On the essentials of drought in a changing climate
1
2020
... 全球气候变化背景下,干旱事件的频率、强度和持续时间均呈现显著增加趋势,对全球陆地生态系统构成了严重威胁[1].干旱事件通过破坏植物的水分平衡,抑制其光合作用与碳同化过程,导致树木生产力下降甚至枯死,使得森林从碳汇向碳源转变[2-3].径向变化是反映树木长期碳积累动态的关键指标,而水热条件作为影响树木生长的关键因子,直接决定了树木固碳能力的强弱[4].因此,研究树木径向变化对气候变化的响应,有助于深入理解树木在短期环境变化(干旱事件)中的适应机制,从而为森林资源有效管理提供科学依据. ...
Triggers of tree mortality under drought
1
2018
... 全球气候变化背景下,干旱事件的频率、强度和持续时间均呈现显著增加趋势,对全球陆地生态系统构成了严重威胁[1].干旱事件通过破坏植物的水分平衡,抑制其光合作用与碳同化过程,导致树木生产力下降甚至枯死,使得森林从碳汇向碳源转变[2-3].径向变化是反映树木长期碳积累动态的关键指标,而水热条件作为影响树木生长的关键因子,直接决定了树木固碳能力的强弱[4].因此,研究树木径向变化对气候变化的响应,有助于深入理解树木在短期环境变化(干旱事件)中的适应机制,从而为森林资源有效管理提供科学依据. ...
Dendrometer and intra-annual tree growth:what kind of information can be inferred?
1
2007
... 全球气候变化背景下,干旱事件的频率、强度和持续时间均呈现显著增加趋势,对全球陆地生态系统构成了严重威胁[1].干旱事件通过破坏植物的水分平衡,抑制其光合作用与碳同化过程,导致树木生产力下降甚至枯死,使得森林从碳汇向碳源转变[2-3].径向变化是反映树木长期碳积累动态的关键指标,而水热条件作为影响树木生长的关键因子,直接决定了树木固碳能力的强弱[4].因此,研究树木径向变化对气候变化的响应,有助于深入理解树木在短期环境变化(干旱事件)中的适应机制,从而为森林资源有效管理提供科学依据. ...
Twentieth century redistribution in climatic drivers of global tree growth
1
2019
... 全球气候变化背景下,干旱事件的频率、强度和持续时间均呈现显著增加趋势,对全球陆地生态系统构成了严重威胁[1].干旱事件通过破坏植物的水分平衡,抑制其光合作用与碳同化过程,导致树木生产力下降甚至枯死,使得森林从碳汇向碳源转变[2-3].径向变化是反映树木长期碳积累动态的关键指标,而水热条件作为影响树木生长的关键因子,直接决定了树木固碳能力的强弱[4].因此,研究树木径向变化对气候变化的响应,有助于深入理解树木在短期环境变化(干旱事件)中的适应机制,从而为森林资源有效管理提供科学依据. ...
Normalized tree water deficit:an automated dendrometer signal to quantify drought stress in trees
1
2025
... 多数研究认为,干旱事件会直接导致树木茎干收缩[5-10],然而近期研究发现,部分树木在经历干旱事件后径向生长量并未出现显著下降[11].这表明,在干旱频率增加的背景下,树木可能正在调整其适应策略,从试图抵抗干旱转向适应干旱并在干旱后快速恢复,体现了其对环境变化的长期适应过程[12-13].为了深入揭示这一生理与生态过程,开展以小时或更高时间分辨率的树木径向变化连续监测至关重要.高分辨率的连续监测数据,不仅记录了形成层分裂与扩张引起的正向生长过程,还能同步捕捉到由茎干水分消耗与补给引起的逆向收缩过程[14],从而完整反映树木应对水分限制的动态生长过程,实现树木生长的昼夜节律和季节动态精细刻画,可为深入理解树木对干旱胁迫的适应机制提供重要支撑. ...
Capturing drought stress signals:the potential of dendrometers for monitoring tree water status
0
2024
Local neighborhood affects stem rehydration under drought:evidence from mixtures of European beech with two different conifers
0
2024
河西走廊荒漠植被归一化指数(NDVI)与降水量的关系
0
2025
黄土丘陵区辽东栎和刺槐树干径向生长与微变化季节动态特征
0
2023
针叶树形成层活动及径向生长监测研究进展
1
2024
... 多数研究认为,干旱事件会直接导致树木茎干收缩[5-10],然而近期研究发现,部分树木在经历干旱事件后径向生长量并未出现显著下降[11].这表明,在干旱频率增加的背景下,树木可能正在调整其适应策略,从试图抵抗干旱转向适应干旱并在干旱后快速恢复,体现了其对环境变化的长期适应过程[12-13].为了深入揭示这一生理与生态过程,开展以小时或更高时间分辨率的树木径向变化连续监测至关重要.高分辨率的连续监测数据,不仅记录了形成层分裂与扩张引起的正向生长过程,还能同步捕捉到由茎干水分消耗与补给引起的逆向收缩过程[14],从而完整反映树木应对水分限制的动态生长过程,实现树木生长的昼夜节律和季节动态精细刻画,可为深入理解树木对干旱胁迫的适应机制提供重要支撑. ...
The 2018 European heatwave led to stem dehydration but not to consistent growth reductions in forests
1
2022
... 多数研究认为,干旱事件会直接导致树木茎干收缩[5-10],然而近期研究发现,部分树木在经历干旱事件后径向生长量并未出现显著下降[11].这表明,在干旱频率增加的背景下,树木可能正在调整其适应策略,从试图抵抗干旱转向适应干旱并在干旱后快速恢复,体现了其对环境变化的长期适应过程[12-13].为了深入揭示这一生理与生态过程,开展以小时或更高时间分辨率的树木径向变化连续监测至关重要.高分辨率的连续监测数据,不仅记录了形成层分裂与扩张引起的正向生长过程,还能同步捕捉到由茎干水分消耗与补给引起的逆向收缩过程[14],从而完整反映树木应对水分限制的动态生长过程,实现树木生长的昼夜节律和季节动态精细刻画,可为深入理解树木对干旱胁迫的适应机制提供重要支撑. ...
Tree post-drought recovery:scenarios,regulatory mechanisms and ways to improve
1
2024
... 多数研究认为,干旱事件会直接导致树木茎干收缩[5-10],然而近期研究发现,部分树木在经历干旱事件后径向生长量并未出现显著下降[11].这表明,在干旱频率增加的背景下,树木可能正在调整其适应策略,从试图抵抗干旱转向适应干旱并在干旱后快速恢复,体现了其对环境变化的长期适应过程[12-13].为了深入揭示这一生理与生态过程,开展以小时或更高时间分辨率的树木径向变化连续监测至关重要.高分辨率的连续监测数据,不仅记录了形成层分裂与扩张引起的正向生长过程,还能同步捕捉到由茎干水分消耗与补给引起的逆向收缩过程[14],从而完整反映树木应对水分限制的动态生长过程,实现树木生长的昼夜节律和季节动态精细刻画,可为深入理解树木对干旱胁迫的适应机制提供重要支撑. ...
Growth and resilience responses of Scots pine to extreme droughts across Europe depend on predrought growth conditions
1
2020
... 多数研究认为,干旱事件会直接导致树木茎干收缩[5-10],然而近期研究发现,部分树木在经历干旱事件后径向生长量并未出现显著下降[11].这表明,在干旱频率增加的背景下,树木可能正在调整其适应策略,从试图抵抗干旱转向适应干旱并在干旱后快速恢复,体现了其对环境变化的长期适应过程[12-13].为了深入揭示这一生理与生态过程,开展以小时或更高时间分辨率的树木径向变化连续监测至关重要.高分辨率的连续监测数据,不仅记录了形成层分裂与扩张引起的正向生长过程,还能同步捕捉到由茎干水分消耗与补给引起的逆向收缩过程[14],从而完整反映树木应对水分限制的动态生长过程,实现树木生长的昼夜节律和季节动态精细刻画,可为深入理解树木对干旱胁迫的适应机制提供重要支撑. ...
Radial stem variations-a source of tree physiological information not fully exploited yet
1
2016
... 多数研究认为,干旱事件会直接导致树木茎干收缩[5-10],然而近期研究发现,部分树木在经历干旱事件后径向生长量并未出现显著下降[11].这表明,在干旱频率增加的背景下,树木可能正在调整其适应策略,从试图抵抗干旱转向适应干旱并在干旱后快速恢复,体现了其对环境变化的长期适应过程[12-13].为了深入揭示这一生理与生态过程,开展以小时或更高时间分辨率的树木径向变化连续监测至关重要.高分辨率的连续监测数据,不仅记录了形成层分裂与扩张引起的正向生长过程,还能同步捕捉到由茎干水分消耗与补给引起的逆向收缩过程[14],从而完整反映树木应对水分限制的动态生长过程,实现树木生长的昼夜节律和季节动态精细刻画,可为深入理解树木对干旱胁迫的适应机制提供重要支撑. ...
祁连山青海云杉物候表型的空间分异及其内在机制
1
2021
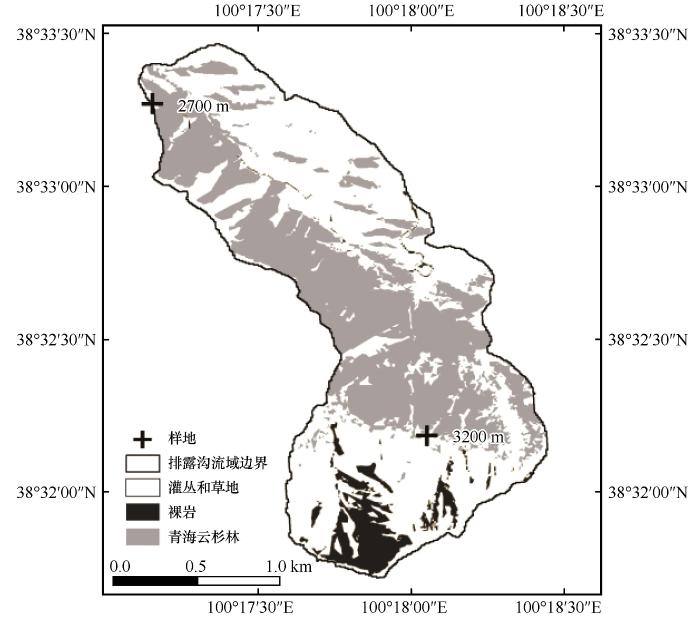
... 祁连山位于青藏高原东北部边缘,是中国西北干旱区山地生态系统的典型代表,在涵养水源、维持区域生态平衡等方面发挥着重要作用[15].近50 a来,该区域年均气温正以0.26 ℃/10a的速率升高,且极端高温天数的年际变率为0.79 d/10a[16-17],季节性干旱事件频发[18].青海云杉(Picea crassifolia)作为该区域的优势种,其稳定性对整个祁连山生态系统的功能发挥至关重要,但是其生长过程对气候变化高度敏感,易受到干旱事件影响[19].然而,目前关于祁连山青海云杉生长的研究多基于年际/年内尺度数据探讨其对环境因子的响应特征[20-22],缺少对干旱事件过后恢复及生长过程的研究,限制了对其环境敏感性及适应性的认识.因此,本研究于2014—2020年在祁连山排露沟流域的青海云杉上下林线3 200 m和2 700 m处分别布设样地,通过收集年内径向变化数据,系统分析青海云杉在干旱胁迫后的恢复和生长过程及其对环境因子的响应,研究结果将有助于深入认识祁连山青海云杉对干旱胁迫的适应策略,为青海云杉林的抚育管理提供依据. ...
近50年来祁连山区气候变化特征研究
1
2009
... 祁连山位于青藏高原东北部边缘,是中国西北干旱区山地生态系统的典型代表,在涵养水源、维持区域生态平衡等方面发挥着重要作用[15].近50 a来,该区域年均气温正以0.26 ℃/10a的速率升高,且极端高温天数的年际变率为0.79 d/10a[16-17],季节性干旱事件频发[18].青海云杉(Picea crassifolia)作为该区域的优势种,其稳定性对整个祁连山生态系统的功能发挥至关重要,但是其生长过程对气候变化高度敏感,易受到干旱事件影响[19].然而,目前关于祁连山青海云杉生长的研究多基于年际/年内尺度数据探讨其对环境因子的响应特征[20-22],缺少对干旱事件过后恢复及生长过程的研究,限制了对其环境敏感性及适应性的认识.因此,本研究于2014—2020年在祁连山排露沟流域的青海云杉上下林线3 200 m和2 700 m处分别布设样地,通过收集年内径向变化数据,系统分析青海云杉在干旱胁迫后的恢复和生长过程及其对环境因子的响应,研究结果将有助于深入认识祁连山青海云杉对干旱胁迫的适应策略,为青海云杉林的抚育管理提供依据. ...
近50 a来祁连山及河西走廊极端气候的时空变化研究
1
2012
... 祁连山位于青藏高原东北部边缘,是中国西北干旱区山地生态系统的典型代表,在涵养水源、维持区域生态平衡等方面发挥着重要作用[15].近50 a来,该区域年均气温正以0.26 ℃/10a的速率升高,且极端高温天数的年际变率为0.79 d/10a[16-17],季节性干旱事件频发[18].青海云杉(Picea crassifolia)作为该区域的优势种,其稳定性对整个祁连山生态系统的功能发挥至关重要,但是其生长过程对气候变化高度敏感,易受到干旱事件影响[19].然而,目前关于祁连山青海云杉生长的研究多基于年际/年内尺度数据探讨其对环境因子的响应特征[20-22],缺少对干旱事件过后恢复及生长过程的研究,限制了对其环境敏感性及适应性的认识.因此,本研究于2014—2020年在祁连山排露沟流域的青海云杉上下林线3 200 m和2 700 m处分别布设样地,通过收集年内径向变化数据,系统分析青海云杉在干旱胁迫后的恢复和生长过程及其对环境因子的响应,研究结果将有助于深入认识祁连山青海云杉对干旱胁迫的适应策略,为青海云杉林的抚育管理提供依据. ...
Spatiotemporal variations and driving mechanisms of flash droughts during 1981-2020 in the Qilian Mountains,China
1
2025
... 祁连山位于青藏高原东北部边缘,是中国西北干旱区山地生态系统的典型代表,在涵养水源、维持区域生态平衡等方面发挥着重要作用[15].近50 a来,该区域年均气温正以0.26 ℃/10a的速率升高,且极端高温天数的年际变率为0.79 d/10a[16-17],季节性干旱事件频发[18].青海云杉(Picea crassifolia)作为该区域的优势种,其稳定性对整个祁连山生态系统的功能发挥至关重要,但是其生长过程对气候变化高度敏感,易受到干旱事件影响[19].然而,目前关于祁连山青海云杉生长的研究多基于年际/年内尺度数据探讨其对环境因子的响应特征[20-22],缺少对干旱事件过后恢复及生长过程的研究,限制了对其环境敏感性及适应性的认识.因此,本研究于2014—2020年在祁连山排露沟流域的青海云杉上下林线3 200 m和2 700 m处分别布设样地,通过收集年内径向变化数据,系统分析青海云杉在干旱胁迫后的恢复和生长过程及其对环境因子的响应,研究结果将有助于深入认识祁连山青海云杉对干旱胁迫的适应策略,为青海云杉林的抚育管理提供依据. ...
Early monsoon failure and mid-summer dryness induces growth cessation of lower range margin Picea crassifolia
1
2018
... 祁连山位于青藏高原东北部边缘,是中国西北干旱区山地生态系统的典型代表,在涵养水源、维持区域生态平衡等方面发挥着重要作用[15].近50 a来,该区域年均气温正以0.26 ℃/10a的速率升高,且极端高温天数的年际变率为0.79 d/10a[16-17],季节性干旱事件频发[18].青海云杉(Picea crassifolia)作为该区域的优势种,其稳定性对整个祁连山生态系统的功能发挥至关重要,但是其生长过程对气候变化高度敏感,易受到干旱事件影响[19].然而,目前关于祁连山青海云杉生长的研究多基于年际/年内尺度数据探讨其对环境因子的响应特征[20-22],缺少对干旱事件过后恢复及生长过程的研究,限制了对其环境敏感性及适应性的认识.因此,本研究于2014—2020年在祁连山排露沟流域的青海云杉上下林线3 200 m和2 700 m处分别布设样地,通过收集年内径向变化数据,系统分析青海云杉在干旱胁迫后的恢复和生长过程及其对环境因子的响应,研究结果将有助于深入认识祁连山青海云杉对干旱胁迫的适应策略,为青海云杉林的抚育管理提供依据. ...
祁连山中东部青海云杉径向生长对气候变暖的响应差异
1
2022
... 祁连山位于青藏高原东北部边缘,是中国西北干旱区山地生态系统的典型代表,在涵养水源、维持区域生态平衡等方面发挥着重要作用[15].近50 a来,该区域年均气温正以0.26 ℃/10a的速率升高,且极端高温天数的年际变率为0.79 d/10a[16-17],季节性干旱事件频发[18].青海云杉(Picea crassifolia)作为该区域的优势种,其稳定性对整个祁连山生态系统的功能发挥至关重要,但是其生长过程对气候变化高度敏感,易受到干旱事件影响[19].然而,目前关于祁连山青海云杉生长的研究多基于年际/年内尺度数据探讨其对环境因子的响应特征[20-22],缺少对干旱事件过后恢复及生长过程的研究,限制了对其环境敏感性及适应性的认识.因此,本研究于2014—2020年在祁连山排露沟流域的青海云杉上下林线3 200 m和2 700 m处分别布设样地,通过收集年内径向变化数据,系统分析青海云杉在干旱胁迫后的恢复和生长过程及其对环境因子的响应,研究结果将有助于深入认识祁连山青海云杉对干旱胁迫的适应策略,为青海云杉林的抚育管理提供依据. ...
Non-structural carbohydrates constrain xylem formation of Picea crassifolia under drought year in the Qilian Mountains of Northwest China
0
2025
Mismatch between primary and secondary growth and its consequences on wood formation in Qinghai spruce
1
2023
... 祁连山位于青藏高原东北部边缘,是中国西北干旱区山地生态系统的典型代表,在涵养水源、维持区域生态平衡等方面发挥着重要作用[15].近50 a来,该区域年均气温正以0.26 ℃/10a的速率升高,且极端高温天数的年际变率为0.79 d/10a[16-17],季节性干旱事件频发[18].青海云杉(Picea crassifolia)作为该区域的优势种,其稳定性对整个祁连山生态系统的功能发挥至关重要,但是其生长过程对气候变化高度敏感,易受到干旱事件影响[19].然而,目前关于祁连山青海云杉生长的研究多基于年际/年内尺度数据探讨其对环境因子的响应特征[20-22],缺少对干旱事件过后恢复及生长过程的研究,限制了对其环境敏感性及适应性的认识.因此,本研究于2014—2020年在祁连山排露沟流域的青海云杉上下林线3 200 m和2 700 m处分别布设样地,通过收集年内径向变化数据,系统分析青海云杉在干旱胁迫后的恢复和生长过程及其对环境因子的响应,研究结果将有助于深入认识祁连山青海云杉对干旱胁迫的适应策略,为青海云杉林的抚育管理提供依据. ...
祁连山中部青海云杉径向变化与树干液流对环境要素的响应
1
2019
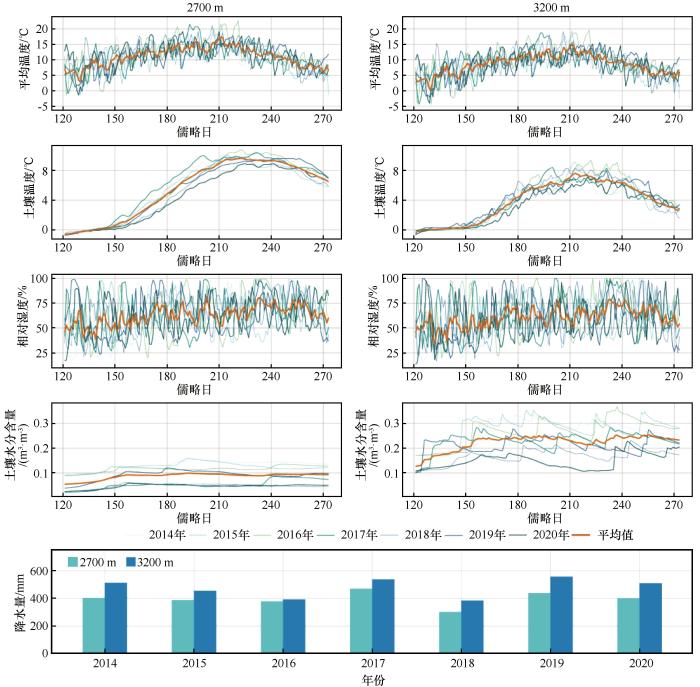
... 祁连山排露沟流域位于祁连山中段北坡,流域面积2.91 km2,属于典型的温带大陆性高寒半干旱气候,冬季寒冷,夏季炎热.根据研究站多年气象监测,年平均气温0.5 ℃,降水量368 mm,蒸散量1 052 mm,相对湿度60%,且降水主要在6—9月,占全年降水的80%左右[23].土壤类型以山地灰褐土和亚高山灌丛草甸土为主.植被类型呈现出典型的海拔梯度分布特征,其中,2 500~3 300 m属于山地森林草原植被带,物种多样性丰富,水热条件适宜灌丛和乔木生长,以寒生、旱生为主,主要有紫花针茅(Stipa purpurea)、甘青针茅(Stipa przewalskyi)等草本,金露梅(Potentilla fruticosa)、山生柳(Salix oritrepha)、小叶忍冬(Lonicera microphylla)等灌木;3 300~3 800 m属于亚高山灌丛草甸植被带,是高山径流的主要涵养区,植被主要有兔耳草(Lagotis glauca)、珠芽蓼(Bistorta vivipara)、高山龙胆(Gentiana algida)、高原毛茛(Ranunculus tanguticus)、西藏嵩草(Kobresia tibetica)、高山嵩草(Kobresia pygmaea)、青海杜鹃(Rhododendron qinghaiense)、紫花碎米荠(Cardamine tangutorum)等.青海云杉是该区域的主要建群种及优势种,多以纯林的形式呈斑块状或条带状格局分布在2 700~3 200 m的阴坡和半阴坡上. ...
Intra-annual stem radial growth of Qinghai spruce and its environmental drivers in the Qilian Mountains,northwestern China
2
2024
... 将Dendrometer监测数据转化为日尺度数据,用于后续分析.树木季节性生长过程一般呈“S”变化趋势,使用Gompertz函数拟合各样树的径向变化,通过对拟合函数求导计算生长速率,确定树木生长开始和结束时间以及最大生长速率发生的时间.本研究中认为生长速率大于4 μm时为生长阶段[24].拟合公式如下: ...
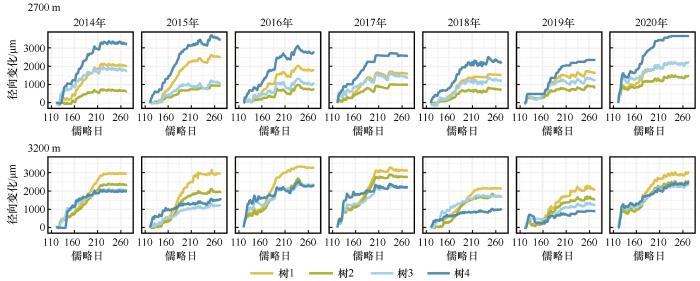
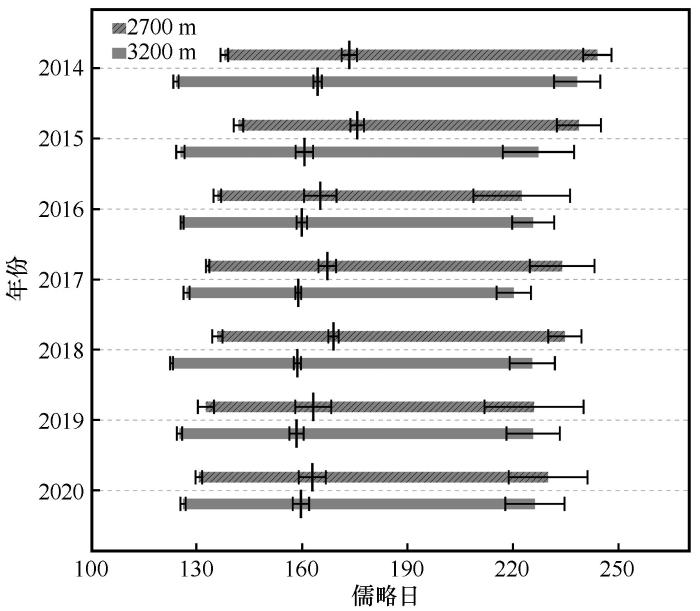
... 树木径向生长的季节性节律是其适应环境条件的综合体现,反映了树木生长开始和结束时间,进而决定了木质部的形成周期与生物量积累效率[27-30].在本研究中,海拔2 700 m和3 200 m处样树的径向生长开始时间存在显著差异,低海拔的生长开始时间普遍早于高海拔,这一现象与样地间的温度差异密切相关.已有研究表明,气温是驱动植物春季物候启动的主要气候因子,较高气温可打破休眠、启动形成层活动[31-32].对青藏高原和欧洲多种针叶树种的研究发现,当气温达到特定阈值时径向生长开始,且偶尔出现的高气温也会促进细胞分裂[24,33-35].一些研究表明,土壤温度对根系活动以及水分吸收有重要影响,从而促进树干细胞液泡膨压增加以及新形成管胞的膨胀[36].当根区土层温度升高时,冻土消融,土壤水分含量提高,根系可利用水分增多,此外,较高的土壤温度有助于夜间根系对水分的吸收,有利于维持树木内部水分平衡[37].因此,气温和土壤温度可能是影响青海云杉径向生长开始的关键因子. ...
Modeling tree water deficit from microclimate:an approach to quantifying drought stress
1
2005
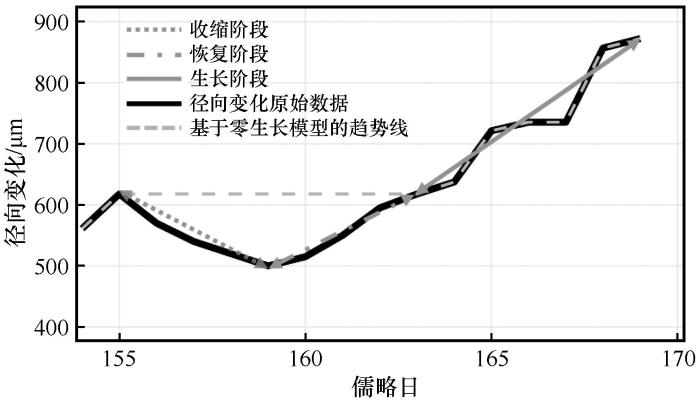
... 基于零生长模型,通过寻找历史时期生长最大值的方法确定树木径向生长趋势线,并将生长趋势线与实测径向变化曲线的差值定义为树木水分亏缺(Tree Water Deficit, TWD),用于反映树木体内水分状况[25-26].根据TWD的增减,将径向变化过程划分为3个生理阶段:收缩阶段(TWD>0,且持续增大),反映了干旱导致树木水分损失加剧的过程;恢复阶段(TWD>0,且持续减小),反映了干旱后树木水分得到补给的过程;生长阶段(TWD=0),反映了树木的生长过程(图2).将径向生长在收缩过程结束后,开始恢复直至回到收缩起始状态的过程定义为一次恢复事件,将随后径向生长至下一个下降拐点之前的过程定义为一次生长事件.根据阶段划分,结合各次事件的持续时间和变化量,使用最小二乘法计算各阶段、各事件的变化速率. ...
TreeNet-the biological drought and growth indicator network
1
2021
... 基于零生长模型,通过寻找历史时期生长最大值的方法确定树木径向生长趋势线,并将生长趋势线与实测径向变化曲线的差值定义为树木水分亏缺(Tree Water Deficit, TWD),用于反映树木体内水分状况[25-26].根据TWD的增减,将径向变化过程划分为3个生理阶段:收缩阶段(TWD>0,且持续增大),反映了干旱导致树木水分损失加剧的过程;恢复阶段(TWD>0,且持续减小),反映了干旱后树木水分得到补给的过程;生长阶段(TWD=0),反映了树木的生长过程(图2).将径向生长在收缩过程结束后,开始恢复直至回到收缩起始状态的过程定义为一次恢复事件,将随后径向生长至下一个下降拐点之前的过程定义为一次生长事件.根据阶段划分,结合各次事件的持续时间和变化量,使用最小二乘法计算各阶段、各事件的变化速率. ...
Number of growth days and not length of the growth period determines radial stem growth of temperate trees
2
2022
... 树木径向生长的季节性节律是其适应环境条件的综合体现,反映了树木生长开始和结束时间,进而决定了木质部的形成周期与生物量积累效率[27-30].在本研究中,海拔2 700 m和3 200 m处样树的径向生长开始时间存在显著差异,低海拔的生长开始时间普遍早于高海拔,这一现象与样地间的温度差异密切相关.已有研究表明,气温是驱动植物春季物候启动的主要气候因子,较高气温可打破休眠、启动形成层活动[31-32].对青藏高原和欧洲多种针叶树种的研究发现,当气温达到特定阈值时径向生长开始,且偶尔出现的高气温也会促进细胞分裂[24,33-35].一些研究表明,土壤温度对根系活动以及水分吸收有重要影响,从而促进树干细胞液泡膨压增加以及新形成管胞的膨胀[36].当根区土层温度升高时,冻土消融,土壤水分含量提高,根系可利用水分增多,此外,较高的土壤温度有助于夜间根系对水分的吸收,有利于维持树木内部水分平衡[37].因此,气温和土壤温度可能是影响青海云杉径向生长开始的关键因子. ...
... 目前,针对决定径向生长结束的因素尚未达成统一共识.多项研究表明,由于秋季温度的快速下降能够改变木质部导管的形态和晚材的形成,温度是影响针叶树生长结束的重要因子[31,44].另一些研究认为,即使在较高温度环境下,水分供给不足也会导致生长提前结束,这是由于降水不足的情况下高温会降低细胞的张力和水势[34-35].最新的研究表明,土壤氮含量升高一方面延缓了木质部细胞壁增厚的完成,另一方面又促进了细胞扩大过程的结束,最终导致生长季提前结束[30].在本研究中,各样树的生长结束时间存在较大差异,且在生长结束后土壤含水量和温度均适宜,降水也相对充足,这表明生长结束不仅受水分供给和温度的影响,还受土壤氮、植被遗传特性等因子的控制[27-28,30]. ...
Increased growing-season productivity drives earlier autumn leaf senescence in temperate trees
1
2020
... 目前,针对决定径向生长结束的因素尚未达成统一共识.多项研究表明,由于秋季温度的快速下降能够改变木质部导管的形态和晚材的形成,温度是影响针叶树生长结束的重要因子[31,44].另一些研究认为,即使在较高温度环境下,水分供给不足也会导致生长提前结束,这是由于降水不足的情况下高温会降低细胞的张力和水势[34-35].最新的研究表明,土壤氮含量升高一方面延缓了木质部细胞壁增厚的完成,另一方面又促进了细胞扩大过程的结束,最终导致生长季提前结束[30].在本研究中,各样树的生长结束时间存在较大差异,且在生长结束后土壤含水量和温度均适宜,降水也相对充足,这表明生长结束不仅受水分供给和温度的影响,还受土壤氮、植被遗传特性等因子的控制[27-28,30]. ...
Radial growth decline of white spruce (Picea glauca) during hot summers without drought:preliminary results from a study site south of a boreal forest border
0
2022
Soil nitrogen drives inverse acclimation of xylem growth cessation to rising temperature in Northern Hemisphere conifers
3
2025
... 树木径向生长的季节性节律是其适应环境条件的综合体现,反映了树木生长开始和结束时间,进而决定了木质部的形成周期与生物量积累效率[27-30].在本研究中,海拔2 700 m和3 200 m处样树的径向生长开始时间存在显著差异,低海拔的生长开始时间普遍早于高海拔,这一现象与样地间的温度差异密切相关.已有研究表明,气温是驱动植物春季物候启动的主要气候因子,较高气温可打破休眠、启动形成层活动[31-32].对青藏高原和欧洲多种针叶树种的研究发现,当气温达到特定阈值时径向生长开始,且偶尔出现的高气温也会促进细胞分裂[24,33-35].一些研究表明,土壤温度对根系活动以及水分吸收有重要影响,从而促进树干细胞液泡膨压增加以及新形成管胞的膨胀[36].当根区土层温度升高时,冻土消融,土壤水分含量提高,根系可利用水分增多,此外,较高的土壤温度有助于夜间根系对水分的吸收,有利于维持树木内部水分平衡[37].因此,气温和土壤温度可能是影响青海云杉径向生长开始的关键因子. ...
... 目前,针对决定径向生长结束的因素尚未达成统一共识.多项研究表明,由于秋季温度的快速下降能够改变木质部导管的形态和晚材的形成,温度是影响针叶树生长结束的重要因子[31,44].另一些研究认为,即使在较高温度环境下,水分供给不足也会导致生长提前结束,这是由于降水不足的情况下高温会降低细胞的张力和水势[34-35].最新的研究表明,土壤氮含量升高一方面延缓了木质部细胞壁增厚的完成,另一方面又促进了细胞扩大过程的结束,最终导致生长季提前结束[30].在本研究中,各样树的生长结束时间存在较大差异,且在生长结束后土壤含水量和温度均适宜,降水也相对充足,这表明生长结束不仅受水分供给和温度的影响,还受土壤氮、植被遗传特性等因子的控制[27-28,30]. ...
... ,30]. ...
Climate change and the regulation of wood formation in trees by temperature
2
2018
... 树木径向生长的季节性节律是其适应环境条件的综合体现,反映了树木生长开始和结束时间,进而决定了木质部的形成周期与生物量积累效率[27-30].在本研究中,海拔2 700 m和3 200 m处样树的径向生长开始时间存在显著差异,低海拔的生长开始时间普遍早于高海拔,这一现象与样地间的温度差异密切相关.已有研究表明,气温是驱动植物春季物候启动的主要气候因子,较高气温可打破休眠、启动形成层活动[31-32].对青藏高原和欧洲多种针叶树种的研究发现,当气温达到特定阈值时径向生长开始,且偶尔出现的高气温也会促进细胞分裂[24,33-35].一些研究表明,土壤温度对根系活动以及水分吸收有重要影响,从而促进树干细胞液泡膨压增加以及新形成管胞的膨胀[36].当根区土层温度升高时,冻土消融,土壤水分含量提高,根系可利用水分增多,此外,较高的土壤温度有助于夜间根系对水分的吸收,有利于维持树木内部水分平衡[37].因此,气温和土壤温度可能是影响青海云杉径向生长开始的关键因子. ...
... 目前,针对决定径向生长结束的因素尚未达成统一共识.多项研究表明,由于秋季温度的快速下降能够改变木质部导管的形态和晚材的形成,温度是影响针叶树生长结束的重要因子[31,44].另一些研究认为,即使在较高温度环境下,水分供给不足也会导致生长提前结束,这是由于降水不足的情况下高温会降低细胞的张力和水势[34-35].最新的研究表明,土壤氮含量升高一方面延缓了木质部细胞壁增厚的完成,另一方面又促进了细胞扩大过程的结束,最终导致生长季提前结束[30].在本研究中,各样树的生长结束时间存在较大差异,且在生长结束后土壤含水量和温度均适宜,降水也相对充足,这表明生长结束不仅受水分供给和温度的影响,还受土壤氮、植被遗传特性等因子的控制[27-28,30]. ...
Photoperiod and temperature as dominant environmental drivers triggering secondary growth resumption in Northern Hemisphere conifers
1
2020
... 树木径向生长的季节性节律是其适应环境条件的综合体现,反映了树木生长开始和结束时间,进而决定了木质部的形成周期与生物量积累效率[27-30].在本研究中,海拔2 700 m和3 200 m处样树的径向生长开始时间存在显著差异,低海拔的生长开始时间普遍早于高海拔,这一现象与样地间的温度差异密切相关.已有研究表明,气温是驱动植物春季物候启动的主要气候因子,较高气温可打破休眠、启动形成层活动[31-32].对青藏高原和欧洲多种针叶树种的研究发现,当气温达到特定阈值时径向生长开始,且偶尔出现的高气温也会促进细胞分裂[24,33-35].一些研究表明,土壤温度对根系活动以及水分吸收有重要影响,从而促进树干细胞液泡膨压增加以及新形成管胞的膨胀[36].当根区土层温度升高时,冻土消融,土壤水分含量提高,根系可利用水分增多,此外,较高的土壤温度有助于夜间根系对水分的吸收,有利于维持树木内部水分平衡[37].因此,气温和土壤温度可能是影响青海云杉径向生长开始的关键因子. ...
Critical temperatures for xylogenesis in conifers of cold climates
1
2008
... 树木径向生长的季节性节律是其适应环境条件的综合体现,反映了树木生长开始和结束时间,进而决定了木质部的形成周期与生物量积累效率[27-30].在本研究中,海拔2 700 m和3 200 m处样树的径向生长开始时间存在显著差异,低海拔的生长开始时间普遍早于高海拔,这一现象与样地间的温度差异密切相关.已有研究表明,气温是驱动植物春季物候启动的主要气候因子,较高气温可打破休眠、启动形成层活动[31-32].对青藏高原和欧洲多种针叶树种的研究发现,当气温达到特定阈值时径向生长开始,且偶尔出现的高气温也会促进细胞分裂[24,33-35].一些研究表明,土壤温度对根系活动以及水分吸收有重要影响,从而促进树干细胞液泡膨压增加以及新形成管胞的膨胀[36].当根区土层温度升高时,冻土消融,土壤水分含量提高,根系可利用水分增多,此外,较高的土壤温度有助于夜间根系对水分的吸收,有利于维持树木内部水分平衡[37].因此,气温和土壤温度可能是影响青海云杉径向生长开始的关键因子. ...
Critical temperatures controlling the phenology and radial growth of Pinus sylvestris var.mongolica on the southern margin of a cold temperate coniferous forest
1
2021
... 目前,针对决定径向生长结束的因素尚未达成统一共识.多项研究表明,由于秋季温度的快速下降能够改变木质部导管的形态和晚材的形成,温度是影响针叶树生长结束的重要因子[31,44].另一些研究认为,即使在较高温度环境下,水分供给不足也会导致生长提前结束,这是由于降水不足的情况下高温会降低细胞的张力和水势[34-35].最新的研究表明,土壤氮含量升高一方面延缓了木质部细胞壁增厚的完成,另一方面又促进了细胞扩大过程的结束,最终导致生长季提前结束[30].在本研究中,各样树的生长结束时间存在较大差异,且在生长结束后土壤含水量和温度均适宜,降水也相对充足,这表明生长结束不仅受水分供给和温度的影响,还受土壤氮、植被遗传特性等因子的控制[27-28,30]. ...
Drought limits wood production of Juniperus przewalskii even as growing seasons lengthens in a cold and arid environment
2
2021
... 树木径向生长的季节性节律是其适应环境条件的综合体现,反映了树木生长开始和结束时间,进而决定了木质部的形成周期与生物量积累效率[27-30].在本研究中,海拔2 700 m和3 200 m处样树的径向生长开始时间存在显著差异,低海拔的生长开始时间普遍早于高海拔,这一现象与样地间的温度差异密切相关.已有研究表明,气温是驱动植物春季物候启动的主要气候因子,较高气温可打破休眠、启动形成层活动[31-32].对青藏高原和欧洲多种针叶树种的研究发现,当气温达到特定阈值时径向生长开始,且偶尔出现的高气温也会促进细胞分裂[24,33-35].一些研究表明,土壤温度对根系活动以及水分吸收有重要影响,从而促进树干细胞液泡膨压增加以及新形成管胞的膨胀[36].当根区土层温度升高时,冻土消融,土壤水分含量提高,根系可利用水分增多,此外,较高的土壤温度有助于夜间根系对水分的吸收,有利于维持树木内部水分平衡[37].因此,气温和土壤温度可能是影响青海云杉径向生长开始的关键因子. ...
... 目前,针对决定径向生长结束的因素尚未达成统一共识.多项研究表明,由于秋季温度的快速下降能够改变木质部导管的形态和晚材的形成,温度是影响针叶树生长结束的重要因子[31,44].另一些研究认为,即使在较高温度环境下,水分供给不足也会导致生长提前结束,这是由于降水不足的情况下高温会降低细胞的张力和水势[34-35].最新的研究表明,土壤氮含量升高一方面延缓了木质部细胞壁增厚的完成,另一方面又促进了细胞扩大过程的结束,最终导致生长季提前结束[30].在本研究中,各样树的生长结束时间存在较大差异,且在生长结束后土壤含水量和温度均适宜,降水也相对充足,这表明生长结束不仅受水分供给和温度的影响,还受土壤氮、植被遗传特性等因子的控制[27-28,30]. ...
Temporal dynamic of wood formation in Pinus cembra along the alpine treeline ecotone and the effect of climate variables
1
2009
... 树木径向生长的季节性节律是其适应环境条件的综合体现,反映了树木生长开始和结束时间,进而决定了木质部的形成周期与生物量积累效率[27-30].在本研究中,海拔2 700 m和3 200 m处样树的径向生长开始时间存在显著差异,低海拔的生长开始时间普遍早于高海拔,这一现象与样地间的温度差异密切相关.已有研究表明,气温是驱动植物春季物候启动的主要气候因子,较高气温可打破休眠、启动形成层活动[31-32].对青藏高原和欧洲多种针叶树种的研究发现,当气温达到特定阈值时径向生长开始,且偶尔出现的高气温也会促进细胞分裂[24,33-35].一些研究表明,土壤温度对根系活动以及水分吸收有重要影响,从而促进树干细胞液泡膨压增加以及新形成管胞的膨胀[36].当根区土层温度升高时,冻土消融,土壤水分含量提高,根系可利用水分增多,此外,较高的土壤温度有助于夜间根系对水分的吸收,有利于维持树木内部水分平衡[37].因此,气温和土壤温度可能是影响青海云杉径向生长开始的关键因子. ...
Influence of climate on seasonal patterns of stem increment of balsam fir in a boreal forest of Québec,Canada
2
2012
... 树木径向生长的季节性节律是其适应环境条件的综合体现,反映了树木生长开始和结束时间,进而决定了木质部的形成周期与生物量积累效率[27-30].在本研究中,海拔2 700 m和3 200 m处样树的径向生长开始时间存在显著差异,低海拔的生长开始时间普遍早于高海拔,这一现象与样地间的温度差异密切相关.已有研究表明,气温是驱动植物春季物候启动的主要气候因子,较高气温可打破休眠、启动形成层活动[31-32].对青藏高原和欧洲多种针叶树种的研究发现,当气温达到特定阈值时径向生长开始,且偶尔出现的高气温也会促进细胞分裂[24,33-35].一些研究表明,土壤温度对根系活动以及水分吸收有重要影响,从而促进树干细胞液泡膨压增加以及新形成管胞的膨胀[36].当根区土层温度升高时,冻土消融,土壤水分含量提高,根系可利用水分增多,此外,较高的土壤温度有助于夜间根系对水分的吸收,有利于维持树木内部水分平衡[37].因此,气温和土壤温度可能是影响青海云杉径向生长开始的关键因子. ...
... 先前的研究普遍认为最大生长速率往往出现在夏至日附近,与光周期密切相关[37-38].然而,本研究发现,2 700 m处树木的最大生长速率在夏至之前出现,而3 200 m处的大多数树木的最大生长速率则在夏至之后出现,这可能是由于不同海拔高度的水热条件差异所致.对高海拔针叶树的研究发现,生长季后期可能会出现水分可利用性降低的状况,树木会提前最大生长速率的出现时间[39].对榛子(Corylus avellana)的研究发现,最大生长速率出现在夏至日前的5周,这种过早达到峰值的生长模式是由于其光饱和点较低,提前进入生长旺盛期来高效利用春末夏初适宜的光照条件,从而规避夏季强光造成的光合效率下降[40-41].对挪威云杉(Norway spruce)的研究表明,最大生长速率发生在夏至日之后,这与最高气温出现时间相吻合[42].此外,另一些研究表明最大生长速率的出现时间会随着海拔的降低而呈提前趋势[43].因此,基于先前的发现与本研究结果,可以推断光周期可能控制着最大生长速率出现的大致时间,但温度、水分条件以及光饱和点等因子会精细化地对其产生影响. ...
Effects of nutrient optimization on intra-annual wood formation in Norway spruce
1
2013
... 先前的研究普遍认为最大生长速率往往出现在夏至日附近,与光周期密切相关[37-38].然而,本研究发现,2 700 m处树木的最大生长速率在夏至之前出现,而3 200 m处的大多数树木的最大生长速率则在夏至之后出现,这可能是由于不同海拔高度的水热条件差异所致.对高海拔针叶树的研究发现,生长季后期可能会出现水分可利用性降低的状况,树木会提前最大生长速率的出现时间[39].对榛子(Corylus avellana)的研究发现,最大生长速率出现在夏至日前的5周,这种过早达到峰值的生长模式是由于其光饱和点较低,提前进入生长旺盛期来高效利用春末夏初适宜的光照条件,从而规避夏季强光造成的光合效率下降[40-41].对挪威云杉(Norway spruce)的研究表明,最大生长速率发生在夏至日之后,这与最高气温出现时间相吻合[42].此外,另一些研究表明最大生长速率的出现时间会随着海拔的降低而呈提前趋势[43].因此,基于先前的发现与本研究结果,可以推断光周期可能控制着最大生长速率出现的大致时间,但温度、水分条件以及光饱和点等因子会精细化地对其产生影响. ...
极端干旱对科尔沁沙地小叶锦鸡儿(Caragana microphylla)灌丛土壤呼吸的影响
1
2025
... 先前的研究普遍认为最大生长速率往往出现在夏至日附近,与光周期密切相关[37-38].然而,本研究发现,2 700 m处树木的最大生长速率在夏至之前出现,而3 200 m处的大多数树木的最大生长速率则在夏至之后出现,这可能是由于不同海拔高度的水热条件差异所致.对高海拔针叶树的研究发现,生长季后期可能会出现水分可利用性降低的状况,树木会提前最大生长速率的出现时间[39].对榛子(Corylus avellana)的研究发现,最大生长速率出现在夏至日前的5周,这种过早达到峰值的生长模式是由于其光饱和点较低,提前进入生长旺盛期来高效利用春末夏初适宜的光照条件,从而规避夏季强光造成的光合效率下降[40-41].对挪威云杉(Norway spruce)的研究表明,最大生长速率发生在夏至日之后,这与最高气温出现时间相吻合[42].此外,另一些研究表明最大生长速率的出现时间会随着海拔的降低而呈提前趋势[43].因此,基于先前的发现与本研究结果,可以推断光周期可能控制着最大生长速率出现的大致时间,但温度、水分条件以及光饱和点等因子会精细化地对其产生影响. ...
Influence of light and shoot development stage on leaf photosynthesis and carbohydrate status during the adventitious root formation in cuttings of Corylus avellana L
1
2015
... 先前的研究普遍认为最大生长速率往往出现在夏至日附近,与光周期密切相关[37-38].然而,本研究发现,2 700 m处树木的最大生长速率在夏至之前出现,而3 200 m处的大多数树木的最大生长速率则在夏至之后出现,这可能是由于不同海拔高度的水热条件差异所致.对高海拔针叶树的研究发现,生长季后期可能会出现水分可利用性降低的状况,树木会提前最大生长速率的出现时间[39].对榛子(Corylus avellana)的研究发现,最大生长速率出现在夏至日前的5周,这种过早达到峰值的生长模式是由于其光饱和点较低,提前进入生长旺盛期来高效利用春末夏初适宜的光照条件,从而规避夏季强光造成的光合效率下降[40-41].对挪威云杉(Norway spruce)的研究表明,最大生长速率发生在夏至日之后,这与最高气温出现时间相吻合[42].此外,另一些研究表明最大生长速率的出现时间会随着海拔的降低而呈提前趋势[43].因此,基于先前的发现与本研究结果,可以推断光周期可能控制着最大生长速率出现的大致时间,但温度、水分条件以及光饱和点等因子会精细化地对其产生影响. ...
Radial stem growth dynamics and phenology of a multi-stemmed species (Corylus avellana L.) across orchards in the Northern and Southern Hemispheres
1
2021
... 先前的研究普遍认为最大生长速率往往出现在夏至日附近,与光周期密切相关[37-38].然而,本研究发现,2 700 m处树木的最大生长速率在夏至之前出现,而3 200 m处的大多数树木的最大生长速率则在夏至之后出现,这可能是由于不同海拔高度的水热条件差异所致.对高海拔针叶树的研究发现,生长季后期可能会出现水分可利用性降低的状况,树木会提前最大生长速率的出现时间[39].对榛子(Corylus avellana)的研究发现,最大生长速率出现在夏至日前的5周,这种过早达到峰值的生长模式是由于其光饱和点较低,提前进入生长旺盛期来高效利用春末夏初适宜的光照条件,从而规避夏季强光造成的光合效率下降[40-41].对挪威云杉(Norway spruce)的研究表明,最大生长速率发生在夏至日之后,这与最高气温出现时间相吻合[42].此外,另一些研究表明最大生长速率的出现时间会随着海拔的降低而呈提前趋势[43].因此,基于先前的发现与本研究结果,可以推断光周期可能控制着最大生长速率出现的大致时间,但温度、水分条件以及光饱和点等因子会精细化地对其产生影响. ...
Intra-annual tracheid production of Norway spruce and Scots pine across a latitudinal gradient in Finland
1
2014
... 先前的研究普遍认为最大生长速率往往出现在夏至日附近,与光周期密切相关[37-38].然而,本研究发现,2 700 m处树木的最大生长速率在夏至之前出现,而3 200 m处的大多数树木的最大生长速率则在夏至之后出现,这可能是由于不同海拔高度的水热条件差异所致.对高海拔针叶树的研究发现,生长季后期可能会出现水分可利用性降低的状况,树木会提前最大生长速率的出现时间[39].对榛子(Corylus avellana)的研究发现,最大生长速率出现在夏至日前的5周,这种过早达到峰值的生长模式是由于其光饱和点较低,提前进入生长旺盛期来高效利用春末夏初适宜的光照条件,从而规避夏季强光造成的光合效率下降[40-41].对挪威云杉(Norway spruce)的研究表明,最大生长速率发生在夏至日之后,这与最高气温出现时间相吻合[42].此外,另一些研究表明最大生长速率的出现时间会随着海拔的降低而呈提前趋势[43].因此,基于先前的发现与本研究结果,可以推断光周期可能控制着最大生长速率出现的大致时间,但温度、水分条件以及光饱和点等因子会精细化地对其产生影响. ...
Cambial phenology in Juniperus przewalskii along different altitudinal gradients in a cold and arid region
1
2018
... 先前的研究普遍认为最大生长速率往往出现在夏至日附近,与光周期密切相关[37-38].然而,本研究发现,2 700 m处树木的最大生长速率在夏至之前出现,而3 200 m处的大多数树木的最大生长速率则在夏至之后出现,这可能是由于不同海拔高度的水热条件差异所致.对高海拔针叶树的研究发现,生长季后期可能会出现水分可利用性降低的状况,树木会提前最大生长速率的出现时间[39].对榛子(Corylus avellana)的研究发现,最大生长速率出现在夏至日前的5周,这种过早达到峰值的生长模式是由于其光饱和点较低,提前进入生长旺盛期来高效利用春末夏初适宜的光照条件,从而规避夏季强光造成的光合效率下降[40-41].对挪威云杉(Norway spruce)的研究表明,最大生长速率发生在夏至日之后,这与最高气温出现时间相吻合[42].此外,另一些研究表明最大生长速率的出现时间会随着海拔的降低而呈提前趋势[43].因此,基于先前的发现与本研究结果,可以推断光周期可能控制着最大生长速率出现的大致时间,但温度、水分条件以及光饱和点等因子会精细化地对其产生影响. ...
Pattern of xylem phenology in conifers of cold ecosystems at the Northern Hemisphere
1
2016
... 目前,针对决定径向生长结束的因素尚未达成统一共识.多项研究表明,由于秋季温度的快速下降能够改变木质部导管的形态和晚材的形成,温度是影响针叶树生长结束的重要因子[31,44].另一些研究认为,即使在较高温度环境下,水分供给不足也会导致生长提前结束,这是由于降水不足的情况下高温会降低细胞的张力和水势[34-35].最新的研究表明,土壤氮含量升高一方面延缓了木质部细胞壁增厚的完成,另一方面又促进了细胞扩大过程的结束,最终导致生长季提前结束[30].在本研究中,各样树的生长结束时间存在较大差异,且在生长结束后土壤含水量和温度均适宜,降水也相对充足,这表明生长结束不仅受水分供给和温度的影响,还受土壤氮、植被遗传特性等因子的控制[27-28,30]. ...
Drought timing and severity affect radial growth of Picea crassifolia at different elevations in the western Qilian Mountains
1
2022
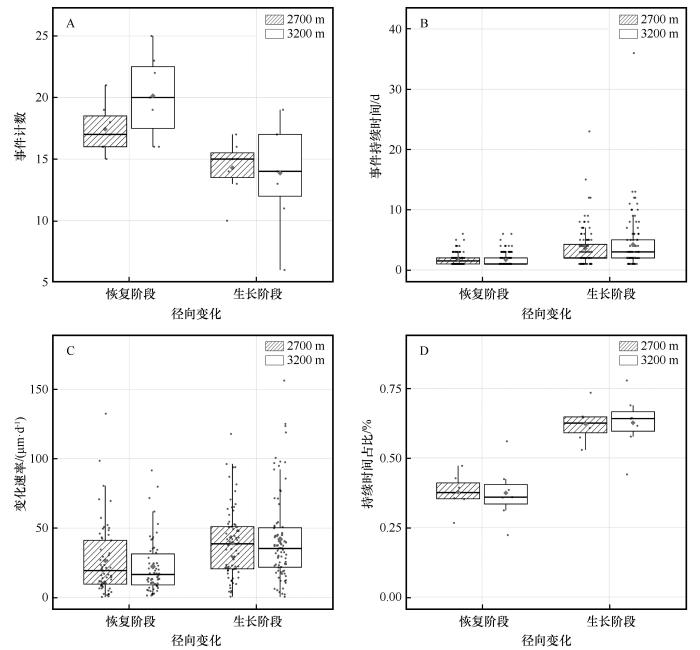
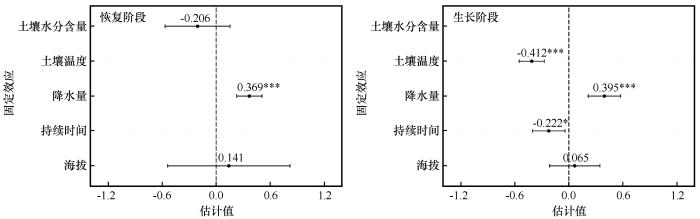
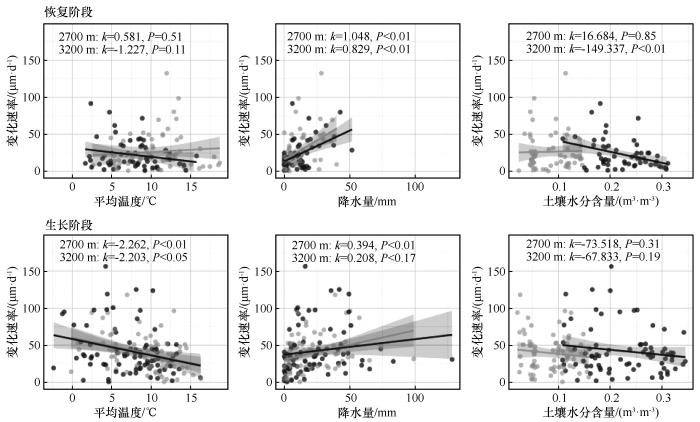
... 树木在经历干旱事件后的恢复与生长过程,反映了其应对环境压力的生理调节机制.本研究发现青海云杉径向生长对干旱的响应存在显著的海拔分异.高海拔样地恢复事件数量更多,反映其对水分改善响应更敏感,径向变化更频繁;而低海拔样地恢复事件数量虽少但变化速率略高,显示其在水分胁迫缓解后具备更快的径向变化恢复能力.因此,在恢复阶段高海拔青海云杉表现出更高的水分敏感性,而低海拔个体则具有更强的恢复力.同时,低海拔样地的生长事件数量更多且径向变化速率更高,表明其在水分条件改善后能迅速提高细胞扩张速率,实现快速生理恢复,对水分波动响应更为敏感;相比之下,高海拔样地变化速率较缓,而生长持续时间占比略高,反映其径向生长更为稳定.因此,在生长阶段低海拔青海云杉表现出更高的水分敏感性,而高海拔则更具抵抗力.这一结果印证了前人关于树木抵抗力随海拔升高而增强的结论[45],但本研究进一步揭示了不同阶段间的差异化响应,反映了树木应对干旱的阶段性及多维性适应特征,这种差异化响应主要受不同阶段、不同海拔的主导环境因子驱动.线性混合效应模型表明,恢复阶段主要由降水事件主导,其通过改善冠层微环境、快速缓解蒸腾胁迫来促进恢复[46];而生长阶段则受水热因子的协同调控,土壤温度的升高会通过增强蒸散作用对径向生长产生显著抑制[47-49].线性拟合结果进一步揭示了主导因子的海拔差异:恢复阶段,低海拔对降水更敏感[46,50],而高海拔对土壤水分含量响应更强[51-52];生长阶段,高海拔和低海拔均对平均气温敏感[53-54].这说明青海云杉的生长在不同海拔均高度依赖水分条件,但主导因子各异,证实了气候变暖正强化高海拔树木对水分依赖的观点[55-56]. ...
Disentangling the effects of vapor pressure deficit on northern terrestrial vegetation productivity
2
2023
... 树木在经历干旱事件后的恢复与生长过程,反映了其应对环境压力的生理调节机制.本研究发现青海云杉径向生长对干旱的响应存在显著的海拔分异.高海拔样地恢复事件数量更多,反映其对水分改善响应更敏感,径向变化更频繁;而低海拔样地恢复事件数量虽少但变化速率略高,显示其在水分胁迫缓解后具备更快的径向变化恢复能力.因此,在恢复阶段高海拔青海云杉表现出更高的水分敏感性,而低海拔个体则具有更强的恢复力.同时,低海拔样地的生长事件数量更多且径向变化速率更高,表明其在水分条件改善后能迅速提高细胞扩张速率,实现快速生理恢复,对水分波动响应更为敏感;相比之下,高海拔样地变化速率较缓,而生长持续时间占比略高,反映其径向生长更为稳定.因此,在生长阶段低海拔青海云杉表现出更高的水分敏感性,而高海拔则更具抵抗力.这一结果印证了前人关于树木抵抗力随海拔升高而增强的结论[45],但本研究进一步揭示了不同阶段间的差异化响应,反映了树木应对干旱的阶段性及多维性适应特征,这种差异化响应主要受不同阶段、不同海拔的主导环境因子驱动.线性混合效应模型表明,恢复阶段主要由降水事件主导,其通过改善冠层微环境、快速缓解蒸腾胁迫来促进恢复[46];而生长阶段则受水热因子的协同调控,土壤温度的升高会通过增强蒸散作用对径向生长产生显著抑制[47-49].线性拟合结果进一步揭示了主导因子的海拔差异:恢复阶段,低海拔对降水更敏感[46,50],而高海拔对土壤水分含量响应更强[51-52];生长阶段,高海拔和低海拔均对平均气温敏感[53-54].这说明青海云杉的生长在不同海拔均高度依赖水分条件,但主导因子各异,证实了气候变暖正强化高海拔树木对水分依赖的观点[55-56]. ...
... [46,50],而高海拔对土壤水分含量响应更强[51-52];生长阶段,高海拔和低海拔均对平均气温敏感[53-54].这说明青海云杉的生长在不同海拔均高度依赖水分条件,但主导因子各异,证实了气候变暖正强化高海拔树木对水分依赖的观点[55-56]. ...
Growth variations of common beech (Fagus sylvatica L.) under different climatic and environmental conditions in Europe:a dendroecological study
1
2003
... 树木在经历干旱事件后的恢复与生长过程,反映了其应对环境压力的生理调节机制.本研究发现青海云杉径向生长对干旱的响应存在显著的海拔分异.高海拔样地恢复事件数量更多,反映其对水分改善响应更敏感,径向变化更频繁;而低海拔样地恢复事件数量虽少但变化速率略高,显示其在水分胁迫缓解后具备更快的径向变化恢复能力.因此,在恢复阶段高海拔青海云杉表现出更高的水分敏感性,而低海拔个体则具有更强的恢复力.同时,低海拔样地的生长事件数量更多且径向变化速率更高,表明其在水分条件改善后能迅速提高细胞扩张速率,实现快速生理恢复,对水分波动响应更为敏感;相比之下,高海拔样地变化速率较缓,而生长持续时间占比略高,反映其径向生长更为稳定.因此,在生长阶段低海拔青海云杉表现出更高的水分敏感性,而高海拔则更具抵抗力.这一结果印证了前人关于树木抵抗力随海拔升高而增强的结论[45],但本研究进一步揭示了不同阶段间的差异化响应,反映了树木应对干旱的阶段性及多维性适应特征,这种差异化响应主要受不同阶段、不同海拔的主导环境因子驱动.线性混合效应模型表明,恢复阶段主要由降水事件主导,其通过改善冠层微环境、快速缓解蒸腾胁迫来促进恢复[46];而生长阶段则受水热因子的协同调控,土壤温度的升高会通过增强蒸散作用对径向生长产生显著抑制[47-49].线性拟合结果进一步揭示了主导因子的海拔差异:恢复阶段,低海拔对降水更敏感[46,50],而高海拔对土壤水分含量响应更强[51-52];生长阶段,高海拔和低海拔均对平均气温敏感[53-54].这说明青海云杉的生长在不同海拔均高度依赖水分条件,但主导因子各异,证实了气候变暖正强化高海拔树木对水分依赖的观点[55-56]. ...
On the ‘divergence problem’in northern forests:a review of the tree-ring evidence and possible causes
0
2008
Reduced sensitivity of recent tree-growth to temperature at high northern latitudes
1
1998
... 树木在经历干旱事件后的恢复与生长过程,反映了其应对环境压力的生理调节机制.本研究发现青海云杉径向生长对干旱的响应存在显著的海拔分异.高海拔样地恢复事件数量更多,反映其对水分改善响应更敏感,径向变化更频繁;而低海拔样地恢复事件数量虽少但变化速率略高,显示其在水分胁迫缓解后具备更快的径向变化恢复能力.因此,在恢复阶段高海拔青海云杉表现出更高的水分敏感性,而低海拔个体则具有更强的恢复力.同时,低海拔样地的生长事件数量更多且径向变化速率更高,表明其在水分条件改善后能迅速提高细胞扩张速率,实现快速生理恢复,对水分波动响应更为敏感;相比之下,高海拔样地变化速率较缓,而生长持续时间占比略高,反映其径向生长更为稳定.因此,在生长阶段低海拔青海云杉表现出更高的水分敏感性,而高海拔则更具抵抗力.这一结果印证了前人关于树木抵抗力随海拔升高而增强的结论[45],但本研究进一步揭示了不同阶段间的差异化响应,反映了树木应对干旱的阶段性及多维性适应特征,这种差异化响应主要受不同阶段、不同海拔的主导环境因子驱动.线性混合效应模型表明,恢复阶段主要由降水事件主导,其通过改善冠层微环境、快速缓解蒸腾胁迫来促进恢复[46];而生长阶段则受水热因子的协同调控,土壤温度的升高会通过增强蒸散作用对径向生长产生显著抑制[47-49].线性拟合结果进一步揭示了主导因子的海拔差异:恢复阶段,低海拔对降水更敏感[46,50],而高海拔对土壤水分含量响应更强[51-52];生长阶段,高海拔和低海拔均对平均气温敏感[53-54].这说明青海云杉的生长在不同海拔均高度依赖水分条件,但主导因子各异,证实了气候变暖正强化高海拔树木对水分依赖的观点[55-56]. ...
Increasing atmospheric dryness reduces boreal forest tree growth
1
2023
... 树木在经历干旱事件后的恢复与生长过程,反映了其应对环境压力的生理调节机制.本研究发现青海云杉径向生长对干旱的响应存在显著的海拔分异.高海拔样地恢复事件数量更多,反映其对水分改善响应更敏感,径向变化更频繁;而低海拔样地恢复事件数量虽少但变化速率略高,显示其在水分胁迫缓解后具备更快的径向变化恢复能力.因此,在恢复阶段高海拔青海云杉表现出更高的水分敏感性,而低海拔个体则具有更强的恢复力.同时,低海拔样地的生长事件数量更多且径向变化速率更高,表明其在水分条件改善后能迅速提高细胞扩张速率,实现快速生理恢复,对水分波动响应更为敏感;相比之下,高海拔样地变化速率较缓,而生长持续时间占比略高,反映其径向生长更为稳定.因此,在生长阶段低海拔青海云杉表现出更高的水分敏感性,而高海拔则更具抵抗力.这一结果印证了前人关于树木抵抗力随海拔升高而增强的结论[45],但本研究进一步揭示了不同阶段间的差异化响应,反映了树木应对干旱的阶段性及多维性适应特征,这种差异化响应主要受不同阶段、不同海拔的主导环境因子驱动.线性混合效应模型表明,恢复阶段主要由降水事件主导,其通过改善冠层微环境、快速缓解蒸腾胁迫来促进恢复[46];而生长阶段则受水热因子的协同调控,土壤温度的升高会通过增强蒸散作用对径向生长产生显著抑制[47-49].线性拟合结果进一步揭示了主导因子的海拔差异:恢复阶段,低海拔对降水更敏感[46,50],而高海拔对土壤水分含量响应更强[51-52];生长阶段,高海拔和低海拔均对平均气温敏感[53-54].这说明青海云杉的生长在不同海拔均高度依赖水分条件,但主导因子各异,证实了气候变暖正强化高海拔树木对水分依赖的观点[55-56]. ...
Moisture availability limits subalpine tree establishment
1
2018
... 树木在经历干旱事件后的恢复与生长过程,反映了其应对环境压力的生理调节机制.本研究发现青海云杉径向生长对干旱的响应存在显著的海拔分异.高海拔样地恢复事件数量更多,反映其对水分改善响应更敏感,径向变化更频繁;而低海拔样地恢复事件数量虽少但变化速率略高,显示其在水分胁迫缓解后具备更快的径向变化恢复能力.因此,在恢复阶段高海拔青海云杉表现出更高的水分敏感性,而低海拔个体则具有更强的恢复力.同时,低海拔样地的生长事件数量更多且径向变化速率更高,表明其在水分条件改善后能迅速提高细胞扩张速率,实现快速生理恢复,对水分波动响应更为敏感;相比之下,高海拔样地变化速率较缓,而生长持续时间占比略高,反映其径向生长更为稳定.因此,在生长阶段低海拔青海云杉表现出更高的水分敏感性,而高海拔则更具抵抗力.这一结果印证了前人关于树木抵抗力随海拔升高而增强的结论[45],但本研究进一步揭示了不同阶段间的差异化响应,反映了树木应对干旱的阶段性及多维性适应特征,这种差异化响应主要受不同阶段、不同海拔的主导环境因子驱动.线性混合效应模型表明,恢复阶段主要由降水事件主导,其通过改善冠层微环境、快速缓解蒸腾胁迫来促进恢复[46];而生长阶段则受水热因子的协同调控,土壤温度的升高会通过增强蒸散作用对径向生长产生显著抑制[47-49].线性拟合结果进一步揭示了主导因子的海拔差异:恢复阶段,低海拔对降水更敏感[46,50],而高海拔对土壤水分含量响应更强[51-52];生长阶段,高海拔和低海拔均对平均气温敏感[53-54].这说明青海云杉的生长在不同海拔均高度依赖水分条件,但主导因子各异,证实了气候变暖正强化高海拔树木对水分依赖的观点[55-56]. ...
Canopy structure modulates the sensitivity of subalpine forest stands to interannual snowpack and precipitation variability
1
2025
... 树木在经历干旱事件后的恢复与生长过程,反映了其应对环境压力的生理调节机制.本研究发现青海云杉径向生长对干旱的响应存在显著的海拔分异.高海拔样地恢复事件数量更多,反映其对水分改善响应更敏感,径向变化更频繁;而低海拔样地恢复事件数量虽少但变化速率略高,显示其在水分胁迫缓解后具备更快的径向变化恢复能力.因此,在恢复阶段高海拔青海云杉表现出更高的水分敏感性,而低海拔个体则具有更强的恢复力.同时,低海拔样地的生长事件数量更多且径向变化速率更高,表明其在水分条件改善后能迅速提高细胞扩张速率,实现快速生理恢复,对水分波动响应更为敏感;相比之下,高海拔样地变化速率较缓,而生长持续时间占比略高,反映其径向生长更为稳定.因此,在生长阶段低海拔青海云杉表现出更高的水分敏感性,而高海拔则更具抵抗力.这一结果印证了前人关于树木抵抗力随海拔升高而增强的结论[45],但本研究进一步揭示了不同阶段间的差异化响应,反映了树木应对干旱的阶段性及多维性适应特征,这种差异化响应主要受不同阶段、不同海拔的主导环境因子驱动.线性混合效应模型表明,恢复阶段主要由降水事件主导,其通过改善冠层微环境、快速缓解蒸腾胁迫来促进恢复[46];而生长阶段则受水热因子的协同调控,土壤温度的升高会通过增强蒸散作用对径向生长产生显著抑制[47-49].线性拟合结果进一步揭示了主导因子的海拔差异:恢复阶段,低海拔对降水更敏感[46,50],而高海拔对土壤水分含量响应更强[51-52];生长阶段,高海拔和低海拔均对平均气温敏感[53-54].这说明青海云杉的生长在不同海拔均高度依赖水分条件,但主导因子各异,证实了气候变暖正强化高海拔树木对水分依赖的观点[55-56]. ...
Tree growth and its climate signal along latitudinal and altitudinal gradients:comparison of tree rings between Finland and the Tibetan Plateau
1
2017
... 树木在经历干旱事件后的恢复与生长过程,反映了其应对环境压力的生理调节机制.本研究发现青海云杉径向生长对干旱的响应存在显著的海拔分异.高海拔样地恢复事件数量更多,反映其对水分改善响应更敏感,径向变化更频繁;而低海拔样地恢复事件数量虽少但变化速率略高,显示其在水分胁迫缓解后具备更快的径向变化恢复能力.因此,在恢复阶段高海拔青海云杉表现出更高的水分敏感性,而低海拔个体则具有更强的恢复力.同时,低海拔样地的生长事件数量更多且径向变化速率更高,表明其在水分条件改善后能迅速提高细胞扩张速率,实现快速生理恢复,对水分波动响应更为敏感;相比之下,高海拔样地变化速率较缓,而生长持续时间占比略高,反映其径向生长更为稳定.因此,在生长阶段低海拔青海云杉表现出更高的水分敏感性,而高海拔则更具抵抗力.这一结果印证了前人关于树木抵抗力随海拔升高而增强的结论[45],但本研究进一步揭示了不同阶段间的差异化响应,反映了树木应对干旱的阶段性及多维性适应特征,这种差异化响应主要受不同阶段、不同海拔的主导环境因子驱动.线性混合效应模型表明,恢复阶段主要由降水事件主导,其通过改善冠层微环境、快速缓解蒸腾胁迫来促进恢复[46];而生长阶段则受水热因子的协同调控,土壤温度的升高会通过增强蒸散作用对径向生长产生显著抑制[47-49].线性拟合结果进一步揭示了主导因子的海拔差异:恢复阶段,低海拔对降水更敏感[46,50],而高海拔对土壤水分含量响应更强[51-52];生长阶段,高海拔和低海拔均对平均气温敏感[53-54].这说明青海云杉的生长在不同海拔均高度依赖水分条件,但主导因子各异,证实了气候变暖正强化高海拔树木对水分依赖的观点[55-56]. ...
Warming,drought,and disturbances lead to shifts in functional composition:a millennial-scale analysis for Amazonian and Andean sites
1
2023
... 树木在经历干旱事件后的恢复与生长过程,反映了其应对环境压力的生理调节机制.本研究发现青海云杉径向生长对干旱的响应存在显著的海拔分异.高海拔样地恢复事件数量更多,反映其对水分改善响应更敏感,径向变化更频繁;而低海拔样地恢复事件数量虽少但变化速率略高,显示其在水分胁迫缓解后具备更快的径向变化恢复能力.因此,在恢复阶段高海拔青海云杉表现出更高的水分敏感性,而低海拔个体则具有更强的恢复力.同时,低海拔样地的生长事件数量更多且径向变化速率更高,表明其在水分条件改善后能迅速提高细胞扩张速率,实现快速生理恢复,对水分波动响应更为敏感;相比之下,高海拔样地变化速率较缓,而生长持续时间占比略高,反映其径向生长更为稳定.因此,在生长阶段低海拔青海云杉表现出更高的水分敏感性,而高海拔则更具抵抗力.这一结果印证了前人关于树木抵抗力随海拔升高而增强的结论[45],但本研究进一步揭示了不同阶段间的差异化响应,反映了树木应对干旱的阶段性及多维性适应特征,这种差异化响应主要受不同阶段、不同海拔的主导环境因子驱动.线性混合效应模型表明,恢复阶段主要由降水事件主导,其通过改善冠层微环境、快速缓解蒸腾胁迫来促进恢复[46];而生长阶段则受水热因子的协同调控,土壤温度的升高会通过增强蒸散作用对径向生长产生显著抑制[47-49].线性拟合结果进一步揭示了主导因子的海拔差异:恢复阶段,低海拔对降水更敏感[46,50],而高海拔对土壤水分含量响应更强[51-52];生长阶段,高海拔和低海拔均对平均气温敏感[53-54].这说明青海云杉的生长在不同海拔均高度依赖水分条件,但主导因子各异,证实了气候变暖正强化高海拔树木对水分依赖的观点[55-56]. ...
Anomalous temperature-growth response of Abies faxoniana to sustained freezing stress along elevational gradients in China's Western Sichuan Province
1
2012
... 树木在经历干旱事件后的恢复与生长过程,反映了其应对环境压力的生理调节机制.本研究发现青海云杉径向生长对干旱的响应存在显著的海拔分异.高海拔样地恢复事件数量更多,反映其对水分改善响应更敏感,径向变化更频繁;而低海拔样地恢复事件数量虽少但变化速率略高,显示其在水分胁迫缓解后具备更快的径向变化恢复能力.因此,在恢复阶段高海拔青海云杉表现出更高的水分敏感性,而低海拔个体则具有更强的恢复力.同时,低海拔样地的生长事件数量更多且径向变化速率更高,表明其在水分条件改善后能迅速提高细胞扩张速率,实现快速生理恢复,对水分波动响应更为敏感;相比之下,高海拔样地变化速率较缓,而生长持续时间占比略高,反映其径向生长更为稳定.因此,在生长阶段低海拔青海云杉表现出更高的水分敏感性,而高海拔则更具抵抗力.这一结果印证了前人关于树木抵抗力随海拔升高而增强的结论[45],但本研究进一步揭示了不同阶段间的差异化响应,反映了树木应对干旱的阶段性及多维性适应特征,这种差异化响应主要受不同阶段、不同海拔的主导环境因子驱动.线性混合效应模型表明,恢复阶段主要由降水事件主导,其通过改善冠层微环境、快速缓解蒸腾胁迫来促进恢复[46];而生长阶段则受水热因子的协同调控,土壤温度的升高会通过增强蒸散作用对径向生长产生显著抑制[47-49].线性拟合结果进一步揭示了主导因子的海拔差异:恢复阶段,低海拔对降水更敏感[46,50],而高海拔对土壤水分含量响应更强[51-52];生长阶段,高海拔和低海拔均对平均气温敏感[53-54].这说明青海云杉的生长在不同海拔均高度依赖水分条件,但主导因子各异,证实了气候变暖正强化高海拔树木对水分依赖的观点[55-56]. ...
Climate signals from tree ring chronologies of the upper and lower treelines in the Dulan region of the northeastern Qinghai‐Tibetan Plateau
1
2006
... 树木在经历干旱事件后的恢复与生长过程,反映了其应对环境压力的生理调节机制.本研究发现青海云杉径向生长对干旱的响应存在显著的海拔分异.高海拔样地恢复事件数量更多,反映其对水分改善响应更敏感,径向变化更频繁;而低海拔样地恢复事件数量虽少但变化速率略高,显示其在水分胁迫缓解后具备更快的径向变化恢复能力.因此,在恢复阶段高海拔青海云杉表现出更高的水分敏感性,而低海拔个体则具有更强的恢复力.同时,低海拔样地的生长事件数量更多且径向变化速率更高,表明其在水分条件改善后能迅速提高细胞扩张速率,实现快速生理恢复,对水分波动响应更为敏感;相比之下,高海拔样地变化速率较缓,而生长持续时间占比略高,反映其径向生长更为稳定.因此,在生长阶段低海拔青海云杉表现出更高的水分敏感性,而高海拔则更具抵抗力.这一结果印证了前人关于树木抵抗力随海拔升高而增强的结论[45],但本研究进一步揭示了不同阶段间的差异化响应,反映了树木应对干旱的阶段性及多维性适应特征,这种差异化响应主要受不同阶段、不同海拔的主导环境因子驱动.线性混合效应模型表明,恢复阶段主要由降水事件主导,其通过改善冠层微环境、快速缓解蒸腾胁迫来促进恢复[46];而生长阶段则受水热因子的协同调控,土壤温度的升高会通过增强蒸散作用对径向生长产生显著抑制[47-49].线性拟合结果进一步揭示了主导因子的海拔差异:恢复阶段,低海拔对降水更敏感[46,50],而高海拔对土壤水分含量响应更强[51-52];生长阶段,高海拔和低海拔均对平均气温敏感[53-54].这说明青海云杉的生长在不同海拔均高度依赖水分条件,但主导因子各异,证实了气候变暖正强化高海拔树木对水分依赖的观点[55-56]. ...




 甘公网安备 62010202000688号
甘公网安备 62010202000688号
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}