中国荒漠主要植物群落调查的意义、现状及方案
1
2023
... 荒漠生态系统是全球极端的陆地生境,占据地球陆地表面积的约三分之一.该生态系统具有强辐射、高温、极端干旱及土壤贫瘠等特点,对植物的生长、分布及生存策略构成严峻挑战[1-2].在长期进化过程中,荒漠植物通过一系列的生理生态适应机制,在严酷环境下维持生存.这些机制涉及形态结构的优化、水力传输的调控、光合系统的调节、抗氧化系统的防御及基因表达的调控,展现出较强的环境适应能力[3-4].近年来,全球气候变化加剧了荒漠化进程,使荒漠植物面临更大的生存压力.因此,深入研究荒漠植物的生理生态适应机制,对于理解植物如何在极端环境中存活,并为生态系统保护与荒漠化治理提供理论支持具有重要意义.本文梳理了近年来荒漠植物在形态适应、水分调控、渗透调节、光合生理、分子机制等方面的研究进展,分析其对环境胁迫的响应策略,并展望未来的研究方向,以期为荒漠生态系统的可持续管理与生态修复提供科学支撑. ...
中国干旱沙区的生态重建与恢复:沙坡头站60年重要研究进展综述
1
2016
... 荒漠生态系统是全球极端的陆地生境,占据地球陆地表面积的约三分之一.该生态系统具有强辐射、高温、极端干旱及土壤贫瘠等特点,对植物的生长、分布及生存策略构成严峻挑战[1-2].在长期进化过程中,荒漠植物通过一系列的生理生态适应机制,在严酷环境下维持生存.这些机制涉及形态结构的优化、水力传输的调控、光合系统的调节、抗氧化系统的防御及基因表达的调控,展现出较强的环境适应能力[3-4].近年来,全球气候变化加剧了荒漠化进程,使荒漠植物面临更大的生存压力.因此,深入研究荒漠植物的生理生态适应机制,对于理解植物如何在极端环境中存活,并为生态系统保护与荒漠化治理提供理论支持具有重要意义.本文梳理了近年来荒漠植物在形态适应、水分调控、渗透调节、光合生理、分子机制等方面的研究进展,分析其对环境胁迫的响应策略,并展望未来的研究方向,以期为荒漠生态系统的可持续管理与生态修复提供科学支撑. ...
荒漠区植物干旱胁迫响应研究进展
1
2024
... 荒漠生态系统是全球极端的陆地生境,占据地球陆地表面积的约三分之一.该生态系统具有强辐射、高温、极端干旱及土壤贫瘠等特点,对植物的生长、分布及生存策略构成严峻挑战[1-2].在长期进化过程中,荒漠植物通过一系列的生理生态适应机制,在严酷环境下维持生存.这些机制涉及形态结构的优化、水力传输的调控、光合系统的调节、抗氧化系统的防御及基因表达的调控,展现出较强的环境适应能力[3-4].近年来,全球气候变化加剧了荒漠化进程,使荒漠植物面临更大的生存压力.因此,深入研究荒漠植物的生理生态适应机制,对于理解植物如何在极端环境中存活,并为生态系统保护与荒漠化治理提供理论支持具有重要意义.本文梳理了近年来荒漠植物在形态适应、水分调控、渗透调节、光合生理、分子机制等方面的研究进展,分析其对环境胁迫的响应策略,并展望未来的研究方向,以期为荒漠生态系统的可持续管理与生态修复提供科学支撑. ...
Dryland mechanisms could widely control ecosystem functioning in a drier and warmer world
1
2022
... 荒漠生态系统是全球极端的陆地生境,占据地球陆地表面积的约三分之一.该生态系统具有强辐射、高温、极端干旱及土壤贫瘠等特点,对植物的生长、分布及生存策略构成严峻挑战[1-2].在长期进化过程中,荒漠植物通过一系列的生理生态适应机制,在严酷环境下维持生存.这些机制涉及形态结构的优化、水力传输的调控、光合系统的调节、抗氧化系统的防御及基因表达的调控,展现出较强的环境适应能力[3-4].近年来,全球气候变化加剧了荒漠化进程,使荒漠植物面临更大的生存压力.因此,深入研究荒漠植物的生理生态适应机制,对于理解植物如何在极端环境中存活,并为生态系统保护与荒漠化治理提供理论支持具有重要意义.本文梳理了近年来荒漠植物在形态适应、水分调控、渗透调节、光合生理、分子机制等方面的研究进展,分析其对环境胁迫的响应策略,并展望未来的研究方向,以期为荒漠生态系统的可持续管理与生态修复提供科学支撑. ...
The cell wall
2
2000
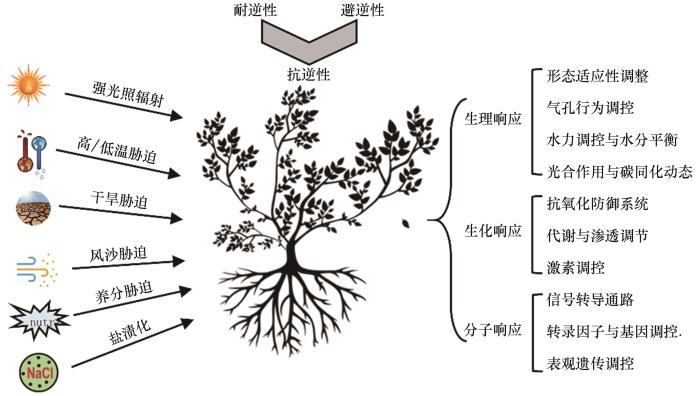
... 逆境是对植物生存生长不利的各种环境因素的总称[5].荒漠植物面临的逆境主要是非生物因子的胁迫,包括强辐射、高温、干旱、风沙侵蚀、养分贫瘠及土壤盐碱化等恶劣环境[6-7].荒漠植物在生长过程中受到这些环境因子的胁迫,生理、生长和发育会受到影响甚至损害,生存与繁殖受制.荒漠逆境类型主要包括:①辐射胁迫,强紫外线与高温促进活性氧爆发,破坏细胞结构与功能;②温度胁迫,如昼夜温差大及极端高温引发的膜脂过氧化与酶活性抑制;③水分胁迫,如长期干旱导致细胞脱水及渗透失衡;④养分限制,荒漠土壤中氮、磷等必需元素匮乏,限制植物代谢与生长;⑤盐碱胁迫,土壤中Na⁺、Cl⁻等离子过量积累干扰离子稳态(图1).植物对逆境的适应性研究是植物生理生态学的重要组成部分.荒漠植物长期暴露在这样的环境条件下,会引发一系列的生理、生化及分子响应,对逆境进行抵抗和忍耐,即植物抗逆性[5].荒漠植物通过形态调节、气孔与水力调控、代谢与渗透调节及分子机制等策略来应对这些逆境,以维持自身的存活和生长. ...
... [5].荒漠植物通过形态调节、气孔与水力调控、代谢与渗透调节及分子机制等策略来应对这些逆境,以维持自身的存活和生长. ...
Plant abiotic stress response and nutrient use efficiency
1
2020
... 逆境是对植物生存生长不利的各种环境因素的总称[5].荒漠植物面临的逆境主要是非生物因子的胁迫,包括强辐射、高温、干旱、风沙侵蚀、养分贫瘠及土壤盐碱化等恶劣环境[6-7].荒漠植物在生长过程中受到这些环境因子的胁迫,生理、生长和发育会受到影响甚至损害,生存与繁殖受制.荒漠逆境类型主要包括:①辐射胁迫,强紫外线与高温促进活性氧爆发,破坏细胞结构与功能;②温度胁迫,如昼夜温差大及极端高温引发的膜脂过氧化与酶活性抑制;③水分胁迫,如长期干旱导致细胞脱水及渗透失衡;④养分限制,荒漠土壤中氮、磷等必需元素匮乏,限制植物代谢与生长;⑤盐碱胁迫,土壤中Na⁺、Cl⁻等离子过量积累干扰离子稳态(图1).植物对逆境的适应性研究是植物生理生态学的重要组成部分.荒漠植物长期暴露在这样的环境条件下,会引发一系列的生理、生化及分子响应,对逆境进行抵抗和忍耐,即植物抗逆性[5].荒漠植物通过形态调节、气孔与水力调控、代谢与渗透调节及分子机制等策略来应对这些逆境,以维持自身的存活和生长. ...
荒漠灌木逆境适应性研究进展
1
2010
... 逆境是对植物生存生长不利的各种环境因素的总称[5].荒漠植物面临的逆境主要是非生物因子的胁迫,包括强辐射、高温、干旱、风沙侵蚀、养分贫瘠及土壤盐碱化等恶劣环境[6-7].荒漠植物在生长过程中受到这些环境因子的胁迫,生理、生长和发育会受到影响甚至损害,生存与繁殖受制.荒漠逆境类型主要包括:①辐射胁迫,强紫外线与高温促进活性氧爆发,破坏细胞结构与功能;②温度胁迫,如昼夜温差大及极端高温引发的膜脂过氧化与酶活性抑制;③水分胁迫,如长期干旱导致细胞脱水及渗透失衡;④养分限制,荒漠土壤中氮、磷等必需元素匮乏,限制植物代谢与生长;⑤盐碱胁迫,土壤中Na⁺、Cl⁻等离子过量积累干扰离子稳态(图1).植物对逆境的适应性研究是植物生理生态学的重要组成部分.荒漠植物长期暴露在这样的环境条件下,会引发一系列的生理、生化及分子响应,对逆境进行抵抗和忍耐,即植物抗逆性[5].荒漠植物通过形态调节、气孔与水力调控、代谢与渗透调节及分子机制等策略来应对这些逆境,以维持自身的存活和生长. ...
植物对荒漠生境的趋同适应
1
2006
... 荒漠生态系统中的植物群落主要由超旱生和旱生植物组成,这些植物通常具有退化或特化的叶片,或者依靠落叶(或落枝)机制适应极端环境.例如,柠条锦鸡儿(Caragana korshinskii)、油蒿(Artemisia ordosica)、沙拐枣(Calligonum mongolicum)和膜果麻黄(Ephedra przewalskii)等典型荒漠植物,在长期进化过程中形成了一系列独特的形态适应特征.植株通常矮小,叶片减少甚至完全退化,以减少水分蒸腾[8].部分植物依赖绿色小枝或茎进行光合作用.例如,梭梭(Haloxylon ammodendron)和沙拐枣的叶片高度退化,以缩小受光面积并减少水分散失,从而适应干热环境,其光合作用主要由同化枝完成[9].对中国干旱半干旱荒漠区28科74属117种自然居群的植物叶片(或同化枝)表皮微形态进行了扫描电镜研究发现,许多荒漠植物的叶或枝上具有发达的保护组织,叶(或同化枝)表皮具有大量的表皮绒毛、角质膜蜡质片层或晶体颗粒、表面瘤状或疣状突起以及相对下陷且密度较低的气孔器,荒漠植物叶片的抗逆性主要依赖于表皮毛、角质层蜡质、表皮凹凸结构、表面突起、混生的附属物以及上下表皮异化特征,通过表皮附属物与表皮结构的相互协调来抵御强光、干旱等不利环境[10].此外,某些植物,如沙拐枣在叶片或枝条表面形成角质层、蜡层或茸毛,并通过特殊的气孔结构调节水分散失,甚至表现出灰白色外观,以减少光吸收和水分蒸腾,在极端环境下,这些植物通常通过休眠或部分落叶(落枝)来减少水分消耗[11].多枝柽柳(Tamarix ramosissima)、梭梭等荒漠植物叶片或枝条呈肉质化,以增强水分储存能力[12],多枝柽柳等植物根系深度或广度远超地上部分,以确保吸收地下深层水源[13]. ...
C4荒漠植物梭梭和沙拐枣在不同水分条件下的光合作用特征
1
2006
... 荒漠生态系统中的植物群落主要由超旱生和旱生植物组成,这些植物通常具有退化或特化的叶片,或者依靠落叶(或落枝)机制适应极端环境.例如,柠条锦鸡儿(Caragana korshinskii)、油蒿(Artemisia ordosica)、沙拐枣(Calligonum mongolicum)和膜果麻黄(Ephedra przewalskii)等典型荒漠植物,在长期进化过程中形成了一系列独特的形态适应特征.植株通常矮小,叶片减少甚至完全退化,以减少水分蒸腾[8].部分植物依赖绿色小枝或茎进行光合作用.例如,梭梭(Haloxylon ammodendron)和沙拐枣的叶片高度退化,以缩小受光面积并减少水分散失,从而适应干热环境,其光合作用主要由同化枝完成[9].对中国干旱半干旱荒漠区28科74属117种自然居群的植物叶片(或同化枝)表皮微形态进行了扫描电镜研究发现,许多荒漠植物的叶或枝上具有发达的保护组织,叶(或同化枝)表皮具有大量的表皮绒毛、角质膜蜡质片层或晶体颗粒、表面瘤状或疣状突起以及相对下陷且密度较低的气孔器,荒漠植物叶片的抗逆性主要依赖于表皮毛、角质层蜡质、表皮凹凸结构、表面突起、混生的附属物以及上下表皮异化特征,通过表皮附属物与表皮结构的相互协调来抵御强光、干旱等不利环境[10].此外,某些植物,如沙拐枣在叶片或枝条表面形成角质层、蜡层或茸毛,并通过特殊的气孔结构调节水分散失,甚至表现出灰白色外观,以减少光吸收和水分蒸腾,在极端环境下,这些植物通常通过休眠或部分落叶(落枝)来减少水分消耗[11].多枝柽柳(Tamarix ramosissima)、梭梭等荒漠植物叶片或枝条呈肉质化,以增强水分储存能力[12],多枝柽柳等植物根系深度或广度远超地上部分,以确保吸收地下深层水源[13]. ...
中国干旱半干旱区荒漠植物叶片(或同化枝)表皮微形态特征
2
2016
... 荒漠生态系统中的植物群落主要由超旱生和旱生植物组成,这些植物通常具有退化或特化的叶片,或者依靠落叶(或落枝)机制适应极端环境.例如,柠条锦鸡儿(Caragana korshinskii)、油蒿(Artemisia ordosica)、沙拐枣(Calligonum mongolicum)和膜果麻黄(Ephedra przewalskii)等典型荒漠植物,在长期进化过程中形成了一系列独特的形态适应特征.植株通常矮小,叶片减少甚至完全退化,以减少水分蒸腾[8].部分植物依赖绿色小枝或茎进行光合作用.例如,梭梭(Haloxylon ammodendron)和沙拐枣的叶片高度退化,以缩小受光面积并减少水分散失,从而适应干热环境,其光合作用主要由同化枝完成[9].对中国干旱半干旱荒漠区28科74属117种自然居群的植物叶片(或同化枝)表皮微形态进行了扫描电镜研究发现,许多荒漠植物的叶或枝上具有发达的保护组织,叶(或同化枝)表皮具有大量的表皮绒毛、角质膜蜡质片层或晶体颗粒、表面瘤状或疣状突起以及相对下陷且密度较低的气孔器,荒漠植物叶片的抗逆性主要依赖于表皮毛、角质层蜡质、表皮凹凸结构、表面突起、混生的附属物以及上下表皮异化特征,通过表皮附属物与表皮结构的相互协调来抵御强光、干旱等不利环境[10].此外,某些植物,如沙拐枣在叶片或枝条表面形成角质层、蜡层或茸毛,并通过特殊的气孔结构调节水分散失,甚至表现出灰白色外观,以减少光吸收和水分蒸腾,在极端环境下,这些植物通常通过休眠或部分落叶(落枝)来减少水分消耗[11].多枝柽柳(Tamarix ramosissima)、梭梭等荒漠植物叶片或枝条呈肉质化,以增强水分储存能力[12],多枝柽柳等植物根系深度或广度远超地上部分,以确保吸收地下深层水源[13]. ...
... 荒漠植物在干旱、高温、强光等极端环境下,形成了多样化的光合碳同化机制,以提高水分利用和碳固定效率.其适应性主要体现为C₃、C₄和CAM光合途径的分化调控:C₄植物通过细胞分工和酶活性增强,提高CO₂浓缩效应,适应极端条件;C₃植物则依赖RuBisCO在温和环境中维持光合活性,尽管存在光呼吸损耗,仍能有效进行碳同化[53].然而对荒漠C3和C4植物研究也有不统一的结论.例如,对腾格里沙漠东南缘优势植物柠条锦鸡儿、油蒿、雾冰藜(Grubovia dasyphylla )和沙米(Agriophyllum squarrosum)进行UV-B辐射处理发现,辐射增强更能引起C4草本植物雾冰藜和沙米光合作用的下降,反而提高了C3灌木半灌木柠条锦鸡儿和油蒿水分利用效率[54].此外,部分荒漠植物(如景天科植物)采用CAM光合途径,通过夜间气孔开放固定CO₂、白天关闭气孔进行有机酸脱羧作用,以减少水分蒸腾损失,提高水分利用效率,这一策略使其能够在极端干旱环境下存活[55].除了光合途径的分化,气孔调控是荒漠植物光合适应的关键机制.通过降低气孔导度以减少蒸腾,大幅提升水分利用效率[53].为平衡CO₂摄取与水分保存,植物通过提高RuBisCO活性、增强叶绿素含量和优化气孔分布等策略,缓解光合作用与失水之间的矛盾[56].此外,叶片结构的可塑性也是荒漠植物适应逆境的重要方式[57].如胡杨的条形叶主要用于维持生长,光合效率低、抗逆性差,随着树体长大,条形叶逐渐被光合效率更高的卵形叶取代,但卵形叶耐光抑制能力较弱,最终被更适应强光、高温和干旱的锯齿叶取代,使胡杨能在极端环境中生存[58].同化枝的发育也是荒漠植物光合适应的重要策略,例如梭梭等植物在极端干旱环境下减少叶片生长,并依赖绿色的同化枝进行光合作用,以降低水分蒸腾损失,提高整体光合能力[10].同样,对塔克拉玛干沙漠防护林梭梭和头状沙拐枣(Calligonum caput-medusae)研究发现,干旱胁迫下这两种植物光合色素活性降低,激发能力下降,表明其以减缓同化枝生长和维持碳同化的方式抵御干旱[59]. ...
3个地理种群蒙古沙拐枣同化枝解剖结构及抗旱性比较
1
2014
... 荒漠生态系统中的植物群落主要由超旱生和旱生植物组成,这些植物通常具有退化或特化的叶片,或者依靠落叶(或落枝)机制适应极端环境.例如,柠条锦鸡儿(Caragana korshinskii)、油蒿(Artemisia ordosica)、沙拐枣(Calligonum mongolicum)和膜果麻黄(Ephedra przewalskii)等典型荒漠植物,在长期进化过程中形成了一系列独特的形态适应特征.植株通常矮小,叶片减少甚至完全退化,以减少水分蒸腾[8].部分植物依赖绿色小枝或茎进行光合作用.例如,梭梭(Haloxylon ammodendron)和沙拐枣的叶片高度退化,以缩小受光面积并减少水分散失,从而适应干热环境,其光合作用主要由同化枝完成[9].对中国干旱半干旱荒漠区28科74属117种自然居群的植物叶片(或同化枝)表皮微形态进行了扫描电镜研究发现,许多荒漠植物的叶或枝上具有发达的保护组织,叶(或同化枝)表皮具有大量的表皮绒毛、角质膜蜡质片层或晶体颗粒、表面瘤状或疣状突起以及相对下陷且密度较低的气孔器,荒漠植物叶片的抗逆性主要依赖于表皮毛、角质层蜡质、表皮凹凸结构、表面突起、混生的附属物以及上下表皮异化特征,通过表皮附属物与表皮结构的相互协调来抵御强光、干旱等不利环境[10].此外,某些植物,如沙拐枣在叶片或枝条表面形成角质层、蜡层或茸毛,并通过特殊的气孔结构调节水分散失,甚至表现出灰白色外观,以减少光吸收和水分蒸腾,在极端环境下,这些植物通常通过休眠或部分落叶(落枝)来减少水分消耗[11].多枝柽柳(Tamarix ramosissima)、梭梭等荒漠植物叶片或枝条呈肉质化,以增强水分储存能力[12],多枝柽柳等植物根系深度或广度远超地上部分,以确保吸收地下深层水源[13]. ...
多枝柽柳与梭梭光合器官形态解剖结构的生态适应性
1
2011
... 荒漠生态系统中的植物群落主要由超旱生和旱生植物组成,这些植物通常具有退化或特化的叶片,或者依靠落叶(或落枝)机制适应极端环境.例如,柠条锦鸡儿(Caragana korshinskii)、油蒿(Artemisia ordosica)、沙拐枣(Calligonum mongolicum)和膜果麻黄(Ephedra przewalskii)等典型荒漠植物,在长期进化过程中形成了一系列独特的形态适应特征.植株通常矮小,叶片减少甚至完全退化,以减少水分蒸腾[8].部分植物依赖绿色小枝或茎进行光合作用.例如,梭梭(Haloxylon ammodendron)和沙拐枣的叶片高度退化,以缩小受光面积并减少水分散失,从而适应干热环境,其光合作用主要由同化枝完成[9].对中国干旱半干旱荒漠区28科74属117种自然居群的植物叶片(或同化枝)表皮微形态进行了扫描电镜研究发现,许多荒漠植物的叶或枝上具有发达的保护组织,叶(或同化枝)表皮具有大量的表皮绒毛、角质膜蜡质片层或晶体颗粒、表面瘤状或疣状突起以及相对下陷且密度较低的气孔器,荒漠植物叶片的抗逆性主要依赖于表皮毛、角质层蜡质、表皮凹凸结构、表面突起、混生的附属物以及上下表皮异化特征,通过表皮附属物与表皮结构的相互协调来抵御强光、干旱等不利环境[10].此外,某些植物,如沙拐枣在叶片或枝条表面形成角质层、蜡层或茸毛,并通过特殊的气孔结构调节水分散失,甚至表现出灰白色外观,以减少光吸收和水分蒸腾,在极端环境下,这些植物通常通过休眠或部分落叶(落枝)来减少水分消耗[11].多枝柽柳(Tamarix ramosissima)、梭梭等荒漠植物叶片或枝条呈肉质化,以增强水分储存能力[12],多枝柽柳等植物根系深度或广度远超地上部分,以确保吸收地下深层水源[13]. ...
三种荒漠灌木根系的构形特征与叶性因子对干旱生境的适应性比较
1
2008
... 荒漠生态系统中的植物群落主要由超旱生和旱生植物组成,这些植物通常具有退化或特化的叶片,或者依靠落叶(或落枝)机制适应极端环境.例如,柠条锦鸡儿(Caragana korshinskii)、油蒿(Artemisia ordosica)、沙拐枣(Calligonum mongolicum)和膜果麻黄(Ephedra przewalskii)等典型荒漠植物,在长期进化过程中形成了一系列独特的形态适应特征.植株通常矮小,叶片减少甚至完全退化,以减少水分蒸腾[8].部分植物依赖绿色小枝或茎进行光合作用.例如,梭梭(Haloxylon ammodendron)和沙拐枣的叶片高度退化,以缩小受光面积并减少水分散失,从而适应干热环境,其光合作用主要由同化枝完成[9].对中国干旱半干旱荒漠区28科74属117种自然居群的植物叶片(或同化枝)表皮微形态进行了扫描电镜研究发现,许多荒漠植物的叶或枝上具有发达的保护组织,叶(或同化枝)表皮具有大量的表皮绒毛、角质膜蜡质片层或晶体颗粒、表面瘤状或疣状突起以及相对下陷且密度较低的气孔器,荒漠植物叶片的抗逆性主要依赖于表皮毛、角质层蜡质、表皮凹凸结构、表面突起、混生的附属物以及上下表皮异化特征,通过表皮附属物与表皮结构的相互协调来抵御强光、干旱等不利环境[10].此外,某些植物,如沙拐枣在叶片或枝条表面形成角质层、蜡层或茸毛,并通过特殊的气孔结构调节水分散失,甚至表现出灰白色外观,以减少光吸收和水分蒸腾,在极端环境下,这些植物通常通过休眠或部分落叶(落枝)来减少水分消耗[11].多枝柽柳(Tamarix ramosissima)、梭梭等荒漠植物叶片或枝条呈肉质化,以增强水分储存能力[12],多枝柽柳等植物根系深度或广度远超地上部分,以确保吸收地下深层水源[13]. ...
Predicting stomatal responses to the environment from the optimization of photosynthetic gain and hydraulic cost
1
2017
... 气孔行为是荒漠植物适应极端环境的重要生理调控机制,直接影响其水分利用效率和生存能力.在荒漠生态系统中,植物常面临水分短缺和高温胁迫,气孔的开闭调节成为控制水分蒸腾与光合作用平衡的关键[14-15].为优化水分利用效率并提高光合碳同化能力,荒漠植物进化出独特的气孔调控策略,包括降低气孔导度、调整气孔密度和优化气孔分布,从而减少蒸腾作用的同时维持一定的CO₂通量[16-17].植物在应对外界环境胁迫时,通过气孔开闭调节气体交换,从而调节植物叶片蒸腾和光合作用,不同的气孔行为反映不同的水分利用策略[18].根据气孔对水势的调节程度,通常将植物划分为等水(isohydry)和非等水(anisohydry)类群[19].等水型的植物能够在水分亏缺时关闭气孔,避免水分继续散失,保持较高水势,属于“保守型”的水分利用策略;而非等水型植物在水分亏缺时仍然保持气孔开放,保证碳获取速率,但叶水势显著下降,属于“索取型”的水分利用策略[20].此外,气孔行为不仅受昼夜节律影响,还会根据环境变化动态调整.例如,对16种荒漠植物的研究发现,一些物种在夜间由非等水势行为转变为等水势行为,反映出植物在不同蒸气压亏缺(VPD)、土壤水分和光照条件下采用不同的气孔导度调节机制,在水分恢复后能够迅速重新开放气孔并恢复光合作用,表现出较强的气孔再开能力,这种适应性对其应对剧烈的水分波动至关重要[21].在这一过程中,植物激素,尤其是乙烯及脱落酸(ABA)等发挥着关键作用.这些作为植物抗逆信号分子,在干旱胁迫下积累,触发气孔关闭机制,从而减少水分流失并维持水分平衡[22]. ...
An unusual strategy of stomatal control in the desert shrub Ammopiptanthus mongolicus
1
2018
... 气孔行为是荒漠植物适应极端环境的重要生理调控机制,直接影响其水分利用效率和生存能力.在荒漠生态系统中,植物常面临水分短缺和高温胁迫,气孔的开闭调节成为控制水分蒸腾与光合作用平衡的关键[14-15].为优化水分利用效率并提高光合碳同化能力,荒漠植物进化出独特的气孔调控策略,包括降低气孔导度、调整气孔密度和优化气孔分布,从而减少蒸腾作用的同时维持一定的CO₂通量[16-17].植物在应对外界环境胁迫时,通过气孔开闭调节气体交换,从而调节植物叶片蒸腾和光合作用,不同的气孔行为反映不同的水分利用策略[18].根据气孔对水势的调节程度,通常将植物划分为等水(isohydry)和非等水(anisohydry)类群[19].等水型的植物能够在水分亏缺时关闭气孔,避免水分继续散失,保持较高水势,属于“保守型”的水分利用策略;而非等水型植物在水分亏缺时仍然保持气孔开放,保证碳获取速率,但叶水势显著下降,属于“索取型”的水分利用策略[20].此外,气孔行为不仅受昼夜节律影响,还会根据环境变化动态调整.例如,对16种荒漠植物的研究发现,一些物种在夜间由非等水势行为转变为等水势行为,反映出植物在不同蒸气压亏缺(VPD)、土壤水分和光照条件下采用不同的气孔导度调节机制,在水分恢复后能够迅速重新开放气孔并恢复光合作用,表现出较强的气孔再开能力,这种适应性对其应对剧烈的水分波动至关重要[21].在这一过程中,植物激素,尤其是乙烯及脱落酸(ABA)等发挥着关键作用.这些作为植物抗逆信号分子,在干旱胁迫下积累,触发气孔关闭机制,从而减少水分流失并维持水分平衡[22]. ...
干旱区荒漠-绿洲过渡带3种典型灌木气孔导度对环境变化的响应及其对蒸腾的调控
1
2021
... 气孔行为是荒漠植物适应极端环境的重要生理调控机制,直接影响其水分利用效率和生存能力.在荒漠生态系统中,植物常面临水分短缺和高温胁迫,气孔的开闭调节成为控制水分蒸腾与光合作用平衡的关键[14-15].为优化水分利用效率并提高光合碳同化能力,荒漠植物进化出独特的气孔调控策略,包括降低气孔导度、调整气孔密度和优化气孔分布,从而减少蒸腾作用的同时维持一定的CO₂通量[16-17].植物在应对外界环境胁迫时,通过气孔开闭调节气体交换,从而调节植物叶片蒸腾和光合作用,不同的气孔行为反映不同的水分利用策略[18].根据气孔对水势的调节程度,通常将植物划分为等水(isohydry)和非等水(anisohydry)类群[19].等水型的植物能够在水分亏缺时关闭气孔,避免水分继续散失,保持较高水势,属于“保守型”的水分利用策略;而非等水型植物在水分亏缺时仍然保持气孔开放,保证碳获取速率,但叶水势显著下降,属于“索取型”的水分利用策略[20].此外,气孔行为不仅受昼夜节律影响,还会根据环境变化动态调整.例如,对16种荒漠植物的研究发现,一些物种在夜间由非等水势行为转变为等水势行为,反映出植物在不同蒸气压亏缺(VPD)、土壤水分和光照条件下采用不同的气孔导度调节机制,在水分恢复后能够迅速重新开放气孔并恢复光合作用,表现出较强的气孔再开能力,这种适应性对其应对剧烈的水分波动至关重要[21].在这一过程中,植物激素,尤其是乙烯及脱落酸(ABA)等发挥着关键作用.这些作为植物抗逆信号分子,在干旱胁迫下积累,触发气孔关闭机制,从而减少水分流失并维持水分平衡[22]. ...
6种荒漠植物叶片气孔性状比较分析
1
2021
... 气孔行为是荒漠植物适应极端环境的重要生理调控机制,直接影响其水分利用效率和生存能力.在荒漠生态系统中,植物常面临水分短缺和高温胁迫,气孔的开闭调节成为控制水分蒸腾与光合作用平衡的关键[14-15].为优化水分利用效率并提高光合碳同化能力,荒漠植物进化出独特的气孔调控策略,包括降低气孔导度、调整气孔密度和优化气孔分布,从而减少蒸腾作用的同时维持一定的CO₂通量[16-17].植物在应对外界环境胁迫时,通过气孔开闭调节气体交换,从而调节植物叶片蒸腾和光合作用,不同的气孔行为反映不同的水分利用策略[18].根据气孔对水势的调节程度,通常将植物划分为等水(isohydry)和非等水(anisohydry)类群[19].等水型的植物能够在水分亏缺时关闭气孔,避免水分继续散失,保持较高水势,属于“保守型”的水分利用策略;而非等水型植物在水分亏缺时仍然保持气孔开放,保证碳获取速率,但叶水势显著下降,属于“索取型”的水分利用策略[20].此外,气孔行为不仅受昼夜节律影响,还会根据环境变化动态调整.例如,对16种荒漠植物的研究发现,一些物种在夜间由非等水势行为转变为等水势行为,反映出植物在不同蒸气压亏缺(VPD)、土壤水分和光照条件下采用不同的气孔导度调节机制,在水分恢复后能够迅速重新开放气孔并恢复光合作用,表现出较强的气孔再开能力,这种适应性对其应对剧烈的水分波动至关重要[21].在这一过程中,植物激素,尤其是乙烯及脱落酸(ABA)等发挥着关键作用.这些作为植物抗逆信号分子,在干旱胁迫下积累,触发气孔关闭机制,从而减少水分流失并维持水分平衡[22]. ...
A stomatal safety-efficiency trade-off constrains responses to leaf dehydration
1
2019
... 气孔行为是荒漠植物适应极端环境的重要生理调控机制,直接影响其水分利用效率和生存能力.在荒漠生态系统中,植物常面临水分短缺和高温胁迫,气孔的开闭调节成为控制水分蒸腾与光合作用平衡的关键[14-15].为优化水分利用效率并提高光合碳同化能力,荒漠植物进化出独特的气孔调控策略,包括降低气孔导度、调整气孔密度和优化气孔分布,从而减少蒸腾作用的同时维持一定的CO₂通量[16-17].植物在应对外界环境胁迫时,通过气孔开闭调节气体交换,从而调节植物叶片蒸腾和光合作用,不同的气孔行为反映不同的水分利用策略[18].根据气孔对水势的调节程度,通常将植物划分为等水(isohydry)和非等水(anisohydry)类群[19].等水型的植物能够在水分亏缺时关闭气孔,避免水分继续散失,保持较高水势,属于“保守型”的水分利用策略;而非等水型植物在水分亏缺时仍然保持气孔开放,保证碳获取速率,但叶水势显著下降,属于“索取型”的水分利用策略[20].此外,气孔行为不仅受昼夜节律影响,还会根据环境变化动态调整.例如,对16种荒漠植物的研究发现,一些物种在夜间由非等水势行为转变为等水势行为,反映出植物在不同蒸气压亏缺(VPD)、土壤水分和光照条件下采用不同的气孔导度调节机制,在水分恢复后能够迅速重新开放气孔并恢复光合作用,表现出较强的气孔再开能力,这种适应性对其应对剧烈的水分波动至关重要[21].在这一过程中,植物激素,尤其是乙烯及脱落酸(ABA)等发挥着关键作用.这些作为植物抗逆信号分子,在干旱胁迫下积累,触发气孔关闭机制,从而减少水分流失并维持水分平衡[22]. ...
Variability among species of stomatal control under fluctuating soil water status and evaporative demand:modelling isohydric and anisohydric behaviours
1
1998
... 气孔行为是荒漠植物适应极端环境的重要生理调控机制,直接影响其水分利用效率和生存能力.在荒漠生态系统中,植物常面临水分短缺和高温胁迫,气孔的开闭调节成为控制水分蒸腾与光合作用平衡的关键[14-15].为优化水分利用效率并提高光合碳同化能力,荒漠植物进化出独特的气孔调控策略,包括降低气孔导度、调整气孔密度和优化气孔分布,从而减少蒸腾作用的同时维持一定的CO₂通量[16-17].植物在应对外界环境胁迫时,通过气孔开闭调节气体交换,从而调节植物叶片蒸腾和光合作用,不同的气孔行为反映不同的水分利用策略[18].根据气孔对水势的调节程度,通常将植物划分为等水(isohydry)和非等水(anisohydry)类群[19].等水型的植物能够在水分亏缺时关闭气孔,避免水分继续散失,保持较高水势,属于“保守型”的水分利用策略;而非等水型植物在水分亏缺时仍然保持气孔开放,保证碳获取速率,但叶水势显著下降,属于“索取型”的水分利用策略[20].此外,气孔行为不仅受昼夜节律影响,还会根据环境变化动态调整.例如,对16种荒漠植物的研究发现,一些物种在夜间由非等水势行为转变为等水势行为,反映出植物在不同蒸气压亏缺(VPD)、土壤水分和光照条件下采用不同的气孔导度调节机制,在水分恢复后能够迅速重新开放气孔并恢复光合作用,表现出较强的气孔再开能力,这种适应性对其应对剧烈的水分波动至关重要[21].在这一过程中,植物激素,尤其是乙烯及脱落酸(ABA)等发挥着关键作用.这些作为植物抗逆信号分子,在干旱胁迫下积累,触发气孔关闭机制,从而减少水分流失并维持水分平衡[22]. ...
Metrics and proxies for stringency of regulation of plant water status (iso/anisohydry):a global data set reveals coordination and trade-offs among water transport traits
1
2019
... 气孔行为是荒漠植物适应极端环境的重要生理调控机制,直接影响其水分利用效率和生存能力.在荒漠生态系统中,植物常面临水分短缺和高温胁迫,气孔的开闭调节成为控制水分蒸腾与光合作用平衡的关键[14-15].为优化水分利用效率并提高光合碳同化能力,荒漠植物进化出独特的气孔调控策略,包括降低气孔导度、调整气孔密度和优化气孔分布,从而减少蒸腾作用的同时维持一定的CO₂通量[16-17].植物在应对外界环境胁迫时,通过气孔开闭调节气体交换,从而调节植物叶片蒸腾和光合作用,不同的气孔行为反映不同的水分利用策略[18].根据气孔对水势的调节程度,通常将植物划分为等水(isohydry)和非等水(anisohydry)类群[19].等水型的植物能够在水分亏缺时关闭气孔,避免水分继续散失,保持较高水势,属于“保守型”的水分利用策略;而非等水型植物在水分亏缺时仍然保持气孔开放,保证碳获取速率,但叶水势显著下降,属于“索取型”的水分利用策略[20].此外,气孔行为不仅受昼夜节律影响,还会根据环境变化动态调整.例如,对16种荒漠植物的研究发现,一些物种在夜间由非等水势行为转变为等水势行为,反映出植物在不同蒸气压亏缺(VPD)、土壤水分和光照条件下采用不同的气孔导度调节机制,在水分恢复后能够迅速重新开放气孔并恢复光合作用,表现出较强的气孔再开能力,这种适应性对其应对剧烈的水分波动至关重要[21].在这一过程中,植物激素,尤其是乙烯及脱落酸(ABA)等发挥着关键作用.这些作为植物抗逆信号分子,在干旱胁迫下积累,触发气孔关闭机制,从而减少水分流失并维持水分平衡[22]. ...
Differential daytime and night-time stomatal behavior in plants from North American deserts
1
2012
... 气孔行为是荒漠植物适应极端环境的重要生理调控机制,直接影响其水分利用效率和生存能力.在荒漠生态系统中,植物常面临水分短缺和高温胁迫,气孔的开闭调节成为控制水分蒸腾与光合作用平衡的关键[14-15].为优化水分利用效率并提高光合碳同化能力,荒漠植物进化出独特的气孔调控策略,包括降低气孔导度、调整气孔密度和优化气孔分布,从而减少蒸腾作用的同时维持一定的CO₂通量[16-17].植物在应对外界环境胁迫时,通过气孔开闭调节气体交换,从而调节植物叶片蒸腾和光合作用,不同的气孔行为反映不同的水分利用策略[18].根据气孔对水势的调节程度,通常将植物划分为等水(isohydry)和非等水(anisohydry)类群[19].等水型的植物能够在水分亏缺时关闭气孔,避免水分继续散失,保持较高水势,属于“保守型”的水分利用策略;而非等水型植物在水分亏缺时仍然保持气孔开放,保证碳获取速率,但叶水势显著下降,属于“索取型”的水分利用策略[20].此外,气孔行为不仅受昼夜节律影响,还会根据环境变化动态调整.例如,对16种荒漠植物的研究发现,一些物种在夜间由非等水势行为转变为等水势行为,反映出植物在不同蒸气压亏缺(VPD)、土壤水分和光照条件下采用不同的气孔导度调节机制,在水分恢复后能够迅速重新开放气孔并恢复光合作用,表现出较强的气孔再开能力,这种适应性对其应对剧烈的水分波动至关重要[21].在这一过程中,植物激素,尤其是乙烯及脱落酸(ABA)等发挥着关键作用.这些作为植物抗逆信号分子,在干旱胁迫下积累,触发气孔关闭机制,从而减少水分流失并维持水分平衡[22]. ...
Ethylene,not ABA,is closely linked to the recovery of gas exchange after drought in four Caragana species
1
2021
... 气孔行为是荒漠植物适应极端环境的重要生理调控机制,直接影响其水分利用效率和生存能力.在荒漠生态系统中,植物常面临水分短缺和高温胁迫,气孔的开闭调节成为控制水分蒸腾与光合作用平衡的关键[14-15].为优化水分利用效率并提高光合碳同化能力,荒漠植物进化出独特的气孔调控策略,包括降低气孔导度、调整气孔密度和优化气孔分布,从而减少蒸腾作用的同时维持一定的CO₂通量[16-17].植物在应对外界环境胁迫时,通过气孔开闭调节气体交换,从而调节植物叶片蒸腾和光合作用,不同的气孔行为反映不同的水分利用策略[18].根据气孔对水势的调节程度,通常将植物划分为等水(isohydry)和非等水(anisohydry)类群[19].等水型的植物能够在水分亏缺时关闭气孔,避免水分继续散失,保持较高水势,属于“保守型”的水分利用策略;而非等水型植物在水分亏缺时仍然保持气孔开放,保证碳获取速率,但叶水势显著下降,属于“索取型”的水分利用策略[20].此外,气孔行为不仅受昼夜节律影响,还会根据环境变化动态调整.例如,对16种荒漠植物的研究发现,一些物种在夜间由非等水势行为转变为等水势行为,反映出植物在不同蒸气压亏缺(VPD)、土壤水分和光照条件下采用不同的气孔导度调节机制,在水分恢复后能够迅速重新开放气孔并恢复光合作用,表现出较强的气孔再开能力,这种适应性对其应对剧烈的水分波动至关重要[21].在这一过程中,植物激素,尤其是乙烯及脱落酸(ABA)等发挥着关键作用.这些作为植物抗逆信号分子,在干旱胁迫下积累,触发气孔关闭机制,从而减少水分流失并维持水分平衡[22]. ...
内蒙古高原荒漠区锦鸡儿属(Caragana)四种植物水力结构特征的比较研究
1
2010
... 荒漠植物的水力结构是其水分传导的关键基础,对抗逆性具有重要影响[23-24].其木质部由导管、管胞、穿孔板和纹孔等组成,构成高效低阻的输水系统[25-26].在干旱胁迫下,水力结构展现出可塑性:有的植物增大导管数量和直径以增强导水能力[27-28],而有的则缩小管胞和纹孔尺寸以控制水分流失[29].荒漠植物通常具备较小直径且壁厚的导管和管胞,有效降低气穴化形成的风险,提高抗栓塞能力[30].许多荒漠植物的导管连通方式较为保守,往往采用梯状或简单穿孔板[31],不仅有助于维持水分连续性,还能在高水势梯度下减少水力失败的可能性[32].荒漠植物中纤维-管胞和纤维-导管协同提升水力安全,其中管胞作为次级通道,抗旱性更强[33].许多荒漠木本植物纹孔常具加厚内缘或带状结构,减少水分流失并提高负压耐受性[34].某些荒漠树种采用带孔穿孔板,其开放面积较小,有助于防止气泡扩散,提高水分输导的稳定性[35].荒漠植物在导管和管胞的排列模式上表现出趋同进化,其较短且密集的水输导单元能够降低水分张力,提高水分的局部再分配能力,维持长期干旱下的水力连续性[36].某些荒漠裸子植物具特化纹孔膜结构,适应极端水势梯度[37].此外,水力结构随器官部位变化:如蛇麻(Humulus lupulus)的基部导水组织增大而顶端缩小[38].表层根趋向形成窄径导管,以降低干旱与冻融栓塞风险[39]. ...
内蒙古高原西部荒漠区锦鸡儿属植物水力结构的变化
1
2015
... 荒漠植物的水力结构是其水分传导的关键基础,对抗逆性具有重要影响[23-24].其木质部由导管、管胞、穿孔板和纹孔等组成,构成高效低阻的输水系统[25-26].在干旱胁迫下,水力结构展现出可塑性:有的植物增大导管数量和直径以增强导水能力[27-28],而有的则缩小管胞和纹孔尺寸以控制水分流失[29].荒漠植物通常具备较小直径且壁厚的导管和管胞,有效降低气穴化形成的风险,提高抗栓塞能力[30].许多荒漠植物的导管连通方式较为保守,往往采用梯状或简单穿孔板[31],不仅有助于维持水分连续性,还能在高水势梯度下减少水力失败的可能性[32].荒漠植物中纤维-管胞和纤维-导管协同提升水力安全,其中管胞作为次级通道,抗旱性更强[33].许多荒漠木本植物纹孔常具加厚内缘或带状结构,减少水分流失并提高负压耐受性[34].某些荒漠树种采用带孔穿孔板,其开放面积较小,有助于防止气泡扩散,提高水分输导的稳定性[35].荒漠植物在导管和管胞的排列模式上表现出趋同进化,其较短且密集的水输导单元能够降低水分张力,提高水分的局部再分配能力,维持长期干旱下的水力连续性[36].某些荒漠裸子植物具特化纹孔膜结构,适应极端水势梯度[37].此外,水力结构随器官部位变化:如蛇麻(Humulus lupulus)的基部导水组织增大而顶端缩小[38].表层根趋向形成窄径导管,以降低干旱与冻融栓塞风险[39]. ...
Linking physiological processes with mangrove forest structure:phosphorus deficiency limits canopy development,hydraulic conductivity and photosynthetic carbon gain in dwarf Rhizophora mangle
1
2006
... 荒漠植物的水力结构是其水分传导的关键基础,对抗逆性具有重要影响[23-24].其木质部由导管、管胞、穿孔板和纹孔等组成,构成高效低阻的输水系统[25-26].在干旱胁迫下,水力结构展现出可塑性:有的植物增大导管数量和直径以增强导水能力[27-28],而有的则缩小管胞和纹孔尺寸以控制水分流失[29].荒漠植物通常具备较小直径且壁厚的导管和管胞,有效降低气穴化形成的风险,提高抗栓塞能力[30].许多荒漠植物的导管连通方式较为保守,往往采用梯状或简单穿孔板[31],不仅有助于维持水分连续性,还能在高水势梯度下减少水力失败的可能性[32].荒漠植物中纤维-管胞和纤维-导管协同提升水力安全,其中管胞作为次级通道,抗旱性更强[33].许多荒漠木本植物纹孔常具加厚内缘或带状结构,减少水分流失并提高负压耐受性[34].某些荒漠树种采用带孔穿孔板,其开放面积较小,有助于防止气泡扩散,提高水分输导的稳定性[35].荒漠植物在导管和管胞的排列模式上表现出趋同进化,其较短且密集的水输导单元能够降低水分张力,提高水分的局部再分配能力,维持长期干旱下的水力连续性[36].某些荒漠裸子植物具特化纹孔膜结构,适应极端水势梯度[37].此外,水力结构随器官部位变化:如蛇麻(Humulus lupulus)的基部导水组织增大而顶端缩小[38].表层根趋向形成窄径导管,以降低干旱与冻融栓塞风险[39]. ...
Irradiance-induced changes in hydraulic architecture
1
2014
... 荒漠植物的水力结构是其水分传导的关键基础,对抗逆性具有重要影响[23-24].其木质部由导管、管胞、穿孔板和纹孔等组成,构成高效低阻的输水系统[25-26].在干旱胁迫下,水力结构展现出可塑性:有的植物增大导管数量和直径以增强导水能力[27-28],而有的则缩小管胞和纹孔尺寸以控制水分流失[29].荒漠植物通常具备较小直径且壁厚的导管和管胞,有效降低气穴化形成的风险,提高抗栓塞能力[30].许多荒漠植物的导管连通方式较为保守,往往采用梯状或简单穿孔板[31],不仅有助于维持水分连续性,还能在高水势梯度下减少水力失败的可能性[32].荒漠植物中纤维-管胞和纤维-导管协同提升水力安全,其中管胞作为次级通道,抗旱性更强[33].许多荒漠木本植物纹孔常具加厚内缘或带状结构,减少水分流失并提高负压耐受性[34].某些荒漠树种采用带孔穿孔板,其开放面积较小,有助于防止气泡扩散,提高水分输导的稳定性[35].荒漠植物在导管和管胞的排列模式上表现出趋同进化,其较短且密集的水输导单元能够降低水分张力,提高水分的局部再分配能力,维持长期干旱下的水力连续性[36].某些荒漠裸子植物具特化纹孔膜结构,适应极端水势梯度[37].此外,水力结构随器官部位变化:如蛇麻(Humulus lupulus)的基部导水组织增大而顶端缩小[38].表层根趋向形成窄径导管,以降低干旱与冻融栓塞风险[39]. ...
Azimuthal variations in xylem structure and water relations in Cork Oak (Quercus suber)
1
2011
... 荒漠植物的水力结构是其水分传导的关键基础,对抗逆性具有重要影响[23-24].其木质部由导管、管胞、穿孔板和纹孔等组成,构成高效低阻的输水系统[25-26].在干旱胁迫下,水力结构展现出可塑性:有的植物增大导管数量和直径以增强导水能力[27-28],而有的则缩小管胞和纹孔尺寸以控制水分流失[29].荒漠植物通常具备较小直径且壁厚的导管和管胞,有效降低气穴化形成的风险,提高抗栓塞能力[30].许多荒漠植物的导管连通方式较为保守,往往采用梯状或简单穿孔板[31],不仅有助于维持水分连续性,还能在高水势梯度下减少水力失败的可能性[32].荒漠植物中纤维-管胞和纤维-导管协同提升水力安全,其中管胞作为次级通道,抗旱性更强[33].许多荒漠木本植物纹孔常具加厚内缘或带状结构,减少水分流失并提高负压耐受性[34].某些荒漠树种采用带孔穿孔板,其开放面积较小,有助于防止气泡扩散,提高水分输导的稳定性[35].荒漠植物在导管和管胞的排列模式上表现出趋同进化,其较短且密集的水输导单元能够降低水分张力,提高水分的局部再分配能力,维持长期干旱下的水力连续性[36].某些荒漠裸子植物具特化纹孔膜结构,适应极端水势梯度[37].此外,水力结构随器官部位变化:如蛇麻(Humulus lupulus)的基部导水组织增大而顶端缩小[38].表层根趋向形成窄径导管,以降低干旱与冻融栓塞风险[39]. ...
Linking root traits to plant physiology and growth in Vahl.seedlings under soil compaction conditions
1
2012
... 荒漠植物的水力结构是其水分传导的关键基础,对抗逆性具有重要影响[23-24].其木质部由导管、管胞、穿孔板和纹孔等组成,构成高效低阻的输水系统[25-26].在干旱胁迫下,水力结构展现出可塑性:有的植物增大导管数量和直径以增强导水能力[27-28],而有的则缩小管胞和纹孔尺寸以控制水分流失[29].荒漠植物通常具备较小直径且壁厚的导管和管胞,有效降低气穴化形成的风险,提高抗栓塞能力[30].许多荒漠植物的导管连通方式较为保守,往往采用梯状或简单穿孔板[31],不仅有助于维持水分连续性,还能在高水势梯度下减少水力失败的可能性[32].荒漠植物中纤维-管胞和纤维-导管协同提升水力安全,其中管胞作为次级通道,抗旱性更强[33].许多荒漠木本植物纹孔常具加厚内缘或带状结构,减少水分流失并提高负压耐受性[34].某些荒漠树种采用带孔穿孔板,其开放面积较小,有助于防止气泡扩散,提高水分输导的稳定性[35].荒漠植物在导管和管胞的排列模式上表现出趋同进化,其较短且密集的水输导单元能够降低水分张力,提高水分的局部再分配能力,维持长期干旱下的水力连续性[36].某些荒漠裸子植物具特化纹孔膜结构,适应极端水势梯度[37].此外,水力结构随器官部位变化:如蛇麻(Humulus lupulus)的基部导水组织增大而顶端缩小[38].表层根趋向形成窄径导管,以降低干旱与冻融栓塞风险[39]. ...
Influence of the water treatment on the xylem anatomy and functionality of current year shoots of Olive Trees
1
2011
... 荒漠植物的水力结构是其水分传导的关键基础,对抗逆性具有重要影响[23-24].其木质部由导管、管胞、穿孔板和纹孔等组成,构成高效低阻的输水系统[25-26].在干旱胁迫下,水力结构展现出可塑性:有的植物增大导管数量和直径以增强导水能力[27-28],而有的则缩小管胞和纹孔尺寸以控制水分流失[29].荒漠植物通常具备较小直径且壁厚的导管和管胞,有效降低气穴化形成的风险,提高抗栓塞能力[30].许多荒漠植物的导管连通方式较为保守,往往采用梯状或简单穿孔板[31],不仅有助于维持水分连续性,还能在高水势梯度下减少水力失败的可能性[32].荒漠植物中纤维-管胞和纤维-导管协同提升水力安全,其中管胞作为次级通道,抗旱性更强[33].许多荒漠木本植物纹孔常具加厚内缘或带状结构,减少水分流失并提高负压耐受性[34].某些荒漠树种采用带孔穿孔板,其开放面积较小,有助于防止气泡扩散,提高水分输导的稳定性[35].荒漠植物在导管和管胞的排列模式上表现出趋同进化,其较短且密集的水输导单元能够降低水分张力,提高水分的局部再分配能力,维持长期干旱下的水力连续性[36].某些荒漠裸子植物具特化纹孔膜结构,适应极端水势梯度[37].此外,水力结构随器官部位变化:如蛇麻(Humulus lupulus)的基部导水组织增大而顶端缩小[38].表层根趋向形成窄径导管,以降低干旱与冻融栓塞风险[39]. ...
Hardwoods:anatomy and functionality of their elements:a short review
1
2024
... 荒漠植物的水力结构是其水分传导的关键基础,对抗逆性具有重要影响[23-24].其木质部由导管、管胞、穿孔板和纹孔等组成,构成高效低阻的输水系统[25-26].在干旱胁迫下,水力结构展现出可塑性:有的植物增大导管数量和直径以增强导水能力[27-28],而有的则缩小管胞和纹孔尺寸以控制水分流失[29].荒漠植物通常具备较小直径且壁厚的导管和管胞,有效降低气穴化形成的风险,提高抗栓塞能力[30].许多荒漠植物的导管连通方式较为保守,往往采用梯状或简单穿孔板[31],不仅有助于维持水分连续性,还能在高水势梯度下减少水力失败的可能性[32].荒漠植物中纤维-管胞和纤维-导管协同提升水力安全,其中管胞作为次级通道,抗旱性更强[33].许多荒漠木本植物纹孔常具加厚内缘或带状结构,减少水分流失并提高负压耐受性[34].某些荒漠树种采用带孔穿孔板,其开放面积较小,有助于防止气泡扩散,提高水分输导的稳定性[35].荒漠植物在导管和管胞的排列模式上表现出趋同进化,其较短且密集的水输导单元能够降低水分张力,提高水分的局部再分配能力,维持长期干旱下的水力连续性[36].某些荒漠裸子植物具特化纹孔膜结构,适应极端水势梯度[37].此外,水力结构随器官部位变化:如蛇麻(Humulus lupulus)的基部导水组织增大而顶端缩小[38].表层根趋向形成窄径导管,以降低干旱与冻融栓塞风险[39]. ...
Vestured pits:do they promote safer water transport?
1
2003
... 荒漠植物的水力结构是其水分传导的关键基础,对抗逆性具有重要影响[23-24].其木质部由导管、管胞、穿孔板和纹孔等组成,构成高效低阻的输水系统[25-26].在干旱胁迫下,水力结构展现出可塑性:有的植物增大导管数量和直径以增强导水能力[27-28],而有的则缩小管胞和纹孔尺寸以控制水分流失[29].荒漠植物通常具备较小直径且壁厚的导管和管胞,有效降低气穴化形成的风险,提高抗栓塞能力[30].许多荒漠植物的导管连通方式较为保守,往往采用梯状或简单穿孔板[31],不仅有助于维持水分连续性,还能在高水势梯度下减少水力失败的可能性[32].荒漠植物中纤维-管胞和纤维-导管协同提升水力安全,其中管胞作为次级通道,抗旱性更强[33].许多荒漠木本植物纹孔常具加厚内缘或带状结构,减少水分流失并提高负压耐受性[34].某些荒漠树种采用带孔穿孔板,其开放面积较小,有助于防止气泡扩散,提高水分输导的稳定性[35].荒漠植物在导管和管胞的排列模式上表现出趋同进化,其较短且密集的水输导单元能够降低水分张力,提高水分的局部再分配能力,维持长期干旱下的水力连续性[36].某些荒漠裸子植物具特化纹孔膜结构,适应极端水势梯度[37].此外,水力结构随器官部位变化:如蛇麻(Humulus lupulus)的基部导水组织增大而顶端缩小[38].表层根趋向形成窄径导管,以降低干旱与冻融栓塞风险[39]. ...
干旱-复水条件下红砂幼苗碳水动态特征研究
1
2024
... 荒漠植物的水力结构是其水分传导的关键基础,对抗逆性具有重要影响[23-24].其木质部由导管、管胞、穿孔板和纹孔等组成,构成高效低阻的输水系统[25-26].在干旱胁迫下,水力结构展现出可塑性:有的植物增大导管数量和直径以增强导水能力[27-28],而有的则缩小管胞和纹孔尺寸以控制水分流失[29].荒漠植物通常具备较小直径且壁厚的导管和管胞,有效降低气穴化形成的风险,提高抗栓塞能力[30].许多荒漠植物的导管连通方式较为保守,往往采用梯状或简单穿孔板[31],不仅有助于维持水分连续性,还能在高水势梯度下减少水力失败的可能性[32].荒漠植物中纤维-管胞和纤维-导管协同提升水力安全,其中管胞作为次级通道,抗旱性更强[33].许多荒漠木本植物纹孔常具加厚内缘或带状结构,减少水分流失并提高负压耐受性[34].某些荒漠树种采用带孔穿孔板,其开放面积较小,有助于防止气泡扩散,提高水分输导的稳定性[35].荒漠植物在导管和管胞的排列模式上表现出趋同进化,其较短且密集的水输导单元能够降低水分张力,提高水分的局部再分配能力,维持长期干旱下的水力连续性[36].某些荒漠裸子植物具特化纹孔膜结构,适应极端水势梯度[37].此外,水力结构随器官部位变化:如蛇麻(Humulus lupulus)的基部导水组织增大而顶端缩小[38].表层根趋向形成窄径导管,以降低干旱与冻融栓塞风险[39]. ...
Intervessel pit membrane thickness as a key determinant of embolism resistance in angiosperm xylem
1
2016
... 荒漠植物的水力结构是其水分传导的关键基础,对抗逆性具有重要影响[23-24].其木质部由导管、管胞、穿孔板和纹孔等组成,构成高效低阻的输水系统[25-26].在干旱胁迫下,水力结构展现出可塑性:有的植物增大导管数量和直径以增强导水能力[27-28],而有的则缩小管胞和纹孔尺寸以控制水分流失[29].荒漠植物通常具备较小直径且壁厚的导管和管胞,有效降低气穴化形成的风险,提高抗栓塞能力[30].许多荒漠植物的导管连通方式较为保守,往往采用梯状或简单穿孔板[31],不仅有助于维持水分连续性,还能在高水势梯度下减少水力失败的可能性[32].荒漠植物中纤维-管胞和纤维-导管协同提升水力安全,其中管胞作为次级通道,抗旱性更强[33].许多荒漠木本植物纹孔常具加厚内缘或带状结构,减少水分流失并提高负压耐受性[34].某些荒漠树种采用带孔穿孔板,其开放面积较小,有助于防止气泡扩散,提高水分输导的稳定性[35].荒漠植物在导管和管胞的排列模式上表现出趋同进化,其较短且密集的水输导单元能够降低水分张力,提高水分的局部再分配能力,维持长期干旱下的水力连续性[36].某些荒漠裸子植物具特化纹孔膜结构,适应极端水势梯度[37].此外,水力结构随器官部位变化:如蛇麻(Humulus lupulus)的基部导水组织增大而顶端缩小[38].表层根趋向形成窄径导管,以降低干旱与冻融栓塞风险[39]. ...
Variation in xylem structure from tropics to tundra:evidence from vestured pits
1
2004
... 荒漠植物的水力结构是其水分传导的关键基础,对抗逆性具有重要影响[23-24].其木质部由导管、管胞、穿孔板和纹孔等组成,构成高效低阻的输水系统[25-26].在干旱胁迫下,水力结构展现出可塑性:有的植物增大导管数量和直径以增强导水能力[27-28],而有的则缩小管胞和纹孔尺寸以控制水分流失[29].荒漠植物通常具备较小直径且壁厚的导管和管胞,有效降低气穴化形成的风险,提高抗栓塞能力[30].许多荒漠植物的导管连通方式较为保守,往往采用梯状或简单穿孔板[31],不仅有助于维持水分连续性,还能在高水势梯度下减少水力失败的可能性[32].荒漠植物中纤维-管胞和纤维-导管协同提升水力安全,其中管胞作为次级通道,抗旱性更强[33].许多荒漠木本植物纹孔常具加厚内缘或带状结构,减少水分流失并提高负压耐受性[34].某些荒漠树种采用带孔穿孔板,其开放面积较小,有助于防止气泡扩散,提高水分输导的稳定性[35].荒漠植物在导管和管胞的排列模式上表现出趋同进化,其较短且密集的水输导单元能够降低水分张力,提高水分的局部再分配能力,维持长期干旱下的水力连续性[36].某些荒漠裸子植物具特化纹孔膜结构,适应极端水势梯度[37].此外,水力结构随器官部位变化:如蛇麻(Humulus lupulus)的基部导水组织增大而顶端缩小[38].表层根趋向形成窄径导管,以降低干旱与冻融栓塞风险[39]. ...
The evolution of water transport in plants:an integrated approach
1
2010
... 荒漠植物的水力结构是其水分传导的关键基础,对抗逆性具有重要影响[23-24].其木质部由导管、管胞、穿孔板和纹孔等组成,构成高效低阻的输水系统[25-26].在干旱胁迫下,水力结构展现出可塑性:有的植物增大导管数量和直径以增强导水能力[27-28],而有的则缩小管胞和纹孔尺寸以控制水分流失[29].荒漠植物通常具备较小直径且壁厚的导管和管胞,有效降低气穴化形成的风险,提高抗栓塞能力[30].许多荒漠植物的导管连通方式较为保守,往往采用梯状或简单穿孔板[31],不仅有助于维持水分连续性,还能在高水势梯度下减少水力失败的可能性[32].荒漠植物中纤维-管胞和纤维-导管协同提升水力安全,其中管胞作为次级通道,抗旱性更强[33].许多荒漠木本植物纹孔常具加厚内缘或带状结构,减少水分流失并提高负压耐受性[34].某些荒漠树种采用带孔穿孔板,其开放面积较小,有助于防止气泡扩散,提高水分输导的稳定性[35].荒漠植物在导管和管胞的排列模式上表现出趋同进化,其较短且密集的水输导单元能够降低水分张力,提高水分的局部再分配能力,维持长期干旱下的水力连续性[36].某些荒漠裸子植物具特化纹孔膜结构,适应极端水势梯度[37].此外,水力结构随器官部位变化:如蛇麻(Humulus lupulus)的基部导水组织增大而顶端缩小[38].表层根趋向形成窄径导管,以降低干旱与冻融栓塞风险[39]. ...
Limits to xylem refilling under negative pressure in and
1
2003
... 荒漠植物的水力结构是其水分传导的关键基础,对抗逆性具有重要影响[23-24].其木质部由导管、管胞、穿孔板和纹孔等组成,构成高效低阻的输水系统[25-26].在干旱胁迫下,水力结构展现出可塑性:有的植物增大导管数量和直径以增强导水能力[27-28],而有的则缩小管胞和纹孔尺寸以控制水分流失[29].荒漠植物通常具备较小直径且壁厚的导管和管胞,有效降低气穴化形成的风险,提高抗栓塞能力[30].许多荒漠植物的导管连通方式较为保守,往往采用梯状或简单穿孔板[31],不仅有助于维持水分连续性,还能在高水势梯度下减少水力失败的可能性[32].荒漠植物中纤维-管胞和纤维-导管协同提升水力安全,其中管胞作为次级通道,抗旱性更强[33].许多荒漠木本植物纹孔常具加厚内缘或带状结构,减少水分流失并提高负压耐受性[34].某些荒漠树种采用带孔穿孔板,其开放面积较小,有助于防止气泡扩散,提高水分输导的稳定性[35].荒漠植物在导管和管胞的排列模式上表现出趋同进化,其较短且密集的水输导单元能够降低水分张力,提高水分的局部再分配能力,维持长期干旱下的水力连续性[36].某些荒漠裸子植物具特化纹孔膜结构,适应极端水势梯度[37].此外,水力结构随器官部位变化:如蛇麻(Humulus lupulus)的基部导水组织增大而顶端缩小[38].表层根趋向形成窄径导管,以降低干旱与冻融栓塞风险[39]. ...
Diversity in conduit and pit structure among extant gymnosperm taxa
1
2021
... 荒漠植物的水力结构是其水分传导的关键基础,对抗逆性具有重要影响[23-24].其木质部由导管、管胞、穿孔板和纹孔等组成,构成高效低阻的输水系统[25-26].在干旱胁迫下,水力结构展现出可塑性:有的植物增大导管数量和直径以增强导水能力[27-28],而有的则缩小管胞和纹孔尺寸以控制水分流失[29].荒漠植物通常具备较小直径且壁厚的导管和管胞,有效降低气穴化形成的风险,提高抗栓塞能力[30].许多荒漠植物的导管连通方式较为保守,往往采用梯状或简单穿孔板[31],不仅有助于维持水分连续性,还能在高水势梯度下减少水力失败的可能性[32].荒漠植物中纤维-管胞和纤维-导管协同提升水力安全,其中管胞作为次级通道,抗旱性更强[33].许多荒漠木本植物纹孔常具加厚内缘或带状结构,减少水分流失并提高负压耐受性[34].某些荒漠树种采用带孔穿孔板,其开放面积较小,有助于防止气泡扩散,提高水分输导的稳定性[35].荒漠植物在导管和管胞的排列模式上表现出趋同进化,其较短且密集的水输导单元能够降低水分张力,提高水分的局部再分配能力,维持长期干旱下的水力连续性[36].某些荒漠裸子植物具特化纹孔膜结构,适应极端水势梯度[37].此外,水力结构随器官部位变化:如蛇麻(Humulus lupulus)的基部导水组织增大而顶端缩小[38].表层根趋向形成窄径导管,以降低干旱与冻融栓塞风险[39]. ...
Effects of limited water availability on xylem transport in liana L
1
2016
... 荒漠植物的水力结构是其水分传导的关键基础,对抗逆性具有重要影响[23-24].其木质部由导管、管胞、穿孔板和纹孔等组成,构成高效低阻的输水系统[25-26].在干旱胁迫下,水力结构展现出可塑性:有的植物增大导管数量和直径以增强导水能力[27-28],而有的则缩小管胞和纹孔尺寸以控制水分流失[29].荒漠植物通常具备较小直径且壁厚的导管和管胞,有效降低气穴化形成的风险,提高抗栓塞能力[30].许多荒漠植物的导管连通方式较为保守,往往采用梯状或简单穿孔板[31],不仅有助于维持水分连续性,还能在高水势梯度下减少水力失败的可能性[32].荒漠植物中纤维-管胞和纤维-导管协同提升水力安全,其中管胞作为次级通道,抗旱性更强[33].许多荒漠木本植物纹孔常具加厚内缘或带状结构,减少水分流失并提高负压耐受性[34].某些荒漠树种采用带孔穿孔板,其开放面积较小,有助于防止气泡扩散,提高水分输导的稳定性[35].荒漠植物在导管和管胞的排列模式上表现出趋同进化,其较短且密集的水输导单元能够降低水分张力,提高水分的局部再分配能力,维持长期干旱下的水力连续性[36].某些荒漠裸子植物具特化纹孔膜结构,适应极端水势梯度[37].此外,水力结构随器官部位变化:如蛇麻(Humulus lupulus)的基部导水组织增大而顶端缩小[38].表层根趋向形成窄径导管,以降低干旱与冻融栓塞风险[39]. ...
Variation in xylem structure and function in stems and roots of trees to 20 m depth
1
2004
... 荒漠植物的水力结构是其水分传导的关键基础,对抗逆性具有重要影响[23-24].其木质部由导管、管胞、穿孔板和纹孔等组成,构成高效低阻的输水系统[25-26].在干旱胁迫下,水力结构展现出可塑性:有的植物增大导管数量和直径以增强导水能力[27-28],而有的则缩小管胞和纹孔尺寸以控制水分流失[29].荒漠植物通常具备较小直径且壁厚的导管和管胞,有效降低气穴化形成的风险,提高抗栓塞能力[30].许多荒漠植物的导管连通方式较为保守,往往采用梯状或简单穿孔板[31],不仅有助于维持水分连续性,还能在高水势梯度下减少水力失败的可能性[32].荒漠植物中纤维-管胞和纤维-导管协同提升水力安全,其中管胞作为次级通道,抗旱性更强[33].许多荒漠木本植物纹孔常具加厚内缘或带状结构,减少水分流失并提高负压耐受性[34].某些荒漠树种采用带孔穿孔板,其开放面积较小,有助于防止气泡扩散,提高水分输导的稳定性[35].荒漠植物在导管和管胞的排列模式上表现出趋同进化,其较短且密集的水输导单元能够降低水分张力,提高水分的局部再分配能力,维持长期干旱下的水力连续性[36].某些荒漠裸子植物具特化纹孔膜结构,适应极端水势梯度[37].此外,水力结构随器官部位变化:如蛇麻(Humulus lupulus)的基部导水组织增大而顶端缩小[38].表层根趋向形成窄径导管,以降低干旱与冻融栓塞风险[39]. ...
树木干旱死亡点预测及致死生理机制研究进展
1
2025
... 面对极端干旱、温度波动和高辐射胁迫等逆境环境,荒漠植物的生存依赖于复杂的水分调控机制,其中水力传导系统在维持植物水分平衡和适应逆境胁迫中发挥着重要作用[40-41].水力传导调控主要涉及根系水分吸收、茎干木质部的水分传导以及叶片蒸腾调节等多层次的结构和生理机制[42-45].针对固沙灌木柠条锦鸡儿和中间锦鸡儿(Caragana liouana)的研究发现,不同土壤水分条件显著影响其水力特性[46].对胡杨(Populus euphratica)幼株根、茎、叶水分传输通道对不同浓度盐胁迫的响应研究发现,盐胁迫下,胡杨可以通过协调导管输水的有效性和安全性来调节木质部的导水率,维持植物正常生长[47].还可以通过水力提升、水力下传及侧向再分配等根系再分配模式缓解土壤干旱对植物的影响[48]. ...
Stomatal and hydraulic redundancy allows woody species to adapt to arid environments
1
2025
... 面对极端干旱、温度波动和高辐射胁迫等逆境环境,荒漠植物的生存依赖于复杂的水分调控机制,其中水力传导系统在维持植物水分平衡和适应逆境胁迫中发挥着重要作用[40-41].水力传导调控主要涉及根系水分吸收、茎干木质部的水分传导以及叶片蒸腾调节等多层次的结构和生理机制[42-45].针对固沙灌木柠条锦鸡儿和中间锦鸡儿(Caragana liouana)的研究发现,不同土壤水分条件显著影响其水力特性[46].对胡杨(Populus euphratica)幼株根、茎、叶水分传输通道对不同浓度盐胁迫的响应研究发现,盐胁迫下,胡杨可以通过协调导管输水的有效性和安全性来调节木质部的导水率,维持植物正常生长[47].还可以通过水力提升、水力下传及侧向再分配等根系再分配模式缓解土壤干旱对植物的影响[48]. ...
Responses of woody plant functional traits to nitrogen addition:a meta-analysis of leaf economics,gas exchange,and hydraulic traits
1
2018
... 面对极端干旱、温度波动和高辐射胁迫等逆境环境,荒漠植物的生存依赖于复杂的水分调控机制,其中水力传导系统在维持植物水分平衡和适应逆境胁迫中发挥着重要作用[40-41].水力传导调控主要涉及根系水分吸收、茎干木质部的水分传导以及叶片蒸腾调节等多层次的结构和生理机制[42-45].针对固沙灌木柠条锦鸡儿和中间锦鸡儿(Caragana liouana)的研究发现,不同土壤水分条件显著影响其水力特性[46].对胡杨(Populus euphratica)幼株根、茎、叶水分传输通道对不同浓度盐胁迫的响应研究发现,盐胁迫下,胡杨可以通过协调导管输水的有效性和安全性来调节木质部的导水率,维持植物正常生长[47].还可以通过水力提升、水力下传及侧向再分配等根系再分配模式缓解土壤干旱对植物的影响[48]. ...
维管植物木质部水分传输过程的影响因素及研究进展
1
2017
... 植物水力传输的效率(水分传导速度)和安全性(抗栓塞能力)以及二者的平衡对于荒漠植物的生存至关重要,直接决定其生长的稳定性和抗逆性[49-50].然而,水力效率与安全性之间存在复杂的权衡关系.提升水力效率有助于增加植物的水分获取速率,但可能使水力系统更脆弱,易受干旱胁迫的影响;相反,增强水力安全性则通常意味着一定的水力效率损失,例如通过减小导管直径或增加纤维组织密度以提升抗栓塞能力[43].研究表明,腾格里沙漠东南缘的8种典型灌木能够通过调节叶片功能特性来优化水力系统,例如通过气孔调控减少蒸腾损失,同时提升叶片水力特性以提高水分传输效率[49].进一步研究发现,柠条锦鸡儿在干旱条件下优先保障生存而非生长,这表明在极端环境下,木质部水力安全性和效率的协同调节是维持植物生存的关键机制[51].同样地,对腾格里沙漠35种荒漠植物的水力功能的研究表明,植物的生存率主要受水力安全性和生长状况的影响.灌木和乔木在水力特性和生长策略上存在显著差异,灌木表现出更强的水力安全性,但导水效率较低,从而在干旱环境下保持较高的生存率.乔木虽具有较高的导水效率,但在严重干旱条件下更容易因栓塞而导致死亡[52]. ...
Evolution of water transport and xylem structure
2003
Deadly predictions in trees
1
2025
... 面对极端干旱、温度波动和高辐射胁迫等逆境环境,荒漠植物的生存依赖于复杂的水分调控机制,其中水力传导系统在维持植物水分平衡和适应逆境胁迫中发挥着重要作用[40-41].水力传导调控主要涉及根系水分吸收、茎干木质部的水分传导以及叶片蒸腾调节等多层次的结构和生理机制[42-45].针对固沙灌木柠条锦鸡儿和中间锦鸡儿(Caragana liouana)的研究发现,不同土壤水分条件显著影响其水力特性[46].对胡杨(Populus euphratica)幼株根、茎、叶水分传输通道对不同浓度盐胁迫的响应研究发现,盐胁迫下,胡杨可以通过协调导管输水的有效性和安全性来调节木质部的导水率,维持植物正常生长[47].还可以通过水力提升、水力下传及侧向再分配等根系再分配模式缓解土壤干旱对植物的影响[48]. ...
固沙灌木柠条锦鸡儿和中间锦鸡儿木质部导水与叶片光合能力对土壤水分的响应
1
2023
... 面对极端干旱、温度波动和高辐射胁迫等逆境环境,荒漠植物的生存依赖于复杂的水分调控机制,其中水力传导系统在维持植物水分平衡和适应逆境胁迫中发挥着重要作用[40-41].水力传导调控主要涉及根系水分吸收、茎干木质部的水分传导以及叶片蒸腾调节等多层次的结构和生理机制[42-45].针对固沙灌木柠条锦鸡儿和中间锦鸡儿(Caragana liouana)的研究发现,不同土壤水分条件显著影响其水力特性[46].对胡杨(Populus euphratica)幼株根、茎、叶水分传输通道对不同浓度盐胁迫的响应研究发现,盐胁迫下,胡杨可以通过协调导管输水的有效性和安全性来调节木质部的导水率,维持植物正常生长[47].还可以通过水力提升、水力下传及侧向再分配等根系再分配模式缓解土壤干旱对植物的影响[48]. ...
胡杨木质部水分传导对盐胁迫的响应与适应
1
2015
... 面对极端干旱、温度波动和高辐射胁迫等逆境环境,荒漠植物的生存依赖于复杂的水分调控机制,其中水力传导系统在维持植物水分平衡和适应逆境胁迫中发挥着重要作用[40-41].水力传导调控主要涉及根系水分吸收、茎干木质部的水分传导以及叶片蒸腾调节等多层次的结构和生理机制[42-45].针对固沙灌木柠条锦鸡儿和中间锦鸡儿(Caragana liouana)的研究发现,不同土壤水分条件显著影响其水力特性[46].对胡杨(Populus euphratica)幼株根、茎、叶水分传输通道对不同浓度盐胁迫的响应研究发现,盐胁迫下,胡杨可以通过协调导管输水的有效性和安全性来调节木质部的导水率,维持植物正常生长[47].还可以通过水力提升、水力下传及侧向再分配等根系再分配模式缓解土壤干旱对植物的影响[48]. ...
胡杨根系水力再分配的模式、大小及其影响因子
1
2014
... 面对极端干旱、温度波动和高辐射胁迫等逆境环境,荒漠植物的生存依赖于复杂的水分调控机制,其中水力传导系统在维持植物水分平衡和适应逆境胁迫中发挥着重要作用[40-41].水力传导调控主要涉及根系水分吸收、茎干木质部的水分传导以及叶片蒸腾调节等多层次的结构和生理机制[42-45].针对固沙灌木柠条锦鸡儿和中间锦鸡儿(Caragana liouana)的研究发现,不同土壤水分条件显著影响其水力特性[46].对胡杨(Populus euphratica)幼株根、茎、叶水分传输通道对不同浓度盐胁迫的响应研究发现,盐胁迫下,胡杨可以通过协调导管输水的有效性和安全性来调节木质部的导水率,维持植物正常生长[47].还可以通过水力提升、水力下传及侧向再分配等根系再分配模式缓解土壤干旱对植物的影响[48]. ...
Hydraulic trade-off and coordination strategies mediated by leaf functional traits of desert shrubs
2
2022
... 植物水力传输的效率(水分传导速度)和安全性(抗栓塞能力)以及二者的平衡对于荒漠植物的生存至关重要,直接决定其生长的稳定性和抗逆性[49-50].然而,水力效率与安全性之间存在复杂的权衡关系.提升水力效率有助于增加植物的水分获取速率,但可能使水力系统更脆弱,易受干旱胁迫的影响;相反,增强水力安全性则通常意味着一定的水力效率损失,例如通过减小导管直径或增加纤维组织密度以提升抗栓塞能力[43].研究表明,腾格里沙漠东南缘的8种典型灌木能够通过调节叶片功能特性来优化水力系统,例如通过气孔调控减少蒸腾损失,同时提升叶片水力特性以提高水分传输效率[49].进一步研究发现,柠条锦鸡儿在干旱条件下优先保障生存而非生长,这表明在极端环境下,木质部水力安全性和效率的协同调节是维持植物生存的关键机制[51].同样地,对腾格里沙漠35种荒漠植物的水力功能的研究表明,植物的生存率主要受水力安全性和生长状况的影响.灌木和乔木在水力特性和生长策略上存在显著差异,灌木表现出更强的水力安全性,但导水效率较低,从而在干旱环境下保持较高的生存率.乔木虽具有较高的导水效率,但在严重干旱条件下更容易因栓塞而导致死亡[52]. ...
... [49].进一步研究发现,柠条锦鸡儿在干旱条件下优先保障生存而非生长,这表明在极端环境下,木质部水力安全性和效率的协同调节是维持植物生存的关键机制[51].同样地,对腾格里沙漠35种荒漠植物的水力功能的研究表明,植物的生存率主要受水力安全性和生长状况的影响.灌木和乔木在水力特性和生长策略上存在显著差异,灌木表现出更强的水力安全性,但导水效率较低,从而在干旱环境下保持较高的生存率.乔木虽具有较高的导水效率,但在严重干旱条件下更容易因栓塞而导致死亡[52]. ...
A clear trade-off between leaf hydraulic efficiency and safety in an aridland shrub during regrowth
1
2021
... 植物水力传输的效率(水分传导速度)和安全性(抗栓塞能力)以及二者的平衡对于荒漠植物的生存至关重要,直接决定其生长的稳定性和抗逆性[49-50].然而,水力效率与安全性之间存在复杂的权衡关系.提升水力效率有助于增加植物的水分获取速率,但可能使水力系统更脆弱,易受干旱胁迫的影响;相反,增强水力安全性则通常意味着一定的水力效率损失,例如通过减小导管直径或增加纤维组织密度以提升抗栓塞能力[43].研究表明,腾格里沙漠东南缘的8种典型灌木能够通过调节叶片功能特性来优化水力系统,例如通过气孔调控减少蒸腾损失,同时提升叶片水力特性以提高水分传输效率[49].进一步研究发现,柠条锦鸡儿在干旱条件下优先保障生存而非生长,这表明在极端环境下,木质部水力安全性和效率的协同调节是维持植物生存的关键机制[51].同样地,对腾格里沙漠35种荒漠植物的水力功能的研究表明,植物的生存率主要受水力安全性和生长状况的影响.灌木和乔木在水力特性和生长策略上存在显著差异,灌木表现出更强的水力安全性,但导水效率较低,从而在干旱环境下保持较高的生存率.乔木虽具有较高的导水效率,但在严重干旱条件下更容易因栓塞而导致死亡[52]. ...
Hydraulic mechanism of limiting growth and maintaining survival of desert shrubs in arid habitats
1
2024
... 植物水力传输的效率(水分传导速度)和安全性(抗栓塞能力)以及二者的平衡对于荒漠植物的生存至关重要,直接决定其生长的稳定性和抗逆性[49-50].然而,水力效率与安全性之间存在复杂的权衡关系.提升水力效率有助于增加植物的水分获取速率,但可能使水力系统更脆弱,易受干旱胁迫的影响;相反,增强水力安全性则通常意味着一定的水力效率损失,例如通过减小导管直径或增加纤维组织密度以提升抗栓塞能力[43].研究表明,腾格里沙漠东南缘的8种典型灌木能够通过调节叶片功能特性来优化水力系统,例如通过气孔调控减少蒸腾损失,同时提升叶片水力特性以提高水分传输效率[49].进一步研究发现,柠条锦鸡儿在干旱条件下优先保障生存而非生长,这表明在极端环境下,木质部水力安全性和效率的协同调节是维持植物生存的关键机制[51].同样地,对腾格里沙漠35种荒漠植物的水力功能的研究表明,植物的生存率主要受水力安全性和生长状况的影响.灌木和乔木在水力特性和生长策略上存在显著差异,灌木表现出更强的水力安全性,但导水效率较低,从而在干旱环境下保持较高的生存率.乔木虽具有较高的导水效率,但在严重干旱条件下更容易因栓塞而导致死亡[52]. ...
Hydraulic safety and growth rather than climate of origin influence survival in desert shrubs and trees
1
2023
... 植物水力传输的效率(水分传导速度)和安全性(抗栓塞能力)以及二者的平衡对于荒漠植物的生存至关重要,直接决定其生长的稳定性和抗逆性[49-50].然而,水力效率与安全性之间存在复杂的权衡关系.提升水力效率有助于增加植物的水分获取速率,但可能使水力系统更脆弱,易受干旱胁迫的影响;相反,增强水力安全性则通常意味着一定的水力效率损失,例如通过减小导管直径或增加纤维组织密度以提升抗栓塞能力[43].研究表明,腾格里沙漠东南缘的8种典型灌木能够通过调节叶片功能特性来优化水力系统,例如通过气孔调控减少蒸腾损失,同时提升叶片水力特性以提高水分传输效率[49].进一步研究发现,柠条锦鸡儿在干旱条件下优先保障生存而非生长,这表明在极端环境下,木质部水力安全性和效率的协同调节是维持植物生存的关键机制[51].同样地,对腾格里沙漠35种荒漠植物的水力功能的研究表明,植物的生存率主要受水力安全性和生长状况的影响.灌木和乔木在水力特性和生长策略上存在显著差异,灌木表现出更强的水力安全性,但导水效率较低,从而在干旱环境下保持较高的生存率.乔木虽具有较高的导水效率,但在严重干旱条件下更容易因栓塞而导致死亡[52]. ...
我国干旱区植物抗旱机理研究进展
4
2010
... 荒漠植物在干旱、高温、强光等极端环境下,形成了多样化的光合碳同化机制,以提高水分利用和碳固定效率.其适应性主要体现为C₃、C₄和CAM光合途径的分化调控:C₄植物通过细胞分工和酶活性增强,提高CO₂浓缩效应,适应极端条件;C₃植物则依赖RuBisCO在温和环境中维持光合活性,尽管存在光呼吸损耗,仍能有效进行碳同化[53].然而对荒漠C3和C4植物研究也有不统一的结论.例如,对腾格里沙漠东南缘优势植物柠条锦鸡儿、油蒿、雾冰藜(Grubovia dasyphylla )和沙米(Agriophyllum squarrosum)进行UV-B辐射处理发现,辐射增强更能引起C4草本植物雾冰藜和沙米光合作用的下降,反而提高了C3灌木半灌木柠条锦鸡儿和油蒿水分利用效率[54].此外,部分荒漠植物(如景天科植物)采用CAM光合途径,通过夜间气孔开放固定CO₂、白天关闭气孔进行有机酸脱羧作用,以减少水分蒸腾损失,提高水分利用效率,这一策略使其能够在极端干旱环境下存活[55].除了光合途径的分化,气孔调控是荒漠植物光合适应的关键机制.通过降低气孔导度以减少蒸腾,大幅提升水分利用效率[53].为平衡CO₂摄取与水分保存,植物通过提高RuBisCO活性、增强叶绿素含量和优化气孔分布等策略,缓解光合作用与失水之间的矛盾[56].此外,叶片结构的可塑性也是荒漠植物适应逆境的重要方式[57].如胡杨的条形叶主要用于维持生长,光合效率低、抗逆性差,随着树体长大,条形叶逐渐被光合效率更高的卵形叶取代,但卵形叶耐光抑制能力较弱,最终被更适应强光、高温和干旱的锯齿叶取代,使胡杨能在极端环境中生存[58].同化枝的发育也是荒漠植物光合适应的重要策略,例如梭梭等植物在极端干旱环境下减少叶片生长,并依赖绿色的同化枝进行光合作用,以降低水分蒸腾损失,提高整体光合能力[10].同样,对塔克拉玛干沙漠防护林梭梭和头状沙拐枣(Calligonum caput-medusae)研究发现,干旱胁迫下这两种植物光合色素活性降低,激发能力下降,表明其以减缓同化枝生长和维持碳同化的方式抵御干旱[59]. ...
... [53].为平衡CO₂摄取与水分保存,植物通过提高RuBisCO活性、增强叶绿素含量和优化气孔分布等策略,缓解光合作用与失水之间的矛盾[56].此外,叶片结构的可塑性也是荒漠植物适应逆境的重要方式[57].如胡杨的条形叶主要用于维持生长,光合效率低、抗逆性差,随着树体长大,条形叶逐渐被光合效率更高的卵形叶取代,但卵形叶耐光抑制能力较弱,最终被更适应强光、高温和干旱的锯齿叶取代,使胡杨能在极端环境中生存[58].同化枝的发育也是荒漠植物光合适应的重要策略,例如梭梭等植物在极端干旱环境下减少叶片生长,并依赖绿色的同化枝进行光合作用,以降低水分蒸腾损失,提高整体光合能力[10].同样,对塔克拉玛干沙漠防护林梭梭和头状沙拐枣(Calligonum caput-medusae)研究发现,干旱胁迫下这两种植物光合色素活性降低,激发能力下降,表明其以减缓同化枝生长和维持碳同化的方式抵御干旱[59]. ...
... 渗透调节是荒漠植物维持细胞水分平衡和渗透稳态的重要机制,尤其在长期干旱和盐胁迫环境下表现得尤为显著[65].非酶抗氧化物质在荒漠植物中广泛存在,这些物质在清除自由基的同时,还兼具渗透调节作用,增强植物对环境胁迫的综合抵御能力[53,66].荒漠植物通过积累脯氨酸、可溶性糖、甜菜碱和无机离子等渗透调节物质降低渗透势,保持细胞内水分含量,进而确保代谢活动的正常进行[67-68].渗透调节物质在保护细胞膜、蛋白质和酶类活性中发挥重要作用.例如红砂在干旱胁迫下大量积累脯氨酸和可溶性糖,以维持细胞渗透势并保护细胞膜的完整性[69].柽柳在盐胁迫下通过积累可溶性糖和脯氨酸,不仅增强了抗氧化能力,还显著提升了对盐碱环境的适应性[63].无机离子的选择性吸收与转运也是荒漠植物应对盐胁迫的重要策略,通过调节钠、钾离子的比值,降低细胞毒性并保持渗透稳态[53].不同荒漠植物的适应策略存在显著差异.研究表明,通过不同渗透势的盐溶液处理两种耐盐性不同的枸杞(Lycium chinense),其在光合色素、质膜透性、渗透调节物质和抗氧化酶活性方面的响应有所不同[70];荒漠藓类植物在干旱和冻融条件下表现出强大的渗透调节能力和抗氧化防御体系,以应对细胞水分丧失和ROS的双重胁迫[71];裸果木(Gymnocarpos przewalskii)则通过调节不同生长季节的抗氧化酶活性和渗透调节物质含量来适应环境中水分条件的变化[72].此外,一些荒漠植物如霸王(Zygophyllum xanthoxylum)和荆芥(Nepeta cataria)在盐胁迫下通过细胞离子平衡调控和非酶抗氧化物积累来提升其抗逆性[73-74]. ...
... [53].不同荒漠植物的适应策略存在显著差异.研究表明,通过不同渗透势的盐溶液处理两种耐盐性不同的枸杞(Lycium chinense),其在光合色素、质膜透性、渗透调节物质和抗氧化酶活性方面的响应有所不同[70];荒漠藓类植物在干旱和冻融条件下表现出强大的渗透调节能力和抗氧化防御体系,以应对细胞水分丧失和ROS的双重胁迫[71];裸果木(Gymnocarpos przewalskii)则通过调节不同生长季节的抗氧化酶活性和渗透调节物质含量来适应环境中水分条件的变化[72].此外,一些荒漠植物如霸王(Zygophyllum xanthoxylum)和荆芥(Nepeta cataria)在盐胁迫下通过细胞离子平衡调控和非酶抗氧化物积累来提升其抗逆性[73-74]. ...
CO2浓度升高和UV-B辐射增强对沙坡头优势植物光合作用的影响
1
2008
... 荒漠植物在干旱、高温、强光等极端环境下,形成了多样化的光合碳同化机制,以提高水分利用和碳固定效率.其适应性主要体现为C₃、C₄和CAM光合途径的分化调控:C₄植物通过细胞分工和酶活性增强,提高CO₂浓缩效应,适应极端条件;C₃植物则依赖RuBisCO在温和环境中维持光合活性,尽管存在光呼吸损耗,仍能有效进行碳同化[53].然而对荒漠C3和C4植物研究也有不统一的结论.例如,对腾格里沙漠东南缘优势植物柠条锦鸡儿、油蒿、雾冰藜(Grubovia dasyphylla )和沙米(Agriophyllum squarrosum)进行UV-B辐射处理发现,辐射增强更能引起C4草本植物雾冰藜和沙米光合作用的下降,反而提高了C3灌木半灌木柠条锦鸡儿和油蒿水分利用效率[54].此外,部分荒漠植物(如景天科植物)采用CAM光合途径,通过夜间气孔开放固定CO₂、白天关闭气孔进行有机酸脱羧作用,以减少水分蒸腾损失,提高水分利用效率,这一策略使其能够在极端干旱环境下存活[55].除了光合途径的分化,气孔调控是荒漠植物光合适应的关键机制.通过降低气孔导度以减少蒸腾,大幅提升水分利用效率[53].为平衡CO₂摄取与水分保存,植物通过提高RuBisCO活性、增强叶绿素含量和优化气孔分布等策略,缓解光合作用与失水之间的矛盾[56].此外,叶片结构的可塑性也是荒漠植物适应逆境的重要方式[57].如胡杨的条形叶主要用于维持生长,光合效率低、抗逆性差,随着树体长大,条形叶逐渐被光合效率更高的卵形叶取代,但卵形叶耐光抑制能力较弱,最终被更适应强光、高温和干旱的锯齿叶取代,使胡杨能在极端环境中生存[58].同化枝的发育也是荒漠植物光合适应的重要策略,例如梭梭等植物在极端干旱环境下减少叶片生长,并依赖绿色的同化枝进行光合作用,以降低水分蒸腾损失,提高整体光合能力[10].同样,对塔克拉玛干沙漠防护林梭梭和头状沙拐枣(Calligonum caput-medusae)研究发现,干旱胁迫下这两种植物光合色素活性降低,激发能力下降,表明其以减缓同化枝生长和维持碳同化的方式抵御干旱[59]. ...
石斛属植物光合碳同化途径地理分异研究进展
1
2015
... 荒漠植物在干旱、高温、强光等极端环境下,形成了多样化的光合碳同化机制,以提高水分利用和碳固定效率.其适应性主要体现为C₃、C₄和CAM光合途径的分化调控:C₄植物通过细胞分工和酶活性增强,提高CO₂浓缩效应,适应极端条件;C₃植物则依赖RuBisCO在温和环境中维持光合活性,尽管存在光呼吸损耗,仍能有效进行碳同化[53].然而对荒漠C3和C4植物研究也有不统一的结论.例如,对腾格里沙漠东南缘优势植物柠条锦鸡儿、油蒿、雾冰藜(Grubovia dasyphylla )和沙米(Agriophyllum squarrosum)进行UV-B辐射处理发现,辐射增强更能引起C4草本植物雾冰藜和沙米光合作用的下降,反而提高了C3灌木半灌木柠条锦鸡儿和油蒿水分利用效率[54].此外,部分荒漠植物(如景天科植物)采用CAM光合途径,通过夜间气孔开放固定CO₂、白天关闭气孔进行有机酸脱羧作用,以减少水分蒸腾损失,提高水分利用效率,这一策略使其能够在极端干旱环境下存活[55].除了光合途径的分化,气孔调控是荒漠植物光合适应的关键机制.通过降低气孔导度以减少蒸腾,大幅提升水分利用效率[53].为平衡CO₂摄取与水分保存,植物通过提高RuBisCO活性、增强叶绿素含量和优化气孔分布等策略,缓解光合作用与失水之间的矛盾[56].此外,叶片结构的可塑性也是荒漠植物适应逆境的重要方式[57].如胡杨的条形叶主要用于维持生长,光合效率低、抗逆性差,随着树体长大,条形叶逐渐被光合效率更高的卵形叶取代,但卵形叶耐光抑制能力较弱,最终被更适应强光、高温和干旱的锯齿叶取代,使胡杨能在极端环境中生存[58].同化枝的发育也是荒漠植物光合适应的重要策略,例如梭梭等植物在极端干旱环境下减少叶片生长,并依赖绿色的同化枝进行光合作用,以降低水分蒸腾损失,提高整体光合能力[10].同样,对塔克拉玛干沙漠防护林梭梭和头状沙拐枣(Calligonum caput-medusae)研究发现,干旱胁迫下这两种植物光合色素活性降低,激发能力下降,表明其以减缓同化枝生长和维持碳同化的方式抵御干旱[59]. ...
作物光合效率对逆境适应及提高途径研究进展
1
2024
... 荒漠植物在干旱、高温、强光等极端环境下,形成了多样化的光合碳同化机制,以提高水分利用和碳固定效率.其适应性主要体现为C₃、C₄和CAM光合途径的分化调控:C₄植物通过细胞分工和酶活性增强,提高CO₂浓缩效应,适应极端条件;C₃植物则依赖RuBisCO在温和环境中维持光合活性,尽管存在光呼吸损耗,仍能有效进行碳同化[53].然而对荒漠C3和C4植物研究也有不统一的结论.例如,对腾格里沙漠东南缘优势植物柠条锦鸡儿、油蒿、雾冰藜(Grubovia dasyphylla )和沙米(Agriophyllum squarrosum)进行UV-B辐射处理发现,辐射增强更能引起C4草本植物雾冰藜和沙米光合作用的下降,反而提高了C3灌木半灌木柠条锦鸡儿和油蒿水分利用效率[54].此外,部分荒漠植物(如景天科植物)采用CAM光合途径,通过夜间气孔开放固定CO₂、白天关闭气孔进行有机酸脱羧作用,以减少水分蒸腾损失,提高水分利用效率,这一策略使其能够在极端干旱环境下存活[55].除了光合途径的分化,气孔调控是荒漠植物光合适应的关键机制.通过降低气孔导度以减少蒸腾,大幅提升水分利用效率[53].为平衡CO₂摄取与水分保存,植物通过提高RuBisCO活性、增强叶绿素含量和优化气孔分布等策略,缓解光合作用与失水之间的矛盾[56].此外,叶片结构的可塑性也是荒漠植物适应逆境的重要方式[57].如胡杨的条形叶主要用于维持生长,光合效率低、抗逆性差,随着树体长大,条形叶逐渐被光合效率更高的卵形叶取代,但卵形叶耐光抑制能力较弱,最终被更适应强光、高温和干旱的锯齿叶取代,使胡杨能在极端环境中生存[58].同化枝的发育也是荒漠植物光合适应的重要策略,例如梭梭等植物在极端干旱环境下减少叶片生长,并依赖绿色的同化枝进行光合作用,以降低水分蒸腾损失,提高整体光合能力[10].同样,对塔克拉玛干沙漠防护林梭梭和头状沙拐枣(Calligonum caput-medusae)研究发现,干旱胁迫下这两种植物光合色素活性降低,激发能力下降,表明其以减缓同化枝生长和维持碳同化的方式抵御干旱[59]. ...
植物叶片形态的生态功能、地理分布与成因
1
2021
... 荒漠植物在干旱、高温、强光等极端环境下,形成了多样化的光合碳同化机制,以提高水分利用和碳固定效率.其适应性主要体现为C₃、C₄和CAM光合途径的分化调控:C₄植物通过细胞分工和酶活性增强,提高CO₂浓缩效应,适应极端条件;C₃植物则依赖RuBisCO在温和环境中维持光合活性,尽管存在光呼吸损耗,仍能有效进行碳同化[53].然而对荒漠C3和C4植物研究也有不统一的结论.例如,对腾格里沙漠东南缘优势植物柠条锦鸡儿、油蒿、雾冰藜(Grubovia dasyphylla )和沙米(Agriophyllum squarrosum)进行UV-B辐射处理发现,辐射增强更能引起C4草本植物雾冰藜和沙米光合作用的下降,反而提高了C3灌木半灌木柠条锦鸡儿和油蒿水分利用效率[54].此外,部分荒漠植物(如景天科植物)采用CAM光合途径,通过夜间气孔开放固定CO₂、白天关闭气孔进行有机酸脱羧作用,以减少水分蒸腾损失,提高水分利用效率,这一策略使其能够在极端干旱环境下存活[55].除了光合途径的分化,气孔调控是荒漠植物光合适应的关键机制.通过降低气孔导度以减少蒸腾,大幅提升水分利用效率[53].为平衡CO₂摄取与水分保存,植物通过提高RuBisCO活性、增强叶绿素含量和优化气孔分布等策略,缓解光合作用与失水之间的矛盾[56].此外,叶片结构的可塑性也是荒漠植物适应逆境的重要方式[57].如胡杨的条形叶主要用于维持生长,光合效率低、抗逆性差,随着树体长大,条形叶逐渐被光合效率更高的卵形叶取代,但卵形叶耐光抑制能力较弱,最终被更适应强光、高温和干旱的锯齿叶取代,使胡杨能在极端环境中生存[58].同化枝的发育也是荒漠植物光合适应的重要策略,例如梭梭等植物在极端干旱环境下减少叶片生长,并依赖绿色的同化枝进行光合作用,以降低水分蒸腾损失,提高整体光合能力[10].同样,对塔克拉玛干沙漠防护林梭梭和头状沙拐枣(Calligonum caput-medusae)研究发现,干旱胁迫下这两种植物光合色素活性降低,激发能力下降,表明其以减缓同化枝生长和维持碳同化的方式抵御干旱[59]. ...
胡杨异形叶光合作用对光强与CO2浓度的响应
1
2014
... 荒漠植物在干旱、高温、强光等极端环境下,形成了多样化的光合碳同化机制,以提高水分利用和碳固定效率.其适应性主要体现为C₃、C₄和CAM光合途径的分化调控:C₄植物通过细胞分工和酶活性增强,提高CO₂浓缩效应,适应极端条件;C₃植物则依赖RuBisCO在温和环境中维持光合活性,尽管存在光呼吸损耗,仍能有效进行碳同化[53].然而对荒漠C3和C4植物研究也有不统一的结论.例如,对腾格里沙漠东南缘优势植物柠条锦鸡儿、油蒿、雾冰藜(Grubovia dasyphylla )和沙米(Agriophyllum squarrosum)进行UV-B辐射处理发现,辐射增强更能引起C4草本植物雾冰藜和沙米光合作用的下降,反而提高了C3灌木半灌木柠条锦鸡儿和油蒿水分利用效率[54].此外,部分荒漠植物(如景天科植物)采用CAM光合途径,通过夜间气孔开放固定CO₂、白天关闭气孔进行有机酸脱羧作用,以减少水分蒸腾损失,提高水分利用效率,这一策略使其能够在极端干旱环境下存活[55].除了光合途径的分化,气孔调控是荒漠植物光合适应的关键机制.通过降低气孔导度以减少蒸腾,大幅提升水分利用效率[53].为平衡CO₂摄取与水分保存,植物通过提高RuBisCO活性、增强叶绿素含量和优化气孔分布等策略,缓解光合作用与失水之间的矛盾[56].此外,叶片结构的可塑性也是荒漠植物适应逆境的重要方式[57].如胡杨的条形叶主要用于维持生长,光合效率低、抗逆性差,随着树体长大,条形叶逐渐被光合效率更高的卵形叶取代,但卵形叶耐光抑制能力较弱,最终被更适应强光、高温和干旱的锯齿叶取代,使胡杨能在极端环境中生存[58].同化枝的发育也是荒漠植物光合适应的重要策略,例如梭梭等植物在极端干旱环境下减少叶片生长,并依赖绿色的同化枝进行光合作用,以降低水分蒸腾损失,提高整体光合能力[10].同样,对塔克拉玛干沙漠防护林梭梭和头状沙拐枣(Calligonum caput-medusae)研究发现,干旱胁迫下这两种植物光合色素活性降低,激发能力下降,表明其以减缓同化枝生长和维持碳同化的方式抵御干旱[59]. ...
梭梭和头状沙拐枣形态及生理生化特性对干旱胁迫的响应
1
2024
... 荒漠植物在干旱、高温、强光等极端环境下,形成了多样化的光合碳同化机制,以提高水分利用和碳固定效率.其适应性主要体现为C₃、C₄和CAM光合途径的分化调控:C₄植物通过细胞分工和酶活性增强,提高CO₂浓缩效应,适应极端条件;C₃植物则依赖RuBisCO在温和环境中维持光合活性,尽管存在光呼吸损耗,仍能有效进行碳同化[53].然而对荒漠C3和C4植物研究也有不统一的结论.例如,对腾格里沙漠东南缘优势植物柠条锦鸡儿、油蒿、雾冰藜(Grubovia dasyphylla )和沙米(Agriophyllum squarrosum)进行UV-B辐射处理发现,辐射增强更能引起C4草本植物雾冰藜和沙米光合作用的下降,反而提高了C3灌木半灌木柠条锦鸡儿和油蒿水分利用效率[54].此外,部分荒漠植物(如景天科植物)采用CAM光合途径,通过夜间气孔开放固定CO₂、白天关闭气孔进行有机酸脱羧作用,以减少水分蒸腾损失,提高水分利用效率,这一策略使其能够在极端干旱环境下存活[55].除了光合途径的分化,气孔调控是荒漠植物光合适应的关键机制.通过降低气孔导度以减少蒸腾,大幅提升水分利用效率[53].为平衡CO₂摄取与水分保存,植物通过提高RuBisCO活性、增强叶绿素含量和优化气孔分布等策略,缓解光合作用与失水之间的矛盾[56].此外,叶片结构的可塑性也是荒漠植物适应逆境的重要方式[57].如胡杨的条形叶主要用于维持生长,光合效率低、抗逆性差,随着树体长大,条形叶逐渐被光合效率更高的卵形叶取代,但卵形叶耐光抑制能力较弱,最终被更适应强光、高温和干旱的锯齿叶取代,使胡杨能在极端环境中生存[58].同化枝的发育也是荒漠植物光合适应的重要策略,例如梭梭等植物在极端干旱环境下减少叶片生长,并依赖绿色的同化枝进行光合作用,以降低水分蒸腾损失,提高整体光合能力[10].同样,对塔克拉玛干沙漠防护林梭梭和头状沙拐枣(Calligonum caput-medusae)研究发现,干旱胁迫下这两种植物光合色素活性降低,激发能力下降,表明其以减缓同化枝生长和维持碳同化的方式抵御干旱[59]. ...
Synergistic effects of exogenous IAA and melatonin on seed priming and physiological biochemistry of three desert plants in saline-alkali soil
1
2024
... 荒漠植物通过复杂的抗氧化系统与精细的渗透调节能力来维持细胞稳态,确保其在恶劣环境下的生存和繁殖.抗氧化系统是荒漠植物应对逆境的第一道防线.在逆境胁迫下,植物体内活性氧(ROS)水平显著升高,威胁细胞膜的完整性及代谢稳定性,荒漠植物通过显著上调抗氧化酶类活性来清除过量ROS,如超氧化物歧化酶(SOD)、过氧化物酶(CAT)和抗坏血酸过氧化物酶(APX)等[60-61],从而能够保护细胞结构的完整性,防止膜脂过氧化对植物生理功能造成损坏[62].如柽柳(Tamarix chinensis)在盐胁迫下显著增强了SOD和CAT活性,降低了膜脂过氧化水平,保证了细胞的正常代谢[63].类似地,梭梭对干旱的代谢和生理适应机制研究表明,不同抗氧化酶的差异性调控提高了梭梭的耐旱潜力[64]. ...
Reactive oxygen species signalling in plant stress responses
1
2022
... 荒漠植物通过复杂的抗氧化系统与精细的渗透调节能力来维持细胞稳态,确保其在恶劣环境下的生存和繁殖.抗氧化系统是荒漠植物应对逆境的第一道防线.在逆境胁迫下,植物体内活性氧(ROS)水平显著升高,威胁细胞膜的完整性及代谢稳定性,荒漠植物通过显著上调抗氧化酶类活性来清除过量ROS,如超氧化物歧化酶(SOD)、过氧化物酶(CAT)和抗坏血酸过氧化物酶(APX)等[60-61],从而能够保护细胞结构的完整性,防止膜脂过氧化对植物生理功能造成损坏[62].如柽柳(Tamarix chinensis)在盐胁迫下显著增强了SOD和CAT活性,降低了膜脂过氧化水平,保证了细胞的正常代谢[63].类似地,梭梭对干旱的代谢和生理适应机制研究表明,不同抗氧化酶的差异性调控提高了梭梭的耐旱潜力[64]. ...
A look into osmotic,ionic,and redox adjustments in wild tomato species under combined salt and water stress
1
2024
... 荒漠植物通过复杂的抗氧化系统与精细的渗透调节能力来维持细胞稳态,确保其在恶劣环境下的生存和繁殖.抗氧化系统是荒漠植物应对逆境的第一道防线.在逆境胁迫下,植物体内活性氧(ROS)水平显著升高,威胁细胞膜的完整性及代谢稳定性,荒漠植物通过显著上调抗氧化酶类活性来清除过量ROS,如超氧化物歧化酶(SOD)、过氧化物酶(CAT)和抗坏血酸过氧化物酶(APX)等[60-61],从而能够保护细胞结构的完整性,防止膜脂过氧化对植物生理功能造成损坏[62].如柽柳(Tamarix chinensis)在盐胁迫下显著增强了SOD和CAT活性,降低了膜脂过氧化水平,保证了细胞的正常代谢[63].类似地,梭梭对干旱的代谢和生理适应机制研究表明,不同抗氧化酶的差异性调控提高了梭梭的耐旱潜力[64]. ...
柽柳组培苗抗氧化酶及渗透调节物质对NaCl胁迫的响应
2
2015
... 荒漠植物通过复杂的抗氧化系统与精细的渗透调节能力来维持细胞稳态,确保其在恶劣环境下的生存和繁殖.抗氧化系统是荒漠植物应对逆境的第一道防线.在逆境胁迫下,植物体内活性氧(ROS)水平显著升高,威胁细胞膜的完整性及代谢稳定性,荒漠植物通过显著上调抗氧化酶类活性来清除过量ROS,如超氧化物歧化酶(SOD)、过氧化物酶(CAT)和抗坏血酸过氧化物酶(APX)等[60-61],从而能够保护细胞结构的完整性,防止膜脂过氧化对植物生理功能造成损坏[62].如柽柳(Tamarix chinensis)在盐胁迫下显著增强了SOD和CAT活性,降低了膜脂过氧化水平,保证了细胞的正常代谢[63].类似地,梭梭对干旱的代谢和生理适应机制研究表明,不同抗氧化酶的差异性调控提高了梭梭的耐旱潜力[64]. ...
... 渗透调节是荒漠植物维持细胞水分平衡和渗透稳态的重要机制,尤其在长期干旱和盐胁迫环境下表现得尤为显著[65].非酶抗氧化物质在荒漠植物中广泛存在,这些物质在清除自由基的同时,还兼具渗透调节作用,增强植物对环境胁迫的综合抵御能力[53,66].荒漠植物通过积累脯氨酸、可溶性糖、甜菜碱和无机离子等渗透调节物质降低渗透势,保持细胞内水分含量,进而确保代谢活动的正常进行[67-68].渗透调节物质在保护细胞膜、蛋白质和酶类活性中发挥重要作用.例如红砂在干旱胁迫下大量积累脯氨酸和可溶性糖,以维持细胞渗透势并保护细胞膜的完整性[69].柽柳在盐胁迫下通过积累可溶性糖和脯氨酸,不仅增强了抗氧化能力,还显著提升了对盐碱环境的适应性[63].无机离子的选择性吸收与转运也是荒漠植物应对盐胁迫的重要策略,通过调节钠、钾离子的比值,降低细胞毒性并保持渗透稳态[53].不同荒漠植物的适应策略存在显著差异.研究表明,通过不同渗透势的盐溶液处理两种耐盐性不同的枸杞(Lycium chinense),其在光合色素、质膜透性、渗透调节物质和抗氧化酶活性方面的响应有所不同[70];荒漠藓类植物在干旱和冻融条件下表现出强大的渗透调节能力和抗氧化防御体系,以应对细胞水分丧失和ROS的双重胁迫[71];裸果木(Gymnocarpos przewalskii)则通过调节不同生长季节的抗氧化酶活性和渗透调节物质含量来适应环境中水分条件的变化[72].此外,一些荒漠植物如霸王(Zygophyllum xanthoxylum)和荆芥(Nepeta cataria)在盐胁迫下通过细胞离子平衡调控和非酶抗氧化物积累来提升其抗逆性[73-74]. ...
Physiological and metabolic adjustments in the xero-halophyte Haloxylon salicornicum conferring drought tolerance
1
2021
... 荒漠植物通过复杂的抗氧化系统与精细的渗透调节能力来维持细胞稳态,确保其在恶劣环境下的生存和繁殖.抗氧化系统是荒漠植物应对逆境的第一道防线.在逆境胁迫下,植物体内活性氧(ROS)水平显著升高,威胁细胞膜的完整性及代谢稳定性,荒漠植物通过显著上调抗氧化酶类活性来清除过量ROS,如超氧化物歧化酶(SOD)、过氧化物酶(CAT)和抗坏血酸过氧化物酶(APX)等[60-61],从而能够保护细胞结构的完整性,防止膜脂过氧化对植物生理功能造成损坏[62].如柽柳(Tamarix chinensis)在盐胁迫下显著增强了SOD和CAT活性,降低了膜脂过氧化水平,保证了细胞的正常代谢[63].类似地,梭梭对干旱的代谢和生理适应机制研究表明,不同抗氧化酶的差异性调控提高了梭梭的耐旱潜力[64]. ...
The role of solute accumulation,osmotic adjustment and changes in cell wall elasticity in drought tolerance in (Lamk)
1
1998
... 渗透调节是荒漠植物维持细胞水分平衡和渗透稳态的重要机制,尤其在长期干旱和盐胁迫环境下表现得尤为显著[65].非酶抗氧化物质在荒漠植物中广泛存在,这些物质在清除自由基的同时,还兼具渗透调节作用,增强植物对环境胁迫的综合抵御能力[53,66].荒漠植物通过积累脯氨酸、可溶性糖、甜菜碱和无机离子等渗透调节物质降低渗透势,保持细胞内水分含量,进而确保代谢活动的正常进行[67-68].渗透调节物质在保护细胞膜、蛋白质和酶类活性中发挥重要作用.例如红砂在干旱胁迫下大量积累脯氨酸和可溶性糖,以维持细胞渗透势并保护细胞膜的完整性[69].柽柳在盐胁迫下通过积累可溶性糖和脯氨酸,不仅增强了抗氧化能力,还显著提升了对盐碱环境的适应性[63].无机离子的选择性吸收与转运也是荒漠植物应对盐胁迫的重要策略,通过调节钠、钾离子的比值,降低细胞毒性并保持渗透稳态[53].不同荒漠植物的适应策略存在显著差异.研究表明,通过不同渗透势的盐溶液处理两种耐盐性不同的枸杞(Lycium chinense),其在光合色素、质膜透性、渗透调节物质和抗氧化酶活性方面的响应有所不同[70];荒漠藓类植物在干旱和冻融条件下表现出强大的渗透调节能力和抗氧化防御体系,以应对细胞水分丧失和ROS的双重胁迫[71];裸果木(Gymnocarpos przewalskii)则通过调节不同生长季节的抗氧化酶活性和渗透调节物质含量来适应环境中水分条件的变化[72].此外,一些荒漠植物如霸王(Zygophyllum xanthoxylum)和荆芥(Nepeta cataria)在盐胁迫下通过细胞离子平衡调控和非酶抗氧化物积累来提升其抗逆性[73-74]. ...
Calligonum mongolicum employs a variety of physiological and biochemical strategies to acclimatize to hyperarid saline deserts
1
2025
... 渗透调节是荒漠植物维持细胞水分平衡和渗透稳态的重要机制,尤其在长期干旱和盐胁迫环境下表现得尤为显著[65].非酶抗氧化物质在荒漠植物中广泛存在,这些物质在清除自由基的同时,还兼具渗透调节作用,增强植物对环境胁迫的综合抵御能力[53,66].荒漠植物通过积累脯氨酸、可溶性糖、甜菜碱和无机离子等渗透调节物质降低渗透势,保持细胞内水分含量,进而确保代谢活动的正常进行[67-68].渗透调节物质在保护细胞膜、蛋白质和酶类活性中发挥重要作用.例如红砂在干旱胁迫下大量积累脯氨酸和可溶性糖,以维持细胞渗透势并保护细胞膜的完整性[69].柽柳在盐胁迫下通过积累可溶性糖和脯氨酸,不仅增强了抗氧化能力,还显著提升了对盐碱环境的适应性[63].无机离子的选择性吸收与转运也是荒漠植物应对盐胁迫的重要策略,通过调节钠、钾离子的比值,降低细胞毒性并保持渗透稳态[53].不同荒漠植物的适应策略存在显著差异.研究表明,通过不同渗透势的盐溶液处理两种耐盐性不同的枸杞(Lycium chinense),其在光合色素、质膜透性、渗透调节物质和抗氧化酶活性方面的响应有所不同[70];荒漠藓类植物在干旱和冻融条件下表现出强大的渗透调节能力和抗氧化防御体系,以应对细胞水分丧失和ROS的双重胁迫[71];裸果木(Gymnocarpos przewalskii)则通过调节不同生长季节的抗氧化酶活性和渗透调节物质含量来适应环境中水分条件的变化[72].此外,一些荒漠植物如霸王(Zygophyllum xanthoxylum)和荆芥(Nepeta cataria)在盐胁迫下通过细胞离子平衡调控和非酶抗氧化物积累来提升其抗逆性[73-74]. ...
干旱胁迫对沙芥和斧形沙芥幼苗生长及抗氧化系统的影响
1
2017
... 渗透调节是荒漠植物维持细胞水分平衡和渗透稳态的重要机制,尤其在长期干旱和盐胁迫环境下表现得尤为显著[65].非酶抗氧化物质在荒漠植物中广泛存在,这些物质在清除自由基的同时,还兼具渗透调节作用,增强植物对环境胁迫的综合抵御能力[53,66].荒漠植物通过积累脯氨酸、可溶性糖、甜菜碱和无机离子等渗透调节物质降低渗透势,保持细胞内水分含量,进而确保代谢活动的正常进行[67-68].渗透调节物质在保护细胞膜、蛋白质和酶类活性中发挥重要作用.例如红砂在干旱胁迫下大量积累脯氨酸和可溶性糖,以维持细胞渗透势并保护细胞膜的完整性[69].柽柳在盐胁迫下通过积累可溶性糖和脯氨酸,不仅增强了抗氧化能力,还显著提升了对盐碱环境的适应性[63].无机离子的选择性吸收与转运也是荒漠植物应对盐胁迫的重要策略,通过调节钠、钾离子的比值,降低细胞毒性并保持渗透稳态[53].不同荒漠植物的适应策略存在显著差异.研究表明,通过不同渗透势的盐溶液处理两种耐盐性不同的枸杞(Lycium chinense),其在光合色素、质膜透性、渗透调节物质和抗氧化酶活性方面的响应有所不同[70];荒漠藓类植物在干旱和冻融条件下表现出强大的渗透调节能力和抗氧化防御体系,以应对细胞水分丧失和ROS的双重胁迫[71];裸果木(Gymnocarpos przewalskii)则通过调节不同生长季节的抗氧化酶活性和渗透调节物质含量来适应环境中水分条件的变化[72].此外,一些荒漠植物如霸王(Zygophyllum xanthoxylum)和荆芥(Nepeta cataria)在盐胁迫下通过细胞离子平衡调控和非酶抗氧化物积累来提升其抗逆性[73-74]. ...
Mechanical wounding improves salt tolerance by maintaining root ion homeostasis in a desert shrub
1
2024
... 渗透调节是荒漠植物维持细胞水分平衡和渗透稳态的重要机制,尤其在长期干旱和盐胁迫环境下表现得尤为显著[65].非酶抗氧化物质在荒漠植物中广泛存在,这些物质在清除自由基的同时,还兼具渗透调节作用,增强植物对环境胁迫的综合抵御能力[53,66].荒漠植物通过积累脯氨酸、可溶性糖、甜菜碱和无机离子等渗透调节物质降低渗透势,保持细胞内水分含量,进而确保代谢活动的正常进行[67-68].渗透调节物质在保护细胞膜、蛋白质和酶类活性中发挥重要作用.例如红砂在干旱胁迫下大量积累脯氨酸和可溶性糖,以维持细胞渗透势并保护细胞膜的完整性[69].柽柳在盐胁迫下通过积累可溶性糖和脯氨酸,不仅增强了抗氧化能力,还显著提升了对盐碱环境的适应性[63].无机离子的选择性吸收与转运也是荒漠植物应对盐胁迫的重要策略,通过调节钠、钾离子的比值,降低细胞毒性并保持渗透稳态[53].不同荒漠植物的适应策略存在显著差异.研究表明,通过不同渗透势的盐溶液处理两种耐盐性不同的枸杞(Lycium chinense),其在光合色素、质膜透性、渗透调节物质和抗氧化酶活性方面的响应有所不同[70];荒漠藓类植物在干旱和冻融条件下表现出强大的渗透调节能力和抗氧化防御体系,以应对细胞水分丧失和ROS的双重胁迫[71];裸果木(Gymnocarpos przewalskii)则通过调节不同生长季节的抗氧化酶活性和渗透调节物质含量来适应环境中水分条件的变化[72].此外,一些荒漠植物如霸王(Zygophyllum xanthoxylum)和荆芥(Nepeta cataria)在盐胁迫下通过细胞离子平衡调控和非酶抗氧化物积累来提升其抗逆性[73-74]. ...
红砂对CO2浓度升高及降水变化的生理生长响应
1
2017
... 渗透调节是荒漠植物维持细胞水分平衡和渗透稳态的重要机制,尤其在长期干旱和盐胁迫环境下表现得尤为显著[65].非酶抗氧化物质在荒漠植物中广泛存在,这些物质在清除自由基的同时,还兼具渗透调节作用,增强植物对环境胁迫的综合抵御能力[53,66].荒漠植物通过积累脯氨酸、可溶性糖、甜菜碱和无机离子等渗透调节物质降低渗透势,保持细胞内水分含量,进而确保代谢活动的正常进行[67-68].渗透调节物质在保护细胞膜、蛋白质和酶类活性中发挥重要作用.例如红砂在干旱胁迫下大量积累脯氨酸和可溶性糖,以维持细胞渗透势并保护细胞膜的完整性[69].柽柳在盐胁迫下通过积累可溶性糖和脯氨酸,不仅增强了抗氧化能力,还显著提升了对盐碱环境的适应性[63].无机离子的选择性吸收与转运也是荒漠植物应对盐胁迫的重要策略,通过调节钠、钾离子的比值,降低细胞毒性并保持渗透稳态[53].不同荒漠植物的适应策略存在显著差异.研究表明,通过不同渗透势的盐溶液处理两种耐盐性不同的枸杞(Lycium chinense),其在光合色素、质膜透性、渗透调节物质和抗氧化酶活性方面的响应有所不同[70];荒漠藓类植物在干旱和冻融条件下表现出强大的渗透调节能力和抗氧化防御体系,以应对细胞水分丧失和ROS的双重胁迫[71];裸果木(Gymnocarpos przewalskii)则通过调节不同生长季节的抗氧化酶活性和渗透调节物质含量来适应环境中水分条件的变化[72].此外,一些荒漠植物如霸王(Zygophyllum xanthoxylum)和荆芥(Nepeta cataria)在盐胁迫下通过细胞离子平衡调控和非酶抗氧化物积累来提升其抗逆性[73-74]. ...
不同外源缓解物质对混合盐胁迫下两种枸杞生理特性的影响
1
2020
... 渗透调节是荒漠植物维持细胞水分平衡和渗透稳态的重要机制,尤其在长期干旱和盐胁迫环境下表现得尤为显著[65].非酶抗氧化物质在荒漠植物中广泛存在,这些物质在清除自由基的同时,还兼具渗透调节作用,增强植物对环境胁迫的综合抵御能力[53,66].荒漠植物通过积累脯氨酸、可溶性糖、甜菜碱和无机离子等渗透调节物质降低渗透势,保持细胞内水分含量,进而确保代谢活动的正常进行[67-68].渗透调节物质在保护细胞膜、蛋白质和酶类活性中发挥重要作用.例如红砂在干旱胁迫下大量积累脯氨酸和可溶性糖,以维持细胞渗透势并保护细胞膜的完整性[69].柽柳在盐胁迫下通过积累可溶性糖和脯氨酸,不仅增强了抗氧化能力,还显著提升了对盐碱环境的适应性[63].无机离子的选择性吸收与转运也是荒漠植物应对盐胁迫的重要策略,通过调节钠、钾离子的比值,降低细胞毒性并保持渗透稳态[53].不同荒漠植物的适应策略存在显著差异.研究表明,通过不同渗透势的盐溶液处理两种耐盐性不同的枸杞(Lycium chinense),其在光合色素、质膜透性、渗透调节物质和抗氧化酶活性方面的响应有所不同[70];荒漠藓类植物在干旱和冻融条件下表现出强大的渗透调节能力和抗氧化防御体系,以应对细胞水分丧失和ROS的双重胁迫[71];裸果木(Gymnocarpos przewalskii)则通过调节不同生长季节的抗氧化酶活性和渗透调节物质含量来适应环境中水分条件的变化[72].此外,一些荒漠植物如霸王(Zygophyllum xanthoxylum)和荆芥(Nepeta cataria)在盐胁迫下通过细胞离子平衡调控和非酶抗氧化物积累来提升其抗逆性[73-74]. ...
冻融过程对荒漠区不同微生境下齿肋赤藓渗透调节物含量和抗氧化酶活力的影响
1
2015
... 渗透调节是荒漠植物维持细胞水分平衡和渗透稳态的重要机制,尤其在长期干旱和盐胁迫环境下表现得尤为显著[65].非酶抗氧化物质在荒漠植物中广泛存在,这些物质在清除自由基的同时,还兼具渗透调节作用,增强植物对环境胁迫的综合抵御能力[53,66].荒漠植物通过积累脯氨酸、可溶性糖、甜菜碱和无机离子等渗透调节物质降低渗透势,保持细胞内水分含量,进而确保代谢活动的正常进行[67-68].渗透调节物质在保护细胞膜、蛋白质和酶类活性中发挥重要作用.例如红砂在干旱胁迫下大量积累脯氨酸和可溶性糖,以维持细胞渗透势并保护细胞膜的完整性[69].柽柳在盐胁迫下通过积累可溶性糖和脯氨酸,不仅增强了抗氧化能力,还显著提升了对盐碱环境的适应性[63].无机离子的选择性吸收与转运也是荒漠植物应对盐胁迫的重要策略,通过调节钠、钾离子的比值,降低细胞毒性并保持渗透稳态[53].不同荒漠植物的适应策略存在显著差异.研究表明,通过不同渗透势的盐溶液处理两种耐盐性不同的枸杞(Lycium chinense),其在光合色素、质膜透性、渗透调节物质和抗氧化酶活性方面的响应有所不同[70];荒漠藓类植物在干旱和冻融条件下表现出强大的渗透调节能力和抗氧化防御体系,以应对细胞水分丧失和ROS的双重胁迫[71];裸果木(Gymnocarpos przewalskii)则通过调节不同生长季节的抗氧化酶活性和渗透调节物质含量来适应环境中水分条件的变化[72].此外,一些荒漠植物如霸王(Zygophyllum xanthoxylum)和荆芥(Nepeta cataria)在盐胁迫下通过细胞离子平衡调控和非酶抗氧化物积累来提升其抗逆性[73-74]. ...
裸果木渗透调节物质和抗氧化酶活性对干旱的响应
1
2020
... 渗透调节是荒漠植物维持细胞水分平衡和渗透稳态的重要机制,尤其在长期干旱和盐胁迫环境下表现得尤为显著[65].非酶抗氧化物质在荒漠植物中广泛存在,这些物质在清除自由基的同时,还兼具渗透调节作用,增强植物对环境胁迫的综合抵御能力[53,66].荒漠植物通过积累脯氨酸、可溶性糖、甜菜碱和无机离子等渗透调节物质降低渗透势,保持细胞内水分含量,进而确保代谢活动的正常进行[67-68].渗透调节物质在保护细胞膜、蛋白质和酶类活性中发挥重要作用.例如红砂在干旱胁迫下大量积累脯氨酸和可溶性糖,以维持细胞渗透势并保护细胞膜的完整性[69].柽柳在盐胁迫下通过积累可溶性糖和脯氨酸,不仅增强了抗氧化能力,还显著提升了对盐碱环境的适应性[63].无机离子的选择性吸收与转运也是荒漠植物应对盐胁迫的重要策略,通过调节钠、钾离子的比值,降低细胞毒性并保持渗透稳态[53].不同荒漠植物的适应策略存在显著差异.研究表明,通过不同渗透势的盐溶液处理两种耐盐性不同的枸杞(Lycium chinense),其在光合色素、质膜透性、渗透调节物质和抗氧化酶活性方面的响应有所不同[70];荒漠藓类植物在干旱和冻融条件下表现出强大的渗透调节能力和抗氧化防御体系,以应对细胞水分丧失和ROS的双重胁迫[71];裸果木(Gymnocarpos przewalskii)则通过调节不同生长季节的抗氧化酶活性和渗透调节物质含量来适应环境中水分条件的变化[72].此外,一些荒漠植物如霸王(Zygophyllum xanthoxylum)和荆芥(Nepeta cataria)在盐胁迫下通过细胞离子平衡调控和非酶抗氧化物积累来提升其抗逆性[73-74]. ...
Na+在霸王适应渗透胁迫中的生理作用
1
2011
... 渗透调节是荒漠植物维持细胞水分平衡和渗透稳态的重要机制,尤其在长期干旱和盐胁迫环境下表现得尤为显著[65].非酶抗氧化物质在荒漠植物中广泛存在,这些物质在清除自由基的同时,还兼具渗透调节作用,增强植物对环境胁迫的综合抵御能力[53,66].荒漠植物通过积累脯氨酸、可溶性糖、甜菜碱和无机离子等渗透调节物质降低渗透势,保持细胞内水分含量,进而确保代谢活动的正常进行[67-68].渗透调节物质在保护细胞膜、蛋白质和酶类活性中发挥重要作用.例如红砂在干旱胁迫下大量积累脯氨酸和可溶性糖,以维持细胞渗透势并保护细胞膜的完整性[69].柽柳在盐胁迫下通过积累可溶性糖和脯氨酸,不仅增强了抗氧化能力,还显著提升了对盐碱环境的适应性[63].无机离子的选择性吸收与转运也是荒漠植物应对盐胁迫的重要策略,通过调节钠、钾离子的比值,降低细胞毒性并保持渗透稳态[53].不同荒漠植物的适应策略存在显著差异.研究表明,通过不同渗透势的盐溶液处理两种耐盐性不同的枸杞(Lycium chinense),其在光合色素、质膜透性、渗透调节物质和抗氧化酶活性方面的响应有所不同[70];荒漠藓类植物在干旱和冻融条件下表现出强大的渗透调节能力和抗氧化防御体系,以应对细胞水分丧失和ROS的双重胁迫[71];裸果木(Gymnocarpos przewalskii)则通过调节不同生长季节的抗氧化酶活性和渗透调节物质含量来适应环境中水分条件的变化[72].此外,一些荒漠植物如霸王(Zygophyllum xanthoxylum)和荆芥(Nepeta cataria)在盐胁迫下通过细胞离子平衡调控和非酶抗氧化物积累来提升其抗逆性[73-74]. ...
荆芥的生长、渗透调节和抗氧化能力对干旱胁迫的响应
1
2020
... 渗透调节是荒漠植物维持细胞水分平衡和渗透稳态的重要机制,尤其在长期干旱和盐胁迫环境下表现得尤为显著[65].非酶抗氧化物质在荒漠植物中广泛存在,这些物质在清除自由基的同时,还兼具渗透调节作用,增强植物对环境胁迫的综合抵御能力[53,66].荒漠植物通过积累脯氨酸、可溶性糖、甜菜碱和无机离子等渗透调节物质降低渗透势,保持细胞内水分含量,进而确保代谢活动的正常进行[67-68].渗透调节物质在保护细胞膜、蛋白质和酶类活性中发挥重要作用.例如红砂在干旱胁迫下大量积累脯氨酸和可溶性糖,以维持细胞渗透势并保护细胞膜的完整性[69].柽柳在盐胁迫下通过积累可溶性糖和脯氨酸,不仅增强了抗氧化能力,还显著提升了对盐碱环境的适应性[63].无机离子的选择性吸收与转运也是荒漠植物应对盐胁迫的重要策略,通过调节钠、钾离子的比值,降低细胞毒性并保持渗透稳态[53].不同荒漠植物的适应策略存在显著差异.研究表明,通过不同渗透势的盐溶液处理两种耐盐性不同的枸杞(Lycium chinense),其在光合色素、质膜透性、渗透调节物质和抗氧化酶活性方面的响应有所不同[70];荒漠藓类植物在干旱和冻融条件下表现出强大的渗透调节能力和抗氧化防御体系,以应对细胞水分丧失和ROS的双重胁迫[71];裸果木(Gymnocarpos przewalskii)则通过调节不同生长季节的抗氧化酶活性和渗透调节物质含量来适应环境中水分条件的变化[72].此外,一些荒漠植物如霸王(Zygophyllum xanthoxylum)和荆芥(Nepeta cataria)在盐胁迫下通过细胞离子平衡调控和非酶抗氧化物积累来提升其抗逆性[73-74]. ...
濒危植物小沙冬青研究进展
1
2019
... 近年来,随着高通量测序、定量代谢组学等多组学技术的发展,荒漠植物在干旱、高温、盐碱及养分匮乏等逆境下的应答机制逐步得到揭示.在代谢调控层面,荒漠植物展现出独特的次生代谢物合成策略.定量代谢组学分析表明,脯氨酸、可溶性糖、甜菜碱等渗透调节物质的积累可维持细胞水势平衡,可维持细胞水势平衡,而黄酮类、酚类化合物等次生代谢物通过清除活性氧(ROS)缓解氧化损伤[75].柠条锦鸡儿通过增强能量代谢和氨基酸代谢等途径应对干旱胁迫,且不同器官采用差异化策略以应对环境挑战[76].微生物-植物互作机制研究荒漠植物耐逆性中的作用日益受到关注.研究表明,根际微生物可调控植物内源激素(如脱落酸和茉莉酸)及碳氮代谢通路,从而增强植物对干旱胁迫的适应能力[77].荒漠植物的内生菌群通过调节宿主植物的转录组模式,影响根系生长和胁迫应答基因的表达,从而提高植物对干旱和盐胁迫的耐受性[78].转录组学研究揭示了荒漠植物的全基因组水平的动态调控机制.例如,研究表明,沙漠耐逆木本植物在干旱和低温胁迫下,水通道蛋白、抗氧化酶、脱落酸信号相关基因及渗透保护蛋白的表达显著上调,表明这些植物主要通过调控水分运输、活性氧清除及ABA信号通路来适应环境胁迫[79].同时研究表明,多个关键转录因子家族如NAC、DREB、bZIP和MYB等,通过调控胁迫应答基因的表达,优化水分管理、抗氧化能力及渗透调节[80].红砂(Reaumuria songarica)类黄酮生物合成关键酶F3H基因在UV-B辐射和干旱胁迫下的调控作用显著,通过增强抗氧化功能,提升植物的适应能力[81].此外,盐生植物盐芥(Eutrema salsugineum)在高盐胁迫下的程序性死亡(PCD)信号通路中,液泡自噬发挥关键作用,抑制ERK1/2-like通路可加剧PCD,而抑制类半胱氨酸蛋白酶3则显著降低细胞死亡,从而揭示了植物在盐胁迫下的调控机制[82].此外,非编码RNA(ncRNA)和表观遗传调控如DNA甲基化及染色质重塑等也在荒漠植物对环境胁迫的适应性调控中发挥重要作用.刘建凤等[83]发现,特定miRNA通过靶向ROS信号通路相关基因的mRNA,调节抗氧化酶活性,增强植物耐逆性.近期多组学分析表明,宽叶中与生长素(IAA)和油菜素内酯(BR)信号传导相关的基因表现出低水平的DNA甲基化和高染色质可及性状态,这有助于这些基因的表达上调,从而促进宽叶的形成和功能分化,使其能够更好地适应干旱环境[84]. ...
沙蓬栽培品系筛选及重要性状初步研究
1
2019
... 近年来,随着高通量测序、定量代谢组学等多组学技术的发展,荒漠植物在干旱、高温、盐碱及养分匮乏等逆境下的应答机制逐步得到揭示.在代谢调控层面,荒漠植物展现出独特的次生代谢物合成策略.定量代谢组学分析表明,脯氨酸、可溶性糖、甜菜碱等渗透调节物质的积累可维持细胞水势平衡,可维持细胞水势平衡,而黄酮类、酚类化合物等次生代谢物通过清除活性氧(ROS)缓解氧化损伤[75].柠条锦鸡儿通过增强能量代谢和氨基酸代谢等途径应对干旱胁迫,且不同器官采用差异化策略以应对环境挑战[76].微生物-植物互作机制研究荒漠植物耐逆性中的作用日益受到关注.研究表明,根际微生物可调控植物内源激素(如脱落酸和茉莉酸)及碳氮代谢通路,从而增强植物对干旱胁迫的适应能力[77].荒漠植物的内生菌群通过调节宿主植物的转录组模式,影响根系生长和胁迫应答基因的表达,从而提高植物对干旱和盐胁迫的耐受性[78].转录组学研究揭示了荒漠植物的全基因组水平的动态调控机制.例如,研究表明,沙漠耐逆木本植物在干旱和低温胁迫下,水通道蛋白、抗氧化酶、脱落酸信号相关基因及渗透保护蛋白的表达显著上调,表明这些植物主要通过调控水分运输、活性氧清除及ABA信号通路来适应环境胁迫[79].同时研究表明,多个关键转录因子家族如NAC、DREB、bZIP和MYB等,通过调控胁迫应答基因的表达,优化水分管理、抗氧化能力及渗透调节[80].红砂(Reaumuria songarica)类黄酮生物合成关键酶F3H基因在UV-B辐射和干旱胁迫下的调控作用显著,通过增强抗氧化功能,提升植物的适应能力[81].此外,盐生植物盐芥(Eutrema salsugineum)在高盐胁迫下的程序性死亡(PCD)信号通路中,液泡自噬发挥关键作用,抑制ERK1/2-like通路可加剧PCD,而抑制类半胱氨酸蛋白酶3则显著降低细胞死亡,从而揭示了植物在盐胁迫下的调控机制[82].此外,非编码RNA(ncRNA)和表观遗传调控如DNA甲基化及染色质重塑等也在荒漠植物对环境胁迫的适应性调控中发挥重要作用.刘建凤等[83]发现,特定miRNA通过靶向ROS信号通路相关基因的mRNA,调节抗氧化酶活性,增强植物耐逆性.近期多组学分析表明,宽叶中与生长素(IAA)和油菜素内酯(BR)信号传导相关的基因表现出低水平的DNA甲基化和高染色质可及性状态,这有助于这些基因的表达上调,从而促进宽叶的形成和功能分化,使其能够更好地适应干旱环境[84]. ...
干旱胁迫下的植物根系-微生物互作体系及其应用
1
2020
... 近年来,随着高通量测序、定量代谢组学等多组学技术的发展,荒漠植物在干旱、高温、盐碱及养分匮乏等逆境下的应答机制逐步得到揭示.在代谢调控层面,荒漠植物展现出独特的次生代谢物合成策略.定量代谢组学分析表明,脯氨酸、可溶性糖、甜菜碱等渗透调节物质的积累可维持细胞水势平衡,可维持细胞水势平衡,而黄酮类、酚类化合物等次生代谢物通过清除活性氧(ROS)缓解氧化损伤[75].柠条锦鸡儿通过增强能量代谢和氨基酸代谢等途径应对干旱胁迫,且不同器官采用差异化策略以应对环境挑战[76].微生物-植物互作机制研究荒漠植物耐逆性中的作用日益受到关注.研究表明,根际微生物可调控植物内源激素(如脱落酸和茉莉酸)及碳氮代谢通路,从而增强植物对干旱胁迫的适应能力[77].荒漠植物的内生菌群通过调节宿主植物的转录组模式,影响根系生长和胁迫应答基因的表达,从而提高植物对干旱和盐胁迫的耐受性[78].转录组学研究揭示了荒漠植物的全基因组水平的动态调控机制.例如,研究表明,沙漠耐逆木本植物在干旱和低温胁迫下,水通道蛋白、抗氧化酶、脱落酸信号相关基因及渗透保护蛋白的表达显著上调,表明这些植物主要通过调控水分运输、活性氧清除及ABA信号通路来适应环境胁迫[79].同时研究表明,多个关键转录因子家族如NAC、DREB、bZIP和MYB等,通过调控胁迫应答基因的表达,优化水分管理、抗氧化能力及渗透调节[80].红砂(Reaumuria songarica)类黄酮生物合成关键酶F3H基因在UV-B辐射和干旱胁迫下的调控作用显著,通过增强抗氧化功能,提升植物的适应能力[81].此外,盐生植物盐芥(Eutrema salsugineum)在高盐胁迫下的程序性死亡(PCD)信号通路中,液泡自噬发挥关键作用,抑制ERK1/2-like通路可加剧PCD,而抑制类半胱氨酸蛋白酶3则显著降低细胞死亡,从而揭示了植物在盐胁迫下的调控机制[82].此外,非编码RNA(ncRNA)和表观遗传调控如DNA甲基化及染色质重塑等也在荒漠植物对环境胁迫的适应性调控中发挥重要作用.刘建凤等[83]发现,特定miRNA通过靶向ROS信号通路相关基因的mRNA,调节抗氧化酶活性,增强植物耐逆性.近期多组学分析表明,宽叶中与生长素(IAA)和油菜素内酯(BR)信号传导相关的基因表现出低水平的DNA甲基化和高染色质可及性状态,这有助于这些基因的表达上调,从而促进宽叶的形成和功能分化,使其能够更好地适应干旱环境[84]. ...
荒漠植物内生菌多样性及其增强农作物抗旱和耐盐性的研究进展
1
2022
... 近年来,随着高通量测序、定量代谢组学等多组学技术的发展,荒漠植物在干旱、高温、盐碱及养分匮乏等逆境下的应答机制逐步得到揭示.在代谢调控层面,荒漠植物展现出独特的次生代谢物合成策略.定量代谢组学分析表明,脯氨酸、可溶性糖、甜菜碱等渗透调节物质的积累可维持细胞水势平衡,可维持细胞水势平衡,而黄酮类、酚类化合物等次生代谢物通过清除活性氧(ROS)缓解氧化损伤[75].柠条锦鸡儿通过增强能量代谢和氨基酸代谢等途径应对干旱胁迫,且不同器官采用差异化策略以应对环境挑战[76].微生物-植物互作机制研究荒漠植物耐逆性中的作用日益受到关注.研究表明,根际微生物可调控植物内源激素(如脱落酸和茉莉酸)及碳氮代谢通路,从而增强植物对干旱胁迫的适应能力[77].荒漠植物的内生菌群通过调节宿主植物的转录组模式,影响根系生长和胁迫应答基因的表达,从而提高植物对干旱和盐胁迫的耐受性[78].转录组学研究揭示了荒漠植物的全基因组水平的动态调控机制.例如,研究表明,沙漠耐逆木本植物在干旱和低温胁迫下,水通道蛋白、抗氧化酶、脱落酸信号相关基因及渗透保护蛋白的表达显著上调,表明这些植物主要通过调控水分运输、活性氧清除及ABA信号通路来适应环境胁迫[79].同时研究表明,多个关键转录因子家族如NAC、DREB、bZIP和MYB等,通过调控胁迫应答基因的表达,优化水分管理、抗氧化能力及渗透调节[80].红砂(Reaumuria songarica)类黄酮生物合成关键酶F3H基因在UV-B辐射和干旱胁迫下的调控作用显著,通过增强抗氧化功能,提升植物的适应能力[81].此外,盐生植物盐芥(Eutrema salsugineum)在高盐胁迫下的程序性死亡(PCD)信号通路中,液泡自噬发挥关键作用,抑制ERK1/2-like通路可加剧PCD,而抑制类半胱氨酸蛋白酶3则显著降低细胞死亡,从而揭示了植物在盐胁迫下的调控机制[82].此外,非编码RNA(ncRNA)和表观遗传调控如DNA甲基化及染色质重塑等也在荒漠植物对环境胁迫的适应性调控中发挥重要作用.刘建凤等[83]发现,特定miRNA通过靶向ROS信号通路相关基因的mRNA,调节抗氧化酶活性,增强植物耐逆性.近期多组学分析表明,宽叶中与生长素(IAA)和油菜素内酯(BR)信号传导相关的基因表现出低水平的DNA甲基化和高染色质可及性状态,这有助于这些基因的表达上调,从而促进宽叶的形成和功能分化,使其能够更好地适应干旱环境[84]. ...
Physiological,transcriptomic and metabolomic insights of three extremophyte woody species living in the multi-stress environment of the Atacama Desert
1
2024
... 近年来,随着高通量测序、定量代谢组学等多组学技术的发展,荒漠植物在干旱、高温、盐碱及养分匮乏等逆境下的应答机制逐步得到揭示.在代谢调控层面,荒漠植物展现出独特的次生代谢物合成策略.定量代谢组学分析表明,脯氨酸、可溶性糖、甜菜碱等渗透调节物质的积累可维持细胞水势平衡,可维持细胞水势平衡,而黄酮类、酚类化合物等次生代谢物通过清除活性氧(ROS)缓解氧化损伤[75].柠条锦鸡儿通过增强能量代谢和氨基酸代谢等途径应对干旱胁迫,且不同器官采用差异化策略以应对环境挑战[76].微生物-植物互作机制研究荒漠植物耐逆性中的作用日益受到关注.研究表明,根际微生物可调控植物内源激素(如脱落酸和茉莉酸)及碳氮代谢通路,从而增强植物对干旱胁迫的适应能力[77].荒漠植物的内生菌群通过调节宿主植物的转录组模式,影响根系生长和胁迫应答基因的表达,从而提高植物对干旱和盐胁迫的耐受性[78].转录组学研究揭示了荒漠植物的全基因组水平的动态调控机制.例如,研究表明,沙漠耐逆木本植物在干旱和低温胁迫下,水通道蛋白、抗氧化酶、脱落酸信号相关基因及渗透保护蛋白的表达显著上调,表明这些植物主要通过调控水分运输、活性氧清除及ABA信号通路来适应环境胁迫[79].同时研究表明,多个关键转录因子家族如NAC、DREB、bZIP和MYB等,通过调控胁迫应答基因的表达,优化水分管理、抗氧化能力及渗透调节[80].红砂(Reaumuria songarica)类黄酮生物合成关键酶F3H基因在UV-B辐射和干旱胁迫下的调控作用显著,通过增强抗氧化功能,提升植物的适应能力[81].此外,盐生植物盐芥(Eutrema salsugineum)在高盐胁迫下的程序性死亡(PCD)信号通路中,液泡自噬发挥关键作用,抑制ERK1/2-like通路可加剧PCD,而抑制类半胱氨酸蛋白酶3则显著降低细胞死亡,从而揭示了植物在盐胁迫下的调控机制[82].此外,非编码RNA(ncRNA)和表观遗传调控如DNA甲基化及染色质重塑等也在荒漠植物对环境胁迫的适应性调控中发挥重要作用.刘建凤等[83]发现,特定miRNA通过靶向ROS信号通路相关基因的mRNA,调节抗氧化酶活性,增强植物耐逆性.近期多组学分析表明,宽叶中与生长素(IAA)和油菜素内酯(BR)信号传导相关的基因表现出低水平的DNA甲基化和高染色质可及性状态,这有助于这些基因的表达上调,从而促进宽叶的形成和功能分化,使其能够更好地适应干旱环境[84]. ...
植物抗旱性鉴定评价方法及抗旱机制研究进展
1
2017
... 近年来,随着高通量测序、定量代谢组学等多组学技术的发展,荒漠植物在干旱、高温、盐碱及养分匮乏等逆境下的应答机制逐步得到揭示.在代谢调控层面,荒漠植物展现出独特的次生代谢物合成策略.定量代谢组学分析表明,脯氨酸、可溶性糖、甜菜碱等渗透调节物质的积累可维持细胞水势平衡,可维持细胞水势平衡,而黄酮类、酚类化合物等次生代谢物通过清除活性氧(ROS)缓解氧化损伤[75].柠条锦鸡儿通过增强能量代谢和氨基酸代谢等途径应对干旱胁迫,且不同器官采用差异化策略以应对环境挑战[76].微生物-植物互作机制研究荒漠植物耐逆性中的作用日益受到关注.研究表明,根际微生物可调控植物内源激素(如脱落酸和茉莉酸)及碳氮代谢通路,从而增强植物对干旱胁迫的适应能力[77].荒漠植物的内生菌群通过调节宿主植物的转录组模式,影响根系生长和胁迫应答基因的表达,从而提高植物对干旱和盐胁迫的耐受性[78].转录组学研究揭示了荒漠植物的全基因组水平的动态调控机制.例如,研究表明,沙漠耐逆木本植物在干旱和低温胁迫下,水通道蛋白、抗氧化酶、脱落酸信号相关基因及渗透保护蛋白的表达显著上调,表明这些植物主要通过调控水分运输、活性氧清除及ABA信号通路来适应环境胁迫[79].同时研究表明,多个关键转录因子家族如NAC、DREB、bZIP和MYB等,通过调控胁迫应答基因的表达,优化水分管理、抗氧化能力及渗透调节[80].红砂(Reaumuria songarica)类黄酮生物合成关键酶F3H基因在UV-B辐射和干旱胁迫下的调控作用显著,通过增强抗氧化功能,提升植物的适应能力[81].此外,盐生植物盐芥(Eutrema salsugineum)在高盐胁迫下的程序性死亡(PCD)信号通路中,液泡自噬发挥关键作用,抑制ERK1/2-like通路可加剧PCD,而抑制类半胱氨酸蛋白酶3则显著降低细胞死亡,从而揭示了植物在盐胁迫下的调控机制[82].此外,非编码RNA(ncRNA)和表观遗传调控如DNA甲基化及染色质重塑等也在荒漠植物对环境胁迫的适应性调控中发挥重要作用.刘建凤等[83]发现,特定miRNA通过靶向ROS信号通路相关基因的mRNA,调节抗氧化酶活性,增强植物耐逆性.近期多组学分析表明,宽叶中与生长素(IAA)和油菜素内酯(BR)信号传导相关的基因表现出低水平的DNA甲基化和高染色质可及性状态,这有助于这些基因的表达上调,从而促进宽叶的形成和功能分化,使其能够更好地适应干旱环境[84]. ...
红砂叶片中基因表达及形态结构特征对干旱胁迫
1
2015
... 近年来,随着高通量测序、定量代谢组学等多组学技术的发展,荒漠植物在干旱、高温、盐碱及养分匮乏等逆境下的应答机制逐步得到揭示.在代谢调控层面,荒漠植物展现出独特的次生代谢物合成策略.定量代谢组学分析表明,脯氨酸、可溶性糖、甜菜碱等渗透调节物质的积累可维持细胞水势平衡,可维持细胞水势平衡,而黄酮类、酚类化合物等次生代谢物通过清除活性氧(ROS)缓解氧化损伤[75].柠条锦鸡儿通过增强能量代谢和氨基酸代谢等途径应对干旱胁迫,且不同器官采用差异化策略以应对环境挑战[76].微生物-植物互作机制研究荒漠植物耐逆性中的作用日益受到关注.研究表明,根际微生物可调控植物内源激素(如脱落酸和茉莉酸)及碳氮代谢通路,从而增强植物对干旱胁迫的适应能力[77].荒漠植物的内生菌群通过调节宿主植物的转录组模式,影响根系生长和胁迫应答基因的表达,从而提高植物对干旱和盐胁迫的耐受性[78].转录组学研究揭示了荒漠植物的全基因组水平的动态调控机制.例如,研究表明,沙漠耐逆木本植物在干旱和低温胁迫下,水通道蛋白、抗氧化酶、脱落酸信号相关基因及渗透保护蛋白的表达显著上调,表明这些植物主要通过调控水分运输、活性氧清除及ABA信号通路来适应环境胁迫[79].同时研究表明,多个关键转录因子家族如NAC、DREB、bZIP和MYB等,通过调控胁迫应答基因的表达,优化水分管理、抗氧化能力及渗透调节[80].红砂(Reaumuria songarica)类黄酮生物合成关键酶F3H基因在UV-B辐射和干旱胁迫下的调控作用显著,通过增强抗氧化功能,提升植物的适应能力[81].此外,盐生植物盐芥(Eutrema salsugineum)在高盐胁迫下的程序性死亡(PCD)信号通路中,液泡自噬发挥关键作用,抑制ERK1/2-like通路可加剧PCD,而抑制类半胱氨酸蛋白酶3则显著降低细胞死亡,从而揭示了植物在盐胁迫下的调控机制[82].此外,非编码RNA(ncRNA)和表观遗传调控如DNA甲基化及染色质重塑等也在荒漠植物对环境胁迫的适应性调控中发挥重要作用.刘建凤等[83]发现,特定miRNA通过靶向ROS信号通路相关基因的mRNA,调节抗氧化酶活性,增强植物耐逆性.近期多组学分析表明,宽叶中与生长素(IAA)和油菜素内酯(BR)信号传导相关的基因表现出低水平的DNA甲基化和高染色质可及性状态,这有助于这些基因的表达上调,从而促进宽叶的形成和功能分化,使其能够更好地适应干旱环境[84]. ...
盐胁迫下盐芥悬浮细胞程序性死亡的信号转导
1
2010
... 近年来,随着高通量测序、定量代谢组学等多组学技术的发展,荒漠植物在干旱、高温、盐碱及养分匮乏等逆境下的应答机制逐步得到揭示.在代谢调控层面,荒漠植物展现出独特的次生代谢物合成策略.定量代谢组学分析表明,脯氨酸、可溶性糖、甜菜碱等渗透调节物质的积累可维持细胞水势平衡,可维持细胞水势平衡,而黄酮类、酚类化合物等次生代谢物通过清除活性氧(ROS)缓解氧化损伤[75].柠条锦鸡儿通过增强能量代谢和氨基酸代谢等途径应对干旱胁迫,且不同器官采用差异化策略以应对环境挑战[76].微生物-植物互作机制研究荒漠植物耐逆性中的作用日益受到关注.研究表明,根际微生物可调控植物内源激素(如脱落酸和茉莉酸)及碳氮代谢通路,从而增强植物对干旱胁迫的适应能力[77].荒漠植物的内生菌群通过调节宿主植物的转录组模式,影响根系生长和胁迫应答基因的表达,从而提高植物对干旱和盐胁迫的耐受性[78].转录组学研究揭示了荒漠植物的全基因组水平的动态调控机制.例如,研究表明,沙漠耐逆木本植物在干旱和低温胁迫下,水通道蛋白、抗氧化酶、脱落酸信号相关基因及渗透保护蛋白的表达显著上调,表明这些植物主要通过调控水分运输、活性氧清除及ABA信号通路来适应环境胁迫[79].同时研究表明,多个关键转录因子家族如NAC、DREB、bZIP和MYB等,通过调控胁迫应答基因的表达,优化水分管理、抗氧化能力及渗透调节[80].红砂(Reaumuria songarica)类黄酮生物合成关键酶F3H基因在UV-B辐射和干旱胁迫下的调控作用显著,通过增强抗氧化功能,提升植物的适应能力[81].此外,盐生植物盐芥(Eutrema salsugineum)在高盐胁迫下的程序性死亡(PCD)信号通路中,液泡自噬发挥关键作用,抑制ERK1/2-like通路可加剧PCD,而抑制类半胱氨酸蛋白酶3则显著降低细胞死亡,从而揭示了植物在盐胁迫下的调控机制[82].此外,非编码RNA(ncRNA)和表观遗传调控如DNA甲基化及染色质重塑等也在荒漠植物对环境胁迫的适应性调控中发挥重要作用.刘建凤等[83]发现,特定miRNA通过靶向ROS信号通路相关基因的mRNA,调节抗氧化酶活性,增强植物耐逆性.近期多组学分析表明,宽叶中与生长素(IAA)和油菜素内酯(BR)信号传导相关的基因表现出低水平的DNA甲基化和高染色质可及性状态,这有助于这些基因的表达上调,从而促进宽叶的形成和功能分化,使其能够更好地适应干旱环境[84]. ...
沙棘抗虫性机制及其分子育种研究进展
1
2020
... 近年来,随着高通量测序、定量代谢组学等多组学技术的发展,荒漠植物在干旱、高温、盐碱及养分匮乏等逆境下的应答机制逐步得到揭示.在代谢调控层面,荒漠植物展现出独特的次生代谢物合成策略.定量代谢组学分析表明,脯氨酸、可溶性糖、甜菜碱等渗透调节物质的积累可维持细胞水势平衡,可维持细胞水势平衡,而黄酮类、酚类化合物等次生代谢物通过清除活性氧(ROS)缓解氧化损伤[75].柠条锦鸡儿通过增强能量代谢和氨基酸代谢等途径应对干旱胁迫,且不同器官采用差异化策略以应对环境挑战[76].微生物-植物互作机制研究荒漠植物耐逆性中的作用日益受到关注.研究表明,根际微生物可调控植物内源激素(如脱落酸和茉莉酸)及碳氮代谢通路,从而增强植物对干旱胁迫的适应能力[77].荒漠植物的内生菌群通过调节宿主植物的转录组模式,影响根系生长和胁迫应答基因的表达,从而提高植物对干旱和盐胁迫的耐受性[78].转录组学研究揭示了荒漠植物的全基因组水平的动态调控机制.例如,研究表明,沙漠耐逆木本植物在干旱和低温胁迫下,水通道蛋白、抗氧化酶、脱落酸信号相关基因及渗透保护蛋白的表达显著上调,表明这些植物主要通过调控水分运输、活性氧清除及ABA信号通路来适应环境胁迫[79].同时研究表明,多个关键转录因子家族如NAC、DREB、bZIP和MYB等,通过调控胁迫应答基因的表达,优化水分管理、抗氧化能力及渗透调节[80].红砂(Reaumuria songarica)类黄酮生物合成关键酶F3H基因在UV-B辐射和干旱胁迫下的调控作用显著,通过增强抗氧化功能,提升植物的适应能力[81].此外,盐生植物盐芥(Eutrema salsugineum)在高盐胁迫下的程序性死亡(PCD)信号通路中,液泡自噬发挥关键作用,抑制ERK1/2-like通路可加剧PCD,而抑制类半胱氨酸蛋白酶3则显著降低细胞死亡,从而揭示了植物在盐胁迫下的调控机制[82].此外,非编码RNA(ncRNA)和表观遗传调控如DNA甲基化及染色质重塑等也在荒漠植物对环境胁迫的适应性调控中发挥重要作用.刘建凤等[83]发现,特定miRNA通过靶向ROS信号通路相关基因的mRNA,调节抗氧化酶活性,增强植物耐逆性.近期多组学分析表明,宽叶中与生长素(IAA)和油菜素内酯(BR)信号传导相关的基因表现出低水平的DNA甲基化和高染色质可及性状态,这有助于这些基因的表达上调,从而促进宽叶的形成和功能分化,使其能够更好地适应干旱环境[84]. ...
Multi-omics analysis reveals spatiotemporal regulation and function of heteromorphic leaves in Populus
1
2023
... 近年来,随着高通量测序、定量代谢组学等多组学技术的发展,荒漠植物在干旱、高温、盐碱及养分匮乏等逆境下的应答机制逐步得到揭示.在代谢调控层面,荒漠植物展现出独特的次生代谢物合成策略.定量代谢组学分析表明,脯氨酸、可溶性糖、甜菜碱等渗透调节物质的积累可维持细胞水势平衡,可维持细胞水势平衡,而黄酮类、酚类化合物等次生代谢物通过清除活性氧(ROS)缓解氧化损伤[75].柠条锦鸡儿通过增强能量代谢和氨基酸代谢等途径应对干旱胁迫,且不同器官采用差异化策略以应对环境挑战[76].微生物-植物互作机制研究荒漠植物耐逆性中的作用日益受到关注.研究表明,根际微生物可调控植物内源激素(如脱落酸和茉莉酸)及碳氮代谢通路,从而增强植物对干旱胁迫的适应能力[77].荒漠植物的内生菌群通过调节宿主植物的转录组模式,影响根系生长和胁迫应答基因的表达,从而提高植物对干旱和盐胁迫的耐受性[78].转录组学研究揭示了荒漠植物的全基因组水平的动态调控机制.例如,研究表明,沙漠耐逆木本植物在干旱和低温胁迫下,水通道蛋白、抗氧化酶、脱落酸信号相关基因及渗透保护蛋白的表达显著上调,表明这些植物主要通过调控水分运输、活性氧清除及ABA信号通路来适应环境胁迫[79].同时研究表明,多个关键转录因子家族如NAC、DREB、bZIP和MYB等,通过调控胁迫应答基因的表达,优化水分管理、抗氧化能力及渗透调节[80].红砂(Reaumuria songarica)类黄酮生物合成关键酶F3H基因在UV-B辐射和干旱胁迫下的调控作用显著,通过增强抗氧化功能,提升植物的适应能力[81].此外,盐生植物盐芥(Eutrema salsugineum)在高盐胁迫下的程序性死亡(PCD)信号通路中,液泡自噬发挥关键作用,抑制ERK1/2-like通路可加剧PCD,而抑制类半胱氨酸蛋白酶3则显著降低细胞死亡,从而揭示了植物在盐胁迫下的调控机制[82].此外,非编码RNA(ncRNA)和表观遗传调控如DNA甲基化及染色质重塑等也在荒漠植物对环境胁迫的适应性调控中发挥重要作用.刘建凤等[83]发现,特定miRNA通过靶向ROS信号通路相关基因的mRNA,调节抗氧化酶活性,增强植物耐逆性.近期多组学分析表明,宽叶中与生长素(IAA)和油菜素内酯(BR)信号传导相关的基因表现出低水平的DNA甲基化和高染色质可及性状态,这有助于这些基因的表达上调,从而促进宽叶的形成和功能分化,使其能够更好地适应干旱环境[84]. ...




 甘公网安备 62010202000688号
甘公网安备 62010202000688号
{kind=link}
{kind=link}