结皮更易产生相对厌氧环境,促进了反硝化作用产生N2O,同时也阻止了N2O通过扩散从土壤进入大气,将N2O转化为N2,因此整体表现为N2O净吸收[10].养分更为贫瘠的风沙土却表现为N2O净排放,即使在土壤最干燥的时期N2O也会排放到大气中[17]. ...
A comprehensive quantification of global nitrous oxide sources and sinks
1
2020
... N2O的排放来源有土壤、海洋和淡水湖泊,以及化石燃料燃烧等,其中土壤是N2O的主要排放源,约占总排放量的60%[18].土壤排放N2O的主要原因是其中含有大量微生物,微生物可通过一系列复杂的生物化学过程产生N2O[19],包括硝化作用(Nitrification)、反硝化作用(Denitrification)、硝酸盐异化还原成铵(Dissimilatory nitrate reduction to ammonium,DNRA)、硝化细菌反硝化作用(Nitrifier denitrification).另外,土壤排放的N2O也可能来自于非生物分解等过程[20-22]. ...
Impacts of nitrogen application rates on the activity and diversity of denitrifying bacteria in the broadbalk wheat experiment
1
2012
... N2O的排放来源有土壤、海洋和淡水湖泊,以及化石燃料燃烧等,其中土壤是N2O的主要排放源,约占总排放量的60%[18].土壤排放N2O的主要原因是其中含有大量微生物,微生物可通过一系列复杂的生物化学过程产生N2O[19],包括硝化作用(Nitrification)、反硝化作用(Denitrification)、硝酸盐异化还原成铵(Dissimilatory nitrate reduction to ammonium,DNRA)、硝化细菌反硝化作用(Nitrifier denitrification).另外,土壤排放的N2O也可能来自于非生物分解等过程[20-22]. ...
Nitrous oxide emissions from soils:how well do we understand the processes and their controls?
3
2013
... N2O的排放来源有土壤、海洋和淡水湖泊,以及化石燃料燃烧等,其中土壤是N2O的主要排放源,约占总排放量的60%[18].土壤排放N2O的主要原因是其中含有大量微生物,微生物可通过一系列复杂的生物化学过程产生N2O[19],包括硝化作用(Nitrification)、反硝化作用(Denitrification)、硝酸盐异化还原成铵(Dissimilatory nitrate reduction to ammonium,DNRA)、硝化细菌反硝化作用(Nitrifier denitrification).另外,土壤排放的N2O也可能来自于非生物分解等过程[20-22]. ...
... 温度对N2O排放的影响有着广泛的微生物基础[20,74].研究表明,自养硝化和异养反硝化作用都是土壤N2O排放的主要原因,二者都是温度敏感型过程[75-80],由不同的微生物类群主导[20,81].干旱草地土壤中AOB或AOA的丰度随温度升高而发生变化[82],这主要由细菌和真菌对温度的敏感性不同所导致.温度是控制与N2O排放相关的微生物功能群体(主要是自养氨氧化菌和异养反硝化菌)的群落特征及其活动的重要因素[83].研究表明,温度可以直接或间接地影响土壤N2O排放[76].温度变化不仅影响微生物的相对丰度和总生物量,也影响其生理过程,即使在微生物群落结构和组成没有发生变化的情况下,与产生和消耗N2O相关基因的表达也可能发生改变;同时温度变化也会对反硝化酶动力学有直接的影响[84].研究发现,随温度升高,N2O排放呈现不同的变化趋势[77-78,85].但大多数研究均表明,N2O排放与土壤温度呈正相关,适宜的温度使得微生物具有较高的新陈代谢速率,加速物质循环. ...
... [20,81].干旱草地土壤中AOB或AOA的丰度随温度升高而发生变化[82],这主要由细菌和真菌对温度的敏感性不同所导致.温度是控制与N2O排放相关的微生物功能群体(主要是自养氨氧化菌和异养反硝化菌)的群落特征及其活动的重要因素[83].研究表明,温度可以直接或间接地影响土壤N2O排放[76].温度变化不仅影响微生物的相对丰度和总生物量,也影响其生理过程,即使在微生物群落结构和组成没有发生变化的情况下,与产生和消耗N2O相关基因的表达也可能发生改变;同时温度变化也会对反硝化酶动力学有直接的影响[84].研究发现,随温度升高,N2O排放呈现不同的变化趋势[77-78,85].但大多数研究均表明,N2O排放与土壤温度呈正相关,适宜的温度使得微生物具有较高的新陈代谢速率,加速物质循环. ...
The microbial nitrogen-cycling network
0
2018
The long-term relationship between microbial metabolism and greenhouse gases
1
2020
... N2O的排放来源有土壤、海洋和淡水湖泊,以及化石燃料燃烧等,其中土壤是N2O的主要排放源,约占总排放量的60%[18].土壤排放N2O的主要原因是其中含有大量微生物,微生物可通过一系列复杂的生物化学过程产生N2O[19],包括硝化作用(Nitrification)、反硝化作用(Denitrification)、硝酸盐异化还原成铵(Dissimilatory nitrate reduction to ammonium,DNRA)、硝化细菌反硝化作用(Nitrifier denitrification).另外,土壤排放的N2O也可能来自于非生物分解等过程[20-22]. ...
Microbial functional genes involved in nitrogen fixation,nitrification and denitrification in forest ecosystems
1
2014
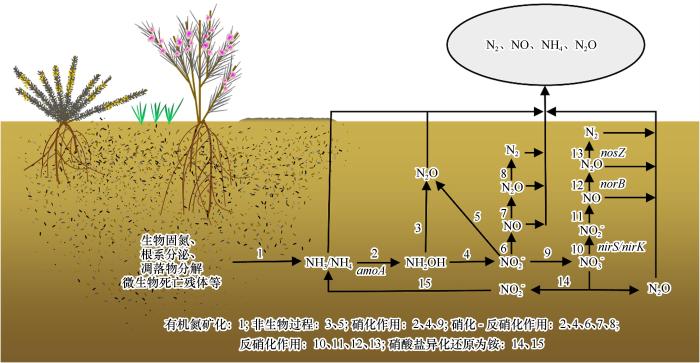
... 土壤N2O的产生过程主要有:①硝化作用,即在氨氧化细菌(AOB)和氨氧化古细菌(AOA)催化下将氨氧化成NO或NO的过程,此过程中生成副产物N2O[23];②硝化-反硝化作用,即将铵氧化成NO,然后直接转化为NO,逐步还原为N2的过程[24];③反硝化作用,即由异养细菌和古菌在厌氧环境下将硝酸盐和亚硝酸盐逐步还原成气态氮化合物的过程,最终产物为NO、N2O或N2[25];④硝酸盐异化还原为铵,即NO先被还原为NO,再进一步还原为铵的过程,此过程会产生N2O[25];⑤非生物过程,即由于NO和NH2OH不稳定的化学性质,通过分解或化学反应生成N2O[24,26].上述N2O产生途径中,硝化和反硝化作用产生的N2O约占土壤释放总量的70%[27]. ...
A review of chemical reactions of nitrification intermediates and their role in nitrogen cycling and nitrogen trace gas formation in soil
2
2016
... 土壤N2O的产生过程主要有:①硝化作用,即在氨氧化细菌(AOB)和氨氧化古细菌(AOA)催化下将氨氧化成NO或NO的过程,此过程中生成副产物N2O[23];②硝化-反硝化作用,即将铵氧化成NO,然后直接转化为NO,逐步还原为N2的过程[24];③反硝化作用,即由异养细菌和古菌在厌氧环境下将硝酸盐和亚硝酸盐逐步还原成气态氮化合物的过程,最终产物为NO、N2O或N2[25];④硝酸盐异化还原为铵,即NO先被还原为NO,再进一步还原为铵的过程,此过程会产生N2O[25];⑤非生物过程,即由于NO和NH2OH不稳定的化学性质,通过分解或化学反应生成N2O[24,26].上述N2O产生途径中,硝化和反硝化作用产生的N2O约占土壤释放总量的70%[27]. ...
... [24,26].上述N2O产生途径中,硝化和反硝化作用产生的N2O约占土壤释放总量的70%[27]. ...
Ecology of denitrification and dissimilatory nitrate reduction to ammonium
2
1988
... 土壤N2O的产生过程主要有:①硝化作用,即在氨氧化细菌(AOB)和氨氧化古细菌(AOA)催化下将氨氧化成NO或NO的过程,此过程中生成副产物N2O[23];②硝化-反硝化作用,即将铵氧化成NO,然后直接转化为NO,逐步还原为N2的过程[24];③反硝化作用,即由异养细菌和古菌在厌氧环境下将硝酸盐和亚硝酸盐逐步还原成气态氮化合物的过程,最终产物为NO、N2O或N2[25];④硝酸盐异化还原为铵,即NO先被还原为NO,再进一步还原为铵的过程,此过程会产生N2O[25];⑤非生物过程,即由于NO和NH2OH不稳定的化学性质,通过分解或化学反应生成N2O[24,26].上述N2O产生途径中,硝化和反硝化作用产生的N2O约占土壤释放总量的70%[27]. ...
... [25];⑤非生物过程,即由于NO和NH2OH不稳定的化学性质,通过分解或化学反应生成N2O[24,26].上述N2O产生途径中,硝化和反硝化作用产生的N2O约占土壤释放总量的70%[27]. ...
The importance of abiotic reactions for nitrous oxide production
1
2015
... 土壤N2O的产生过程主要有:①硝化作用,即在氨氧化细菌(AOB)和氨氧化古细菌(AOA)催化下将氨氧化成NO或NO的过程,此过程中生成副产物N2O[23];②硝化-反硝化作用,即将铵氧化成NO,然后直接转化为NO,逐步还原为N2的过程[24];③反硝化作用,即由异养细菌和古菌在厌氧环境下将硝酸盐和亚硝酸盐逐步还原成气态氮化合物的过程,最终产物为NO、N2O或N2[25];④硝酸盐异化还原为铵,即NO先被还原为NO,再进一步还原为铵的过程,此过程会产生N2O[25];⑤非生物过程,即由于NO和NH2OH不稳定的化学性质,通过分解或化学反应生成N2O[24,26].上述N2O产生途径中,硝化和反硝化作用产生的N2O约占土壤释放总量的70%[27]. ...
Contribution of fungi to soil nitrous oxide emission and their research methods:a review
1
2014
... 土壤N2O的产生过程主要有:①硝化作用,即在氨氧化细菌(AOB)和氨氧化古细菌(AOA)催化下将氨氧化成NO或NO的过程,此过程中生成副产物N2O[23];②硝化-反硝化作用,即将铵氧化成NO,然后直接转化为NO,逐步还原为N2的过程[24];③反硝化作用,即由异养细菌和古菌在厌氧环境下将硝酸盐和亚硝酸盐逐步还原成气态氮化合物的过程,最终产物为NO、N2O或N2[25];④硝酸盐异化还原为铵,即NO先被还原为NO,再进一步还原为铵的过程,此过程会产生N2O[25];⑤非生物过程,即由于NO和NH2OH不稳定的化学性质,通过分解或化学反应生成N2O[24,26].上述N2O产生途径中,硝化和反硝化作用产生的N2O约占土壤释放总量的70%[27]. ...
Steps towards a mechanistic model of global soil nitric oxide emissions:implementation and space based-constraints
1
2012
... 在全球范围内,荒漠区已经被确定为含氮气体重要的“源”[28],究其原因,主要是发生自然降水后微生物代谢活性增强,促进了氮的快速周转[29].尽管荒漠区土壤中含有的营养物质(C、N)有限,但这种氮的快速周转依然使其成为重要的N2O排放“源”. ...
Soil and vegetation-atmosphere exchange of NO,NH3,and N2O from field measurements in a semi arid grazed ecosystem in Senegal
3
2017
... 在全球范围内,荒漠区已经被确定为含氮气体重要的“源”[28],究其原因,主要是发生自然降水后微生物代谢活性增强,促进了氮的快速周转[29].尽管荒漠区土壤中含有的营养物质(C、N)有限,但这种氮的快速周转依然使其成为重要的N2O排放“源”. ...
... N
2O emission fluxes and emissions in different studies
Table 1| 生境类型 | 植被类型 | 土壤类型 | 观测 时间 | 年排放量 /(kg·hm-2) | 生长季排放量 占全年比重/% | 排放通量 /(μg·m-2·h-1) | 参考 文献 |
|---|
| 荒漠 | 藻类 | 结皮土 | 全年 | -0.19 | — | -2.12(均值) | [10] |
| 藻类 | 结皮土 | 季节性 | — | — | -18.5~9.8 | [33] |
| 苔藓 | 结皮土 | 全年 | -3.87 | — | -4.42(均值) | [10] |
| 苔藓 | 结皮土 | 季节性 | — | — | -15.5~4.5 | [33] |
| 混生 | 结皮土 | 全年 | -0.27 | — | -3.12(均值) | [10] |
| 草本 | 砂质土 | 全年 | 0.13 | — | 0~3.46 | [32] |
| 牧草 | 砂质土 | 全年 | 0.13 | 43 | 1.49(均值) | [30] |
| 稀疏灌木,草本 | 砂质土 | 夏季 | — | — | 0~8 | [11] |
| 干旱草原 | 草本 | 砂质土 | 全年 | 0.22 | 53 | -0.7~10.3 | [34] |
| 草本 | 砂质土 | 生长季 | — | — | 8.58(均值) | [35] |
| 半干旱草地 | 草本 | 砂质土 | 生长季 | — | — | -0.14~46.62 | [36] |
| 草本 | 细壤土 | 生长季 | — | — | 0~400 | [37] |
| 半干旱草原 | 草本 | 盐碱土 | 生长季 | 13.1 | 90 | 18.7(均值) | [38] |
| 牧草 | 砂质土 | 生长季 | — | — | 19.8(均值) | [29] |
| 草本 | 砂质土 | 生长季 | — | — | -0.34~0.12 | [39] |
| 草本 | 砂质土 | 生长季 | — | — | 2.8~3.9 | [40] |
| 草本 | 砂质土 | 生长季 | — | — | 3.4~12 | [41] |
| 苔藓 | 结皮土 | 生长季 | — | — | -0.42~0.84 | [42] |
| 草本 | 砂质土 | 生长季 | — | — | 4(均值) | [43] |
“—”代表无相关数据. ...
... 土壤中不同粒径组成造成土壤通透性差异较大,这对硝化作用和反硝化作用以及N2O扩散速率均有重要影响[55],同时不同粒径土壤团聚体中养分差异对N2O产生也有重要影响[56].荒漠区土壤砂粒含量普遍较高,但在不同生境斑块(如隐花植物、草本和灌木)中砂粒含量有所不同,其中流沙中砂粒含量最高、黏粒含量最低[10,29,57].相关研究表明,黏粒含量高的土壤N2O排放量较高,这主要与黏粒含量高的土壤保水能力强、孔隙度低以及O2含量低,有利于反硝化作用有关[55].此外,不同粒径土壤团聚体对N2O的生成也有不同影响[56],并且N2O排放量的研究结果存在差异[58-59],这主要与不同粒径的土壤团聚体在通透性、营养物质含量和微生物分布等方面不同有关[60]. ...
Are annual nitrous oxide fluxes sensitive to warming and increasing precipitation in the Gurbantunggut Desert?
7
2021
... 在新疆古尔班通古特荒漠草原的研究表明,自然气候条件下N2O排放通量(以N计量)较低(0~3.46 μg·m-2·h-1)[30],同时,此研究中2月的N2O通量为负值(-0.93 μg·m-2·h-1),表现为N2O的净吸收,这主要与土壤冰冻或积雪覆盖期N2O无法扩散至空气中,通过反硝化作用进一步还原为N2有关[30].另一项在内蒙古半干旱温带草地的研究同样表明,自然气候条件下N2O排放速率变化较大,有时表现为负值(-0.36~3.46 μg·m-2·h-1)[31]. ...
... [30].另一项在内蒙古半干旱温带草地的研究同样表明,自然气候条件下N2O排放速率变化较大,有时表现为负值(-0.36~3.46 μg·m-2·h-1)[31]. ...
... N2O排放量在不同研究中具有明显的差异性,如在古尔班通古特沙漠进行1年和连续3年的野外观测研究表明,土壤N2O年排放量为0.13 kg·hm-2[30,32].然而,在腾格里沙漠东南缘的研究表明,生物土壤结皮(Biological soil crust,BSC)的N2O年排放量为负值(平均为-1.44 kg·hm-2),表现为N2O年吸收[10],这可能主要与结皮性质有关,相对致密且渗水性低的生物结皮不利于N2O向大气扩散,土壤微生物促使N2O向N2转化.不同研究中,土壤N2O排放速率及排放量见表1[10-11,33-43]. ...
... N
2O emission fluxes and emissions in different studies
Table 1| 生境类型 | 植被类型 | 土壤类型 | 观测 时间 | 年排放量 /(kg·hm-2) | 生长季排放量 占全年比重/% | 排放通量 /(μg·m-2·h-1) | 参考 文献 |
|---|
| 荒漠 | 藻类 | 结皮土 | 全年 | -0.19 | — | -2.12(均值) | [10] |
| 藻类 | 结皮土 | 季节性 | — | — | -18.5~9.8 | [33] |
| 苔藓 | 结皮土 | 全年 | -3.87 | — | -4.42(均值) | [10] |
| 苔藓 | 结皮土 | 季节性 | — | — | -15.5~4.5 | [33] |
| 混生 | 结皮土 | 全年 | -0.27 | — | -3.12(均值) | [10] |
| 草本 | 砂质土 | 全年 | 0.13 | — | 0~3.46 | [32] |
| 牧草 | 砂质土 | 全年 | 0.13 | 43 | 1.49(均值) | [30] |
| 稀疏灌木,草本 | 砂质土 | 夏季 | — | — | 0~8 | [11] |
| 干旱草原 | 草本 | 砂质土 | 全年 | 0.22 | 53 | -0.7~10.3 | [34] |
| 草本 | 砂质土 | 生长季 | — | — | 8.58(均值) | [35] |
| 半干旱草地 | 草本 | 砂质土 | 生长季 | — | — | -0.14~46.62 | [36] |
| 草本 | 细壤土 | 生长季 | — | — | 0~400 | [37] |
| 半干旱草原 | 草本 | 盐碱土 | 生长季 | 13.1 | 90 | 18.7(均值) | [38] |
| 牧草 | 砂质土 | 生长季 | — | — | 19.8(均值) | [29] |
| 草本 | 砂质土 | 生长季 | — | — | -0.34~0.12 | [39] |
| 草本 | 砂质土 | 生长季 | — | — | 2.8~3.9 | [40] |
| 草本 | 砂质土 | 生长季 | — | — | 3.4~12 | [41] |
| 苔藓 | 结皮土 | 生长季 | — | — | -0.42~0.84 | [42] |
| 草本 | 砂质土 | 生长季 | — | — | 4(均值) | [43] |
“—”代表无相关数据. ...
... 荒漠土壤N2O排放具有明显的季节动态.在中国北方荒漠草原为期1年的观测结果表明,N2O排放通量整体表现为春夏季节较高(最大值为14.79 μg· m-2·h-1),冬季排放量较低甚至为负(最低值为-0.93 μg·m-2·h-1)[30],这主要是由于生长季土壤在多次降水脉冲后反复经历干湿循环过程,干旱阶段土壤微生物主要以硝化作用产生N2O;短暂湿润阶段土壤微生物主要以反硝化作用产生N2O,同时适宜的温度也有助于增加微生物活性,因此生长季荒漠土壤产生较多的N2O(占全年的43%~90%)[44];非生长季排放的N2O较少,主要与土壤含水量低、温度较低、微生物活性较低有关,同时冬季表层土壤冰冻或者被积雪覆盖导致N2O无法向大气扩散,故N2O排放减少[10]. ...
... 土壤中NH-N和NO-N是硝化细菌和反硝化细菌利用的底物,二者在土壤中的含量是影响N2O排放的关键因素[14].有研究表明,土壤中NH-N和NO-N含量增加将显著促进N2O的排放,使N2O年排放量增加55%~133%[30];同样,高有机氮含量(通过矿化作用将其转化为无机氮)也会促进N2O排放[15].但是在半干旱草原的模拟实验表明土壤氮含量增加对N2O排放的促进作用有限,施加氮肥(NH4NO3)会增加土壤N2O排放量(与不施肥相比约增加30%);土壤NO-N和NH-N含量的增加对N2O排放的促进作用弱于预期,这在很大程度上由植物-微生物对氮素的竞争所致,植物吸收了大部分的氮素[1]. ...
... 目前,有关降水变化如何影响土壤N2O排放通量的研究大多聚焦于农田和森林生态系统[62-64].研究表明,模拟增加降水的试验处理会刺激土壤排放N2O[65].也有学者持相反的观点,认为增加降水不会改变N2O排放[66],这主要取决于降水是否为该生态系统产生N2O的限制因子,如果降水是产生N2O的限制因子,增加降水会促使N2O排放;反之,则不会显著影响N2O排放.在新疆古尔班通古特荒漠草原的研究表明,增加降水(5 mm)未观察到N2O排放量的显著变化[30].但也有研究表明,通过模拟增加降水可以增加或减少N2O通量[31,67-68].荒漠土壤对降水响应不显著的原因可能是:一方面由于高温和较高的蒸散率,非有效降水对荒漠土壤水分影响的持续时间较短[32],并且土壤短暂的湿润过程无法保证微生物具有较高碳氮矿化的活性,营养物质积累较少;另一方面,自然降水后土壤水分渗透压的快速变化使得微生物死亡,导致参与N2O产生途径的微生物数目减少;土壤水分降低促进硝化作用与抑制反硝化作用相抵消[69]. ...
Response of N2O emission to water and nitrogen addition in temperate typical steppe soil in Inner Mongolia,China
2
2015
... 在新疆古尔班通古特荒漠草原的研究表明,自然气候条件下N2O排放通量(以N计量)较低(0~3.46 μg·m-2·h-1)[30],同时,此研究中2月的N2O通量为负值(-0.93 μg·m-2·h-1),表现为N2O的净吸收,这主要与土壤冰冻或积雪覆盖期N2O无法扩散至空气中,通过反硝化作用进一步还原为N2有关[30].另一项在内蒙古半干旱温带草地的研究同样表明,自然气候条件下N2O排放速率变化较大,有时表现为负值(-0.36~3.46 μg·m-2·h-1)[31]. ...
... 目前,有关降水变化如何影响土壤N2O排放通量的研究大多聚焦于农田和森林生态系统[62-64].研究表明,模拟增加降水的试验处理会刺激土壤排放N2O[65].也有学者持相反的观点,认为增加降水不会改变N2O排放[66],这主要取决于降水是否为该生态系统产生N2O的限制因子,如果降水是产生N2O的限制因子,增加降水会促使N2O排放;反之,则不会显著影响N2O排放.在新疆古尔班通古特荒漠草原的研究表明,增加降水(5 mm)未观察到N2O排放量的显著变化[30].但也有研究表明,通过模拟增加降水可以增加或减少N2O通量[31,67-68].荒漠土壤对降水响应不显著的原因可能是:一方面由于高温和较高的蒸散率,非有效降水对荒漠土壤水分影响的持续时间较短[32],并且土壤短暂的湿润过程无法保证微生物具有较高碳氮矿化的活性,营养物质积累较少;另一方面,自然降水后土壤水分渗透压的快速变化使得微生物死亡,导致参与N2O产生途径的微生物数目减少;土壤水分降低促进硝化作用与抑制反硝化作用相抵消[69]. ...
Fluxes of N2O,CH4 and soil respiration as affected by water and nitrogen addition in a temperate desert
3
2019
... N2O排放量在不同研究中具有明显的差异性,如在古尔班通古特沙漠进行1年和连续3年的野外观测研究表明,土壤N2O年排放量为0.13 kg·hm-2[30,32].然而,在腾格里沙漠东南缘的研究表明,生物土壤结皮(Biological soil crust,BSC)的N2O年排放量为负值(平均为-1.44 kg·hm-2),表现为N2O年吸收[10],这可能主要与结皮性质有关,相对致密且渗水性低的生物结皮不利于N2O向大气扩散,土壤微生物促使N2O向N2转化.不同研究中,土壤N2O排放速率及排放量见表1[10-11,33-43]. ...
... N
2O emission fluxes and emissions in different studies
Table 1| 生境类型 | 植被类型 | 土壤类型 | 观测 时间 | 年排放量 /(kg·hm-2) | 生长季排放量 占全年比重/% | 排放通量 /(μg·m-2·h-1) | 参考 文献 |
|---|
| 荒漠 | 藻类 | 结皮土 | 全年 | -0.19 | — | -2.12(均值) | [10] |
| 藻类 | 结皮土 | 季节性 | — | — | -18.5~9.8 | [33] |
| 苔藓 | 结皮土 | 全年 | -3.87 | — | -4.42(均值) | [10] |
| 苔藓 | 结皮土 | 季节性 | — | — | -15.5~4.5 | [33] |
| 混生 | 结皮土 | 全年 | -0.27 | — | -3.12(均值) | [10] |
| 草本 | 砂质土 | 全年 | 0.13 | — | 0~3.46 | [32] |
| 牧草 | 砂质土 | 全年 | 0.13 | 43 | 1.49(均值) | [30] |
| 稀疏灌木,草本 | 砂质土 | 夏季 | — | — | 0~8 | [11] |
| 干旱草原 | 草本 | 砂质土 | 全年 | 0.22 | 53 | -0.7~10.3 | [34] |
| 草本 | 砂质土 | 生长季 | — | — | 8.58(均值) | [35] |
| 半干旱草地 | 草本 | 砂质土 | 生长季 | — | — | -0.14~46.62 | [36] |
| 草本 | 细壤土 | 生长季 | — | — | 0~400 | [37] |
| 半干旱草原 | 草本 | 盐碱土 | 生长季 | 13.1 | 90 | 18.7(均值) | [38] |
| 牧草 | 砂质土 | 生长季 | — | — | 19.8(均值) | [29] |
| 草本 | 砂质土 | 生长季 | — | — | -0.34~0.12 | [39] |
| 草本 | 砂质土 | 生长季 | — | — | 2.8~3.9 | [40] |
| 草本 | 砂质土 | 生长季 | — | — | 3.4~12 | [41] |
| 苔藓 | 结皮土 | 生长季 | — | — | -0.42~0.84 | [42] |
| 草本 | 砂质土 | 生长季 | — | — | 4(均值) | [43] |
“—”代表无相关数据. ...
... 目前,有关降水变化如何影响土壤N2O排放通量的研究大多聚焦于农田和森林生态系统[62-64].研究表明,模拟增加降水的试验处理会刺激土壤排放N2O[65].也有学者持相反的观点,认为增加降水不会改变N2O排放[66],这主要取决于降水是否为该生态系统产生N2O的限制因子,如果降水是产生N2O的限制因子,增加降水会促使N2O排放;反之,则不会显著影响N2O排放.在新疆古尔班通古特荒漠草原的研究表明,增加降水(5 mm)未观察到N2O排放量的显著变化[30].但也有研究表明,通过模拟增加降水可以增加或减少N2O通量[31,67-68].荒漠土壤对降水响应不显著的原因可能是:一方面由于高温和较高的蒸散率,非有效降水对荒漠土壤水分影响的持续时间较短[32],并且土壤短暂的湿润过程无法保证微生物具有较高碳氮矿化的活性,营养物质积累较少;另一方面,自然降水后土壤水分渗透压的快速变化使得微生物死亡,导致参与N2O产生途径的微生物数目减少;土壤水分降低促进硝化作用与抑制反硝化作用相抵消[69]. ...
Reference for different sensitivities of greenhouse gases effluxes to warming climate among types of desert biological soil crust
4
2022
... N2O排放量在不同研究中具有明显的差异性,如在古尔班通古特沙漠进行1年和连续3年的野外观测研究表明,土壤N2O年排放量为0.13 kg·hm-2[30,32].然而,在腾格里沙漠东南缘的研究表明,生物土壤结皮(Biological soil crust,BSC)的N2O年排放量为负值(平均为-1.44 kg·hm-2),表现为N2O年吸收[10],这可能主要与结皮性质有关,相对致密且渗水性低的生物结皮不利于N2O向大气扩散,土壤微生物促使N2O向N2转化.不同研究中,土壤N2O排放速率及排放量见表1[10-11,33-43]. ...
... N
2O emission fluxes and emissions in different studies
Table 1| 生境类型 | 植被类型 | 土壤类型 | 观测 时间 | 年排放量 /(kg·hm-2) | 生长季排放量 占全年比重/% | 排放通量 /(μg·m-2·h-1) | 参考 文献 |
|---|
| 荒漠 | 藻类 | 结皮土 | 全年 | -0.19 | — | -2.12(均值) | [10] |
| 藻类 | 结皮土 | 季节性 | — | — | -18.5~9.8 | [33] |
| 苔藓 | 结皮土 | 全年 | -3.87 | — | -4.42(均值) | [10] |
| 苔藓 | 结皮土 | 季节性 | — | — | -15.5~4.5 | [33] |
| 混生 | 结皮土 | 全年 | -0.27 | — | -3.12(均值) | [10] |
| 草本 | 砂质土 | 全年 | 0.13 | — | 0~3.46 | [32] |
| 牧草 | 砂质土 | 全年 | 0.13 | 43 | 1.49(均值) | [30] |
| 稀疏灌木,草本 | 砂质土 | 夏季 | — | — | 0~8 | [11] |
| 干旱草原 | 草本 | 砂质土 | 全年 | 0.22 | 53 | -0.7~10.3 | [34] |
| 草本 | 砂质土 | 生长季 | — | — | 8.58(均值) | [35] |
| 半干旱草地 | 草本 | 砂质土 | 生长季 | — | — | -0.14~46.62 | [36] |
| 草本 | 细壤土 | 生长季 | — | — | 0~400 | [37] |
| 半干旱草原 | 草本 | 盐碱土 | 生长季 | 13.1 | 90 | 18.7(均值) | [38] |
| 牧草 | 砂质土 | 生长季 | — | — | 19.8(均值) | [29] |
| 草本 | 砂质土 | 生长季 | — | — | -0.34~0.12 | [39] |
| 草本 | 砂质土 | 生长季 | — | — | 2.8~3.9 | [40] |
| 草本 | 砂质土 | 生长季 | — | — | 3.4~12 | [41] |
| 苔藓 | 结皮土 | 生长季 | — | — | -0.42~0.84 | [42] |
| 草本 | 砂质土 | 生长季 | — | — | 4(均值) | [43] |
“—”代表无相关数据. ...
... [
33]
| 混生 | 结皮土 | 全年 | -0.27 | — | -3.12(均值) | [10] |
| 草本 | 砂质土 | 全年 | 0.13 | — | 0~3.46 | [32] |
| 牧草 | 砂质土 | 全年 | 0.13 | 43 | 1.49(均值) | [30] |
| 稀疏灌木,草本 | 砂质土 | 夏季 | — | — | 0~8 | [11] |
| 干旱草原 | 草本 | 砂质土 | 全年 | 0.22 | 53 | -0.7~10.3 | [34] |
| 草本 | 砂质土 | 生长季 | — | — | 8.58(均值) | [35] |
| 半干旱草地 | 草本 | 砂质土 | 生长季 | — | — | -0.14~46.62 | [36] |
| 草本 | 细壤土 | 生长季 | — | — | 0~400 | [37] |
| 半干旱草原 | 草本 | 盐碱土 | 生长季 | 13.1 | 90 | 18.7(均值) | [38] |
| 牧草 | 砂质土 | 生长季 | — | — | 19.8(均值) | [29] |
| 草本 | 砂质土 | 生长季 | — | — | -0.34~0.12 | [39] |
| 草本 | 砂质土 | 生长季 | — | — | 2.8~3.9 | [40] |
| 草本 | 砂质土 | 生长季 | — | — | 3.4~12 | [41] |
| 苔藓 | 结皮土 | 生长季 | — | — | -0.42~0.84 | [42] |
| 草本 | 砂质土 | 生长季 | — | — | 4(均值) | [43] |
“—”代表无相关数据. ...
... 有关不同类型生物土壤结皮的研究表明,藓类结皮、藻类结皮和混生结皮均表现为N2O负通量,即N2O净吸收,吸收量的大小为藓类结皮>混生结皮>藻类结皮[10].也有研究发现,藻类结皮和藓类结皮生长季(4—10月)的吸收通量显著高于非生长季[33].藓类结皮更易产生相对厌氧环境,促进了反硝化作用产生N2O,同时也阻止了N2O通过扩散从土壤进入大气,将N2O转化为N2,因此整体表现为N2O净吸收[10].养分更为贫瘠的风沙土却表现为N2O净排放,即使在土壤最干燥的时期N2O也会排放到大气中[17]. ...
Influence of increasing temperature and nitrogen input on greenhouse gas emissions from a desert steppe soil in Inner Mongolia
1
2011
... N
2O emission fluxes and emissions in different studies
Table 1| 生境类型 | 植被类型 | 土壤类型 | 观测 时间 | 年排放量 /(kg·hm-2) | 生长季排放量 占全年比重/% | 排放通量 /(μg·m-2·h-1) | 参考 文献 |
|---|
| 荒漠 | 藻类 | 结皮土 | 全年 | -0.19 | — | -2.12(均值) | [10] |
| 藻类 | 结皮土 | 季节性 | — | — | -18.5~9.8 | [33] |
| 苔藓 | 结皮土 | 全年 | -3.87 | — | -4.42(均值) | [10] |
| 苔藓 | 结皮土 | 季节性 | — | — | -15.5~4.5 | [33] |
| 混生 | 结皮土 | 全年 | -0.27 | — | -3.12(均值) | [10] |
| 草本 | 砂质土 | 全年 | 0.13 | — | 0~3.46 | [32] |
| 牧草 | 砂质土 | 全年 | 0.13 | 43 | 1.49(均值) | [30] |
| 稀疏灌木,草本 | 砂质土 | 夏季 | — | — | 0~8 | [11] |
| 干旱草原 | 草本 | 砂质土 | 全年 | 0.22 | 53 | -0.7~10.3 | [34] |
| 草本 | 砂质土 | 生长季 | — | — | 8.58(均值) | [35] |
| 半干旱草地 | 草本 | 砂质土 | 生长季 | — | — | -0.14~46.62 | [36] |
| 草本 | 细壤土 | 生长季 | — | — | 0~400 | [37] |
| 半干旱草原 | 草本 | 盐碱土 | 生长季 | 13.1 | 90 | 18.7(均值) | [38] |
| 牧草 | 砂质土 | 生长季 | — | — | 19.8(均值) | [29] |
| 草本 | 砂质土 | 生长季 | — | — | -0.34~0.12 | [39] |
| 草本 | 砂质土 | 生长季 | — | — | 2.8~3.9 | [40] |
| 草本 | 砂质土 | 生长季 | — | — | 3.4~12 | [41] |
| 苔藓 | 结皮土 | 生长季 | — | — | -0.42~0.84 | [42] |
| 草本 | 砂质土 | 生长季 | — | — | 4(均值) | [43] |
“—”代表无相关数据. ...
Emissions of N2O from Sonoran desert and effluent-irrigated grass ecosytems
1
1998
... N
2O emission fluxes and emissions in different studies
Table 1| 生境类型 | 植被类型 | 土壤类型 | 观测 时间 | 年排放量 /(kg·hm-2) | 生长季排放量 占全年比重/% | 排放通量 /(μg·m-2·h-1) | 参考 文献 |
|---|
| 荒漠 | 藻类 | 结皮土 | 全年 | -0.19 | — | -2.12(均值) | [10] |
| 藻类 | 结皮土 | 季节性 | — | — | -18.5~9.8 | [33] |
| 苔藓 | 结皮土 | 全年 | -3.87 | — | -4.42(均值) | [10] |
| 苔藓 | 结皮土 | 季节性 | — | — | -15.5~4.5 | [33] |
| 混生 | 结皮土 | 全年 | -0.27 | — | -3.12(均值) | [10] |
| 草本 | 砂质土 | 全年 | 0.13 | — | 0~3.46 | [32] |
| 牧草 | 砂质土 | 全年 | 0.13 | 43 | 1.49(均值) | [30] |
| 稀疏灌木,草本 | 砂质土 | 夏季 | — | — | 0~8 | [11] |
| 干旱草原 | 草本 | 砂质土 | 全年 | 0.22 | 53 | -0.7~10.3 | [34] |
| 草本 | 砂质土 | 生长季 | — | — | 8.58(均值) | [35] |
| 半干旱草地 | 草本 | 砂质土 | 生长季 | — | — | -0.14~46.62 | [36] |
| 草本 | 细壤土 | 生长季 | — | — | 0~400 | [37] |
| 半干旱草原 | 草本 | 盐碱土 | 生长季 | 13.1 | 90 | 18.7(均值) | [38] |
| 牧草 | 砂质土 | 生长季 | — | — | 19.8(均值) | [29] |
| 草本 | 砂质土 | 生长季 | — | — | -0.34~0.12 | [39] |
| 草本 | 砂质土 | 生长季 | — | — | 2.8~3.9 | [40] |
| 草本 | 砂质土 | 生长季 | — | — | 3.4~12 | [41] |
| 苔藓 | 结皮土 | 生长季 | — | — | -0.42~0.84 | [42] |
| 草本 | 砂质土 | 生长季 | — | — | 4(均值) | [43] |
“—”代表无相关数据. ...
Linking NO and N2O emission pulses with the mobilization of mineral and organic N upon rewetting dry soils
2
2017
... N
2O emission fluxes and emissions in different studies
Table 1| 生境类型 | 植被类型 | 土壤类型 | 观测 时间 | 年排放量 /(kg·hm-2) | 生长季排放量 占全年比重/% | 排放通量 /(μg·m-2·h-1) | 参考 文献 |
|---|
| 荒漠 | 藻类 | 结皮土 | 全年 | -0.19 | — | -2.12(均值) | [10] |
| 藻类 | 结皮土 | 季节性 | — | — | -18.5~9.8 | [33] |
| 苔藓 | 结皮土 | 全年 | -3.87 | — | -4.42(均值) | [10] |
| 苔藓 | 结皮土 | 季节性 | — | — | -15.5~4.5 | [33] |
| 混生 | 结皮土 | 全年 | -0.27 | — | -3.12(均值) | [10] |
| 草本 | 砂质土 | 全年 | 0.13 | — | 0~3.46 | [32] |
| 牧草 | 砂质土 | 全年 | 0.13 | 43 | 1.49(均值) | [30] |
| 稀疏灌木,草本 | 砂质土 | 夏季 | — | — | 0~8 | [11] |
| 干旱草原 | 草本 | 砂质土 | 全年 | 0.22 | 53 | -0.7~10.3 | [34] |
| 草本 | 砂质土 | 生长季 | — | — | 8.58(均值) | [35] |
| 半干旱草地 | 草本 | 砂质土 | 生长季 | — | — | -0.14~46.62 | [36] |
| 草本 | 细壤土 | 生长季 | — | — | 0~400 | [37] |
| 半干旱草原 | 草本 | 盐碱土 | 生长季 | 13.1 | 90 | 18.7(均值) | [38] |
| 牧草 | 砂质土 | 生长季 | — | — | 19.8(均值) | [29] |
| 草本 | 砂质土 | 生长季 | — | — | -0.34~0.12 | [39] |
| 草本 | 砂质土 | 生长季 | — | — | 2.8~3.9 | [40] |
| 草本 | 砂质土 | 生长季 | — | — | 3.4~12 | [41] |
| 苔藓 | 结皮土 | 生长季 | — | — | -0.42~0.84 | [42] |
| 草本 | 砂质土 | 生长季 | — | — | 4(均值) | [43] |
“—”代表无相关数据. ...
... 水分是荒漠生态系统不同时空尺度各种生物过程的重要驱动因子.降水作为主要的水分来源,由于其间断性和不可预知性,导致土壤水分与养分等关键资源的获得也呈不连续的脉冲状态[70].不同频率的降水脉冲导致土壤经历干湿循环,当土壤干燥且含水量降低时,碳氮可利用性和微生物活性较低,营养物质周转受到限制,N2O排放处于较低水平;土壤复湿后,碳氮可利用性和微生物活性较高,并激发土壤生物化学过程,并促进N2O排放[36,71].土壤湿度增加形成的厌氧条件也促进了反硝化作用产生N2O.同样,土壤频繁地经历干旱和湿润过程,也使硝化作用和反硝化作用交替进行,从而促进了N2O的产生和氮矿化(硝态氮和铵态氮的积累)[44,72].室内培养实验结果表明,不同降水频率(8天/18天)处理与恒湿处理相比,能够显著激发土壤N2O排放,排放量平均增加15倍[72],主要原因是干湿交替增加了微生物的死亡并促进土壤释放营养物质,使得土壤中氮的矿化量增加[73]. ...
Moisture pulses,trace gas emissions and soil C and N in cheatgrass and native grass-dominated sagebrush-steppe in Wyoming,USA
1
2008
... N
2O emission fluxes and emissions in different studies
Table 1| 生境类型 | 植被类型 | 土壤类型 | 观测 时间 | 年排放量 /(kg·hm-2) | 生长季排放量 占全年比重/% | 排放通量 /(μg·m-2·h-1) | 参考 文献 |
|---|
| 荒漠 | 藻类 | 结皮土 | 全年 | -0.19 | — | -2.12(均值) | [10] |
| 藻类 | 结皮土 | 季节性 | — | — | -18.5~9.8 | [33] |
| 苔藓 | 结皮土 | 全年 | -3.87 | — | -4.42(均值) | [10] |
| 苔藓 | 结皮土 | 季节性 | — | — | -15.5~4.5 | [33] |
| 混生 | 结皮土 | 全年 | -0.27 | — | -3.12(均值) | [10] |
| 草本 | 砂质土 | 全年 | 0.13 | — | 0~3.46 | [32] |
| 牧草 | 砂质土 | 全年 | 0.13 | 43 | 1.49(均值) | [30] |
| 稀疏灌木,草本 | 砂质土 | 夏季 | — | — | 0~8 | [11] |
| 干旱草原 | 草本 | 砂质土 | 全年 | 0.22 | 53 | -0.7~10.3 | [34] |
| 草本 | 砂质土 | 生长季 | — | — | 8.58(均值) | [35] |
| 半干旱草地 | 草本 | 砂质土 | 生长季 | — | — | -0.14~46.62 | [36] |
| 草本 | 细壤土 | 生长季 | — | — | 0~400 | [37] |
| 半干旱草原 | 草本 | 盐碱土 | 生长季 | 13.1 | 90 | 18.7(均值) | [38] |
| 牧草 | 砂质土 | 生长季 | — | — | 19.8(均值) | [29] |
| 草本 | 砂质土 | 生长季 | — | — | -0.34~0.12 | [39] |
| 草本 | 砂质土 | 生长季 | — | — | 2.8~3.9 | [40] |
| 草本 | 砂质土 | 生长季 | — | — | 3.4~12 | [41] |
| 苔藓 | 结皮土 | 生长季 | — | — | -0.42~0.84 | [42] |
| 草本 | 砂质土 | 生长季 | — | — | 4(均值) | [43] |
“—”代表无相关数据. ...
氮沉降、放牧和极端降水对羊草草地N2O排放的影响机制研究
1
2021
... N
2O emission fluxes and emissions in different studies
Table 1| 生境类型 | 植被类型 | 土壤类型 | 观测 时间 | 年排放量 /(kg·hm-2) | 生长季排放量 占全年比重/% | 排放通量 /(μg·m-2·h-1) | 参考 文献 |
|---|
| 荒漠 | 藻类 | 结皮土 | 全年 | -0.19 | — | -2.12(均值) | [10] |
| 藻类 | 结皮土 | 季节性 | — | — | -18.5~9.8 | [33] |
| 苔藓 | 结皮土 | 全年 | -3.87 | — | -4.42(均值) | [10] |
| 苔藓 | 结皮土 | 季节性 | — | — | -15.5~4.5 | [33] |
| 混生 | 结皮土 | 全年 | -0.27 | — | -3.12(均值) | [10] |
| 草本 | 砂质土 | 全年 | 0.13 | — | 0~3.46 | [32] |
| 牧草 | 砂质土 | 全年 | 0.13 | 43 | 1.49(均值) | [30] |
| 稀疏灌木,草本 | 砂质土 | 夏季 | — | — | 0~8 | [11] |
| 干旱草原 | 草本 | 砂质土 | 全年 | 0.22 | 53 | -0.7~10.3 | [34] |
| 草本 | 砂质土 | 生长季 | — | — | 8.58(均值) | [35] |
| 半干旱草地 | 草本 | 砂质土 | 生长季 | — | — | -0.14~46.62 | [36] |
| 草本 | 细壤土 | 生长季 | — | — | 0~400 | [37] |
| 半干旱草原 | 草本 | 盐碱土 | 生长季 | 13.1 | 90 | 18.7(均值) | [38] |
| 牧草 | 砂质土 | 生长季 | — | — | 19.8(均值) | [29] |
| 草本 | 砂质土 | 生长季 | — | — | -0.34~0.12 | [39] |
| 草本 | 砂质土 | 生长季 | — | — | 2.8~3.9 | [40] |
| 草本 | 砂质土 | 生长季 | — | — | 3.4~12 | [41] |
| 苔藓 | 结皮土 | 生长季 | — | — | -0.42~0.84 | [42] |
| 草本 | 砂质土 | 生长季 | — | — | 4(均值) | [43] |
“—”代表无相关数据. ...
Climate change reduces the net sink of CH4 and N2O in a semiarid grassland
1
2013
... N
2O emission fluxes and emissions in different studies
Table 1| 生境类型 | 植被类型 | 土壤类型 | 观测 时间 | 年排放量 /(kg·hm-2) | 生长季排放量 占全年比重/% | 排放通量 /(μg·m-2·h-1) | 参考 文献 |
|---|
| 荒漠 | 藻类 | 结皮土 | 全年 | -0.19 | — | -2.12(均值) | [10] |
| 藻类 | 结皮土 | 季节性 | — | — | -18.5~9.8 | [33] |
| 苔藓 | 结皮土 | 全年 | -3.87 | — | -4.42(均值) | [10] |
| 苔藓 | 结皮土 | 季节性 | — | — | -15.5~4.5 | [33] |
| 混生 | 结皮土 | 全年 | -0.27 | — | -3.12(均值) | [10] |
| 草本 | 砂质土 | 全年 | 0.13 | — | 0~3.46 | [32] |
| 牧草 | 砂质土 | 全年 | 0.13 | 43 | 1.49(均值) | [30] |
| 稀疏灌木,草本 | 砂质土 | 夏季 | — | — | 0~8 | [11] |
| 干旱草原 | 草本 | 砂质土 | 全年 | 0.22 | 53 | -0.7~10.3 | [34] |
| 草本 | 砂质土 | 生长季 | — | — | 8.58(均值) | [35] |
| 半干旱草地 | 草本 | 砂质土 | 生长季 | — | — | -0.14~46.62 | [36] |
| 草本 | 细壤土 | 生长季 | — | — | 0~400 | [37] |
| 半干旱草原 | 草本 | 盐碱土 | 生长季 | 13.1 | 90 | 18.7(均值) | [38] |
| 牧草 | 砂质土 | 生长季 | — | — | 19.8(均值) | [29] |
| 草本 | 砂质土 | 生长季 | — | — | -0.34~0.12 | [39] |
| 草本 | 砂质土 | 生长季 | — | — | 2.8~3.9 | [40] |
| 草本 | 砂质土 | 生长季 | — | — | 3.4~12 | [41] |
| 苔藓 | 结皮土 | 生长季 | — | — | -0.42~0.84 | [42] |
| 草本 | 砂质土 | 生长季 | — | — | 4(均值) | [43] |
“—”代表无相关数据. ...
Soil nitrogen and greenhouse gas dynamics in a temperate grassland under experimental warming and defoliation
1
2019
... N
2O emission fluxes and emissions in different studies
Table 1| 生境类型 | 植被类型 | 土壤类型 | 观测 时间 | 年排放量 /(kg·hm-2) | 生长季排放量 占全年比重/% | 排放通量 /(μg·m-2·h-1) | 参考 文献 |
|---|
| 荒漠 | 藻类 | 结皮土 | 全年 | -0.19 | — | -2.12(均值) | [10] |
| 藻类 | 结皮土 | 季节性 | — | — | -18.5~9.8 | [33] |
| 苔藓 | 结皮土 | 全年 | -3.87 | — | -4.42(均值) | [10] |
| 苔藓 | 结皮土 | 季节性 | — | — | -15.5~4.5 | [33] |
| 混生 | 结皮土 | 全年 | -0.27 | — | -3.12(均值) | [10] |
| 草本 | 砂质土 | 全年 | 0.13 | — | 0~3.46 | [32] |
| 牧草 | 砂质土 | 全年 | 0.13 | 43 | 1.49(均值) | [30] |
| 稀疏灌木,草本 | 砂质土 | 夏季 | — | — | 0~8 | [11] |
| 干旱草原 | 草本 | 砂质土 | 全年 | 0.22 | 53 | -0.7~10.3 | [34] |
| 草本 | 砂质土 | 生长季 | — | — | 8.58(均值) | [35] |
| 半干旱草地 | 草本 | 砂质土 | 生长季 | — | — | -0.14~46.62 | [36] |
| 草本 | 细壤土 | 生长季 | — | — | 0~400 | [37] |
| 半干旱草原 | 草本 | 盐碱土 | 生长季 | 13.1 | 90 | 18.7(均值) | [38] |
| 牧草 | 砂质土 | 生长季 | — | — | 19.8(均值) | [29] |
| 草本 | 砂质土 | 生长季 | — | — | -0.34~0.12 | [39] |
| 草本 | 砂质土 | 生长季 | — | — | 2.8~3.9 | [40] |
| 草本 | 砂质土 | 生长季 | — | — | 3.4~12 | [41] |
| 苔藓 | 结皮土 | 生长季 | — | — | -0.42~0.84 | [42] |
| 草本 | 砂质土 | 生长季 | — | — | 4(均值) | [43] |
“—”代表无相关数据. ...
Responses of greenhouse gas fluxes to climate extremes in a semiarid grassland
1
2016
... N
2O emission fluxes and emissions in different studies
Table 1| 生境类型 | 植被类型 | 土壤类型 | 观测 时间 | 年排放量 /(kg·hm-2) | 生长季排放量 占全年比重/% | 排放通量 /(μg·m-2·h-1) | 参考 文献 |
|---|
| 荒漠 | 藻类 | 结皮土 | 全年 | -0.19 | — | -2.12(均值) | [10] |
| 藻类 | 结皮土 | 季节性 | — | — | -18.5~9.8 | [33] |
| 苔藓 | 结皮土 | 全年 | -3.87 | — | -4.42(均值) | [10] |
| 苔藓 | 结皮土 | 季节性 | — | — | -15.5~4.5 | [33] |
| 混生 | 结皮土 | 全年 | -0.27 | — | -3.12(均值) | [10] |
| 草本 | 砂质土 | 全年 | 0.13 | — | 0~3.46 | [32] |
| 牧草 | 砂质土 | 全年 | 0.13 | 43 | 1.49(均值) | [30] |
| 稀疏灌木,草本 | 砂质土 | 夏季 | — | — | 0~8 | [11] |
| 干旱草原 | 草本 | 砂质土 | 全年 | 0.22 | 53 | -0.7~10.3 | [34] |
| 草本 | 砂质土 | 生长季 | — | — | 8.58(均值) | [35] |
| 半干旱草地 | 草本 | 砂质土 | 生长季 | — | — | -0.14~46.62 | [36] |
| 草本 | 细壤土 | 生长季 | — | — | 0~400 | [37] |
| 半干旱草原 | 草本 | 盐碱土 | 生长季 | 13.1 | 90 | 18.7(均值) | [38] |
| 牧草 | 砂质土 | 生长季 | — | — | 19.8(均值) | [29] |
| 草本 | 砂质土 | 生长季 | — | — | -0.34~0.12 | [39] |
| 草本 | 砂质土 | 生长季 | — | — | 2.8~3.9 | [40] |
| 草本 | 砂质土 | 生长季 | — | — | 3.4~12 | [41] |
| 苔藓 | 结皮土 | 生长季 | — | — | -0.42~0.84 | [42] |
| 草本 | 砂质土 | 生长季 | — | — | 4(均值) | [43] |
“—”代表无相关数据. ...
Biocrusts modulate responses of nitrous oxide and methane soil fluxes to simulated climate change in a mediterranean dryland
1
2020
... N
2O emission fluxes and emissions in different studies
Table 1| 生境类型 | 植被类型 | 土壤类型 | 观测 时间 | 年排放量 /(kg·hm-2) | 生长季排放量 占全年比重/% | 排放通量 /(μg·m-2·h-1) | 参考 文献 |
|---|
| 荒漠 | 藻类 | 结皮土 | 全年 | -0.19 | — | -2.12(均值) | [10] |
| 藻类 | 结皮土 | 季节性 | — | — | -18.5~9.8 | [33] |
| 苔藓 | 结皮土 | 全年 | -3.87 | — | -4.42(均值) | [10] |
| 苔藓 | 结皮土 | 季节性 | — | — | -15.5~4.5 | [33] |
| 混生 | 结皮土 | 全年 | -0.27 | — | -3.12(均值) | [10] |
| 草本 | 砂质土 | 全年 | 0.13 | — | 0~3.46 | [32] |
| 牧草 | 砂质土 | 全年 | 0.13 | 43 | 1.49(均值) | [30] |
| 稀疏灌木,草本 | 砂质土 | 夏季 | — | — | 0~8 | [11] |
| 干旱草原 | 草本 | 砂质土 | 全年 | 0.22 | 53 | -0.7~10.3 | [34] |
| 草本 | 砂质土 | 生长季 | — | — | 8.58(均值) | [35] |
| 半干旱草地 | 草本 | 砂质土 | 生长季 | — | — | -0.14~46.62 | [36] |
| 草本 | 细壤土 | 生长季 | — | — | 0~400 | [37] |
| 半干旱草原 | 草本 | 盐碱土 | 生长季 | 13.1 | 90 | 18.7(均值) | [38] |
| 牧草 | 砂质土 | 生长季 | — | — | 19.8(均值) | [29] |
| 草本 | 砂质土 | 生长季 | — | — | -0.34~0.12 | [39] |
| 草本 | 砂质土 | 生长季 | — | — | 2.8~3.9 | [40] |
| 草本 | 砂质土 | 生长季 | — | — | 3.4~12 | [41] |
| 苔藓 | 结皮土 | 生长季 | — | — | -0.42~0.84 | [42] |
| 草本 | 砂质土 | 生长季 | — | — | 4(均值) | [43] |
“—”代表无相关数据. ...
Effects of increasing precipitation and nitrogen deposition on CH4 and N2O fluxes and ecosystem respiration in a degraded steppe in Inner Mongolia,China
2
2013
... N2O排放量在不同研究中具有明显的差异性,如在古尔班通古特沙漠进行1年和连续3年的野外观测研究表明,土壤N2O年排放量为0.13 kg·hm-2[30,32].然而,在腾格里沙漠东南缘的研究表明,生物土壤结皮(Biological soil crust,BSC)的N2O年排放量为负值(平均为-1.44 kg·hm-2),表现为N2O年吸收[10],这可能主要与结皮性质有关,相对致密且渗水性低的生物结皮不利于N2O向大气扩散,土壤微生物促使N2O向N2转化.不同研究中,土壤N2O排放速率及排放量见表1[10-11,33-43]. ...
... N
2O emission fluxes and emissions in different studies
Table 1| 生境类型 | 植被类型 | 土壤类型 | 观测 时间 | 年排放量 /(kg·hm-2) | 生长季排放量 占全年比重/% | 排放通量 /(μg·m-2·h-1) | 参考 文献 |
|---|
| 荒漠 | 藻类 | 结皮土 | 全年 | -0.19 | — | -2.12(均值) | [10] |
| 藻类 | 结皮土 | 季节性 | — | — | -18.5~9.8 | [33] |
| 苔藓 | 结皮土 | 全年 | -3.87 | — | -4.42(均值) | [10] |
| 苔藓 | 结皮土 | 季节性 | — | — | -15.5~4.5 | [33] |
| 混生 | 结皮土 | 全年 | -0.27 | — | -3.12(均值) | [10] |
| 草本 | 砂质土 | 全年 | 0.13 | — | 0~3.46 | [32] |
| 牧草 | 砂质土 | 全年 | 0.13 | 43 | 1.49(均值) | [30] |
| 稀疏灌木,草本 | 砂质土 | 夏季 | — | — | 0~8 | [11] |
| 干旱草原 | 草本 | 砂质土 | 全年 | 0.22 | 53 | -0.7~10.3 | [34] |
| 草本 | 砂质土 | 生长季 | — | — | 8.58(均值) | [35] |
| 半干旱草地 | 草本 | 砂质土 | 生长季 | — | — | -0.14~46.62 | [36] |
| 草本 | 细壤土 | 生长季 | — | — | 0~400 | [37] |
| 半干旱草原 | 草本 | 盐碱土 | 生长季 | 13.1 | 90 | 18.7(均值) | [38] |
| 牧草 | 砂质土 | 生长季 | — | — | 19.8(均值) | [29] |
| 草本 | 砂质土 | 生长季 | — | — | -0.34~0.12 | [39] |
| 草本 | 砂质土 | 生长季 | — | — | 2.8~3.9 | [40] |
| 草本 | 砂质土 | 生长季 | — | — | 3.4~12 | [41] |
| 苔藓 | 结皮土 | 生长季 | — | — | -0.42~0.84 | [42] |
| 草本 | 砂质土 | 生长季 | — | — | 4(均值) | [43] |
“—”代表无相关数据. ...
Wetting-drying cycles do not increase organic carbon and nitrogen mineralization in soils with straw amendment
2
2017
... 荒漠土壤N2O排放具有明显的季节动态.在中国北方荒漠草原为期1年的观测结果表明,N2O排放通量整体表现为春夏季节较高(最大值为14.79 μg· m-2·h-1),冬季排放量较低甚至为负(最低值为-0.93 μg·m-2·h-1)[30],这主要是由于生长季土壤在多次降水脉冲后反复经历干湿循环过程,干旱阶段土壤微生物主要以硝化作用产生N2O;短暂湿润阶段土壤微生物主要以反硝化作用产生N2O,同时适宜的温度也有助于增加微生物活性,因此生长季荒漠土壤产生较多的N2O(占全年的43%~90%)[44];非生长季排放的N2O较少,主要与土壤含水量低、温度较低、微生物活性较低有关,同时冬季表层土壤冰冻或者被积雪覆盖导致N2O无法向大气扩散,故N2O排放减少[10]. ...
... 水分是荒漠生态系统不同时空尺度各种生物过程的重要驱动因子.降水作为主要的水分来源,由于其间断性和不可预知性,导致土壤水分与养分等关键资源的获得也呈不连续的脉冲状态[70].不同频率的降水脉冲导致土壤经历干湿循环,当土壤干燥且含水量降低时,碳氮可利用性和微生物活性较低,营养物质周转受到限制,N2O排放处于较低水平;土壤复湿后,碳氮可利用性和微生物活性较高,并激发土壤生物化学过程,并促进N2O排放[36,71].土壤湿度增加形成的厌氧条件也促进了反硝化作用产生N2O.同样,土壤频繁地经历干旱和湿润过程,也使硝化作用和反硝化作用交替进行,从而促进了N2O的产生和氮矿化(硝态氮和铵态氮的积累)[44,72].室内培养实验结果表明,不同降水频率(8天/18天)处理与恒湿处理相比,能够显著激发土壤N2O排放,排放量平均增加15倍[72],主要原因是干湿交替增加了微生物的死亡并促进土壤释放营养物质,使得土壤中氮的矿化量增加[73]. ...
Initial soil formation by biocrusts:nitrogen demand and clay protection control microbial necromass accrual and recycling
1
2022
... 土壤有机碳是影响异养硝化细菌和反硝化细菌活性的重要因素[14].荒漠土壤有机碳的主要来源为植物凋落物分解、根系分泌和微生物分泌、死亡残体释放[45].自然降水后,雨水促进植物和生物结皮生长,有利于植物根系、生物结皮对土壤有机碳的输入;同时,土壤水势增加使得微生物承受较高的渗透压力,其中一部分微生物死亡释放出有机物质,另一部分通过释放大量易分解含碳化合物(包括小分子含碳化合物和可溶性碳)来适应迅速增加的土壤水势.有机碳可为微生物提供重要碳源[16],有机碳供给增加一方面可加快微生物繁殖,消耗氧气从而形成厌氧环境促进反硝化作用发生,另一方面有机碳作为能量来源,促进微生物新陈代谢和物质循环,同时也为反硝化细菌提供电子从而促进反硝化作用[46]. ...
Variation in N2O emission and N2O related microbial functional genes in straw-and biochar-amended and non-amended soils
1
2019
... 土壤有机碳是影响异养硝化细菌和反硝化细菌活性的重要因素[14].荒漠土壤有机碳的主要来源为植物凋落物分解、根系分泌和微生物分泌、死亡残体释放[45].自然降水后,雨水促进植物和生物结皮生长,有利于植物根系、生物结皮对土壤有机碳的输入;同时,土壤水势增加使得微生物承受较高的渗透压力,其中一部分微生物死亡释放出有机物质,另一部分通过释放大量易分解含碳化合物(包括小分子含碳化合物和可溶性碳)来适应迅速增加的土壤水势.有机碳可为微生物提供重要碳源[16],有机碳供给增加一方面可加快微生物繁殖,消耗氧气从而形成厌氧环境促进反硝化作用发生,另一方面有机碳作为能量来源,促进微生物新陈代谢和物质循环,同时也为反硝化细菌提供电子从而促进反硝化作用[46]. ...
Denitrification in the top-soil of managed grasslands in the netherlands in relation to soil type and fertilizer level
1
1994
... 土壤pH是影响硝化作用和反硝化作用的重要因素,进而也影响土壤N2O排放.当土壤pH为5.6~8.0时,硝化速率随pH的升高而增大.当土壤pH从4.7增加到6.5时,硝化速率增加可达3~5倍[47],此时N2O主要由硝化作用产生;当土壤pH较低时,N2O成为反硝化作用的主要产物,其原因是低pH能抑制N2O还原酶的活性,从而增大N2O在反硝化产物中的比例. ...
Heterotrophic nitrification of organic N and its contribution to nitrous oxide emissions in soils
1
2015
... pH影响参与氮转化的相关过程,如反硝化、自养和异养硝化,这些生物过程都与N2O的产生有关[48].有研究指出,增加土壤pH促进N2O向N2的转化,整体表现为低N2O排放[15],主要原因在于pH影响细菌和真菌活性,大多数最适宜反硝化微生物生长的pH为6.0~8.0的中性环境,在pH较高或较低时反硝化作用速率较低[49-50].同时,pH也会通过调控AOA和AOB的群落结构影响N2O通量,例如AOB在中性、碱性土壤中占主导,而AOA在酸性土壤中占主导[51]. ...
Insights into the effect of soil pH on N2O and N2 emissions and denitrifier community size and activity
1
2010
... pH影响参与氮转化的相关过程,如反硝化、自养和异养硝化,这些生物过程都与N2O的产生有关[48].有研究指出,增加土壤pH促进N2O向N2的转化,整体表现为低N2O排放[15],主要原因在于pH影响细菌和真菌活性,大多数最适宜反硝化微生物生长的pH为6.0~8.0的中性环境,在pH较高或较低时反硝化作用速率较低[49-50].同时,pH也会通过调控AOA和AOB的群落结构影响N2O通量,例如AOB在中性、碱性土壤中占主导,而AOA在酸性土壤中占主导[51]. ...
1
1989
... pH影响参与氮转化的相关过程,如反硝化、自养和异养硝化,这些生物过程都与N2O的产生有关[48].有研究指出,增加土壤pH促进N2O向N2的转化,整体表现为低N2O排放[15],主要原因在于pH影响细菌和真菌活性,大多数最适宜反硝化微生物生长的pH为6.0~8.0的中性环境,在pH较高或较低时反硝化作用速率较低[49-50].同时,pH也会通过调控AOA和AOB的群落结构影响N2O通量,例如AOB在中性、碱性土壤中占主导,而AOA在酸性土壤中占主导[51]. ...
Multi-factorial drivers of ammonia oxidizer communities:evidence from a national soil survey
1
2013
... pH影响参与氮转化的相关过程,如反硝化、自养和异养硝化,这些生物过程都与N2O的产生有关[48].有研究指出,增加土壤pH促进N2O向N2的转化,整体表现为低N2O排放[15],主要原因在于pH影响细菌和真菌活性,大多数最适宜反硝化微生物生长的pH为6.0~8.0的中性环境,在pH较高或较低时反硝化作用速率较低[49-50].同时,pH也会通过调控AOA和AOB的群落结构影响N2O通量,例如AOB在中性、碱性土壤中占主导,而AOA在酸性土壤中占主导[51]. ...
A comparison of NO and N2O production by the autotrophic nitrifier nitrosomonas-europaea and the heterotrophic nitrifier alcaligenes-faecalis
1
1993
... 土壤O2含量是影响土壤N2O排放的重要因素,参与硝化作用和反硝化作用的微生物对O2含量有一定的需求.硝化作用是严格的好氧过程,硝化微生物及相关酶类必须在有氧条件下才能正常进行硝化作用,随土壤O2含量降低,可增加硝化作用产物中N2O的比例[52];相反,反硝化作用则是厌氧过程,仅在低氧或无氧的条件下发生,O2含量主要通过影响与反硝化作用相关的酶活性,进而影响N2O生成速率及其在反硝化产物中的比例.研究表明,当土壤中的O2含量低于5 mL·L-1时反硝化微生物的活性显著增加[53],且反硝化微生物对O2含量的耐受范围较广,可在不同O2含量下产生N2O[54]. ...
Denitrification:production and consumption of nitric oxide
1
1994
... 土壤O2含量是影响土壤N2O排放的重要因素,参与硝化作用和反硝化作用的微生物对O2含量有一定的需求.硝化作用是严格的好氧过程,硝化微生物及相关酶类必须在有氧条件下才能正常进行硝化作用,随土壤O2含量降低,可增加硝化作用产物中N2O的比例[52];相反,反硝化作用则是厌氧过程,仅在低氧或无氧的条件下发生,O2含量主要通过影响与反硝化作用相关的酶活性,进而影响N2O生成速率及其在反硝化产物中的比例.研究表明,当土壤中的O2含量低于5 mL·L-1时反硝化微生物的活性显著增加[53],且反硝化微生物对O2含量的耐受范围较广,可在不同O2含量下产生N2O[54]. ...
Ammonia oxidation pathways and nitrifier denitrification are significant sources of N2O and NO under low oxygen availability
1
2013
... 土壤O2含量是影响土壤N2O排放的重要因素,参与硝化作用和反硝化作用的微生物对O2含量有一定的需求.硝化作用是严格的好氧过程,硝化微生物及相关酶类必须在有氧条件下才能正常进行硝化作用,随土壤O2含量降低,可增加硝化作用产物中N2O的比例[52];相反,反硝化作用则是厌氧过程,仅在低氧或无氧的条件下发生,O2含量主要通过影响与反硝化作用相关的酶活性,进而影响N2O生成速率及其在反硝化产物中的比例.研究表明,当土壤中的O2含量低于5 mL·L-1时反硝化微生物的活性显著增加[53],且反硝化微生物对O2含量的耐受范围较广,可在不同O2含量下产生N2O[54]. ...
农田土壤N2O排放的主要影响因素及减排措施研究进展
2
2009
... 土壤中不同粒径组成造成土壤通透性差异较大,这对硝化作用和反硝化作用以及N2O扩散速率均有重要影响[55],同时不同粒径土壤团聚体中养分差异对N2O产生也有重要影响[56].荒漠区土壤砂粒含量普遍较高,但在不同生境斑块(如隐花植物、草本和灌木)中砂粒含量有所不同,其中流沙中砂粒含量最高、黏粒含量最低[10,29,57].相关研究表明,黏粒含量高的土壤N2O排放量较高,这主要与黏粒含量高的土壤保水能力强、孔隙度低以及O2含量低,有利于反硝化作用有关[55].此外,不同粒径土壤团聚体对N2O的生成也有不同影响[56],并且N2O排放量的研究结果存在差异[58-59],这主要与不同粒径的土壤团聚体在通透性、营养物质含量和微生物分布等方面不同有关[60]. ...
... [55].此外,不同粒径土壤团聚体对N2O的生成也有不同影响[56],并且N2O排放量的研究结果存在差异[58-59],这主要与不同粒径的土壤团聚体在通透性、营养物质含量和微生物分布等方面不同有关[60]. ...
Water pulses and biogeochemical cycles in arid and semiarid ecosystems
2
2004
... 土壤中不同粒径组成造成土壤通透性差异较大,这对硝化作用和反硝化作用以及N2O扩散速率均有重要影响[55],同时不同粒径土壤团聚体中养分差异对N2O产生也有重要影响[56].荒漠区土壤砂粒含量普遍较高,但在不同生境斑块(如隐花植物、草本和灌木)中砂粒含量有所不同,其中流沙中砂粒含量最高、黏粒含量最低[10,29,57].相关研究表明,黏粒含量高的土壤N2O排放量较高,这主要与黏粒含量高的土壤保水能力强、孔隙度低以及O2含量低,有利于反硝化作用有关[55].此外,不同粒径土壤团聚体对N2O的生成也有不同影响[56],并且N2O排放量的研究结果存在差异[58-59],这主要与不同粒径的土壤团聚体在通透性、营养物质含量和微生物分布等方面不同有关[60]. ...
... [56],并且N2O排放量的研究结果存在差异[58-59],这主要与不同粒径的土壤团聚体在通透性、营养物质含量和微生物分布等方面不同有关[60]. ...
Four years of experimental climate change modifies the microbial drivers of N2O fluxes in an upland grassland ecosystem
1
2012
... 土壤中不同粒径组成造成土壤通透性差异较大,这对硝化作用和反硝化作用以及N2O扩散速率均有重要影响[55],同时不同粒径土壤团聚体中养分差异对N2O产生也有重要影响[56].荒漠区土壤砂粒含量普遍较高,但在不同生境斑块(如隐花植物、草本和灌木)中砂粒含量有所不同,其中流沙中砂粒含量最高、黏粒含量最低[10,29,57].相关研究表明,黏粒含量高的土壤N2O排放量较高,这主要与黏粒含量高的土壤保水能力强、孔隙度低以及O2含量低,有利于反硝化作用有关[55].此外,不同粒径土壤团聚体对N2O的生成也有不同影响[56],并且N2O排放量的研究结果存在差异[58-59],这主要与不同粒径的土壤团聚体在通透性、营养物质含量和微生物分布等方面不同有关[60]. ...
Effects of transient anaerobic conditions in the presence of acetylene on subsequent aerobic respiration and N2O emission by soil aggregates
1
2005
... 土壤中不同粒径组成造成土壤通透性差异较大,这对硝化作用和反硝化作用以及N2O扩散速率均有重要影响[55],同时不同粒径土壤团聚体中养分差异对N2O产生也有重要影响[56].荒漠区土壤砂粒含量普遍较高,但在不同生境斑块(如隐花植物、草本和灌木)中砂粒含量有所不同,其中流沙中砂粒含量最高、黏粒含量最低[10,29,57].相关研究表明,黏粒含量高的土壤N2O排放量较高,这主要与黏粒含量高的土壤保水能力强、孔隙度低以及O2含量低,有利于反硝化作用有关[55].此外,不同粒径土壤团聚体对N2O的生成也有不同影响[56],并且N2O排放量的研究结果存在差异[58-59],这主要与不同粒径的土壤团聚体在通透性、营养物质含量和微生物分布等方面不同有关[60]. ...
Soil aggregates as biogeochemical reactors and implications for soil-atmosphere exchange of greenhouse gases:a concept
1
2019
... 土壤中不同粒径组成造成土壤通透性差异较大,这对硝化作用和反硝化作用以及N2O扩散速率均有重要影响[55],同时不同粒径土壤团聚体中养分差异对N2O产生也有重要影响[56].荒漠区土壤砂粒含量普遍较高,但在不同生境斑块(如隐花植物、草本和灌木)中砂粒含量有所不同,其中流沙中砂粒含量最高、黏粒含量最低[10,29,57].相关研究表明,黏粒含量高的土壤N2O排放量较高,这主要与黏粒含量高的土壤保水能力强、孔隙度低以及O2含量低,有利于反硝化作用有关[55].此外,不同粒径土壤团聚体对N2O的生成也有不同影响[56],并且N2O排放量的研究结果存在差异[58-59],这主要与不同粒径的土壤团聚体在通透性、营养物质含量和微生物分布等方面不同有关[60]. ...
Small-scale heterogeneity in carbon dioxide,nitrous oxide and methane production from aggregates of a cultivated sandy-loam soil
1
2008
... 土壤中不同粒径组成造成土壤通透性差异较大,这对硝化作用和反硝化作用以及N2O扩散速率均有重要影响[55],同时不同粒径土壤团聚体中养分差异对N2O产生也有重要影响[56].荒漠区土壤砂粒含量普遍较高,但在不同生境斑块(如隐花植物、草本和灌木)中砂粒含量有所不同,其中流沙中砂粒含量最高、黏粒含量最低[10,29,57].相关研究表明,黏粒含量高的土壤N2O排放量较高,这主要与黏粒含量高的土壤保水能力强、孔隙度低以及O2含量低,有利于反硝化作用有关[55].此外,不同粒径土壤团聚体对N2O的生成也有不同影响[56],并且N2O排放量的研究结果存在差异[58-59],这主要与不同粒径的土壤团聚体在通透性、营养物质含量和微生物分布等方面不同有关[60]. ...
High N2O emissions in dry ecosystems
2
2013
... 荒漠区植被具有明显的斑块性,其主要类型包括:一是由草本和灌木组成的植被类型;二是生物土壤结皮斑块,主要由地衣、苔藓、藻类等和土壤微生物组成,其特征是存在透水性弱的土壤表层.在荒漠区中一些豆科植物,如柠条(Caragana korshinskii)、花棒(Hedysarum scoparium)等及微生物(固氮蓝藻)本身具有固氮功能,为土壤贡献了显著的氮[61],这对N2O排放也有重要影响.研究表明,灌木土壤,如矮梭梭属(Hammada sp.)、附药蓬属(Noaea sp.)和刺地榆属(Sarcopoterium sp.)的N2O排放量大于混生结皮土壤(由藻类和苔藓组成)[61]. ...
... [61]. ...
Global agriculture and nitrous oxide emissions
1
2012
... 目前,有关降水变化如何影响土壤N2O排放通量的研究大多聚焦于农田和森林生态系统[62-64].研究表明,模拟增加降水的试验处理会刺激土壤排放N2O[65].也有学者持相反的观点,认为增加降水不会改变N2O排放[66],这主要取决于降水是否为该生态系统产生N2O的限制因子,如果降水是产生N2O的限制因子,增加降水会促使N2O排放;反之,则不会显著影响N2O排放.在新疆古尔班通古特荒漠草原的研究表明,增加降水(5 mm)未观察到N2O排放量的显著变化[30].但也有研究表明,通过模拟增加降水可以增加或减少N2O通量[31,67-68].荒漠土壤对降水响应不显著的原因可能是:一方面由于高温和较高的蒸散率,非有效降水对荒漠土壤水分影响的持续时间较短[32],并且土壤短暂的湿润过程无法保证微生物具有较高碳氮矿化的活性,营养物质积累较少;另一方面,自然降水后土壤水分渗透压的快速变化使得微生物死亡,导致参与N2O产生途径的微生物数目减少;土壤水分降低促进硝化作用与抑制反硝化作用相抵消[69]. ...
A global meta-analysis of nitrous oxide emission from drip irrigated cropping system
0
2021
In-depth analysis of N2O fluxes in tropical forest soils of the Congo Basin combining isotope and functional gene analysis
1
2021
... 目前,有关降水变化如何影响土壤N2O排放通量的研究大多聚焦于农田和森林生态系统[62-64].研究表明,模拟增加降水的试验处理会刺激土壤排放N2O[65].也有学者持相反的观点,认为增加降水不会改变N2O排放[66],这主要取决于降水是否为该生态系统产生N2O的限制因子,如果降水是产生N2O的限制因子,增加降水会促使N2O排放;反之,则不会显著影响N2O排放.在新疆古尔班通古特荒漠草原的研究表明,增加降水(5 mm)未观察到N2O排放量的显著变化[30].但也有研究表明,通过模拟增加降水可以增加或减少N2O通量[31,67-68].荒漠土壤对降水响应不显著的原因可能是:一方面由于高温和较高的蒸散率,非有效降水对荒漠土壤水分影响的持续时间较短[32],并且土壤短暂的湿润过程无法保证微生物具有较高碳氮矿化的活性,营养物质积累较少;另一方面,自然降水后土壤水分渗透压的快速变化使得微生物死亡,导致参与N2O产生途径的微生物数目减少;土壤水分降低促进硝化作用与抑制反硝化作用相抵消[69]. ...
Precipitation and nitrogen application stimulation soil nitrous oxide emission
1
2021
... 目前,有关降水变化如何影响土壤N2O排放通量的研究大多聚焦于农田和森林生态系统[62-64].研究表明,模拟增加降水的试验处理会刺激土壤排放N2O[65].也有学者持相反的观点,认为增加降水不会改变N2O排放[66],这主要取决于降水是否为该生态系统产生N2O的限制因子,如果降水是产生N2O的限制因子,增加降水会促使N2O排放;反之,则不会显著影响N2O排放.在新疆古尔班通古特荒漠草原的研究表明,增加降水(5 mm)未观察到N2O排放量的显著变化[30].但也有研究表明,通过模拟增加降水可以增加或减少N2O通量[31,67-68].荒漠土壤对降水响应不显著的原因可能是:一方面由于高温和较高的蒸散率,非有效降水对荒漠土壤水分影响的持续时间较短[32],并且土壤短暂的湿润过程无法保证微生物具有较高碳氮矿化的活性,营养物质积累较少;另一方面,自然降水后土壤水分渗透压的快速变化使得微生物死亡,导致参与N2O产生途径的微生物数目减少;土壤水分降低促进硝化作用与抑制反硝化作用相抵消[69]. ...
Effects of frequency and intensity of drying-rewetting cycles on Hydrocotyle vulgaris growth and greenhouse gas emissions from wetland microcosms
1
2018
... 目前,有关降水变化如何影响土壤N2O排放通量的研究大多聚焦于农田和森林生态系统[62-64].研究表明,模拟增加降水的试验处理会刺激土壤排放N2O[65].也有学者持相反的观点,认为增加降水不会改变N2O排放[66],这主要取决于降水是否为该生态系统产生N2O的限制因子,如果降水是产生N2O的限制因子,增加降水会促使N2O排放;反之,则不会显著影响N2O排放.在新疆古尔班通古特荒漠草原的研究表明,增加降水(5 mm)未观察到N2O排放量的显著变化[30].但也有研究表明,通过模拟增加降水可以增加或减少N2O通量[31,67-68].荒漠土壤对降水响应不显著的原因可能是:一方面由于高温和较高的蒸散率,非有效降水对荒漠土壤水分影响的持续时间较短[32],并且土壤短暂的湿润过程无法保证微生物具有较高碳氮矿化的活性,营养物质积累较少;另一方面,自然降水后土壤水分渗透压的快速变化使得微生物死亡,导致参与N2O产生途径的微生物数目减少;土壤水分降低促进硝化作用与抑制反硝化作用相抵消[69]. ...
Soil nitrogen transformation responses to seasonal precipitation changes are regulated by changes in functional microbial abundance in a subtropical forest
1
2017
... 目前,有关降水变化如何影响土壤N2O排放通量的研究大多聚焦于农田和森林生态系统[62-64].研究表明,模拟增加降水的试验处理会刺激土壤排放N2O[65].也有学者持相反的观点,认为增加降水不会改变N2O排放[66],这主要取决于降水是否为该生态系统产生N2O的限制因子,如果降水是产生N2O的限制因子,增加降水会促使N2O排放;反之,则不会显著影响N2O排放.在新疆古尔班通古特荒漠草原的研究表明,增加降水(5 mm)未观察到N2O排放量的显著变化[30].但也有研究表明,通过模拟增加降水可以增加或减少N2O通量[31,67-68].荒漠土壤对降水响应不显著的原因可能是:一方面由于高温和较高的蒸散率,非有效降水对荒漠土壤水分影响的持续时间较短[32],并且土壤短暂的湿润过程无法保证微生物具有较高碳氮矿化的活性,营养物质积累较少;另一方面,自然降水后土壤水分渗透压的快速变化使得微生物死亡,导致参与N2O产生途径的微生物数目减少;土壤水分降低促进硝化作用与抑制反硝化作用相抵消[69]. ...
Effects of precipitation exclusion on N2O emissions in a savanna ecosystem in SW China
1
2018
... 目前,有关降水变化如何影响土壤N2O排放通量的研究大多聚焦于农田和森林生态系统[62-64].研究表明,模拟增加降水的试验处理会刺激土壤排放N2O[65].也有学者持相反的观点,认为增加降水不会改变N2O排放[66],这主要取决于降水是否为该生态系统产生N2O的限制因子,如果降水是产生N2O的限制因子,增加降水会促使N2O排放;反之,则不会显著影响N2O排放.在新疆古尔班通古特荒漠草原的研究表明,增加降水(5 mm)未观察到N2O排放量的显著变化[30].但也有研究表明,通过模拟增加降水可以增加或减少N2O通量[31,67-68].荒漠土壤对降水响应不显著的原因可能是:一方面由于高温和较高的蒸散率,非有效降水对荒漠土壤水分影响的持续时间较短[32],并且土壤短暂的湿润过程无法保证微生物具有较高碳氮矿化的活性,营养物质积累较少;另一方面,自然降水后土壤水分渗透压的快速变化使得微生物死亡,导致参与N2O产生途径的微生物数目减少;土壤水分降低促进硝化作用与抑制反硝化作用相抵消[69]. ...
Linkage of plant and abiotic properties to the abundance and activity of N-cycling microbial communities in Tibetan permafrost-affected regions
2
2019
... 目前,有关降水变化如何影响土壤N2O排放通量的研究大多聚焦于农田和森林生态系统[62-64].研究表明,模拟增加降水的试验处理会刺激土壤排放N2O[65].也有学者持相反的观点,认为增加降水不会改变N2O排放[66],这主要取决于降水是否为该生态系统产生N2O的限制因子,如果降水是产生N2O的限制因子,增加降水会促使N2O排放;反之,则不会显著影响N2O排放.在新疆古尔班通古特荒漠草原的研究表明,增加降水(5 mm)未观察到N2O排放量的显著变化[30].但也有研究表明,通过模拟增加降水可以增加或减少N2O通量[31,67-68].荒漠土壤对降水响应不显著的原因可能是:一方面由于高温和较高的蒸散率,非有效降水对荒漠土壤水分影响的持续时间较短[32],并且土壤短暂的湿润过程无法保证微生物具有较高碳氮矿化的活性,营养物质积累较少;另一方面,自然降水后土壤水分渗透压的快速变化使得微生物死亡,导致参与N2O产生途径的微生物数目减少;土壤水分降低促进硝化作用与抑制反硝化作用相抵消[69]. ...
... 许多与氮循环相关的功能基因,包括amoA (AOA和AOB)、nirS、nirK、norB和nosZ,是指示土壤N2O排放的潜在有效指标[69,84,90-91].其中,amoA (AOA)和amoB (AOB)基因编码蛋白为氨单加氧酶,主要参与土壤中的氨氧化过程[91];nirS和nirK基因编码的蛋白为硝酸还原酶,其参与了NO向NO的转换,是反硝化作用(以及N2O生成)中的重要步骤[92];norB基因编码NO还原酶,该酶参与生成N2O;nosZ基因编码N2O还原酶,其丰度是指示N2O转化为N2的有效指标.因此,在厌氧条件下N2O排放量减少与nosZ基因表达增加有关[93]. ...
Negative effects of multiple global change factors on soil microbial diversity
1
2021
... 水分是荒漠生态系统不同时空尺度各种生物过程的重要驱动因子.降水作为主要的水分来源,由于其间断性和不可预知性,导致土壤水分与养分等关键资源的获得也呈不连续的脉冲状态[70].不同频率的降水脉冲导致土壤经历干湿循环,当土壤干燥且含水量降低时,碳氮可利用性和微生物活性较低,营养物质周转受到限制,N2O排放处于较低水平;土壤复湿后,碳氮可利用性和微生物活性较高,并激发土壤生物化学过程,并促进N2O排放[36,71].土壤湿度增加形成的厌氧条件也促进了反硝化作用产生N2O.同样,土壤频繁地经历干旱和湿润过程,也使硝化作用和反硝化作用交替进行,从而促进了N2O的产生和氮矿化(硝态氮和铵态氮的积累)[44,72].室内培养实验结果表明,不同降水频率(8天/18天)处理与恒湿处理相比,能够显著激发土壤N2O排放,排放量平均增加15倍[72],主要原因是干湿交替增加了微生物的死亡并促进土壤释放营养物质,使得土壤中氮的矿化量增加[73]. ...
Responses of soil nitrogen and phosphorus cycling to drying and rewetting cycles:a meta-analysis
1
2020
... 水分是荒漠生态系统不同时空尺度各种生物过程的重要驱动因子.降水作为主要的水分来源,由于其间断性和不可预知性,导致土壤水分与养分等关键资源的获得也呈不连续的脉冲状态[70].不同频率的降水脉冲导致土壤经历干湿循环,当土壤干燥且含水量降低时,碳氮可利用性和微生物活性较低,营养物质周转受到限制,N2O排放处于较低水平;土壤复湿后,碳氮可利用性和微生物活性较高,并激发土壤生物化学过程,并促进N2O排放[36,71].土壤湿度增加形成的厌氧条件也促进了反硝化作用产生N2O.同样,土壤频繁地经历干旱和湿润过程,也使硝化作用和反硝化作用交替进行,从而促进了N2O的产生和氮矿化(硝态氮和铵态氮的积累)[44,72].室内培养实验结果表明,不同降水频率(8天/18天)处理与恒湿处理相比,能够显著激发土壤N2O排放,排放量平均增加15倍[72],主要原因是干湿交替增加了微生物的死亡并促进土壤释放营养物质,使得土壤中氮的矿化量增加[73]. ...
Compaction effects on CO2 and N2O production during drying and rewetting of soil
2
2009
... 水分是荒漠生态系统不同时空尺度各种生物过程的重要驱动因子.降水作为主要的水分来源,由于其间断性和不可预知性,导致土壤水分与养分等关键资源的获得也呈不连续的脉冲状态[70].不同频率的降水脉冲导致土壤经历干湿循环,当土壤干燥且含水量降低时,碳氮可利用性和微生物活性较低,营养物质周转受到限制,N2O排放处于较低水平;土壤复湿后,碳氮可利用性和微生物活性较高,并激发土壤生物化学过程,并促进N2O排放[36,71].土壤湿度增加形成的厌氧条件也促进了反硝化作用产生N2O.同样,土壤频繁地经历干旱和湿润过程,也使硝化作用和反硝化作用交替进行,从而促进了N2O的产生和氮矿化(硝态氮和铵态氮的积累)[44,72].室内培养实验结果表明,不同降水频率(8天/18天)处理与恒湿处理相比,能够显著激发土壤N2O排放,排放量平均增加15倍[72],主要原因是干湿交替增加了微生物的死亡并促进土壤释放营养物质,使得土壤中氮的矿化量增加[73]. ...
... [72],主要原因是干湿交替增加了微生物的死亡并促进土壤释放营养物质,使得土壤中氮的矿化量增加[73]. ...
Effects of drying-rewetting frequency on soil carbon and nitrogen transformations
1
2002
... 水分是荒漠生态系统不同时空尺度各种生物过程的重要驱动因子.降水作为主要的水分来源,由于其间断性和不可预知性,导致土壤水分与养分等关键资源的获得也呈不连续的脉冲状态[70].不同频率的降水脉冲导致土壤经历干湿循环,当土壤干燥且含水量降低时,碳氮可利用性和微生物活性较低,营养物质周转受到限制,N2O排放处于较低水平;土壤复湿后,碳氮可利用性和微生物活性较高,并激发土壤生物化学过程,并促进N2O排放[36,71].土壤湿度增加形成的厌氧条件也促进了反硝化作用产生N2O.同样,土壤频繁地经历干旱和湿润过程,也使硝化作用和反硝化作用交替进行,从而促进了N2O的产生和氮矿化(硝态氮和铵态氮的积累)[44,72].室内培养实验结果表明,不同降水频率(8天/18天)处理与恒湿处理相比,能够显著激发土壤N2O排放,排放量平均增加15倍[72],主要原因是干湿交替增加了微生物的死亡并促进土壤释放营养物质,使得土壤中氮的矿化量增加[73]. ...
O-producing microbial communities in soils:What matters for their functioning?
1
2
... 温度对N2O排放的影响有着广泛的微生物基础[20,74].研究表明,自养硝化和异养反硝化作用都是土壤N2O排放的主要原因,二者都是温度敏感型过程[75-80],由不同的微生物类群主导[20,81].干旱草地土壤中AOB或AOA的丰度随温度升高而发生变化[82],这主要由细菌和真菌对温度的敏感性不同所导致.温度是控制与N2O排放相关的微生物功能群体(主要是自养氨氧化菌和异养反硝化菌)的群落特征及其活动的重要因素[83].研究表明,温度可以直接或间接地影响土壤N2O排放[76].温度变化不仅影响微生物的相对丰度和总生物量,也影响其生理过程,即使在微生物群落结构和组成没有发生变化的情况下,与产生和消耗N2O相关基因的表达也可能发生改变;同时温度变化也会对反硝化酶动力学有直接的影响[84].研究发现,随温度升高,N2O排放呈现不同的变化趋势[77-78,85].但大多数研究均表明,N2O排放与土壤温度呈正相关,适宜的温度使得微生物具有较高的新陈代谢速率,加速物质循环. ...
1
1977
... 温度对N2O排放的影响有着广泛的微生物基础[20,74].研究表明,自养硝化和异养反硝化作用都是土壤N2O排放的主要原因,二者都是温度敏感型过程[75-80],由不同的微生物类群主导[20,81].干旱草地土壤中AOB或AOA的丰度随温度升高而发生变化[82],这主要由细菌和真菌对温度的敏感性不同所导致.温度是控制与N2O排放相关的微生物功能群体(主要是自养氨氧化菌和异养反硝化菌)的群落特征及其活动的重要因素[83].研究表明,温度可以直接或间接地影响土壤N2O排放[76].温度变化不仅影响微生物的相对丰度和总生物量,也影响其生理过程,即使在微生物群落结构和组成没有发生变化的情况下,与产生和消耗N2O相关基因的表达也可能发生改变;同时温度变化也会对反硝化酶动力学有直接的影响[84].研究发现,随温度升高,N2O排放呈现不同的变化趋势[77-78,85].但大多数研究均表明,N2O排放与土壤温度呈正相关,适宜的温度使得微生物具有较高的新陈代谢速率,加速物质循环. ...
The potential for feedback effects induced by global warming on emissions of nitrous oxide by soils
1
1997
... 温度对N2O排放的影响有着广泛的微生物基础[20,74].研究表明,自养硝化和异养反硝化作用都是土壤N2O排放的主要原因,二者都是温度敏感型过程[75-80],由不同的微生物类群主导[20,81].干旱草地土壤中AOB或AOA的丰度随温度升高而发生变化[82],这主要由细菌和真菌对温度的敏感性不同所导致.温度是控制与N2O排放相关的微生物功能群体(主要是自养氨氧化菌和异养反硝化菌)的群落特征及其活动的重要因素[83].研究表明,温度可以直接或间接地影响土壤N2O排放[76].温度变化不仅影响微生物的相对丰度和总生物量,也影响其生理过程,即使在微生物群落结构和组成没有发生变化的情况下,与产生和消耗N2O相关基因的表达也可能发生改变;同时温度变化也会对反硝化酶动力学有直接的影响[84].研究发现,随温度升高,N2O排放呈现不同的变化趋势[77-78,85].但大多数研究均表明,N2O排放与土壤温度呈正相关,适宜的温度使得微生物具有较高的新陈代谢速率,加速物质循环. ...
Immediate and adaptational temperature effects on nitric oxide production and nitrous oxide release from nitrification and denitrification in two soils
1
1999
... 温度对N2O排放的影响有着广泛的微生物基础[20,74].研究表明,自养硝化和异养反硝化作用都是土壤N2O排放的主要原因,二者都是温度敏感型过程[75-80],由不同的微生物类群主导[20,81].干旱草地土壤中AOB或AOA的丰度随温度升高而发生变化[82],这主要由细菌和真菌对温度的敏感性不同所导致.温度是控制与N2O排放相关的微生物功能群体(主要是自养氨氧化菌和异养反硝化菌)的群落特征及其活动的重要因素[83].研究表明,温度可以直接或间接地影响土壤N2O排放[76].温度变化不仅影响微生物的相对丰度和总生物量,也影响其生理过程,即使在微生物群落结构和组成没有发生变化的情况下,与产生和消耗N2O相关基因的表达也可能发生改变;同时温度变化也会对反硝化酶动力学有直接的影响[84].研究发现,随温度升高,N2O排放呈现不同的变化趋势[77-78,85].但大多数研究均表明,N2O排放与土壤温度呈正相关,适宜的温度使得微生物具有较高的新陈代谢速率,加速物质循环. ...
N2O emission rates in a California meadow soil are influenced by fertilizer level,soil moisture and the community structure of ammonia-oxidizing bacteria
3
2009
... 温度对N2O排放的影响有着广泛的微生物基础[20,74].研究表明,自养硝化和异养反硝化作用都是土壤N2O排放的主要原因,二者都是温度敏感型过程[75-80],由不同的微生物类群主导[20,81].干旱草地土壤中AOB或AOA的丰度随温度升高而发生变化[82],这主要由细菌和真菌对温度的敏感性不同所导致.温度是控制与N2O排放相关的微生物功能群体(主要是自养氨氧化菌和异养反硝化菌)的群落特征及其活动的重要因素[83].研究表明,温度可以直接或间接地影响土壤N2O排放[76].温度变化不仅影响微生物的相对丰度和总生物量,也影响其生理过程,即使在微生物群落结构和组成没有发生变化的情况下,与产生和消耗N2O相关基因的表达也可能发生改变;同时温度变化也会对反硝化酶动力学有直接的影响[84].研究发现,随温度升高,N2O排放呈现不同的变化趋势[77-78,85].但大多数研究均表明,N2O排放与土壤温度呈正相关,适宜的温度使得微生物具有较高的新陈代谢速率,加速物质循环. ...
... 温度和土壤湿度变化会影响微生物群落特征(群落结构和丰度,以及与N2O排放相关的功能基因)从而影响N2O排放,而在不同土壤中N2O排放的响应模式不同[78-80,82,94-97].例如,已被证明温度变化会改变草原[78,97]、草甸[94]、森林[96]以及农田土壤[95]中的AOA和AOB的群落结构,相关研究表明无论是在amoA基因丰度还是微生物群落结构方面,AOA都与研究地点的土壤N2O排放呈正相关[83];同时,在实验处理条件(不同土壤湿度)下,AOA在土壤N2O排放中发挥了重要作用[81,98].各功能基因参与荒漠土壤N2O形成过程如图1所示. ...
... [78,97]、草甸[94]、森林[96]以及农田土壤[95]中的AOA和AOB的群落结构,相关研究表明无论是在amoA基因丰度还是微生物群落结构方面,AOA都与研究地点的土壤N2O排放呈正相关[83];同时,在实验处理条件(不同土壤湿度)下,AOA在土壤N2O排放中发挥了重要作用[81,98].各功能基因参与荒漠土壤N2O形成过程如图1所示. ...
Influence of temperature on the composition and activity of denitrifying soil communities
0
2010
Long-term organic and inorganic fertilization alters temperature sensitivity of potential N2O emissions and associated microbes
2
2016
... 温度对N2O排放的影响有着广泛的微生物基础[20,74].研究表明,自养硝化和异养反硝化作用都是土壤N2O排放的主要原因,二者都是温度敏感型过程[75-80],由不同的微生物类群主导[20,81].干旱草地土壤中AOB或AOA的丰度随温度升高而发生变化[82],这主要由细菌和真菌对温度的敏感性不同所导致.温度是控制与N2O排放相关的微生物功能群体(主要是自养氨氧化菌和异养反硝化菌)的群落特征及其活动的重要因素[83].研究表明,温度可以直接或间接地影响土壤N2O排放[76].温度变化不仅影响微生物的相对丰度和总生物量,也影响其生理过程,即使在微生物群落结构和组成没有发生变化的情况下,与产生和消耗N2O相关基因的表达也可能发生改变;同时温度变化也会对反硝化酶动力学有直接的影响[84].研究发现,随温度升高,N2O排放呈现不同的变化趋势[77-78,85].但大多数研究均表明,N2O排放与土壤温度呈正相关,适宜的温度使得微生物具有较高的新陈代谢速率,加速物质循环. ...
... 温度和土壤湿度变化会影响微生物群落特征(群落结构和丰度,以及与N2O排放相关的功能基因)从而影响N2O排放,而在不同土壤中N2O排放的响应模式不同[78-80,82,94-97].例如,已被证明温度变化会改变草原[78,97]、草甸[94]、森林[96]以及农田土壤[95]中的AOA和AOB的群落结构,相关研究表明无论是在amoA基因丰度还是微生物群落结构方面,AOA都与研究地点的土壤N2O排放呈正相关[83];同时,在实验处理条件(不同土壤湿度)下,AOA在土壤N2O排放中发挥了重要作用[81,98].各功能基因参与荒漠土壤N2O形成过程如图1所示. ...
Microbial regulation of terrestrial nitrous oxide formation:understanding the biological pathways for prediction of emission rates
2
2015
... 温度对N2O排放的影响有着广泛的微生物基础[20,74].研究表明,自养硝化和异养反硝化作用都是土壤N2O排放的主要原因,二者都是温度敏感型过程[75-80],由不同的微生物类群主导[20,81].干旱草地土壤中AOB或AOA的丰度随温度升高而发生变化[82],这主要由细菌和真菌对温度的敏感性不同所导致.温度是控制与N2O排放相关的微生物功能群体(主要是自养氨氧化菌和异养反硝化菌)的群落特征及其活动的重要因素[83].研究表明,温度可以直接或间接地影响土壤N2O排放[76].温度变化不仅影响微生物的相对丰度和总生物量,也影响其生理过程,即使在微生物群落结构和组成没有发生变化的情况下,与产生和消耗N2O相关基因的表达也可能发生改变;同时温度变化也会对反硝化酶动力学有直接的影响[84].研究发现,随温度升高,N2O排放呈现不同的变化趋势[77-78,85].但大多数研究均表明,N2O排放与土壤温度呈正相关,适宜的温度使得微生物具有较高的新陈代谢速率,加速物质循环. ...
... 温度和土壤湿度变化会影响微生物群落特征(群落结构和丰度,以及与N2O排放相关的功能基因)从而影响N2O排放,而在不同土壤中N2O排放的响应模式不同[78-80,82,94-97].例如,已被证明温度变化会改变草原[78,97]、草甸[94]、森林[96]以及农田土壤[95]中的AOA和AOB的群落结构,相关研究表明无论是在amoA基因丰度还是微生物群落结构方面,AOA都与研究地点的土壤N2O排放呈正相关[83];同时,在实验处理条件(不同土壤湿度)下,AOA在土壤N2O排放中发挥了重要作用[81,98].各功能基因参与荒漠土壤N2O形成过程如图1所示. ...
Ammonia-oxidizing bacteria respond to multifactorial global change
2
2004
... 温度对N2O排放的影响有着广泛的微生物基础[20,74].研究表明,自养硝化和异养反硝化作用都是土壤N2O排放的主要原因,二者都是温度敏感型过程[75-80],由不同的微生物类群主导[20,81].干旱草地土壤中AOB或AOA的丰度随温度升高而发生变化[82],这主要由细菌和真菌对温度的敏感性不同所导致.温度是控制与N2O排放相关的微生物功能群体(主要是自养氨氧化菌和异养反硝化菌)的群落特征及其活动的重要因素[83].研究表明,温度可以直接或间接地影响土壤N2O排放[76].温度变化不仅影响微生物的相对丰度和总生物量,也影响其生理过程,即使在微生物群落结构和组成没有发生变化的情况下,与产生和消耗N2O相关基因的表达也可能发生改变;同时温度变化也会对反硝化酶动力学有直接的影响[84].研究发现,随温度升高,N2O排放呈现不同的变化趋势[77-78,85].但大多数研究均表明,N2O排放与土壤温度呈正相关,适宜的温度使得微生物具有较高的新陈代谢速率,加速物质循环. ...
... 温度和土壤湿度变化会影响微生物群落特征(群落结构和丰度,以及与N2O排放相关的功能基因)从而影响N2O排放,而在不同土壤中N2O排放的响应模式不同[78-80,82,94-97].例如,已被证明温度变化会改变草原[78,97]、草甸[94]、森林[96]以及农田土壤[95]中的AOA和AOB的群落结构,相关研究表明无论是在amoA基因丰度还是微生物群落结构方面,AOA都与研究地点的土壤N2O排放呈正相关[83];同时,在实验处理条件(不同土壤湿度)下,AOA在土壤N2O排放中发挥了重要作用[81,98].各功能基因参与荒漠土壤N2O形成过程如图1所示. ...
The response patterns of community traits of N2O emission-related functional guilds to temperature across different arable soils under inorganic fertilization
2
2017
... 温度对N2O排放的影响有着广泛的微生物基础[20,74].研究表明,自养硝化和异养反硝化作用都是土壤N2O排放的主要原因,二者都是温度敏感型过程[75-80],由不同的微生物类群主导[20,81].干旱草地土壤中AOB或AOA的丰度随温度升高而发生变化[82],这主要由细菌和真菌对温度的敏感性不同所导致.温度是控制与N2O排放相关的微生物功能群体(主要是自养氨氧化菌和异养反硝化菌)的群落特征及其活动的重要因素[83].研究表明,温度可以直接或间接地影响土壤N2O排放[76].温度变化不仅影响微生物的相对丰度和总生物量,也影响其生理过程,即使在微生物群落结构和组成没有发生变化的情况下,与产生和消耗N2O相关基因的表达也可能发生改变;同时温度变化也会对反硝化酶动力学有直接的影响[84].研究发现,随温度升高,N2O排放呈现不同的变化趋势[77-78,85].但大多数研究均表明,N2O排放与土壤温度呈正相关,适宜的温度使得微生物具有较高的新陈代谢速率,加速物质循环. ...
... 温度和土壤湿度变化会影响微生物群落特征(群落结构和丰度,以及与N2O排放相关的功能基因)从而影响N2O排放,而在不同土壤中N2O排放的响应模式不同[78-80,82,94-97].例如,已被证明温度变化会改变草原[78,97]、草甸[94]、森林[96]以及农田土壤[95]中的AOA和AOB的群落结构,相关研究表明无论是在amoA基因丰度还是微生物群落结构方面,AOA都与研究地点的土壤N2O排放呈正相关[83];同时,在实验处理条件(不同土壤湿度)下,AOA在土壤N2O排放中发挥了重要作用[81,98].各功能基因参与荒漠土壤N2O形成过程如图1所示. ...
Warming-induced enhancement of soil N2O efflux linked to distinct response times of genes driving N2O production and consumption
2
2014
... 温度对N2O排放的影响有着广泛的微生物基础[20,74].研究表明,自养硝化和异养反硝化作用都是土壤N2O排放的主要原因,二者都是温度敏感型过程[75-80],由不同的微生物类群主导[20,81].干旱草地土壤中AOB或AOA的丰度随温度升高而发生变化[82],这主要由细菌和真菌对温度的敏感性不同所导致.温度是控制与N2O排放相关的微生物功能群体(主要是自养氨氧化菌和异养反硝化菌)的群落特征及其活动的重要因素[83].研究表明,温度可以直接或间接地影响土壤N2O排放[76].温度变化不仅影响微生物的相对丰度和总生物量,也影响其生理过程,即使在微生物群落结构和组成没有发生变化的情况下,与产生和消耗N2O相关基因的表达也可能发生改变;同时温度变化也会对反硝化酶动力学有直接的影响[84].研究发现,随温度升高,N2O排放呈现不同的变化趋势[77-78,85].但大多数研究均表明,N2O排放与土壤温度呈正相关,适宜的温度使得微生物具有较高的新陈代谢速率,加速物质循环. ...
... 许多与氮循环相关的功能基因,包括amoA (AOA和AOB)、nirS、nirK、norB和nosZ,是指示土壤N2O排放的潜在有效指标[69,84,90-91].其中,amoA (AOA)和amoB (AOB)基因编码蛋白为氨单加氧酶,主要参与土壤中的氨氧化过程[91];nirS和nirK基因编码的蛋白为硝酸还原酶,其参与了NO向NO的转换,是反硝化作用(以及N2O生成)中的重要步骤[92];norB基因编码NO还原酶,该酶参与生成N2O;nosZ基因编码N2O还原酶,其丰度是指示N2O转化为N2的有效指标.因此,在厌氧条件下N2O排放量减少与nosZ基因表达增加有关[93]. ...
The effects of temperature,water-filled pore space and land use on N2O emissions from an imperfectly drained gleysol
1
2001
... 温度对N2O排放的影响有着广泛的微生物基础[20,74].研究表明,自养硝化和异养反硝化作用都是土壤N2O排放的主要原因,二者都是温度敏感型过程[75-80],由不同的微生物类群主导[20,81].干旱草地土壤中AOB或AOA的丰度随温度升高而发生变化[82],这主要由细菌和真菌对温度的敏感性不同所导致.温度是控制与N2O排放相关的微生物功能群体(主要是自养氨氧化菌和异养反硝化菌)的群落特征及其活动的重要因素[83].研究表明,温度可以直接或间接地影响土壤N2O排放[76].温度变化不仅影响微生物的相对丰度和总生物量,也影响其生理过程,即使在微生物群落结构和组成没有发生变化的情况下,与产生和消耗N2O相关基因的表达也可能发生改变;同时温度变化也会对反硝化酶动力学有直接的影响[84].研究发现,随温度升高,N2O排放呈现不同的变化趋势[77-78,85].但大多数研究均表明,N2O排放与土壤温度呈正相关,适宜的温度使得微生物具有较高的新陈代谢速率,加速物质循环. ...
反硝化细菌研究进展
2
2010
... 微生物作为生态系统的分解者,在氮循环过程中发挥着核心作用.AOA和AOB是硝化作用的主要驱动者,二者也有着不同的生态位分化,AOA在氮含量低、酸性土壤条件下发挥主要作用,而AOB则相反[86].参与反硝化的微生物广泛分布于古细菌、细菌和真菌中,而在酵母菌和放线菌中也有可参与反硝化作用的微生物类群[86].同时,也发现部分好氧反硝化细菌,如副球菌属(Paracocccus sp)和芽孢杆菌属(Bacillus sp)等[87]. ...
... [86].同时,也发现部分好氧反硝化细菌,如副球菌属(Paracocccus sp)和芽孢杆菌属(Bacillus sp)等[87]. ...
好氧反硝化菌及其在生物处理与修复中的应用研究进展
1
2011
... 微生物作为生态系统的分解者,在氮循环过程中发挥着核心作用.AOA和AOB是硝化作用的主要驱动者,二者也有着不同的生态位分化,AOA在氮含量低、酸性土壤条件下发挥主要作用,而AOB则相反[86].参与反硝化的微生物广泛分布于古细菌、细菌和真菌中,而在酵母菌和放线菌中也有可参与反硝化作用的微生物类群[86].同时,也发现部分好氧反硝化细菌,如副球菌属(Paracocccus sp)和芽孢杆菌属(Bacillus sp)等[87]. ...
苜蓿种植年限对土壤硝化潜势和氨氧化微生物丰度的影响
1
2020
... 有研究关注微生物生物量碳/氮与土壤N2O排放的关系.结果表明:仅微生物生物量碳与土壤N2O排放具有显著正相关关系[88],但也有研究表明仅微生物生物量氮与土壤N2O排放具有显著正相关关系[89],这主要与营养物质含量是否有利于微生物代谢,使其更高效地参与物质循环有关. ...
短期放牧对半干旱草地生态系统CO2和N2O排放的影响
1
2018
... 有研究关注微生物生物量碳/氮与土壤N2O排放的关系.结果表明:仅微生物生物量碳与土壤N2O排放具有显著正相关关系[88],但也有研究表明仅微生物生物量氮与土壤N2O排放具有显著正相关关系[89],这主要与营养物质含量是否有利于微生物代谢,使其更高效地参与物质循环有关. ...
Environmental variables better explain changes in potential nitrification and denitrification activities than microbial properties in fertilized forest soils
1
2019
... 许多与氮循环相关的功能基因,包括amoA (AOA和AOB)、nirS、nirK、norB和nosZ,是指示土壤N2O排放的潜在有效指标[69,84,90-91].其中,amoA (AOA)和amoB (AOB)基因编码蛋白为氨单加氧酶,主要参与土壤中的氨氧化过程[91];nirS和nirK基因编码的蛋白为硝酸还原酶,其参与了NO向NO的转换,是反硝化作用(以及N2O生成)中的重要步骤[92];norB基因编码NO还原酶,该酶参与生成N2O;nosZ基因编码N2O还原酶,其丰度是指示N2O转化为N2的有效指标.因此,在厌氧条件下N2O排放量减少与nosZ基因表达增加有关[93]. ...
Microbial abundances predict methane and nitrous oxide fluxes from a windrow composting system
2
2017
... 许多与氮循环相关的功能基因,包括amoA (AOA和AOB)、nirS、nirK、norB和nosZ,是指示土壤N2O排放的潜在有效指标[69,84,90-91].其中,amoA (AOA)和amoB (AOB)基因编码蛋白为氨单加氧酶,主要参与土壤中的氨氧化过程[91];nirS和nirK基因编码的蛋白为硝酸还原酶,其参与了NO向NO的转换,是反硝化作用(以及N2O生成)中的重要步骤[92];norB基因编码NO还原酶,该酶参与生成N2O;nosZ基因编码N2O还原酶,其丰度是指示N2O转化为N2的有效指标.因此,在厌氧条件下N2O排放量减少与nosZ基因表达增加有关[93]. ...
... [91];nirS和nirK基因编码的蛋白为硝酸还原酶,其参与了NO向NO的转换,是反硝化作用(以及N2O生成)中的重要步骤[92];norB基因编码NO还原酶,该酶参与生成N2O;nosZ基因编码N2O还原酶,其丰度是指示N2O转化为N2的有效指标.因此,在厌氧条件下N2O排放量减少与nosZ基因表达增加有关[93]. ...
Cell biology and molecular basis of denitrification
1
1997
... 许多与氮循环相关的功能基因,包括amoA (AOA和AOB)、nirS、nirK、norB和nosZ,是指示土壤N2O排放的潜在有效指标[69,84,90-91].其中,amoA (AOA)和amoB (AOB)基因编码蛋白为氨单加氧酶,主要参与土壤中的氨氧化过程[91];nirS和nirK基因编码的蛋白为硝酸还原酶,其参与了NO向NO的转换,是反硝化作用(以及N2O生成)中的重要步骤[92];norB基因编码NO还原酶,该酶参与生成N2O;nosZ基因编码N2O还原酶,其丰度是指示N2O转化为N2的有效指标.因此,在厌氧条件下N2O排放量减少与nosZ基因表达增加有关[93]. ...
Mixed grazing and clipping is beneficial to ecosystem recovery but may increase potential N2O emissions in a semi-arid grassland
1
2017
... 许多与氮循环相关的功能基因,包括amoA (AOA和AOB)、nirS、nirK、norB和nosZ,是指示土壤N2O排放的潜在有效指标[69,84,90-91].其中,amoA (AOA)和amoB (AOB)基因编码蛋白为氨单加氧酶,主要参与土壤中的氨氧化过程[91];nirS和nirK基因编码的蛋白为硝酸还原酶,其参与了NO向NO的转换,是反硝化作用(以及N2O生成)中的重要步骤[92];norB基因编码NO还原酶,该酶参与生成N2O;nosZ基因编码N2O还原酶,其丰度是指示N2O转化为N2的有效指标.因此,在厌氧条件下N2O排放量减少与nosZ基因表达增加有关[93]. ...
Effects of temperature and fertilizer on activity and community structure of soil ammonia oxidizers
2
2003
... 温度和土壤湿度变化会影响微生物群落特征(群落结构和丰度,以及与N2O排放相关的功能基因)从而影响N2O排放,而在不同土壤中N2O排放的响应模式不同[78-80,82,94-97].例如,已被证明温度变化会改变草原[78,97]、草甸[94]、森林[96]以及农田土壤[95]中的AOA和AOB的群落结构,相关研究表明无论是在amoA基因丰度还是微生物群落结构方面,AOA都与研究地点的土壤N2O排放呈正相关[83];同时,在实验处理条件(不同土壤湿度)下,AOA在土壤N2O排放中发挥了重要作用[81,98].各功能基因参与荒漠土壤N2O形成过程如图1所示. ...
... [94]、森林[96]以及农田土壤[95]中的AOA和AOB的群落结构,相关研究表明无论是在amoA基因丰度还是微生物群落结构方面,AOA都与研究地点的土壤N2O排放呈正相关[83];同时,在实验处理条件(不同土壤湿度)下,AOA在土壤N2O排放中发挥了重要作用[81,98].各功能基因参与荒漠土壤N2O形成过程如图1所示. ...
Growth,activity and temperature responses of ammonia-oxidizing archaea and bacteria in soil microcosms
1
2008
... 温度和土壤湿度变化会影响微生物群落特征(群落结构和丰度,以及与N2O排放相关的功能基因)从而影响N2O排放,而在不同土壤中N2O排放的响应模式不同[78-80,82,94-97].例如,已被证明温度变化会改变草原[78,97]、草甸[94]、森林[96]以及农田土壤[95]中的AOA和AOB的群落结构,相关研究表明无论是在amoA基因丰度还是微生物群落结构方面,AOA都与研究地点的土壤N2O排放呈正相关[83];同时,在实验处理条件(不同土壤湿度)下,AOA在土壤N2O排放中发挥了重要作用[81,98].各功能基因参与荒漠土壤N2O形成过程如图1所示. ...
Nitrifiers and denitrifiers respond rapidly to changed moisture and increasing temperature in a pristine forest soil
1
2010
... 温度和土壤湿度变化会影响微生物群落特征(群落结构和丰度,以及与N2O排放相关的功能基因)从而影响N2O排放,而在不同土壤中N2O排放的响应模式不同[78-80,82,94-97].例如,已被证明温度变化会改变草原[78,97]、草甸[94]、森林[96]以及农田土壤[95]中的AOA和AOB的群落结构,相关研究表明无论是在amoA基因丰度还是微生物群落结构方面,AOA都与研究地点的土壤N2O排放呈正相关[83];同时,在实验处理条件(不同土壤湿度)下,AOA在土壤N2O排放中发挥了重要作用[81,98].各功能基因参与荒漠土壤N2O形成过程如图1所示. ...
Effects of climate warming and elevated CO2 on autotrophic nitrification and nitrifiers in dryland ecosystems
2
2016
... 温度和土壤湿度变化会影响微生物群落特征(群落结构和丰度,以及与N2O排放相关的功能基因)从而影响N2O排放,而在不同土壤中N2O排放的响应模式不同[78-80,82,94-97].例如,已被证明温度变化会改变草原[78,97]、草甸[94]、森林[96]以及农田土壤[95]中的AOA和AOB的群落结构,相关研究表明无论是在amoA基因丰度还是微生物群落结构方面,AOA都与研究地点的土壤N2O排放呈正相关[83];同时,在实验处理条件(不同土壤湿度)下,AOA在土壤N2O排放中发挥了重要作用[81,98].各功能基因参与荒漠土壤N2O形成过程如图1所示. ...
... ,97]、草甸[94]、森林[96]以及农田土壤[95]中的AOA和AOB的群落结构,相关研究表明无论是在amoA基因丰度还是微生物群落结构方面,AOA都与研究地点的土壤N2O排放呈正相关[83];同时,在实验处理条件(不同土壤湿度)下,AOA在土壤N2O排放中发挥了重要作用[81,98].各功能基因参与荒漠土壤N2O形成过程如图1所示. ...
Isotopic signatures of N2O produced by ammonia-oxidizing archaea from soils
1
2014
... 温度和土壤湿度变化会影响微生物群落特征(群落结构和丰度,以及与N2O排放相关的功能基因)从而影响N2O排放,而在不同土壤中N2O排放的响应模式不同[78-80,82,94-97].例如,已被证明温度变化会改变草原[78,97]、草甸[94]、森林[96]以及农田土壤[95]中的AOA和AOB的群落结构,相关研究表明无论是在amoA基因丰度还是微生物群落结构方面,AOA都与研究地点的土壤N2O排放呈正相关[83];同时,在实验处理条件(不同土壤湿度)下,AOA在土壤N2O排放中发挥了重要作用[81,98].各功能基因参与荒漠土壤N2O形成过程如图1所示. ...
A multi-criteria based review of models that predict environmental impacts of land use-change for perennial energy crops on water,carbon and nitrogen cycling
1
2013
... 利用模型模拟N2O排放能较全面地掌握和预测其排放量,但在应用前需进行实验验证.目前,有关模拟N2O排放的模型多应用于农田生态系统,相关模型有DNDC(Denitrification-decomposition model)模型、Ecosys(Ecosystem model)模型和WNMM(Water and nitrogen management model)模型[99];其中,DNDC模型应用最为广泛,其需要的参数类型主要包括气象数据、土壤性质、植被类型和农田管理措施4个方面[100],由于荒漠土壤植被稀疏也无土壤管理措施,单独利用DNDC模型不适用于模拟荒漠土壤N2O排放;相对而言WNMM模型主要用于干旱半干旱气候条件下农田土壤的N2O排放模拟,更适合于模拟荒漠土壤N2O排放,该模型需参数类型也主要包括气象数据、土壤性质、植被类型等,但着重关注水肥管理的影响[101].由于荒漠地区植被类型单一并无人工水肥管理等,因此仅采用WNMM模型具有较多不确定因素.目前已有研究采用DNDC模型模拟荒漠区温室气体排放,发现该模型误差平均在20%以内[102],因此在模型选用时可采取多模型结合(如DNDC模型+WNMM模型)和实验数据进行验证,以提高模型模拟的准确性. ...
Modeling trace gas emissions from agricultural ecosystems
1
2000
... 利用模型模拟N2O排放能较全面地掌握和预测其排放量,但在应用前需进行实验验证.目前,有关模拟N2O排放的模型多应用于农田生态系统,相关模型有DNDC(Denitrification-decomposition model)模型、Ecosys(Ecosystem model)模型和WNMM(Water and nitrogen management model)模型[99];其中,DNDC模型应用最为广泛,其需要的参数类型主要包括气象数据、土壤性质、植被类型和农田管理措施4个方面[100],由于荒漠土壤植被稀疏也无土壤管理措施,单独利用DNDC模型不适用于模拟荒漠土壤N2O排放;相对而言WNMM模型主要用于干旱半干旱气候条件下农田土壤的N2O排放模拟,更适合于模拟荒漠土壤N2O排放,该模型需参数类型也主要包括气象数据、土壤性质、植被类型等,但着重关注水肥管理的影响[101].由于荒漠地区植被类型单一并无人工水肥管理等,因此仅采用WNMM模型具有较多不确定因素.目前已有研究采用DNDC模型模拟荒漠区温室气体排放,发现该模型误差平均在20%以内[102],因此在模型选用时可采取多模型结合(如DNDC模型+WNMM模型)和实验数据进行验证,以提高模型模拟的准确性. ...
A spatially referenced water and nitrogen management model (WNMM) for (irrigated) intensive cropping systems in the North China Plain
1
2007
... 利用模型模拟N2O排放能较全面地掌握和预测其排放量,但在应用前需进行实验验证.目前,有关模拟N2O排放的模型多应用于农田生态系统,相关模型有DNDC(Denitrification-decomposition model)模型、Ecosys(Ecosystem model)模型和WNMM(Water and nitrogen management model)模型[99];其中,DNDC模型应用最为广泛,其需要的参数类型主要包括气象数据、土壤性质、植被类型和农田管理措施4个方面[100],由于荒漠土壤植被稀疏也无土壤管理措施,单独利用DNDC模型不适用于模拟荒漠土壤N2O排放;相对而言WNMM模型主要用于干旱半干旱气候条件下农田土壤的N2O排放模拟,更适合于模拟荒漠土壤N2O排放,该模型需参数类型也主要包括气象数据、土壤性质、植被类型等,但着重关注水肥管理的影响[101].由于荒漠地区植被类型单一并无人工水肥管理等,因此仅采用WNMM模型具有较多不确定因素.目前已有研究采用DNDC模型模拟荒漠区温室气体排放,发现该模型误差平均在20%以内[102],因此在模型选用时可采取多模型结合(如DNDC模型+WNMM模型)和实验数据进行验证,以提高模型模拟的准确性. ...
基于DNDC模型的干旱区水稻田温室气体排放实验研究:以乌鲁木齐市米东区为例
1
2021
... 利用模型模拟N2O排放能较全面地掌握和预测其排放量,但在应用前需进行实验验证.目前,有关模拟N2O排放的模型多应用于农田生态系统,相关模型有DNDC(Denitrification-decomposition model)模型、Ecosys(Ecosystem model)模型和WNMM(Water and nitrogen management model)模型[99];其中,DNDC模型应用最为广泛,其需要的参数类型主要包括气象数据、土壤性质、植被类型和农田管理措施4个方面[100],由于荒漠土壤植被稀疏也无土壤管理措施,单独利用DNDC模型不适用于模拟荒漠土壤N2O排放;相对而言WNMM模型主要用于干旱半干旱气候条件下农田土壤的N2O排放模拟,更适合于模拟荒漠土壤N2O排放,该模型需参数类型也主要包括气象数据、土壤性质、植被类型等,但着重关注水肥管理的影响[101].由于荒漠地区植被类型单一并无人工水肥管理等,因此仅采用WNMM模型具有较多不确定因素.目前已有研究采用DNDC模型模拟荒漠区温室气体排放,发现该模型误差平均在20%以内[102],因此在模型选用时可采取多模型结合(如DNDC模型+WNMM模型)和实验数据进行验证,以提高模型模拟的准确性. ...




 甘公网安备 62010202000688号
甘公网安备 62010202000688号
{kind=link}
{kind=link}